

Abstract
In East Asian traditional medicine, Pulsatillae Radix (PR) is widely used to treat amoebic dysentery and renowned for its anti-inflammatory effects. This study aimed to confirm evidence regarding the potential therapeutic effect of PR on Crohn's disease using a system network level based in silico approach. Study results showed that the compounds in PR are highly connected to Crohn's disease related pathways, biological processes, and organs, and these findings were confirmed by compound-target network, target-pathway network, and gene ontology analysis. Most compounds in PR have been reported to possess anti-inflammatory, anticancer, and antioxidant effects, and we found that these compounds interact with multiple targets in a synergetic way. Furthermore, the mRNA expressions of genes targeted by PR are elevated significantly in immunity-related organ tissues, small intestine, and colon. Our results suggest that the anti-inflammatory and repair and immune system enhancing effects of PR might have therapeutic impact on Crohn's disease.
1. Introduction
Inflammatory bowel disease (IBD) may be categorized clinically as Crohn's disease or ulcerative colitis [1]. Crohn's disease usually causes a variety of systemic symptoms, which include chronic inflammation of the bowel [2]. Although any part of the digestive tract from mouth to anus may be affected, usually the small intestine (ileum) and the large intestine (colon) are involved; ileocolic Crohn's accounts for 50% of cases, ileal Crohn's for 30%, and colic Crohn's for 20% of cases [3]. Symptoms vary though its common manifestations include persistent diarrhea, rectal bleeding, abdominal cramps, and pain, though fever, extreme fatigue, and weight loss are also common [1, 2, 4]. Constipation is also a frequent symptom and can lead to bowel obstruction and, thus, increase the risk of bowel cancer. Complications involving other than the gastrointestinal (GI) tract include anemia, arthritis, liver disease, eye inflammation, and skin rashes [2, 4].
According to a statistical report issued by the Health Insurance Review and Assessment Service in Korea in 2016, the number of Crohn's disease patients increased from 13,920 in 2011 to 18,332 in 2015, an average annual increase of 7.1%. Furthermore, over the same period, total medical cost has increased by 19.4% annually, and more than half of patients are in 20s or 30s. A systematic review about the economic and quality-of-life burden of Crohn's disease reported that, in the USA, Germany, France, UK, Italy, and Spain, in all countries combined, Crohn's medical costs totaled €30 billion annually and that patient quality of life was substantially diminished by the physical, emotional, and social effects of the disease [5].
Although Crohn's disease is a global health problem [5], its pathology remains poorly understood [1, 6]. Nevertheless, it has been established that its etiology is associated with complex interactions between environmental, immune, microbial, and genetic factors [4, 7], though a number of authors have suggested that the primary defect in Crohn's disease is one of relative immunodeficiency [6, 8, 9].
A systematic review of publications from 1947 to 2013 involving controlled studies of herbal therapies in IBD indicated that at least 50 percent of IBD patients used some form of complementary and alternative medicine (CAM), since most herbal therapies had been reported to have anti-inflammatory effects and plausible action mechanisms in IBD with minimal adverse effects [10]. Moreover, herbal medicines are being increasingly used and requested by IBD patients not only in Asia, but also in Western countries [11].
Of the anti-inflammatory herbal medicines, Pulsatillae Radix (Baekduong, PR) is worth considering as a potential treatment for Crohn's disease as it was widely used in traditional medicine to treat amoebic dysentery and has also been demonstrated scientifically to have anti-inflammatory effects experimentally [12, 13]. PR is traditional Korean herbal medicine prepared from the roots of Pulsatilla koreana Nakai and contains several phytochemicals, including anemonin, hederagenin, oleanolic acid, and deoxypodophyllotoxin [14, 15]. Some experimental study results have shown that PR has various biological activities. For instance, aqueous and ethanol extracts of PR have been reported to demonstrate anticancer effects in anaplastic thyroid cancer [16], methanol extract of PR was found to have anti-inflammatory effects in lipopolysaccharide (LPS) exposed rats [17], and PR has also been reported to inhibit adipocyte differentiation and to suppress adipogenesis [13].
According to the meridian tropism theory of traditional medicine, the effects of PR reach to the stomach meridian and the large intestine meridian, which is in accord with the organs commonly affected by Crohn's disease. In terms of the selection of traditional medication, meridian tropism theory plays an important role, as it is one of the core principles of traditional medicine [18]. Based on meridian tropism theory, each herbal medicine possesses different affinities for certain organs and meridians of the body [19].
In the present study, we sought to confirm the therapeutic effects of PR in Crohn's disease using system level analysis and a network based in silico approach. A schematic of the network pharmacological study is summarized in Figure 1.
Figure 1.

Workflow: network analysis, relevant organ location network analysis, gene ontology, and pathway analysis were performed on active compounds identified in Pulsatillae Radix (PR) by screening and target fishing.
2. Material and Methods
2.1. Identification of Active Compounds
According to the Traditional Chinese Medicine Systems Pharmacology (TCMSP, http://ibts.hkbu.edu.hk/LSP/tcmsp.php) database (a free phytochemical database of herbal medicine), PR contains 57 identified compounds. Parameters related to absorption, distribution, metabolism, and excretion (ADME), namely, human drug-likeness (DL) [20], oral bioavailability (OB) [21], and Caco-2 permeability (Caco-2) [22], were employed to filter out potential active compounds.
2.1.1. Drug-Likeness Evaluation
DL helps filter out “drug-like” compounds in oriental herbs, as DL represents a qualitative concept for valuations based on how “drug-like” prospective compound is [23]. Accordingly, a high DL may lead to a greater possibility of therapeutic success, and compounds with a higher DL value are more likely to possess certain biological properties [24]. Calculations of DL in the TCMSP database are based on the Tanimoto coefficient formula [25] as follows:
| (1) |
where A represents the molecular parameters of herbal compounds and B is the average molecular parameter of all compounds in the DrugBank database (http://www.drugbank.ca/) [26]. In the present study, we excluded compounds with a DL value of < 0.18.
2.1.2. Oral Bioavailability (OB) Prediction
Oral bioavailability (OB) is defined as the absorption ratio of an active compound into the systemic circulation, which represents convergence of the ADME process [21]. OB values are dependent on drug dissolution in the gastrointestinal (GI) tract, intestinal and hepatic first-pass metabolisms, and intestinal membrane permeation, and, thus, OB is considered a major pharmacokinetic parameter for drug evaluations [24]. In this study, the OB threshold was set as ≥ 15%.
2.1.3. Caco-2 Permeability Screening
Caco-2 permeability is used to predict the absorption of an orally administered drug [22]. Surface absorptivity of the small intestine is maximized by villi and microvilli; for this reason, orally administered drugs are mostly absorbed in the small intestine [27]. Moreover, the movement of orally administered drugs across the intestinal epithelial barrier determines the rate and extent of human absorption and ultimately affects drug bioavailability [28]. In the present study, compounds with OB, DL, and Caco-2 values of >15%, >0.18, and >−0.4, respectively, were regarded as active and subjected to analysis. In addition, we included some compounds with lower ADME profile than above thresholds, for the reason that those were reported to possess anti-inflammatory, antioxidant, anticancer, and antibacterial effects. This study was about only one single herb, and for this reason we did not use a high threshold of ADME profile to filter potential active compounds. Instead, we lowered the standard of OB in order to analyze the most potential targets of PR.
2.2. Target Fishing
Molecular targets of filtered potential active compounds were found in the TCMSP [29], Similarity Ensemble Approach (SEA, http://sea.bkslab.org), and the Binding Database (http://www.bindingdb.org). In addition, filtered compound-target interaction profile mapping was performed using the UniProt database (http://www.uniprot.org/) [30].
2.3. Gene Ontology (GO) Analysis
Biological process (BP) of gene ontology (GO) analysis was employed to determine the biological properties of target genes [31]. GO annotation provides statistical analyses on gene function information. In this research, GO BP terms with P values < 0.01 were employed and the data was collected using the DAVID 6.8 Gene Functional Classification Tool (http://david.abcc.ncifcrf.gov/).
2.4. Network Construction and Analysis
In order to understand the multiscale interactions between the active compounds of PR and targets, two types of networks were built: (1) a compound-target network (C-T network), in which nodes represented either compounds or target proteins, and edges indicated compound-target connections; and (2) a target-pathway network (T-P network), which was used to extract pathways from the KEGG database (http://www.genome.jp/kegg/) and to filter out terms highly associated with Crohn's disease and P values of < 0.01. Related targets were mapped onto relevant pathways to produce the T-P network. Both networks were generated in Cytoscape 3.4.0, an open-source biological network visualization and data integration software package [32].
