

Summary
Embryonic stem cells (ESCs) are characterized by the pluripotent capacity to generate all embryonic lineages. Here, we show that ESCs can occupy a spectrum of distinct transcriptional and epigenetic states in response to varied extrinsic conditions. This spectrum broadly corresponds to a developmental continuum of pluripotency and is coupled with a gradient of increasing global DNA methylation. Each pluripotent state is linked with activation of distinct classes of transposable elements (TEs), which in turn influence ESCs through generating chimeric transcripts. Moreover, varied ESC culture parameters differentially license heterogeneous activation of master lineage regulators, including Sox1, Gata4, or Blimp1, and influence differentiation. Activation of Blimp1 is prevalent in 2i (without LIF) conditions, and marks a dynamic primordial germ cell (PGC)-like sub-state that is directly repressed by Klf4 downstream of LIF/STAT3 signaling. Thus, extrinsic cues establish a spectrum of pluripotent states, in part by modulating sub-populations, as well as directing the transcriptome, epigenome, and TE.
Keywords: pluripotency, chimeric transcripts, DNA methylation, PGC, transposable element, STAT3, embryonic stem cell, imprints, KLF4, heterogeneity
Graphical Abstract
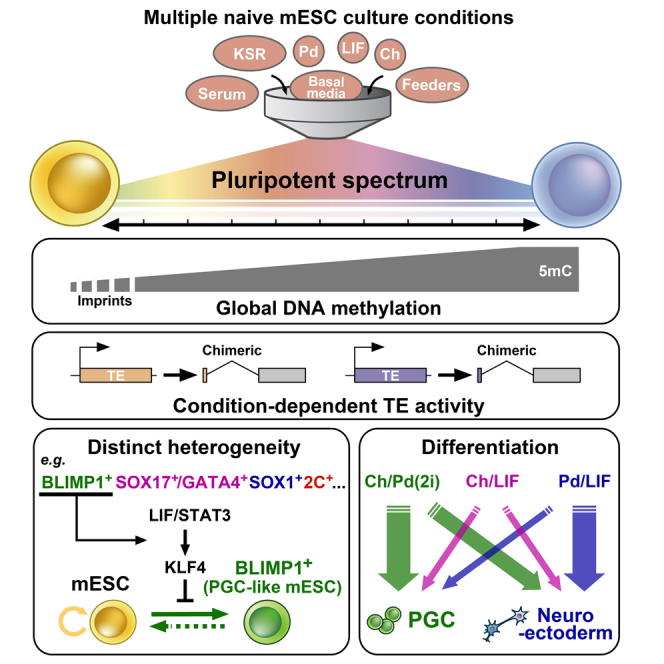
Highlights
-
•
Diverse culture parameters establish a spectrum of ESC pluripotency
-
•
Activation of distinct transposable elements in each pluripotent state
-
•
ESC conditions influence DNA methylation and response to differentiation cues
-
•
Distinct heterogeneities including a PGC-like state regulated by LIF and KLF4
In this article Surani and colleagues report a spectrum of pluripotent ESC states and characterize the distinct epigenomes and transcriptomes. They find each pluripotent condition is linked with specific transposable elements, influences differentiation, and supports distinct ESC sub-populations. They further trace the signaling principle which regulates one sub-population (a PGC-like state) to show it is modulated by KLF4 downstream of LIF/STAT3.
Introduction
Pluripotency is a transient state established during mammalian preimplantation development, and is characterized by the capacity to give rise to all fetal lineages. The pluripotent state can be propagated indefinitely through derivation of embryonic stem cells (ESCs) or via reprogramming strategies (De Los Angeles et al., 2015). Mouse ESCs are considered to exhibit naive pluripotency, which reflects their ability to contribute to all embryonic lineages upon re-introduction into a blastocyst, as well as other key hallmarks such as a derestricted epigenome and two active X chromosomes in female cells (Hackett and Surani, 2014). The preservation of naive pluripotency in ESCs is underpinned by expression of a network of auto-regulatory transcription factors, including Oct4, Sox2, Nanog, and Tfcp2l1, which are themselves sustained by extrinsic signaling cues (Dunn et al., 2014). An alternative, and possibly more developmentally advanced state of pluripotency, can also be propagated, as exemplified by epiblast stem cells (EpiSCs) and human ESCs (hESCs) in conventional culture conditions. These are typically classified as being in a primed pluripotent state that is poised to initiate lineage decisions (Nichols and Smith, 2009).
A broader array of pluripotent states is, however, likely. For example, culture using inhibitors of MAPK signaling and GSK3β (termed 2i) together with LIF (2i/L), render mouse ESCs in a relatively uniform naive state that is molecularly and epigenetically distinct from ESCs in conventional serum/LIF medium (Ying et al., 2008, Marks et al., 2012, Leitch et al., 2013). Any two of the 2i/L components in various combinations also yield naive ESCs, which may occupy distinct phases of pluripotency (Wray et al., 2010). Multiple other pluripotent conformations could also arise depending on the derivation strategy, available metabolites, and the precise signaling regime supplied, which may reflect distinct spatial or temporal identities (Blaschke et al., 2013, Tonge et al., 2014, Wu et al., 2015, Huang et al., 2014, Irie et al., 2015, Weinberger et al., 2016). Collectively, this suggests that a naive/primed duality model may not capture the broad complexity of pluripotency in vitro and possibly in vivo. Instead a graded spectrum of pluripotent states may emerge that exhibit distinct molecular and functional properties (Hackett and Surani, 2014, Wu and Izpisua Belmonte, 2015).
Heterogeneity in pluripotent stem cell populations is also prevalent, in part driven by dynamic sub-states. ESCs in conventional serum/LIF conditions interconvert between several configurations that include naive and primed sub-populations (Torres-Padilla and Chambers, 2014, Klein et al., 2015). Naive ESCs cultured in 2i/LIF are also apparently composed of sub-populations (Kolodziejczyk et al., 2015, Morgani et al., 2013), while EpiSCs and hESCs are also known to be highly heterogeneous (Cahan and Daley, 2013). This implies that dynamic heterogeneity may be a fundamental feature of pluripotent stem cells. Indeed, mouse ESCs continually transit through a Zscan4+ sub-state that promotes transient DNA demethylation and telomere rejuvenation, with the latter being essential for sustained pluripotent viability (Zalzman et al., 2010, Eckersley-Maslin et al., 2016). Passing through sub-states can therefore imprint a significant memory, implying that the prevalence of sub-populations could have broad functional implications for the whole ESC population.
It has recently emerged that successive stages of early mammalian development are linked with expression of distinct classes of transposable element (TE) (Goke et al., 2015). For example, MERVL elements and their cognate MT2 LTR become active specifically at the 2-cell (2C) stage in mice, while HERVK is active from the 8-cell stage in human embryos (Peaston et al., 2004, Grow et al., 2015). These elements can significantly influence expression of nearby genes. For example, TE can act as co-opted promoters that splice to downstream genes thereby generating “chimeric transcripts” (Macfarlan et al., 2012). In addition, TEs can affect gene expression through promoting open chromatin configurations, production of long noncoding RNAs (lncRNA), or by acting as enhancers (Thompson et al., 2016). The impact of TEs may be particularly prevalent in pluripotent cells, since TEs are under selective pressure to be active in pluripotent or germline phases in order to propagate transgenerationally (Bourque et al., 2008). Indeed, HERVH elements have a key role in contributing to the pluripotency network in hESCs (Wang et al., 2014). Thus, TEs represent a relatively unexplored regulatory source for the establishment and control of alternate pluripotent states.
