

Abstract
NZ17074 (N1), an arenicin-3 derivative isolated from the lugworm, has potent antibacterial activity and is cytotoxic. To reduce its cytotoxicity, seven N1 analogues with different structures were designed by changing their disulfide bonds, hydrophobicity, or charge. The “rocket” analogue-N2 and the “kite” analogue-N6 have potent activity and showed lower cytotoxicity in RAW264.7 cells than N1. The NMR spectra revealed that N1, N2, and N6 adopt β-sheet structures stabilized by one or two disulfide bonds. N2 and N6 permeabilized the outer/inner membranes of E. coli, but did not permeabilize the inner membranes of S. enteritidis. N2 and N6 induced E. coli and S. enteritidis cell cycle arrest in the I-phase and R-phase, respectively. In E. coli and in S. enteritidis, 18.7–43.8% of DNA/RNA/cell wall synthesis and 5.7–61.8% of DNA/RNA/protein synthesis were inhibited by the two peptides, respectively. Collapsed and filamentous E. coli cells and intact morphologies of S. enteritidis cells were observed after treatment with the two peptides. Body weight doses from 2.5–7.5 mg/kg of N2 and N6 enhanced the survival rate of peritonitis- and endotoxemia-induced mice; reduced the serum IL-6, IL-1β and TNF-α levels; and protected mice from lipopolysaccharide-induced lung injury. These data indicate that N2 and N6, through multiple selective actions, may be promising dual-function candidates as novel antimicrobial and anti-endotoxin peptides.
Introduction
Escherichia coli and Salmonella enteritidis are major food-borne pathogens that often cause outbreaks of diarrheal diseases and extraintestinal infections in both animals and humans throughout the world1. The World Health Organization (WHO) has estimated that 1.7–2.5 million human deaths occur worldwide every year2. Antibiotics such as β-lactams, fluoroquinolones, aminoglycosides, and trimethoprim-sulfamethoxazole are the primary antimicrobials used to treat these infectious diseases2–4. However, resistance of E. coli and S. enteritidis to these antibiotics is definitively on the rise3, 5. Treatment with these antibiotic therapies also causes a three- to 20-fold increase in the total concentration of lipopolysaccharide (LPS) rapidly released from cell walls of gram-negative bacteria, and subsequently induces a variety of acute and chronic diseases including sepsis, fatal septic shock, ulcerative colitis, and systemic inflammatory response syndrome6–8. The annual incidence of sepsis has been roughly estimated at 15–19 million cases globally, and the mortality of severe sepsis reaches 30–50% worldwide9, 10. Although monoclonal antibodies or other antiendotoxin chemical drugs have been examined in a few clinical studies to combat LPS-induced diseases, these therapies have been largely unsuccessful due to either high host toxicity or limited effects11, 12. The absence of efficient therapies makes studies seeking novel LPS-neutralizing agents of the utmost importance.
The removal of gram-negative bacteria by antimicrobial peptides (AMPs) may be an effective strategy to control and prevent worsening antibiotic resistance and LPS-induced pathophysiological responses8, 13. It has been demonstrated that several α-helical AMPs, such as cathelicidin (LL-37) and KSL-W, can effectively neutralize Porphyromonas gingivalis and E. coli LPS and inhibit LPS-induced inflammatory cytokine production in vitro 13, 14. Bedran et al.15 reported that LL-37 and β-sheet peptide hBD-3 displayed anti-inflammatory activities when applied individually and synergistically to a three-dimensional co-culture model of gingival epithelial cells and fibroblasts15. Due to their small sizes (<30 amino acid residues) and excellent binding ability to the lipid bilayer of bacterial membranes, β-sheet AMPs such as s-thanatin are also well-established as promising candidates for the treatment of multidrug-resistant bacterial infections and neutralization of LPS16, 17.
Recent studies have mainly focused on developing β-sheet AMPs that have fewer or no Cys residues, as they are easier and less expensive to produce18. It has been reported that there are three main categories of β-sheet AMPs that have: (i) antimicrobial activity that is independent of the number of disulfide bonds, for example human β-defensins 1–3, cryptdin-4, μ-conotoxins, and thanatin19–23; (ii) have enhanced antimicrobial activity in the presence of fewer disulfide bonds such as HNP-1 and cryptdin-4 variants19, 24; and (iii) have decreased antimicrobial activity in the presence of fewer or no disulfide bonds, such as PM1-S, human β-defensin 3, human α-defensin HNP-1, and arenicin-125, 26. This difference is most likely related to the position of cationic residues in the disulfide loop of AMPs. For example, the location of “K” or “R” residues in disulfide bonds could be an important determining factor for their antimicrobial activity20. On the other hand, intramolecular disulfide bond locations of several AMPs are also very important for antimicrobial activity27. Harwig et al. designed PG-1 variants that lacked one pair of disulfide bonds - a “bullet” analogue (Cys → Ala at position of 8 and 13, disulfide bond distal to the turn) - and a “kite” analogue (Cys → Ala at position 6 and 15, disulfide bond proximal to the turn) and found that, compared with PG-1, the “kite” analogue had a greater loss of antimicrobial activity than the “bullet” analogue27.
Marine peptide NZ17074 (N1), a variant of arenicin-3 (Tyr5 → Asn, Tyr17 → His) isolated from the marine invertebrate lugworm Arenicola marina, exhibits potent antimicrobial activity against gram-negative bacteria and fungi, and it is currently undergoing preclinical studies28–30. However, N1 has a significant degree of toxicity in normal eukaryotic cells29, 30. Additionally, the role of disulfide bonds in modulating the antimicrobial activity of N1 is unclear. Thus, modification of N1 is required to generate more therapeutically valuable molecules. It has been found that several residues (Val6, 8, 13, 15, Tyr9, and Asn11, 21) of N1 are essential for antibacterial activity31. In the present study, to reduce cell cytotoxicity, determine the role of disulfide bonds, evaluate structure–activity relationships and improve antimicrobial activity, modification of N1 was performed based on the replacement of multiple residues by the hydrophobic/cationic residues Ala/Lys and the removal of disulfide bonds or the addition of hydrophobic Trp residue at the 1st, 3rd, 4th, 5th, 7th, 12th, 16th, and 20th positions, which generated four types of N1 variants: (i) “rocket” analogues with two disulfide bonds (Gly1,12 → Ala; Trp4, Asn5 → Ala), (ii) a “kite” analogue with one disulfide bond (Cys3,20 → Ala), (iii) a “bullet” analogue with one disulfide bond (Cys7,16 → Ala), and iv) other linear analogues without disulfide bonds (∆Cys3,7,16,20; Cys3,7,16,20 → Ala). These peptides were chemically synthesized and their antimicrobial activity against gram-negative bacteria was examined. Multiple possible levels of selective antimicrobial actions of N2 and N6 against E. coli and S. enteritidis were further elucidated. Additionally, the in vivo antibacterial activity of N2 and N6 was evaluated in murine peritonitis models induced by E. coli and S. enteritidis, respectively, as well as endotoxin neutralizing activity in a murine endotoxemia model challenged with LPS.
Results
Peptides designed based on structural determinants
Seven N1 analogues were obtained by changing disulfide bonds, hydrophobicity, or charges. According to the number and location of disulfide bonds, they are divided into the following classes: “rocket” analogues with two disulfide bonds (N2: Gly1,12 → Ala; N3: Trp4,Asn5 → Ala; N4: Trp0), a “kite” analogue with a Cys7-Cys16 disulfide bond (N6: Cys3,20 → Ala), a “bullet” analogue with a Cys3-Cys20 disulfide bond (N7: Cys7,16 → Ala), and other disulfide bond-free linear analogues (N5: ∆Cys3,7,16,20; N8: Cys3,7,16,20 → Ala) (Supplementary Fig. S1). As shown in Table 1, the PI, charge, hydrophilicity, aliphatic index, antibacterial activity score, and α-helix values of N4 were highly similar to those of the parent peptide-N1. The net charge of N3 was higher than that of N1 and the other analogues. The N3, N6, N7, and N8 peptides had a higher α-helix content (28.6%), and N2 and N5 contained a lower α-helix content (9.5% and 11.8%) than N1. The β-sheet contents of N5 to N8 deceased from 71.4% to 57.1%. Hydrophobicity of N2 and N3 increased from −0.133 to −0.033, indicating that they are more hydrophobic than N1 due to the addition of a methyl group by Gly/Gln to Ala residue substitution at positions 1, 5, and 12. The aliphatic indices of N2 to N8, with the exception of N4, ranged from 60.00 to 74.29, which were higher than that of N1. The Boman index of the N1 analogues, except for N5, deceased from 2.61 to 2.43 kcal/mol, indicating higher antibacterial activity. N3 had the highest antibacterial activity score, while N5 and N8 had the lowest score.
Table 1.
Sequences and physicochemical properties of N1 and its analogues.
| Amino acid sequence | MW (Da) | PI | Charge | GRAVY | AI | BI (kcal/mol) | AAS | Secondary structure (%) | ||
|---|---|---|---|---|---|---|---|---|---|---|
| α-helix | β-sheet | |||||||||
| Parent peptide | ||||||||||
| N1 (NZ17074) | GFCWNVCVYRNGVRVCHRRCN | 2452.0 | 9.37 | +4 | −0.243 | 55.24 | 2.66 | 0.245 | 0 | 81.0 |
| Hydrophobicity/charge modification: “rocket” | ||||||||||
| N2 | AFCWNVCVYRNAVRVCHRRCN | 2570.0 | 9.38 | +4 | −0.033 | 64.76 | 2.58 | 0.121 | 9.5 | 81.0 |
| N3 | GFCKAVCVYRNGVRVCHRRCN | 2440.9 | 9.69 | +5 | −0.133 | 60.00 | 2.63 | 0.413 | 28.6 | 81.0 |
| N4 | WGFCWNVCVYRNGVRVCHRRCN | 2728.1 | 9.37 | +4 | −0.273 | 52.73 | 2.43 | 0.273 | 0 | 86.4 |
| S-S modification: “kite”/“bullet”/“linear” | ||||||||||
| N5 | GF_WNV_VYRNGVRV_HRR_N | 2129.4 | 12.00 | +4 | −0.888 | 68.24 | 3.59 | −0.053 | 11.8 | 70.6 |
| N6 | GFAWNVCVYRNGVRVCHRRAN | 2477.8 | 10.72 | +4 | −0.310 | 64.76 | 2.61 | 0.229 | 28.6 | 71.4 |
| N7 | GFCWNVAVYRNGVRVAHRRCN | 2477.8 | 10.72 | +4 | −0.310 | 64.76 | 2.61 | 0.229 | 28.6 | 66.7 |
| N8 | GFAWNVAVYRNGVRVAHRRAN | 2413.7 | 12.00 | +4 | −0.376 | 74.29 | 2.56 | −0.007 | 28.6 | 57.1 |
Note: MW: molecular weight; PI: isoelectric point; GRAVY: grand average of hydropathicity; underlined residues: the modified residues. AI: aliphatic index; BI: Boman index; AAS: antibacterial activity score; S-S: disulfide bonds. N1, N2, N3 and N4: the “rocket” analogues with two disulfide bonds of Cys3-20 and Cys7-16; N6: the “kite” analogue with one disulfide bond of Cys7-16; N7: the “bullet” analogue with one disulfide bond of Cys3-20; N5 and N8: the other analogues without disulfide bonds.
