

Abstract
Increasing evidence underscores the interesting ability of tryptophan to regulate immune responses. However, the exact mechanisms of tryptophan’s immune regulation remain to be determined. Tryptophan catabolism via the kynurenine pathway is known to play an important role in tryptophan’s involvement in immune responses. Interestingly, quinolinic acid, which is a neurotoxic catabolite of the kynurenine pathway, is the major pathway for the de novo synthesis of nicotinamide adenine dinucleotide (NAD+). Recent studies have shown that NAD+, a natural coenzyme found in all living cells, regulates immune responses and creates homeostasis via a novel signaling pathway. More importantly, the immunoregulatory properties of NAD+ are strongly related to the overexpression of tryptophan hydroxylase 1 (Tph1). This review provides recent knowledge of tryptophan and NAD+ and their specific and intriguing roles in the immune system. Furthermore, it focuses on the mechanisms by which tryptophan regulates NAD+ synthesis as well as innate and adaptive immune responses.
Keywords: Nicotinamide adenine dinucleotide, NAD+, immune responses, tryptophan, kynurenine pathway
Introduction
Tryptophan was initially discovered by Hopkins and Cole in 1901 after isolation from casein protein.1 Tryptophan is 1 of the 8 essential amino acids that cannot be synthesized by the human body.2 Although tryptophan has the lowest concentration in the body of all the essential amino acids, only very low concentrations are required for healthy nutrition.2 Tryptophan has been extensively studied due to its important metabolic functions, which include the synthesis of hormones such as kynurenine, serotonin, tryptamine, melatonin, and essential coenzymes, particularly nicotinamide adenine dinucleotide (NAD+), that are known to play critical physiological functions.2,3
Tryptophan has been associated with several clinical disorders. Historically, tryptophan was associated with pellagra, a dermatological condition associated with deficiency in tryptophan and niacin.4,5 Most notably, tryptophan is involved in behavioral changes through synthesis of serotonin and tryptamine, 2 major players in neuropsychiatric disorders.2,6 Tryptophan deprivation induces depression, separation anxiety,7,8 chronic fatigue syndrome, and fibromyalgia. Furthermore, increasing evidence shows the role of tryptophan in brain tumor pathogenesis9,10 and other cancer types, such as colorectal, breast, and bladder cancers.7,11–13
Interestingly, tryptophan deprivation also sensitizes activated T-cells to apoptosis in novo14 and favors the induction of regulatory T-cells (Tregs) via dendritic cells.15,16 Although the role of tryptophan in regulating the immune system has been previously described,17 research in this area has been scarce until recently. Several recent publications highlighted the involvement of tryptophan in inflammation, immune activation, and immune tolerance,11,13,18–20 raising awareness of tryptophan catabolism and its role in the immune system.
Kynurenine is an important biosynthetic pathway in tryptophan catabolism.21 Moreover, most of the immunologic function accredited to tryptophan was attributed to the kynurenine pathway.3,18,22,23 More importantly, the kynurenine pathway is essential for the de novo synthesis of NAD+24 and acts as a substrate for several other molecules.25,26 Of note, the NAD+ levels have been described as very volatile; therefore, the de novo synthesis of NAD+ plays an important role in maintaining adequate NAD+ levels.27 Quinolinic acid (QUIN), a product of tryptophan degradation, was found to be essential for de novo synthesis of NAD+ and may serve as a precursor of NAD+ in the tryptophan/kynurenine pathway. Although QUIN is neurotoxic,9,22,28 recent publications have underscored the capacity of NAD+ to regulate T-cell fate and promote homeostasis29–31 independent of the classical pathways that have considered cytokine environment and transcription factors as the major regulator of CD4+ T-cell differentiation and adaptive immune response.
The kynurenine pathway is initiated by 2 rate-limiting enzymes: indoleamine 2,3-dioxygenase (IDO) and tryptophan 2,3-dioxygenase (TDO), which are essential in tryptophan catabolism. Indoleamine 2,3-dioxygenase has been found in many immune cells, such as dendritic cells (DCs) and macrophages,32,33 whereas TDO has been found in the liver and neurons.32 Moreover, both IDO34 and TDO11 are present in several malignancies such as prostate, colorectal, pancreatic, and breast cancers. Increasing evidence points toward an important role of IDO in the regulation of the immune system.12,17,35 Interestingly, IDO is involved in the activation of the protein aryl hydrocarbon receptor (AhR),32 a ligand-activated transcription factor that regulates the immune system and inflammatory responses.36–38 Indeed, on activation, AhR can promote the differentiation of type 1 regulatory cells (Tr1)39,40 and control Treg/TH17 differentiation,37,41 critical CD4+ T-cell subsets involved in immunosuppression or pro-inflammatory responses. Furthermore, IDO promotes long-term tolerance by DCs through transforming growth factor β (TGF-β) in a noninflammatory context in novo.42,43
Collectively, these findings indicate the existence of other pathways that regulate the immune system. These alternative pathways play key roles in a myriad of diseases and may serve as an alternative to develop novel therapeutic approaches. In this mini-review, we discuss the current knowledge of the increasing evidence of tryptophan and its role in immune regulation, with an overview of the kynurenine/QUIN/NAD+ pathway and emphasize missing links for which more research is needed. We also discuss possible models and key players in immune regulation via the kynurenine/QUIN/NAD+ pathway.
