

Abstract
Circadian rhythms are generated by an intrinsic cellular mechanism that controls a large array of physiological and metabolic processes. There is erosion in the robustness of circadian rhythms during aging, and disruption of the clock by genetic ablation of specific genes is associated with aging-related features. Importantly, environmental conditions are thought to modulate the aging process. For example, caloric restriction is a very strong environmental effector capable of delaying aging. Intracellular pathways implicating nutrient sensors, such as SIRTs and mTOR complexes, impinge on cellular and epigenetic mechanisms that control the aging process. Strikingly, accumulating evidences indicate that these pathways are involved in both the modulation of the aging process and the control of the clock. Hence, innovative therapeutic strategies focused at controlling the circadian clock and the nutrient sensing pathways might beneficially influence the negative effects of aging.
Introduction
Along evolution, all life forms have adapted to a 24 hours day–night cycle and anticipate circadian (from Latin circa diem, approximately a day) fluctuations in the environment. The central clock is localized in the suprachiasmatic nucleus (SCN) of the hypothalamus that is daily synchronized by the light stimulus through the retinohypothalamic tract (RTH). Moreover, non-photic stimulus such as nutritional inputs involving feeding time and diet composition can also entrain the clock [1,2,3•,4]. In addition to the SCN, peripheral clocks are localized in other brain areas, including nuclei within the hypothalamus and the hippocampus, as well as in peripheral tissues including the liver, kidney, heart, among others. These are synchronized in concert with the central clock, but are also influenced by additional stimuli, including food, hormones, among others. At the molecular level the clock consists of a network of interlocked transcriptional-translational feedback loops. The core molecular gears of this machinery are the transcription factors Circadian Locomotor Output Cycles Kaput (CLOCK) and Brain and Muscle ARNT-Like 1 (BMAL1), which heterodimerize and bind E-box promoter elements on the genome, activating a large number of clock-controlled genes (CCGs). Within the CCGs, the Period 1-3 (Per1, Per2 and Per3) and Cryptochrome 1-2 (Cry1 and Cry2) genes encode clock proteins that associate together and repress CLOCK–BMAL1, inhibiting their own expression by a negative autoregulatory feedback loop.
A number of CCGs encode for transcription factors, including D-box binding protein (DBP), thyrotroph embryonic factor (TEF), retinoic acid-related orphan receptor a (RORα) and reverse erythroblastosis virus α and β (REV-ERBα/β). DBP and TEF bind D-boxes, while RORα and REV-ERBα/β bind Reb-Erb/ROR promoter elements, thereby inducing additional circadian waves in expression of downstream genes. It is estimated that the circadian machinery controls the cyclic expression of about 10–20% of genes in any given cell [5], although recent findings indicate that many more genes may become circadian depending on nutritional and metabolic inputs [6••]. Importantly, epigenetic control appears to play a central role in the harmonic organization of circadian transcription [5,7,8,9••]. The circadian clock participates in homeostatic control by governing a number of physiological processes, metabolism, behavior, endocrine regulation, and cellular pathways. Hence disturbances in the circadian clock have been associated to various pathologies including, obesity, type-2 diabetes, feeding disorders, sleep disorders, Alzheimer's disease and psychiatric disorders.
The circadian clock in aging
It is common knowledge that elderly individuals have difficulties of sleeping at night, and they wake up at early morning [10]. Indeed, disruption of the circadian clock has been associated with several age-related pathologies including sleep disorders, diabetes, cancer, and memory reduction [10]. For example, daily fluctuations in the levels of hormones (including melatonin and cortisol), body temperature, and the sleep-wakefulness cycle are modified during aging. These changes lead to disrupted cycles, including reduction in the amplitude and phase shifts [11]. Moreover, in an intriguing experiment, cultured fibroblast expressing a luciferase reporter under the control of Bmal1 promoter was treated with serum from young and old individuals. This resulted in shortening of the circadian period and phase advance in the cells, as measured by luminescence, suggesting that a circulating factor present in the serum of older individuals is altering the cellular rhythms. Interestingly, the serum factor from old people is not melatonin non-cortisol, since there were no differences in the levels of these hormones from young and older blood donors [12].
