

Abstract
Climate change might pose an additional threat to the already vulnerable giant panda (Ailuropoda melanoleuca). Effective conservation efforts require projections of vulnerability of the giant panda in facing climate change and proactive strategies to reduce emerging climate‐related threats. We used the maximum entropy model to assess the vulnerability of giant panda to climate change in the Qinling Mountains of China. The results of modeling included the following findings: (1) the area of suitable habitat for giant pandas was projected to decrease by 281 km2 from climate change by the 2050s; (2) the mean elevation of suitable habitat of giant panda was predicted to shift 30 m higher due to climate change over this period; (3) the network of nature reserves protect 61.73% of current suitable habitat for the species, and 59.23% of future suitable habitat; (4) current suitable habitat mainly located in Chenggu, Taibai, and Yangxian counties (with a total area of 987 km2) was predicted to be vulnerable. Assessing the vulnerability of giant panda provided adaptive strategies for conservation programs and national park construction. We proposed adaptation strategies to ameliorate the predicted impacts of climate change on giant panda, including establishing and adjusting reserves, establishing habitat corridors, improving adaptive capacity to climate change, and strengthening monitoring of giant panda.
Keywords: adaptive conservation strategies, Maxent, nature reserve, suitable habitat, vulnerability
1. Introduction
Rapid climate change has been widely recognized as a major threat to biodiversity (Cramer et al., 2014). Compelling evidence has already been presented of the effects of climate change on geographic distributions (Ancillotto, Santini, Ranc, Maiorano, & Russo, 2016; Molina‐Martínez et al., 2016), population dynamics (Auer & Martin, 2013; Lehikoinen et al., 2016), phenological phase (Lučan, Weiser, & Hanák, 2013; Yang & Rudolf, 2010), and evolution (Charmantier & Gienapp, 2014; Koen, Bowman, Murray, & Wilson, 2014), and these impacts are predicted to be exacerbated in future (Rinawati, Stein, & Lindner, 2013; Urban, 2015). Projected change rates of climate are now getting faster than they were in the past (IPCC, 2014). If global warming is not effectively controlled, a mean increase in global temperature of >2.0°C could be the result (2.0°C is defined as “dangerous”; UNFCCC, 2015), and 15%–35% of global species could be committed to extinction (Thomas et al., 2004). Although the impact of climate change on the extent and rate of species extinction is still controversial, it is clear that the trend of global warming will accelerate the extinction risk for species (Malcolm, Liu, Neilson, Hansen, & Hannah, 2006; Pereira et al., 2010; Urban, 2015).
Faced with an irrefutable crisis of biodiversity loss, it is imperative to assess the vulnerability of species to future climate change, and adopt conservation strategies to mitigate the harmful impacts of climate change on these species (Heikkinen, Luoto, Leikola, Pöyry, & Settele, 2010; Polaina, Revilla, & González‐Suárez, 2016; Tuberville, Andrews, Sperry, & Grosse, 2015; Williams, Shoo, Isaac, Hoffmann, & Langham, 2008). Assessments of species vulnerability to climate change are usually based on available information of the species being assessed (Glick, Stein, & Edelson, 2011; Pacifici et al., 2015; Rowland, Davison, & Graumlich, 2011). A few tools and approaches have been developed to assess species’ vulnerability to climate change, such as vulnerability indices (Bagne, Friggens, & Finch, 2011; Foden et al., 2013; Young et al., 2015), mechanistic distribution models (Kearney & Porter, 2009; Monahan, 2009), and bioclimatic envelope models (Lawler, Shafer, & Bancroft, 2010; Pearson et al., 2014). Bioclimatic envelope models are one of the most common approaches, because they generally require only robust data on species ranges and an associated climate database (Rowland et al., 2011). Spatial shifts in climatically suitable habitat under climate change scenarios are then forecasted (Kane, Burkett, Kloper, & Sewall, 2013; Rowland et al., 2011; Thuiller, Lavorel, & Araújo, 2005). Identifying species’ potential range shifts is crucial for management and conservation of vulnerable species in a changing climate (Heikkinen et al., 2010).
The giant panda (Ailuropoda melanoleuca) is probably one of the world's most treasured endangered species (Wei et al., 2015). Its habitat is currently restricted to six isolated mountain ranges in Sichuan, Shaanxi, and Gansu provinces in south‐central China (State Forestry Administration, 2015). The giant panda was listed as an endangered species by the International Union for Conservation of Nature (IUCN) in 1996 (IUCN, 1996) due to their limited geographic range, the risk of small and isolated populations, low reproductive rates, habitat loss, and diet specialization (Swaisgood, Wang, & Wei, 2016; Wang, Ye, Skidmore, & Toxopeus, 2010; Wei et al., 2015). A narrow geographic distribution makes them highly susceptible to climate change (Liu, Guang, Dai, Li, & Gong, 2016; Songer, Delion, Biggs, & Huang, 2012). Over the past decades, the Chinese government implemented many conservation programs to protect giant panda, such as establishment of reserves (State Forestry Administration, 2015), the panda monitoring project (Wei et al., 2015), and the Grain‐to‐Green program (Li et al., 2013). From 1988 to 2015, the population of giant panda grew from 1,114 to 1,864 (State Forestry Administration, 2015), and the species has been downlisted from “Endangered” to “Vulnerable” in the IUCN Red List of Threatened Species (Swaisgood et al., 2016). The Chinese government announced that giant panda conservation programs will continue and will establish national parks in the giant panda's range to specifically strengthen further conservation of giant panda (State Forestry Administration, 2016a). Therefore, a major motivation for assessing the vulnerability of giant panda is to provide adaptive strategies for conservation programs and development of national parks to reduce effectively potential climate‐related threats to the species.
In this study, we use the maximum entropy model (i.e., Maxent, Phillips, Anderson, & Schapire, 2006) to predict the habitat suitability, to assess vulnerability of the giant panda to climate change, and to identify the potential refuges and corridors. Furthermore, we propose the conservation strategies for the species and provide fundamental information for establishing giant panda national parks in the Qinling Mountains of China.
2. Methods
2.1. Study area
The study area is located in the Qinling Mountains (106°30′–108°05′E, 32°40′–34°35′N) in Shaanxi Province in China. The Qinling Mountains are characterized by a specific geographic system in terms of topography and climate, and include the boundary between the temperate and subtropical zones (Zhao, Zhang, & Dong, 2014). The mountains rise from 222 m to 3,734 m, with a gentler gradient on the southern slope; however, their northern face is generally steep (Pan et al., 2001). Regarding the differences in climate between northern and southern China, the southern slope is generally warmer and moister than the northern face, and climatic conditions vary with elevation gradient (Pan et al., 2001). Deciduous broadleaf and subtropical evergreen forests mainly inhabit at low elevation; temperate broadleaf and subalpine coniferous forests inhabit at mid‐elevation; and subalpine scrub meadows inhabit at high elevation (State Forestry Administration, 2006). A population of 345 giant pandas was estimated to inhabit the Qinling Mountains (State Forestry Administration, 2015). A total of 19 nature reserves (including eight national and nine provincial nature reserves, and two at the application stage) have been established to protect giant panda and their habitat in this region (State Forestry Administration, 2015).
2.2. Data preparation
Locations of giant panda's signs (including feces, dens, bed sites, and footprints, N = 273) were obtained from the 3rd National Survey Report on Giant Panda in China (State Forestry Administration, 2006; Figure 1). Model parameters require an unbiased sample; therefore, we filtered the presence points by randomly choosing one record per 1 × 1 km cell. To correct the effect of sample selection bias on predictive performance (Phillips et al., 2009), we created 10,000 random points as target‐group background points in our study area and the random points were generated from any forest in the Qinling Mountains (the forest data were derived from global land cover, which were interpreted by the United Nations Food and Agricultural Organization; Charmantier & Gienapp, 2014; Tateshi et al., 2014).
Figure 1.