Tryptophan and Its Role in Immune Responses
Tryptophan has been implicated in many metabolic processes, including the metabolic synthesis of major proteins, such as kynurenine, serotonin, tryptamine, melatonin, and other coenzymes, particularly NAD/NADP (NAD phosphate).2 Although these metabolic processes are critical, the role of tryptophan in the immune system is still poorly understood.
The kynurenine pathway is a major route for the catabolism of tryptophan and has an important role in the immune system.17,21,32,44 The kynurenine pathway is initiated by cleavage of the indole ring, which is initiated by tryptophan 2,3-dioxygenase 2 (TDO-2), indoleamine 2,3-dioxygenase 1 (IDO-1), or IDO-2, which shares similarities with IDO-1 but has limited catabolic activity compared with TDO-2 and IDO-1.22,45–47 TDO-2 has been found primarily in the liver and central nervous system (CNS), but recent publications have identified TDO-2 in different types of cancer.10,23 TDO-2 suppresses antitumor immune responses and promotes tumor cell survival.48 Furthermore, TDO-2 promotes metastasis in breast cancer49,50 and the progression of lung cancer in vivo.51 Moreover, enzymatically active TDO-2 is overexpressed in a significant proportion of human tumors.10 TDO-2 overexpression has been correlated with poor prognosis in high-grade gliomas.45 Current immunosuppressive therapies aim to inhibit TDO-2.11 Pilotte et al10 were able to restore the ability to reject TDO-expressing tumors using a self-developed TDO-inhibitor (compound 680C91). Furthermore, Crosignani et al52 presented their preclinical assessment of a TDO-2-inhibitor (iTeos) in the 29th Annual Meeting of the Society for Immunotherapy Cancer, with similar results on a p815 mastocytoma tumor model overexpressing TDO-2.
Contrary to TDO-2, IDO-1 is found extrahepatically and is present in several immune cells.35,53,54 Indeed, IDO-1 is present in human tumor cells and can prevent tumor cell rejection due to a lack of accumulation of specific T-cells.34 Moreover, IDO-1 is upregulated by several pro-inflammatory molecules.55,56 The interaction between IDO-1 and DCs has been the most studied to date. The rapid switch in function from immunogenic to tolerogenic activity is dependent on IFN-γ, whereas the tolerogenic phenotype is conferred by IDO-1. Indeed, IFN-γ is a potent activator of gene expression and enzymatic activity of IDO-1.57,58 The tolerogenic phenotype induced by IDO-1 plays a double role in immune responses. Indoleamine 2,3-dioxygenase expands human CD4+CD25highFoxp3+ Tregs by promoting maturation of DCs.59 Second, IDO-expressing DCs suppressed allograft rejection by expansion of CD4+CD25highFoxp3+ Tregs.33 In contrast, IDO promotes tolerance toward tumor cells and can promote resistance toward immunotherapies.60 More importantly, tumors expressing Foxp3+/IDO+ are associated with more advanced disease.44 Interestingly, therapies targeting IDO have been shown to be effective against different types of cancer by measuring messenger RNA.12,61,62 Similar interactions between IDO and other cells of the immune system have been described. Among these immune cells, we found interactions with macrophages,54,63 mast cells,32,42 and Tregs.64,65
The initial activation models for the kynurenine pathway showed a redundant activation through TDO-2 and IDO-1.22 However, present evidence suggests specific roles for each of these enzymes in the tryptophan catabolic pathway. As Puccetti notes, in the early events of systemic inflammation, cortisol induces the release of acute phase proteins such as TDO-2. TDO-2 release activates the AhR, as explained below, transcribing interleukin (IL)-10 and IDO-1. Finally, the transcription of IL-10 and IDO-1 will create homeostasis.47 This model indicates different gene activation processes and roles for TDO-2 and IDO-1.47,66
Immune Implications of the AhR and Tryptophan Catabolic Enzymes
We have recently shown that kynurenine was able to promote mast cell activation by activating the AhR.32 As previously reported, activation of AhR has been linked to activation of Tr1 and to control of Tregs and TH17 differentiation.39–41 Interestingly, most immune cells—T-cells, B cells, macrophages, DCs, and natural killer cells—express AhR.67 These findings suggest an important role of AhR in regulation of the immune system.