The notion that the circadian clock might modulate the rate of aging is further supported by early studies made in rodents showing that the graft of fetal SCN to a model of aged hamsters (known as taus/+) restores behavioral rhythmicity and extends the life span [13]. In line with this observation, the transplantation of pineal gland, from young to old mice, prolongs life span [14]. In this context, the hormone melatonin produced in the pineal gland is a strong synchronizer. The synthesis and secretion of melatonin fluctuate with the day/night cycles, and are under the control of sympathetic innervation from the SCN [15–17]. Moreover, in addition to its chronobiological activities, melatonin also exhibits immunomodulatory, neuroendocrine and antioxidant activities; all of these contribute to its anti-aging properties [18]. Notably, the amplitude of plasmatic melatonin rhythms shows progressive reduction with aging. Additionally, melatonin operates as direct scavenger for free radicals and induces the expression and activity of antioxidant enzymes [19]. In this context, a functional relationship between anti-oxidative mechanisms and the circadian clock has been established. Several anti-oxidative enzyme activities and low molecular weight antioxidants follow circadian rhythms [19]. Moreover, experimental evidence shows that reactive-oxygen species (ROS) also follow circadian fluctuations [19]. Hence, it has been suggested that the circadian clock regulates mechanisms to protect the organism from oxidative stress [20]. For example, recent observations reveal that the dimer CLOCK:BMAL1 regulates the expression of the transcription factor NRF2 in the lung, which in turn, drives the circadian transcription of antioxidant genes. Some of these include the glutamate-cysteine ligase modifier subunit (Gclm) and the glutathione S-transferase A3 (Gsta3). Hence, the clock machinery modulates the circadian anti-oxidant response in lungs [21]. Importantly, results obtained from transgenic animals have confirmed the interplay of the circadian clock and aging and have also provided useful mechanistic leads for exploration. Specifically, ablation of the Bmal1 gene (define) by homologous recombination in the mouse results in accelerated aging and increased levels of ROS in several tissues [22]. Finally, additional evidence has revealed some of the mechanisms that are behind the age-related pathologies associated to circadian disruption. Specifically, as discussed below, cellular energy/nutrient sensing mechanisms seem to be critical in both circadian control and the aging process [23–25].
SIRT1: linking the clock with metabolism
The enzyme SIRT1, a NAD+ dependent deacetylase, targets a large variety of proteins, both histones and non-histones, including FOXO1, PGC1α, p53, E2F1, PPARγ, STAT3 and SCREBP-1c [19,20] (define). The histone deacetylase (HDAC) activity of SIRT1 oscillates in a circadian manner, rhythmically deacetylating histone H3 K9/K14 at promoters of CCGs, and the circadian core proteins BMAL1 and PER2 (define) [7,8]. Additionally, genetic ablation of Sirt1 or pharmacological inhibition of SIRT1 provokes disturbances in circadian cycles, both in cultured cells and in vivo [26,27]. It has been suggested that the activity of SIRT1 counterbalances the rhythmic histone acetyltransferase (HAT) function of CLOCK, although other HATs are likely to be implicated [28]. Importantly, it has been found that the activity of SIRT1 is modulated by in a circadian manner by its cofactor NAD+ [8]. NAD+ can be synthesized de novo from tryptophan or by the salvage pathway [29]. NAD+ can be used for energy transferring with the subsequent production of ATP in the mitochondria, or it is used in regulatory functions as a cofactor for NAD+-consuming enzymes. Remarkably, the circadian clock acts on the synthesis of NAD+, controlling the circadian expression of the nicotinamide phosphoribosyltransferase (Nampt) gene, a CCG that encodes the key rate-limiting enzyme in the salvage pathway [26,30]. Thus, the circadian feedback transcriptional loop is tightly linked to an enzymatic feedback loop (Figure 1). Furthermore, the NAD+/NADH ratio apparently changes with feeding in the hypothalamus and peripheral tissues such as the liver, possibly inducing cyclic activation of SIRT1 enzymatic activity [31,32]. Importantly, recent evidence has shown that the levels of NAD+ decrease during aging, hence impacting SIRT1 activity [33••].
Figure 1.

The NAD+ salvage pathway and its control by the circadian clock. The biosynthesis of NAD+ follows a circadian pattern, which is caused by the circadian expression of NAMPT, a rate-limiting enzyme in the NAD+ biosynthetic salvage pathway. The Nampt gene contains E-boxes in its promoter, leading to direct transcriptional control by the dimer CLOCK:BMAL1. The fluctuating levels of NAD+ modulate the activity of SIRT1 which in turn regulates the transcriptional activity of CLOCK:BMAL1 on their targets genes. During the aging the levels of NAD+ decreases and might alter the circadian rhythms of clock-controlled genes (CCGs)
SIRT1: linking the clock with aging
The hypothalamus controls a wide array of physiological functions that are modulated by aging. For example, the hypothalamus controls the whole organism energy balance, and malfunctioning of this control can trigger aging-associated metabolic disturbances such as obesity and type-2 diabetes. The hypothalamus also controls the sleep/wakefulness cycle that is altered in elderly individuals [10,34]. Importantly, the deciphering of the cellular mechanisms within these neuronal circuits has revealed the importance of nutrient-sensing pathways in the neuronal responses toward the body's energy balance and aging. Among these sensors, the role of SIRT1 has been analyzed in different hypothalamic nuclei. SIRT1 is necessary for the adaptation to fasting, and in conditions of caloric restriction, it triggers lipid mobilization from the adipose tissue, a switch from glucose to lipid oxidation in skeletal muscle and liver, and an increase in hepatic glucose production [34]. SIRT1 is necessary in neurons of the ventro-medial hypothalamus (VMH) and arcuate nucleus (ARC), to control glucose metabolism and lipid metabolism in peripheral tissues whose unbalance are hallmark of aging [34,35]. SIRT1 has been reported to protect against Alzheimer's disease, amyotrophic lateral sclerosis and axonal degeneration. Importantly, a recent study demonstrated that mice overexpressing SIRT1 in the brain, particularly in the hypothalamic nuclei dorsomedial hypothalamus (DMH) and lateral hypothalamus (LH) show increased lifespan, mitochondrial function in skeletal muscle and improved sleep quality [36•]. This ‘age-delayed’ phenotype is correlated with an increase of neuronal activity of the DMH and LH through an increment in the expression of the orexin type 2 receptor (Ox2r) [36•]. Importantly, in the SCN, SIRT1 modulates the central clock, in a process that appears to become less efficient in aged animals [37••]. This modulation comprises a direct deacetylation of BMAL1 by SIRT1 [32,38] through PGC-1α and NAMPT [37••] (define).