2.5. Target Organ Location Network
Tissue-specific patterns of mRNA expression can indicate important associations with particular biological events or gene functions [33]. Thus, to explore the beneficial effects of PR on Crohn's disease, it was important to determine the tissue mRNA expression profiles of target proteins at the organ level [34]. The target organ location network was used with the Dataset: GeneAtlas U133A, gcrma (http://biogps.org). The BioGPS database provides expression data acquired by direct measurements of gene expressions by microarrays analysis [35]. First, the mRNA expression patterns of each target gene in 84 parts of organ tissues were obtained. Second, average values were calculated for each gene. Third, above average mRNA expressions in relevant organ tissues were extracted and arranged by frequency. Finally, a target organ location network was constructed using Cytoscape 3.4.0 and organ-specific, Crohn's disease related, gene overexpression data.
2.6. GEO2R Analysis
Using Gene Expression Omnibus (GEO, https://www.ncbi.nlm.nih.gov/geo), we compared mRNA expression pattern of normal groups and Crohn's disease groups. GEOquery and limma R packages of GEO2R tool were employed to identify highly expressed genes.
2.7. Network Pathway
In order to elucidate the action mechanisms of PR in Crohn's disease, filtered target proteins were input into the pathway map of inflammatory bowel disease acquired from the Kyoto Encyclopedia of Genes and Genomes (KEGG, http://www.kegg.jp/) database.
3. Results
3.1. Identification of Active Compounds
Of the 57 compounds (as shown in Supplementary Table S1 in Supplementary Material available online at https://doi.org/10.1155/2017/4198035) in PR acquired from the TCMSP, excluding compounds with no target information, 19 compounds with a known target met the criteria OB ≥ 15%, Caco-2 ≥ −0.4, and DL ≥ 0.18. Additionally 13 compounds reported to have anti-inflammatory, antioxidant, anticancer, and antibacterial effects were added, and finally 32 compounds were analyzed (as shown in Table 1).
Table 1.
32 potential active compounds of Pulsatillae Radix.
| ID | Active compounds | Structure | OB (%) | Caco-2 | DL |
|---|---|---|---|---|---|
| C1 | 3-Methylcoumarin |
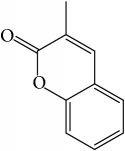
|
19.66 | 1.27 | 0.05 |
|
| |||||
| C2 | 5,6,7-Trimethoxycoumarin |
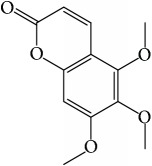
|
32.54 | 0.94 | 0.12 |
|
| |||||
| C3 | AIDS045703 |
|
21.37 | 0.55 | 0.87 |
|
| |||||
| C4 | Androstane-3,11,17-triol |
|
13.19 | −0.04 | 0.38 |
|
| |||||
| C5 | Anemosapogenin |
|
17.87 | 0.07 | 0.77 |
|
| |||||
| C6 | Anemoside A3 |
|
15.46 | −1.6 | 0.15 |
|
| |||||
| C7 | Aureusidin |
|
53.42 | 0.07 | 0.24 |
|
| |||||
| C8 | Beta-sitosterol |
|
36.91 | 1.32 | 0.75 |
|
| |||||
| C9 | Betulonic acid |
|
16.83 | 0.65 | 0.78 |
|
| |||||
| C10 | Campesterol |
|
5.57 | 1.6 | 0.72 |
|
| |||||
| C11 | Cauloside A |
|
6.84 | −0.82 | 0.4 |
|
| |||||
| C12 | Cernuoside |
|
2.69 | −1.51 | −2.18 |
|
| |||||
| C13 | Dauricine (8CI) |
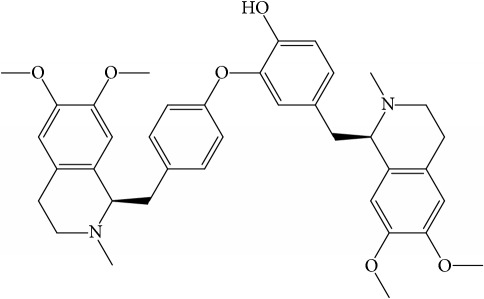
|
23.65 | 0.9 | 0.37 |
|
| |||||
| C14 | Ergosterol |
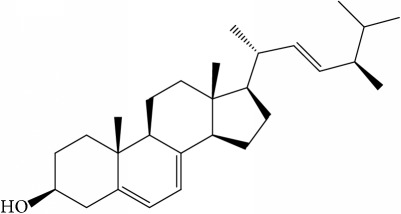
|
14.29 | 1.47 | 0.72 |
|
| |||||
| C15 | Fraxinol |
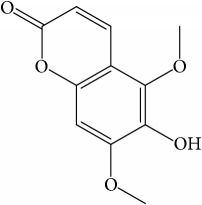
|
24.19 | 0.7 | 0.1 |
|
| |||||
| C16 | Hederagenol |
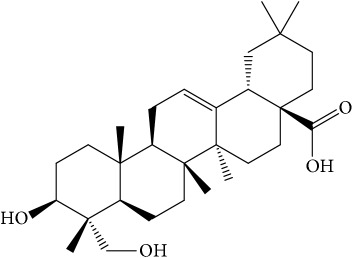
|
22.42 | 0.1 | 0.74 |
|
| |||||
| C17 | Isorhamnetin |
|
49.6 | 0.31 | 0.31 |
|
| |||||
| C18 | LAN |
|
42.12 | 1.52 | 0.75 |
|
| |||||
| C19 | Lignoceric acid |
|
14.9 | 1.24 | 0.33 |
|
| |||||
| C20 | Mairin |
|
55.38 | 0.73 | 0.78 |
|
| |||||
| C21 | Oleanolic acid |
|
29.02 | 0.59 | 0.76 |
|
| |||||
| C22 | Oleanolic acid deriv. |
|
14.24 | 0.65 | 0.7 |
|
| |||||
| C23 | Pinoresinol |
|
4.25 | 0.52 | 0.52 |
|
| |||||
| C24 | Pulchinenoside A_qt |
|
16.91 | 0.12 | 0.77 |
|
| |||||
| C25 | Scoparone |
|
74.75 | 0.85 | 0.09 |
|
| |||||
| C26 | Sitogluside |
|
20.63 | −0.14 | 0.62 |
|
| |||||
| C27 | Sitosteryl acetate |
|
40.39 | 1.39 | 0.85 |
|
| |||||
| C28 | Stigmasterol |
|
43.83 | 1.44 | 0.76 |
|
| |||||
| C29 | Tricosanoic acid |
|
15.29 | 1.18 | 0.3 |
|
| |||||
| C30 | Ursolic acid |
|
16.77 | 0.67 | 0.75 |
|
| |||||
| C31 | ZINC01615307 |
|
56.38 | 0.53 | 0.87 |
|
| |||||
| C32 | β-Sitosterol |
|
5.84 | 1.42 | 0.71 |
A number of these 32 compounds have been shown experimentally to have various biological activities. For example, antioxidative effect of cernuoside (C12; DL = 0.79, OB = 2.69, Caco-2 = −1.51) was experimentally identified [36]. Pinoresinol (C23; DL = 0.52, OB = 4.25, Caco-2 = 0.52) was reported to have anti-inflammatory properties [37]. β-Sitosterol (C32; DL = 0.71, OB = 5.84, Caco-2 = 1.42) and campesterol (C23; DL = 0.72, OB = 5.57, Caco-2 = 1.6) were reported to have the protecting effect by natural and synthetic antioxidants during heating [38]. Antiallergic effects of scoparone (C25; DL = 0.09, OB = 74.75, Caco-2 = 0.85) was experimentally demonstrated in rat model, which attenuates IgE-mediated allergic response in mast cells [39]. Aureusidin (C7; DL = 0.24, OB = 53.42, Caco-2 = 0.07) was reported to have marked antioxidant activity and to be useful for the treatment of several diseases [40, 41], and anemosapogenin (C5; DL = 0.77, OB = 17.87, Caco-2 = 0.07) has antitumor effects [42, 43]. Betulonic acid (C9; DL = 0.78, OB = 16.83, Caco-2 = 0.65) possesses various medical effects, such as antiviral (HIV-1), anticancer, and immunomodulatory activities [44]. Cauloside (C11; DL = 0.4, OB = 6.84, Caco-2 = −0.82) from blue cohosh was reported to inhibit proinflammatory cytokine induction by LPS [45]. Dauricine (C13; DL = 0.9, OB = 23.65, Caco-2 = 0.37) from Asiatic moonseed was reported to have significant antibacterial and anti-inflammatory effects [46], and ergosterol (C14; DL = 0.72, OB = 14.29, Caco-2 = 1.47) [47] from the mushroom and isorhamnetin (C17; DL = 0.31, OB = 49.6, Caco-2 = 0.31) [48] were both found to have anti-inflammatory effects. Furthermore, oleanolic acid (C21; DL = 0.76, OB = 29.02, Caco-2 = 0.59) and ursolic acids (C30; DL = 0.75, OB = 16.77, Caco-2 = 0.67) have been reported to have antioxidative and anti‐inflammatory effect [49, 50]. As mentioned above, PR contains many compounds, which are ubiquitous in plants, herbs, and fruits, with anti-inflammatory, anticancer, and antioxidative effects.