Here, we identify a spectrum of ESC states and characterize the distinct transcriptional networks and epigenome at each node. Distinct classes of TEs are active between pluripotent conformations and influence the emergent transcriptome. Strikingly, some naive culture conditions license dynamic activation of master regulators for a specific primary germ layer (endoderm or ectoderm) or primordial germ cells (PGCs). Mechanistically, we identify Klf4 and LIF/STAT3 as the key regulators of a Blimp1+ PGC-like ESC state. Overall we report that ESC populations can occupy a continuum of transcriptional states, in part through accessing sub-states under certain culture parameters.
Results
Distinct Transcriptional States of Pluripotency
We initially sought to define the transcriptional variation across a broad range of pluripotent ESC states, with the expectation that any underlying differences may influence differentiation, particularly toward PGC fate. We selected nine culture parameters capable of supporting naive pluripotency, as judged by chimera contribution, and transited male (XY) low-passage (<p13) 129X1/SvJ (129) or C57BL/6J (B6) ESCs into each condition for ≥5 passages. The culture parameters were designed to test the influence of multiple extrinsic factors including combinations of GSK3β inhibition/WNT activation (CHIR99021, hereafter Ch), MEK inhibition (PD0325901, hereafter Pd), STAT3 activity (LIF), BMP/undefined signaling (serum), vitamin C/undefined signaling (knockout serum replacement, hereafter KSR), basal medium, feeders, and genetic background, on the overall state of pluripotency (Figure 1A). We observed no overt karyotype alterations during transition to each condition, as judged by indicator chromosomes (Figure S1A; D'Hulst et al., 2013).
Figure 1.

A Transcriptional Spectrum of Pluripotent States
(A) ESC culture conditions assayed in this study.
(B) Heatmap showing expression of key pluripotency genes by RNA-seq.
(C) Global transcriptional correlation between all genetic backgrounds and culture parameters.
(D) Principal-component analysis (PCA) showing the relationship between pluripotent ESC states.
(E) Loadings for PCA from (D) showing representative highly weighted genes that drive separation along each component. The histogram shows the distribution of all gene weightings.
(F) PCA analysis showing in vivo pluripotent stages and the full ESC spectrum (left) or defined ESC states separated by genetic background (right).
(G) Fluorescence-activated cell sorting (FACS) plot showing the response of Stella-GFP and MERVL-tdTomato (2C) ESCs in 2i/L to serum.
RNA sequencing (RNA-seq) revealed that ESCs in each condition expressed high levels of key pluripotency genes Oct4, Sox2, and Nanog, supporting their pluripotent status (Figure 1B). Some naive pluripotency genes varied among culture parameters, however, presumably reflecting the presence or absence of their direct signaling regulators. For example, Klf4 is downregulated in 2i conditions that lack LIF, consistent with Klf4 being a direct LIF/STAT3 target (Hall et al., 2009), while Tfcp2l1 is repressed in Pd/L, likely owing to attenuated WNT transduction (Ye et al., 2013). Pluripotent markers Stella and Rex1 primarily exhibited reciprocal background-dependent expression; with Rex1 elevated in 129 ESCs and Stella upregulated in B6 ESCs (Figure 1B). In general, ESCs in conventional serum/LIF (S/L) conditions expressed lower overall levels of naive markers, particularly when feeder free. Thus, while the broad network of naive pluripotency genes is comparable among ESC conditions, the precise transcriptional levels of each component vary downstream of culture parameters and genetic background.
The correlation of global gene expression patterns revealed two broad clusters; a response to combinations of 2i/L components, or ESCs under S/L and Pd/L conditions (Figures 1C and S1B). We found 3,048 genes exhibited robust differential expression (Log2(FC) > 2; adjusted p < 0.01) in pairwise comparisons between all states, and these assembled into distinct co-expression clusters associated with specific gene ontologies (Figure S1C). In addition, 136 and 82 genes are robustly linked with 129- or B6-specific expression, respectively, which we confirmed using multiple independent 129 and B6 ESC lines (Figure S2A).
To further investigate the relationship between each pluripotent condition we applied principal-component analysis (PCA). Strikingly, we observed a clear separation of ESC states along the first component (PC1) that appeared to reflect an ordering of developmental progression; ranging from 2i/L (with or without serum/KSR) through to feeder-free S/L (Figures 1D and S1D). By contrast PC2 separated ESCs according to genetic background (129 versus B6), while PC3 apparently segregated ESCs cultured in the presence or absence of serum/KSR, irrespective of other signaling influences (Figure 1D). To validate these interpretations, we examined the gene loadings along each principal component to determine the key driver genes that separate ESC states. Notably, negative PC1 values were strongly weighted by naive-associated genes, and in particular Tfcp2l1 and Prdm14. In contrast, positive PC1 scores were driven by primed and early developmental genes including Fgf5, Otx2, and Lefty1 (Figure 1E). This is consistent with separation along PC1 reflecting a spectrum of pluripotent ESC states that range from most naive to most developmentally progressed, with conditions such as Ch/L establishing intermediary states of naive pluripotency.
To investigate how this apparent pluripotent spectrum relates to embryonic ontogeny, we compared the ESC transcriptomes with publically available in vivo developmental stages (Boroviak et al., 2015). The first component here reflected technical differences but the analysis still recapitulated the continuum of ESC states, and, strikingly, they broadly correspond to developmental progression of the pluripotent epiblast lineage (Figure 1F). Specifically, the most naive ESC states cluster closest with embryonic day 3.5 (E3.5) inner cell mass, whereas ESC states predicted to be further along the pluripotent spectrum become progressively more comparable with E4.5 and, to some degree, E5.5 epiblasts (Figure 1F).
We next examined the other principal components that separate ESC states and observed that gene loading along PC2, which delineates genetic background, exhibited no significant gene class enrichment, albeit some neuroectodermal genes such as Olig1 are linked with B6 ESCs (Figure 1E). This implies that, although background has significant influence on the precise ESC transcriptome, no specific gene category predominates. Finally, gene loadings for PC3 revealed strong weighting for BMP targets Id1 and Id2, along with mesendoderm genes, consistent with separation of ESC transcriptomes according to the presence of BMP-rich serum or KSR. However notably, the most significant PC3 weightings (nine out of ten top genes) correspond to genes preferentially expressed in 2-cell embryos, including Zscan4 and Tcstv3, suggesting the 2C program is the single most affected pathway downstream of serum/KSR supplementation (Figure 1E).