An additional aspect that links the circadian clock to the aging process is the control of energy metabolism in the mitochondria. Importantly, an unbiased analysis of the circadian acetylome has revealed that the large majority of clock-controlled acetylation relates to mitochondrial proteins [39•]. This his finding is in keeping with the involvement of SIRT3, a sirtuin that targets mitochondrial proteins, in clock control. SIRT3 appears to generate rhythms in the acetylation and activity of oxidative enzymes and respiration in the mitochondria of the liver [40••]. Some of these enzymes are discussed in McMurray in this issue. However, it is unclear whether this mechanism is influenced during aging. Nevertheless, SIRT3 regulates mitochondrial anti-oxidative stress in hematopoietic stem cells, being required to maintain mitochondrial homeostasis during oxidative stress or aging [41•]. Furthermore, it has been observed that in conditions of critical oxidative stress, cells respond by resetting the circadian clock, activating anti-oxidant pathways through BMAL1, heat shock factor 1 (HSF1) and casein kinase II (CK2) [42]. Interestingly, SIRT1 also participates in mitochondrial anti-aging function by modulating the nuclear-mitochondrial communication. It appears that the decrease in NAD+ levels during aging affects SIRT1 activity and thereby the control of nuclear-coded mitochondrial-genes [33••]. Hence, the decreased levels in NAD+ might impinge negatively on mitochondrial homeostasis by inhibiting the activity of sirtuins (Figure 2).
Figure 2.

Aging alters mitochondrial homeostasis. During aging the NAD+ synthesis decreases and consequently impairs the sirtuins activity, which have two consequences in the mitochondrial function: (a) The reduction of SIRT1 activity provokes the activation of HNF1α which in turn inhibits the transcription factor cMYC necessary to activate the transcription of Tfam which regulates the expression of mitochondrial genes. (b) Reduction of SIRT3 activity alters the function of mitochondrial proteins.
Nutrient sensing and feeding restriction: the clock-aging interaction
The concept that genetic components control aging derives from various observations, including that different species display different extents of longevity. Moreover, the manifest role played by environment on the aging process illustrates the presence of an epigenetic component [43]. In this context, caloric restriction (CR) is a powerful environmental intervention that delays the effects of aging in experimental models such as yeast, worms, flies and mice [23,44]. Importantly, CR experimental protocols are generally accompanied by temporal feeding. Indeed, the nutrient/metabolic sensor system that acts on the generation of beneficial cellular adaptations during CR and consequently delaying aging, also modulates the central clock [37••, 45•] (Table 1). These notions support the concept that the circadian clock is positioned downstream of metabolic sensors, acting as an effector and thus controlling cellular and physiological homeostasis. For example, mice fed with high fat diet (HFD) only during the mice's activity period show less body weight and better glucose metabolism accompanied by more robust circadian rhythms, despite their consumption of the same quantity of calories than the ad libitum fed mice [3•]. Thus, the synchronization of the circadian clock by the environment appears critical for the correct control of cellular and body homeostasis. In this context, recent experimental data in humans have shown the association between short sleep duration and the development of metabolic unbalances [46–48].
