

Supplemental Digital Content is available in the text.
Keywords: aging, angiogenesis, dedifferentiation, development, endothelial cells, erythropoiesis, fibroblasts, myocardial infarction, regeneration, reprogramming, progenitor cells, SOX17, telomerase
Abstract
Background:
The mechanisms underlying the dedifferentiation and lineage conversion of adult human fibroblasts into functional endothelial cells have not yet been fully defined. Furthermore, it is not known whether fibroblast dedifferentiation recapitulates the generation of multipotent progenitors during embryonic development, which give rise to endothelial and hematopoietic cell lineages. Here we established the role of the developmental transcription factor SOX17 in regulating the bilineage conversion of fibroblasts by the generation of intermediate progenitors.
Methods:
CD34+ progenitors were generated after the dedifferentiation of human adult dermal fibroblasts by overexpression of pluripotency transcription factors. Sorted CD34+ cells were transdifferentiated into induced endothelial cells and induced erythroblasts using lineage-specific growth factors. The therapeutic potential of the generated cells was assessed in an experimental model of myocardial infarction.
Results:
Induced endothelial cells expressed specific endothelial cell surface markers and also exhibited the capacity for cell proliferation and neovascularization. Induced erythroblasts expressed erythroid surface markers and formed erythroid colonies. Endothelial lineage conversion was dependent on the upregulation of the developmental transcription factor SOX17, whereas suppression of SOX17 instead directed the cells toward an erythroid fate. Implantation of these human bipotential CD34+ progenitors into nonobese diabetic/severe combined immunodeficiency (NOD-SCID) mice resulted in the formation of microvessels derived from human fibroblasts perfused with mouse and human erythrocytes. Endothelial cells generated from human fibroblasts also showed upregulation of telomerase. Cell implantation markedly improved vascularity and cardiac function after myocardial infarction without any evidence of teratoma formation.
Conclusions:
Dedifferentiation of fibroblasts to intermediate CD34+ progenitors gives rise to endothelial cells and erythroblasts in a SOX17-dependent manner. These findings identify the intermediate CD34+ progenitor state as a critical bifurcation point, which can be tuned to generate functional blood vessels or erythrocytes and salvage ischemic tissue.
Direct conversion of somatic cells such as fibroblasts into other cell types for organ and tissue regeneration holds great therapeutic promise.1 Studies have shown that fibroblasts can be reprogrammed into hepatocyte-like cells, cardiomyocytes, neurons, endothelial cells (ECs), and erythroid progenitors.2–6 Direct conversion of fibroblasts to regenerative therapeutic cells is thus not only more expedient than the intermediate generation of iPSCs, which requires further differentiation into specific lineage cells, but also avoids risk of teratoma formation caused by any residual pluripotent cells.7 Two general strategies are used to convert fibroblasts into ECs. One relies on dedifferentiation, in which the pluripotency-inducing transcription factors OCT-4, KLF4, SOX2, and c-MYC are expressed to partially dedifferentiate fibroblasts, and this is followed by applying endothelial growth factors such as vascular endothelial growth factor to induce terminal EC differentiation.8–10 The second approach relies on the targeted overexpression of developmental transcription factors, such as ETV2, known to regulate formation of ECs.11,12
However, little is known about the underlying mechanisms that enable reprogramming and whether these 2 strategies—partial dedifferentiation and directed vascular transcription factor overexpression—can be combined to induce more efficient EC generation. The advantage of the dedifferentiation strategy is that it may generate multipotent progenitors that can in turn be directed into multiple regenerative cell lineages. ECs generated during embryonic development are derived from mesodermal progenitors and hemangioblasts, which are a short-lived cell population that generates both endothelial and hematopoietic lineage cells.13–15 Thus, we surmised that if dedifferentiation of adult fibroblasts recapitulates embryonic development, then generation of fibroblast-derived progenitors may also provide a means of forming new blood vessels along with hematopoietic cells, including erythrocytes. Human hemangioblasts reportedly have distinct subpopulations expressing different surface markers such as CD34.16–18 Here, we demonstrated that dedifferentiation of human fibroblasts generated bipotential CD34+ progenitors capable of differentiating into ECs and erythroblasts. The transcription factor SOX17 functioned as a tunable rheostat-like switch in CD34+ cells, giving rise to ECs and the erythroid lineage. Further, we demonstrated that implantation of CD34+ progenitors generated functional human blood vessels and improved cardiac function in infarcted mouse hearts.
Methods
Cell Culture and Transfection
Normal human adult dermal fibroblast cells (Lonza, CC-2511) and human neonatal dermal fibroblast cells (Lonza, CC-2509) were cultured in DMEM (Life Technologies, 21063) containing 10% FBS, 50 U/mL penicillin and 50 mg/mL streptomycin, 2.25 mM L-glutamine, and 2.25 mM nonessential amino acids. Fibroblasts (5.0×105/100 mm dish) and PLAT-A cells (Cell Biolabs Inc. RV-102) (5.0×106/100 mm dish) were plated in DMEM medium without antibiotics; 100 mm dishes were plated with PLAT-A cells, 1 dish per plasmid, each encoding OCT4, KLF4, SOX2, and c-MYC (OKSM; all obtained from Addgene and were gifts from Shinya Yamanaka: pMXs-hOCT3/4, Addgene plasmid #17217; pMXs-hSOX2, Addgene plasmid #17218; pMXs-hKLF4, Addgene plasmid #117219; pMXs-hc-MYC, Addgene plasmid #17220), as well as green fluorescent protein (GFP) as a control. On day 0, Fugene HD transfection reagent (Roche, E2311) and 10 μg of the appropriate plasmid DNA were added to one 100 mm dish cultured with PLAT-A cells. The virus-containing medium was collected and filtered from dishes of transfected PLAT-A cells on days 1 and 2 for titration. The medium of human fibroblasts was replaced with virus-containing medium on days 1 and 2. On day 3, the medium was switched to DMEM/F12 (Life Technologies, 11330), 20% KnockOut serum replacement (Life Technologies, 10828), 10 ng ml−1 bFGF (R&D Systems, 233-FB), 1 mM GlutaMAX, 0.1 mM nonessential amino acids, and 55 μM β-mercaptoethanol with medium changes every day for 5 days to continue the dedifferentiation phase. The fibroblasts transduced by OKSM and maintained in this medium were referred to as dedifferentiated fibroblasts. The protocol is shown in Figure 1A. All cell lines were maintained in an incubator (37°C, 5% CO2).
Figure 1.
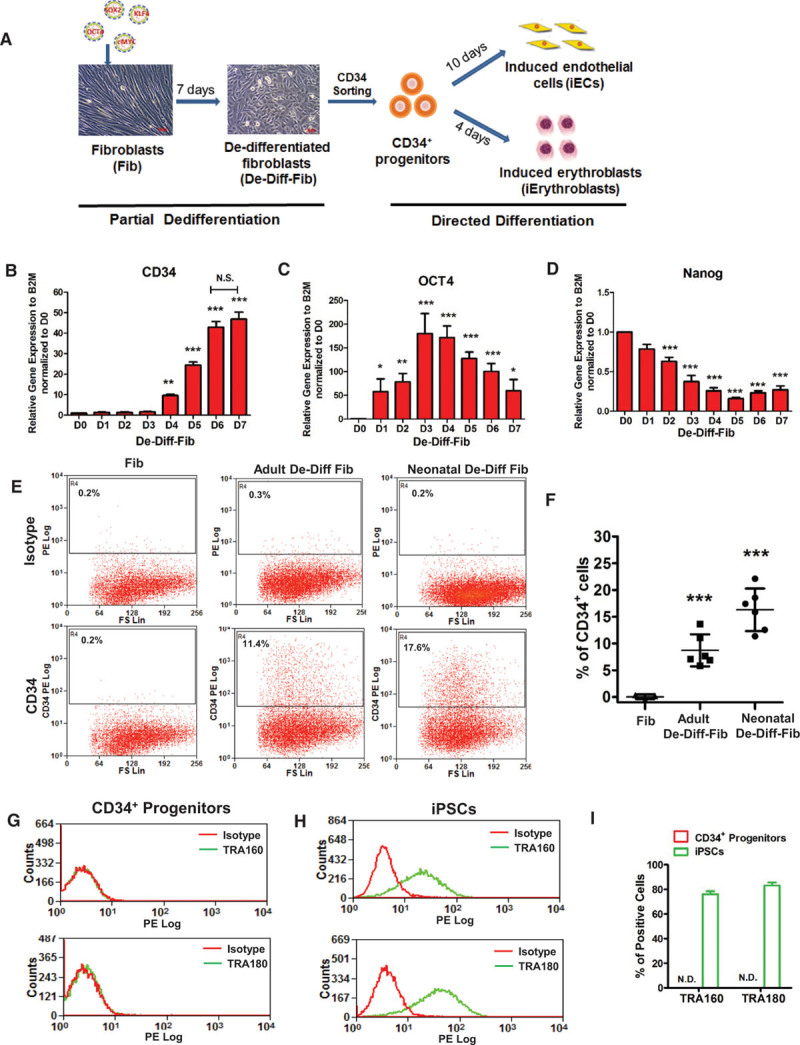
Generating CD34+progenitors from human dermal fibroblasts. A, Schematic of converting human dermal fibroblasts into induced endothelial cells (iECs) and induced Erythroblasts (iErythroblasts) by sorted CD34+ progenitors from dedifferentiated fibroblasts (De-Diff-Fib) at day 7. B through D, Quantitative polymerase chain reaction (PCR) analysis of the mesodermal marker CD34 (B), pluripotency markers OCT4 (C), and Nanog (D) during adult fibroblast dedifferentiation from days 0 to 7 (n=4). E, Representative flow-cytometry plots of the mesodermal marker CD34+ in fibroblasts, adult De-Diff-Fib, and neonatal De-Diff-Fib. F, Quantification and statistical analysis of CD34+ cells in fibroblasts (n=3), adult De-Diff-Fib (n=6), and neonatal De-Diff-Fib (n=6) from flow cytometry. G through I, Representative flow-cytometry overlay plots of the pluripotency markers: human TRA-160 and TRA-180 in CD34+ progenitors (G) and iPSCs (H) and quantification of TRA-160 or TRA-180-positive cells (I) in CD34+ progenitors and iPSCs. Data are presented as mean±SE, *P<0.05, **P<0.01, ***P<0.001, compared with day 0 in B, C, and D, compared with fibroblasts in F, using repeated measures 1-way analysis of variance in B, C, and D and using 1-way analysis in F. N.D. indicates not detected in I.
Isolation of CD34+ Progenitors From Dedifferentiated Fibroblasts
We identified the CD34+ progenitors by CD34, a marker of human progenitors, including endothelial and hematopoietic progenitors.18 We isolated CD34+ cells generated during dedifferentiation of fibroblast with anti-CD34–conjugated magnetic beads (Miltenyi, 130-046) according to manufacturer’s instructions with slight modifications. Briefly, ≤109 cells were incubated at 4°C with 100 μl of Fc-blocking solution and 100 μl anti-CD34 magnetic beads in a total volume of 500 μl FACS blocking buffer. After 30 minutes, cells were sorted by 2 consecutive rounds of column separation through applying magnetic activated cell sorting (MACS, Miltenyi Biotec) separation magnets to increase purity.