AhR is a transcription factor in the cytosol that is activated on ligand binding.67,68 AhR is a key regulator of xenobiotic toxicity, such as 2,3,7,8,9-tetracholodibenzo-p-dioxin (TCDD) and dioxin, the most prominent member of the environmental pollutants.36,67 However, AhR has also been identified as a key player in several signaling cascades and other cell biological events. Most likely, one of the most interesting and newly identified aspects of AhR is its involvement in immune regulation and tolerance.
AhR has been shown to regulate CD4+CD25+Foxp3+ Tregs as well as TH17 cell fate, which are 2 major subsets of the adaptive immune system; extensive studies have underscored the importance of Tregs and TH17 cell balance.69–71 Indeed, Tregs are responsible for homeostasis and immune tolerance and are very robust immunosuppressive cells, whereas TH17 cells are considered to be pro-inflammatory cells involved in a myriad of diseases including autoimmunity.72,73 Interestingly, AhR has been found to control Tregs and TH17 cell differentiation in experimental autoimmune encephalomyelitis (EAE),41 which are the most commonly used experimental model for multiple sclerosis. In this study, the authors found that AhR activation by TCDD controls Treg development in vivo, suppressing the severity of EAE. Furthermore, activation of AhR via another ligand, 6-formylindolo 3,2-b carbazole (FICZ), increased TH17 differentiation, thereby increasing the severity of EAE.41 Interestingly, FICZ has been shown to control Tregs via the activation of epidermal DCs when keratinocytes were exposed to UV-B light.74 Indeed, UV-B light activation of AhR is believed to generate FICZ in the skin.75 FICZ and TCDD have shown exert different effects on TH17/Treg development either in vitro or in vivo.76 It is now clear that AhR modulates the immune system in response to ligands of diverse origin.37 Furthermore, AhR ligands affect several pathways and responses depending on the tissue/cellular context.36 There is a range of potential physiological ligands for AhR.77 However, the exact molecular mechanisms remain elusive and are subject of current research.
The role of AhR as a mediator of homeostasis and immune tolerance is further supported by its involvement in the regulation of type 1 Tregs (Tr1). Tr1 cells have the capacity to coproduce IFN-γ and IL-1030,78 and have emerged as an important immunosuppressive CD4+ T-cell subset that dampens inflammatory responses, promotes transplant tolerance, and prevents tissue damage.29,79,80 Gandhi et al40 observed induction of Tr1 cells by activation of AhR through TCDD in human cells. Interestingly, in the same study, they found that adding TGF-β1 to the activation of AhR through TCDD induced functional Tregs.40 Independent of the type of regulatory cell that is activated, AhR activation through TCDD induces a significantly higher expression of IL-10 in human cells.40 Simultaneously, Apetoh and Quintana39 corroborated the findings from Gandhi. Furthermore, they showed that AhR interacts with c-Maf, a transcription factor shown to regulate synthesis of IL-10,81 to contribute to Tr1-cell differentiation.39 The high expression of IL-10 could protect animals from EAE. Tullius et al29 and Wang et al31 have also shown that IL-10 is capable of promoting homeostasis and protecting from EAE via the direct activation of Tr1 cells.
As previously mentioned, the metabolites generated from the tryptophan/IDO/TDO pathway act as ligands for AhR.67 Indeed, the IDO-1/AhR interaction appears to be a more effective pathway for 2 main reasons: it is self-regulated because AhR presides over the regulatory proteolysis of IDO-1, and it acts in a feed-forward loop initiated by kynurenine, followed by AhR activation, promoting IDO-1 transcription.47 Interestingly, the IDO-1/AhR axis is very dependent on the presence or absence IL-6, an essential pro-inflammatory cytokine for the induction of TH17 cells, and it can be antagonized by IL-10.82
The absence of IL-6 is paramount for the IDO-1/AhR axis to be effective in IL-10 production and to create homeostasis.36 However, in the early phases of inflammation, IL-6 is required as an initial defense mechanism.83,84 During the early phase of inflammation, the source of kynurenine must be from somewhere other than the IDO-1/AhR axis.47 Once the IDO-1/AhR axis is started through an external kynurenine source, AhR can repress IL-6 via the transcription of IL-10.36 An interesting model suggested by Puccetti indicates cortisol as an outsource for kynurenine through TDO-2.47 Cortisol is released with pro-inflammatory cytokines in the early phases of inflammation and releases acute phase proteins by the liver, one of which is TDO-2.47,48,85 Interestingly, clinical trials indicated that cortisol and IL-6 increased the percentages of Tregs.84 Furthermore, both cortisol and IL-6 can produce high amounts of TDO-2 by the liver activating the kynurenine pathway.86 Indeed, other sources for the activation of AhR might be involved in this process and warrant further investigation.