Table 1. Nutrient sensors linking the clock system to the aging process.
| Protein | Circadian function | Aging phenotype | Reference |
|---|---|---|---|
| SIRT1 | Regulates the circadian clock by BMAL1 and PER2 deacetylation. Activates BMAL1 and CLOCK in the SCN in young mice | Modulates mitochondrial function through NAD+ levels in young mice | [7,8,33••,37••] |
| SIRT3 | Modulates the circadian activity of the mitochondria by rhythms in the acetylation and activation of oxidative enzymes | In stems cells reverts the effect of aging-oxidative stress in mitochondria activating the anti-oxidative defense system | [40••,41•] |
| mTOR | Modulates rhythmically the translational control in circadian genes through 4E-BP1 | Its inhibition extends lifespan in mice | [24,45•,49] |
| AMPK | Phosphorylates and destabilizes CRY1 altering the circadian rhythms in mice | Activated under low ATP levels, inhibits mTOR, and its pharmacological activation extend lifespan in mice | [52,55] |
mTOR: another defense against aging
The mammalian target of rapamycin, mTOR, has been involved in the control of aging and caloric restriction effects. mTOR is a kinase that integrates nutrients, stress, growth and energy status inputs with cellular metabolic resources to balance catabolic and anabolic pathways. mTOR is found in two multi-protein complexes named mTORC1 and mTORC2 [49]. Caloric restriction reduces mTORC1 activity, and pharmacological or genetic disruption of mTORC1 is sufficient to extend lifespan in invertebrates and mice under normal caloric conditions [44]. Interestingly, a role for mTOR in the circadian clock has been shown. In the SCN, mTOR shows circadian activity, and photic signals activate mTOR signaling, which in turn promotes the translation of Vip (vasoactive intestinal particle) mNA by repressing 4E-BP1 [45•]. Finally, the clock seems to modulate negatively the mTOR pathway by a mechanism that involves BMAL1 [25]. These data stress the intrinsic cooperation between the photic and non-photic stimuli (i.e. feeding restriction) to modulate the circadian clock through nutrient sensor pathways such as SIRT1, AMPK, and mTOR, which in turn could be modulated by the circadian clock, generating a loop that could be both beneficial or prejudicial for healthy aging, depending on environmental factors (Table 1 and Figure 3).
Figure 3.
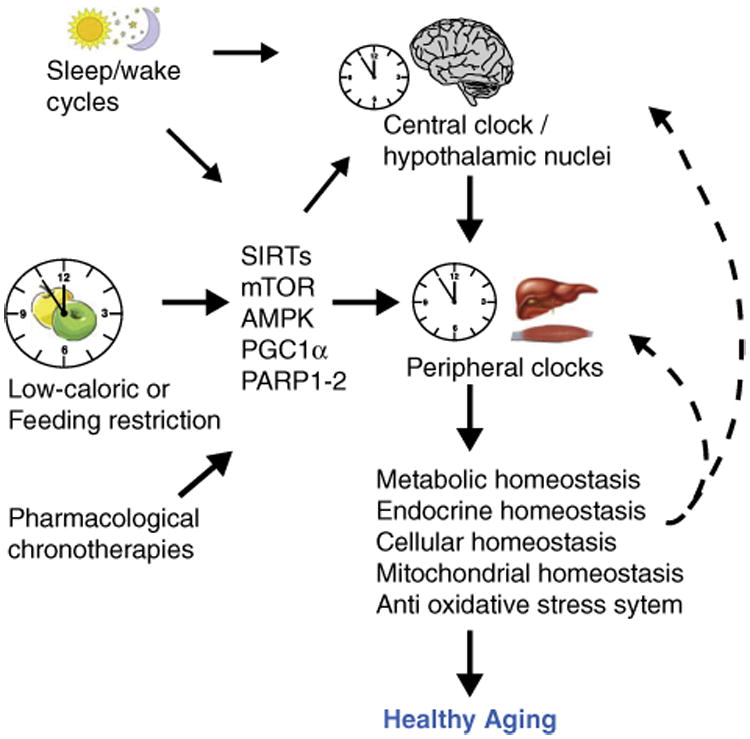
Nutrient sensors ‘sense’ the environmental conditions that modulate the circadian clock and the aging process. Healthy environment, such as enough sleeping time and low caloric diet/scheduled feeding, modulates nutrient sensors localized in the brain and peripheral tissues. These in turn synchronize the circadian clocks. As a consequence, the activation of anti-aging mechanisms improves the homeostasis at different levels promoting healthy aging. The different physiological conditions might decelerate or accelerate aging.
Concluding remarks
Despite the advances in medicine during the past century, a significant increment in the morbidity of noninfectious diseases such as cancer, type-2 diabetes, obesity and mental illness is evident. This trend has been associated to a diminution in the quality of life. Studies during the past years have permitted to identify key molecular mechanisms behind the aging process. In this respect, the remarkable role of cellular nutrient sensing pathways has allowed the development of strategies including genetic, diet protocols (such as the reduction in calories and/or the use of most appropriate meal schedules), and pharmacological approaches.