3.2. Target Fishing
These 32 identified active compounds interact with 182 target proteins (Table 2); that is, on average, they interact with 5.7 target genes, which does much to explain the polypharmacological and synergistic effects of PR on multiple targets [51].
Table 2.
Related targets of Pulsatillae Radix.
| UniProt ID | Target | Gene name |
|---|---|---|
| Q92887 | Canalicular multispecific organic anion transporter 1 | ABCC2 |
| P12821 | Angiotensin-converting enzyme | ACE |
| P22303 | Acetylcholinesterase | ACHE |
| P24666 | Low molecular weight phosphotyrosine protein phosphatase | ACP1 |
| P00326 | Alcohol dehydrogenase 1C | ADH1C |
| P35348 | Alpha-1A adrenergic receptor | ADRA1A |
| P35368 | Alpha-1B adrenergic receptor | ADRA1B |
| P25100 | Alpha-1D adrenergic receptor | ADRA1D |
| P08913 | Alpha-2A adrenergic receptor | ADRA2A |
| P18825 | Alpha-2C adrenergic receptor | ADRA2C |
| P08588 | Beta-1 adrenergic receptor | ADRB1 |
| P07550 | Beta-2 adrenergic receptor | ADRB2 |
| P15121 | Aldose reductase | AKR1B1 |
| O60218 | Aldo-keto reductase family 1 member B10 | AKR1B10 |
| P09917 | Arachidonate 5-lipoxygenase | ALOX5 |
| P09923 | Intestinal-type alkaline phosphatase | ALPI |
| P04746 | Pancreatic alpha-amylase | AMY2A |
| P10275 | Androgen receptor | AR |
| P15336 | Cyclic AMP-dependent transcription factor ATF-2 | ATF2 |
| P05023 | Sodium/potassium-transporting ATPase subunit alpha-1 | ATP1A1 |
| P15291 | Beta-1,4-galactosyltransferase 1 | B4GALT1 |
| Q07812 | Apoptosis regulator BAX | BAX |
| P10415 | Apoptosis regulator Bcl-2 | BCL2 |
| Q07817 | Bcl-2-like protein 1 | BCL2L1 |
| O15392 | Baculoviral IAP repeat-containing protein 5 | BIRC5 |
| O43570 | Carbonic anhydrase 12 | CA12 |
| Q8N1Q1 | Carbonic anhydrase 13 | CA13 |
| P00918 | Carbonic anhydrase II | CA2 |
| P22748 | Carbonic anhydrase IV | CA4 |
| P35218 | Carbonic anhydrase 5A, mitochondrial | CA5A |
| Q16790 | Carbonic anhydrase VI | CA9 |
| P62158 | Calmodulin | CALM1 |
| P29466 | Caspase-1 | CASP1 |
| P42574 | Caspase-3 | CASP3 |
| Q14790 | Caspase-8 | CASP8 |
| P55211 | Caspase-9 | CASP9 |
| P13500 | C-C motif chemokine 2 | CCL2 |
| P20248 | Cyclin-A2 | CCNA2 |
| P24385 | G1/S-specific cyclin-D1 | CCND1 |
| P30279 | G1/S-specific cyclin-D2 | CCND2 |
| P60033 | CD81 antigen | CD81 |
| P24941 | Cell division protein kinase 2 | CDK2 |
| P11802 | Cell division protein kinase 4 | CDK4 |
| Q00534 | Cell division protein kinase 6 | CDK6 |
| P38936 | Cyclin-dependent kinase inhibitor 1 | CDKN1A |
| O14757 | Serine/threonine-protein kinase Chk1 | CHEK1 |
| P11229 | Muscarinic acetylcholine receptor M1 | CHRM1 |
| P08172 | Muscarinic acetylcholine receptor M2 | CHRM2 |
| P20309 | Muscarinic acetylcholine receptor M3 | CHRM3 |
| P08173 | Muscarinic acetylcholine receptor M4 | CHRM4 |
| Q15822 | Neuronal acetylcholine receptor subunit alpha-2 | CHRNA2 |
| P36544 | Neuronal acetylcholine receptor protein, alpha-7 chain | CHRNA7 |
| O15111 | NF-kappa-B inhibitor alpha | CHUK |
| P16220 | Cyclic AMP-responsive element-binding protein 1 | CREB1 |
| P15509 | Granulocyte-macrophage colony-stimulating factor | CSF2RA |
| P17538 | Chymotrypsinogen B | CTRB1 |
| P07858 | Cathepsin B | CTSB |
| P10145 | Interleukin-8 | CXCL8 |
| Q16850 | Lanosterol 14-alpha demethylase | CYP51A1 |
| Q9UBM7 | 7-Dehydrocholesterol reductase | DHCR7 |
| P27487 | Dipeptidyl peptidase IV | DPP4 |
| P21728 | Dopamine D1 receptor | DRD1 |
| Q9NRD8 | Dual oxidase 2 | DUOX2 |
| Q6UWV6 | Intestinal alkaline sphingomyelinase | ENPP7 |
| Q99814 | Endothelial PAS domain-containing protein 1 | EPAS1 |
| P03372 | Estrogen receptor | ESR1 |
| Q92731 | Estrogen receptor beta | ESR2 |
| P00742 | Coagulation factor Xa | F10 |
| P00734 | Thrombin | F2 |
| P08709 | Coagulation factor VII | F7 |
| P48023 | Tumor necrosis factor ligand superfamily member 6 | FASLG |
| P49327 | Fatty acid synthase | FASN |
| P05230 | Fibroblast growth factor 1 | FGF1 |
| P09038 | Heparin-binding growth factor 2 | FGF2 |
| P01100 | Proto-oncogene c-Fos | FOS |
| P14867 | Gamma-aminobutyric acid receptor subunit alpha-1 | GABRA1 |
| P47869 | Gamma-aminobutyric-acid receptor alpha-2 subunit | GABRA2 |
| P34903 | Gamma-aminobutyric-acid receptor alpha-3 subunit | GABRA3 |
| P31644 | Gamma-aminobutyric-acid receptor alpha-5 subunit | GABRA5 |
| P17677 | Neuromodulin | GAP43 |
| Q8TDU6 | G-protein coupled bile acid receptor 1 | GPBAR1 |
| P42262 | Glutamate receptor 2 | GRIA2 |
| P49841 | Glycogen synthase kinase-3 beta | GSK3B |
| Q9UII4 | Probable E3 ubiquitin-protein ligase HERC5 | HERC5 |
| P09601 | Heme oxygenase 1 | HMOX1 |
| P01112 | GTPase HRas | HRAS |
| P28845 | Corticosteroid 11-beta-dehydrogenase isozyme 1 | HSD11B1 |
| P80365 | Corticosteroid 11-beta-dehydrogenase isozyme 2 | HSD11B2 |
| P08238 | Heat shock protein HSP 90 | HSP90AB1 |
| P28223 | 5-Hydroxytryptamine 2A receptor | HTR2A |
| P46098 | 5-Hydroxytryptamine receptor 3A | HTR3A |
| P05362 | Intercellular adhesion molecule 1 | ICAM1 |
| P01857 | Ig gamma-1 chain C region | IGHG1 |
| P01584 | Interleukin-1 beta | IL1B |
| P60568 | Interleukin-2 | IL2 |
| P05231 | Interleukin-6 | IL6 |
| O15357 | Phosphatidylinositol-3,4,5-trisphosphate 5-phosphatase 2 | INPPL1 |
| P05412 | Transcription factor AP-1 | JUN |
| Q12809 | Potassium voltage-gated channel subfamily H member 2 | KCNH2 |
| Q12791 | Calcium-activated potassium channel subunit alpha 1 | KCNMA1 |
| O75164 | Lysine-specific demethylase 4A | KDM4A |
| P35968 | Vascular endothelial growth factor receptor 2 | KDR |
| Q99732 | Lipopolysaccharide-induced tumor necrosis factor-alpha factor | LITAF |
| P09960 | Leukotriene A-4 hydrolase | LTA4H |
| P21397 | Amine oxidase [flavin-containing] A | MAOA |
| P27338 | Amine oxidase [flavin-containing] B | MAOB |
| P11137 | Microtubule-associated protein 2 | MAP2 |
| Q16539 | Mitogen-activated protein kinase 14 | MAPK14 |
| P45983 | Mitogen-activated protein kinase 8 | MAPK8 |
| Q13387 | C-Jun-amino-terminal kinase-interacting protein 2 | MAPK8IP2 |
| Q07820 | Induced myeloid leukemia cell differentiation protein Mcl-1 | MCL1 |
| P14174 | L-Dopachrome tautomerase | MIF |
| P03956 | Interstitial collagenase | MMP1 |
| P09238 | Stromelysin-2 | MMP10 |
| P08253 | 72 kDa type IV collagenase | MMP2 |
| P08254 | Stromelysin-1 | MMP3 |
| P14780 | Matrix metalloproteinase-9 | MMP9 |
| P42345 | Serine/threonine-protein kinase mTOR | MTOR |
| P14598 | Neutrophil cytosol factor 1 | NCF1 |
| Q15788 | Nuclear receptor coactivator 1 | NCOA1 |
| Q15596 | Nuclear receptor coactivator 2 | NCOA2 |
| Q9GZQ4 | Neuromedin-U receptor 2 | NMUR2 |
| P35228 | Nitric oxide synthase, inducible | NOS2 |
| P29474 | Nitric oxide synthase, endothelial | NOS3 |
| P15559 | NAD(P)H dehydrogenase [quinone] 1 | NQO1 |
| P04150 | Glucocorticoid receptor | NR3C1 |
| P08235 | Mineralocorticoid receptor | NR3C2 |
| P01111 | GTPase NRas | NRAS |
| P78380 | Oxidized low-density lipoprotein receptor 1 | OLR1 |
| P35372 | Mu-type opioid receptor | OPRM1 |
| P07237 | Protein disulfide-isomerase | P4HB |
| Q14432 | CGMP-inhibited 3′,5′-cyclic phosphodiesterase A | PDE3A |