Immunoregulatory Properties of NAD+ and Tryptophan
The major catabolic pathway of tryptophan is the kynurenine pathway, which ultimately leads to biosynthesis of NAD+,18 a vital cofactor that can rewire metabolism, activate sirtuins, and maintain mitochondrial fitness.87
Nicotinamide adenine dinucleotide was originally discovered by Sir Arthur Harden and his colleagues in 1906.88 However, its key role in hydrogen transfer in biochemical reactions was not discovered until 1935.87 Nicotinamide adenine dinucleotide plays an important role in energy metabolism, but the roles of NAD+ have expanded beyond its initial function as a coenzyme linking cellular metabolism to several signaling and transcriptional events.27,29,30,87,89,90
One of the most interesting newly discovered roles of NAD+ is probably its involvement in homeostasis and immune responses. Data increasingly support the notion of NAD+-regulating immune responses. For example, NAD+ regulates cytokine production of peripheral blood lymphocytes and monocytes.91,92 Based on these findings, we investigated the role of NAD+ and T-cells. We found that intraperitoneal injections of NAD+ protected mice against autoimmune diseases and prolonged allograft survival after skin transplantation.29,30 In our studies, NAD+ induced a systemic increase in IL-10 despite the reduction in Tregs and increase in TH17 cells. The increased IL-10 production was mediated via the immunosuppressive Tr1 cells. Moreover, NAD+ induced T-cell differentiation, despite the cytokine milieu and well-established transcription factors. Interestingly, a genome-wide gene expression profile assessed by microarray analysis showed tryptophan hydroxylase 1 (Tph1) to be upregulated under the influence of NAD+, mostly described on mast cells. Collectively, our data indicate NAD+ as a potent immune regulator through tryptophan catabolism.
Several publications have indicated a strong relation between NAD+ and tryptophan, particularly with the kynurenine pathway.27,89,93,94 Furthermore, NAD+ is essential for cells under physiological stress to avoid cell damage.17 Nicotinamide adenine dinucleotide can be synthesized through niacin, which is typically present in an adequate diet2,87,94 and is the main source of NAD+ synthesis or de novo through tryptophan and the kynurenine pathway.27,95
De novo synthesis of NAD+ starts with the catalysis of tryptophan by either TDO-2 or IDO-195 into formylkynurenine followed by conversion into l-kynurenine. Three more enzymatic reactions described elsewhere9 lead to the conversion into 2-amino-3-carboxymuconate semialdehyde (ACMS). ACMS can go either through an enzymatic pathway leading to total oxidation or through a nonenzymatic cyclization to QUIN. QUIN is essential for de novo synthesis of NAD+.17,87 Interestingly, QUIN has been shown to be neurotoxic.28,96 Moreover, QUIN accumulates in human glioma cells and is associated with malignant phenotypes.97 Furthermore, QUIN was found to confer resistance of gliomas to oxidative stress. The cytotoxic properties of QUIN in the brain have extensively been discussed in the literature, but very little is known about its role in immune responses. Nicotinamide adenine dinucleotide has been found to induce programmed cell death and hinder Tregs.98,99 Although we were able to confirm that NAD+ induces apoptosis of naïve CD4+ T-cells and reduces the number of Tregs,29 CD4+ T-cells that were differentiated into TH1, TH2, TH17, and iTreg were protected against apoptosis by NAD+.
Our previous studies showed the importance of the tryptophan/NAD+ pathway in inflammation, including autoimmune diseases and transplantation. These studies were performed in the context of antigen and antigen presentation. Our unpublished data indicate that the NAD+/tryptophan signaling pathway can trigger innate and adaptive immune responses in the absence of antigen. This is consistent with previous studies reporting that tryptophan administration promotes wound healing.100 However, the effects of exogenous tryptophan in immune responses have not been well studied. Interestingly, a previous study by Swanson et al101 showed that exogenous addition of tryptophan to recover lung interstitial antigen-presenting cells (APCs)–induced T-cell proliferation in mixed leukocyte reactions. Of note, the addition of exogenous tryptophan can restore pathogens and replication of some bacteria and viruses43; thus, further studies are required to elucidate the role of exogenous tryptophan in immune responses.
Antigen-presenting cells play a unique role in the regulation of adaptive immune responses due to their ability to capture, process, and present antigens via their major histocompatibility complex (MHC) cell surface molecule to the T-cell receptor (TCR) to mount a restricted MHC immune response.102–104 DCs are still considered the major APCs bridging innate and adaptive immune responses.105,106 The mode of action of DCs is mediated through at least 3 signals: (1) TCR activation, (2) activation of co-stimulatory molecules, and (3) secretion of chemokines and pro-inflammatory cytokines.105 In addition, DCs can regulate innate and adaptive immune responses by recognizing pathogen-associated molecular patterns, such as microbial nucleic acids, lipoproteins, and carbohydrates, or damage-associated molecular patterns that are released from injured cells via intracellular or surface-expressed pattern recognition receptors (PRRs).107 Thus, our unpublished data, indicating that the tryptophan/NAD+ pathway can trigger the immune system in the absence of pathogen invasion or PRRs’ activation, suggest a novel cellular and molecular signaling pathway distinct from the classical ag-APCs (DCs)-MHCII-TCR signaling machinery. It would therefore be interesting to further investigate the path used by tryptophan/NAD+, particularly the cells and molecules involved.