N2/N6 have potent antimicrobial activity, low hemolysis and cytotoxicity, and no antimicrobial resistance
Antimicrobial activity. As shown in Table 2, the “rocket” N2 analogue, with two disulfide bonds (Gly1,12 → Ala), exhibited the strongest antimicrobial activity against gram-negative bacteria with minimal inhibitory concentration (MIC) values ranging from 0.25 to 1 μg/ml against E. coli, Salmonella, and Pseudomonas strains. Compared to N1, the “rocket” N3 (Trp4,Asn5 → Ala) showed relatively weaker antimicrobial activity against bacteria and Candida albicans. The additional introduction of Trp1 did not significantly affect antimicrobial activity of the “rocket” N4, indicating that Trp1 is not important for antimicrobial activity. Disulfide-free linear N5 (∆Cys3,7,16,20) had the lowest antimicrobial activity against Salmonella, Pseudomonas, and S. suis (with MICs > 64 μg/ml), which may be related to its higher hydrophilicity. The antimicrobial activity of the “kite” N6 (Cys3,20 → Ala) with one disulfide bond, was similar to that of N1; the “bullet” N7 (Cys7,16 → Ala) with one disulfide bond, had higher antimicrobial activity against several gram-positive bacteria than N1, but lower activity against gram-negative bacteria. Alanine substitution of four Cys residues largely decreased the antimicrobial activity of N8 (Cys3,7,16,20 → Ala) against bacteria, with an approximate 4–128-fold increase in MICs. This indicated that a single disulfide bond was sufficient to restore antimicrobial potency and that the Cys7-Cys16 disulfide bond rather than the Cys3-Cys20 one is indispensable for N1 antimicrobial activity.
Table 2.
The MIC values of N1 and its analogues.
| Strains | MIC (μg/ml) | |||||||
|---|---|---|---|---|---|---|---|---|
| N1 | N2 | N3 | N4 | N5 | N6 | N7 | N8 | |
| Gram-negative bacteria | ||||||||
| Escherichia coli CVCC195a | 0.5–1 | 0.25 | 1 | 1 | 64 | 0.5 | 2 | 32 |
| E. coli CVCC1515a | 0.25 | 0.5 | 1 | 1 | 128 | 1 | 2 | 16 |
| E. coli CICC21530 (serotype O157:H7)b | 0.5 | 0.25 | 1 | 1 | 64 | 0.5 | 2 | 16 |
| Salmonella typhimurium ATCC14028c | 0.5 | 1 | 4 | 2 | 128 | 2 | 8 | >64 |
| S. enteritidis CVCC3377a | 0.25 | 0.125 | 0.5 | 0.25 | 4 | 0.25 | 0.5 | 1 |
| S. pullorum CVCC1789a | 0.25 | 0.5 | 2 | 1 | >128 | 0.5 | 4 | >128 |
| S. pullorum CVCC1802a | 0.25 | 0.25 | 1 | 2 | >128 | 0.5 | 4 | 32 |
| S. choleraesuis CVCC503a | 0.5 | 0.5 | 4 | NA | >128 | 2 | 16 | >64 |
| Pseudomonas aeruginosa CICC10419b | 2 | 2 | >16 | NA | >128 | 8 | >64 | >128 |
| P. aeruginosa CICC21630b | 4 | 2 | 16 | 4 | >128 | 4 | >64 | >128 |
| Gram-positive bacteria | ||||||||
| Staphylococcus aureus ATCC43300c | 16 | 16 | >16 | 8 | >128 | 16 | >64 | >64 |
| S. aureus ATCC25923c | 0.5–1 | 0.25 | 0.25 | 1 | 2 | 0.25 | 0.5 | 2 |
| S. suis CVCC606a | 16 | 16 | 16 | NA | >128 | 16 | 8 | >128 |
| Enterococcus faecium CMCC1.2136d | >16 | 4 | 8 | NA | 64 | 4 | 2 | 16 |
| Bacillus subtilis ATCC6633c | 0.25 | 0.5 | 2 | 1 | 64 | 0.5 | 2 | 4 |
| Listeria ivanovii ATCC19119c | 128 | 4 | 8 | NA | 128 | NA | 2 | NA |
| Fungi | ||||||||
| Candida albicans CGMCC2.2411d | 16 | 32 | >16 | NA | >128 | 64 | >16 | NA |
Note: NA: no detection. aChina Veterinary Culture Collection Center (CVCC); bChina Center of Industrial Culture Collection (CICC); cAmerican Type Culture Collection (ATCC); dNational Center Center for Medical Culture Collection (CMCC). Data were representative of three independent experiments.
Low hemolysis, cytotoxicity, and no antimicrobial resistance
As shown in Fig. 1A, the hemolysis of N2 and N6 at a concentration of 128 μg/ml was 0.4% and 0.04%, respectively, which was lower than that of the parent peptide N1 (3%)29. Even at a concentration of 256 μg/ml, the hemolysis of N2 and N6 was 3.8% and 1.9%, respectively, indicating that the two peptides have very low hemolytic activity against murine erythrocytes. The cytotoxicity of N2 and N6 in murine peritoneal RAW264.7 macrophage cells was determined via the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay (Fig. 1B). The cell survival of N2 and N6 at a concentration range of 64–128 μg/ml was higher than that of N1, indicating that both N2 and N6 have lower cytotoxicity than N1 (from 25.3% to 37.9%). N2 demonstrated the lowest cytotoxicity (from 3.3% to 5.8%) and N6 (from 9.6% to 11.8%) had moderate cytotoxicity. These results suggested that the application of N2 and N6 at high concentrations can cause mild disruption of eukaryotic cells.
Figure 1.

Hemolytic activity, cytotoxicity and stability of N2 and N6. (A) Hemolytic activity of N2 and N6 against murine erythrocytes. (B) Cytotoxicity of the peptides against RAW264.7 cells. (C,D,E) The effects of temperature (C), pH (D) and proteases (E) on the antibacterial activity of N2 and N6 in vitro against S. enteritidis CVCC3377 cells. The results are given as the mean ± SEM (n = 3).
Additionally, after 18 serial passages in the presence of N2 or N6, the MIC values did not change and no mutants of E. coli resistant to N2 or N6 were observed (data not shown).
N2/N6 exhibited adequate thermal, alkaline pH, and anti-pepsin stability
The antimicrobial activity of N2 and N6 against E. coli and S. enteritidis was not affected upon exposure to 20, 40, 60, or 80 °C temperatures, but a small reduction of 2.2–16.5% for E. coli and of 2.8–10.9% for S. enteritidis was observed after incubation at 100 °C for 1 h (Fig. 1C, Supplementary Fig. S2A). N2 and N6 were stable between pH 8.0 and pH 10.0, but very susceptible to disruption between pH 2.0 and pH 4.0 (Fig. 1C, Supplementary Fig. S2B). Additionally, the inhibitory activity of N2 and N6 decreased by 40–100% for E. coli and by 51.4–100% for S. enteritidis after incubation for 4 h with trypsin and papain, and by 20.3–26.4% for E. coli and 13.3–17.7% for S. enteritidis after treatment with pepsin (Fig. 1E, Supplementary Fig. S2C).
Structures of N2/N6 analyzed by circular dichroism (CD) and nuclear magnetic resonance (NMR)
CD
The anionic surfactant sodium dodecyl sulfate (SDS) is often used to mimic the cell membrane environment. To analyze the structural features of N2/N6, the CD spectra of peptides were measured in ddH2O and 10–40 mM SDS, respectively. The secondary structures of N2/N6 in ddH2O were characterized predominantly by antiparallel strands (56.4–57.9%), α-helices (19.6–22.0%), and β-turns (21.2–21.9%) with a characteristic positive maximum at 190 nm and 230 nm and negative minimum at 205 nm (Fig. 2A). Both N2 and N6 showed a significant increase in α-helices (>80.3%), higher than the parent peptide N1 (<54.6%), and decrease in antiparallel strands (<0.8%) in SDS (Supplementary Table S1). This result indicated that SDS is favorable to form α-helical structures.
Figure 2.
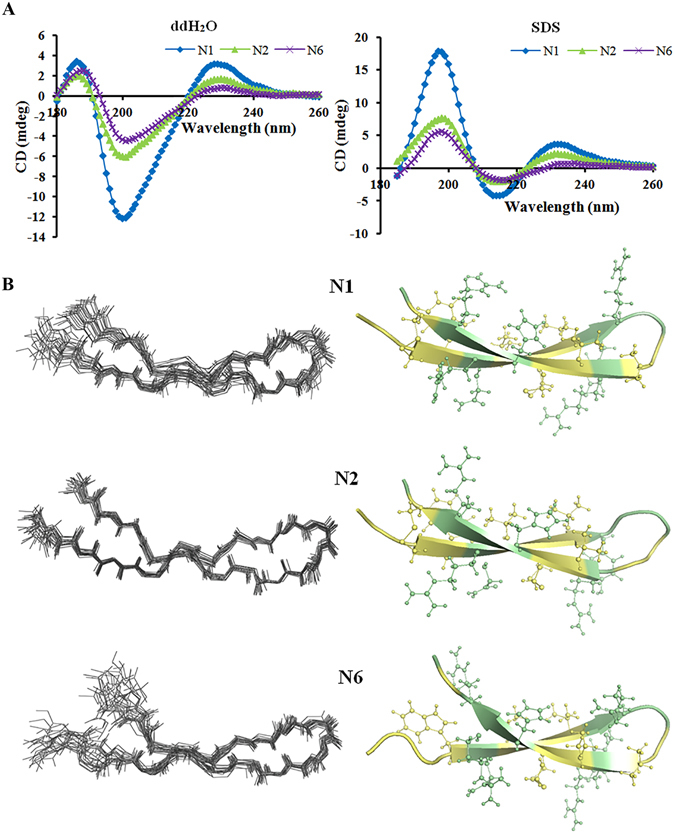
Structures of N1, N2, and N6. (A) CD spectra of N1, N2 and N6. (B) NMR analysis of N1, N2 and N6. Left panel shows the sets of the 20-best structures, superimposed over the backbone atoms. Only backbone atoms are shown. Right panel shows the ribbon representations of the peptides. The side chains on the β-sheet strands are represented by balls and sticks. The hydrophobicity and hydrophilicity of the residues are color-coded pale yellow and pale-green, respectively. The figure was prepared using the program Pymol.
NMR
From 100 calculated structures, 20 with the lowest energy were selected to undergo water refinement, and the refined structures were used to represent the peptide structure in aqueous solution. The NMR-determined structures of N1, N2 and N6 are shown in Fig. 2B as superimposed backbone traces of the ensembles and their representative ribbon diagrams. The ribbons were generated using model 2 of N1, model 20 of N2 and model 19 of N6, which were the medoid structures of each ensemble. The structural statistics are listed in Table 3. Not surprisingly, N1, N2 and N6 adopt a β-hairpin structure (Fig. 2B) stabilized by one (N6, Cys7-Cys16) or two (N1 and N2, Cys7-Cys16 proximal to the β-sheet, and Cys3-Cys20 distal to the β-sheet) disulfide bonds and several inter-strand hydrogen bonds, which are determined both in the structure calculation process and by analyzing the temperature coefficients (Supplementary Table S2). The antiparallel β-strands are Cys3-Arg10 and Val13-Cys20 for N1, Cys3-Try9 and Arg14-Cys20 for N2, and Trp4-Arg10 and Val13-Arg19 for N6 (Fig. 2B). The Gly to Ala mutation in N2 disturbs the tip of the β-hairpin and twists the connecting loop slightly, possibly to shield the larger hydrophobic side chain of Ala. Supplementary Fig. S3 illustrates the surface of each peptide with composing amino acids color-coded as hydrophobic or hydrophilic. N1, N2 and N6 showed impressive characteristics of amphipathicity. The hydrophobic side is dominated by F2, W4, V6, C7 and V8, while the hydrophilic side is composed mainly of R10, N11, H17, R18, R19 and N21. The electrostatic map indicated that N1 and N6 possess large cationic areas with very limited negative charge (Supplementary Fig. S4). However, N2 somehow has a greater negative charge and a disrupted positive-charge area. The absence of one disulfide bond in N6 shortens each of the β-sheet-forming strands by one residue (A3 and A20) and makes the residues at the N and C terminal more flexible, as suggested by larger amide proton temperature coefficients (Supplementary Table S2). The double disulfide bonds do not guarantee a more thermo-stable structure, as seen in Fig. 1C. However, these analogues do show pH stability in an alkaline pH range (Fig. 1D) and more protection against pepsin and papain but not trypsin.
Table 3.