To further investigate this and to test the PCA predictions we generated ESCs carrying dual reporters for 2C gene activity and Stella, which are expected to respond reciprocally to serum (see PC3) (Figure 1E). Consistently, we found that addition of serum to 2i/L reduced the fraction of Stella-positive ESCs by 4.0-fold, while concurrently increasing the fraction of 2C-positive ESCs by 3.7-fold (Figures 1G and S1E). In summary, we identify a spectrum of multiple naive pluripotent states that emerge in response to distinct combinations of extrinsic signaling cues. This spectrum appears to correspond to a developmental progression of pluripotency. The emergent ESC states are further dispersed depending on genetic background and/or additional supplements, such as serum or KSR. Moreover, the spectrum may in part reflect different sub-population identities and frequencies between culture parameters, as exemplified by 2C and Stella-positive ESCs.
Retrotransposon Activation Is Linked with Pluripotent State
Stage-specific activation of distinct TEs has been observed during successive stages of early embryonic development (Goke et al., 2015). To investigate whether each state along the ESC pluripotent spectrum is also linked with a specific signature of TE activity, we examined differentially expressed repeat families. We found 64 significantly altered families (total n = 1,110) in pairwise comparisons, of which 51 belonged to the LTR class of retrotransposons, including MERVL, MT2, and IAPLTR3 (Figures 2A and S2B). Several TE families are preferentially active in a specific ESC condition, for example LTR9 and L1M3d in Ch/L and S/L_F, respectively. Interestingly, PCA analysis of TE expression ordered the samples into a highly comparable arrangement as gene-based PCA, apparently recapitulating the pluripotent continuum (Figure 2B).
Figure 2.

Transposable Element Activation across the Pluripotent Spectrum
(A) Relative proportion of differentially expressed transposable element (TE) families between pairwise comparisons of all conditions.
(B) PCA showing the relationship between pluripotent states based on differential TE activation.
(C) Expression of selected LTR and LINE elements across the pluripotent continuum.
(D) Expression of the lncRNA Miat.
(E–G) Genome view showing RNA-seq tracks of detected chimeric transcripts that originate at an upstream LTR and splice to (E) Tcstv3, (F) Cyp2b23, and (G) Ash2l.
(H) Boxplots showing the median global DNA methylation (5-mC) level by bisulfite sequencing across various genomic features in the indicated culture condition and genetic background.
(I) Heatmap showing DNA methylation level at genomic imprints.
Examining loading of retrotransposons in the PCA revealed that families including IAPEy-int and RLTR45 strongly weighted the most naive-like ESC states (2i/L, 2i), whereas higher expression of MER50B and the LINE family L1M6B was linked with more developmentally progressed pluripotent identities (Figure 2C). The strong weighting of IAPEy-int is noteworthy as this family shares closest homology with HERVK elements, which are implicated in influencing human pluripotent cells (Grow et al., 2015). We additionally found 70 differentially expressed lncRNA between ESC conformations, many of which originate at TEs (Figure S2C). Among these the lncRNA Miat, which is involved in a feedback loop with pluripotency-associated factors (Sheik Mohamed et al., 2010), is progressively upregulated in each pluripotent state along the spectrum (Figure 2D).
Next, we asked whether activation of distinct sets of TE is directly linked with altered gene expression. We initially noted that between defined conditions (serum-free), 2i ESCs preferentially activated MT2 and MERVL elements, as well as many 2C-associated genes that can derive from such repeats as chimeric transcripts. Among these, expression of the telomere regulators Tcstv3 and Zscan4, and the metabolism gene Arg2, is highly elevated in 2i. Analyzing spliced RNA-seq reads revealed that gene upregulation in 2i is often a direct consequence of activation of upstream MERVL and MT2 elements, which initiate chimeric transcripts (Figure 2E).
Using our RNA-seq datasets to detect additional spliced junctions, we identified a further 637 chimeric transcripts across the pluripotent conditions, which derive from activated TEs upstream of the first annotated exon. Many of these are active or repressed in only a subset of pluripotent states. For example an LTRIS2 element upstream of Phf11d is responsible for a chimeric transcript, but this TE is preferentially repressed in Pd/L ESCs. As a consequence Phf11d mRNA is significantly downregulated in the Pd/L pluripotent conformation (Figure S3A). Similarly, activation of adjacent EtnERV2, RMER16, and PD1D10 elements upon KSR addition drives strong expression of the downstream Cyp2b23 gene (>30-fold transcriptional upregulation) specifically in 2i/L_K ESCs (Figure 2F). The potential significance of such TE-based regulatory mechanisms is underscored by our observation that a chimeric transcript derived from an RLTR12C element drives primary expression of the essential pluripotency gene Ash2l in ESCs (Wan et al., 2012; Figure 2G). Other key genes including Grb2 and Zfp640 also appear to be at least partly expressed from an upstream TE (data not shown). Taken together our data imply that each state of naive pluripotency is associated with a distinct repertoire of transcriptionally active retrotransposons, particularly among the LTR class. In some cases this is directly responsible for modulating gene transcription through chimeric transcripts. Notably, this links altered retrotransposon activity and overall ESC state.
DNA Methylation across the Pluripotent Spectrum
Global DNA hypomethylation is intimately associated with naive pluripotency, whereas increasing DNA methylation (5-methylcytosine, 5-mC) levels are generally linked with primed pluripotency and lineage commitment (Leitch et al., 2013). We used shallow-coverage whole-genome bisulfite sequencing to investigate 5-mC across the pluripotent spectrum. Genome-wide 5-mC levels were lowest in 2i/L (18%–37%) and most elevated in S/L_KD (78%–83%) ESCs, consistent with previous reports (Habibi et al., 2013, Ficz et al., 2013, Hackett et al., 2013a). However rather than falling into binary hypo- or hypermethylated status, other pluripotent states exhibited a gradient of progressive global 5-mC, with Pd/L (53%–61%) and Ch/L (69%–75%) associated with intermediate DNA methylation levels, for example (Figure 2H). Thus, similar to the continuum of transcriptional states, ESCs acquire a spectrum of epigenetic states.
The trend of DNA methylation was preserved across distinct genomic features such as promoters, enhancers, and repetitive elements (Figures 2H and S3B). An exception, however, is genomic imprints. These were relatively stable across most conditions (particularly in 129 background), but exhibited atrophy in 2i/L and erasure in 2i/L+KSR culture (Figure 2I). This latter observation may reflect the presence of vitamin C in KSR, which directly enhances TET activity, previously linked with erasure of imprints (Yamaguchi et al., 2013, Hackett et al., 2013b, Blaschke et al., 2013, Piccolo et al., 2013). In summary, the 5-mC epigenome is established across a wide spectrum in ESCs, ranging from hypo- to hypermethylated, with the precise level correlated to the developmental progression of the underlying pluripotent transcriptome and, to some extent, the presence of MEK inhibitor (Pd). The transcriptional level of genes known to influence DNA methylation levels, such as Dnmt3a, Uhrf1, Tet1, and Prdm14, did not definitively correlate with global 5-mC levels, however (Figure S3C). This may indicate that the overall epigenetic state in ESCs is regulated by a complex interplay between gene expression, available metabolites, and post-transcriptional control, such as recently shown for UHRF1 (von Meyenn et al., 2016).