Tryptophan and NAD+ in the CNS
Active interactions between the CNS and the immune system are essential to maintain CNS integrity and homeostasis. The intact blood-brain barrier allows traffic of peripheral immune cells, CNS neurons and glia regulate macrophage and lymphocyte responses, and microglia play key roles in both innate and adaptive immune responses, providing the CNS with the strategic means to respond to pathogens. Thus, the past 30 years have seen significant changes in notions of CNS immune privilege. The immune implications of tryptophan have begun to extend into the CNS as well. Tryptophan is also the sole precursor of an important neurotransmitter, serotonin, or 5-hydroxytryptophan. Serotonin is synthesized from tryptophan through hydroxylation and decarboxylation, processes that are catalyzed by tryptophan hydroxylase (Tph) and the aromatic acid decarboxylase, with Tph being the rate-limiting step in the synthesis. Of the 2 isoforms of Tph, Tph1 is expressed both in the periphery (skin, enterochromaffin cells of the gut, mast cells, and the pineal gland) and in the CNS, and Tph2 is expressed in neurons of the central, peripheral, and enteric nervous systems. Indeed, tph1-null mice have gross abnormalities such as an overall size reduction, altered morphology of the rhombencephalon regions and neopallial cortex, alterations in gait dynamics, and reduced motivation locomotion.108 Tph2-null mice show alterations in developmental milestones, brain-to-body weight ratio, and substantial deficits in numerous validated tests of social interaction and communication with highly repetitive and compulsive behaviors.109 There appears to be no substantial overlap in the expression of the 2 tph isoforms by adulthood. Dietary tryptophan can also regulate serotonin synthesis in the brain and can affect mood, behavior, and cognition. Serotonin synthesis is thus an important tryptophan pathway and is a topic of intense research due to its broad impact as a neuromodulator. Multiple lines of evidence support its association with numerous neuropsychiatric conditions such as Parkinson disease, schizophrenia, depression, anxiety, and, of great significance, autism.
Autism, which is characterized as both a neurodevelopmental and a neuropsychiatric disease, has also distinctly evolved into a disease of the immune system.110–112 One or more core behavioral symptoms, such as impaired social recognition, reduced social interactions, communication deficits, increased anxiety, motor stereotypies, repetitive behaviors, insistence on sameness, and restricted interests, identify autism. The Centers for Disease Control and Prevention estimates that ~1 in 88 children is diagnosed with autism each year, more than those diagnosed annually with cancer, diabetes, and AIDS combined. Currently, approximately $35 billion in direct and indirect care costs are spent annually for autism.113 The annual cost of caring for patients with autism is believed to reach $500 billion by 2025, with outside estimates reaching $1 trillion.114 Therefore, it is critical to gain mechanistic insights into multiple pathways coexisting in autism that will lay the foundation for new therapeutic opportunities.
Close associations between the serotonin and kynurenic pathways occur in tryptophan metabolism that are strongly influenced by the activity of the immune system. Quinolinic acid, a downstream product of the kynurenic pathway, is the structural precursor of NAD+, a critical energy carrier in mitochondria. Therefore, decreased tryptophan metabolism via either serotonin or QUIN will alter brain development, neuroimmune activity, and mitochondrial function. Interestingly, decreased tryptophan metabolism affecting both serotonin and kynurenic pathways has been linked to autism. Even a short-term reduction in the levels of tryptophan exacerbates repetitive, compulsive behaviors (eg, flapping, rocking, and whirling) in drug-free adults with autism.115 Serotonin in the blood is stored in platelets that undergo endocytosis via the serotonin transporter. Abnormal blood levels of serotonin produce autistic-like behaviors, such as impaired social communication and avoidance of new situations.116 Children with autism show a diminished capacity of whole-brain or regional brain serotonin synthesis that affects language production and sensory integration.117,118 There are reports of reductions in not only serotonin concentrations but also the kynurenic acid levels in the cerebrospinal fluid of autistic children.119,120 Diminished levels of NAD+ and plasma tryptophan in parallel with increased levels of oxidative stress have been reported in children with autism.121,122 In addition, urinary levels of tryptophan are reduced in patients with autism.123
Taken together, these results illustrate the critical need for a better understanding of the detailed mechanistic actions of tryptophan and NAD+ from cellular, molecular, and immunologic perspectives at distinct developmental time points in the CNS, which will aid in designing novel treatment strategies and bring new hope for neuroimmunological diseases such as autism. In addition, changes in tryptophan metabolism and pathway intermediates may serve in the development of new diagnostic assays for CNS immunologic disorders.