Differentiation of CD34+ Progenitors Into Induced ECs (iECs)
Isolated CD34+ progenitors were plated in 0.1% gelatin–coated plates (2×105/6 well), cultured in endothelial medium EGM-2 (CC-4176), and supplemented with 50 ng/mL BMP4 (R&D Systems, 314-BP) and 50 ng/mL vascular endothelial growth factor (R&D Systems, 293-VE) to induce endothelial lineage specification, and the medium was changed every other day. On reaching ≈90% confluence, cells were split 1:3. Primary human aortic ECs were purchased from Lonza (CC-2535) and cultured in EGM-2 medium as control cells.
Differentiation of CD34+ Progenitors Into Induced Erythroblasts (iErythroblasts)
To generate erythroid cells, sorted CD34+ progenitors were plated in 0.1% gelatin-coated plates (1.0×106/well in a 6-well plate) and cultured for 4 days in erythroblast induction medium, which consisted of 49% IMDM (Life Technologies, 12440), 49% Ham’s F12 (Life Technologies, 11765), 1% ITSX, 1% lipid (Life Technologies, 11905-031), 5 mg/100 mL I-ascorbic acid (Sigma, A8960), 500 mg/100 mL BSA (Sigma, A7030), and 200uM 1-thioglycerol (Sigma, M6145), supplemented with 100 ng/mL SCF (PeproTech, 300–07), 10 ng/mL interleukin-3 (PeproTech, 200–03), 5 U/mL EPO (R&D Systems, 287-TC), 40 ng/mL insulin-like growth factor-1(Sigma I-3769), 1 μM Dexamethsone (Sigma, D2915), and 50 ng/mL Flt3-L (PeproTech, 300–19) (Figure 1A). Unlike adherent ECs, induced erythroblasts detached on generation and were collected as nonadherent cell fraction at day 4 for characterization.
Matrigel Plug In Vivo Angiogenesis Assay
To test the ability of iECs to form blood vessels in vivo, we performed an in vivo gel plug assay.19 Nonobese diabetic/severe combined immunodeficiency (NOD-SCID) mice were used to avoid the rejection of human cells. Anesthesia was induced using a mixture of xylazine (10 mg/kg) and ketamine (100 mg/kg) in saline IP. A total of 1×106 cells (either CD34+ progenitors, fibroblasts, or aortic ECs) were suspended in 200 μl of cold growth factor-reduced Matrigel Matrix (Corning, 356234). We then injected the cell-containing Matrigel solutions subcutaneously in the abdomen. The mice were euthanized after 1 week implantation, and the Matrigel plugs were removed by a wide excision of the skin, fixed in 10% formalin overnight, embedded in paraffin, and sectioned. Five sections from each plug were randomly selected to quantify human vessels. Quantification of vessel density in angiogenesis plugs was shown per section per plug. Each mouse was injected with 3 plugs containing iECs, fibroblasts, or mature ECs. Each data point represents 1 plug from 1 mouse.
Myocardial Infarction (MI) Surgery and Intramyocardial Injection of Cells
The MI surgery is described in detail in the online-only Data Supplement. For the cell implantation, 40 μl of concentrated cells −1×106 CD34+ progenitors or fibroblasts suspended in growth factor-reduced Matrigel (Matrigel basement-membrane matrix from BD) were injected into the border zone of the infarct at 2 to 3 different areas using a Gastight 1710 syringe (Hamilton) immediately after ligation. The injected cells had been prelabeled with lentiviral GFP. The surgeon was blinded to the nature of the injected cells. Two weeks later, hearts were retrograde perfused with 4% paraformaldehyde by the aorta, fixed in 4% paraformaldehyde for 4 hours, incubated in 30% sucrose overnight, embedded in OCT, frozen, and cryosectioned. Ten sections with injected GFP cells from each heart were used for quantification of human CD31+ vessels. Quantification of vessel density was shown as vessels per section per heart.
Experimental Animals
Immune deficient NOD-SCID mice were purchased from Jackson Laboratory (NOD.Cg-Prkdcscid Il2rgtm1Wjl/SzJ, strain 005557). All animal experiments were conducted in accordance with National Institutes of Health guidelines for the care and use of live animals and were approved by the Institutional Animal Care and Use Committee of the University of Illinois.
Statistics
Cell culture studies on fibroblasts and endothelial cells were performed with commercially obtained primary cells from multiple batches (Lonza) and are biological replicates. Values were reported as mean±SE. The statistical analysis was performed with GraphPad Prism and is indicated in the figure legends. Bonferroni corrections were used for analysis of variance with multiple comparisons. Statistical significance was set at P<0.05.
Results
Dedifferentiation of Human Fibroblasts Into CD34+ Progenitors
CD34+ progenitors were generated by expressing the Yamanaka factors OCT4, KLF4, SOX2, and c-MYC (OKSM) in human fibroblasts, followed by 7 days of dedifferentiation (Figure 1A). These fibroblast-derived cells were referred to as dedifferentiated fibroblasts, and we subsequently applied endothelial or erythroid lineage growth factors to differentiate them into ECs or erythroblasts (Figure 1A and Figure I in the online-only Data Supplement). As shown in Figure 1B, expression of CD34 gradually increased during the dedifferentiation period and reached a plateau at days 6 to 7. OCT4 mRNA levels peaked at days 3 to 4 and then steadily decreased (Figure 1C), indicating that pluripotency was circumvented. Expression of SOX2, c-MYC, and KLF4 exhibited similar expression changes as OCT4. We also assessed the expression of NANOG, an endogenous pluripotency factor, which was not part of the OKSM cocktail. NANOG was downregulated during fibroblast dedifferentiation (Figure 1D), confirming that short-term induction of 7 days did not induce pluripotency. We used day 7 cells for subsequent studies because CD34 expression had reached a plateau, and there was no evidence of pluripotency at this stage. Assessing generation of CD34+ cells by flow cytometry, we found that dedifferentiation of both human neonatal and adult dermal fibroblasts gave rise to CD34+ cells, with higher CD34+ yield from neonatal compared with adult fibroblasts (16.3±2% versus 8.7±1%; Figure 1E and 1F). We then focused on human adult dermal fibroblasts because of their potential utility for vascular regeneration in patients.
The CD34+ cells were also evaluated for their clonal expansion potential and the pluripotency surface markers TRA-160 and TRA-180, endothelial surface markers VE-cadherin and CD31, and the erythroblast/erythrocyte surface marker CD235a by flow cytometry. Plating of individual CD34+ cells did not result in any formation of colonies. We also found that CD34+ cells did not express markers of pluripotency (Figure 1G through 1I), endothelial differentiation (Figure IIA and IIB in the online-only Data Supplement), or erythroblast differentiation (Figure IIC and IID in the online-only Data Supplement), demonstrating that the cells were undifferentiated progenitors.
Conversion of Fibroblast-Derived CD34+ Progenitors Into ECs
We isolated CD34+ progenitors from the dedifferentiated fibroblast population using magnetic bead sorting and exposed the cells to an EC lineage induction medium consisting of standard endothelial growth medium supplemented with the EC differentiation factors vascular endothelial growth factor and BMP-4 for 10 days. During the induction phase, gene expression of EC markers VE-cadherin, VWF, CD31, and FLK1 progressively increased (Figure 2A). The induction of CD34+ cells was terminated on day 10 when EC marker levels plateaued (data not shown). Flow cytometry of cells at day 10 demonstrated that 7.8±3% of cells were positive for VE-cadherin, 8.4±3% of cells for CD31, and 4.9±1% of cells for both surface markers (Figure 2B and 2C). We found 92.9±1.2% of VE-cadherin+ iECs continued to express CD34 after endothelial differentiation (Figure IIE in the online-only Data Supplement). Western blotting established upregulation of VE-cadherin, CD31, and FLK1 in these generated cells, which we termed iECs (Figure 2D). Immunocytochemistry staining demonstrated that the bulk of VE-cadherin was localized in the cytoplasm in these nonconfluent iECs as opposed to the cell surface membrane where VE-cadherin is typically found in confluent adherent ECs (Figure 2E). The relatively high-cytosolic VE-cadherin expression also explained why only 8% of iECs were positive for VE-cadherin by flow cytometry as it monitors the cell surface expressed protein. Because VE-cadherin transinteractions between cells stabilize VE-cadherin at the cell membrane and promote formation of adherens junctions,20 we next cocultured the iECs with primary adult human aortic ECs. Confocal imaging demonstrated that this increased the membrane localization of VE-cadherin such that iECs formed characteristic adherens junctions by homotypic interactions (Figure 2F). We also examined the gene expression levels of the endothelial markers VE-cadherin, CD31, FLK1, and VWF in CD31+ bead-sorted iECs at day 10 of induction. The iEC mRNA levels of these endothelial markers were ≈50% of those in mature human ECs but 100- to 1000-fold higher than those in fibroblasts and CD34+ progenitors (Figure IIIA through IIID in the online-only Data Supplement).
Figure 2.