Naive ESC Populations Exhibit Lineage-Associated Co-activation
To investigate the pluripotent spectrum in more detail, we selected four defined (serum-free) conditions (2i/L, 2i, Ch/L, and Pd/L) associated with a clear naive signature and robust chimera contribution (Dunn et al., 2014). We confirmed naive status by using the ΔPE Oct4-GFP reporter, which marks activation of the naive-specific distal Oct4 enhancer in ESCs. All states exhibited a single peak of GFP, implying relatively uniform naiveté among these populations (Figure 3A). To examine any underlying molecular differences between the naive states, we identified 1,056 differentially expressed genes (Log2(FC) > 2; p < 0.01) in all pairwise comparisons. These genes were primarily linked with differential metabolic processes, but surprisingly also included several markers of alternate lineage fates. Indeed, gene ontology analysis of genes upregulated uniquely in only one condition suggested activation of divergent germ layer programs, such as neurogenesis-related processes in Pd/L (Figure 3B).
Figure 3.

Distinct Naive ESC Conditions Activate Master Lineage Regulators
(A) FACS analysis showing near-uniform activation of the ΔPE Oct4-GFP naive pluripotency reporter in ESCs under defined conditions.
(B) Heatmap of differentially expressed genes in pairwise comparisons between naive ESC states. Selected genes from pathways enriched by gene ontology are shown.
(C) Heatmap showing expression of master regulator genes for each of the three primary germ layers and the germline (PGC).
(D) Immunofluorescence staining for master regulators of the germline (BLIMP1), endoderm (SOX17), and neuroectoderm (PAX6) in defined ESC conditions. Scale bars, 50 μm.
(E) FACS plots showing Sox1-GFP activation upon neural induction from indicated ESC conditions.
(F) qRT-PCR showing expression of indicated genes in ΔPE Oct4-GFP ESCs and upon induction of epiblast-like cells (EpiLCs) for 2 days. n = 3 biological replicates each assayed in technical triplicate; error bars, SEM.
(G) Representative examples of primordial germ cell (PGC)-like cell (PGCLC) induction from ancestral ESC conditions. Shown is the percentage of PGCLCs as determined by FACS. Scale bars, 200 μm.
To explore this further we inspected expression of master regulators for the three primary germ layers; endoderm, mesoderm, and ectoderm, as well as the germline, among the four naive states. Remarkably, we found that pluripotent ESCs in Pd/L strongly and specifically co-expressed multiple master (neuro)ectoderm genes such as Sox1, Pax6, and NeuroD1, but not markers of other lineages, in both 129 and B6 backgrounds (Figure 3C). By contrast, Ch/L ESCs upregulated master regulators for endoderm specification including Sox17, Gata4, and Gata6, as well as definitive endoderm markers FoxA2 and Hnf1b. Finally, 2i conditions exhibited a clear PGC signature, uniquely co-activating the three key PGC specification genes Blimp1, Prdm14, and Ap2γ, and some primitive streak/mesodermal genes such as Mixl1. Unsupervised hierarchical clustering further revealed that 2i, Ch/L, and Pd/L each segregate separately from all other pluripotent conditions based only on expression of germline, endoderm, and neuroectoderm master regulators, respectively (Figure S4A). This highlights their exclusive overall state with respect to lineage-associated expression. Notably, however, master primary germ layer genes were near undetectable in 2i/L ESCs (Figure 3C).
We next sought confirmation for activation at the protein level by immunostaining. This revealed robust detection of the key germline-determinant BLIMP1 in 2i, endoderm regulators SOX17 and GATA4 in Ch/L, and neuroectoderm regulators PAX6 and OTX2 in Pd/L, but near-undetectable levels in reciprocal conditions (Figures 3D and S4B). NANOG was strongly detected under all parameters. In general, lineage-associated expression was heterogeneous, possibly reflecting dynamic expression. Using the Fucci reporter, we also noted altered cell-cycle dynamics between pluripotent states, which can influence lineage-bias in stem cells (data not shown) (Pauklin and Vallier, 2013). Overall, the tested pluripotent parameters appear to differentially license activation of lineage-specific master regulators, with a subset of ESCs shuttling into positive state.
To investigate whether lineage-associated expression influences cell fate, we seeded ESCs in N2B27 medium without cytokines, which preferentially induces neuroectoderm specification, but is also permissive for other lineages (Ying et al., 2003). Using Sox1-GFP ESCs that report on acquisition of neuroectoderm fate, we observed that cells from the ancestral Pd/L condition activated GFP both earlier and with greater maximal efficiency (>80%) than ESCs from other initial parameters. Moreover Ch/L maintained ESCs, which exhibited some activation of endoderm regulators, were relatively resistant to acquiring a Sox1-GFP+ fate (18%–31%) (Figure 3E).
We next asked whether an alternative strategy of differentiation is also influenced by initial pluripotent state by inducing PGC fate. PGC specification proceeds through induction of naive ESCs into epiblast-like cells (EpiLCs), which closely parallel post-implantation epiblast cells, and subsequently specification of PGC-like cells (PGCLCs) in embryoids (Hayashi et al., 2011). All examined ESC states formed morphologically equivalent EpiLCs, which expressed comparable levels of “primed” markers such as Fgf5 and Dnmt3b, while concurrently repressing naive genes including Nanog and Klf4 (Figures 3F, S5A, and S5B). The differential expression of most lineage regulators also equalized during EpiLC induction. Nonetheless, we observed a marked difference in PGCLC specification from EpiLCs depending on the predecessor condition of ESCs.
Specifically, PGCLCs were specified with high efficiency from ancestral 2i/L ESCs and, in particular, from 2i ESCs. In contrast, PGCLC specification from initial Ch/L and Pd/L states was significantly impaired, as judged by detection of ΔPE Oct4-GFP and Blimp1-GFP (Figures 3G and S5C). PGCLCs derived from all ESC culture parameters exhibited appropriate germline gene expression signatures, suggesting that they all acquire PGC fate, but differ in their specification efficiency (Figure S5D). Thus, the initial pluripotent ESC condition establishes an enduring memory that affects subsequent differentiation potential, despite an apparently normalizing intermediate EpiLC step. Collectively, these analyses imply that the initial pluripotent parameters can influence the rate and efficiency of directed differentiation toward distinct lineages. This appears to be partly correlated with the differential licensing of lineage-associated programs between ESC conditions, albeit other factors also likely contribute.
ESCs Transit through a Blimp1+ PGC-like State
We next sought to investigate the nature of lineage-associated programs in naive ESCs, focusing on the apparent germline state enriched in 2i. We employed a GFP reporter for the key PGC specifier Blimp1, which faithfully reports on endogenous BLIMP1 expression in ESCs (Figure 4A). While ESCs in all four naive conditions express the undifferentiated marker CD31, ESCs in 2i contained a greater proportion of Blimp1-GFP+ (Figure 4B).
Figure 4.

ESCs Enter a Blimp1-Positive PGC-like State
(A) Immunofluorescence staining of ESCs showing co-activation of Blimp1-GFP and endogenous BLIMP1. Scale bar, 50 μm.
(B) FACS plots of ESCs in indicated conditions showing 2i most strongly activates Blimp1 expression.
(C) Gene expression of naive pluripotency markers (upper) and PGC markers in sorted Blimp1−, Blimp1low, and Blimp1high ESC fractions by qRT-PCR. n = 3 values obtained from technical replicates; error bars, SEM.