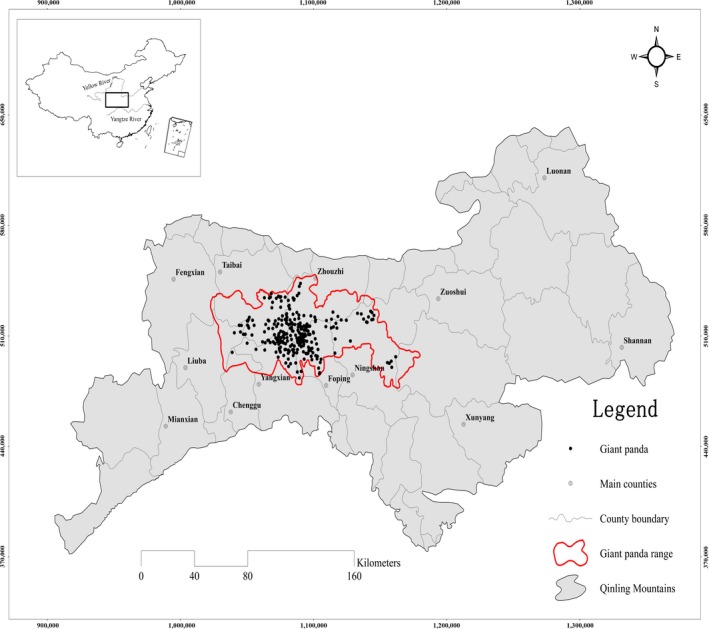
Distribution of giant pandas in Qinling Mountains
Nineteen bioclimatic variables at 30 s resolution (~1 km) were obtained from the WorldClim database (Hijmans, Cameron, Parra, Jones, & Jarvis, 2005) for current climatic (average for 1950–2000) and future climatic scenarios for the 2050s (average for 2041–2060; available at http://www.worldclim.org/version1). The future climate data applied in this study comprised of IPCC‐CMIP5 climate projection from the Met Office Hadley Center for climate change coupled model (HadGEM2‐AO) under the representative concentration pathway (RCP) 4.5 (Baek et al., 2013). For the 2050s, the average increase in global temperature of 0.9–2.0°C under RCP4.5 would fall within a 2°C global warming limit (UNFCCC, 2015). The time horizon of the 2050s was selected for being a date far enough in future for significant changes to have occurred (Young et al., 2012).
Other environmental variables were also used to construct the panda distribution models (Fan et al., 2011; Loucks et al., 2003). The densities of rivers, roads, and settlements were obtained from a 1:1,000,000 map of China (National Geomatics Center of China, data are available at http://atgcc.sbsm.gov.cn). Elevation data were derived from a digital elevation model with a resolution of 30 s, obtained from the WorldClim database (Appendix 1). Because nonclimate variables (i.e., densities of roads, rivers, and settlements) were not available for the 2050s, we used the same variables in projections for the 2050s.
All spatial layers were resampled into resolution of 1 × 1 km and projected to an equal area projection (Asia North Albers Equal Area Conic) using ArcGIS 10.1 (ESRI Inc., Redlands, CA, USA). We then calculated correlation coefficients between variables and eliminated one variable from each pair that was strongly correlated (|r| > .8; Cord, Klein, Mora, & Dech, 2014; Lemke, Hulme, Brown, & Tadesse, 2011). Thirteen variables (annual precipitation, annual temperature range, density of rivers, density of roads, density of settlements, elevation, mean diurnal range, min. temperature of coldest month, precipitation of warmest quarter, precipitation seasonality, precipitation of driest quarter, precipitation of driest month, and temperature constancy) which were the most biologically meaningful for giant pandas were retained (Appendix 2; Li, Xu, Wong, Qiu, & Li, 2015; Songer et al., 2012). Subsequently, we first input thirteen environmental variables layer into the Maxent model. Then, we input the set of most important variables based on permutation importance obtained from first model output, to construct the giant panda finally distribution model, and rerun the Maxent models.
2.3. Habitat suitability model
We used the Maxent software (version 3.3.3k) to build the habitat suitability model for the giant panda. This approach is considered one of the best performing algorithms in predicting species distribution with presence‐only data (Elith, Phillips, Hastie, Dudík, & Chee, 2011). It has been extensively applied to project species range shifts under climate change (Li, Clinton, et al., 2015; Lei, Xu, Cui, Guang, & Ding, 2014; Songer et al., 2012). Maxent estimates species distributions by finding the probability distribution of maximum entropy, subject to the constraints of the data that are available (Phillips et al., 2006). Maxent also estimates the importance of variables and contributions representing the degree to which each variable has contributed to the model, based on jackknife tests. We divided the occurrence data of giant panda into training sets (75%) for model building, and testing sets (25%) for model evaluation, and conducted a subsample procedure (Khatchikian, Sangermano, Kendell, & Livdahl, 2011; Wisz et al., 2008) to evaluate the habitat suitability model by performing 15 replications in Maxent.
Model performance was measured using the area under the receiver operating characteristic curve (AUC). An AUC value closer to 1 represents near perfect performance of the model (Phillips et al., 2006). The output of Maxent comprised continuous values between 0 and 1 that were considered as probabilities of species’ occurrence. We then convert these values to presence and absence predictions, based on the threshold values that maximized training sensitivity plus specificity (Liu, Berry, Dawson, & Pearson, 2005; Songer et al., 2012). The cells with probability values above the threshold value were selected as suitable habitat for the giant panda. We then removed patches <4 km2 and >0.5 km distance from the nearest patch based on the minimum home range size and the average daily dispersal ability of specie (Pan et al., 2001). A Mann–Whitney U test was used to examine the difference in mean elevation of suitable habitat between current and the 2050s. Statistical analyses were conducted using the SPSS 19.0 software (IBM Inc., USA).
2.4. Gap analysis of nature reserves
The Gap analysis of protection of biodiversity is a powerful and efficient step to first assess the protection of biodiversity on a coarse‐filter scale (Scott et al., 1993). The current and projected suitable habitat were overlapped with the boundaries of established nature reserve networks, to explore the conservation effectiveness of these reserves in protecting giant pandas under climate change (Feeley & Silman, 2016).
2.5. Vulnerability assessment
The identification of vulnerable habitat of species under climate change scenario is important for decision‐making in adaptive conservation management (Guisan, Tingley, Baumgartner, Naujokaitis‐Lewis, & Sucliffe, 2013). Suitable habitat changes between the current and the 2050s illustrate the locations that likely would be vulnerable, categorized as follows:
Unchanged suitable habitat: the area where suitable habitat overlapped between current and the 2050s;
Vulnerable habitat: the area where current suitable habitat transferred to unsuitable habitat by the 2050s;
New increased suitability habitat: the area where current unsuitable habitat changed to suitable habitat by the 2050s;
Unsuitable habitat: the area where unsuitable habitat overlapped between current and the 2050s.
We used three indicators to assess the impacts of climate change on giant panda: (1) percentage of area change (AC); (2) percentage of loss area of current suitable habitat (SHc); and (3) percentage of increased area of the 2050s’ suitable habitat (SHf). Indicators calculated as follows:
| (1) |
| (2) |
| (3) |
In these formulas, A f is area of projected suitable habitat for pandas under the 2050s’ climatic scenario; A c is the area of projected current suitable habitat; A fc is the overlapped distribution space between current and the 2050s (Duan, Kong, Huang, Vaerla, & Ji, 2016; Levinsky, Skov, Svenning, & Rahbek, 2007; Thuiller, Lavorel, Araújo, Sykes, & Prentice, 2005).
3. Results
3.1. Species distribution model
In the Maxent model, 256 presence points and nine variables were finally used as model parameters to construct the giant panda distribution model. The average training AUC was 0.967 ± 0.001, and the average testing AUC was 0.961 ± 0.005. The permutation importance of variables in the model as ranked from the highest to the lowest were as follows: density of rivers (34.6%), annual precipitation (30.4%), mean diurnal range (15.4%), precipitation seasonality (5.3%), precipitation of warmest quarter (4.4%), density of roads (3.6%), annual temperature range (3.1%), density of settlements (2.0%), and temperature constancy (1.3%; Figure 2). The average threshold for the probability of presence at maximum training sensitivity plus specificity was .1434. We then defined the cells with probability values greater than .1434 as suitable habitat for giant panda.
Figure 2.