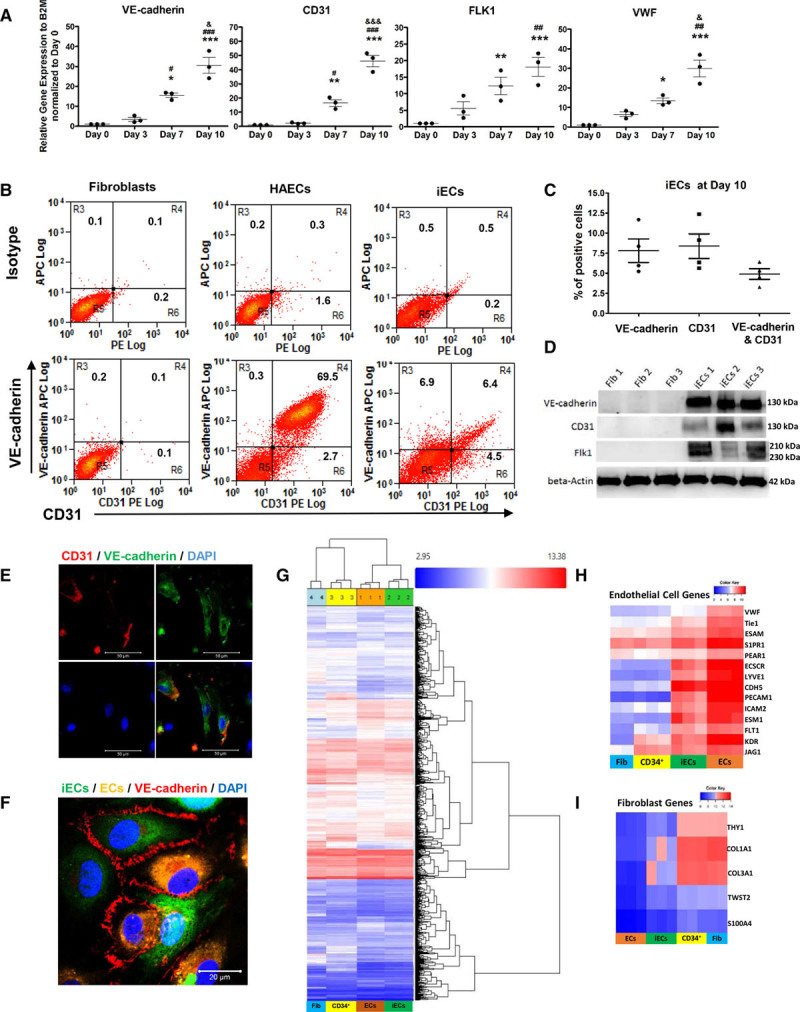
Induction of endothelial cells. A, Quantitative polymerase chain reaction (PCR) analysis of endothelial-specific genes VE-cadherin, CD31, VWF, and FLK1 in induced endothelial cells (iECs) from days 0 to 10, normalized to day 0 (n=3). B, Representative flow-cytometry plots of endothelial markers VE-cadherin and CD31 at day 10 in human aortic ECs and iECs. C, Quantification of VE-cadherin+ and CD31+ cells in iECs at day 10 from flow cytometry (n=4). D, Immunoblots of the endothelial-specific proteins human VE-cadherin, CD31, and FLK1 in iECs and fibroblasts; β-Actin served as a control. E, Representative micrographs of immunocytochemistry staining of human VE-cadherin (green) and human CD31 (red) of iECs at day 10; nuclei were stained by DAPI. Scale bars: 50 µm. F, Confocal micrographs of immunofluorescence staining of human VE-cadherin (red) in iECs (labeled by CellTracker Green, Invitrogen Catalog #: C7025TM) co-cultured with ECs (labeled by CellTracker Orange, Invitrogen Catalog #: C2927) for 2 days. Scale bars: 20 µm. G–I, Analysis of cDNA microarrays in adult fibroblasts, CD34+ progenitors (labeled as CD34+), iECs, and ECs heat map of global gene using hierarchical clustering analysis (G), heat map of endothelial genes (H), and fibroblast genes (I). The color bars indicate gene expression level in log2 scale. Blue to red color represents low to high expression level. Data are presented as mean±SE; *P<0.05, **P<0.01, ***P<0.001, compared with day 0. #P<0.05, ##P<0.01, ###P<0.001, compared with day 3; &P<0.05, &&&P<0.05 compared with day 7 using repeated measures 1-way analysis in A.
We next performed a transcriptomic analysis of adult fibroblasts, fibroblast-derived CD34+ progenitors, iECs, and primary human ECs. A heat map of global gene expression revealed that iECs clustered with ECs, whereas CD34+ progenitors clustered with fibroblasts (Figure 2G). Thus, iECs exhibited a gene expression profile similar to adult primary ECs, whereas fibroblast-derived CD34+ progenitors retained fibroblast signatures. Heat maps of EC- and fibroblast-specific genes indicated that gain of EC identify and loss of fibroblast signatures occurred during the transition from CD34+ cells to iECs (Figure 2H and 2I).
Fibroblast-Derived iECs Are Functional ECs
Key functions of ECs are to form a restrictive and dynamic barrier and upregulate cell surface adhesion molecules in response to inflammatory stimuli. Thus, we assessed transendothelial electric resistance in a confluent monolayer of iECs as a measure of the barrier function. In response to thrombin, we observed a significant decrease in transendothelial electric resistance of iECs, while monolayers of parent fibroblasts showed no such response (Figure 3A and 3B and Figure IV in the online-only Data Supplement). It is important to note that iECs exhibited restoration of the barrier, a physiological adaptive response seen in native endothelium. We also demonstrated that iECs formed capillary networks in 2D-Matrigel (Figure 3C and 3D), whereas fibroblasts predominately remained as single cells or cell clusters. To evaluate responses to the proinflammatory cytokine tumor necrosis factor-α, we assessed expression of adhesion molecules VCAM1 and E-Selectin. iECs responded with upregulation of VCAM1 and E-selectin proteins (Figure 3E) and mRNA (Figure 3F) in the same manner as primary human ECs.
Figure 3.
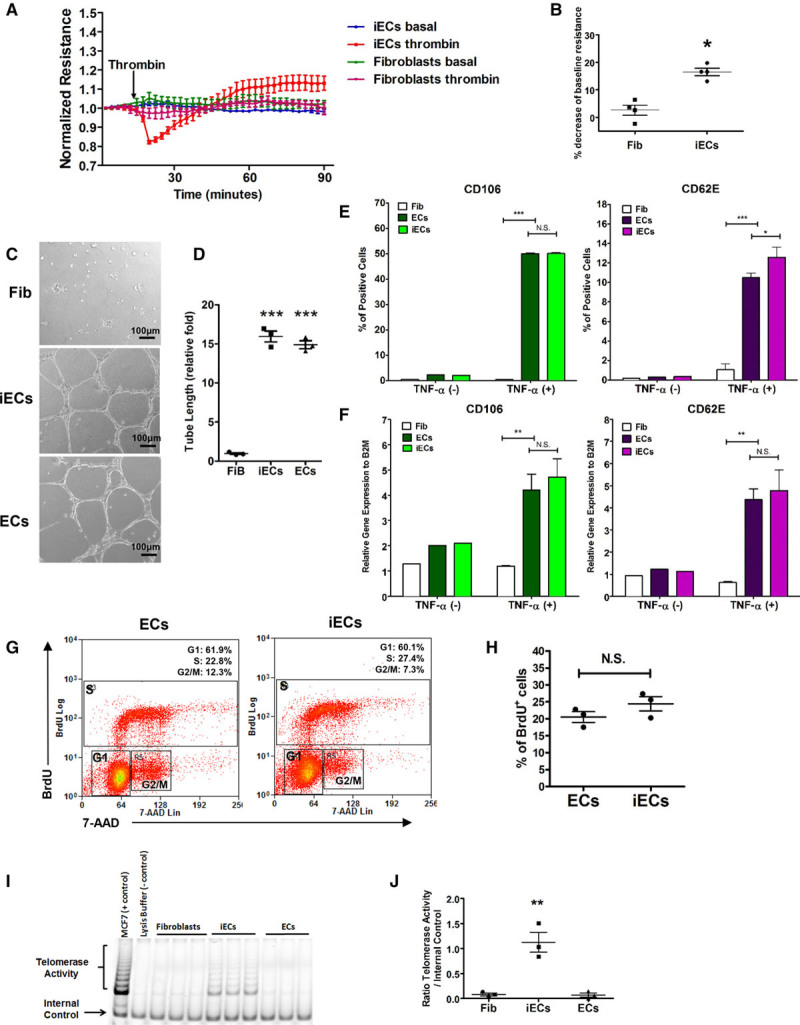
Characterization of induced endothelial cells (iECs). A and B, The permeability of iECs in response to thrombin. Transendothelial electric resistance (TER) was measured for 90 minutes in fibroblasts and iECs. Thrombin (3.5µg/mL, 0.5 U) was added as indicated. The time course of resistance in fibroblasts and iECs normalized to baseline (A) and statistical analysis (B) of percentage of deduction of resistance in response to thrombin in fibroblasts and iECs (n=4). C and D, In vitro capillary network formation of iECs and fibroblasts on Matrigel. The representative images (C) and statistical analysis (D) of tube network formation in fibroblasts, iECs, and ECs (n=3). E and F, The inflammation response of iECs. FACS analysis (E) and quantitative realtime-polymerase chain reaction (PCR) measurement (F) for CD106 and CD62E on tumor necrosis factor-α (TNF-α) activated ECs and iECs (n=3). G and H, The proliferation of iECs. Representative flow-cytometry plots (G) of the proliferation capacity of iECs and ECs as assessed by BrdU uptake and statistical analysis (H) of the percentage of BrdU+ cells in iECs and ECs (n=3). I and J, The telomerase activity of iECs. Telomerase activity was measured by a telomeric repeat amplification protocol (TRAP) assay. PCR amplification results of TRAP (I) from positive control, negative control, fibroblasts, ECs, and iECs; and statistical analysis (J) of telomerase activity in above cell types (n=3). Data are presented as mean±SE; *P<0.05, **P<0.01, ***P<0.001, N.S. P>0.05; compared with fibroblasts in B, D through F, and J; compared with ECs in E, F, and H using Student t test in B and H, 1-way analysis of variance in J and 2-way analysis in E and F.
Proliferation Potential and Telomerase Expression in iECs
Because ECs’ ability to proliferate is required for vascular regeneration, we assessed proliferative potential of iECs. As shown in Figure 3G and 3H, 20% to 30% of iECs were labeled with BrdU, a value similar to the proliferation rate of primary human ECs. Cell expansion induces cell senescence and results in cell cycle arrest because of continuous shortening of telomeres.21–23 Therefore, we assessed whether the generated iECs represented an aging cell population. We determined telomerase activity because it maintains telomere length and protects against premature senescence in embryonic and neonatal cells.21–23 We observed that telomerase activity of iECs, as assessed by a telomeric repeat amplification protocol, was significantly greater compared with adult fibroblasts or primary adult human ECs (Figure 3I and 3J). However, despite higher telomerase activity, iECs did not show significantly elevated expression of the pluripotency transcription factor OCT4 or the pro- to oncogene c-MYC compared with fibroblasts (Figure VA and VB in the online-only Data Supplement).
Fibroblasts Also Give Rise to Erythroblasts by Intermediate CD34+ Progenitors
Fibroblast-derived CD34+ progenitors were cultured in erythroblast induction medium for 4 days to assess the generation of erythroblasts. Unlike iECs, which remained adherent to the substrate, emerging erythroblasts detached from the CD34+ cell layer. Hema 3-staining cytospin preparations of iErythroblasts showed morphology reflecting different stages of maturation from basophilic erythroblasts to polychromatic erythroblasts (Figure 4A). To address whether iErythroblasts expressed an erythroid marker, we stained the cytospin preparations for the erythroid marker glycophorin CD235a. We found that 7.1±1% of nucleated were CD235a+ (Figure 4B and 4C). Flow cytometry demonstrated that 4.2%±0.8% of cells undergoing erythroblast induction were CD235a+ (Figure 4D and 4E). Gene expression analysis by quantitative polymerase chain reaction showed expression of human hemoglobin beta and hemoglobin gamma, confirming the erythroid nature of the generated human cells (Figure 4F). The expression levels of hematopoietic transcriptional factors –RUNX1 and PU.1, as well as the erythroblast transcription factor GATA1,24,25 were also markedly enhanced when compared with fibroblasts (Figure 4G). Moreover, erythroid colony-forming-cell assay demonstrated formation of distinct erythroid colonies (Figure 4H and 4I).
Figure 4.