Structure statistics for 20 conformers of the three peptides N1, N2 and N6.
| N1 | N2 | N6 | |
|---|---|---|---|
| Unambiguous distance restraints | |||
| Intrar-residue(i-j = 0) | 150 | 169 | 115 |
| Sequential (|i-j| = 1) | 68 | 85 | 81 |
| Medium range (2 ≤ |i-j| ≤ 4) | 16 | 37 | 19 |
| Long range (|i-j| ≥ 5) | 51 | 75 | 34 |
| Ambiguous distance restraints | 6 | 11 | 10 |
| Hydrogen bond | 9 | 7 | 3 |
| Disulfide bond | 2 | 2 | 1 |
| Total | 302 | 386 | 263 |
| Dihedral angle restraints | |||
| Φ | 17 | 17 | 17 |
| Ψ | 17 | 17 | 17 |
| Total | 34 | 34 | 34 |
| Mean r.m.s. deviations from the experimental restraints | |||
| Distance (Å) | 1.73e-2 ± 5.48e-3 | 2.27e-2 ± 2.65e-3 | 2.54e-2 ± 3.25e-3 |
| Dihedral angle (°) | 2.14e-1 ± 1.14e-1 | 3.23e-1 ± 8.70e-2 | 4.30e-1 ± 1.09e-1 |
| Mean r.m.s. deviations from idealized covalent geometry | |||
| Bond (Å) | 3.57e-3 ± 2.24e-4 | 3.99e-3 ± 2.03e-4 | 3.97e-3 ± 1.99e-4 |
| Angle (°) | 4.01e-1 ± 2.28e-2 | 4.78e-1 ± 2.12e-2 | 4.41e-1 ± 2.64e-2 |
| Improper (°) | 8.82e-1 ± 1.43e-1 | 1.03 ± 1.23e-1 | 1.04 ± 1.37e-1 |
| Ramachandran plot | |||
| most favorable regions | 92.6% | 87.9% | 85.9% |
| additional allowed regions | 7.4% | 12.1% | 14.1% |
| generously allowed regions | 0% | 0% | 0% |
| Atomic r.m.s. differences (Å)b | |||
| Backbone heavy atoms | 0.76 (0.54) | 0.49 (0.19) | 1.29 (0.20) |
| All heavy atoms | 1.27 (1.15) | 0.93 (0.75) | 1.94 (0.95) |
None of the structures exhibits distance violations greater than 0.3 Å or dihedral angle violations greater than 4˚. aThe program PROCHECK (http://www.biochem.ucl.ac.uk/~roman/procheck_nmr/procheck_nmr.html) was used to assess the overall quality of the structures; bThe numbers in the parenthesis are the rms differences for the well-ordered region (residues 3–20 for N1, residues 2–20 for N2, and residues 3–19 for N6).
N2/N6 permeabilized model membranes and cell membranes and disrupted membrane potentials
Model membranes
To gain insights into the mechanism of action of N2 and N6, the release of the fluorescent dye calcein from model bacterial and mammalian membranes was measured with a Tecan Infinite M200 PRO microplate reader. As shown in Fig. 3A, N2 and N6 induced 3.0–10.5% and 6.3–21.9% dye leakage from phosphatidylcholine (POPC)/phosphatidylglycerol (POPG) large unilamellar vesicles (LUVs) (imitating bacterial cell membranes) at 1×, 2×, and 4× MIC against E. coli cells, which was higher than the parent peptide N1. N2 and N6 induced less dye leakage (0.6–19.9% and 0.4–15.4%) from POPC/cholesterol LUVs mimicking eukaryotic cell membranes at concentrations from 64 to 256 μg/ml compared to N1 (2.5–25.3%) and polymyxin B (PMB) (12.3–51.0%). This suggested that N2 and N6 have more potent abilities to permeabilize cell membranes of gram-negative bacteria and lower cytotoxicity against eukaryotic cells than N1.
Figure 3.
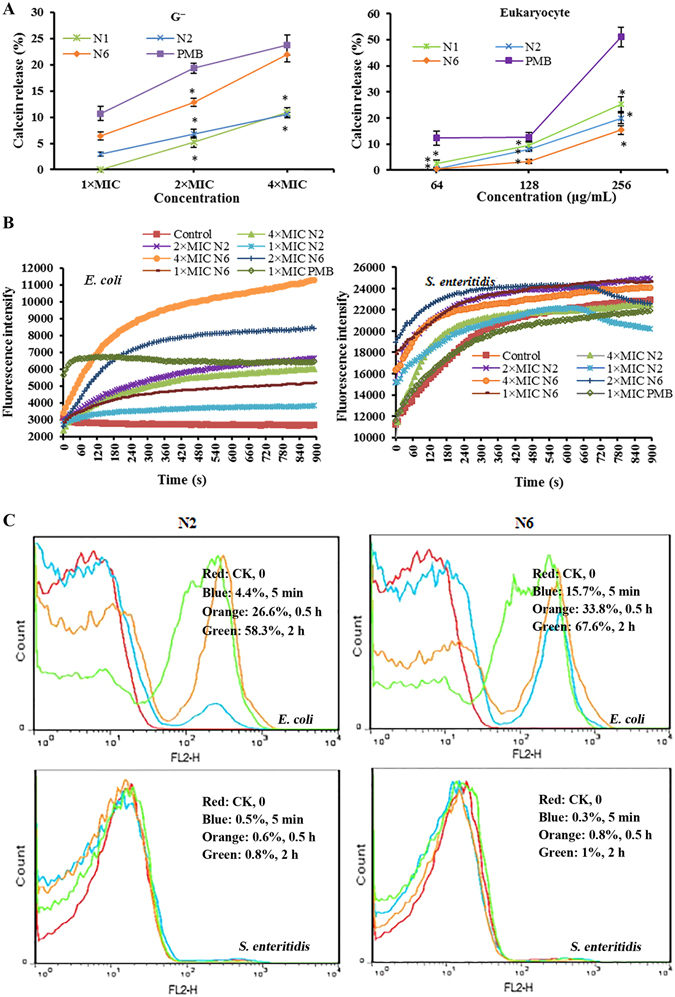
Penetration of biofilms by N2 and N6. (A) Calcein leakage from POPC/POPG = 4:1 (gram-negative) and POPC/cholesterol = 1:1 (eukaryotic) model membranes induced by different concentrations of N2 and N6, respectively. The results are given as the mean ± SEM (n = 3). Differences between groups were determined by one-way ANOVA followed by SPSS analysis. *p < 0.05 compared to the PMB group. (B) Outer membrane permeabilization assays of N2 and N6. (C) Flow cytometric analysis of PI-staining in E. coli CVCC195 and S. enteritidis CVCC3377 cells treated with 1× MIC N2 and N6, respectively. Red line: no peptide, negative control; Blue line: treatment with peptides for 5 min; Orange line: treatment with peptides for 0.5 h; Green line: treatment with peptides for 2 h.
Outer membrane permeability
The ability of peptides to permeabilize outer membranes was measured with the N-phenyl-1-naphthylamine (NPN) fluorescent dye. As shown in Fig. 3B, both N2 and N6 induced a time-dependent and concentration-dependent increase in NPN fluorescence in both E. coli and S. enteritidis cells. The addition of 1× and 2× MIC of N2 or N6 to E. coli cells caused a small increase in fluorescence intensity, indicating slow outer membrane permeability of peptides. However, permeabilization of S. enteritidis began within 1 min after treatment with N2 or N6 and a one-fold increase in fluorescence was observed, indicating that the two peptides can instantly permeabilize the outer membrane of S. enteritidis cells and that N6 has higher penetrating ability than N2.
Inner membrane permeability
The red fluorescent dye propidium iodide (PI), which is blocked on the outside of intact membranes, can penetrate damaged cell membranes and intercalate into nucleic acid. The fluorescent intensity of PI indicates the level of cell membrane integrity. In the absence of peptides, 99% of cells exhibited no PI staining, indicating that membranes were intact (Fig. 3C). After treatment with N2 and N6 for 5 min−2 h, the percentages of PI-permeable E. coli cells were 4.4–58.3% and 15.7–67.6%, respectively. This suggested that the inner membrane of E. coli is disrupted after 5 min of treatment with N2 and N6. However, both N2 and N6 did not disrupt the inner membrane of S. enteritidis, which showed <1% PI-staining.
Membrane potential
The changes in membrane potential were examined using rhodamine-123, which accumulates on the inner surface of intact membranes. If the membrane is disrupted, the potential dissipates and fluorescence diminishes32. As shown in Supplementary Fig. S5, cellular fluorescence of E. coli cells treated with N2 or N6 for 2 h was elevated in a concentration-dependent manner compared to that of control cells without treatment (CK). S. enteritidis cells treated with 1×, 2×, and 4× MIC N2 or N6 had a slight increase in cellular fluorescence. This result indicated that both N2 and N6 caused plasma membrane potential depolarization in E. coli, but that the inner membrane of S. enteritidis is intact and is not disrupted by N2 and N6, consistent with investigations of the inner membrane permeability.
Intracellular ROS accumulation
Reactive oxygen species (ROS) are a trigger that induce cell apoptosis and have destructive actions on DNA and proteins30. To determine if ROS are a mechanism of bacteria-killing, we used the ROS-sensitive dye dihydrorhodamine-123 (DHR-123) to monitor the production and accumulation of intracellular ROS after incubation with peptides. The results showed that N2 and N6 did not induce changes in fluorescence in E. coli (Supplementary Fig. S6A and B). However, S. enteritidis cells exhibited higher ROS levels than untreated cells after treatment with N2 or N6 (Supplementary Fig. S6C and D). This result indicated that N2 and N6 promoted ROS generation in S. enteritidis cells, which may be the main mechanism of killing them.
Morphological changes observed by electron microscopy after treatment with N2/N6
Scanning electron microscopy (SEM) observations
E. coli CVCC195 and S. enteritidis CVCC3377 cells were treated for 2 h with 4× MIC N2 and N6, respectively, and observed by SEM to investigate cellular morphological changes. As shown in Fig. 4A, the non-treated E. coli cells exhibited intact smooth surfaces, while the cells treated with peptides underwent considerable structural changes. After treatment with N2, over 50% filamentous cells with filiferous substances outside of E. coli cells were produced. Collapsed and filamentous E. coli cells were observed after treatment with N6. For S. enteritidis, several holes on the outer membrane of cells were observed after treatment with N2 and N6, but overall cell morphology had no change.
Figure 4.

Electron microscopy analysis of E. coli CVCC195 and S. enteritidis CVCC3377 cells treated with N2 and N6. Bacteria in mid-logarithmic growth phases were treated with peptides at 4× MIC for 2 h. (A) SEM images of E. coli CVCC195 and S. enteritidis CVCC3377 cells treated with N2 and N6. (B) TEM images of E. coli CVCC195 and S. enteritidis CVCC3377 cells treated with N2 and N6.
Transmission electron microscopy (TEM) observations
The effects of N2 and N6 on E. coli CVCC195 and S. enteritidis CVCC3377 cells were also visualized using TEM. As shown in Fig. 4B, the untreated control E. coli CVCC195 and S. enteritidis CVCC3377 cells displayed normal morphology and intact cell membranes, and the cytoplasm appeared to have homogeneous electron density. After treatment with 4× MIC N2 or N6 for 2 h, cell membrane disruption and leakage of cellular contents (approximately 40%) were observed in E. coli. For S. enteritidis, cell swelling and ghost cells were observed after treatment with N2 and N6. The cytoplasm displayed a heterogeneous electron density, the cell morphology was deformed, and approximately 15% cell lysis occurred.
N2/N6 bound to bacterial genomic DNA, disrupted cell cycles, and inhibited macromolecular synthesis
DNA gel movement retardation
In an attempt to clarify the molecular mechanism of action, the DNA-binding properties of N2 and N6 were examined by analyzing electrophoretic mobility of DNA bands at various mass ratios of peptides to DNA on an agarose gel. N2 and N6 inhibited the migration of genomic DNA from E. coli at mass ratios greater than 0.1 and 0.15 (Fig. 5A), respectively. The migration of genomic DNA from S. enteritidis was suppressed by N2 and N6 at mass ratios greater than 0.2 and 0.15, respectively. This result indicated that N2 and N6 can bind to bacterial DNA.