| P16284 | Platelet endothelial cell adhesion molecule | PECAM1 |
| P06401 | Progesterone receptor | PGR |
| P42336 | Phosphatidylinositol 4,5-bisphosphate 3-kinase catalytic subunit alpha isoform | PIK3CA |
| P48736 | Phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit, gamma isoform | PIK3CG |
| P11309 | Proto-oncogene serine/threonine-protein kinase Pim-1 | PIM1 |
| P61925 | cAMP-dependent protein kinase inhibitor alpha | PKIA |
| P00749 | Urokinase-type plasminogen activator | PLAU |
| P19174 | 1-Phosphatidylinositol 4,5-bisphosphate phosphodiesterase gamma-1 | PLCG1 |
| P09884 | DNA polymerase alpha catalytic subunit | POLA1 |
| P06746 | DNA polymerase beta | POLB |
| P54098 | DNA polymerase catalytic subunit | POLG |
| P27169 | Serum paraoxonase/arylesterase 1 | PON1 |
| Q03181 | Peroxisome proliferator activated receptor delta | PPARD |
| P37231 | Peroxisome proliferator activated receptor gamma | PPARG |
| P17612 | mRNA of PKA catalytic subunit C-alpha | PRKACA |
| P17252 | Protein kinase C alpha type | PRKCA |
| P05129 | Protein kinase C gamma type | PRKCG |
| P07477 | Trypsin-1 | PRSS1 |
| P43115 | Prostaglandin E2 receptor EP3 subtype | PTGER3 |
| O14684 | Prostaglandin E synthase | PTGES |
| P23219 | Prostaglandin G/H synthase 1 | PTGS1 |
| P35354 | Prostaglandin G/H synthase 2 | PTGS2 |
| P18031 | Protein-tyrosine phosphatase 1B | PTPN1 |
| P17706 | T-cell protein-tyrosine phosphatase | PTPN2 |
| P29350 | Hematopoietic cell protein-tyrosine phosphatase | PTPN6 |
| P11217 | Glycogen phosphorylase, muscle form | PYGM |
| Q04206 | Transcription factor p65 | RELA |
| P19793 | Retinoic acid receptor RXR-alpha | RXRA |
| Q14524 | Sodium channel protein type 5 subunit alpha | SCN5A |
| P16581 | E-selectin | SELE |
| Q9HAS3 | Solute carrier family 28 member 3 | SLC28A3 |
| O43826 | Glucose-6-phosphate translocase | SLC37A4 |
| P13866 | Sodium/glucose cotransporter 1 | SLC5A1 |
| P31639 | Sodium/glucose cotransporter 2 | SLC5A2 |
| Q9NY91 | Low affinity sodium-glucose cotransporter | SLC5A4 |
| P23975 | Sodium-dependent noradrenaline transporter | SLC6A2 |
| Q01959 | Sodium-dependent dopamine transporter | SLC6A3 |
| P31645 | Sodium-dependent serotonin transporter | SLC6A4 |
| Q9Y6L6 | Solute carrier organic anion transporter family member 1B1 | SLCO1B1 |
| P40763 | Signal transducer and activator of transcription 3 | STAT3 |
| P59538 | Taste receptor type 2 member 31 | TAS2R31 |
| P01137 | Transforming growth factor beta-1 | TGFB1 |
| P01375 | Tumor necrosis factor | TNF |
| P11388 | DNA topoisomerase II alpha | TOP2A |
| P04637 | Cellular tumor antigen p53 | TP53 |
| P17752 | Tryptophan 5-hydroxylase 1 | TPH1 |
| P14679 | Tyrosinase | TYR |
| P17643 | 5,6-dihydroxyindole-2-carboxylic acid oxidase | TYRP1 |
| P15692 | Vascular endothelial growth factor A | VEGFA |
| P47989 | Xanthine dehydrogenase/oxidase | XDH |
3.3. GO Analysis
For the filtered 182 target genes, 469 biological process terms with P values of < 0.01 were sorted using the functional annotation chart of the DAVID 6.8 Gene Functional Classification Tool and P values were adjusted using Benjamini-Hochberg method. This process resulted in the identification of 25 biological process terms. GO analysis showed that the 182 genes were highly related to inflammation, proliferation, oxidation reduction, and the regulations of apoptosis and signal transduction (Figure 2).
Figure 2.

GO analysis: 25 biological process (BP) of gene ontology (GO) terms sorted by P value < 0.01. Counts of genes and P value related to each BP terms are shown. The y-axis represents BP terms for the target genes, and the x-axis shows counts of genes and −log10(P value).
In detail, phosphatidylinositol-4, 5-bisphosphate 3-kinase catalytic subunit, gamma isoform (PIK3CG), interleukin-6 (IL6), tumor necrosis factor (TNF), C-C motif chemokine 2 (CCL2), prostaglandin E2 receptor EP3 subtype (PTGER3), oxidized low-density lipoprotein receptor 1 (OLR1), prostaglandin G/H synthase 2 (PTGS2), and others are related to “inflammatory response.”
Androgen receptor (AR), interleukin-6 (IL6), heparin-binding growth factor 2 (FGF2), GTPase HRas (HRAS), hematopoietic cell protein-tyrosine phosphatase (PTPN6), and signal transducer and activator of transcription 3 (STAT3) are related to both the “positive regulation and negative regulation of cell proliferation.”
Xanthine dehydrogenase/oxidase (XDH), 5,6-dihydroxyindole-2-carboxylic acid oxidase (TYRP1), prostaglandin G/H synthase 2 (PTGS2), neutrophil cytosol factor 1 (NCF1), lanosterol 14-alpha demethylase (CYP51A1), amine oxidase [flavin-containing] A (MAOA), dual oxidase 2 (DUOX2), and others are associated with “oxidation-reduction process.”
Bcl-2-like protein 1 (BCL2L1), interleukin-6 (IL6), mitogen-activated protein kinase 8 (MAPK8), and cellular tumor antigen p53 (TP53) are associated with both the “positive regulation and negative regulation of apoptotic process.”
To summarize, it is likely that the therapeutic effect of PR in Crohn's disease is due to its anti-inflammatory and repair process and immune system enhancing effects.
3.4. Network Construction and Analysis
To visualize more conveniently the multitargeted effects of PR, network analysis was used to investigate its actions within the context of the whole human genome [52, 53]. As shown in Figure 3, constructed (A) C-T and (B) T-P network demonstrated multicompound and multitargeted effects and relations between various pathways and targets. Circular nodes represent compounds and targets in the C-T network and triangles and circular nodes show pathways and compounds in the T-P network. In both networks, node size was regulated by degree centrality and edges showed interactions between nodes.
Figure 3.

C-T network and T-P network: (a) is a compound-target (C-T) network, and nodes represent compounds and targets; and (b) shows a target-pathway (T-P) network, and circular nodes represent compounds and triangles pathways. Node size is related to the degree and edges demonstrate interactions between nodes.
The C-T network showed 415 interactions between the 182 targets and 32 active compounds of PR. Ursolic acid (C30, degree = 55) had the highest number of interactions with targets, followed by beta-sitosterol (C8, degree = 37) and isorhamnetin (C17, degree = 36), and, thus, these results demonstrated that single molecules can target multiple receptors [54]. Likewise, prostaglandin G/H synthase 2 (PTGS2, degree = 15) displayed the most affinitive connections with compounds, followed by prostaglandin G/H synthase 1 (PTGS1, degree = 13) and nuclear receptor coactivator 2 (NCOA2, degree = 11). According to betweenness centrality results, protein-tyrosine phosphatase 1B (PTPN1, betweenness = 0.11) was second highest followed by PTGS2 (betweenness = 0.12). 28 (88%) Of the 32 active compounds were connected with more than two targets and 86 (47%) of the 182 targets interact with more than one compound. This network analysis results clearly demonstrated the multitargeting natures of herbal compounds; besides it showed that ursolic acid (C30) is the most essential compound in PR.