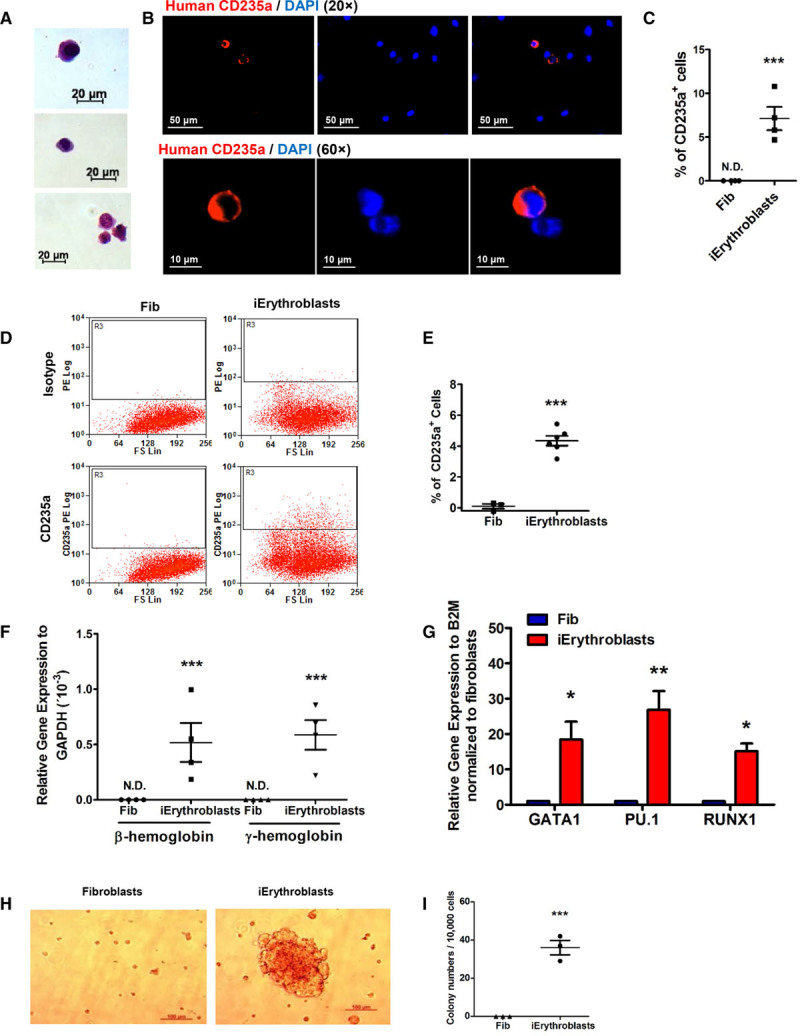
Induction and characterization of induced Erythroblasts (iErythroblasts). A, Representative Hema 3 staining images of iErythroblasts. Scale bars: 20 µm. B and C, Immunocytochemistry analysis of erythrocyte markers CD235a in iErythroblasts: representative immunofluorescence images of iErythroblasts (B) and statistical analysis (C) of percentage of CD235a+ cells in fibroblasts and iErythroblasts (n=4). Scale bars: 20 µm. D and E, FACS analysis of erythrocyte markers CD235a. Representative flow-cytometry plots (D) and statistical analysis (E) of percentage of CD235a+ cells in iErythroblasts and fibroblasts (n=6). F, Quantitative polymerase chain reaction (PCR) analysis of hemoglobin β and hemoglobin γ by Taqman assay in the fibroblasts and iErythroblasts (n=3). G, Quantitative PCR analysis of transcriptional factors of hematopoietic cells GATA1, RUNX1, and PU.1 in fibroblasts and iErythroblasts (n=4). H and I, Erythroid colony-forming cell (CFC) assays. Representative erythroid colony images (H) and statistical analysis (I) of erythroid colony numbers of fibroblasts and iErythroblasts (n=3). Data are presented as mean±SE; *P<0.05, **P<0.01, ***P<0.001, compared with fibroblasts using the Student t test in C, E, and I and the Student t test with multiple comparisons (Bonferroni correction) in F and G.
SOX17 Mediates Generation of iECs From CD34+ Progenitors
We next evaluated expression of vascular developmental genes during iEC induction as shown in a quantitative real-time polymerase chain reaction heat map (Figure 5A). We observed that the transcriptional regulator SOX17 (a member of Sry-transcription factor family) was significantly upregulated during iECs induction. The transcription factor ETV2 (also known as ER71), which when overexpressed induced fibroblast-to-endothelial transition,11,12 was not significantly upregulated during lineage conversion of CD34+ progenitors to iECs.
Figure 5.
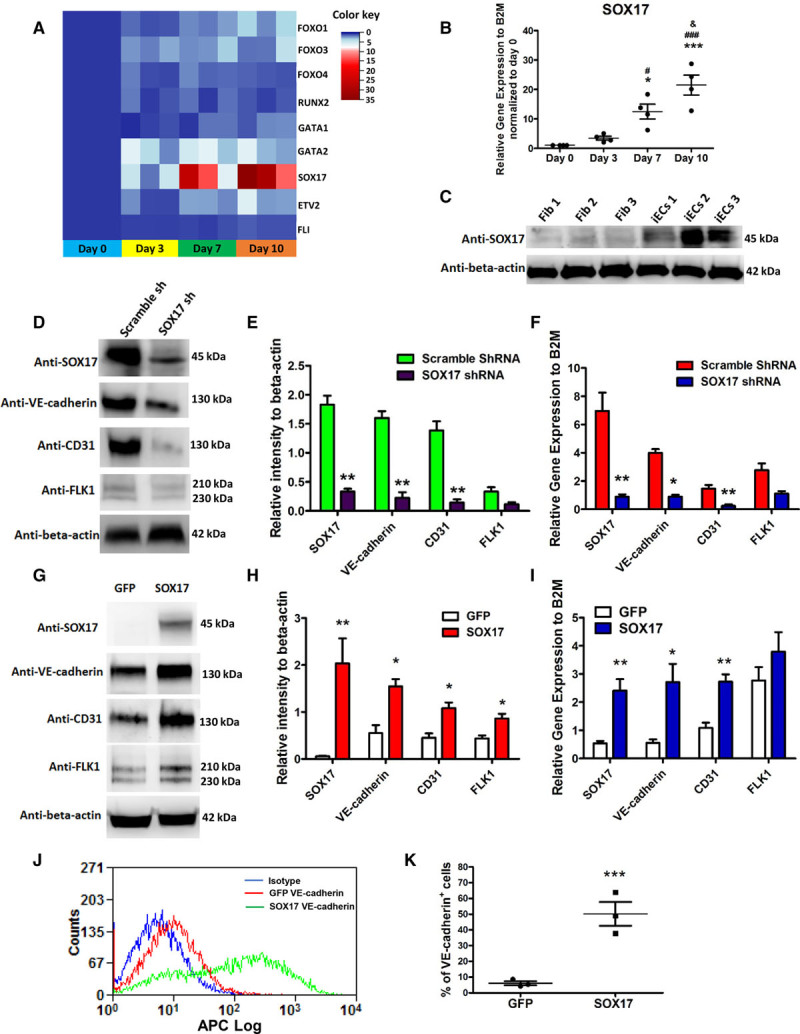
SOX17 is required for the induction of induced endothelial cells (iECs) from CD34+ progenitors. A, Heat map of endothelial developmental genes during iEC induction (iECs at days 0, 3, 7, and 10) by quantitative real-time polymerase chain reaction (PCR). The color key indicates gene expression level relative to B2M (beta-2-microglobulin) normalized to iECs at day 0. Blue to red color represents low to high expression level. B and C, The expression level of SOX17 in iECs. The time course of mRNA level of SOX17 during the induction of iECs (n=4). B, Immunoblot of SOX17 in adult fibroblasts and iECs (C). D through F, The representative immunoblots (D) and their statistical analysis (E), and the quantitative PCR analysis (F) of SOX17 and EC-specific markers VE-cadherin, CD31, and FLK1 in iECs infected with lentivirus-expressing SOX17-shRNA or scramble shRNA (n=4). G–I, The representative immunoblots (G) and their statistical analysis (H), and the quantitative PCR analysis (I) of SOX17 and EC-specific markers VE-cadherin, CD31, and FLK1 in iECs infected with retrovirus-expressing SOX17-GFP or GFP (n=4). J and K, Representative flow-cytometry overlay plots of the endothelial markers VE-cadherin (J) and statistical analysis (K) of percentage of VE-cadherin–positive cells in iECs overexpressed with SOX17-GFP or GFP. Data are presented as mean±SE; *P<0.05, ***P<0.001, compared with day 0 in A; #P<0.05, ###P<0.001, compared with day 3 in A; &P<0.05 compared with day 7 in A. *P<0.05, **P<0.01, compared with control in E, F, H, and I. Using repeated measures 1-way analysis of variance in B, the Student t test with multiple comparisons (Bonferroni Correction) in E, F, H, and I, and the Student t test in K.
SOX17, a member of the Sry-related high-mobility group domain family F (SoxF) transcriptional factors, is a key regulator of EC and hematopoietic development during embryogenesis. Thus, we next investigated whether the transcription factor SOX17 identified earlier was required for conversion of CD34+ progenitors to iECs. SOX17 progressively increased during the 10-day induction period (Figure 5B). Immunoblotting confirmed upregulation of SOX17 in iECs compared with fibroblasts (Figure 5C). To assess whether expression of SOX17 was required for induction of iECs, lentiviral shRNAs were used to deplete SOX17 during EC induction phase. CD34+ progenitors were infected with lentiviruses encoding SOX17 shRNA or scramble shRNA on day 1 with 95% transduction efficiency (Figure VIA through VIC in the online-only Data Supplement). Using multiple shRNA constructs, we identified a SOX17-shRNA, which achieved >90% SOX17 depletion (Figure VID in the online-only Data Supplement). Depletion of SOX17 prevented the differentiation of CD34+ progenitors to iECs as demonstrated by suppression of the EC-specific markers (VE-cadherin, CD31, and FLK1) at both mRNA and protein levels when compared with scrambled shRNA (Figure 5D through 5F).
Based on these findings, we next determined whether overexpression of SOX17 would enhance EC lineage conversion. CD34+ progenitors undergoing conversion were transduced with retroviruses encoding SOX17-GFP or GFP on day 1. We observed significant increases in mRNA and protein levels of VE-cadherin, CD31, and FLK1 when compared with controls (Figure 5G through 5I). Quantitative analysis with flow cytometry showed that overexpression of SOX17 enhanced EC differentiation ≈9-fold as evidenced by increased fraction of VE-cadherin–positive cells when compared with using only growth factors for EC conversion (50.9±3% versus 6.1±2%) (Figure 5J and 5K). Thus, conversion of CD34+ progenitors into iECs was markedly enhanced by activation of the transcription factor SOX17.