Figure 5.

Interaction of N2 and N6 with bacterial DNA and biomacromolecule synthesis. (A) Interaction of N2 and N6 with bacterial genomic DNA by a gel migration assay. M: DNA marker λDNA/HindIII; 1–8: the mass ratios of N2/ DNA were 0, 0.05, 0.1, 0.15, 0.2, 0.25, 0.3, and 0.4; 9–16: the mass ratios of N6/DNA were 0, 0.05, 0.1, 0.15, 0.2, 0.25, 0.3, and 0.4. (B) Impact of N2 and N6 on macromolecular synthesis in E. coli and S. enteritidis. Incorporation of 3H-thymidine (DNA), 3H-uridine (RNA), 3H-glucosamine (peptidoglycan), and 3H-leucine (protein) was determined in cells treated with 1× MIC N1, N2, and N6, respectively. Ciprofloxacin (Cip, 8× MIC), rifampicin (Rif, 4× MIC), vancomycin (Van, 2× MIC), and erythromycin (Ery, 2× MIC) were used as controls. The results are given as the mean ± SEM (n = 3).
Cell cycle arrest at the I-/R-phase
Changes in DNA frequency distribution histograms of bacterial cells exposed to N2 and N6 for different times are shown in Supplementary Table S3 and Supplementary Figs S7 and S8. With incubation of E. coli cells with N2 and N6 for 0.5 h, the proportion of cells in I-phase appeared to increase from 33.4 to 41.9% and to 39.4%, respectively. The population of I-phase cells increased to 50.6% and 46.5% with incubation for 2 h in the presence of N2 and N6, respectively. In contrast, decreases in the percentages of cells in the R- and D-phases were observed. This indicated that N2 and N6 could induce E. coli cell cycle arrest in the I-phase. Exposure to N2 and N6 for 5 min−2 h in S. enteritidis cells led to an increase in the percentage of R-phase cells in a time-dependent manner (from 65.0% to 80.6% and from 66.8% to 82.3%, respectively) and a corresponding reduction in the percentage of cells in the I-phase (from 26.0% to 15.8% and from 15.8% to 5.2%, respectively) and D-phase (from 8.0% to 3.6% and 17.4% to 5.5%, respectively). This indicated that N2 and N6 induced cell cycle arrest in the R-phase in S. enteritidis cells.
Inhibition of DNA and protein synthesis
The incorporation of radioactive precursors into DNA (3H-thymidine), RNA (3H-uridine), protein (3H-leucine), and peptidoglycan (3H-glucosamine) was measured to evaluate the effects of N2 and N6 on macromolecular synthesis in E. coli CVCC195 and S. enteritidis CVCC3377 cells. As shown in Fig. 5B, a significant inhibition of 3H-thymidine (28.6%) and 3H-uridine (18.7%) incorporation was observed with exposure of E. coli cells to 1× MIC N2 for 15 min, suggesting that N2 is a DNA and RNA synthesis inhibitor. N6 induced a decrease in DNA (43.8%) and cell wall (19.6%) synthesis, suggesting that N6 is a DNA and cell wall synthesis inhibitor. In S. enteritidis, however, N2 caused an inhibition in the incorporation of radioactive precursors into DNA (35.5%) and protein (17.0%), and N6 inhibited DNA (61.8%) and RNA (5.7%) biosynthesis.
N2/N6 bound to LPS and protected mice from lethal challenge with LPS
Harboring predicted LPS-binding sites
3D-structures of N1, N2, and N6 peptides were docked onto LPS to obtain models of the peptide/LPS complexes (Fig. 6A). Interactions between N1 and LPS were presumably stabilized by hydrogen bonding between positively charged side chains of the His9, Arg14, and Arg19 basic residues and the fatty acid chains MYR-1014, GLC-1007, and KDO-1003 of LPS (Fig. 6A). The Trp4, Arg10, Arg14, and Asn21 residues in N2 appear to form a number of salt-bridges and hydrogen bonds with the fatty acid chains FTT-1010, 63-α-D-glucosyl-maltotriose GMH-1005, MYR-1014, PA1-1000 and the monophosphate group of the lipid A moiety of LPS. Arg19 forms a hydrogen bond with the 2-Keto-3-deoxyoctonate (KDO-1003), which belongs to the core polysaccharide of LPS (Fig. 6A). The N6-LPS docked complex reveals probable hydrogen bonds or salt bridges between side chains of the Arg10/Arg19/Asn21 residues and FTT-1010/glucose GLC-1006/PA1-1000 of LPS (Fig. 6A). This indicated that N1, N2, and N6 have different LPS-binding sites and different modes of action.
Figure 6.
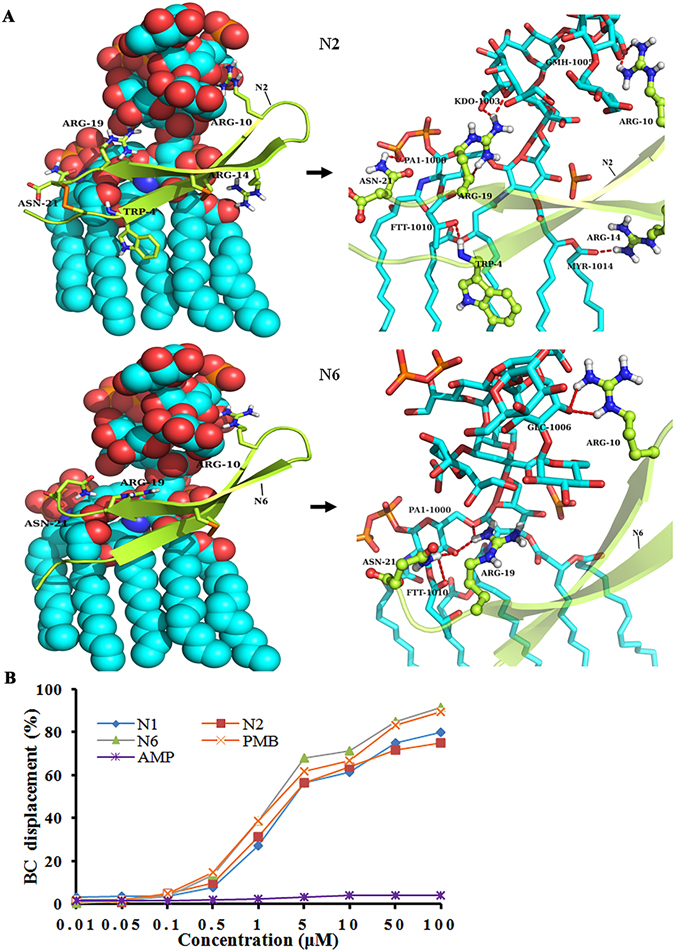
Potential docking of N2 and N6 with LPS. (A) The complex structure of N2 and N6 and LPS (left) and detailed view of the binding pocket (right). N2 and N6 are shown as cartoons and the key residues (N2: Trp4, Arg10, Arg14, Arg19, and Asn21; N6: Arg10, Arg19, and Asn21) are shown as sticks and balls. In this proposed autodock-generated model, the fatty acid chains FTT-1010, 63-α-D-glucosyl-maltotriose GMH-1005, MYR-1014, PA1-1000, and the monophosphate group that belongs to the lipid A moiety form hydrogen bonds and salt bridges to Trp4, Arg10, Arg14, and Asn21 of N2, while Arg19 forms a hydrogen bond with the 2-Keto-3-deoxyoctonate (KDO-1003), which belongs to the core polysaccharide of LPS. The fatty acid chain FTT-1010, glucose GLC-1006, and PA1-1000, respectively, form hydrogen bonds to Arg10, Arg19, and Asn21 of N6. (B) Binding affinity of N2 and N6 to LPS. Ampicillin (AMP) and PMB were used as negative and positive controls, respectively.
LPS binding affinity in vitro
To reveal the mechanism of N1 analogues, binding to LPS was determined using the BODIPY’-TR-cadaverine (BC) displacement assay. PMB, a positive control33, showed potent LPS binding affinity (Fig. 6B), whereas ampicillin did not displace BC and bind to LPS (Fig. 6B). Similarly to PMB, N1, N2, and N6 induced a significant dose-dependent probe displacement, reflecting their high capacity to bind to LPS. The BC displacement percentage of N6 (91.5%) at 100 μM was greater than that of N1 (79.8%), N2 (75.0%), and PMB (89.3%), indicating that N6 has the highest binding affinity to LPS.
N2/N6 protected mice from lethal challenge with E. coli and S. enteritidis or LPS
E. coli-induced peritonitis
To evaluate the therapeutic activity of N2 and N6 in a peritonitis model, mice were intraperitoneally inoculated with E. coli CVCC1515 and S. enteritidis CICC22956 strains, respectively. Mice in the uninoculated control group survived throughout the experimental period (Fig. 7A). The untreated mice began to die 8 h after inoculation with E. coli, and all untreated mice died within 48 h. After treatment with 1.25 and 2.5 mg/kg N2, the survival rates of mice were 66.7% and 100%, respectively, which was higher than for N6 at the same doses (16.7% and 66.7%). The survival rate of mice treated with 5 mg/kg N6 was 100%. The survival rates of mice treated with 0.31 and 0.625 mg/kg PMB were 66.7% and 100%, respectively (Fig. 7A).
Figure 7.

Effect of N2 and N6 on bacteria-induced and LPS-induced responses in vivo. (A) Survival of mice treated with N2 and N6 in E. coli, S. enteritidis, and LPS lethal models. Mice were intraperitoneally injected with E. coli (2.5 × 108 CFU/ml), S. enteritidis (1 × 106 CFU/ml), and LPS (30 mg/kg of body weight), respectively, followed by injection with N2, N6 or PMB at 0.5 and 8 h. Survival was recorded for 7 days. (B) Inhibition of LPS-induced cytokine release in C57BL/6 mice by N2 and N6. The results are given as the mean ± SEM. Differences between groups were determined by one-way ANOVA followed by SPSS analysis (n = 3 per group). *p < 0.05 compared to the LPS-treated group. (C) Protection of lungs from damage by LPS. Lung specimens were collected from sacrificed mice after LPS injection and treatment. The sections were stained with hematoxylin and eosin. V: vessel; A: alveoli; T: trachea. 1: alveolar septum thickening; 2: pulmonary alveolar collapse; 3: inflammatory cell infiltration.
S. enteritidis-induced peritonitis
In another peritonitis model, mice were injected with N2/N6 (5 mg/kg and 7.5 mg/kg) or PMB (0.31 and 0.625 mg/kg) at 0.5 h and 8 h after challenge with S. enteritidis. Mice in the uninoculated control group survived throughout the experimental period (Fig. 7A). All mice without treatment died within 72 h after inoculation with S. enteritidis. After treatment with 5 mg/kg N2 and N6, the survival rates of mice were 83.3% and 66.7%, respectively. All mice treated with 7.5 mg/kg N2 or N6 survived. The survival rates of mice treated with 0.31 and 0.625 mg/kg PMB were 83.3% and 100%, respectively (Fig. 7A).
LPS-induced endotoxemia
The LPS-detoxifying activity of N2 and N6 was evaluated in mice challenged with LPS. At 0.5 h and 8 h after challenge with LPS, mice were treated with N2/N6 (2.5 and 5 mg/kg) or PMB (10 and 15 mg/kg). Mice in an unchallenged control group did not die during the experimental period (Fig. 7A). Mice without treatment began to die at 8 h after challenge with LPS, and all untreated mice were dead within 24 h. After treatment with 2.5 mg/kg N2 and N6, the survival rates of mice were 66.7% and 33.3%, respectively. Meanwhile, mice administered 5 mg/kg N2 and N6 improved their survival rates to 100%. The survival rates of mice treated with 10 and 15 mg/kg PMB were 16.7% and 100%, respectively (Fig. 7A). This result indicated that both N2 and N6 can protect mice from lethal LPS challenge in vivo, and have better therapeutic efficacy than PMB.