(D) FACS showing that purified Blimp1+ and Blimp1− ESC fractions reacquire the initial equilibrium of Blimp1-GFP activity in the population after 4 days.
(E) Capacity of single or pools of five Blimp1+ or Blimp1− ESCs maintained in 2i/L to contribute to embryonic chimeras. ET, embryo transfers.
(F) Representative images of chimera contribution at E6.5 of constitutive H2B-tdTomato ESCs from Blimp1+ or Blimp1− fractions. Scale bars, 200 μm.
(G) Alkaline phosphatase-positive colony formation by sorted Blimp1+ and Blimp1− ESCs. Quantification of independent ESC lines in the right panel. Scale bar, 5 mm.
To determine the identity of this population we isolated Blimp1-high, -low, or -negative ESCs, and found that all fractions express equivalent levels of the pluripotency genes, Oct4, Sox2, and Nanog, by qRT-PCR. Strikingly however, there was a highly significant upregulation of PGC markers Nanos3, Ap2γ, Blimp1, and Stella, specifically in the Blimp1+ ESCs (Figure 4C). This was paralleled by strong repression of Uhrf1 and Klf4, which are coordinately silenced uniquely in the PGC lineage. In contrast, expression of primitive endoderm genes Gata4, Sox7, and Hex, which are also associated with BLIMP1 (Ohinata et al., 2008), were not altered. These data are consistent with Blimp1+ ESCs acquiring a PGC-like transcriptional conformation (Figure S5E). We next considered that Blimp1+ ESCs could either reversibly transit between positive and negative status in culture, or represent a static population. To determine this we fluorescence-activated cell sorted and re-plated Blimp1 -positive and -negative cells. After 4 days both isolated sub-populations reacquired equivalent levels of Blimp1+ cells, indicating that at least some ESCs can enter and exit Blimp1+ status at a frequency that rapidly leads to population equilibrium (Figure 4D).
To characterize Blimp1+ ESCs functionally, we inserted a constitutive H2B-tdTomato cassette into the Blimp1-GFP ESC line. We then introduced either 1-cell or 5-cells from 2i/L culture into blastocysts to examine the capacity for contribution to chimeras. Whereas Blimp1− ESCs robustly integrated into chimeras (12% for 1-cell and 67% of embryos for 5-cell injections), Blimp1+ ESCs contributed poorly (0% for 1-cell and 7% of embryos for 5-cell injections), despite both populations expressing high levels of naive pluripotency genes (Figures 4E and 4F). We next tested colony-formation capacity and found that Blimp1+ ESCs generated significantly fewer alkaline-positive colonies after re-plating than Blimp1− ESCs (Figure 4G). These results are consistent with observations that PGCs cannot contribute to chimeras or directly form colonies, despite expressing naive pluripotency genes (Leitch et al., 2014). The combined data support the conclusion that ESCs transiently acquire a PGC-like status, with the frequency elevated in 2i-only conditions.
Klf4 Regulates a Germline Configuration in ESCs
To understand how extrinsic signals affect dynamic ESC sub-populations we sought to investigate the regulatory principles that modulate the PGC-like ESC status. The observation that Blimp1+ are most prevalent in the 2i without LIF implies that LIF may repress entry into the germline program. To test this we found that inhibition of LIF targets JAK, but not phosphatidylinositol 3-kinase, restored the fraction of Blimp1-GFP+ cells in 2i/L to 2i levels, suggesting that JAK/STAT3 is the critical germline-repressive pathway downstream of LIF (Figure 5A). JAK/STAT3 is known to activate multiple direct targets in ESCs, including Klf4, Gbx2, Tfcp2l1, and Klf2. Forced expression of these factors using ESCs maintained in 2i revealed Klf4 most strongly downregulates Blimp1 activation to levels comparable with 2i/L (Figure 5B). Subsequent immunostaining revealed that KLF4 expression within ESC colonies is inversely correlated with BLIMP1, supporting a direct relationship (Figure 5C).
Figure 5.

Klf4 Represses the Blimp1-Positive State in ESCs Downstream of STAT3
(A) Percentage of Blimp1+ ESCs in 2i/L with or without the JAK kinase inhibitor or the phosphatidylinositol 3-kinase (PI3K) inhibitor.
(B) Percentage of Blimp1+ ESCs in 2i with forced expression of the indicated LIF target gene (A and B). n = 3 biological replicates each assayed in technical triplicate; error bars, SEM.
(C) Immunofluorescence staining of Blimp1-GFP and KLF4 in ESC colonies showing mutually exclusive expression. Scale bar, 50 μm.
(D) Immunofluorescence showing knockout (KO) of Klf4 in independent ESC lines and undetectable effects on pluripotency gene NANOG. Scale bars, 50 μm.
(E) qRT-PCR analysis of gene expression in Klf4 KO ESCs. n = 3 biological replicates each assayed in technical triplicate.
(F) FACS analysis showing a significant increase in the Blimp1+ fraction in Klf4 KO ESCs.
(G) Western blot showing KLF4 transgene rescue to WT-equivalent levels (−) or overexpression (+) in Klf4 KO cells.
(H) Re-expressing KLF4 re-imposes repression of the Blimp1+ ESC state in a dose-dependent manner. n = 3 biological replicates each assayed in technical triplicate; error bars, SEM.
To investigate this possibility we generated Klf4−/− ESCs carrying Blimp1-GFP using CRISPR targeting. Mutant ESCs formed colonies, proliferated normally, and maintained strong expression of NANOG (Figure 5D). Moreover, qRT-PCR profiling showed that expression of naive markers was unaltered between wild-type (WT) and Klf4−/− ESCs, with the exception of modest upregulation of Klf2 (Figure 5E). Importantly, in the absence of Klf4, activation of Blimp1-GFP in 2i/L increased markedly to levels similar to 2i, suggesting that KLF4 is the critical mediator sufficient to drive LIF-dependent repression of the PGC program in ESCs (Figure 5F). To examine whether re-introduction of Klf4 rescued this, we made use of a destabilized Klf4 construct whereby exogenous KLF4 is stabilized by addition of Shield1, enabling tuning of protein levels. In the absence of Shield1, the levels of exogenous KLF4 reached levels comparable with WT, while addition of Shield1 led to a significant overexpression (Figure 5G). We found that expressing Klf4 at WT levels in knockout ESCs re-imposed repression of the Blimp1+ PGC-like state in 2i/L. Moreover, elevating KLF4 levels further by addition of Shield1 further reduced the fraction of Blimp1+ ESCs to <0.5% (Figure 5H), with KLF4 chromatin immunoprecipitation-qPCR of the Blimp1 promoter suggesting that this may be an indirect effect (Figure S5G). Thus, Klf4 acts downstream of LIF/STAT3 signaling in ESCs to block dynamic entry into a germline-associated configuration.