Furthermore, recent evidence shows a clear link between the circadian clock and nutrient sensors that ultimately may modulate anti-aging and homeostatic mechanisms. Hence, therapeutic strategies that would include circadian regulation of metabolism and physiology are to be taken seriously into consideration [50]. Attempts to modulate these pathways through chronotherapy are likely to enhance their anti-aging properties and dampen possible side effects [23,51,52]. Finally, taking into consideration the notion of integrons in aging [53] and the observation that key tissues are able to influence the whole body homeostasis [36•, 41•, 54] (Figure 3), it is conceivable that monitoring the aging-rate in these key tissues would substantially improve the healthy aging in the whole organism.
Acknowledgments
We thank all the members of the Sassone-Corsi and Borrelli laboratories for the discussions and insights. Work in the Center for Epigenetics and Metabolism is supported by the National Institute of Health, the Merieux Fondation and INSERM (Institut National de la Sante et Recherche Medicale, France). R.O.-S. is supported by a fellowship from the Government of Mexico and by the Della Martin Foundation.
References and recommended reading
Papers of particular interest, published within the period of review, have been highlighted as:
• of special interest
•• of outstanding interest
- 1.Damiola LMNF, Preitner N, Kornmann B, Fleury-Olela F, Schibler U. Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus. Genes Dev. 2000;14:2950–2961. doi: 10.1101/gad.183500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Saper CB, Scammell TE, Lu J. Hypothalamic regulation of sleep and circadian rhythms. Nature. 2005;437:1257–1263. doi: 10.1038/nature04284. [DOI] [PubMed] [Google Scholar]
- 3•.Hatori M, Vollmers C, Zarrinpar A, DiTacchio L, Bushong EA, Gill S, Leblanc M, Chaix A, Joens M, Fitzpatrick JA, et al. Time-restricted feeding without reducing caloric intake preventsmetabolic diseases in mice fed a high-fat diet. Cell Metab. 2012;15:848–860. doi: 10.1016/j.cmet.2012.04.019. Time-restricted feeding in mice protects against metabolic disturbances under high-fat feeding and promotes robustness in the circadian and metabolic rhythms. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Stephan FK. The other circadian system: food as a zeitgeber. J Biol Rhythm. 2002;17:284–292. doi: 10.1177/074873040201700402. [DOI] [PubMed] [Google Scholar]
- 5.Aguilar-Arnal L, Sassone-Corsi P. The circadian epigenome:how metabolism talks to chromatin remodeling. Curr Opin Cell Biol. 2013;25:170–176. doi: 10.1016/j.ceb.2013.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6••.Eckel-Mahan KL, Patel VR, de Mateo S, Orozco-Solis R, Ceglia NJ, Sahar S, Dilag-Penilla SA, Dyar KA, Baldi P, Sassone-Corsi P. Reprogramming of the circadian clock by nutritional challenge. Cell. 2013;155:1464–1478. doi: 10.1016/j.cell.2013.11.034. Nutritional challenge induces a reorganization in metabolic pathways and circadian transcription by molecular mechanisms involving epigenetic control. These metabolic and transcriptional changes are reversible depending on the caloric content of food. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Asher G, Gatfield D, Stratmann M, Reinke H, Dibner C, Kreppel F, Mostoslavsky R, Alt FW, Schibler U. SIRT1 regulates circadianclock gene expression through PER2 deacetylation. Cell. 2008;134:317–328. doi: 10.1016/j.cell.2008.06.050. [DOI] [PubMed] [Google Scholar]
- 8.Nakahata Y, Kaluzova M, Grimaldi B, Sahar S, Hirayama J, Chen D, Guarente LP, Sassone-Corsi P. The NAD+-dependentdeacetylase SIRT1 modulates CLOCK-mediatedchromatin remodeling and circadian control. Cell. 2008;134:329–340. doi: 10.1016/j.cell.2008.07.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9••.Aguilar-Arnal L, Hakim O, Patel VR, Baldi P, Hager GL, Sassone-Corsi P. Cycles in spatial and temporal chromosomal organization driven by the circadian clock. Nat Struct Mol Biol. 2013;20:1206–1213. doi: 10.1038/nsmb.2667. First demonstration by chromosome-capture technology that control of circadian gene expression is obtained through ‘circadian interactomes’, structural hubs of nuclear organization in which several co-regulated genes are physically associated and regulated by the circadian clock. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Kondratova AA, Kondratov RV. The circadian clock and pathology of the ageing brain. Nat Rev Neurosci. 2012;13:325–335. doi: 10.1038/nrn3208. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Hofman MA, Swaab DF. Living by the clock: the circadian pacemaker in older people. Ageing Res Rev. 2006;5:33–51. doi: 10.1016/j.arr.2005.07.001. [DOI] [PubMed] [Google Scholar]