Inhibition of inflammatory cytokine production
To further explore the effects of N2/N6 on inflammatory cytokines, an endotoxemia mouse model was constructed by intraperitoneal injection with 10 mg/kg LPS followed by treatment with peptides or antibiotics. As shown in Fig. 7B, similarly to PMB, N2 and N6 significantly reduced the release of tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), and interleukin-1β (IL-1β). The levels of TNF-α, IL-6, and IL-1β in endotoxemia mice treated with N2 (111.9, 630.3, and 33.3 pg/ml, respectively), N6 (127.2, 350.1, and 127.2 pg/ml, respectively) or PMB (74.0, 514.9, and 242.8 pg/ml, respectively) were significantly lower than those of the corresponding LPS-challenged control groups (394.9, 1026.8, and 292.9 pg/ml, respectively). This indicated that both N2 and N6 can inhibit the production of proinflammatory cytokines.
Protection of LPS-induced lung injury
To explore if N2 and N6 can protect mice from LPS-induced lung injury, lung tissues were dissected and examined 7 d after treatment. As shown in Fig. 7C, no pathological change was observed in the lungs of mice injected only with PBS, whereas acute injury, including fibrosis, alveolar septum distention, inflammatory cells, and red blood cell infiltration, was found in the untreated LPS-challenged mice. In contrast, after treatment with 5 mg/kg N2 and N6, there was less apparent damage in the lungs of treated mice. At a high dose of 7.5 mg/kg N2 or N6, no obvious pathological changes were found in the murine lung tissues. The efficacy of N2 and N6 was higher than that of PMB. These data demonstrated that both N2 and N6 may be potential therapeutics for E. coli and S. enteritidis infection and LPS-induced endotoxemia.
Discussion
In this work, a series of N1 analogues were designed by changing disulfide bonds, hydrophobicity, or charge to reduce cell cytotoxicity, investigate the effects of disulfide bonds on antimicrobial activity, and develop novel cell-selective β-sheet AMPs with improved antimicrobial activity. Among these analogues, the “rocket” analogue N2, with two disulfide bonds, and the “kite” analogue N6, with one disulfide bond, were found to be the most active against both E. coli and S. enteritidis (Table 2), and multiple levels of selective antimicrobial mechanisms of the two peptides and detoxifying activity were systematically investigated in vitro and in vivo.
It has been demonstrated that an increase in the hydrophobic moment and net charge leads to an increase in antibacterial activity of AMPs34. In our study, the favorable effect of residue replacement on the activity of N2 may result from increased hydrophobicity due to the replacement of the Gly residue at positions 1 and 12 in N1 by Ala (Table 2; Supplementary Fig. S3). On the one hand, the Gly → Ala substitution resulted in a conformational perturbation of approximately 9.5% of the α-helical structure formation (Table 1, Fig. 2) due to the introduction of a β-carbon into the 1 and 12 positions of N235, which controls the peptide flexibility and contributes to optimal antimicrobial activity (Table 2)36. On the other hand, the Gly → Ala substitution led to a marked decrease in the thermal stability of N2 at 100 °C (Fig. 1C), indicating an increase in free energy from the Ala substitution, as observed or other peptides35, 36. Additionally, compared to N1, a more positively charged N-terminus (from +4 to +5) of the “rocket” N3 analogue, through Lys replacement at position Trp4, decreased activity (Table 2), while the substitution of Asn at position 5 by Ala led to an increase in hydrophobicity (from −0.243 to −0.113) (Table 1). This may be attributed to destabilization of the pore due to increased repulsion among peptide monomers or to an imbalance between positively charged amino acids and hydrophobicity37, 38, although the correlation with activity is not fully clarified. Introduction of Trp at the N-terminus of N1 gave rise to the “rocket” N4 analogue with a potent hydrophobic nature (from −0.243 to −0.273) which showed relatively weaker antimicrobial activity than N1, indicating that the Trp1 side chain is unimportant for activity. Linear N5 and N8 analogues exhibited the lowest hydrophobicity (−0.888 and −0.376) and antibacterial activity (Tables 1, 2). Therefore, these results demonstrated that hydrophobicity is very important for the full activity of N1 peptides.
Due to the small size of AMPs, it is thought that disulfide bonds are critical for maintaining the 3D-folding and structure of the molecule. However, several AMPs, such as hBD3 and μ-conotoxins, have shown that disulfide bonds are not absolutely necessary for antimicrobial activity against pathogens20, 22. Furthermore, variants of HNP-1 and cryptdin-4 with fewer disulfide bonds are more antimicrobial than the wild-type peptide against E. coli, S. aureus, and P. aeruginosa 19, 24. In this study, the antimicrobial activity of the “kite” analogue N6 was almost unaffected by the deletion of the first disulfide bond (Table 2), indicating that the first disulfide bond (Cys3-Cys20) in N1 is not required for its antimicrobial activity. However, the second disulfide bond (Cys7-Cys16) in the “bullet” analogue N7 is indispensable for its antimicrobial activity. Linear N5 and N8 analogues without disulfide bonds lost considerable activity against all tested bacterial strains. These data suggest that the second disulfide bond (Cys7-Cys16), which is proximal to the β-sheet, plays a major role in stabilizing the β-sheet structure within N1 and maintaining the antibacterial activity, in contrast to the PG-1 variants27. Perhaps this is related to differences in the nature, the structure, and the spacing between their disulfide bonds.
Our previous study has demonstrated that N1 can interact electrostatically with the cell membrane of E. coli, permeabilize the outer and inner membrane, cause depolarization and leakage of contents, and eventually lead to cell death30. In this study, N2 and N6 exhibited remarkable specificity for particular AMP–bacteria pairings. Both N2 and N6 more strongly permeabilized a model bacterial membrane than did N1, the outer and inner membrane of E. coli, and the outer but not the inner membrane of S. enteritidis (Fig. 3C). This may be related to more conformational flexibility in the N- and C-terminal regions of N2 and N6 (Fig. 2B), additionally conferring less toxicity than N1 (Fig. 1A and B), which is consistent with other AMPs such as LAK120, LAK80 and their analogues39. Furthermore, the inner membrane potential of S. enteritidis is rarely changed by the two peptides and more than 99% of the inner membrane integrity was maintained (Supplementary Fig. S6C). This is consistent with a previous report in which pleurocidin and its analogues did not cause inner membrane permeabilization despite inhibiting intracellular macromolecular synthesis in E. coli 40. Moreover, we demonstrated that the activity of the two peptides against S. enteritidis cells was associated with the accumulation of ROS, which is consistent with the results of a previous study which found that ROS accumulation induced by N1 may play an important role in apoptosis30. These results suggest that the nature of AMPs and the characteristics of the bacterial cell membrane determine the antimicrobial actions of AMPs41.
Another significant effect of N2 and N6 on E. coli and S. enteritidis was evident from their DNA-binding properties. The migration of genomic DNA from E. coli and S. enteritidis was inhibited by N2 and N6 when a DNA mass ratio was greater than 0.1 (Fig. 5A), which is similar to our previous report that N1 bound tightly to DNA by inserting base pairs and changing the DNA conformation30.
The cell wall is a shape-maintaining element in bacteria42. The collapse of E. coli cells treated with N6 was observed in SEM images (Fig. 4A), which seems to be a consequence of cell wall synthesis inhibition by N6 (Fig. 5B). Additionally, both N2 and N6 inhibited other macromolecular synthesis in bacterial cells (Fig. 5B). The inhibition of DNA synthesis in E. coli cells was accompanied by slightly delayed cessation of RNA (N2) and cell wall (N6) synthesis (Fig. 5B). This effect of N2 on E. coli was similar to that observed for N1 (30). However, synthesis of DNA, RNA, and proteins in S. enteritidis treated with N2 and N6 seems to be affected in a direct way (Fig. 5B) but probably by different mechanisms, as confirmed by SEM images showing intact cell morphology (Fig. 4A). This is consistent with recently published data for pyrrhocoricin and oncocin which function by repressing translation or by binding to the 70S ribosome43, 44.
LPS, a highly conserved major component of the outer membrane of gram-negative bacteria, is released spontaneously and rapidly during bacterial growth and after exposure to antibiotics7, 45. It is the primary trigger of endotoxemia because it stimulates monocytes and macrophages to produce large amounts of proinflammatory mediators, such as TNF-α and IL-67, 46, 47. Dixon et al. reported that the synthetic AMP KSL-W can bind to the LPS of E. coli and various oral bacteria14. Our previous study has demonstrated that N1 can bind LPS and reduce the release of proinflammatory cytokines in LPS-induced acute peritonitis in mice30. In this study, BC probe displacement and docking calculation confirmed binding of N2 and N6 with LPS through the formation of hydrogen bonds and salt bridges (Fig. 6 and Supplementary Fig. S9). It is speculated that the binding of N2 and N6 with LPS may initially interrupt the interaction of LPS with its toll-like receptor, and subsequently inhibit the release of LPS-induced IL-6, IL-1β and TNF-α in mice, thereby effectively protecting mice from acute lung injury (Fig. 7B and C). Moreover, N2 and N6 enhanced the survival rates of peritonitis- and endotoxemia-induced mice, which were significantly superior to PMB in the endotoxemia model (Fig. 7A). Together, these results suggest that, similarly to N1, both N2 and N6, having low cytotoxicity and no antimicrobial resistance, are potential antibacterial and endotoxemia therapeutics.
In this study, we designed a series of N1 analogues based on disulfide bonds, hydrophobicity, and charge. Both N2 and N6 showed potent antimicrobial activity against E. coli and S. enteritidis, with N2 having higher activity than N1. N2 and N6 had lower hemolysis and cytotoxic effects than N1 due to increased flexibility in the N- and C-terminal regions. Both N2 and N6 disrupted the outer and inner membranes of E. coli, induced cell cycle arrest at the I-phase, and inhibited DNA/RNA/cell wall synthesis in E. coli. For S. enteritidis, the outer membrane rather than the inner membrane was permeabilized, cell cycle was arrested at the R-phase, and DNA/RNA/protein synthesis was inhibited by N2 and N6. N2 and N6 protected mice from mortality induced by bacterial infection (E. coli and S. enteritidis) and E. coli LPS. Our findings suggest that N2 and N6 are promising candidates for further research as new antimicrobial and anti-endotoxin agents.
Materials and Methods
Peptide design
The N1 analogues were designed based on the replacement of Gly, Cys and Gln residues by the hydrophobic/cationic residues Ala/Lys and the removal of disulfide bonds or the addition of a hydrophobic Trp residue at positions 1, 3, 4, 5, 7, 12, 16, and 20 (Table 1). The physicochemical properties of these peptides were calculated with Protparam (http://web.expasy.org/protparam/) and APD3, Antimicrobial Peptide Calculator and Predictor (http://aps.unmc.edu/AP/prediction/prediction_main.php). Three dimensional (3D) molecular structures of N1 and its analogues were made using the PyMOL molecular modeling system (http://www.pymol.org/). All peptides with 90% purity were synthesized by the ChinaPeptides Co. Ltd. (Shanghai, China).
Antimicrobial activity
The antimicrobial activity of peptides was determined by the microbroth dilution method48. The bacterial strains were incubated in Mueller-Hinton (MH) medium at 37 °C overnight until the mid-log phase (OD600 nm reading of 0.5). Bacteria were diluted to 1 × 105 CFU/ml and added into 96-well plates, followed by the addition of serial dilutions of peptides (from 0.0625 to 128 μg/ml). The plates were incubated at 37 °C for 18–24 h. The MIC value was determined as the lowest peptide concentration at which no bacterial growth was observed. All tests were conducted in triplicate.
Hemolysis
The hemolytic activity of N2 and N6 was evaluated by determining the amount of hemoglobin that was released from fresh mouse erythrocytes29. The blood cells were washed three times with 10 mM PBS (pH 7.4) and centrifuged at 1,500 rpm for 10 min at room temperature. In brief, 100 μl of erythrocyte solution (8%, v/v) was mixed with 100 μl of a series of peptide solutions and incubated for 1 h at 37 °C. The mixture was then centrifuged at 1,500 rpm for 5 min, and the absorbance of supernatants was measured at 540 nm. Values of 0 and 100% hemolysis were determined in PBS and 0.1% Triton X-100, respectively. Three replicates were performed for each condition. Hemolysis (%) = [Abs540 nm of the treated sample − Abs540 nm of the negative control)/(Abs540 nm of positive control − Abs540 nm of negative control)] × 100.