Conclusions
Increasing evidence indicates the important role of tryptophan catabolism in immune responses via the kynurenine pathway in vivo.51,124,125 However, the exact mechanisms remain unknown. Based on the findings shown here, NAD+ might be responsible for the newly found functions of tryptophan in immune responses. In particular, de novo synthesis of NAD+ through QUIN, one of the catabolites of the kynurenine pathway, appears to have an important role in immune regulation and tolerance (Figure 1). However, tryptophan catabolites appear to have contradictory functions, depending on where they are involved and warrant further investigation. Furthermore, mast cells, although they are not typical APCs, might play an important role in the tryptophan/QUIN/NAD+ pathway regulating T-cell activation. Clearly, the field of immunology shaped by tryptophan and NAD+ is poised for several exciting discoveries in the future.
Figure 1.
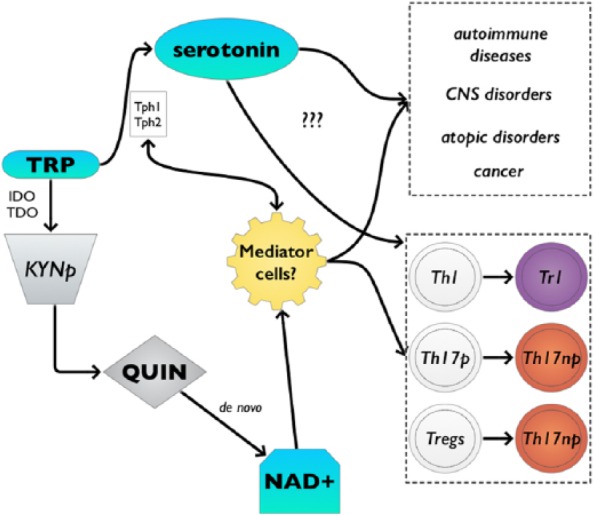
Newfound functions of tryptophan in immune responses might be influenced by NAD+. De novo synthesis of NAD+ through QUIN appears to have an important role in immune regulation and tolerance. CNS indicates central nervous system; IDO, indoleamine 2,3-dyoxygenase; KYNp, kynurenine pathway; NAD+, nicotinamide adenine dinucleotide; QUIN, quinolinic acid; TDO, tryptophan 2,3-dyoxygenase; Tph1, tryptophan hydroxylase 1; Tph2, tryptophan hydroxylase 2; Tregs, regulatory T-cells; TRP, tryptophan.
Footnotes
FUNDING: The author(s) disclosed receipt of the following financial support for the research, authorship, and/or publication of this article: This project was funded by the “Queen Elizabeth Hospital Birmingham (QEHB) Charity.”
PEER REVIEW: Six peer reviewers contributed to the peer review report. Reviewers’ reports totaled 1714 words, excluding any confidential comments to the academic editor.
DECLARATION OF CONFLICTING INTERESTS: The author(s) declared no potential conflicts of interest with respect to the research, authorship, and/or publication of this article.
Author Contributions
Conceived the concepts: HRCB, AE. Wrote or contributed to the writing of the manuscript: HRCB, AV, AE. Made critical revisions: AV, AE. Reviewed and approved the final manuscript: HRCB, AE.
REFERENCES
- 1.Hopkins FG, Cole SW. A contribution to the chemistry of proteids: part I. A preliminary study of a hitherto undescribed product of tryptic digestion. J Physiol (Lond) 1901;27:418–428. doi: 10.1113/jphysiol.1901.sp000880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Richard DM, Dawes MA, Mathias CW, Acheson A, Hill-Kapturczak N, Dougherty DM. L-tryptophan: basic metabolic functions, behavioral research and therapeutic indications. Int J Tryptophan Res. 2009;2:45–60. doi: 10.4137/ijtr.s2129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Orhan F, Bhat M, Sandberg K, et al. Tryptophan metabolism along the kynurenine pathway downstream of toll-like receptor stimulation in peripheral monocytes. Scand J Immunol. 2016;84:262–271. doi: 10.1111/sji.12479. [DOI] [PubMed] [Google Scholar]
- 4.Bender DA. Biochemistry of tryptophan in health and disease. Mol Aspects Med. 1983;6:101–197. doi: 10.1016/0098-2997(83)90005-5. [DOI] [PubMed] [Google Scholar]
- 5.Blankfield A. A brief historic overview of clinical disorders associated with tryptophan: the relevance to chronic fatigue syndrome (CFS) and fibromyalgia (FM) Int J Tryptophan Res. 2012;5:27–32. doi: 10.4137/IJTR.S10085. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Lichters M, Brunnlieb C, Nave G. The influence of serotonin deficiency on choice deferral and the compromise effect. J Marketing Res. 2016;53:183–198. [Google Scholar]
- 7.Rose DP. Aspects of tryptophan metabolism in health and disease: a review. J Clin Pathol. 1972;25:17–25. doi: 10.1136/jcp.25.1.17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Gracia-Rubio I, Moscoso-Castro M, Pozo OJ, Marcos J, Nadal R, Valverde O. Maternal separation induces neuroinflammation and long-lasting emotional alterations in mice. Prog Neuropsychopharmacol Biol Psychiatry. 2016;65:104–117. doi: 10.1016/j.pnpbp.2015.09.003. [DOI] [PubMed] [Google Scholar]