- 12.Pagani L, Schmitt K, Meier F, Izakovic J, Roemer K, Viola A, Cajochen C, Wirz-Justice A, Brown SA, Eckert A. Serum factors in older individuals change cellular clock properties. Proc Natl Acad Sci U S A. 2011;108:7218–7223. doi: 10.1073/pnas.1008882108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hurd MW, Ralph MR. The significance of circadian organization for longevity in the golden hamster. J Biol Rhythm. 1998;13:430–436. doi: 10.1177/074873098129000255. [DOI] [PubMed] [Google Scholar]
- 14.Lesnikov VA, Pierpaoli W. Pineal cross-transplantation (old-to-young and vice versa) as evidence for an endogenous aging clock. Ann N YAcad Sci. 1994;719:456–460. doi: 10.1111/j.1749-6632.1994.tb56850.x. [DOI] [PubMed] [Google Scholar]
- 15.Foulkes NS, Borjigin J, Snyder SH, Sassone-Corsi P. Rhythmic transcription: the molecular basis of circadian melatonin synthesis. Trends Neurosci. 1997;20:487–492. doi: 10.1016/s0166-2236(97)01109-0. [DOI] [PubMed] [Google Scholar]
- 16.Haus E. Chronobiology in the endocrine system. Adv Drug Deliv Rev. 2007;59:985–1014. doi: 10.1016/j.addr.2007.01.001. [DOI] [PubMed] [Google Scholar]
- 17.Perreau-Lenz S, Kalsbeek A, Van Der Vliet J, Pévet P, Buijs RM. In vivo evidence for a controlled offset of melatonin synthesis at dawn by the suprachiasmatic nucleus in the rat. Neuroscience. 2005;130:797–803. doi: 10.1016/j.neuroscience.2004.10.014. [DOI] [PubMed] [Google Scholar]
- 18.Poeggeler B. Melatonin, aging, and age-related diseases: perspectives for prevention, intervention, and therapy. Endocrine. 2005;27:201–212. doi: 10.1385/ENDO:27:2:201. [DOI] [PubMed] [Google Scholar]
- 19.Hardeland R, Coto-Montes A, Poeggeler B. Circadian rhythms, oxidative stress, and antioxidative defense mechanisms. Chronobiol Int. 2003;20:921–962. doi: 10.1081/cbi-120025245. [DOI] [PubMed] [Google Scholar]
- 20.Stangherlin A, Reddy AB. Regulation of circadian clocks by redox homeostasis. J Biol Chem. 2013;288:26505–26511. doi: 10.1074/jbc.R113.457564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Pekovic-Vaughan V, Gibbs J, Yoshitane H, Yang N, Pathiranage D, Guo B, Sagami A, Taguchi K, Bechtold D, Loudon A, et al. The circadian clock regulates rhythmic activation of the NRF2/glutathione-mediated antioxidant defense pathway to modulate pulmonary fibrosis. Genes Dev. 2014;28:548–560. doi: 10.1101/gad.237081.113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Kondratov RV, Kondratova AA, Gorbacheva VY, Vykhovanets OV, Antoch MP. Early aging and age-related pathologies in mice deficient in BMAL1, the core component of the circadian clock. Genes Dev. 2006;20:1868–1873. doi: 10.1101/gad.1432206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Fontana L, Partridge L, Longo VD. Extending healthy life span — from yeast to humans. Science. 2010;328:321–326. doi: 10.1126/science.1172539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Bellet MM, Orozco-Solis R, Sahar S, Eckel-Mahan K, Sassone-Corsi P. The time of metabolism: NAD+, SIRT1, and the circadian clock. Cold Spring Harb Symp Quant Biol. 2011;76:31–38. doi: 10.1101/sqb.2011.76.010520. [DOI] [PubMed] [Google Scholar]
- 25.Khapre RV, Kondratova AA, Patel S, Dubrovsky Y, Wrobel M, Antoch MP, Kondratov RV. BMAL1-dependent regulation of the mTOR signaling pathway delays aging. Aging (Albany, NY) 2014;6:48–57. doi: 10.18632/aging.100633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Nakahata Y, Sahar S, Astarita G, Kaluzova M, Sassone-Corsi P. Circadian control of the NAD+ salvage pathway by CLOCK-SIRT1. Science. 2009;324:654–657. doi: 10.1126/science.1170803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Bellet MM, Nakahata Y, Boudjelal M, Watts E, Mossakowska DE, Edwards KA, Cervantes M, Astarita G, Loh C, Ellis JL, et al. Pharmacological modulation of circadian rhythms by synthetic activators of the deacetylase SIRT1. Proc Natl Acad Sci USA. 2013;110:3333–3338. doi: 10.1073/pnas.1214266110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Masri S, Sassone-Corsi P. Plasticity and specificity of the circadian epigenome. Nat Neurosci. 2010;13:1324–1329. doi: 10.1038/nn.2668. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Revollo JR, Grimm AA, Imai S. The NAD biosynthesis pathway mediated by nicotinamide phosphoribosyltransferase regulates Sir2 activity in mammalian cells. J Biol Chem. 2004;279:50754–50763. doi: 10.1074/jbc.M408388200. [DOI] [PubMed] [Google Scholar]