Cytotoxicity
The colorimetric MTT assay was used to determine the effect of peptides on the viability of murine peritoneal RAW264.7 macrophage cells49. Cells (5 × 103 cells/well) were added into 96-well microtiter plates and incubated in a humidified 5% (v/v) CO2/air environment at 37 °C for 24 h. A series of peptide solutions were added and incubated for 48 h. Untreated cells were used as controls. The MTT solution was added into plates, incubated for 4 h, and then removed from plates. Dimethyl sulfoxide (DMSO) was added into plates and the absorbance was measured at 570 nm with a spectrophotometer. The rate of inhibition of cell proliferation was calculated using the following formula: Growth inhibition (%) = (Abs570 nm of control − Abs570 nm of treated sample)/Abs570 nm of control × 100.
Antimicrobial resistance
Antimicrobial resistance of N2 and N6 was measured by the sequential passaging method50. E. coli cultures (106 CFU/ml) were treated with N2 and N6 (from 0.25× to 8× MIC), respectively. At 24 h intervals, cultures from the second-highest concentrations (OD ≥ 0.5) were diluted 1:100 into fresh MH broth containing N2 or N6. This serial passaging was repeated daily for 18 days. Any cultures that grew at higher than the MIC levels were passaged on peptide-free MH agar plates and the MIC was then determined by broth microdilution. This was repeated three times.
Sensitivity of N2/N6 to temperature, pH, and proteases
To determine effects of temperature on antibacterial activity, N2 and N6 were incubated for 1 h at 4, 20, 40, 60, 80 and 100 °C in PBS. The pH stability of N2 and N6 was determined after 3 h incubation in 100 mM glycine-HCl buffer (pH 2.0), sodium acetate buffer (pH 4.0), sodium phosphate buffer (pH 6.0), Tris-HCl buffer (pH 8.0), or glycine-NaOH buffer (pH 10.0). In addition, N2 and N6 were incubated for 4 h at 37 °C in pepsin (3000 U/mg, pH 2.0), papain (500 U/mg, pH 6.0), and trypsin (250 U/mg, pH 8.0) (10:1, w/w) solutions, respectively. The antibacterial activity of peptides was estimated against E. coli CVCC195 and S. enteritidis CVCC3377 cells by a microdilution plate assay48. All tests were conducted in triplicate.
CD and NMR analysis of N2/N6
The secondary structure of peptides was investigated in ddH2O and SDS solutions by CD spectroscopy51. The CD spectra of N2 and N6 were measured via a MOS-450 spectropolarimeter (Bio-Logic, Grenoble, France) using a 1.0 mm path-length cuvette. Peptides were dissolved in ddH2O or SDS solutions (10–40 mM). The spectra of peptides were recorded from 180 to 260 nm at 25 °C at a scanning speed of 100 nm/min with a step resolution of 2.0 nm and an integration time of 2 s. Data were analyzed using CDNN software.
The structures of N1, N2 and N6 in 90% H2O/10% D2O were determined by solution state NMR. All NMR experiments were acquired at 298 K on an Agilent DD2 600 MHz spectrometer equipped with a cold-probe. For each peptide, approximately 5 mg of solid powder was dissolved in 500 µl aqueous solvent (10% D2O, 90% H2O, v/v) to make the NMR samples. A series of 1D and 2D spectra, including 1H, 1H-1H TOCSY, 1H-1H NOESY, 1H-13C HSQC, and 1H-15N HSQC, were acquired for the structure elucidation. Amide proton temperature coefficients were derived from four TOCSY experiments at different temperatures (293, 298, 303 and 308 K) for investigating intramolecular hydrogen bonds. All TOCSY experiments were carried out using an 80 ms MLEV-17 spin-lock with a field strength of 6839 Hz. The mixing time for each NOESY experiment was 150 ms. The spectra widths were 7396.4 * 7396.4 Hz for the 1H-1H, 7396.4 * 21124.9 Hz for the 1H-13C and 7396.4 * 1945.6 Hz for the 1H-15N correlation spectra. 256, 140 and 128 complex points were collected for the indirect proton, carbon and nitrogen dimensions, respectively, and 512 complex points were collected for the directly observing dimension. The chemical shifts were referenced to DSS at 0.00 ppm for the proton and indirectly for the carbon and nitrogen dimensions52. All NMR data were processed using NMRPipe and analyzed with the CcpNmr program suite53. Signal assignment was obtained by following the method developed by Wuthrich, and inter-proton distance restraints were derived from the NOESY spectrum. Structure calculation and NOE assignment were performed simultaneously by using the programs CNS and ARIA254, 55. Hydrogen bonding restraints were generated by analyzing the NOE patterns, backbone secondary chemical shifts as well as the amide proton temperature coefficients. Backbone dihedral angle restraints (Φ and Ψ angles) were derived using the program DANGLE incorporated in the CcpNmr suite56. A total of 100 structures were calculated, and the 20 structures with the lowest total energy were selected to perform a refinement procedure in water. The protein structure ensemble was displayed and analyzed with the software Pymol (http://www.pymol.org/).
Effects of N2/N6 on model membranes, cell membranes, and membrane potential
Dye leakage experiments from model membranes
Different lipids (Avanti Polar Linpids, Inc.) were used to prepare model membranes composed of POPC/ POPG (4:1 molar ratio) and POPC/cholesterol (1:1) by the extrusion method as previously described57. POPC/POPG and POPC/cholesterol LUVs were used to mimic gram-negative bacterial and eukaryotic membranes, respectively. Lipid suspensions (500 μM) loaded with calcein (50 μM) were incubated with peptides at 1×, 2×, and 4× MIC against E. coli cells or 2-fold dilution concentrations (0.25–256 μg/ml). After incubation for 5 min, the fluorescence (F) (λexcitation = 492 nm, λemission = 518 nm) was measured on a microplate reader. The calcein leakage released from LUVs treated with N2 or N6 was determined as % Leakage = (F − F0)/(Ft − F0) × 100, wherein Ft is fluorescence released from vesicles treated with 1% Triton X-100 and F0 is the fluorescence of each sample at T = 058.
Outer membrane permeability assay
The outer membrane permeabilization activity of N2 and N6 was determined using the fluorescent dye NPN assay59. Mid-log phase E. coli CVCC195 and S. enteritidis CVCC3377 cells were collected by centrifugation and washed twice, then suspended in HEPES buffer (pH 7.4) to an OD600 nm of 0.4. Cell suspensions and NPN solutions (10 μM) were added into 96-well black plates, followed by the addition of peptide solutions (1×, 2×, and 4× MIC). Fluorescence intensity was recorded until no further increase was observed with a microplate reader (excitation/emission, 328/438 nm). The cells treated with PMB and PBS were used as positive and negative controls, respectively.
Inner membrane permeability assay
Mid-log phase E. coli CVCC195 and S. enteritidis CVCC3377 cells (1 × 108 CFU/ml) were incubated with or without 1× MIC peptide solutions at 37 °C for 5, 30, and 120 min. After washing twice with PBS, cells were then incubated with 50 μg/ml PI for 15 min and analyzed using a FACS Calibur Flow Cytometer (BD, USA)46.
Membrane potential assay
The effect of peptides on bacterial membrane potentials was measured with the lipophilic fluorescent dye rhodamine-12360, 61. Mid-log phase E. coli CVCC195 and S. enteritidis CVCC3377 cells were harvested by centrifugation and resuspended in PBS (1 × 108 CFU/ml). Then, 1×, 2×, and 4× MIC N2 or N6 solutions were incubated for 2 h with the above cells at 37 °C. After washing twice with PBS, rhodamine-123 was added and incubated at room temperature for 10 min in the dark. Fluorescence intensity was analyzed using a flow cytometer. Untreated cells were used as a blank control.
Intracellular ROS accumulation
Intracellular ROS accumulation was measured using the fluorescent dye DHR-123. E. coli CVCC195 and S. enteritidis CVCC3377 cells (1 × 108 CFU/ml) were incubated for 2 h at 37 °C with 1×, 2×, and 4× MIC N2 or N6, washed with PBS and stained with DHR-123 (5 μg/ml). The fluorescence of the dyes was measured using a flow cytometer.
Electron microscopy
Mid-log phase E. coli CVCC195 and S. enteritidis CVCC3377 cells (1 × 108 CFU/ml) were incubated with 4× MIC N2 or N6 for 2 h at 37 °C. The bacterial cells were then centrifuged and washed three times with 0.1 M PBS (pH 7.2) and fixed overnight at 4 °C with 2.5% glutaraldehyde. After washing twice with PBS, the cells were post-fixed with 1% osmium tetroxide (OsO4) for 2 h, dehydrated in a graded ethanol series (from 50% to 100%) for 15 min each time, and dried by CO2. Gold-palladium was sputtered on samples and observed on a QUANTA200 SEM (FEI, Philips, Netherlands)58.
After fixation with 2.5% glutaraldehyde at 4 °C overnight, bacterial cells were post-fixed with 1% OsO4 for 1 h and washed three times with PBS. The cells were then dehydrated for 7 min each time with a graded acetone series (50%, 70%, 85%, 95% and 100%). The samples were immersed in mixtures of acetone and resin (3:1, 1:1 and 1:3, respectively) for 40 min each time and in a pure epoxy resin overnight in a dryer. The samples were then embedded in capsules containing embedding medium, polymerized at 45 °C for 3 h and at 65 °C for 24 h, respectively. Thin sections were prepared using an ultramicrotome and stained with 1% uranyl acetate and visualized by a JEM1400 (JEDL, Tokyo, Japan)59.
Effects of N2/N6 on bacterial genomic DNA migration, cell cycle and macromolecular synthesis
DNA gel migration assay
The genomic DNA of S. enteritidis was incubated for 10 min at room temperature with different concentrations of N2 and N6 in binding buffer (5% glycerol, 10 mM Tris-HCl, pH 8.0, 20 mM KCl, 1 mM EDTA, 1 mM dithiothreitol and 50 mg/ml BSA). The mass ratios of peptides and DNA were 0, 0.05, 0.1, 0.15, 0.2, 0.25, 0.3, and 0.4. The migration of DNA was assessed by electrophoresis on a 1% agarose gel58, 59.
Cell cycle analysis
Mid-log phase S. enteritidis CVCC3377 cells (1 × 108 CFU/ml) were incubated with 1× MIC at 37 °C for 5 min, 30 min and 120 min59. After centrifugation and washing twice with PBS, the cells were fixed in precooled ethanol overnight at 4 °C. The cells were then harvested, washed, and resuspended in PBS. After incubation with RNAse A (0.5 mg/ml) at room temperature for 15 min, the cells were stained with PI solution for 15 min and examined using a flow cytometer.
Macromolecular synthesis
Effects of N2 and N6 on DNA, RNA, protein and peptidoglycan synthesis were determined in S. enteritidis by measuring incorporation of radioactive precursors of L-[methyl-3H] thymidine, [5-3H] uridine, L-[3,4,5-3H] leucine, and D-[6-3H(N)] glucosamine hydrochloride50. Mid-log phase S. enteritidis CVCC3377 cells (105 CFU/ml) were incubated with 1× MIC N2, N6 or antibiotics at 37 °C for 15 min. Radioactive precursors were added into cells (1 μCi/ml) and shaken at 37 °C for 20 min. Ice-cold 25% trichloroacetic acid (TCA) was added and placed on ice for 30 min. The cells were centrifuged and washed twice with ice-cold 25% TCA. After drying, the samples were mixed with scintillation fluid and counted for their radioactivity using a MicroBeta 1450 scintillation counter (Perkin Elmer)62.