In addition, 40 pathways related to Crohn's disease were extracted to construct the T-P network. According to degree centrality, “pathways in cancer” (degree = 49) had the highest number of connections with the targets, followed by the “PI3K-Akt signaling pathway” (degree = 34) and “hepatitis B” (degree = 33). In the same manner, phosphatidylinositol 4,5-bisphosphate 3-kinase catalytic subunit alpha isoform (PIK3CA, degree = 33), phosphatidylinositol 4,5-bisphosphate 3-kinase catalytic subunit gamma isoform (PIK3CG, degree = 33), and nuclear factor NF-κB p65 subunit (RELA, degree = 25) demonstrate higher affinitive connections with pathways. Betweenness centrality and degree centrality results were similar; there was only little difference. “Pathways in cancer” (betweenness = 0.19) had the highest betweenness among the targets, which concurred with degree centrality, and this was followed by “neuroactive ligand-receptor interaction” (betweenness = 0.15) and “PI3K-Akt signaling pathway” (betweenness = 0.10). Regarding highest betweenness targets, PIK3CA (betweenness = 0.10), PIK3CG (betweenness = 0.10), and prostaglandin E2 receptor EP3 subtype (PTGER3, betweenness = 0.04) showed most affinitive connections with pathways.
3.5. Target Organ Location Network
The tissue mRNA expression profiles of target genes at the organ level were investigated to identify effects of PR on Crohn's disease. No mRNA expression information of three of genes, muscarinic acetylcholine receptor M1 (CHRM1), G-protein coupled receptor TGR-1 (NMUR2), and taste receptor type 2 member 31 (TAS2R31), was found in the BioGPS. In total, the mRNA expression profiles of 179 of 182 genes were analyzed (Supplementary Table S2). 158 Of the genes displayed above average mRNA expressions in 17 relevant organ tissues, namely, in BDCA4+ dendritic cells, bone marrow, CD14+ monocytes, CD19+ B cells, CD33+ myeloid cells, CD34+ hematopoietic stem cells, CD4+ T cells, CD56+ NK cells, CD8+ T cells, colon, colorectal adenocarcinoma, liver, lymph nodes, lymphoblasts, small intestine, smooth muscle, and thymus. In addition, these 17 organ tissues, retina, prefrontal cortex, pineal, amygdala, cardiac myocyte, heart, whole blood, and other tissues, were also associated with relatively high mRNA expressions. Networks of the tissue mRNA expressions of 158 target genes and PR compounds are shown in Figure 4, nodes represent organs and genes, and node sizes indicate the number of interactions between nodes.
Figure 4.

Target organ location network: the tissue-specific patterns of the mRNA expressions of targets in organs related to the immune system, colon, and small intestine. Nodes represent targets and organ locations. Node size is relative to the degree. Gene expression data was based on gene expression microarrays analysis results in BioGPS.
In detail, among 84 organ tissues, CD33+ myeloid showed the most overexpressed mRNA expression, 104 genes of 158 target genes were overexpressed in CD33+ myeloid, followed by 97 genes in lymphoblasts, 95 in each of smooth muscle and CD34+ hematopoietic stem cells, 91 genes in liver and CD56+ NK cell, 84 in bone marrow and colorectal adenocarcinoma, 82 in BDCA4+ dendritic cells, 75 in CD14+ monocytes, 73 in small intestine, 72 in CD4+ T cells, 70 in CD19+ B cells, 67 in colon, 38 in CD8+ T cells, 23 in thymus, and 22 in lymph nodes. It is evident that most genes were overexpressed in several organs at the same time.
Additionally, intestinal alkaline sphingomyelinase (ENPP7), DNA polymerase catalytic subunit (POLG), and carbonic anhydrase 13 (CA13) recorded beyond average mRNA expressions in all 17 organs. Furthermore, more than 146 (92%) of 158 target genes were overexpressed in two or more organ tissues, suggesting that these organs and compounds in PR are closely related. Furthermore, because the above 17 organs are highly related to immunity, our study results indicate that the therapeutic effects of PR on Crohn's disease are due to its targeting and activating the immune system.
The other 21 target genes, such as acetylcholinesterase (ACHE), pancreatic alpha-amylase (AMY2A), and muscarinic acetylcholine receptor M3 (CHRM3), did not show above average mRNA expression in the 17 organs.
3.6. GEO2R Analysis
Comparison data between normal tissue and Crohn's disease patients' mRNA expression pattern from Gene Expression Omnibus (GEO) was collected. We employed GEOquery and limma R packages of GEO2R tool to identify highly expressed genes in 6 datasets. Dataset accession numbers are as follows: GSE24287, GSE60083, GSE6731, GSE36807, GSE68570, and GSE72780. To sum up, 86 normal samples and 149 Crohn's disease samples were analyzed in each dataset. GEO2R presented the top 250 highly expressed genes in Crohn's disease group compared to the control group and we deleted overlaps, so, in the end, 1182 genes were sorted out.
We found out that there were 23 common genes (Table 3) between target genes of PR and highly expressed genes of Crohn's disease dataset from GEO.
Table 3.
Highly expressed targets in Crohn's disease.
| UniProt ID | Target | Gene name |
|---|---|---|
| Q92887 | Canalicular multispecific organic anion transporter 1 | ABCC2 |
| O60218 | Aldo-keto reductase family 1 member B10 | AKR1B10 |
| P09917 | Arachidonate 5-lipoxygenase | ALOX5 |
| P62158 | Calmodulin | CALM1 |
| Q00534 | Cell division protein kinase 6 | CDK6 |
| P10145 | Interleukin-8 | CXCL8 |
| Q99814 | Endothelial PAS domain-containing protein 1 | EPAS1 |
| P01100 | Proto-oncogene c-Fos | FOS |
| P47869 | Gamma-aminobutyric-acid receptor alpha-2 subunit | GABRA2 |
| P49841 | Glycogen synthase kinase-3 beta | GSK3B |
| P09601 | Heme oxygenase 1 | HMOX1 |
| P05362 | Intercellular adhesion molecule 1 | ICAM1 |
| P01584 | Interleukin-1 beta | IL1B |
| P05231 | Interleukin-6 | IL6 |
| P03956 | Interstitial collagenase | MMP1 |
| Q15788 | Nuclear receptor coactivator 1 | NCOA1 |
| P35228 | Nitric oxide synthase, inducible | NOS2 |
| P01111 | GTPase NRas | NRAS |
| P11309 | Proto-oncogene serine/threonine-protein kinase Pim-1 | PIM1 |
| P61925 | cAMP-dependent protein kinase inhibitor alpha | PKIA |
| P00749 | Urokinase-type plasminogen activator | PLAU |
| P35354 | Prostaglandin G/H synthase 2 | PTGS2 |
| P40763 | Signal transducer and activator of transcription 3 | STAT3 |
| P17752 | Tryptophan 5-hydroxylase 1 | TPH1 |
3.7. Network Pathway
In order to investigate further the effect of PR in Crohn's disease, we performed pathway enrichment analysis (Figure 5). Using the IBD pathway provided by the KEGG pathway database, we confirmed the pathway mapping effect of PR in Crohn's disease by inputting the filtered human target genes into the pathway. The KEGG pathway was constructed according to the current knowledge of the pathogenesis IBD.
Figure 5.

Network pathway: pathway enrichment analysis was performed. 182 filtered target genes were mapped into the Crohn's disease pathway from the Kyoto Encyclopedia of Genes and Genomes (KEGG) to confirm the possible effect pathway of Pulsatillae Radix (PR) on Crohn's disease.
The synthesis of inflammatory cytokines, such as IL-1, IL-6 and TNF-α, is mediated by NF-κB, which is a key regulator of inflammation [55, 56]. We found that oleanolic acid derivative (C22) targets IL-1; ursolic acid (C30) targets all of IL-1, IL-6, and TNF; isorhamnetin (C17), scoparone (C25), and ursolic acid (C30) target nuclear factor NF-κB p65 subunit (RELA); and scoparone (C25) and ursolic acid (C30) target NF-κB inhibitor alpha (CHUK), which suggests that these compounds affect NF-κB activity. Cernuoside (C12) targets IL-2. Beta-sitosterol (C8) targets transforming growth factor beta-1 (TGFB1) and transcription factor AP-1 (JUN), and ursolic acid (C30) also targets transcription factor AP-1 (JUN) and signal transducer and activator of transcription 3 (STAT3). Furthermore, NF-κB and AP-1 in combination are highly related to the initial inflammatory response and to the development of acquired immunity [57]. Moreover, IL-6-mediated STAT3 activation on mucosal T cells may has been suggested to play a role in the development of IBD [58].