- 9.Adams S, Braidy N, Bessesde A, et al. The kynurenine pathway in brain tumor pathogenesis. Cancer Res. 2012;72:5649–5657. doi: 10.1158/0008-5472.CAN-12-0549. [DOI] [PubMed] [Google Scholar]
- 10.Pilotte L, Larrieu P, Stroobant V, et al. Reversal of tumoral immune resistance by inhibition of tryptophan 2,3-dioxygenase. Proc Natl Acad Sci U S A. 2012;109:2497–2502. doi: 10.1073/pnas.1113873109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Puccetti P, Fallarino F, Italiano A, et al. Accumulation of an endogenous tryptophan-derived metabolite in colorectal and breast cancers. PLoS ONE. 2015;10:e0122046. doi: 10.1371/journal.pone.0122046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Popov A, Schultze JL. IDO-expressing regulatory dendritic cells in cancer and chronic infection. J Mol Med. 2008;86:145–160. doi: 10.1007/s00109-007-0262-6. [DOI] [PubMed] [Google Scholar]
- 13.Platten M, Wick W, Van den Eynde BJ. Tryptophan catabolism in cancer: beyond IDO and tryptophan depletion. Cancer Res. 2012;72:5435–5440. doi: 10.1158/0008-5472.CAN-12-0569. [DOI] [PubMed] [Google Scholar]
- 14.Lee GK, Park HJ, Macleod M, Chandler P, Munn DH, Mellor AL. Tryptophan deprivation sensitizes activated T-cells to apoptosis prior to cell division. Immunology. 2002;107:452–460. doi: 10.1046/j.1365-2567.2002.01526.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Kushwah R, Hu J. Role of dendritic cells in the induction of regulatory T-cells. Cell Biosci. 2010;1:20. doi: 10.1186/2045-3701-1-20. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Maldonado RA, von Andrian UH. How tolerogenic dendritic cells induce regulatory T-cells. Adv Immunol. 2009;108:111–165. doi: 10.1016/B978-0-12-380995-7.00004-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Moffett JR, Namboodiri MA. Tryptophan and the immune response. Immunol Cell Biol. 2003;81:247–265. doi: 10.1046/j.1440-1711.2003.t01-1-01177.x. [DOI] [PubMed] [Google Scholar]
- 18.Wang Q, Liu D, Song P, Zou MH. Tryptophan-kynurenine pathway is dysregulated in inflammation, and immune activation. Front Biosci. 2015;20:1116–1143. doi: 10.2741/4363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Pertovaara M, Raitala A, Uusitalo H, et al. Mechanisms dependent on tryptophan catabolism regulate immune responses in primary Sjogren’s syndrome. Clin Exp Immunol. 2005;142:155–161. doi: 10.1111/j.1365-2249.2005.02889.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.van Baren N, Van den Eynde BJ. Tumoral immune resistance mediated by enzymes that degrade tryptophan. Cancer Immunol Res. 2015;3:978–985. doi: 10.1158/2326-6066.CIR-15-0095. [DOI] [PubMed] [Google Scholar]
- 21.Schröcksnadel K, Wirleitner B, Winkler C, Fuchs D. Monitoring tryptophan metabolism in chronic immune activation. Clin Chim Acta. 2006;364:82–90. doi: 10.1016/j.cca.2005.06.013. [DOI] [PubMed] [Google Scholar]
- 22.Jones SP, Guillemin G, Brew B. The kynurenine pathway in stem cell biology. Int J Tryptophan Res. 2013;6:57–66. doi: 10.4137/IJTR.S12626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Michelhaugh SK, Varadarajan K, Guastella AR, Mittal S. Role of kynurenine pathway in neuro-oncology. In: Mittal S, editor. Targeting the Broadly Pathogenic Kynurenine Pathway. Cham: Springer International Publishing; 2015. pp. 287–295. [Google Scholar]
- 24.Grant RS, Passey R, Matanovic G, Smythe G, Kapoor V. Evidence for increased de novo synthesis of NAD in immune-activated RAW264.7 macrophages: a self-protective mechanism? Arch Biochem Biophys. 1999;372:1–7. doi: 10.1006/abbi.1999.1381. [DOI] [PubMed] [Google Scholar]
- 25.Berger SJ, Sudar DC, Berger NA. Metabolic consequences of DNA damage: DNA damage induces alterations in glucose metabolism by activation of poly (ADP-ribose) polymerase. Biochem Biophys Res Commun. 1986;134:227–232. doi: 10.1016/0006-291x(86)90551-6. [DOI] [PubMed] [Google Scholar]