SOX17 Inhibits Generation of iErythroblasts From CD34+ Progenitors
We next examined whether SOX17 also regulates transition of CD34+ progenitors, the point of bifurcation, into iErythroblasts. Here we found that the expression of SOX17 increased in CD34+ progenitors when compared with the parent fibroblasts but progressively decreased during the induction of iErythroblasts (Figure 6A), consistent with the role of SOX17 in favoring EC fate conversion at the CD34+ progenitor bifurcation state described earlier. To explore the role of SOX17 in iErythroblast generation, SOX17 was depleted using shRNA in fibroblasts before dedifferentiation. Depletion of SOX17 increased the expression of the hematopoietic transcription factors RUNX1 and PU.1 as well as erythropoiesis-specific regulator GATA1 during erythroblast induction (Figure 6B). Flow cytometry confirmed increased erythropoiesis on SOX17 depletion trending toward increased erythropoiesis (Figure 6C and 6D). In addition, we observed that depleting SOX17 significantly increased the formation of erythroid colonies (Figure 6E and 6F), consistent with the role of SOX17 in directing the fate of CD34+ progenitors to either iECs or erythroblasts; that is, the presence of SOX17 favored the iEC lineage, whereas its downregulation enhanced erythroid specification, indicative of the rheostat function of SOX17 at the CD34 bifurcation point.
Figure 6.
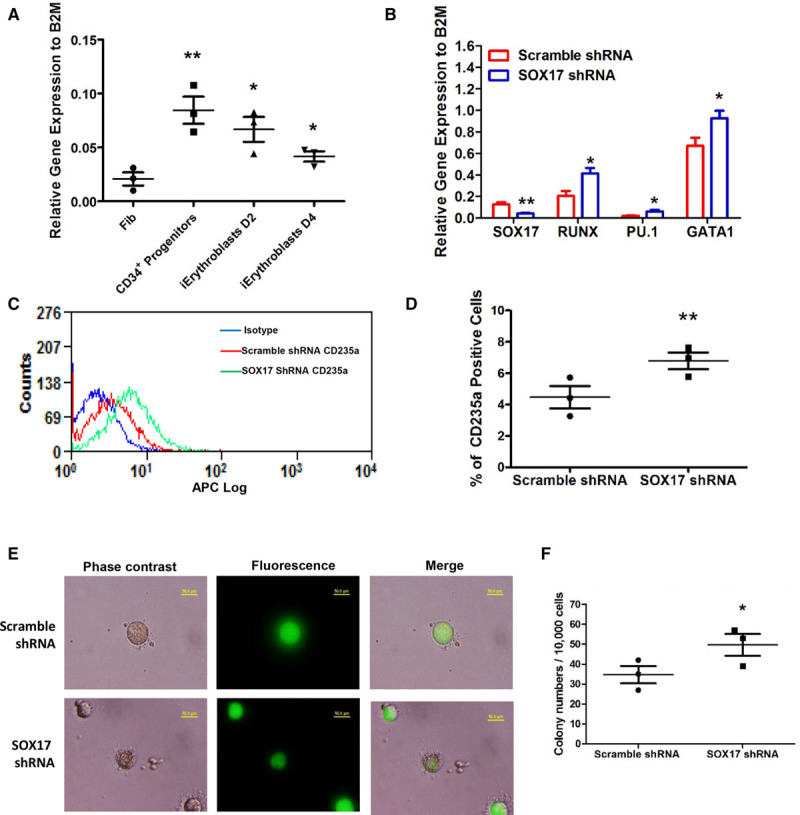
Depletion of SOX17 facilitates the induction of erythroblasts from CD34+ progenitors. A, Quantitative polymerase chain reaction (PCR) analysis of SOX17 in fibroblasts, CD34+ progenitors, and induced Erythroblasts (iErythroblasts) at days 2 and 4 (n=3). B, The gene expression levels of hematopoietic transcription factors (GATA1, PU.1, RUNX) and SOX17 in iErythroblasts infected with lentivirus-expressing SOX17-1shRNA using scramble shRNA as control. C and D, Representative flow-cytometry overlay plots of the erythroblasts/erythrocytes markers human CD235a (C), and statistical analysis (D) of percentage of CD235a+ cells in iErythroblasts infected with lentivirus-expressing SOX17-shRNA or scramble shRNA. E and F, Representative images (E) and statistical analysis (F) of erythroid colony-forming cell (CFC) assays in iErythroblasts infected with SOX17-shRNA or scramble shRNA. The infected cells expressed GFP because the pLL3.7-GFP lentivectors were applied to express SOX17-shRNA or scramble shRNA. Data are presented as mean±SE; *P<0.05, **P<0.01, compared with fibroblasts in A, compared with scramble shRNA in B, D, and F, using 1-way analysis of variance in A, Student t test with multiple comparisons (Bonferroni Correction) in B, and the Student t test in D and F.
CD34+ Progenitors Form Blood Vessels and Erythrocytes In Vivo and Restore Myocardial Function in Mice
We next assessed the in vivo significance of human fibroblast-derived CD34+ progenitors in giving rise to iECs and iErythroblasts. We injected growth factor-reduced Matrigel containing CD34+ progenitors, fibroblasts, or primary human ECs subcutaneously into immune deficient NOD-SCID mice. Matrigel plugs explanted after 1 week showed that implantation of either primary aortic human ECs (controls) or CD34+ progenitors induced formation of blood vessels, whereas blood vessels did not form after implantation of fibroblasts alone (Figure 7A through 7C)
Figure 7.
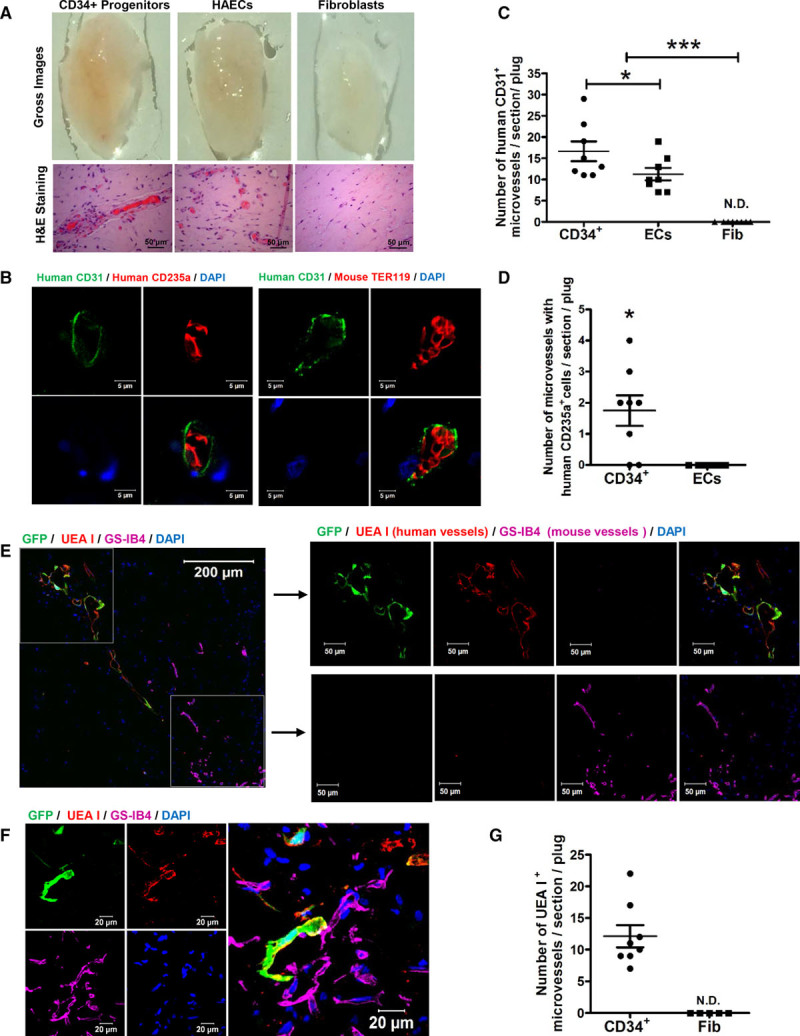
In vivo angiogenesis of fibroblast-derived CD34+ progenitors. A, Representative gross images (upper) and H&E staining images of section (lower) of explanted gels at day 7 of the angiogenesis assay. B–D, Representative confocal micrographs (B) of immunohistochemistry staining of human CD235a (red, left) or mouse TER119 (red, right) and anti-human CD31 (green) in the explanted plugs of CD34+ progenitors. Nuclei stained by DAPI. Scale bars, 5 µm. C and D, Quantification and statistical analysis of human vessels (CD31+) in the explanted plugs of CD34+ progenitors, endothelial cells (ECs), or fibroblasts (C), and the composition of vessels (CD31+) with human RBC (CD235a+) in the explanted plugs of CD34+ progenitors and ECs (D). N.D.indicates not detected. E through G, The analysis of functionality of human vessels formed by GFP-labeled CD34+ progenitors in the angiogenesis plugs. UEA I (labels human endothelium) and GS-IB4 (labels mouse endothelium) were intravenously injected 1 hour before explantation of plugs. Representative confocal images of human vessels (red) and mouse host vessels (magenta) (E), and integration of human vessels (red) and mouse host vessels (magenta) in the plugs implanted with human CD34+ progenitors (F). The quantification of UEA I+ vessels in the plugs implanted with CD34+ progenitors and ECs (G). Data are presented as mean±SE; *P<0.05, ***P<0.001, compared with fibroblasts in C, compared with ECs in C, D, and G, using 1-way analysis of variance in C and the Student t test in D and G.
To evaluate whether the ECs lining the newly formed blood vessels and erythrocytes in the Matrigel plug were derived from the implanted human CD34+ progenitors or host mouse vasculature, we used a human-specific CD31 antibody to stain human vessels. We also used a human-specific CD235a antibody and a mouse-specific TER119 antibody to stain human and mouse erythrocytes, respectively, after confirming the human species specificity (Figure VII in the online-only Data Supplement). Staining of explanted Matrigel plugs containing CD34+ progenitors demonstrated that vessels were positive for human CD31 and had greater angiogenesis than those implanted with mature ECs (Figure 7B and 7C). We further found that a small fraction of erythrocytes within the vessels stained positive for human CD235a (Figure 7D). We observed that the newly formed human blood vessels predominantly contained murine erythrocytes, indicating communication between the host mouse vasculature and newly formed vessels derived from human CD34+ progenitors. It is important to note that Matrigel plugs containing primary human ECs showed presence of erythrocytes derived only from host mice (Figure VIIIA and VIIIB in the online-only Data Supplement).
To further demonstrate the functionality of formed human vessels, fluorescence-labeled lectins that stain species-specific endothelium were injected intravenously before harvesting Matrigel implants containing GFP-labeled CD34+ progenitors or GFP-labeled fibroblasts. We used Rhodamine-labeled UEA I to assess functional human endothelium and DyLight 649-labeled GS-IB4 for mouse endothelium.26 Colocalization of GFP and UEA I+ demonstrated the functionality of CD34+ progenitor-derived human vessels (Figure 7E), whereas areas without GFP+/UEA I+ cells predominantly consisted of GS-IB4+ host mouse vessels (Figure 7E). It is important to note that we identified selected areas of close proximity between human CD34+ progenitor cells and mouse host GS-IB4+ vessels (Figure 7F and Movie I in the online-only Data Supplement), which likely represented communications between the host vasculature and neovessels from the implanted cells, thus explaining why systemic intravenous injection of a human-specific lectin into the mice was able to stain the CD34+ progenitor-derived human vessels. No UEA I+ human vessels were detected in plugs implanted with fibroblasts (Figure 7G). We also found examples of GFP+ erythrocytes (Figure IX in the online-only Data Supplement), further confirming the capacity of CD34+ progenitors to mature into erythrocytes in vivo.