4. Discussion
In this study, a network pharmacology method with active compounds filtration, multiple drug target prediction, gene ontology, network analysis, relevant organ location network, and pathway enrichment analysis were employed to determine the targets of PR in relation to Crohn's disease. Our study shows that PR is highly connected to the pathways, biological processes, and organs of Crohn's disease. A pharmacological approach was used to identify the actions of PR at the systems network level.
In this study, pathway mapping result showed that the target genes of PR overlap more with Crohn's disease than with ulcerative colitis. Experimental study also suggested that the markers of both diseases are different from each other [59, 60]. The clinical symptoms of these diseases also differ; for instance, Crohn's disease affect any region of the entire gastrointestinal (GI) tract and all layers of the bowel wall, whereas ulcerative colitis affects only the mucosa and submucosa of colon [61]. Furthermore, ulcerative colitis can be cured by surgery, but Crohn's disease of any part of GI tract tends to relapse after surgery [62]. For this reason, long term management using herbal medicines might be highly recommendable treatment option for Crohn's disease, since herbal medicines have advantages for managing chronic diseases [63, 64].
In addition, IBD is usually referred to as an autoimmune disorder [65], whereas Crohn's disease does not meet the criteria of an autoimmune disorder; rather it is associated with immune deficiency or a secondary immune response to altered intestinal microbiota [65]. Furthermore, ulcerative colitis is a mucosal disease where autoimmune autoantibodies are commonly detected [66], whereas Crohn's disease is a transmural disease, in which pathological changes in gut wall are thought to result from submucosal inflammatory changes [67]. Accordingly, the areas targeted for treatment in these two diseases should be differentiated.
In the present study, we focused on the use of PR as a potential therapy for Crohn's disease. However, herb pairs and combinations are more commonly prescribed and are regarded to be more effective and safer [68]. In terms of the Gunsinjwasa theory of traditional medicine combinations, there are four different roles for each herb in the formula. First, the major component targets the main symptom. Second, the supportive component assists the effect of the major component or targets the secondary symptoms. Third, the neutralizing component allays the side effects or toxins of the major and the supportive component. Fourth, the deliver/retaining component guides the medicine to the targeting part of the body [69, 70]. This combination principle enables not only the enhancement of synergistic medicinal effects but also potentially reduces toxicities [69]. In order to induce better effects and reduce toxicities, an extended analysis of the Pulsatillae Radix (Baekduong, PR), Phellodendri Cortex (Hwangbaek), Coptidis Rhizoma (Hwangryeon), and Citrus reticulata (Jinpi) herb combination (a widely prescribed formula, known as Baekduong decoction) should be explored in the future.
Through GO analysis, we found out that targets of PR are associated with liver diseases such as hepatitis B, hepatitis C, and nonalcoholic fatty liver. In addition, the mRNA expression of 91 of 179 genes in liver was overexpressed according to the target organ location network result. Liver inflammation is as common extraintestinal symptom of Crohn's disease [71], and the number of liver abscesses in Crohn's disease patients is 15 to 20 times higher than that found in the general population [72]. Besides biochemical liver dysfunction [73] and hepatic fibrosis [74] are also frequently found in Crohn's disease, and a number of drugs used to treat IBD have been reported to be associated with liver injury [75]. Furthermore, in an experimental study using a mice model of Crohn's disease-like ileitis, it was found that TLR9 plays an important role in hepatic involvement in IBD [71]. More detailed pathways and relations between liver and Crohn's disease should be discussed in the future.
This study demonstrated that 73 and 67 of 158 targets of PR were highly expressed in small intestine and colon, respectively, results which were accordance with not only the organs commonly affected by Crohn's disease but also the properties of PR in terms of meridian tropism theory. However stronger evidence with another research design is required to support this result.
We confirmed a multicompound and multitarget interaction through the C-T network, which showed that 88% of the active compounds were connected with more than two targets and 47% of the targets interacted with more than one compound. Although it demonstrated the synergetic network of multitarget actions, we should explore more differentiated drug action based on degree centrality and how different the drug actions are when more compounds target one gene in simultaneous way compared to the case of one compound targeting one gene.
5. Conclusion
Previous studies indicate that most compounds in PR have anti-inflammatory, anticancer, and antioxidant effects. In the present study, we found that these compounds interact with multiple targets in a synergetic manner and that PR is highly connected to Crohn's disease related pathways, biological processes, and organs. C-T and T-P network results demonstrated complex multicompound and multitarget drug actions and the relations between targets and various pathways. Furthermore, target genes were found to be overexpressed in organs highly related to immunity. These findings suggest the anti-inflammatory effects of PR, and its enhancements of repair processes and immune system might be of therapeutic benefit in Crohn's disease.
Supplementary Material
Supplementary Table S1 showed the ADME profiles of 57 compounds in PR, and supplementary Table S2 demonstrated the mRNA expression profiles of 179 of 182 genes.
Acknowledgments
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (NRF-2015R1D1A1A01059994).
Conflicts of Interest
The authors have no conflicts of interest to declare.
References
- 1.Baumgart D. C. Crohn's Disease and Ulcerative Colitis: From Epidemiology and Immunobiology to a Rational Diagnostic and Therapeutic Approach. Springer Science & Business Media; 2012. [Google Scholar]
- 2.Baumgart D. C., Sandborn W. J. Crohn's disease. The Lancet. 2012;380(9853):1590–1605. doi: 10.1016/S0140-6736(12)60026-9. [DOI] [PubMed] [Google Scholar]
- 3.Tan W. C., Allan R. N. Diffuse jejunoileitis of Crohn's disease. Gut. 1993;34(10):1374–1378. doi: 10.1136/gut.34.10.1374. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dessein R., Chamaillard M., Danese S. Innate immunity in Crohn’s disease: the reverse side of the medal. Journal of Clinical Gastroenterology. 2008;42(supplement 3):S144–S147. doi: 10.1097/MCG.0b013e3181662c90. [DOI] [PubMed] [Google Scholar]
- 5.Floyd D. N., Langham S., Séverac H. C., Levesque B. G. The economic and quality-of-life burden of crohn’s disease in Europe and the United States, 2000 to 2013: a systematic review. Digestive Diseases and Sciences. 2016;60(2):299–312. doi: 10.1007/s10620-014-3368-z. [DOI] [PubMed] [Google Scholar]
- 6.Marks D. J. B., Rahman F. Z., Sewell G. W., Segal A. W. Crohn's disease: an immune deficiency state. Clinical Reviews in Allergy & Immunology. 2010;38(1):20–31. doi: 10.1007/s12016-009-8133-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Stefanelli T., Malesci A., Repici A., Vetrano S., Danese S. New insights into inflammatory bowel disease pathophysiology: paving the way for novel therapeutic targets. Current Drug Targets. 2008;9(5):413–418. doi: 10.2174/138945008784221170. [DOI] [PubMed] [Google Scholar]
- 8.Hayee B., Rahman F. Z., Sewell G., Smith A. M., Segal A. W. Crohn's disease as an immunodeficiency. Expert Review of Clinical Immunology. 2010;6(4):585–596. doi: 10.1586/eci.10.32. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ananthakrishnan A. N., Xavier R. J., Podolsky D. K. Inflammatory bowel diseases: pathogenesis. Yamada's Textbook of Gastroenterology. 2016:1364–1377. doi: 10.1002/9781119077633. [DOI] [Google Scholar]
- 10.Ng S. C., Lam Y. T., Tsoi K. K. F., Chan F. K. L., Sung J. J. Y., Wu J. C. Y. Systematic review: the efficacy of herbal therapy in inflammatory bowel disease. Alimentary Pharmacology and Therapeutics. 2013;38(8):854–863. doi: 10.1111/apt.12464. [DOI] [PubMed] [Google Scholar]
- 11.Sałaga M., Zatorski H., Sobczak M., Chen C., Fichna J. Chinese herbal medicines in the treatment of IBD and colorectal cancer: a review. Current Treatment Options in Oncology. 2014;15(3):405–420. doi: 10.1007/s11864-014-0288-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Kim S.-H. Anti-inflammatory effect of Pulsatilla koreana. pp. 1–49, 2008.
- 13.Hyo-Jin L. Y.-M. K. Pulsatilla koreana ethanol extract suppress adipocyte differentiation and adipogenesis via down-regulation of PPAR-γ and C/EBPs. pp. 1–47, 2014.