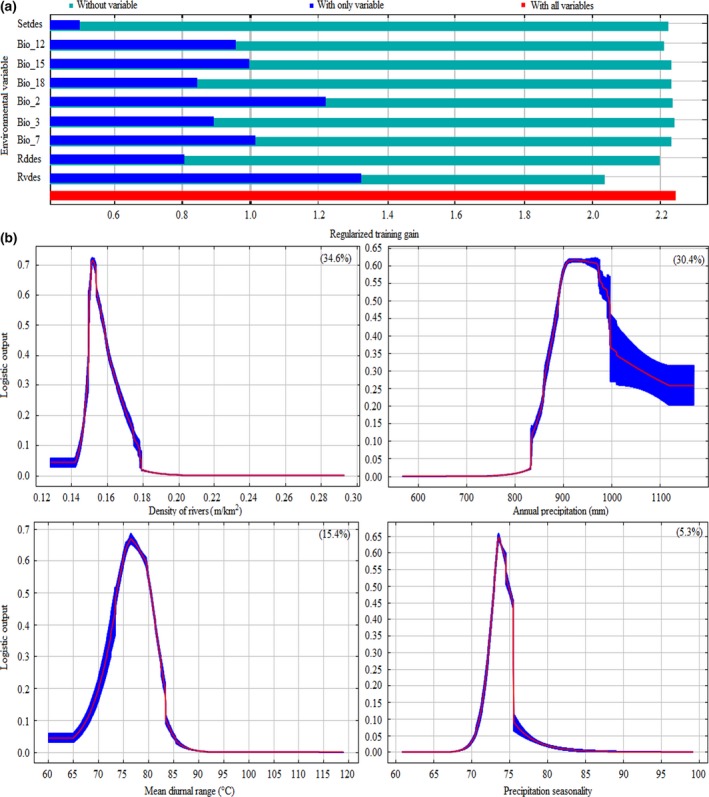
Results of Maxent models: (a) Jackknife test of variable importance. Codes of the variables are found in Appendix 1; and (b) marginal response curves of environmental variables in Maxent model of giant panda
3.2. Suitable habitat change
Under the current conditions, area of suitable habitat for giant panda in the Qinling Mountains was 4,810 km2. Current suitable habitat for giant panda is distributed in Chenggu, Foping, Liuba, Ningshan, Taibai, Yangxian, and Zhouzhi counties. For the 2050s, a reduction to 4,529 km2 (AC = −5.8%) in the area of suitable habitat was projected, and mainly distributed among Foping, Ningshan, Taibai, Yangxian, and Zhouzhi counties (Figure 3). Climate change would result in the shift of suitable habitat for giant panda to higher elevations. The mean elevation of suitable habitat in the 2050s was projected to be 1,870.57 ± 418.57 m, which was significantly higher (Z = −3.877, p = .000) than that of current suitability habitat (1,837.41 ± 432.24 m).
Figure 3.
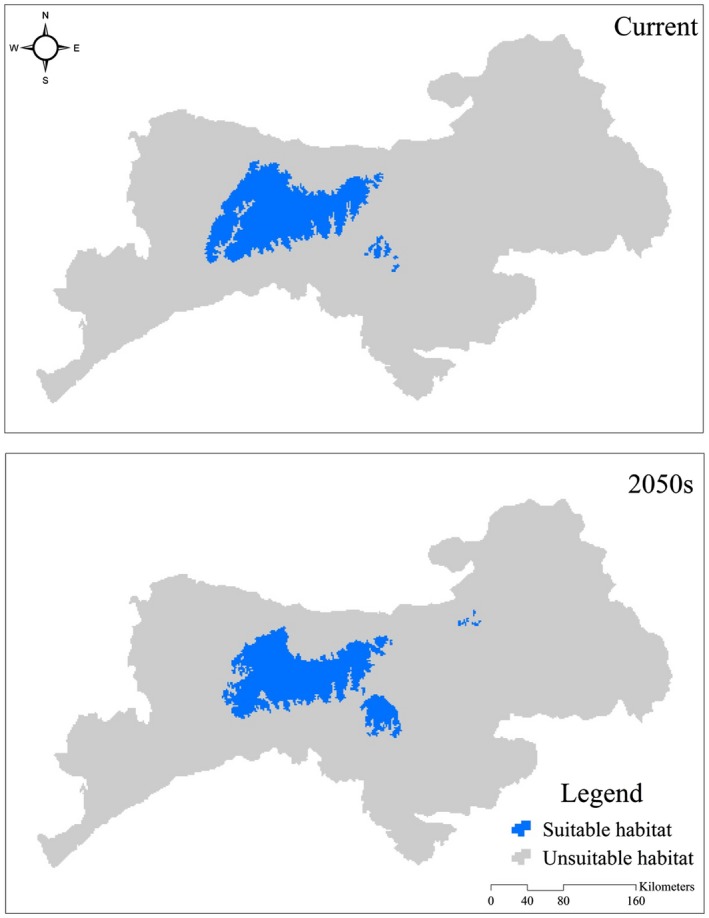
Predicted current and the 2050s’ suitable habitat for giant panda in Qinling Mountains
3.3. Gap analysis of nature reserve network
Nature reserves protect 61.73% of current suitable habitat and 59.23% of suitable habitat in the 2050s (Table 1, Figure 4). In the 2050s, giant panda suitable habitat is predicted to suffer loss in nine nature reserves, among which Banqiao (AC = −21.66%), Motianling (AC = −100%), Niangniangshan (AC = −22.20%), Panlong (AC = −35.44%), Sangyuan (AC = −92.46%), Tiabaishan (AC = −22.83%), and Zhouzhi (AC = −9.87%) nature reserves estimated to suffer the greatest loss of suitable habitat in the 2050s. Climate change will increase the extent of the panda's distribution, mainly in the Huangguanshan, Pingheliang, and Yingzuishi nature reserves (Table 1).
Table 1.
Projected change in suitable habitat of giant pandas in nature reserves
| Nature reserve | Suitable habitat area/(km2) | Percentage of area change (AC) | |
|---|---|---|---|
| Current | 2050s | ||
| 1—Banqiao | 309.20 | 242.23 | −21.66 |
| 2—Changqing | 307.21 | 295.96 | −3.66 |
| 3—Foping | 300.00 | 300.00 | 0.00 |
| 4—Guanyinshan | 148.44 | 148.44 | 0.00 |
| 5—Hanzhongzhuhuan | 0.00 | 0.00 | — |
| 6—Huangbaiyuan | 210.79 | 210.79 | 0.00 |
| 7—Huangguanshan | 81.28 | 129.75 | 59.64 |
| 8—Laoxiancheng | 120.57 | 120.57 | 0.00 |
| 9—Motianling | 47.56 | 0.00 | −100.00 |
| 10—Niangniangshan | 96.49 | 75.24 | −22.02 |
| 11—Niubeiliang | 0.00 | 0.00 | — |
| 12—Panlong | 158.13 | 102.10 | −35.44 |
| 13—Pingheliang | 12.36 | 58.54 | 373.63 |
| 14—Sangyuan | 120.05 | 9.05 | −92.46 |
| 15—Taibainiuweihe | 106.98 | 102.77 | −3.94 |
| 16—Taibaishan | 348.93 | 269.28 | −22.83 |
| 17—Tianhuashan | 273.31 | 285.90 | 4.60 |
| 18—Yingzuishi | 5.78 | 41.33 | 615.41 |
| 19—Zhouzhi | 322.31 | 290.50 | −9.87 |
Figure 4.