To assess the teratoma-forming capacity of CD34+ progenitors, immune deficient NOD-SCID mice were implanted with CD34+ progenitors or iPSCs (positive control) and observed for 8 weeks. We found that the implantation of iPSCs for 8 weeks formed teratomas, whereas the CD34+ progenitors did not form any teratomas, thus underscoring their safety and lack of pluripotency (Figure XA through XD in the online-only Data Supplement).
We next used a mouse MI model to assess whether CD34+ progenitors enhance perfusion and cardiac function. We injected a Matrigel suspension containing either CD34+ progenitors or control fibroblasts in the peri-infarct area after left coronary artery ligation. Masson trichrome staining of a cross-section of hearts on day 14 after MI showed reduced fibrosis in hearts injected with CD34+ progenitors (Figure 8A). Heart weight/body weight ratio was also decreased in mice receiving CD34+ progenitors (Figure 8B), indicating suppression of adverse remodeling after MI. Echocardiography demonstrated significantly improved cardiac contractility as measured by ejection fraction (Figure 8C) and fractional shortening (Figure 8D) at 1 and 2 weeks after MI in hearts receiving CD34+ progenitors when compared with hearts receiving control fibroblasts. CD34+ progenitor implantation also improved morphological and hemodynamic indices as assessed by echocardiography (Table I in the online-only Data Supplement). CD34+ cells were transduced with GFP before transplantation to track the fate of implanted cells. Immunofluorescence demonstrated that the implantation of CD34+ progenitors resulted in formation of human CD31+ blood vessels, whereas implantation of human fibroblasts formed none (Figure XIA through XIC in the online-only Data Supplement). We also observed that human CD31+ blood vessels were integrated with host mouse CD31+ vasculature (Figure XIB and Movie II in the online-only Data Supplement).
Figure 8.
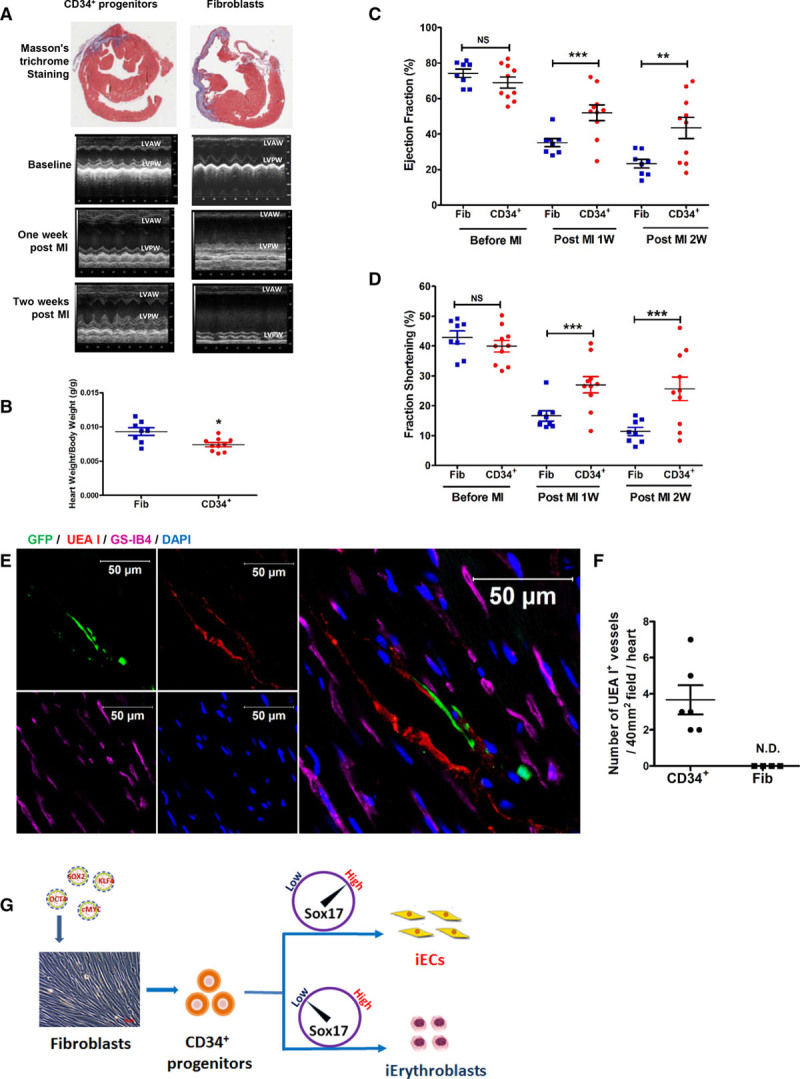
Regenerative potential of fibroblast-derived CD34+ progenitors after myocardial infarction (MI). A, Images of Masson’s trichrome staining of myocardial cross-sections 2 weeks after MI and M-mode echocardiography images at baseline, 1 week after MI, and 2 weeks after MI. B–D, The statistical analysis of heart weight/body weight (B) 2 weeks after MI with CD34+ progenitor or fibroblast treatment, ejection fraction (C), and fraction shortening (D) at baseline (before MI), 1 week after MI, and 2 weeks after MI with CD34+ progenitor or fibroblast treatment.(E, UEA I and GS-IB4 (intravenously injected 1 hour before heart explantation). Representative confocal micrographs of myocardial cross-section 2 weeks after MI implanted with GFP-prelabeled human CD34+ progenitor treatment. Nuclei stained with DAPI (blue). Scale bars, 50 µm. F, Human vessels (UEA I+, red) were quantified per 40 mm2 visual field in sections of the heart that were in proximity to the injection sites 2 weeks after MI with human CD34+ progenitor (n=6) or fibroblast (n=4) treatment. N.D. indicates not detected. Data are presented as mean±SE; *P<0.05, **P<0.01, ***P<0.001, compared with fibroblasts using the Student t test in B and F, and repeated measures 2-way analysis of variance in C and D. G, Overview of a differential role of SOX17 in the induction of endothelial cells and erythroblasts from CD34+ progenitors.
To further confirm the functionality of human neovessels in the myocardium, fluorescence-labeled UEA I and GS-IB4 were intravenously injected before harvesting the heart. We observed that GFP colocalized with UEA I (Figure 8E and 8F) demonstrated that implantation of CD34+ progenitors formed functional human vessels in the myocardium after MI.
Discussion
Here we demonstrate a novel approach for the reprogramming of human fibroblasts into ECs that also gives rise to erythroblasts by the dedifferentiation into intermediate CD34+ progenitors. We identified the developmental transcription factor SOX17 as a tunable rheostat-like switch converting CD34+ cells to both lineages. Implantation of human CD34+ progenitors in immune deficient NOD-SCID mice gave rise to human ECs capable of forming blood vessels carrying erythrocytes, markedly enhancing human microvessel generation and cardiac function after MI. Unlike pluripotent stem cells, fibroblasts dedifferentiated to CD34+ progenitors did not form any teratomas after implantation, thus underscoring their safety for translational applications in vascular engineering and revascularization. It is important to note that systemic intravenous injections of human and mouse lectins into the recipient mice demonstrated that the neovessels formed by the implanted human fibroblast-derived CD34+ progenitor cells are functional and communicate with the host mouse vasculature.
Previous approaches to generating ECs from fibroblasts involved the ectopic expression of EC-specific transcription factors,11,12 which have relatively low yield and persistence of fibroblast markers, thus limiting their therapeutic potential. Other approaches involved dedifferentiation by Yamanaka factors without genetically directing cells toward the EC lineage.8–10 In the present study, we show a third strategy involving the generation of intermediate dedifferentiated CD34+ progenitors and then genetically modulating the transcription factor SOX17 to direct lineage fate to iECs and iErythroblasts. Thus, in this method, the intermediate CD34+ progenitors can be directed to form both iECs and erythroid lineage cells with therapeutic potential. After purification for CD34+ cells using magnetic beads, not all cells expressed CD34 by subsequent flow-cytometric analysis, which may be indicative of CD34 internalization or differences in the specificity and sensitivity between magnetic bead sorting and flow-cytometry analysis.
To define transcriptional mechanisms underlying the generation of iECs and erythrocytes, we identified by transcriptomic analysis of CD34+ cells the requisite role of SOX17 as an essential transcription factor responsible for the conversion. SOX17, a member of the Sry-related high-mobility group domain family F (SoxF) transcriptional factor,27 is a key regulator of endothelial and hematopoietic developmental during embryogenesis.28–32 We showed that SOX17 was required for endothelial lineage conversion and that its overexpression markedly enhanced the generation of iECs from fibroblasts. Further, depletion of SOX17 facilitated the generation of erythrocytes. The regulatory role of SOX17 at the CD34 bifurcation point thus specified the conversion to iEC and iErythroblast fates. This finding is reminiscent of the role of SOX17 during embryonic development, during which SOX17 regulates EC to hematopoietic fate switch.33 Specifically, loss of SOX17-mediated repression of hematopoietic transcription factors Runx1 and Gata2 prevented EC generation and augmented blood cell generation.33
The described method of iEC and iErythroblast generation has a distinct advantage in that it delineates for the first time the generation of cells capable of forming blood vessels and erythrocytes in vivo. We demonstrated that transplantation of CD34+ progenitors in subcutaneous Matrigel plugs induced formation of blood vessels comprised of human ECs and perfused with human erythrocytes. A likely mechanism by which transplanted CD34+ progenitors give rise to both cells may be the release of endothelial differentiation cues in ischemic niche such as vascular endothelial growth factor and the presence of erythropoietin in the circulating blood,34–37 which in turn regulate SOX17 expression to optimize generation of both cells.
Our transcriptomic analysis of CD34+ progenitors and iECs indicated that iECs exhibited the gene expression profile similar to adult primary ECs, whereas CD34+ progenitors retained fibroblast signatures such as collagen1a and collagen3a. Gain of EC identity and concomitant loss of fibroblast identity occurred during the transition from the CD34+ progenitor to the iEC state. Although CD34+ progenitors exhibited residual fibroblast gene expression, implantation of CD34+ progenitors in the infarcted myocardium reduced fibrosis, suggesting that the imprinted fibroblast memory was lost in situ in the ischemic environment.