Discussion
Our study reports a spectrum of distinct transcriptional states of pluripotency that appear to order from most naive associated to most developmentally advanced. The spectrum reflects a response to culture conditions, which influence the overall ESC transcriptome and epigenome, as well as controlling access to sub-populations. The continuum of ESC states is coupled with a gradient of increasing global DNA methylation levels, with the more hypomethylated states most closely linked with a naive signature. Moreover, alternate pluripotent conformations are linked with activation of at least some distinct classes of TEs. This, in turn, appears to influence the transcriptome, potentially through a number of routes, such as opening local chromatin structure, acting as enhancers, or forming chimeric transcripts (Chuong et al., 2016). We observed that the latter mechanism is prevalent, with chimeric transcript expression often being restricted to a unique or subsets of pluripotent conditions.
Stage-specific activation of TEs has been observed during early embryonic development, and is thought to have been co-opted to drive gene regulatory networks in a stepwise manner (Thompson et al., 2016). In this way, sequential TE activation may coordinately regulate groups of genes required for successive stages of ontogeny. An example of this can be seen at the 2C embryonic stage, where multiple key genes are activated as chimeric transcripts from MT2 and MERVL elements (Macfarlan et al., 2012). The observation that the distinct ESC conformations observed here are also linked with differential TE activity suggests that TEs may contribute to overall pluripotent status. Notably it is possible that the gradient of epigenomic states across conditions partially underpins differential TE expression, as TEs are generally more prominently activated in hypomethylated ESCs here, and transition between pluripotent/epigenetic states is also linked with distinct TE responses (Walter et al., 2016). Moreover, repeat elements activated in the more derestricted epigenetic states along the spectrum, such as LTR45 and IAP, correlate well with TEs activated upon deletion of epigenetic regulators (Reichmann et al., 2012). This collectively implies a direct relationship between the epigenome, TE activity, and overall cell state.
Whether alternate in vitro pluripotent states have any functional significance is an important question. Indeed, despite inducing broad transcriptional/epigenetic differences, the assayed culture conditions are still all capable of supporting naive pluripotency, implying that activation of the core pluripotency network overcomes wider gene expression variance. Nevertheless, the initial naive condition may subtly influence subsequent differentiation efficiency. For example, ESC propagation in Pd/L led to a poor capacity to generate the germline in vitro, but by contrast was highly efficient for the induction of nascent neuroectoderm. Likewise, culture in 2i without LIF supported high PGCLC induction, whereas Ch/L impaired such specification. It may, therefore, be important to consider the initial ESC culture condition when optimizing directed differentiation approaches, as it can leave a legacy that influences subsequent cell fate efficiency. This would imply that there is no optimal pluripotent condition to induce every cell fate with maximal efficiency, but rather a certain degree of pre-existing “bias” might be exploited according to the required endpoint. Indeed, this has recently been demonstrated by utilizing naive-like hESCs that modestly co-express primitive streak/mesodermal regulators to directly generate human PGCLCs with high efficiency (Irie et al., 2015). A further issue to consider is the apparent atrophy of imprints after extended 2i/L culture, which would be predicted to impact the developmental potential of ESCs.
A key contributor to the distinct transcriptomes between culture conditions is differential heterogeneity, as revealed at the protein level. This extends to master lineage regulators such as SOX17, GATA4, PAX6, OTX2, SOX1, and BLIMP1. Using Blimp1+ cells as a paradigm, we observed that this heterogeneity is transient, and marks a sub-population of ESCs in naive conditions with PGC-like properties. This includes strong upregulation of important germline markers Stella, Nanos3, Ap2y, and Blimp1, downregulation of Uhrf1 and Klf4, and poor performance in pluripotency assays despite strong expression of core pluripotency genes, which are all consistent with PGC properties. We show that activation of the PGC-like state in ESCs is directly repressed by LIF/STAT3 signaling. The critical LIF/STAT3 target is Klf4, which unlike other naive pluripotency genes, is repressed in PGCs in vivo (Nagamatsu et al., 2013), presumably to enable activation of the incipient germline program. Activation of Blimp1 in ESCs may be a consequence of the negative feedback loop that operates on LIF/STAT3 signaling (Starr et al., 1997), which would promote windows of Klf4 downregulation and transient activation of a PGC-like program. The prevalence of such cell-state dynamics in ESC populations may leave an enduring molecular memory, perhaps by enhancer priming, which influences subsequent cell responses to inductive cues. Alternatively, differences between culture conditions may reflect other parameters, such as differential metabolic profiles or chromatin states. The extent to which these possibilities are responsible for defining ESC states and responses merits further investigation.
Experimental Procedures
All husbandry and experiments involving mice were authorized by a UK Home Office Project License 80/2637 and carried out in a Home Office-designated facility.
ESCs were maintained on gelatin-coated dishes using culture medium as specified in Figure 1A, for at least five passages prior to experimental analysis. ESCs were maintained in a humidified 37°C chamber supplemented with 5% CO2 and passaged every 2–3 days with TrpLE. RNA-seq was performed on independent replicate samples using the TruSeq RNA Library Preparation v. 2.0 Kit (Illumina). Whole-genome bisulfite sequencing was carried out using the EZ DNA Methylation Gold Kit (Zymo Research) and Ovation Ultralow Methyl-seq Kit (NuGEN). Libraries were sequenced on a HiSeq 1500 or 2500. Differentiation assays were performed as described previously (Ying et al., 2003, Hayashi et al., 2011). CRISPR-mediated knockout of Klf4 was achieved using dual gRNAs and the Cas9 nickase to target a critical portion of exon 3. A complete description of all methods and bioinformatics analysis is listed in the Supplemental Experimental Procedures.
Author Contributions
J.A.H. designed the study, performed experiments and bioinformatics, and wrote the manuscript. T.K. designed the study and performed experiments. S.D. performed bioinformatics analysis. M.A.S. designed and supervised the study.
Acknowledgments
We thank members of the Surani lab and Dr Harry Leitch for critical reading of the manuscript and helpful discussions on the project. The Sox1-GFP ESCs were a kind gift from Prof. Austin Smith. The 2C::tdTomato construct was a kind gift from Dr Todd Macfarlan. Funding for this study came from a Wellcome Trust program grant (to M.A.S.), and Cancer Research UK (C6946/A14492)/Wellcome Trust (092096) core grants.
Published: June 6, 2017
Footnotes
Supplemental Information includes Supplemental Experimental Procedures and five figures and can be found with this article online at http://dx.doi.org/10.1016/j.stemcr.2017.05.014.
Accession Numbers
All data have been submitted to GEO under accession GEO: GSE98517.