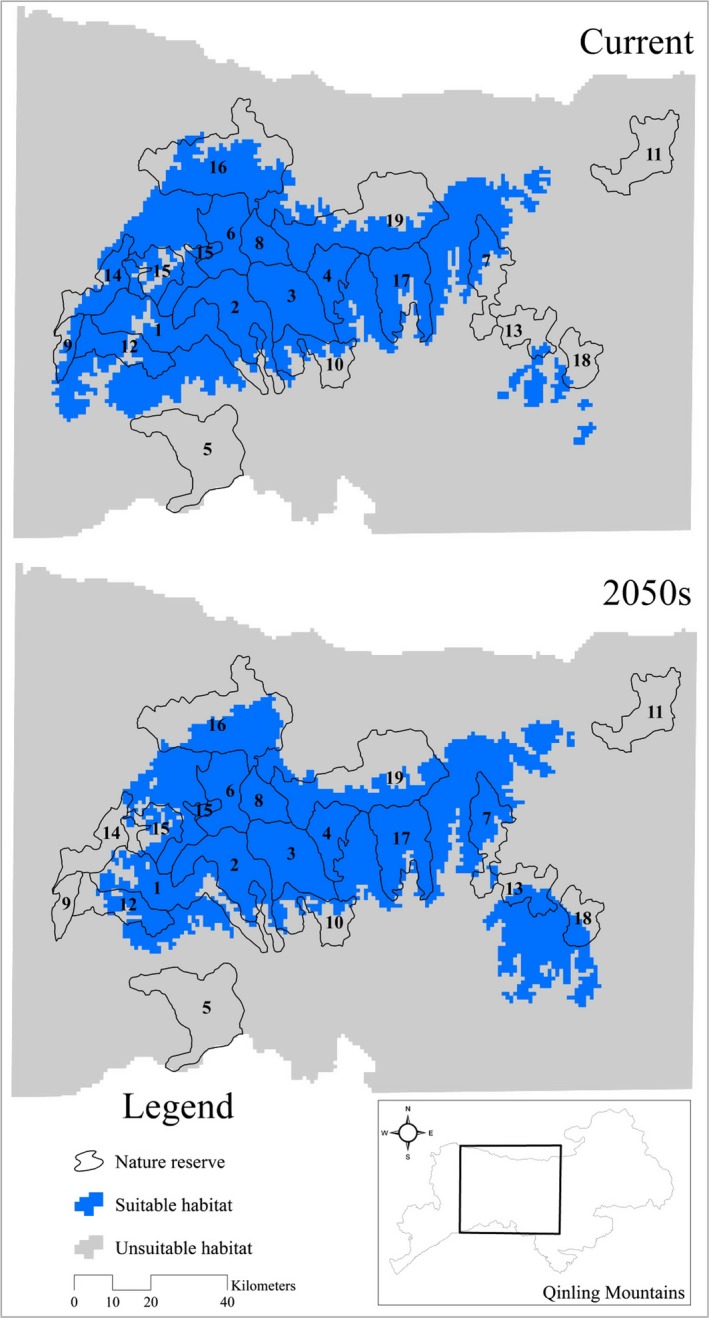
Gap analysis of the giant panda in Qinling Mountains. Codes of the reserves: 1—Banqiao, 2—Changqing, 3—Foping, 4—Guanyinshan, 5—Hangzhongzhuhuan, 6—Huangbaiyuan, 7—Huangguanshan, 8—Laoxiancheng, 9—Motianling, 10—Niangniangshan, 11—Niubeiliang, 12—Panlong, 13—Pingheliang, 14—Sanyuan, 15—Taibainiuweihe, 16—Taibaishan, 17—Tianhuashan, 18—Yingzuishi, 19—Zhouzhi
3.4. Vulnerability assessment
Our predicted 3,823 km2 of unchanged suitable habitat is mainly distributed in Foping, Ningshan, Taibai, Yangxian, and Zhouzhi counties. We predicted 987 km2 (SHc = 20.52%) of current suitable habitat distributed in Chenggu, Taibai, and Yangxian counties is expected to become vulnerable habitat. Interestingly, our results also revealed that there was an increase in the extent of suitable habitat (706 km2, SHf = 15.89%) in Ningshan country (Figure 5).
Figure 5.
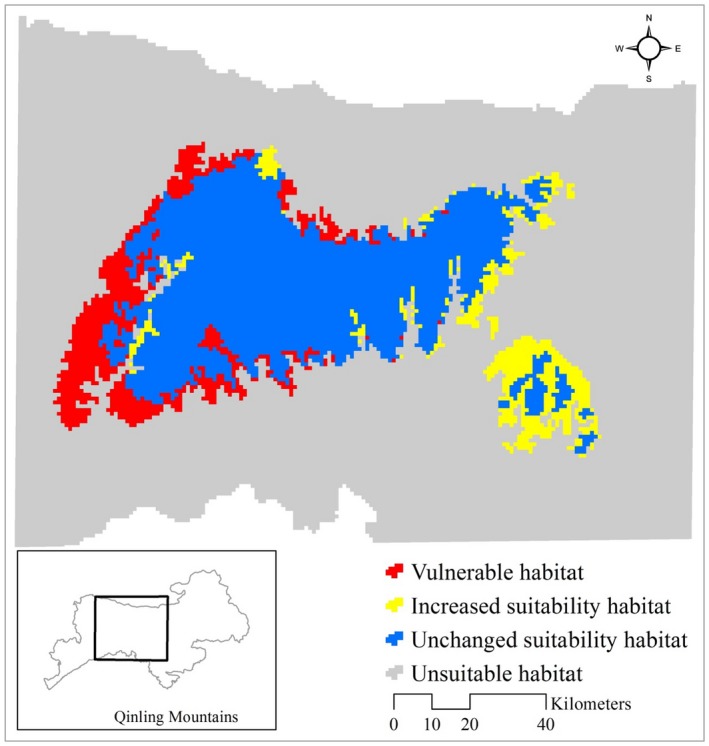
Vulnerability analysis of giant panda suitable habitat in Qinling Mountains
4. Discussion
Over the past several decades, giant pandas have been exposed to several threats to their survival, such as bamboo flowering, extensive poaching, and habitat destruction (Li, Guo, Yang, Wang, & Niemelä, 2003; Pan et al., 2001). However, the Chinese government has conducted giant panda conservation programs, and many of the key threats have been mitigated (Wei et al., 2015; Zhu et al., 2013). At present, human disturbances (e.g., roads, construction projects, ecotourism, and environmental pollutants) and climate change are considered as the paramount threats that degrade and fragment panda habitat (Wei et al., 2015). Particularly, the impacts of climate change on giant panda may have negative impacts on current conservation efforts (Shen, Pimm, Feng, Ren, & Liu, 2015). Therefore, assessing vulnerability is the key step to develop proactive strategies to reduce the impacts of climate change on the giant panda.
Based on the model output, under a mild climate change scenario (RCP 4.5), 20.52% (SHc) of current suitable habitat of giant panda is projected to transfer to unsuitable habitat, particularly in the southwestern region of the Qinling Mountains (i.e., Chenggu and Liuba counties). Climate change associated with suitable habitat fragmentation would present another conservation challenge for this species (Holyoak & Heath, 2016; Li, Clinton, et al., 2015). Current habitat connectivity in southwestern portion of Qinling Mountains is relatively low, and these areas are predicted to experience greatest loss by the 2050s due to climate change, thereby emphasizing the need for a regional conservation strategy for giant panda conservation to protect these areas, and constructing migration corridors to facilitate the dispersal of southwestern populations to large core areas. Fortunately, Ningshan county is predicted to have considerable areas of newly suitable habitat for giant panda. However, migration into the new areas may be impeded by both natural and artificial barriers (e.g., rivers, roads, and human settlements; Fan et al., 2011; Sun et al., 2007). Therefore, proactive measures for habitat restoration should be taken to protect and improve the habitat for the species, and construct migration corridors to facilitate the dispersal of a greater number of giant panda to these new suitable habitats (that currently have a relatively small population of giant panda; Sun et al., 2007).
An assessment of the impact of climate change on species is a critical initial step in implementing the adaptation planning process (Rowland et al., 2011). Some nature reserves, among which planning had been done decades in advance, need to be re‐evaluated considering climate change (Bellard, Bertelsmeier, Leadley, Thuiller, & Courchamp, 2012; Hansen, Hoffman, Drews, & Mielbrecht, 2010). Our results revealed that the loss of giant panda suitable habitat would affect the conservation effectiveness of the existing giant panda reserves. These reserves do not adequately protect the current suitable habitat of giant panda, nor will they protect future potential suitable habitat. Coping strategies to deal with potential threats, particularly in those nature reserves (i.e., Banqiao, Motianling, Panlong, Sanyuan) that would suffer the greatest loss of suitable habitat under future climate change, require further in‐depth study. Meanwhile, three provincial nature reserves located in Ningshan county are also urgent need to improve their conservation effectiveness against climate change, because they currently support a small population of giant panda (Sun et al., 2007), but are isolated from the network of large reserves, and have low levels of protection (Figure 4).
Vulnerability assessments can provide information about the locations that are vulnerable to climate change (Levinsky et al., 2007) and broadscale guidance to direct conservation efforts (Dubois, Caldas, Boshoven, & Delach, 2011; Rowland et al., 2011). Based on our vulnerability assessment, protection needs to prioritize habitat in which the maximum effects of climate change are predicted to occur, namely the vulnerable areas. These regions are predicted to suffer from large range contractions under climate change and present the greatest risk to the persistence of giant panda in the 2050s. Furthermore, vulnerability assessments are able to identify the potential climatic refuges for giant panda within Qinling Mountains range, namely unchanged and new increased suitability habitat (Ashcroft, 2010; Li et al., 2016), and these areas may facilitate species persistence during periods of climatic stress.
4.1. Conservation implications
As a flagship species in China, the government of China has listed the giant panda in the key program of biodiversity conservation (Ministry of Environmental Protection et al., 2011), and will conduct giant panda conservation programs and establish national parks specifically for protecting the species in Shaanxi, Sichuan, and Gansu provinces (State Forestry Administration, 2016b). Thus, assessment of vulnerability provided key information in designing effective adaptation strategies to cope with the impacts of future climate change for national parks development. Our results suggest the following adaptation strategies to ameliorate the predicted impacts of climate change on giant panda in Qinling Mountains:
4.1.1. Establishing new reserves
Gap analysis showed the distribution of current suitable habitat in Foping, Ningshan, and Taibai counties is largely unprotected, leaving significant gaps in the conservation network, and suitable habitat distributed in these areas will be discrete and fragmentated by the 2050s (Figure 6). Therefore, new reserves need to be established in these regions to improve the connectivity of habitat.