Previous studies described the role of ETV2 (ER71) in inducing fibroblast to endothelial conversion.11,12 However, we did not find its significant upregulation. Our approach was guided by unbiased transcriptomic analysis of endogenous transcription factors upregulated during dedifferentiation and lineage conversion. Although endogenous ETV2 was not upregulated during dedifferentiation, its overexpression can direct fibroblasts toward EC lineage.11,12 Our results are in accord with an interesting lineage tracing study showing that p53 signaling mediated in situ conversion of endogenous fibroblasts to ECs leads to improved cardiac function after MI,38 although these studies did not address whether there was an intermediate CD34+ progenitor cell responsible for EC conversion. Recent studies also show that innate immunity is a critical regulator for reprogramming of fibroblasts into the pluripotent state as well as transdifferentiating fibroblasts to ECs.39,40 Because MI activates inflammatory pathways and innate immunity, it is possible that this serves as a cue for implanted CD34+ cells to initiate in situ reprogramming to iECs and iErythroblasts.
SOX17, together with Notch1, regulates the embryonic EC-hematopoietic fate transition.33,41 SOX17 prevented hematopoietic cell fate of hemangioblasts and hemogenic ECs by repressing the expression of hematopoietic transcription factors, whereas loss of SOX17 instructed differentiation toward hematopoietic fate.33 We did not examine the role of Notch1 in regulating CD34+ cell to iEC transition because our transcriptomic analysis showed that it did not increase during transdifferentiation. In addition, SOX17 was selectively expressed in mouse embryonic and adult arterial ECs, where it was required for maintaining arterial identity42 and integrity43 consistent with our finding of the essential importance of SOX17 in mediating generation of iECs. It has also been recently shown that SOX17 expression can enhance the maturation and engraftment of endothelial cells derived from amniotic cells.44 It is therefore possible that the SOX17 expression in fibroblast-derived ECs similarly helped to mature and engraft the ECs derived from the progenitors we implanted.
In addition to identifying the bilineage differentiation potential of fibroblast-derived CD34+ progenitors and obligatory role of SOX17 in iEC and iErythroblast generation, we found that dedifferentiation upregulated telomerase activity of iECs. This finding may be the result of the initial introduction of pluripotency factors that activated embryonic transcriptional programs. Activation of telomerase may be important for autologous cell therapy, which requires extensive in vitro cell expansion because telomeres undergo shortening during cell expansion.21–23 We found that dedifferentiation in fact upregulated telomerase in the CD34+ progenitors undergoing lineage conversion, which may induce generation of blood vessels with greater survival advantage in ischemic tissue. It is interesting to note that the dedifferentiation of adult ECs to vascular progenitors has also been shown to generate cells with significant vascularization potential,45 which may reflect some resilience to senescence.
We cannot exclude the possibility that the CD34+ progenitor cells derived from fibroblasts contain subpopulations of cells that may be poised to give rise only to endothelial or erythroid lineage cells. However, our flow-cytometric analysis of the CD34+ population did not show any evidence of endothelial or erythroid markers, suggesting that they are uncommitted at this stage.
In summary, our studies establish a novel role for the transcription factor SOX17 as a critical endogenous regulator of the transition from fibroblasts to ECs and to erythrocytes by the dedifferentiation to CD34+ progenitors (Figure 8G). Dedifferentiation of adult fibroblasts in part recapitulates embryonic development as evidenced by the generation of progenitors that can give rise to endothelial and erythroid cells and express telomerase. CD34+ progenitors in situ gave rise to functional human ECs and erythrocytes and displayed profound vasculogenesis and restored cardiac function after transplantation, thus highlighting the potential therapeutic role of such progenitors.
Acknowledgments
L.Z., A.J., Z.H., G.M., P.T., and B.H. performed the experiments. L.Z, A.B.M., and J.R. designed the experiments. L.Z., A.J., B.H., and J.R. analyzed and interpreted the data. L.Z., A.B.M., and J.R. wrote the article. The authors thank Jillian Koziel for assistance in performing the telomerase activity assays and Yang Yuan for assistance in histological analysis.
Sources of Funding
This work was supported in part by National Institutes of Health grants R01GM094220, R01HL118068, T32HL007829, and R01HL090152.
Disclosures
None.
Supplementary Material
Footnotes
The online-only Data Supplement is available with this article at http://circ.ahajournals.org/lookup/suppl/doi:10.1161/CIRCULATIONAHA.116.025722/-/DC1.
Circulation is available at http://circ.ahajournals.org.
Clinical Perspective
What Is New?
Adult human skin fibroblasts can be dedifferentiated to intermediate CD34+ progenitors, which in turn give rise to endothelial cells and erythrocytes.
Lineage conversion of fibroblasts by partial dedifferentiation recapitulates in part the embryonic development of the vasculature as evidenced by upregulation of the anti-aging enzyme telomerase and the bilineage potential of the generated progenitors.
The transcription factor SOX17 functions as a switch that regulates cell fate of CD34+ progenitors toward an endothelial versus erythroid lineage.
Implanted fibroblast-derived CD34+ progenitors stably engraft to form functional human blood vessels in mice that improve cardiac function after myocardial infarction.
What Are the Clinical Implications?
Dedifferentiation of fibroblasts to multipotent progenitors can generate patient-specific functional endothelial cells.
When human fibroblast-derived progenitors are implanted in a mouse model of myocardial infarction, they are able to improve cardiac function.
The molecular switch SOX17 provides a means to optimize the generation of endothelial cells for vascular tissue regeneration or disease modeling.
The upregulation of telomerase suggests that fibroblast dedifferentiation may avoid premature aging of the cells during the lineage conversion process, which could be especially helpful when deriving personalized endothelial cells from skin fibroblasts of older patients.
References
- 1.Zhang L, Malik AB, Rehman J. Reprogramming fibroblasts to endothelial cells: converted or born again? Circulation. 2014;130:1136–1138. doi: 10.1161/CIRCULATIONAHA.114.012540. doi: 10.1161/CIRCULATIONAHA.114.012540. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Capellera-Garcia S, Pulecio J, Dhulipala K, Siva K, Rayon-Estrada V, Singbrant S, Sommarin MN, Walkley CR, Soneji S, Karlsson G, Raya Á, Sankaran VG, Flygare J. Defining the minimal factors required for erythropoiesis through direct lineage conversion. Cell Rep. 2016;15:2550–2562. doi: 10.1016/j.celrep.2016.05.027. doi: 10.1016/j.celrep.2016.05.027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Efe JA, Hilcove S, Kim J, Zhou H, Ouyang K, Wang G, Chen J, Ding S. Conversion of mouse fibroblasts into cardiomyocytes using a direct reprogramming strategy. Nat Cell Biol. 2011;13:215–222. doi: 10.1038/ncb2164. doi: 10.1038/ncb2164. [DOI] [PubMed] [Google Scholar]
- 4.Huang P, He Z, Ji S, Sun H, Xiang D, Liu C, Hu Y, Wang X, Hui L. Induction of functional hepatocyte-like cells from mouse fibroblasts by defined factors. Nature. 2011;475:386–389. doi: 10.1038/nature10116. doi: 10.1038/nature10116. [DOI] [PubMed] [Google Scholar]
- 5.Ieda M, Fu JD, Delgado-Olguin P, Vedantham V, Hayashi Y, Bruneau BG, Srivastava D. Direct reprogramming of fibroblasts into functional cardiomyocytes by defined factors. Cell. 2010;142:375–386. doi: 10.1016/j.cell.2010.07.002. doi: 10.1016/j.cell.2010.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Vierbuchen T, Ostermeier A, Pang ZP, Kokubu Y, Südhof TC, Wernig M. Direct conversion of fibroblasts to functional neurons by defined factors. Nature. 2010;463:1035–1041. doi: 10.1038/nature08797. doi: 10.1038/nature08797. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Panopoulos AD, Ruiz S, Izpisua Belmonte JC. iPSCs: induced back to controversy. Cell Stem Cell. 2011;8:347–348. doi: 10.1016/j.stem.2011.03.003. doi: 10.1016/j.stem.2011.03.003. [DOI] [PubMed] [Google Scholar]
- 8.Kurian L, Sancho-Martinez I, Nivet E, Aguirre A, Moon K, Pendaries C, Volle-Challier C, Bono F, Herbert JM, Pulecio J, Xia Y, Li M, Montserrat N, Ruiz S, Dubova I, Rodriguez C, Denli AM, Boscolo FS, Thiagarajan RD, Gage FH, Loring JF, Laurent LC, Izpisua Belmonte JC. Conversion of human fibroblasts to angioblast-like progenitor cells. Nat Methods. 2013;10:77–83. doi: 10.1038/nmeth.2255. doi: 10.1038/nmeth.2255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Li J, Huang NF, Zou J, Laurent TJ, Lee JC, Okogbaa J, Cooke JP, Ding S. Conversion of human fibroblasts to functional endothelial cells by defined factors. Arterioscler Thromb Vasc Biol. 2013;33:1366–1375. doi: 10.1161/ATVBAHA.112.301167. doi: 10.1161/ATVBAHA.112.301167. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Margariti A, Winkler B, Karamariti E, Zampetaki A, Tsai TN, Baban D, Ragoussis J, Huang Y, Han JD, Zeng L, Hu Y, Xu Q. Direct reprogramming of fibroblasts into endothelial cells capable of angiogenesis and reendothelialization in tissue-engineered vessels. Proc Natl Acad Sci U S A. 2012;109:13793–13798. doi: 10.1073/pnas.1205526109. doi: 10.1073/pnas.1205526109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Han JK, Chang SH, Cho HJ, Choi SB, Ahn HS, Lee J, Jeong H, Youn SW, Lee HJ, Kwon YW, Cho HJ, Oh BH, Oettgen P, Park YB, Kim HS. Direct conversion of adult skin fibroblasts to endothelial cells by defined factors. Circulation. 2014;130:1168–1178. doi: 10.1161/CIRCULATIONAHA.113.007727. doi: 10.1161/CIRCULATIONAHA.113.007727. [DOI] [PubMed] [Google Scholar]