Supplemental Information
References
- Blaschke K., Ebata K.T., Karimi M.M., Zepeda-Martinez J.A., Goyal P., Mahapatra S., Tam A., Laird D.J. Vitamin C induces Tet-dependent DNA demethylation and a blastocyst-like state in ES cells. Nature. 2013;500:222–226. doi: 10.1038/nature12362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Boroviak T., Loos R., Lombard P., Okahara J., Behr R., Sasaki E., Nichols J., Smith A. Lineage-specific profiling delineates the emergence and progression of naive pluripotency in mammalian embryogenesis. Dev. Cell. 2015;35:366–382. doi: 10.1016/j.devcel.2015.10.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bourque G., Leong B., Vega V.B., Chen X., Lee Y.L., Srinivasan K.G., Chew J.L., Ruan Y. Evolution of the mammalian transcription factor binding repertoire via transposable elements. Genome Res. 2008;18:1752–1762. doi: 10.1101/gr.080663.108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cahan P., Daley G.Q. Origins and implications of pluripotent stem cell variability and heterogeneity. Nat. Rev. Mol. Cell Biol. 2013;14:357–368. doi: 10.1038/nrm3584. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chuong E.B., Elde N.C., Feschotte C. Regulatory activities of transposable elements: from conflicts to benefits. Nat. Rev. Genet. 2016;18:71–86. doi: 10.1038/nrg.2016.139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- D'Hulst C., Parvanova I., Tomoiaga D., Sapar M.L., Feinstein P. Fast quantitative real-time PCR-based screening for common chromosomal aneuploidies in mouse embryonic stem cells. Stem Cell Reports. 2013;1:350–359. doi: 10.1016/j.stemcr.2013.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Los Angeles A., Ferrari F., Xi R., Fujiwara Y., Benvenisty N., Deng H., Hochedlinger K., Jaenisch R. Hallmarks of pluripotency. Nature. 2015;525:469–478. doi: 10.1038/nature15515. [DOI] [PubMed] [Google Scholar]
- Dunn S.J., Martello G., Yordanov B., Emmott S., Smith A.G. Defining an essential transcription factor program for naive pluripotency. Science. 2014;344:1156–1160. doi: 10.1126/science.1248882. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eckersley-Maslin M.A., Svensson V., Krueger C., Stubbs T.M., Giehr P., Krueger F., Miragaia R.J., Kyriakopoulos C. MERVL/Zscan4 network activation results in transient genome-wide DNA demethylation of mESCs. Cell Rep. 2016;17:179–192. doi: 10.1016/j.celrep.2016.08.087. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ficz G., Hore T.A., Santos F., Lee H.J., Dean W., Arand J., Krueger F., Oxley D. FGF signaling inhibition in ESCs drives rapid genome-wide demethylation to the epigenetic ground state of pluripotency. Cell Stem Cell. 2013;13:351–359. doi: 10.1016/j.stem.2013.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goke J., Lu X., Chan Y.S., Ng H.H., Ly L.H., Sachs F., Szczerbinska I. Dynamic transcription of distinct classes of endogenous retroviral elements marks specific populations of early human embryonic cells. Cell Stem Cell. 2015;16:135–141. doi: 10.1016/j.stem.2015.01.005. [DOI] [PubMed] [Google Scholar]
- Grow E.J., Flynn R.A., Chavez S.L., Bayless N.L., Wossidlo M., Wesche D.J., Martin L., Ware C.B. Intrinsic retroviral reactivation in human preimplantation embryos and pluripotent cells. Nature. 2015;522:221–225. doi: 10.1038/nature14308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Habibi E., Brinkman A.B., Arand J., Kroeze L.I., Kerstens H.H., Matarese F., Lepikhov K., Gut M. Whole-genome bisulfite sequencing of two distinct interconvertible DNA methylomes of mouse embryonic stem cells. Cell Stem Cell. 2013;13:360–369. doi: 10.1016/j.stem.2013.06.002. [DOI] [PubMed] [Google Scholar]
- Hackett J.A., Surani M.A. Regulatory principles of pluripotency: from the ground state up. Cell Stem Cell. 2014;15:416–430. doi: 10.1016/j.stem.2014.09.015. [DOI] [PubMed] [Google Scholar]
- Hackett J.A., Dietmann S., Murakami K., Down T.A., Leitch H.G., Surani M.A. Synergistic mechanisms of DNA demethylation during transition to ground-state pluripotency. Stem Cell Reports. 2013;1:518–531. doi: 10.1016/j.stemcr.2013.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hackett J.A., Sengupta R., Zylicz J.J., Murakami K., Lee C., Down T.A., Surani M.A. Germline DNA demethylation dynamics and imprint erasure through 5-hydroxymethylcytosine. Science. 2013;339:448–452. doi: 10.1126/science.1229277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall J., Guo G., Wray J., Eyres I., Nichols J., Grotewold L., Morfopoulou S., Humphreys P. Oct4 and LIF/Stat3 additively induce Kruppel factors to sustain embryonic stem cell self-renewal. Cell Stem Cell. 2009;5:597–609. doi: 10.1016/j.stem.2009.11.003. [DOI] [PubMed] [Google Scholar]
- Hayashi K., Ohta H., Kurimoto K., Aramaki S., Saitou M. Reconstitution of the mouse germ cell specification pathway in culture by pluripotent stem cells. Cell. 2011;146:519–532. doi: 10.1016/j.cell.2011.06.052. [DOI] [PubMed] [Google Scholar]
- Huang K., Maruyama T., Fan G. The naive state of human pluripotent stem cells: a synthesis of stem cell and preimplantation embryo transcriptome analyses. Cell Stem Cell. 2014;15:410–415. doi: 10.1016/j.stem.2014.09.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Irie N., Weinberger L., Tang W.W., Kobayashi T., Viukov S., Manor Y.S., Dietmann S., Hanna J.H. SOX17 is a critical specifier of human primordial germ cell fate. Cell. 2015;160:253–268. doi: 10.1016/j.cell.2014.12.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klein A.M., Mazutis L., Akartuna I., Tallapragada N., Veres A., Li V., Peshkin L., Weitz D.A. Droplet barcoding for single-cell transcriptomics applied to embryonic stem cells. Cell. 2015;161:1187–1201. doi: 10.1016/j.cell.2015.04.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kolodziejczyk A.A., Kim J.K., Tsang J.C., Ilicic T., Henriksson J., Natarajan K.N., Tuck A.C., Gao X. Single cell RNA-sequencing of pluripotent states unlocks modular transcriptional variation. Cell Stem Cell. 2015;17:471–485. doi: 10.1016/j.stem.2015.09.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leitch H.G., Mcewen K.R., Turp A., Encheva V., Carroll T., Grabole N., Mansfield W., Nashun B. Naive pluripotency is associated with global DNA hypomethylation. Nat. Struct. Mol. Biol. 2013;20:311–316. doi: 10.1038/nsmb.2510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leitch H.G., Okamura D., Durcova-Hills G., Stewart C.L., Gardner R.L., Matsui Y., Papaioannou V.E. On the fate of primordial germ cells injected into early mouse embryos. Dev. Biol. 2014;385:155–159. doi: 10.1016/j.ydbio.2013.11.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Macfarlan T.S., Gifford W.D., Driscoll S., Lettieri K., Rowe H.M., Bonanomi D., Firth A., Singer O. Embryonic stem cell potency fluctuates with endogenous retrovirus activity. Nature. 2012;487:57–63. doi: 10.1038/nature11244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Marks H., Kalkan T., Menafra R., Denissov S., Jones K., Hofemeister H., Nichols J., Kranz A. The transcriptional and epigenomic foundations of ground state pluripotency. Cell. 2012;149:590–604. doi: 10.1016/j.cell.2012.03.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morgani S.M., Canham M.A., Nichols J., Sharov A.A., Migueles R.P., Ko M.S., Brickman J.M. Totipotent embryonic stem cells arise in ground-state culture conditions. Cell Rep. 2013;3:1945–1957. doi: 10.1016/j.celrep.2013.04.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nagamatsu G., Kosaka T., Saito S., Honda H., Takubo K., Kinoshita T., Akiyama H., Sudo T. Induction of pluripotent stem cells from primordial germ cells by single reprogramming factors. Stem Cells. 2013;31:479–487. doi: 10.1002/stem.1303. [DOI] [PubMed] [Google Scholar]