- 30.Ramsey KM, Yoshino J, Brace CS, Abrassart D, Kobayashi Y, Marcheva B, Hong HK, Chong JL, Buhr ED, Lee C, et al. Circadian clock feedback cycle through NAMPT-mediated NAD+ biosynthesis. Science. 2009;324:651–654. doi: 10.1126/science.1171641. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Dibner C, Schibler U, Albrecht U. The mammalian circadian timing system: organization and coordination of central and peripheral clocks. Ann Rev Physiol. 2010;72:517–549. doi: 10.1146/annurev-physiol-021909-135821. [DOI] [PubMed] [Google Scholar]
- 32.Cakir I, Perello M, Lansari O, Messier NJ, Vaslet CA, Nillni EA. Hypothalamic Sirt1 regulates food intake in a rodent modelsystem. PLoS ONE. 2009;4:e8322. doi: 10.1371/journal.pone.0008322. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33••.Gomes Ana P, Price Nathan L, Ling Alvin JY, Moslehi Javid J, Montgomery MK, Rajman L, White James P, Teodoro João S, Wrann Christiane D, Hubbard Basil P, et al. Declining NAD+ induces a pseudohypoxic state disrupting nuclear-mitochondrial communication during aging. Cell. 2013;155:1624–1638. doi: 10.1016/j.cell.2013.11.037. Decline in SIRT1 activity by the reduction in NAD+ levels during aging provokes a reduction in the expression of mitochondrial-encoded genes leading to mitochondrial malfunction. Increasing the NAD+ levels restores the nucleus–mitochondria communication reversing the malfunction in aged mice. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Ramadori G, Fujikawa T, Anderson J, Berglund Eric D, Frazao R, Michan S, Vianna Claudia R, Sinclair David A, Elias Carol F, Coppari R. SIRT1 deacetylase in SF1 neurons protects against metabolic imbalance. Cell Metabol. 2011;14:301–312. doi: 10.1016/j.cmet.2011.06.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Ramadori G, Fujikawa T, Fukuda M, Anderson J, Morgan DA, Mostoslavsky R, Stuart RC, Perello M, Vianna CR, Nillni EA, et al. SIRT1 deacetylase in POMC neurons is required for homeostatic defenses against diet-induced obesity. Cell Metab. 2010;12:78–87. doi: 10.1016/j.cmet.2010.05.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36•.Satoh A, Brace Cynthia S, Rensing N, Cliften P, Wozniak David F, Herzog Erik D, Yamada Kelvin A, Imai Si. Sirt1 extends life span and delays aging in mice through the regulation of Nk2 homeobox 1 in the DMH and LH. Cell Metabol. 2013;18:416–430. doi: 10.1016/j.cmet.2013.07.013. This work demonstrates the participation of the hypothalamic SIRT1 in aging and longevity, through increasing Ox2r within hypothalamic nuclei DMH and LH. An axe involving sleeping time and muscular physiology is proposed to explain the reduced aging phenotype in these mice. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37••.Chang HC, Guarente L. SIRT1 mediates central circadian control in the SCN by a mechanism that decays with aging. Cell. 2013;153:1448–1460. doi: 10.1016/j.cell.2013.05.027. This study indicates that SIRT1 controls the central clock by activating BMAL1 and CLOCK via PGC1α and NAMPT. Importantly, this mechanism decays during aging, and the overexpression of SIRT1 in the brain protects against the loss of circadian function. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Hirayama J, Sahar S, Grimaldi B, Tamaru T, Takamatsu K, Nakahata Y, Sassone-Corsi P. CLOCK-mediated acetylation of BMAL1 controls circadian function. Nature. 2007;450:1086–1090. doi: 10.1038/nature06394. [DOI] [PubMed] [Google Scholar]
- 39•.Masri S, Patel VR, Eckel-Mahan KL, Peleg S, Forne I, Ladurner AG, Baldi P, Imhof A, Sassone-Corsi P. Circadian acetylome reveals regulation of mitochondrial metabolic pathways. Proc Natl Acad Sci USA. 2013;110:3339–3344. doi: 10.1073/pnas.1217632110. The analysis of the liver circadian acetylome by mass spectrometry revealed that the clock drives the function of a number of mitochondrial metabolic pathways through the rhythmic acetylation of mitochondrial proteins. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40••.Peek CB, Affinati AH, Ramsey KM, Kuo HY, Yu W, Sena LA, Ilkayeva O, Marcheva B, Kobayashi Y, Omura C, et al. Circadian clock NAD+ cycle drives mitochondrial oxidative metabolism in mice. Science. 2013:342. doi: 10.1126/science.1243417. Circadian oscillation of NAD+ controls the activity of SIRT3, which in turn controls the acetylation and activity of mitochondrial enzymes, leading to regulation of oxidative metabolism. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41•.Brown K, Xie S, Qiu X, Mohrin M, Shin J, Liu Y, Zhang D, Scadden David T, Chen D. SIRT3 reverses aging-associated degeneration. Cell Rep. 2013;3:319–327. doi: 10.1016/j.celrep.2013.01.005. SIRT3 regulates the stress response in stem cells, a critical process under stress or aging. Overactivation of SIRT3 enhances the regenerative capacity in old stem cells. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Tamaru T, Hattori M, Ninomiya Y, Kawamura G, Vares G, Honda K, Mishra DP, Wang B, Benjamin I, Sassone-Corsi P, et al. ROS stress resets circadian clocks to coordinate pro-survival signals. PLoS ONE. 2013;8:e82006. doi: 10.1371/journal.pone.0082006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Jirtle RL, Skinner MK. Environmental epigenomics and disease susceptibility. Nat Rev Genet. 2007;8:253–262. doi: 10.1038/nrg2045. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Johnson SC, Rabinovitch PS, Kaeberlein M. mTOR is a key modulator of ageing and age-related disease. Nature. 2013;493:338–345. doi: 10.1038/nature11861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45•.Cao R, Robinson B, Xu H, Gkogkas C, Khoutorsky A, Alain T, Yanagiya A, Nevarko T, Liu Andrew C, Amir S, et al. Translational control of entrainment and synchrony of the suprachiasmatic circadian clock by mTOR/4E-BP1. Signal Neuron. 2013;79:712–724. doi: 10.1016/j.neuron.2013.06.026. The authors show that mTOR controls the circadian activity of 4E-BP1, which in turn represses the mRNA translation of Vip. They also show that in Mtor+/− mice VIP expression is decreased and accompanied by defects in the circadian entrainment of the SCN. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Möller-Levet CS, Archer SN, Bucca G, Laing EE, Slak A, Kabiljo R, Lo JCY, Santhi N, von Schantz M, Smith CP, et al. Effects of insufficient sleep on circadian rhythmicity and expression amplitude of the human blood transcriptome. Proc Natl Acad Sci USA. 2013;110:E1132–E1141. doi: 10.1073/pnas.1217154110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Markwald RR, Melanson EL, Smith MR, Higgins J, Perreault L, Eckel RH, Wright KP. Impact of insufficient sleep on total daily energy expenditure, food intake, and weight gain. Proc Natl Acad Sci USA. 2013;110:5695–5700. doi: 10.1073/pnas.1216951110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Buxton OM, Cain SW, O'Connor SP, Porter JH, Duffy JF, Wang W, Czeisler CA, Shea SA. Adverse metabolic consequences in humans of prolonged sleep restriction combined with circadian disruption. Sci Transl Med. 2012;4:129ra143. doi: 10.1126/scitranslmed.3003200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Lamming DW, Ye L, Katajisto P, Goncalves MD, Saitoh M, Stevens DM, Davis JG, Salmon AB, Richardson A, Ahima RS, et al. Rapamycin-induced insulin resistance is mediated by mTORC2 loss and uncoupled from longevity. Science. 2012;335:1638–1643. doi: 10.1126/science.1215135. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sahar S, Sassone-Corsi P. Metabolism and cancer: the circadian clock connection. Nat Rev Cancer. 2009;9:886–896. doi: 10.1038/nrc2747. [DOI] [PubMed] [Google Scholar]
- 51.Johnson SC, Yanos ME, Kayser EB, Quintana A, Sangesland M, Castanza A, Uhde L, Hui J, Wall VZ, Gagnidze A, et al. mTOR inhibition alleviates mitochondrial disease in a mouse model of Leigh syndrome. Science. 2013;342:1524–1528. doi: 10.1126/science.1244360. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Anisimov VN. Metformin for aging and cancer prevention. Aging (Albany, NY) 2010;2:760–774. doi: 10.18632/aging.100230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Dillin A, Gottschling DE, Nyström T. The good and the bad of being connected: the integrons of aging. Curr Opin Cell Biol. 2014;26:107–112. doi: 10.1016/j.ceb.2013.12.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Zhang G, Li J, Purkayastha S, Tang Y, Zhang H, Yin Y, Li B, Liu G, Cai D. Hypothalamic programming of systemic ageing involving IKK-beta, NF-kappaB and GnRH. Nature. 2013;497:211–216. doi: 10.1038/nature12143. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Lamia KA, Sachdeva UM, DiTacchio L, Williams EC, Alvarez JG, Egan DF, Vasquez DS, Juguilon H, Panda S, Shaw RJ, et al. AMPK regulates the circadian clock by cryptochrome phosphorylation and degradation. Science. 2009;326:437–440. doi: 10.1126/science.1172156. [DOI] [PMC free article] [PubMed] [Google Scholar]