- 14.Li W., Ding Y., Sun Y. N., et al. Oleanane-type triterpenoid saponins from the roots of Pulsatilla koreana and their apoptosis-inducing effects on HL-60 human promyelocytic leukemia cells. Archives of Pharmacal Research. 2013;36(6):768–774. doi: 10.1007/s12272-013-0042-5. [DOI] [PubMed] [Google Scholar]
- 15.Li W., Ding Y., Sun Y. N., et al. Triterpenoid saponins of pulsatilla koreana root have inhibition effects of tumor necrosis factor-α secretion in lipopolysaccharide-induced RAW264.7 cells. Chemical and Pharmaceutical Bulletin. 2013;61(4):471–476. doi: 10.1248/cpb.c12-01034. [DOI] [PubMed] [Google Scholar]
- 16.Park B. H., Jung K. H., Son M. K., et al. Antitumor activity of Pulsatilla koreana extract in anaplastic thyroid cancer via apoptosis and anti-angiogenesis. Molecular Medicine Reports. 2013;7(1):26–30. doi: 10.3892/mmr.2012.1166. [DOI] [PubMed] [Google Scholar]
- 17.Lee S. H., Lee E., Ko Y. T. Anti-inflammatory effects of a methanol extract from Pulsatilla koreana in lipopolysaccharide-exposed rats. BMB Reports. 2012;45(6):371–376. doi: 10.5483/BMBRep.2012.45.6.018. [DOI] [PubMed] [Google Scholar]
- 18.Zhao Y., Li Y., Wang X., Sun W. J. The experimental study of Cortex Eucommiae on meridian tropsim: the distribution study of aucubin in rat tissues. Journal of Pharmaceutical and Biomedical Analysis. 2008;46(2):368–373. doi: 10.1016/j.jpba.2007.09.028. [DOI] [PubMed] [Google Scholar]
- 19.Chang Y.-X., Sun Y.-G., Li J., et al. The experimental study of Astragalus membranaceus on meridian tropsim: The distribution study of astragaloside IV in rat tissues. Journal of Chromatography B: Analytical Technologies in the Biomedical and Life Sciences. 2012;911:71–75. doi: 10.1016/j.jchromb.2012.10.024. [DOI] [PubMed] [Google Scholar]
- 20.Shen M., Tian S., Li Y., et al. Drug-likeness analysis of traditional Chinese medicines: 1. property distributions of drug-like compounds, non-drug-like compounds and natural compounds from traditional Chinese medicines. Journal of Cheminformatics. 2012;4(1, article 31) doi: 10.1186/1758-2946-4-31. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Xu X., Zhang W., Huang C., et al. A novel chemometric method for the prediction of human oral bioavailability. International Journal of Molecular Sciences. 2012;13(6):6964–6982. doi: 10.3390/ijms13066964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Hubatsch I., Ragnarsson E. G. E., Artursson P. Determination of drug permeability and prediction of drug absorption in Caco-2 monolayers. Nature Protocols. 2007;2(9):2111–2119. doi: 10.1038/nprot.2007.303. [DOI] [PubMed] [Google Scholar]
- 23.Tao W., Xu X., Wang X., et al. Network pharmacology-based prediction of the active ingredients and potential targets of Chinese herbal Radix Curcumae formula for application to cardiovascular disease. Journal of Ethnopharmacology. 2013;145(1):1–10. doi: 10.1016/j.jep.2012.09.051. [DOI] [PubMed] [Google Scholar]
- 24.Zhang J., Li Y., Chen S.-S., et al. Systems pharmacology dissection of the anti-inflammatory mechanism for the medicinal herb Folium eriobotryae. International Journal of Molecular Sciences. 2015;16(2):2913–2941. doi: 10.3390/ijms16022913. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Willett P., Barnard J. M., Downs G. M. Chemical similarity searching. Journal of Chemical Information and Computer Sciences. 1998;38(6):983–996. doi: 10.1021/ci9800211. [DOI] [Google Scholar]
- 26.Liu H., Wang J., Zhou W., Wang Y., Yang L. Systems approaches and polypharmacology for drug discovery from herbal medicines: an example using licorice. Journal of Ethnopharmacology. 2013;146(3):773–793. doi: 10.1016/j.jep.2013.02.004. [DOI] [PubMed] [Google Scholar]
- 27.Pang K. S. Modeling of intestinal drug absorption: Roles of transporters and metabolic enzymes (for the gillette review series) Drug Metabolism & Disposition. 2003;31(12):1507–1519. doi: 10.1124/dmd.31.12.1507. [DOI] [PubMed] [Google Scholar]
- 28.Pei T., Zheng C., Huang C., et al. Systematic understanding the mechanisms of vitiligo pathogenesis and its treatment by Qubaibabuqi formula. Journal of Ethnopharmacology. 2016;190:272–287. doi: 10.1016/j.jep.2016.06.001. [DOI] [PubMed] [Google Scholar]
- 29.Yu H., Chen J., Xu X., et al. A systematic prediction of multiple drug-target interactions from chemical, genomic, and pharmacological data. PLoS ONE. 2012;7(5) doi: 10.1371/journal.pone.0037608.e37608 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Wu C. H., Apweiler R., Bairoch A., et al. The Universal Protein Resource (UniProt): an expanding universe of protein information. Nucleic Acids Research. 2006;34(Database issue):D187–D191. doi: 10.1093/nar/gkj161. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Bindea G., Mlecnik B., Hackl H., et al. ClueGO: a Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics. 2009;25(8):1091–1093. doi: 10.1093/bioinformatics/btp101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Smoot M. E., Ono K., Ruscheinski J., Wang P. L., Ideker T. Cytoscape 2.8: new features for data integration and network visualization. Bioinformatics. 2011;27(3):431–432. doi: 10.1093/bioinformatics/btq675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Pan J.-B., Hu S.-C., Shi D., et al. PaGenBase: a pattern gene database for the global and dynamic understanding of gene function. PLoS ONE. 2013;8(12) doi: 10.1371/journal.pone.0080747.e80747 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Zhang W., Tao Q., Guo Z., et al. Systems pharmacology dissection of the integrated treatment for cardiovascular and gastrointestinal disorders by traditional chinese medicine. Scientific Reports. 2016;6:p. 32400. doi: 10.1038/srep32400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Prevoršek Z., Gorjanc G., Paigen B., Horvat S. Congenic and bioinformatics analyses resolved a major-effect Fob3b QTL on mouse Chr 15 into two closely linked loci. Mammalian Genome. 2010;21(3-4):172–185. doi: 10.1007/s00335-010-9252-z. [DOI] [PubMed] [Google Scholar]
- 36.Senthil Kumar K., Kumaresan R. A comparative study on the antioxidant properties of bractein and cernuoside by the DFT method. Monatshefte fur Chemie. 2013;144(10):1513–1524. doi: 10.1007/s00706-013-1024-5. [DOI] [Google Scholar]
- 37.Guo T., Deng Y.-X., Xie H., et al. Antinociceptive and anti-inflammatory activities of ethyl acetate fraction from Zanthoxylum armatum in mice. Fitoterapia. 2011;82(3):347–351. doi: 10.1016/j.fitote.2010.11.004. [DOI] [PubMed] [Google Scholar]
- 38.Kmiecik D., Korczak J., Rudzińska M., Kobus-Cisowska J., Gramza-Michałowska A., Hȩś M. β-Sitosterol and campesterol stabilisation by natural and synthetic antioxidants during heating. Food Chemistry. 2011;128(4):937–942. doi: 10.1016/j.foodchem.2011.03.122. [DOI] [PubMed] [Google Scholar]
- 39.Choi Y. H., Yan G. H. Anti-allergic effects of scoparone on mast cell-mediated allergy model. Phytomedicine. 2009;16(12):1089–1094. doi: 10.1016/j.phymed.2009.05.003. [DOI] [PubMed] [Google Scholar]
- 40.Kumar K. S., Kumaresan R. A quantum chemical study on the antioxidant properties of aureusidin and bracteatin. International Journal of Quantum Chemistry. 2011;111(15):4483–4496. doi: 10.1002/qua.22964. [DOI] [Google Scholar]
- 41.Roussaki M., Gaitanarou A., Diamanti P. C., et al. Encapsulation of the natural antioxidant aureusidin in biodegradable PLA nanoparticles. Polymer Degradation and Stability. 2014;108:182–187. doi: 10.1016/j.polymdegradstab.2014.08.004. [DOI] [Google Scholar]
- 42.Yanping Q. Effects of velvet for the tumor and immune function in tumor-bearing mice. Journal of Heilongjiang Bayi Agricultural University. 2012;1:p. 17. [Google Scholar]
- 43.Dan F., Bin Z. C. The anti-tumor effect of the extract from Radix Pulsatillae. Chinese Journal of Hospital Pharmacy. 2003;9:p. 009. [Google Scholar]