Figure 6.
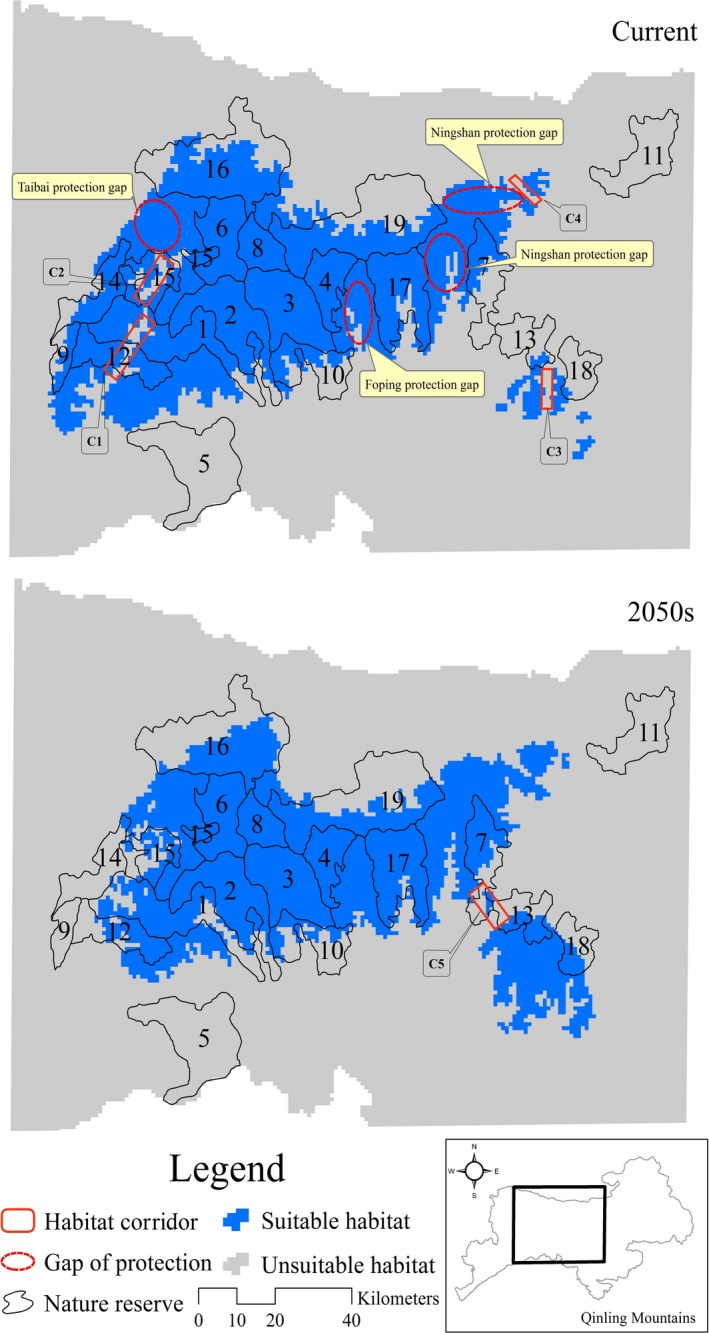
Protection gaps and habitat corridors giant pandas in Qinling Mountains. C1–C5 indicates habitat corridors
4.1.2. Adjusting reserves
An adjustment of range to the existing nature reserves also might be necessary, where habitat shift is observed within the reserves and in their vicinities. For example, it might be necessary to enlarge the protected area of Huangguanshan to connect with Tianhuashan nature reserve. Similarly, increase in area in Pingheliang and Yingzuishi nature reserves may be needed, to protect their surrounding suitable habitat of giant panda.
4.1.3. Establishing habitat corridors
Establishing migration corridors in juncture of Chenggu, Taibai and Yangxian counties (C1 and C2), and Ningshan county (C5; Figure 6) to increase chances for the small population of these areas to larger suitable areas, and enable giant panda to escapes from unsuitable climatic conditions. We also need to establish habitat corridors in Ningshan county (C3 and C4) to enhance habitat connectivity in these areas.
4.1.4. Improving adaptive capacity to climate change
Reducing nonclimate stressors (such as invasive species, human activities, pollution, disease, and other stressors) will improve the impact on the ability of specie to adapt to climate change (Gross, Watson, Woodley, Welling, & Harmon, 2015). Such as invasive species, anticipatory actions might focus on identifying invasive species likely to expand their ranges in response to climate change, and establishing early‐detection and rapid response protocols designed to keep them from invading sensitive areas.
4.1.5. Strengthening monitoring on giant panda
Many nature reserves just started to consider strategies to adapt to climate change when they made their master plans. We do not fully understand how giant panda will respond to those strategies and what management measures might be effective. Therefore, a standardized monitoring program is necessary for nature reserves to collect information of climate change impacts on panda and monitor the responses of the species to the strategies.
Conflict of Interest
None declared.
Acknowledgments
This research was supported by National Science & Technology Pillar Program during the Twelfth Five‐year Plan Period (No. 2013BAC09B02). We thank Dr. Minghao Gong for helping us collect occurrence data of the giant panda. We thank Dr. Yuguang Zhang and Li Yang for valuable suggestions for the manuscript.
Appendix 1. The code and description of environmental variables, the min, max, mean, and SD values are calculated over the entire Qinling Mountains geographic extent
1.1.
| Variable | Description | Unit | Type | Min | Max | Mean | SD |
|---|---|---|---|---|---|---|---|
| Bio1 | Annual mean temperature | °C | Continuous | −1.3 | 16.9 | 11.1 | 2.5 |
| Bio2 | Mean diurnal range | °C | Continuous | 6.5 | 11.4 | 8.9 | 0.8 |
| Bio3 | Temperature constancy | Continuous | 21.0 | 31.0 | 26.8 | 1.4 | |
| Bio4 | Temperature seasonality | Continuous | 6,904.0 | 9,944.0 | 8,272.1 | 413.7 | |
| Bio5 | Max temperature of warmest month | °C | Continuous | 12.3 | 33.7 | 27.2 | 2.9 |
| Bio6 | Min temperature of coldest month | °C | Continuous | −15.6 | 1.1 | −5.4 | 2.2 |
| Bio7 | Annual temperature range | °C | Continuous | 27.9 | 39.1 | 32.6 | 1.7 |
| Bio8 | Mean temperature of wettest quarter | °C | Continuous | 6.9 | 26.3 | 20.4 | 2.7 |
| Bio9 | Mean temperature of driest quarter | °C | Continuous | −10.6 | 6.5 | 0.0 | 2.3 |
| Bio10 | Mean temperature of warmest quarter | °C | Continuous | 7.8 | 27.5 | 21.6 | 2.7 |
| Bio11 | Mean temperature of coldest quarter | °C | Continuous | −10.6 | 6.5 | 0.0 | 2.3 |
| Bio12 | Annual precipitation | mm | Continuous | 549.0 | 1,119.0 | 818.4 | 73.2 |
| Bio13 | Precipitation of wettest month | mm | Continuous | 97.0 | 197.0 | 149.4 | 13.0 |
| Bio14 | Precipitation of driest month | mm | Continuous | 4.0 | 13.0 | 7.4 | 1.8 |
| Bio15 | Precipitation seasonality | Continuous | 64.0 | 96.0 | 72.1 | 4.6 | |
| Bio16 | Precipitation of wettest quarter | mm | Continuous | 280.0 | 559.0 | 417.0 | 41.1 |
| Bio17 | Precipitation of driest quarter | mm | Continuous | 14.0 | 46.0 | 27.6 | 6.1 |
| Bio18 | Precipitation of warmest quarter | mm | Continuous | 247.0 | 492.0 | 369.3 | 33.0 |
| Bio19 | Precipitation of coldest quarter | mm | Continuous | 14.0 | 46.0 | 27.6 | 6.1 |
| Alt | Elevation above sea level | m | Continuous | 189.0 | 3,637.0 | 1,225.1 | 479.8 |
| Rddes | Density of roads | km/km2 | Continuous | 0.1 | 0.7 | 0.3 | 0.1 |
| Rvdes | Density of rivers | m/km2 | Continuous | 0.1 | 0.3 | 0.2 | 0.0 |
| Setdes | Density of settlements | #/km2 | Continuous | 0.3 | 1.4 | 0.7 | 0.1 |
Appendix 2. Analysis of correlation coefficient of the environmental variables
2.1.