- 12.Morita R, Suzuki M, Kasahara H, Shimizu N, Shichita T, Sekiya T, Kimura A, Sasaki K, Yasukawa H, Yoshimura A. ETS transcription factor ETV2 directly converts human fibroblasts into functional endothelial cells. Proc Natl Acad Sci U S A. 2015;112:160–165. doi: 10.1073/pnas.1413234112. doi: 10.1073/pnas.1413234112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hirschi KK. Hemogenic endothelium during development and beyond. Blood. 2012;119:4823–4827. doi: 10.1182/blood-2011-12-353466. doi: 10.1182/blood-2011-12-353466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Lancrin C, Sroczynska P, Stephenson C, Allen T, Kouskoff V, Lacaud G. The haemangioblast generates haematopoietic cells through a haemogenic endothelium stage. Nature. 2009;457:892–895. doi: 10.1038/nature07679. doi: 10.1038/nature07679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Palis J, Robertson S, Kennedy M, Wall C, Keller G. Development of erythroid and myeloid progenitors in the yolk sac and embryo proper of the mouse. Development. 1999;126:5073–5084. doi: 10.1242/dev.126.22.5073. [DOI] [PubMed] [Google Scholar]
- 16.Lu SJ, Feng Q, Caballero S, Chen Y, Moore MA, Grant MB, Lanza R. Generation of functional hemangioblasts from human embryonic stem cells. Nat Methods. 2007;4:501–509. doi: 10.1038/nmeth1041. doi: 10.1038/nmeth1041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Peters A, Burridge PW, Pryzhkova MV, Levine MA, Park TS, Roxbury C, Yuan X, Péault B, Zambidis ET. Challenges and strategies for generating therapeutic patient-specific hemangioblasts and hematopoietic stem cells from human pluripotent stem cells. Int J Dev Biol. 2010;54:965–990. doi: 10.1387/ijdb.093043ap. doi: 10.1387/ijdb.093043ap. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sidney LE, Branch MJ, Dunphy SE, Dua HS, Hopkinson A. Concise review: evidence for CD34 as a common marker for diverse progenitors. Stem Cells. 2014;32:1380–1389. doi: 10.1002/stem.1661. doi: 10.1002/stem.1661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Paul JD, Coulombe KL, Toth PT, Zhang Y, Marsboom G, Bindokas VP, Smith DW, Murry CE, Rehman J. SLIT3-ROBO4 activation promotes vascular network formation in human engineered tissue and angiogenesis in vivo. J Mol Cell Cardiol. 2013;64:124–131. doi: 10.1016/j.yjmcc.2013.09.005. doi: 10.1016/j.yjmcc.2013.09.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Daneshjou N, Sieracki N, van Nieuw Amerongen GP, Conway DE, Schwartz MA, Komarova YA, Malik AB. Rac1 functions as a reversible tension modulator to stabilize VE-cadherin trans-interaction. J Cell Biol. 2015;208:23–32. doi: 10.1083/jcb.201409108. doi: 10.1083/jcb.201409108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Armanios M. Telomeres and age-related disease: how telomere biology informs clinical paradigms. J Clin Invest. 2013;123:996–1002. doi: 10.1172/JCI66370. doi: 10.1172/JCI66370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Blasco MA. Telomere length, stem cells and aging. Nat Chem Biol. 2007;3:640–649. doi: 10.1038/nchembio.2007.38. doi: 10.1038/nchembio.2007.38. [DOI] [PubMed] [Google Scholar]
- 23.Rando TA, Chang HY. Aging, rejuvenation, and epigenetic reprogramming: resetting the aging clock. Cell. 2012;148:46–57. doi: 10.1016/j.cell.2012.01.003. doi: 10.1016/j.cell.2012.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Takeuchi M, Fuse Y, Watanabe M, Andrea CS, Takeuchi M, Nakajima H, Ohashi K, Kaneko H, Kobayashi-Osaki M, Yamamoto M, Kobayashi M. LSD1/KDM1A promotes hematopoietic commitment of hemangioblasts through downregulation of Etv2. Proc Natl Acad Sci U S A. 2015;112:13922–13927. doi: 10.1073/pnas.1517326112. doi: 10.1073/pnas.1517326112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Hewitt KJ, Sanalkumar R, Johnson KD, Keles S, Bresnick EH. Epigenetic and genetic mechanisms in red cell biology. Curr Opin Hematol. 2014;21:155–164. doi: 10.1097/MOH.0000000000000034. doi: 10.1097/MOH.0000000000000034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Kang KT, Allen P, Bischoff J. Bioengineered human vascular networks transplanted into secondary mice reconnect with the host vasculature and re-establish perfusion. Blood. 2011;118:6718–6721. doi: 10.1182/blood-2011-08-375188. doi: 10.1182/blood-2011-08-375188. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Park C, Kim TM, Malik AB. Transcriptional regulation of endothelial cell and vascular development. Circ Res. 2013;112:1380–1400. doi: 10.1161/CIRCRESAHA.113.301078. doi: 10.1161/CIRCRESAHA.113.301078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Choi KD, Vodyanik MA, Togarrati PP, Suknuntha K, Kumar A, Samarjeet F, Probasco MD, Tian S, Stewart R, Thomson JA, Slukvin II. Identification of the hemogenic endothelial progenitor and its direct precursor in human pluripotent stem cell differentiation cultures. Cell Rep. 2012;2:553–567. doi: 10.1016/j.celrep.2012.08.002. doi: 10.1016/j.celrep.2012.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Clarke RL, Yzaguirre AD, Yashiro-Ohtani Y, Bondue A, Blanpain C, Pear WS, Speck NA, Keller G. The expression of Sox17 identifies and regulates haemogenic endothelium. Nat Cell Biol. 2013;15:502–510. doi: 10.1038/ncb2724. doi: 10.1038/ncb2724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lee SH, Lee S, Yang H, Song S, Kim K, Saunders TL, Yoon JK, Koh GY, Kim I. Notch pathway targets proangiogenic regulator Sox17 to restrict angiogenesis. Circ Res. 2014;115:215–226. doi: 10.1161/CIRCRESAHA.115.303142. doi: 10.1161/CIRCRESAHA.115.303142. [DOI] [PubMed] [Google Scholar]
- 31.Nakajima-Takagi Y, Osawa M, Oshima M, Takagi H, Miyagi S, Endoh M, Endo TA, Takayama N, Eto K, Toyoda T, Koseki H, Nakauchi H, Iwama A. Role of SOX17 in hematopoietic development from human embryonic stem cells. Blood. 2013;121:447–458. doi: 10.1182/blood-2012-05-431403. doi: 10.1182/blood-2012-05-431403. [DOI] [PubMed] [Google Scholar]
- 32.Niakan KK, Ji H, Maehr R, Vokes SA, Rodolfa KT, Sherwood RI, Yamaki M, Dimos JT, Chen AE, Melton DA, McMahon AP, Eggan K. Sox17 promotes differentiation in mouse embryonic stem cells by directly regulating extraembryonic gene expression and indirectly antagonizing self-renewal. Genes Dev. 2010;24:312–326. doi: 10.1101/gad.1833510. doi: 10.1101/gad.1833510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lizama CO, Hawkins JS, Schmitt CE, Bos FL, Zape JP, Cautivo KM, Borges Pinto H, Rhyner AM, Yu H, Donohoe ME, Wythe JD, Zovein AC. Repression of arterial genes in hemogenic endothelium is sufficient for haematopoietic fate acquisition. Nat Commun. 2015;6:7739. doi: 10.1038/ncomms8739. doi: 10.1038/ncomms8739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Arany Z, Foo SY, Ma Y, Ruas JL, Bommi-Reddy A, Girnun G, Cooper M, Laznik D, Chinsomboon J, Rangwala SM, Baek KH, Rosenzweig A, Spiegelman BM. HIF-independent regulation of VEGF and angiogenesis by the transcriptional coactivator PGC-1alpha. Nature. 2008;451:1008–1012. doi: 10.1038/nature06613. doi: 10.1038/nature06613. [DOI] [PubMed] [Google Scholar]
- 35.Lemus-Varela ML, Flores-Soto ME, Cervantes-Munguía R, Torres-Mendoza BM, Gudiño-Cabrera G, Chaparro-Huerta V, Ortuño-Sahagún D, Beas-Zárate C. Expression of HIF-1 alpha, VEGF and EPO in peripheral blood from patients with two cardiac abnormalities associated with hypoxia. Clin Biochem. 2010;43:234–239. doi: 10.1016/j.clinbiochem.2009.09.022. doi: 10.1016/j.clinbiochem.2009.09.022. [DOI] [PubMed] [Google Scholar]
- 36.Rankin EB, Biju MP, Liu Q, Unger TL, Rha J, Johnson RS, Simon MC, Keith B, Haase VH. Hypoxia-inducible factor-2 (HIF-2) regulates hepatic erythropoietin in vivo. J Clin Invest. 2007;117:1068–1077. doi: 10.1172/JCI30117. doi: 10.1172/JCI30117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Semenza GL, Nejfelt MK, Chi SM, Antonarakis SE. Hypoxia-inducible nuclear factors bind to an enhancer element located 3’ to the human erythropoietin gene. Proc Natl Acad Sci U S A. 1991;88:5680–5684. doi: 10.1073/pnas.88.13.5680. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Ubil E, Duan J, Pillai IC, Rosa-Garrido M, Wu Y, Bargiacchi F, Lu Y, Stanbouly S, Huang J, Rojas M, Vondriska TM, Stefani E, Deb A. Mesenchymal-endothelial transition contributes to cardiac neovascularization. Nature. 2014;514:585–590. doi: 10.1038/nature13839. doi: 10.1038/nature13839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lee J, Sayed N, Hunter A, Au KF, Wong WH, Mocarski ES, Pera RR, Yakubov E, Cooke JP. Activation of innate immunity is required for efficient nuclear reprogramming. Cell. 2012;151:547–558. doi: 10.1016/j.cell.2012.09.034. doi: 10.1016/j.cell.2012.09.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Sayed N, Wong WT, Ospino F, Meng S, Lee J, Jha A, Dexheimer P, Aronow BJ, Cooke JP. Transdifferentiation of human fibroblasts to endothelial cells: role of innate immunity. Circulation. 2015;131:300–309. doi: 10.1161/CIRCULATIONAHA.113.007394. doi: 10.1161/CIRCULATIONAHA.113.007394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Gama-Norton L, Ferrando E, Ruiz-Herguido C, Liu Z, Liu Z, Guiu J, Islam AB, Lee SU, Yan M, Guidos CJ, López-Bigas N, Maeda T, Espinosa L, Kopan R, Bigas A. Notch signal strength controls cell fate in the haemogenic endothelium. Nat Commun. 2015;6:8510. doi: 10.1038/ncomms9510. doi: 10.1038/ncomms9510. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Corada M, Orsenigo F, Morini MF, Pitulescu ME, Bhat G, Nyqvist D, Breviario F, Conti V, Briot A, Iruela-Arispe ML, Adams RH, Dejana E. Sox17 is indispensable for acquisition and maintenance of arterial identity. Nat Commun. 2013;4:2609. doi: 10.1038/ncomms3609. doi: 10.1038/ncomms3609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Lee S, Kim IK, Ahn JS, Woo DC, Kim ST, Song S, Koh GY, Kim HS, Jeon BH, Kim I. Deficiency of endothelium-specific transcription factor Sox17 induces intracranial aneurysm. Circulation. 2015;131:995–1005. doi: 10.1161/CIRCULATIONAHA.114.012568. doi: 10.1161/CIRCULATIONAHA.114.012568. [DOI] [PubMed] [Google Scholar]
- 44.Schachterle W, Badwe CR, Palikuqi B, Kunar B, Ginsberg M, Lis R, Yokoyama M, Elemento O, Scandura JM, Rafii S. Sox17 drives functional engraftment of endothelium converted from non-vascular cells. Nat Commun. 2017;8:13963. doi: 10.1038/ncomms13963. doi: 10.1038/ncomms13963. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Yin L, Ohanyan V, Pung YF, Delucia A, Bailey E, Enrick M, Stevanov K, Kolz CL, Guarini G, Chilian WM. Induction of vascular progenitor cells from endothelial cells stimulates coronary collateral growth. Circ Res. 2012;110:241–252. doi: 10.1161/CIRCRESAHA.111.250126. doi: 10.1161/CIRCRESAHA.111.250126. [DOI] [PMC free article] [PubMed] [Google Scholar]