- Nichols J., Smith A. Naive and primed pluripotent states. Cell Stem Cell. 2009;4:487–492. doi: 10.1016/j.stem.2009.05.015. [DOI] [PubMed] [Google Scholar]
- Ohinata Y., Sano M., Shigeta M., Yamanaka K., Saitou M. A comprehensive, non-invasive visualization of primordial germ cell development in mice by the Prdm1-mVenus and Dppa3-ECFP double transgenic reporter. Reproduction. 2008;136:503–514. doi: 10.1530/REP-08-0053. [DOI] [PubMed] [Google Scholar]
- Pauklin S., Vallier L. The cell-cycle state of stem cells determines cell fate propensity. Cell. 2013;155:135–147. doi: 10.1016/j.cell.2013.08.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peaston A.E., Evsikov A.V., Graber J.H., de Vries W.N., Holbrook A.E., Solter D., Knowles B.B. Retrotransposons regulate host genes in mouse oocytes and preimplantation embryos. Dev. Cell. 2004;7:597–606. doi: 10.1016/j.devcel.2004.09.004. [DOI] [PubMed] [Google Scholar]
- Piccolo F.M., Bagci H., Brown K.E., Landeira D., Soza-Ried J., Feytout A., Mooijman D., Hajkova P. Different roles for Tet1 and Tet2 proteins in reprogramming-mediated erasure of imprints induced by EGC fusion. Mol. Cell. 2013;49:1023–1033. doi: 10.1016/j.molcel.2013.01.032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reichmann J., Crichton J.H., Madej M.J., Taggart M., Gautier P., Garcia-Perez J.L., Meehan R.R., Adams I.R. Microarray analysis of LTR retrotransposon silencing identifies Hdac1 as a regulator of retrotransposon expression in mouse embryonic stem cells. PLoS Comput. Biol. 2012;8:e1002486. doi: 10.1371/journal.pcbi.1002486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sheik Mohamed J., Gaughwin P.M., Lim B., Robson P., Lipovich L. Conserved long noncoding RNAs transcriptionally regulated by Oct4 and Nanog modulate pluripotency in mouse embryonic stem cells. RNA. 2010;16:324–337. doi: 10.1261/rna.1441510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Starr R., Willson T.A., Viney E.M., Murray L.J., Rayner J.R., Jenkins B.J., Gonda T.J., Alexander W.S. A family of cytokine-inducible inhibitors of signalling. Nature. 1997;387:917–921. doi: 10.1038/43206. [DOI] [PubMed] [Google Scholar]
- Thompson P.J., Macfarlan T.S., Lorincz M.C. Long terminal repeats: from parasitic elements to building blocks of the transcriptional regulatory repertoire. Mol. Cell. 2016;62:766–776. doi: 10.1016/j.molcel.2016.03.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tonge P.D., Corso A.J., Monetti C., Hussein S.M., Puri M.C., Michael I.P., Li M., Lee D.S. Divergent reprogramming routes lead to alternative stem-cell states. Nature. 2014;516:192–197. doi: 10.1038/nature14047. [DOI] [PubMed] [Google Scholar]
- Torres-Padilla M.E., Chambers I. Transcription factor heterogeneity in pluripotent stem cells: a stochastic advantage. Development. 2014;141:2173–2181. doi: 10.1242/dev.102624. [DOI] [PubMed] [Google Scholar]
- von Meyenn F., Iurlaro M., Habibi E., Liu N.Q., Salehzadeh-Yazdi A., Santos F., Petrini E., Milagre I. Impairment of DNA methylation maintenance is the main cause of global demethylation in naive embryonic stem cells. Mol. Cell. 2016;62:983. doi: 10.1016/j.molcel.2016.06.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Walter M., Teissandier A., Perez-Palacios R., Bourc'his D. An epigenetic switch ensures transposon repression upon dynamic loss of DNA methylation in embryonic stem cells. Elife. 2016;5:e11418. doi: 10.7554/eLife.11418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wan M., Liang J., Xiong Y., Shi F., Zhang Y., Lu W., He Q., Yang D., Chen R., Liu D. The trithorax group protein Ash2l is essential for pluripotency and maintaining open chromatin in embryonic stem cells. J Biol. Chem. 2012;288:5039–5048. doi: 10.1074/jbc.M112.424515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang J., Xie G., Singh M., Ghanbarian A.T., Rasko T., Szvetnik A., Cai H., Besser D. Primate-specific endogenous retrovirus-driven transcription defines naive-like stem cells. Nature. 2014;516:405–409. doi: 10.1038/nature13804. [DOI] [PubMed] [Google Scholar]
- Weinberger L., Ayyash M., Novershtern N., Hanna J.H. Dynamic stem cell states: naive to primed pluripotency in rodents and humans. Nat. Rev. Mol. Cell Biol. 2016;17:155–169. doi: 10.1038/nrm.2015.28. [DOI] [PubMed] [Google Scholar]
- Wray J., Kalkan T., Smith A.G. The ground state of pluripotency. Biochem. Soc. Trans. 2010;38:1027–1032. doi: 10.1042/BST0381027. [DOI] [PubMed] [Google Scholar]
- Wu J., Izpisua Belmonte J.C. Dynamic pluripotent stem cell states and their applications. Cell Stem Cell. 2015;17:509–525. doi: 10.1016/j.stem.2015.10.009. [DOI] [PubMed] [Google Scholar]
- Wu J., Okamura D., Li M., Suzuki K., Luo C., Ma L., He Y., Li Z. An alternative pluripotent state confers interspecies chimaeric competency. Nature. 2015;521:316–321. doi: 10.1038/nature14413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaguchi S., Shen L., Liu Y., Sendler D., Zhang Y. Role of Tet1 in erasure of genomic imprinting. Nature. 2013;504:460–464. doi: 10.1038/nature12805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ye S., Li P., Tong C., Ying Q.L. Embryonic stem cell self-renewal pathways converge on the transcription factor Tfcp2l1. EMBO J. 2013;32:2548–2560. doi: 10.1038/emboj.2013.175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ying Q.L., Stavridis M., Griffiths D., Li M., Smith A. Conversion of embryonic stem cells into neuroectodermal precursors in adherent monoculture. Nat. Biotechnol. 2003;21:183–186. doi: 10.1038/nbt780. [DOI] [PubMed] [Google Scholar]
- Ying Q.L., Wray J., Nichols J., Batlle-Morera L., Doble B., Woodgett J., Cohen P., Smith A. The ground state of embryonic stem cell self-renewal. Nature. 2008;453:519–523. doi: 10.1038/nature06968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zalzman M., Falco G., Sharova L.V., Nishiyama A., Thomas M., Lee S.L., Stagg C.A., Hoang H.G., Yang H.T., Indig F.E. Zscan4 regulates telomere elongation and genomic stability in ES cells. Nature. 2010;464:858–863. doi: 10.1038/nature08882. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.