| Variables | Bio1 | Bio2 | Bio3 | Bio4 | Bio5 | Bio6 | Bio7 | Bio8 | Bio9 | Bio10 | Bio11 | Bio12 | Bio13 | Bio14 | Bio15 | Bio16 | Bio17 | Bio18 | Bio19 | Alt | Rddes | Rvdes | Setdes |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Bio1 | 1.000 | ||||||||||||||||||||||
| Bio2 | .717 | 1.000 | |||||||||||||||||||||
| Bio3 | .719 | .839 | 1.000 | ||||||||||||||||||||
| Bio4 | .365 | .660 | .182 | 1.000 | |||||||||||||||||||
| Bio5 | .981 | .808 | .723 | .524 | 1.000 | ||||||||||||||||||
| Bio6 | .912 | .416 | .601 | −.022 | .823 | 1.000 | |||||||||||||||||
| Bio7 | .513 | .863 | .471 | .943 | .663 | .120 | 1.000 | ||||||||||||||||
| Bio8 | .989 | .770 | .699 | .494 | .996 | .848 | .622 | 1.000 | |||||||||||||||
| Bio9 | .975 | .599 | .714 | .150 | .917 | .976 | .316 | .933 | 1.000 | ||||||||||||||
| Bio10 | .982 | .784 | .689 | .534 | .997 | .824 | .656 | .999 | .916 | 1.000 | |||||||||||||
| Bio11 | .975 | .599 | .714 | .150 | .917 | .976 | .316 | .933 | 1.000 | .916 | 1.000 | ||||||||||||
| Bio12 | −.447 | −.652 | −.380 | −.663 | −.558 | −.172 | −.748 | −.499 | −.300 | −.529 | −.300 | 1.000 | |||||||||||
| Bio13 | −.413 | −.565 | −.298 | −.618 | −.523 | −.177 | −.680 | −.471 | −.283 | −.495 | −.283 | .875 | 1.000 | ||||||||||
| Bio14 | −.339 | .114 | −.110 | .338 | −.261 | −.559 | .281 | −.251 | −.437 | −.234 | −.437 | .168 | .179 | 1.000 | |||||||||
| Bio15 | .016 | −.352 | −.023 | −.626 | −.094 | .298 | −.556 | −.103 | .154 | −.123 | .154 | .021 | .182 | −.803 | 1.000 | ||||||||
| Bio16 | −.450 | −.762 | −.405 | −.837 | −.587 | −.093 | −.903 | −.540 | −.268 | −.573 | −.268 | .908 | .871 | −.165 | .424 | 1.000 | |||||||
| Bio17 | −.288 | .115 | −.103 | .343 | −.215 | −.493 | .274 | −.198 | −.379 | −.183 | −.379 | .258 | .214 | .973 | −.862 | −.112 | 1.000 | ||||||
| Bio18 | −.397 | −.661 | −.256 | −.880 | −.547 | −.059 | −.879 | −.501 | −.209 | −.537 | −.209 | .844 | .895 | −.069 | .452 | .940 | −.057 | 1.000 | |||||
| Bio19 | −.288 | .115 | −.103 | .343 | −.215 | −.493 | .274 | −.198 | −.379 | −.183 | −.379 | .258 | .214 | .973 | −.862 | −.112 | 1.000 | −.057 | 1.000 | ||||
| Alt | −.940 | −.775 | −.681 | −.529 | −.957 | −.780 | −.644 | −.962 | −.875 | −.962 | −.875 | .469 | .438 | .159 | .184 | .542 | .104 | .508 | .104 | 1.000 | |||
| Rddes | .139 | .075 | −.087 | .285 | .185 | .074 | .226 | .164 | .075 | .178 | .075 | −.397 | −.424 | −.310 | .119 | −.296 | −.326 | −.378 | −.326 | −.153 | 1.000 | ||
| Rvdes | .351 | .606 | .332 | .678 | .442 | .057 | .697 | .438 | .211 | .457 | .211 | −.331 | −.237 | .460 | −.577 | −.547 | .470 | −.470 | .470 | −.486 | .200 | 1.000 | |
| Setdes | .370 | .230 | .163 | .236 | .369 | .311 | .235 | .377 | .333 | .379 | .333 | −.310 | −.281 | −.152 | .002 | −.295 | −.168 | −.272 | −.168 | −.378 | .489 | .206 | 1.000 |
Li J, Liu F, Xue Y, Zhang Y, Li D. Assessing vulnerability of giant pandas to climate change in the Qinling Mountains of China. Ecol Evol. 2017;7:4003–4015. https://doi.org/10.1002/ece3.2981
References
- Ancillotto, L. , Santini, L. , Ranc, N. , Maiorano, L. , & Russo, D. (2016). Extraordinary range expansion in a common bat: The potential roles of climate change and urbanization. Science Nature, 103, 15. [DOI] [PubMed] [Google Scholar]
- Ashcroft, M. B. (2010). Identifying refugia from climate change. Journal of Biogeography, 37, 1047–1413. [Google Scholar]
- Auer, S. K. , & Martin, T. E. (2013). Climate change has indirect effects on resource use and overlap among coexisting bird species with negative consequences for their reproductive success. Global Change Biology, 19, 411–419. [DOI] [PubMed] [Google Scholar]
- Baek, H. J. , Lee, J. , Lee, H. S. , Hyun, Y. K. , Cho, C. H. , Kwon, W. T. , … Byun, Y. H. (2013). Climate change in the 21st century simulated by HadGME2‐AO under representative conservation pathways. Asia‐Pacific Journal of Atmospheric Sciences, 49, 603–618. [Google Scholar]
- Bagne, K. E. , Friggens, M. M. , & Finch, D. M. (2011). A System for Assessing Vulnerability of Species (SAVS) to Climate Change. Gen. Tech. Rep. RMRS‐GTR‐257. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. [Google Scholar]
- Bellard, C. , Bertelsmeier, C. , Leadley, P. , Thuiller, W. , & Courchamp, F. (2012). Impact of climate change on the future of biodiversity. Ecology Letters, 15, 365–377. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charmantier, A. , & Gienapp, P. (2014). Climate change and timing of avian breeding and migration: Evolutionary versus plastic change. Evolutionary Applications, 7, 15–28. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cord, A. F. , Klein, D. , Mora, F. , & Dech, S. (2014). Comparing the suitability of classified land cover and remote sensing variables for modeling distribution patterns of plants. Ecological Modelling, 272, 129–140. [Google Scholar]
- Cramer, W. , Yohe, G. W. , Auffhammer, M. , Huggel, U. , Molau, U. , da Silva Dias, M. A. F. , … Tibig, L. (2014). Detection and attribution of observed impacts In Field C. B., Barros V. R., Dokken D.J., Mach K. J., Mastrandrea M. D., Bilir T. E., … White L. L. (Eds.), Climate change 2014: Impacts, adaptation, and vulnerability. Part A: Global and Sectoral Aspect Contribution of Working Group II to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (pp. 979–1037). Cambridge, UK and New York, NY: Cambridge University Press. [Google Scholar]
- Duan, R. , Kong, X. , Huang, M. , Vaerla, S. , & Ji, X. (2016). The potential effects of climate change on amphibian distribution, range fragmentation and turnover in China. PeerJ, 4, e2185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dubois, N. , Caldas, A. , Boshoven, J. , & Delach, A. (2011). Integrating climate change vulnerability assessments into adaptation planning: a case study using the NatureServe Climate Change Vulnerability Index to inform conservation planning for species in Florida [final report]. Defender of Wildlife, Washington, DC. [Google Scholar]
- Elith, J. , Phillips, S. , Hastie, T. , Dudík, M. , & Chee, Y. E. (2011). A statistical explanation of MaxEnt for ecologists. Diversity and Distribution, 17, 43–57. [Google Scholar]
- Fan, J. T. , Li, J. S. , Quan, Z. J. , Wu, X. P. , Hu, L. L. , & Yang, Q. P. (2011). Impact of road construction on giant panda's habitat and its carrying capacity in Qinling Mountains. Acta Ecologica Sinica, 31, 145–149. [Google Scholar]
- Feeley, K. , & Silman, M. R. (2016). Disappearing climates will limit the efficacy of Amazonian protected area. Diversity and Distributions, 22, 1081–1084. [Google Scholar]
- Foden, W. B. , Butchart, S. H. M. , Stuart, S. N. , Vié, J.‐C. , Akçakaya, H. R. , Angulo, A. , … Mace, G. M. (2013). Identifying the world's most climate change vulnerable species: A systematic trait‐based assessment of all birds, amphibians and corals. PLoS One, 8, e65427. doi:10.1371/journal.pone.0056427 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glick, P. , Stein, B. A. , & Edelson, N. A. (2011). Scanning the conservation horizon: A guide to climate change vulnerability assessment. Washington, DC: National Wildlife Federation. [Google Scholar]
- Gross, J. , Watson, J. , Woodley, S. , Welling, L. , & Harmon, D. (2015). Responding to climate change: Guidance for protected area managers and planners. Switzerland: IUCN. [Google Scholar]