- 44.Vasilevsky S. F., Govdi A. I., Sorokina I. V., et al. Rapid access to new bioconjugates of betulonic acid via click chemistry. Bioorganic and Medicinal Chemistry Letters. 2011;21(1):62–65. doi: 10.1016/j.bmcl.2010.11.072. [DOI] [PubMed] [Google Scholar]
- 45.Lee Y., Jung J.-C., Ali Z., Khan I. A., Oh S. Anti-inflammatory effect of triterpene saponins isolated from blue cohosh (caulophyllum thalictroides) Evidence-based Complementary and Alternative Medicine. 2012;2012:8. doi: 10.1155/2012/798192.798192 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Chen N., Ji Y.-B., Song D.-X., Xu C.-R., Song H., Li J. Effects of Dauricine concentration in asiatic moonseed by different extraction solvents and methods. Applied Mechanics and Materials. 2013;411–414:3162–3165. doi: 10.4028/www.scientific.net/AMM.411-414.3162. [DOI] [Google Scholar]
- 47.Ma L., Chen H., Dong P., Lu X. Anti-inflammatory and anticancer activities of extracts and compounds from the mushroom Inonotus obliquus. Food Chemistry. 2013;139(1–4):503–508. doi: 10.1016/j.foodchem.2013.01.030. [DOI] [PubMed] [Google Scholar]
- 48.Boesch-Saadatmandi C., Loboda A., Wagner A. E., et al. Effect of quercetin and its metabolites isorhamnetin and quercetin-3-glucuronide on inflammatory gene expression: role of miR-155. Journal of Nutritional Biochemistry. 2011;22(3):293–299. doi: 10.1016/j.jnutbio.2010.02.008. [DOI] [PubMed] [Google Scholar]
- 49.Liu J. Oleanolic acid and ursolic acid: research perspectives. Journal of Ethnopharmacology. 2005;100(1-2):92–94. doi: 10.1016/j.jep.2005.05.024. [DOI] [PubMed] [Google Scholar]
- 50.Tsai S.-J., Yin M.-C. Antioxidative and anti-inflammatory protection of oleanolic acid and ursolic acid in PC12 cells. Journal of Food Science. 2008;73(7):H174–H178. doi: 10.1111/j.1750-3841.2008.00864.x. [DOI] [PubMed] [Google Scholar]
- 51.Liu J., Pei T., Mu J., et al. Systems pharmacology uncovers the multiple mechanisms of Xijiao Dihuang decoction for the treatment of viral hemorrhagic fever. Evidence-Based Complementary and Alternative Medicine. 2016;2016:17. doi: 10.1155/2016/9025036.9025036 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Berger S. I., Iyengar R. Network analyses in systems pharmacology. Bioinformatics. 2009;25(19):2466–2472. doi: 10.1093/bioinformatics/btp465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Li P., Qi L.-W., Liu E.-H., Zhou J.-L., Wen X.-D. Analysis of Chinese herbal medicines with holistic approaches and integrated evaluation models. TrAC—Trends in Analytical Chemistry. 2008;27(1):66–77. doi: 10.1016/j.trac.2007.11.005. [DOI] [Google Scholar]
- 54.Espinoza-Fonseca L. M. The benefits of the multi-target approach in drug design and discovery. Bioorganic and Medicinal Chemistry. 2006;14(4):896–897. doi: 10.1016/j.bmc.2005.09.011. [DOI] [PubMed] [Google Scholar]
- 55.Tak P. P., Firestein G. S. NF-κB: a key role in inflammatory diseases. Journal of Clinical Investigation. 2001;107(1):7–11. doi: 10.1172/JCI11830. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Xiao W., Hodge D. R., Wang L., Yang X., Zhang X., Farrar W. L. NF-κB activates IL-6 expression through cooperation with c-Jun and IL6-AP1 site, but is independent of its IL6-NFκB regulatory site in autocrine human multiple myeloma cells. Cancer Biology & Therapy. 2004;3(10):1007–1017. doi: 10.4161/cbt.3.10.1141. [DOI] [PubMed] [Google Scholar]
- 57.Glass C. K., Saijo K. Nuclear receptor transrepression pathways that regulate inflammation in macrophages and T cells. Nature Reviews Immunology. 2010;10(5):365–376. doi: 10.1038/nri2748. [DOI] [PubMed] [Google Scholar]
- 58.Mitsuyama K., Matsumoto S., Masuda J., et al. Therapeutic strategies for targeting the IL-6/STAT3 cytokine signaling pathway in inflammatory bowel disease. Anticancer Research. 2007;27(6A):3749–3756. [PubMed] [Google Scholar]
- 59.Yamazaki K., Umeno J., Takahashi A., et al. A genome-wide association study identifies 2 susceptibility loci for Crohn's disease in a Japanese population. Gastroenterology. 2013;144(4):781–788. doi: 10.1053/j.gastro.2012.12.021. [DOI] [PubMed] [Google Scholar]
- 60.Costa F., Mumolo M. G., Ceccarelli L., et al. Calprotectin is a stronger predictive marker of relapse in ulcerative colitis than in Crohn's disease. Gut. 2005;54(3):364–368. doi: 10.1136/gut.2004.043406. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Ament M. E. Inflammatory disease of the colon: ulcerative colitis and Crohn's colitis. The Journal of Pediatrics. 1975;86(3):322–334. doi: 10.1016/S0022-3476(75)80958-9. [DOI] [PubMed] [Google Scholar]
- 62.Kristo I., Stift A., Bergmann M., Riss S. Surgical recurrence in Crohn's disease: are we getting better? World Journal of Gastroenterology. 2015;21(20):6097–6100. doi: 10.3748/wjg.v21.i20.6097. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Elder C., Ritenbaugh C., Aickin M., et al. Reductions in pain medication use associated with traditional Chinese medicine for chronic pain. Issues. 2016;16:18–23. doi: 10.7812/tpp/12.967. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Tan M., Win M., Khan S. A. The use of complementary and alternative medicine in chronic pain patients in Singapore: a single-centre study. Annals of the Academy of Medicine, Singapore. 2013;42(3):133–137. [PubMed] [Google Scholar]
- 65.Behr M. A., Divangahi M., Lalande J.-D. What's in a name? The (mis)labelling of Crohn's as an autoimmune disease. The Lancet. 2010;376(9736):202–203. doi: 10.1016/S0140-6736(10)60282-6. [DOI] [PubMed] [Google Scholar]
- 66.Das K. M., Biancone L. Is IBD an autoimmune disorder? Inflammatory Bowel Diseases. 2008;14(S2):S97–S101. doi: 10.1002/ibd.20723. [DOI] [PubMed] [Google Scholar]
- 67.Crohn B. B., Ginzburg L., Oppenheimer G. D. Regional ileitis: a pathologic and clinical entity. Journal of the American Medical Association. 1932;99(16):1323–1329. doi: 10.1001/jama.1932.02740680019005. [DOI] [Google Scholar]
- 68.Riaz A., Khan R. A., Ahmed S., Afroz S. Assessment of acute toxicity and reproductive capability of a herbal combination. Pakistan Journal of Pharmaceutical Sciences. 2010;23(3):291–294. [PubMed] [Google Scholar]
- 69.Kim H. U., Ryu J. Y., Lee J. O., Lee S. Y. A systems approach to traditional oriental medicine. Nature Biotechnology. 2016;33(3):264–268. doi: 10.1038/nbt.3167. [DOI] [PubMed] [Google Scholar]
- 70.Hsieh H.-Y., Chiu P.-H., Wang S.-C. Epigenetics in traditional chinese pharmacy: a bioinformatic study at pharmacopoeia scale. Evidence-based Complementary and Alternative Medicine. 2011;2011:10. doi: 10.1093/ecam/neq050.816714 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Omenetti S., Brogi M., Garg R., et al. Essential role for Toll-like receptor 9 in the pathogenesis of liver inflammation in a murine model of Crohns disease-like ileitis (P3147) The Journal of Immunology. 2013;190(1 supplement):35–43. [Google Scholar]
- 72.Mir-Madjlessi S. H., McHenry M. C., Farmer R. G. Liver abscess in Crohn's disease. Report of four cases and review of the literature. Gastroenterology. 1986;91(4):987–993. doi: 10.1016/0016-5085(86)90704-3. [DOI] [PubMed] [Google Scholar]
- 73.Perrett A. D., Higgins G., Johnston H. H., et al. The liver in Crohn's disease. Gastroenterology. 1971;40(158):187–209. doi: 10.1093/oxfordjournals.qjmed.a067265. [DOI] [PubMed] [Google Scholar]
- 74.Eade M. N., Cooke W. T., Williams I. A. Liver disease in crohn's disease: a study of 100 consecutive patients. Scandinavian Journal of Gastroenterology. 1971;6(3):199–204. doi: 10.3109/00365527109180693. [DOI] [PubMed] [Google Scholar]
- 75.Carvalheiro J., Mendes S., Sofia C. Infliximab induced liver injury in Crohn's disease: a challenging diagnosis. Journal of Crohn's and Colitis. 2014;8(5):436–437. doi: 10.1016/j.crohns.2013.11.013. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Supplementary Table S1 showed the ADME profiles of 57 compounds in PR, and supplementary Table S2 demonstrated the mRNA expression profiles of 179 of 182 genes.