Docking of N2/N6 with LPS
The NMR structures of N1, N2 and N6 were docked onto LPS by the program AutoDock 4.263. Peptides were used as ligands and backbones were kept rigid, and almost all side chains were defined as flexible. LPS was defined as a receptor and was kept rigid. Grid maps representing LPS were constructed using 70 × 80 × 80 points, with a grid spacing of 0.375 Å and grid center of the H2 atom of the glucosamine II (GlcN II) residue in lipid A. Docking of peptides with LPS was generated by using a Lamarchian genetic algorithm (LGA) with a translation step of 0.2 Å, a quaternion step of 5°, and a torsion step of 5°. The maximum number of energy evaluations increased to 15,000,000. One hundred LGA docking runs were performed.
LPS binding affinity in vitro
The binding affinity of N2 and N6 to the lipid A region of LPS was determined by the fluorescence probe BC displacement method64. Equal volumes of 40 μg/ml LPS and 10 μM BC in 50 mM Tris buffer (pH 7.4) were mixed, added into 96-well plates and incubated in the dark for 4 h at room temperature. Different concentrations of N2, N6 or antibiotics were added and kept in the dark for 1 h at room temperature. Fluorescence was then measured using a microplate reader (excitation/emission, 580/620 nm). The occupancy rate was calculated by the following equation: BC displacement rate (%) = (1 − (F0 − F)/(F0 − Fmax)) × 100, wherein F0 is the fluorescence intensity of the solution of BC alone; Fmax is the fluorescence intensity in the presence of LPS at the saturation concentration, and F is the intensity of LPS:BC mixtures at varying displacer concentrations.
Efficacy of N2/N6 in vivo
Mice were purchased from the Beijing Vital River Laboratory Animal Technology Co. Ltd. and were housed under standard conditions and a 12:12 h light/dark cycle. All animal experiments were performed in accordance with the Animal Care and Use Committee of the Feed Research Institute of the Chinese Academy of Agricultural Sciences (CAAS), and protocols have been reviewed and approved by the Laboratory Animal Ethical Committee and its Inspection of the Feed Research Institute of CAAS (AEC-CAAS-20090609).
Bacteria-induced peritonitis models in mice
Six-week-old female ICR mice (approximately 20 g, six mice per group) were intraperitoneally injected with E. coli CVCC1515 (2.5 × 108 CFU/ml, 1 ml) and S. enteritidis CICC22956 (1 × 106 CFUF/ml, 1 ml) strains, respectively, followed by intraperitoneal injection with N2 and N6 (1.25, 2.5, 5, and 7.5 mg/kg of body weight, 0.2 ml) or PMB (0.31, 0.625, and 1.25 mg/kg of body weight, 0.2 ml) at 0.5 and 8 h. Mice injected with only bacteria or saline served as negative (untreated) or blank (unchallenged) controls, respectively. Survival of mice was recorded every 12 h and followed for 7 days.
LPS-induced endotoxemia model in mice
Eight-week-old SPF male C57BL/6 mice (20 g ± 5 g, six mice per group) were injected intraperitoneally with E. coli 0111:B4 LPS (30 mg/kg body weight, Sigma). The mice were injected intraperitoneally with 0.2 ml of N2, N6 (2.5 and 5 mg/kg) or PMB (10 and 15 mg/kg) at 0.5 h and 8 h after challenge with LPS. Mice injected with PBS were used as the negative control. The mouse survival rate was recorded for 7 days.
Nine mice per group were injected intraperitoneally with N2 and N6 (5 mg/kg) or PMB (15 mg/kg) 0.5 h after challenge with 10 mg/kg LPS. Blood was collected in a heparin sodium tube at 2 h or 8 h after LPS injection and incubated for 30 min at 37 °C and then overnight at 4 °C. After centrifugation at 3,000 rpm for 10 min at 4 °C, serum was collected from supernatant. The cytokine levels of TNF-α, IL-6 and IL-1β were detected by the Jiaxuan Biotech. Co. Ltd. (Beijing, China) using an enzyme linked immunosorbent assay (ELISA).
The mice were injected intraperitoneally with N2 and N6 (5 and 7.5 mg/kg) or PMB (15 mg/kg) at 0.5 h and 8 h after challenge with LPS (30 mg/kg). At 7 d after LPS injection, lungs were dissected out and removed, washed with PBS, and fixed in 4% paraformaldehyde at 4 °C for 24 h. The lung tissues were then washed with PBS, dehydrated with a graded ethanol series (75–95%), infiltrated with xylene, and embedded in paraffin wax. Sections were cut, stained with hematoxylin and eosin, and observed by a light microscope. Mice injected with only LPS or PBS served as a negative or blank controls.
Statistical analysis
All data are presented as the mean ± SEM (standard error of the mean). Comparisons among multiple groups were performed by one-way ANOVA and SPSS analysis. A p-value of <0.05 was considered statistically significant.
Electronic supplementary material
Acknowledgements
This study was financially supported by the National Natural Science Foundation of China (grants No. 31372346, No. 31672467, No. 31572444, and No. 31572445), the Project of the National Support Program for Science and Technology in China (No. 2013BAD10B02), the Special Fund for Agro- scientific Research in the Public Interest in China (grant No. 201403047), and the AMP Direction of National Innovation Program of Agricultural Science and Technology in CAAS (CAAS- ASTIP-2013- FRI-02).
Author Contributions
X.M.W., D.T. and J.H.W. conceived and designed the research. N.Y., R.Y.M., Y.H., and X.W. developed the CD, FACS and SEM methods, and interpreted the results. X.H.L. performed the NMR experiment. Z.Z.L. provided help and support with MIC and mice experiments. N.Y., X.H.L., X.M.W. and J.H.W. wrote the manuscript. All authors reviewed the manuscript.
Competing Interests
The authors declare that they have no competing interests.
Footnotes
Na Yang, Xuehui Liu and Da Teng contributed equally to this work.
Electronic supplementary material
Supplementary information accompanies this paper at doi:10.1038/s41598-017-03664-2
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Xiumin Wang, Email: wangxiumin@caas.cn.
Jianhua Wang, Email: wangjianhua@caas.cn.
References
- 1.Zhao T, Zhao P, Doyle MP. Inactivation of Salmonella and Escherichia coli O157:H7 on lettuce and poultry skin by combinations of levulinic acid and sodium dodecyl sulfate. J Food Prot. 2009;72:928–936. doi: 10.4315/0362-028X-72.5.928. [DOI] [PubMed] [Google Scholar]
- 2.Wu WH, et al. Aptasensors for rapid detection of Escherichia coli O157:H7 and Salmonella typhimurium. Nanoscale Res Lett. 2012;7:658. doi: 10.1186/1556-276X-7-658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Tang J, Wang L, Xi Y, Liu G. A three-year survey of the antimicrobial resistance of microorganisms at a Chinese hospital. Exp Ther Med. 2016;11:731–736. doi: 10.3892/etm.2016.2983. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Allocati N, Masulli M, Alexeyev MF, Di I. C. Escherichia coli in Europe: an overview. Int J Environ Res Public Health. 2013;10:6235–6254. doi: 10.3390/ijerph10126235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Qu M, et al. Prevalence and antibiotic resiance of bacterial pathogens isolated from childhood diarrhea in Beijing, China (2010–2014) Gut Pathog. 2016;8:31. doi: 10.1186/s13099-016-0116-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hurley JC. Antibiotic-induced release of endotoxin: a reappraisal. Clin Infect Dis. 1992;15:840–854. doi: 10.1093/clind/15.5.840. [DOI] [PubMed] [Google Scholar]
- 7.Langevelde P, Kwappenberg KM, Groeneveld PH, Mattie H, Dissel JT. Antibiotic-induced lipopolysaccharide (LPS) release from Salmonella typhi: delay between killing by ceftazidime and imipenem and release of LPS. Antimicrob Agents Chemother. 1998;42:739–743. doi: 10.1128/aac.42.4.739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Li P, Wohland T, Ho B, Ding JL. Perturbation of Lipopolysaccharide (LPS) Micelles by Sushi 3 (S3) antimicrobial peptide. The importance of an intermolecular disulfide bond in S3 dimer for binding, disruption, and neutralization of LPS. J Biol Chem. 2004;279:50150–50156. doi: 10.1074/jbc.M405606200. [DOI] [PubMed] [Google Scholar]
- 9.Płóciennikowska A, Hromada-Judycka A, Borzęcka K, Kwiatkowska K. Co-operation of TLR4 and raft proteins in LPS-induced pro-inflammatory signaling. Cell Mol Life Sci. 2015;72:557–581. doi: 10.1007/s00018-014-1762-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Mebazaa A, et al. Designing phase 3 sepsis trials: application of learned experiences from critical care trials in acute heart failure. J Intensive Care. 2016;4:24. doi: 10.1186/s40560-016-0151-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Maher S, McClean S. Investigation of the cytotoxicity of eukaryotic and prokaryotic antimicrobial peptides in intestinal epithelial cells in vitro. Biochem Pharmacol. 2006;71:1289–1298. doi: 10.1016/j.bcp.2006.01.012. [DOI] [PubMed] [Google Scholar]
- 12.Rotem S, Mor A. Antimicrobial peptidemimics for improved therapeutic properties. Biochim Biophys Acta. 2009;1788:1582–1592. doi: 10.1016/j.bbamem.2008.10.020. [DOI] [PubMed] [Google Scholar]
- 13.Walters SM, Dubey VS, Jeffrey NR, Dixon DR. Antibiotic-induced Porphyromonas gingivalis LPS release and inhibition of LPS-stimulated cytokines by antimicrobial peptides. Peptides. 2010;9:1649–1653. doi: 10.1016/j.peptides.2010.06.001. [DOI] [PubMed] [Google Scholar]
- 14.Dixon DR, Karimi-Naser L, Darveau RP, Leung KP. The anti-endotoxic effects of the KSL-W decapeptide on Escherichia coli O55:B5 and various oral lipopolysaccharides. J Periodontal Res. 2008;43:422–430. doi: 10.1111/j.1600-0765.2007.01067.x. [DOI] [PubMed] [Google Scholar]
- 15.Bedran TB, Mayer MP, Spolidorio DP, Grenier D. Synergistic anti-inflammatory activity of the antimicrobial peptides human beta-defensin-3 (hBD-3) and cathelicidin (LL-37) in a three-dimensional co-culture model of gingival epithelial cells and fibroblasts. PLoS One. 2014;9:e106766. doi: 10.1371/journal.pone.0106766. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Wu GQ, et al. Interaction of antimicrobial peptide s-thanatin with lipopolysaccharide in vitro and in an experimental mouse model of septic shock caused by a multidrug-resistant clinical isolate of Escherichia coli. Int J Antimicrob Agents. 2010;35:250–254. doi: 10.1016/j.ijantimicag.2009.11.009. [DOI] [PubMed] [Google Scholar]
- 17.Edwards IA, et al. Contribution of amphipathicity and hydrophobicity to the antimicrobial activity and cytotoxicity of β-hairpin peptides. ACS Infect Dis. 2016;2:442–450. doi: 10.1021/acsinfecdis.6b00045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Ramamoorthy A. Deletion of all cysteines in tachyplesin I abolishes hemolytic activity and retains antimicrobial activity and lipopolysaccharide selective binding. Biochemistry. 2006;45:6529–6540. doi: 10.1021/bi052629q. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Maemoto A, et al. Functional analysisof the alpha-defensin disulfide array in mouse cryptdin-4. J Biol Chem. 2004;279:44188–44196. doi: 10.1074/jbc.M406154200. [DOI] [PubMed] [Google Scholar]
- 20.Sharma H, Nagaraj R. Antimicrobial activity of human β-defensin 4 analogs: insights into the role of disulfide linkages in modulating activity. Peptides. 2012;38:255–265. doi: 10.1016/j.peptides.2012.08.024. [DOI] [PubMed] [Google Scholar]
- 21.Krishnakumari V, Singh S, Nagaraj R. Antibacterial activities of synthetic peptides corresponding to the carboxy-terminal region of human beta-defensins 1–3. Peptides. 2006;27:2607–2613. doi: 10.1016/j.peptides.2006.06.004. [DOI] [PubMed] [Google Scholar]
- 22.Hoover DM, Wu Z, Tucker K, Lu W, Lubkowski J. Antimicrobial characterization of human beta-defensin 3 derivatives. Antimicrob Agents Chemother. 2003;47:2804–2809. doi: 10.1128/AAC.47.9.2804-2809.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Ma B, et al. The disulfide bond of the peptide thanatin is dispensible for its antimicrobial activity in vivo and in vitro. Antimicrob Agents Chemother. 2016;60:4283–4289. doi: 10.1128/AAC.00041-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Mandal M, Nagaraj R. Antibacterial activities and conformations of synthetic alpha-defensin HNP-1 and analogs with one, two and three disulfide bridges. J Peptide Res. 2002;59:95. doi: 10.1034/j.1399-3011.2002.01945.x. [DOI] [PubMed] [Google Scholar]
- 25.Powers JP, Rozek A, Hancock RE. Structure-activity relationships for the beta-hairpin cationic antimicrobial peptide polyphemusin I. Biochim Biophys Acta. 2004;1698:239–250. doi: 10.1016/j.bbapap.2003.12.009. [DOI] [PubMed] [Google Scholar]
- 26.Lee JU, et al. Solution structures and biological functions of the antimicrobial peptide, arenicin-1, and its linear derivative. Biopolymers. 2007;88:208–216. doi: 10.1002/bip.20700. [DOI] [PubMed] [Google Scholar]
- 27.Harwig SS, et al. Intramolecular disulfide bonds enhance the antimicrobial and lytic activities of protegrins at physiological sodium chloride concentrations. Eur J Biochem. 1996;240:352–357. doi: 10.1111/j.1432-1033.1996.0352h.x. [DOI] [PubMed] [Google Scholar]
- 28.Fox JL. Antimicrobial peptides stage a comeback. Nat Biotechnol. 2013;31:379–382. doi: 10.1038/nbt.2572. [DOI] [PubMed] [Google Scholar]
- 29.Wang XJ, et al. Candidacidal mechanism of the arenicin-3-derived peptide NZ17074 from Arenia marina. Appl Microbiol Biotechnol. 2014;98:7387–7398. doi: 10.1007/s00253-014-5784-6. [DOI] [PubMed] [Google Scholar]
- 30.Wang, X. M. et al. 2016. Combined systems approaches reveal a multistage mode of action of a marine antimicrobial peptide against pathogenic Escherichia coli and its protective effect against bacterial peritonitis and endotoxemia. Antimicrob Agents Chermother61, e01056–16 (2017). [DOI] [PMC free article] [PubMed]
- 31.Hoegenhaug, H-H. K. et al. Antimicrobial peptide variants and polynucleotides encoding same. US 20110306750Al.