- Guisan, A. , Tingley, R. , Baumgartner, J. B. , Naujokaitis‐Lewis, I. , & Sucliffe, P. R. (2013). Predicting species distributions for conservation decisions. Ecology Letters, 16, 1424–1435. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hansen, L. , Hoffman, J. , Drews, C. , & Mielbrecht, E. (2010). Designing climate‐smart conservation: Guidance and case studies. Conservation Biology, 24, 63–69. [DOI] [PubMed] [Google Scholar]
- Heikkinen, R. K. , Luoto, M. , Leikola, N. , Pöyry, J. , & Settele, J. (2010). Assessing the vulnerability of European butterflies to climate change using multiple criteria. Biodiversity and Conservation, 19, 695–723. [Google Scholar]
- Hijmans, R. J. , Cameron, S. E. , Parra, J. L. , Jones, P. G. , & Jarvis, A. (2005). Very high resolution interpolated climate surfaces for global land areas. International Journal of Climatology, 25, 1965–1978. [Google Scholar]
- Holyoak, M. , & Heath, S. K. (2016). The integration of climate change, spatial dynamics, and habitat fragmentation: A conceptual overview. Integrative Zoology, 11, 40–59. [DOI] [PubMed] [Google Scholar]
- IPCC (Intergovernmental Panel on Climate Change) (2014). Climate Change 2014: Synthesis Report. Contribution of Working Group I, II, and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change. Geneva, Switzerland: Author. [Google Scholar]
- IUCN (International Union for Conservation of Nature and Natural Resources) (1996). 1996 IUCN Red List of Threatened Animals. Gland, Switzerland and Cambridge, UK: Author. [Google Scholar]
- Kane, A. , Burkett, T. C. , Kloper, S. , & Sewall, J. (2013). Virginia's climate modeling and species vulnerability assessment: How climate data can inform management and conservation. Reston, VA: National Wildlife Federation. [Google Scholar]
- Kearney, M. , & Porter, W. (2009). Mechanistic niche modeling: Combining physiological and spatial data to predict species’ ranges. Ecology Letters, 12, 334–350. [DOI] [PubMed] [Google Scholar]
- Khatchikian, C. , Sangermano, F. , Kendell, D. , & Livdahl, T. (2011). Evaluation of species distribution model algorithms for fine‐scale container‐breeding mosquito risk prediction. Medical and Veterinary Entomology, 25, 268–275. doi:10.1111/j1365‐2915.2010.00935.x [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koen, E. L. , Bowman, J. , Murray, D. L. , & Wilson, P. J. (2014). Climate change reduces genetic diversity of Canada Lynx at trailing range edge. Ecography, 37, 375–762. [Google Scholar]
- Lawler, J. J. , Shafer, S. L. , & Bancroft, B. A. (2010). Projected climate impacts for the amphibians of the Western Hemisphere. Conservation Biology, 24, 38–50. [DOI] [PubMed] [Google Scholar]
- Lehikoinen, A. , Foppen, R. P. B. , Heldbjer, H. , Lindström, Ă. , van Manen, W. , Piirainen, S. , … Butchart, S. H. M. (2016). Large‐scale climatic drivers of regional winter bird population trends. Diversity and Distributions, 22, 1163–1173. [Google Scholar]
- Lei, J. C. , Xu, H. G. , Cui, P. , Guang, Q. W. , & Ding, H. (2014). The potential effects of climate change on suitable habitat for the Sichuan hill partridge (Arborophila rufipectus, Boulton): Based on the maximum entropy modeling. Polish Journal of Ecology, 62, 771–787. [Google Scholar]
- Lemke, D. , Hulme, P. E. , Brown, J. A. , & Tadesse, W. (2011). Distribution modeling of Japanese honeysuckle (Lonicera japonica) invasion in the Cumberland Plateau and Mountain Region, USA. Forest Ecology and Management, 262, 139–149. [Google Scholar]
- Levinsky, I. , Skov, F. , Svenning, J. C. , & Rahbek, C. (2007). Potential impacts of climate change on the distribution and diversity patterns of European mammals. Biodiversity and Conservation, 16, 3803–3816. [Google Scholar]
- Li, X. Y. , Clinton, N. , Si, Y. L. , Liao, J. S. , Liang, L. , & Gong, P. (2015). Projected impacts of climate change on protected birds and nature reserves in China. Chinese Science Bulletin, 60, 1644–1653. [Google Scholar]
- Li, Y. , Guo, Z. , Yang, Q. , Wang, Y. , & Niemelä, J. (2003). The implications of poaching for giant panda conservation. Biological Conservation, 111, 125–136. [Google Scholar]
- Li, J. , McCarthy, T. M. , Wang, H. , Weckworth, B. V. , Schaller, G. B. , Mishra, C. , … Beissinger, S. R. (2016). Climate refugia of snow leopards in High Asia. Biological Conservation, 203, 188–196. [Google Scholar]
- Li, Y. , Viña, A. , Yang, W. , Chen, X. D. , Zhang, J. D. , Ouyang, Z. Y. , … Liu, J. G. (2013). Effects of conservation policies on forest cover change in giant panda habitat regions, China. Land Use Policy, 33, 42–53. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li, R. Q. , Xu, M. , Wong, M. H. G. , Qiu, S. , & Li, X. H. (2015). Climate change threatens giant panda protection in the 21st century. Biological Conservation, 182, 93–101. [Google Scholar]
- Liu, C. , Berry, P. M. , Dawson, T. P. , & Pearson, R. G. (2005). Selecting thresholds of occurrence in the prediction of species distribution. Ecography, 28, 385–393. [Google Scholar]
- Liu, G. , Guang, T. P. , Dai, Q. , Li, H. X. , & Gong, M. H. (2016). Impacts of temperature on giant panda habitat in the north Minshan Mountains. Ecology and Evolution, 6, 987–996. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Loucks, C. , Zhi, L. , Dinerstein, E. , Dajun, W. , Dali, F. , & Hao, W. (2003). The giant pandas of the Qinling Mountains, China: A case study in designing conservation landscapes for elevational migrants. Conservation Biology, 17, 558–565. [Google Scholar]
- Lučan, R. K. , Weiser, M. , & Hanák, V. (2013). Contrasting effects of climate change on the timing of reproduction and reproductive success of a temperate insectivorous bat. Journal of Zoology, 290, 151–159. [Google Scholar]
- Malcolm, J. R. , Liu, C. , Neilson, R. P. , Hansen, L. , & Hannah, L. (2006). Global warming and extinctions of endemic species from biodiversity hotspots. Conservation Biology, 20, 538–548. [DOI] [PubMed] [Google Scholar]
- Ministry of Environmental Protection , The Central Propaganda Department , Ministry of Forestry Affairs , National Development and Reform Commission , Ministry of Education , Ministry of Science and Technology , … Guangming Daily Press (2011). Ministries of joint‐drafting china national biodiversity conservation strategy and action plan (73 pp.). Beijing, China: China Environmental Science Press. [Google Scholar]
- Molina‐Martínez, A. , León‐Cortés, J. L. , Regan, H. M. , Lewis, O. T. , Navarrete, D. , & Caballero, U. (2016). Changes in butterfly distributions and species assemblages on a Neotropical mountain range in response to global warming and anthropogenic land use. Diversity and Distributions, 22, 1085–1098. [Google Scholar]
- Monahan, W. B. (2009). A mechanistic niche model for measuring species’ distributional response temperature gradients. PLoS One, 4, e7921. doi:10.1371/journal.pone.0007921 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pacifici, M. , Foden, W. B. , Visconti, P. , Watson, J. E. M. , Butchart, S. H. M. , Kovacs, K. M. , … Rondinini, C. (2015). Assessing species vulnerability to climate change. Nature Climate Change, 5, 215–224. [Google Scholar]
- Pan, W. S. , Lu, Z. , Zhu, X. J. , Wang, D. J. , Wang, H. , Long, Y. , … Zhou, X. (2001). A chance for lasting survival. Beijing, China: Peking University Press. [Google Scholar]
- Pearson, R. G. , Stanton, J. C. , Shoemaker, K. T. , Aiello‐Lammens, M. E. , Ersts, P. J. , Horning, N. , … Akçakaya, H. R. (2014). Life history and spatial traits predict extinction risk due to climate change. Nature Climate Change, 4, 217–221. [Google Scholar]
- Pereira, H. M. , Leadley, P. W. , Proenca, V. , Alkemade, R. , Scharlemann, J. P. W. , Fernandez‐Manjarrés, J. F. , … Walpole, M. (2010). Scenarios for global biodiversity in the 21st century. Science, 330, 1496–1501. [DOI] [PubMed] [Google Scholar]
- Phillips, S. J. , Anderson, R. P. , & Schapire, R. E. (2006). Maximum entropy modeling of species geographic distributions. Ecological Modelling, 190, 231–259. [Google Scholar]
- Phillips, S. J. , Dudík, M. , Elith, J. , Graham, G. H. , Lehmann, A. L. , Leathwick, J. , & Ferrier, S. (2009). Sample selection bias and presence‐only distribution models: Implications for background and pseudo‐absence data. Ecological Applications, 19, 181–197. [DOI] [PubMed] [Google Scholar]
- Polaina, E. , Revilla, E. , & González‐Suárez, M. (2016). Putting susceptibility on map to improve conservation planning, an example with terrestrial mammals. Diversity and Distributions, 22, 881–892. [Google Scholar]
- Rinawati, F. , Stein, K. , & Lindner, A. (2013). Climate change impacts on biodiversity—The setting of a lingering global crisis. Diversity, 5, 114–123. [Google Scholar]
- Rowland, E. L. , Davison, J. E. , & Graumlich, L. J. (2011). Approaches to evaluating climate change impacts on species: A guide to initiating the adaptation plan process. Environmental Management, 47, 322–337. [DOI] [PubMed] [Google Scholar]
- Scott, J. M. , Davis, F. , Csuti, B. , Noss, R. , Butterfield, B. , Groves, C. , … Wright, R. G. (1993). Gap analysis: A geographic approach to protection of biological diversity. Wildlife Monographs, 123, 3–41. [Google Scholar]
- Shen, G. Z. , Pimm, S. L. , Feng, C. Y. , Ren, G. F. , & Liu, Y. P. (2015). Climate change challenges the current conservation strategy for the giant panda. Biological Conservation, 190, 43–50. [Google Scholar]
- Songer, M. , Delion, M. , Biggs, A. , & Huang, Q. (2012). Modeling impacts of climate change on giant panda habitat. International Journal of Ecology, 22, 48. [Google Scholar]
- State Forestry Administration (2006). The 3rd National Survey Report on Giant Panda in China. Beijing, China: Science Press. [Google Scholar]
- State Forestry Administration (2015). Release of the Fourth National Survey Report on Giant Panda in China. Author, Beijing, China: Retrieved from http://www.forestry.gov.cn/main/69/content-743562.html [Google Scholar]
- State Forestry Administration (2016a). Qinling protection should be open national park model. Retrieved from http://www.forestry.gov.cn/main/72/content-912580.html
- State Forestry Administration (2016b). Three provinces to build joint national park to protect pandas. Retrieved from http://english.forestry.gov.cn/index.php/giant-panda/1249-three-provinces-to-build-joint-national-park-to-protect-pandas
- Sun, C. Q. , Jin, X. L. , Yuan, W. , Wang, W. Y. , Cao, Y. H. , & Lu, X. R. (2007). The 3rd Comprehensive Survey Report on Giant Panda in Shaanxi Province. Xi'an, China: Xi'an Map Press. [Google Scholar]
- Swaisgood, R. , Wang, D. , & Wei, F. (2016). Ailuropoda melanoleuca. The IUCN Red list of Threatened Species 2016: e.T712A45033386. https://doi.org/10.2305/IUCN.UK.2016-2.RLTS.T712A45033386.en
- Tateshi, R. , Thanh, H. N. , Kobayashi, T. , Alsaaideh, B. , Tana, G. , & Xuan, P. D. (2014). Production of land cover data‐GLCNMO2008. Journal of Geography and Geology, 6, 99–122. [Google Scholar]
- Thomas, C. D. , Cameron, A. , Green, R. E. , Bakkenes, M. , Beaumont, L. J. , Collingham, Y. C. , … Williams, S. E. (2004). Extinction risk from climate change. Nature, 427, 145–148. [DOI] [PubMed] [Google Scholar]
- Thuiller, W. , Lavorel, S. , & Araújo, M. B. (2005). Niche properties and geographical extent as predictors of species sensitivity to climate change. Global Ecology and Biogeography, 14, 347–357. [Google Scholar]
- Thuiller, W. , Lavorel, S. , Araújo, M. B. , Sykes, M. T. , & Prentice, I. C. (2005). Climate change threats to plant diversity in Europe. Proceedings of the National Academy Sciences of the United States of America, 102, 8245–8250. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tuberville, T. D. , Andrews, K. M. , Sperry, J. H. , & Grosse, A. M. (2015). Use of the NatureServe climate change vulnerability index as an assessment tool for reptiles and amphibians: Lessons learned. Environmental Management, 56, 822–834. [DOI] [PubMed] [Google Scholar]
- UNFCCC (2015). Adoption of the Paris agreement. Report No. FCCC/CP/2015/L.9/Rev.1. Retrieved from http://unfccc.int/resource/docs/2015/cop21/eng/l09r01.pdf
- Urban, M. C. (2015). Accelerating extinction risk from climate change. Science, 348, 571–573. [DOI] [PubMed] [Google Scholar]
- Wang, T. J. , Ye, X. P. , Skidmore, A. K. , & Toxopeus, A. G. (2010). Characterizing the spatial distribution of giant pandas (Ailuropoda melanoleuca) in fragmented forest landscapes. Journal of Biogeography, 37, 865–878. [Google Scholar]
- Wei, F. W. , Swaisgood, R. , Hu, Y. B. , Nie, Y. G. , Li, Y. , Zhang, Z. J. , … Zhu, L. F. (2015). Progress in the ecology and conservation of giant pandas. Conservation Biology, 29, 1497–1507. [DOI] [PubMed] [Google Scholar]
- Williams, S. E. , Shoo, L. P. , Isaac, J. L. , Hoffmann, A. A. , & Langham, G. (2008). Towards an integrated framework for assessing the vulnerability of species to climate change. PloS Biology, 6, 2621–2626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wisz, M. S. , Hijmans, R. J. , Li, J. , Peterson, A. T. , Graham, C. H. , Guisan, A. , & NECAS Predicting Species Distributions Working Group (2008). Effects of sample size on the performance of species distribution models. Diversity and Distributions, 14, 763–773. [Google Scholar]
- Yang, L. H. , & Rudolf, V. H. W. (2010). Phenology, ontogeny and the effects of climate change on the timing of species interactions. Ecology Letters, 13, 1–10. [DOI] [PubMed] [Google Scholar]
- Young, B. E. , Byers, E. , Hammerson, G. , Frances, A. , Oliver, L. , & Treher, A. (2015). Guidelines for Using the NatureServe Climate Change Vulnerability Index (Release 3.0, April 2015). Arlington, VA: NatureServe. [Google Scholar]
- Young, B. E. , Hall, K. R. , Byers, E. , Gravuer, K. , Hammerson, G. , Redder, A. , & Szabo, K. (2012). Rapid assessment of plant and animal vulnerability to climate change In Brodie J. F., Post E. & Doak D. (Eds.), Wildlife conservation in a changing climate (pp. 129–150). Chicago, IL: University of Chicago Press. [Google Scholar]
- Zhao, N. X. , Zhang, X. M. , & Dong, W. (2014). Field guide to nature history and wildlife of Qinling Mountains. Beijing, China: Encyclopedia of China Publishing House. [Google Scholar]
- Zhu, L. F. , Hu, Y. B. , Qi, D. W. , Wu, H. , Zhan, X. J. , Zhang, Z. J. , … Wei, F. W. (2013). Genetic consequences of historical anthropogenic and ecological events on giant panda. Ecology, 94, 2346–2357. [DOI] [PubMed] [Google Scholar]