- 32.Breeuwer P, Abee T. Assessment of the membrane potential, intracellular pH and respiration of bacteria employing fluorescence techniques. Mol Microbial Ecol Manual. 2004;8:1563–1580. doi: 10.1007/978-1-4020-2177-0_8. [DOI] [Google Scholar]
- 33.David SA, Balasubramanian KA, Mathan VI, Balaram P. Analysis of the binding of polymyxin B to endotoxic lipid A and core glycolipid using a fluorescent displacement probe. Biochim Biophys Acta. 1992;1165:147–152. doi: 10.1016/0005-2760(92)90180-4. [DOI] [PubMed] [Google Scholar]
- 34.Rekdal Ø, Andersen J, Vorland LH, Svendsen JS. Construction and synthesis of lactoferricin derivatives with enhanced antibacterial activity. J Peptide Sci. 1999;5:32–45. doi: 10.1002/(SICI)1099-1387(199901)5:1<32::AID-PSC172>3.0.CO;2-9. [DOI] [Google Scholar]
- 35.Long CG, Li MH, Baum J, Brodsky B. Nuclear magnetic resonance and circular dichroism studies of a triple-helical peptide with a glycine substitution. J Mol Biol. 1992;225:1–4. doi: 10.1016/0022-2836(92)91020-P. [DOI] [PubMed] [Google Scholar]
- 36.Yang N, et al. Modification and characterization of a new recombinant marine antimicrobial peptide N2. Process Biochem. 2016;51:734–739. doi: 10.1016/j.procbio.2016.03.005. [DOI] [Google Scholar]
- 37.Andreu D, Rivas L. Animal antimicrobial peptides: an overview. Biopolymers. 1998;47:415–433. doi: 10.1002/(SICI)1097-0282(1998)47:6<415::AID-BIP2>3.0.CO;2-D. [DOI] [PubMed] [Google Scholar]
- 38.Oren Z, Hong J, Shai Y. A repertoire of novel antibacterial diastereomeric peptides with selective cytolytic activity. J Biol Chem. 1997;272:14643–14649. doi: 10.1074/jbc.272.23.14643. [DOI] [PubMed] [Google Scholar]
- 39.Vermeer LS, et al. Conformational flexibility determines selectivity and antibacterial, antiplasmodial, and anticancer potency of cationic α-helical peptides. J Biol Chem. 2012;287:34120–34133. doi: 10.1074/jbc.M112.359067. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Patrzykat A, Friedrich CL, Zhang L, Mendoza cV, Hancock RE. Sublethal concentrations of pleurocidin-derived antimicrobial peptides inhibit macromolecular synthesis in Escherichia coli. Antimicrob Agents Chemother. 2002;46:605–614. doi: 10.1128/AAC.46.3.605-614.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Liang JF, Kim SC. Not only the nature of peptide but also the characteristics of cell membrane determine the antimicrobial mechanism of a peptide. J Pept Res. 1999;53:518–522. doi: 10.1034/j.1399-3011.1999.00051.x. [DOI] [PubMed] [Google Scholar]
- 42.Scheffers DJ, Pinho MG. Bacterial cell wall synthesis: new insights from localization studies. Microbiol Mol Biol Rev. 2005;69:585–607. doi: 10.1128/MMBR.69.4.585-607.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Taniguchi M, et al. Pyrrhocoricin, a proline-rich antimicrobial peptide derived from insect, inhibits the translation process in the cell-free Escherichia coli protein synthesis system. J Biosci Bioeng. 2016;121:591–598. doi: 10.1016/j.jbiosc.2015.09.002. [DOI] [PubMed] [Google Scholar]
- 44.Roy RN, Lomakin IB, Gagnon MG, Steitz TA. The mechanism of inhibition of protein synthesis by the proline-rich peptide oncocin. Nat Struct Mol Biol. 2015;22:466–469. doi: 10.1038/nsmb.3031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Jackson JJ, Kropp H. Beta-Lactam antibiotic-induced release of free endotoxin: in vitro comparison of penicillin-binding protein (PBP) 2-specific imipenem and PBP 3-specific ceftazidime. J Infect Dis. 1992;165:1033–1041. doi: 10.1093/infdis/165.6.1033. [DOI] [PubMed] [Google Scholar]
- 46.Bone RC. The pathogenesis of sepsis. Ann Intern Med. 1991;115:457–469. doi: 10.7326/0003-4819-115-6-457. [DOI] [PubMed] [Google Scholar]
- 47.Michie HR. Detection of circulating tumor necrosis factor after endotoxin administration. N Engl J Med. 1988;318:1481–1486. doi: 10.1056/NEJM198806093182301. [DOI] [PubMed] [Google Scholar]
- 48.Zhang Y, et al. High expression of a plectasin-derived peptide NZ2114 in Pichia pastoris and its pharmacodynamics, postantibiotic and synergy against Staphylococcus aureus. Appl Microbiol Biotechnol. 2014;98:681–694. doi: 10.1007/s00253-013-4881-2. [DOI] [PubMed] [Google Scholar]
- 49.Jiao J, et al. GAP-initiated constitutive expression of a novel plectasin-derived peptide MP1106 by Pichia pastoris and its activity against Streptococcus suis. Process Biochem. 2015;50:253–261. doi: 10.1016/j.procbio.2014.12.019. [DOI] [Google Scholar]
- 50.Ling LL, et al. A new antibiotic kills pathogens without detectable resistance. Nature. 2015;517:455–459. doi: 10.1038/nature14098. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Pavia KE, Spinella SA, Elmore DE. Novel histone-derived antimicrobial peptides use different antimicrobial mechanisms. Biochim Biophys Acta. 2012;1818:869–876. doi: 10.1016/j.bbamem.2011.12.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Markley JL, et al. Recommendations for the presentation of NMR structures of proteins and nucleic acids–IUPAC-IUBMB-IUPAB Inter-Union Task Group on the standardization of data bases of protein and nucleic acid structures determined by NMR spectroscopy. Eur J Biochem. 1998;256:1–15. doi: 10.1046/j.1432-1327.1998.2560001.x. [DOI] [PubMed] [Google Scholar]
- 53.Vranken WF, et al. The CCPN data model for NMR spectroscopy: development of a software pipeline. Proteins. 2005;59:687–696. doi: 10.1002/prot.20449. [DOI] [PubMed] [Google Scholar]
- 54.Brunger AT. Version 1.2 of the Crystallography and NMR system. Nat Protoc. 2007;2:2728–2733. doi: 10.1038/nprot.2007.406. [DOI] [PubMed] [Google Scholar]
- 55.Rieping W, et al. ARIA2: automated NOE assignment and data integration in NMR structure calculation. Bioinformatics. 2007;23:381–382. doi: 10.1093/bioinformatics/btl589. [DOI] [PubMed] [Google Scholar]
- 56.Cheung MS, Maguire ML, Stevens TJ, Broadhurst RW. DANGLE: A Bayesian inferential method for predicting protein backbone dihedral angles and secondary structure. J Magn Reson. 2010;202:223–233. doi: 10.1016/j.jmr.2009.11.008. [DOI] [PubMed] [Google Scholar]
- 57.Torcato IM, et al. The antimicrobial activity of Sub3 is dependent on membrane binding and cell-penetrating ability. Chembiochem. 2013;14:2013–2022. doi: 10.1002/cbic.201300274. [DOI] [PubMed] [Google Scholar]
- 58.Alfred RL, Palombo EA, Panozzo JF, Bhave M. The antimicrobial domains of wheat puroindolines are cell-penetrating peptides with possible intracellular mechanisms of action. PLoS One. 2013;8:e75488. doi: 10.1371/journal.pone.0075488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Teng D, et al. A dual mechanism involved in membrane and nucleic acid disruption of AvBD103b, a new avian defensin from the king penguin, against Salmonella enteritidis CVCC3377. Appl Microbiol Biotechnol. 2014;98:8313–8325. doi: 10.1007/s00253-014-5898-x. [DOI] [PubMed] [Google Scholar]
- 60.Warnes SL, Caves V, Keevil CW. Mechanism of copper surface toxicity in Escherichia coli O157: H7 and Salmonella involves immediate membrane depolarization followed by slower rate of DNA destruction which differs from that observed for Gram-positive bacteria. Environ Microbiol. 2012;14:1730–1743. doi: 10.1111/j.1462-2920.2011.02677.x. [DOI] [PubMed] [Google Scholar]
- 61.Hakansson AP, Roche-Hakansson H, Mossberg AK, Svanborg C. Apoptosis-like death in bacteria induced by HAMLET, a human milk lipid-protein complex. PLoS One. 2011;6:e17717. doi: 10.1371/journal.pone.0017717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Xiong YQ, Bayer AS, Yeaman MR. Inhibition of intracellular macromolecular synthesis in Staphylococcus aureus by thrombin-induced platelet microbicidal proteins. J Infect Dis. 2002;185:348–356. doi: 10.1086/338514. [DOI] [PubMed] [Google Scholar]
- 63.Kushibiki T, et al. Interaction between tachyplesin I, an antimicrobial peptide derived from horseshoe crab, and lipopolysaccharide. Biochim Biophys Acta. 2014;1844:527–534. doi: 10.1016/j.bbapap.2013.12.017. [DOI] [PubMed] [Google Scholar]
- 64.Sautrey G, et al. New amphiphilic neamine derivatives active against resistant Pseudomonas aeruginosa and their interactions with lipopolysaccharides. Antimicrob Agents Chemother. 2014;58:4420–4430. doi: 10.1128/AAC.02536-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.