- 26.Berger F, Ramírez-Hernández MH, Ziegler M. The new life of a centenarian: signalling functions of NAD(P) Trends Biochem Sci. 2004;29:111–118. doi: 10.1016/j.tibs.2004.01.007. [DOI] [PubMed] [Google Scholar]
- 27.Braidy N, Guillemin GJ, Grant R. Effects of kynurenine pathway inhibition on NAD+ metabolism and cell viability in human primary astrocytes and neurons. Int J Tryptophan Res. 2011;4:29–37. doi: 10.4137/IJTR.S7052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Guillemin GJ. Quinolinic acid, the inescapable neurotoxin. FEBS J. 2012;279:1356–1365. doi: 10.1111/j.1742-4658.2012.08485.x. [DOI] [PubMed] [Google Scholar]
- 29.Tullius SG, Cetina Biefer HR, Li S, et al. NAD+ protects against EAE by regulating CD4+ T-cell differentiation. Nat Commun. 2014;5:5101. doi: 10.1038/ncomms6101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Elkhal A, Cetina Biefer HR, Heinbokel T, et al. NAD+ regulates Treg cell fate and promotes allograft survival via a systemic IL-10 production that is CD4+ CD25+ Foxp3+ T-cells independent. Sci Rep. 2016;6:22325. doi: 10.1038/srep22325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Wang J, Zhao C, Kong P, et al. Treatment with NAD+ inhibited experimental autoimmune encephalomyelitis by activating AMPK/SIRT1 signaling pathway and modulating Th1/Th17 immune responses in mice. Int Immunopharmacol. 2016;39:287–294. doi: 10.1016/j.intimp.2016.07.036. [DOI] [PubMed] [Google Scholar]
- 32.Kawasaki H, Chang HW, Tseng HC, et al. A tryptophan metabolite, kynurenine, promotes mast cell activation through aryl hydrocarbon receptor. Allergy. 2014;69:445–452. doi: 10.1111/all.12346. [DOI] [PubMed] [Google Scholar]
- 33.Xie FT, Cao JS, Zhao J, Yu Y, Qi F, Dai XC. IDO expressing dendritic cells suppress allograft rejection of small bowel transplantation in mice by expansion of Foxp3+ regulatory T-cells. Transpl Immunol. 2015;33:69–77. doi: 10.1016/j.trim.2015.05.003. [DOI] [PubMed] [Google Scholar]
- 34.Uyttenhove C, Pilotte L, Théate I, et al. Evidence for a tumoral immune resistance mechanism based on tryptophan degradation by indoleamine 2,3-dioxygenase. Nat Med. 2003;9:1269–1274. doi: 10.1038/nm934. [DOI] [PubMed] [Google Scholar]
- 35.Yan Y, Zhang G-X, Gran B, et al. IDO upregulates regulatory T-cells via tryptophan catabolite and suppresses encephalitogenic T-cell responses in experimental autoimmune encephalomyelitis. J Immunol. 2010;185:5953–5961. doi: 10.4049/jimmunol.1001628. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Stockinger B, Meglio PD, Gialitakis M, Duarte JH. The aryl hydrocarbon receptor: multitasking in the immune system. Annu Rev Immunol. 2014;32:403–432. doi: 10.1146/annurev-immunol-032713-120245. [DOI] [PubMed] [Google Scholar]
- 37.Quintana FJ, Sherr DH. Aryl hydrocarbon receptor control of adaptive immunity. Pharmacol Rev. 2013;65:1148–1161. doi: 10.1124/pr.113.007823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Quintana FJ. The aryl hydrocarbon receptor: a molecular pathway for the environmental control of the immune response. Immunology. 2013;138:183–189. doi: 10.1111/imm.12046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Apetoh L, Quintana FJ, Pot C, et al. The aryl hydrocarbon receptor interacts with c-Maf to promote the differentiation of type 1 regulatory T-cells induced by IL-27. Nat Immunol. 2010;11:854–861. doi: 10.1038/ni.1912. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Gandhi R, Kumar D, Burns EJ, et al. Activation of the aryl hydrocarbon receptor induces human type 1 regulatory T-cell-like and Foxp3+ regulatory T-cells. Nat Immunol. 2010;11:846–853. doi: 10.1038/ni.1915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Quintana FJ, Basso AS, Iglesias AH, et al. Control of T(reg) and T(H)17 cell differentiation by the aryl hydrocarbon receptor. Nature. 2008;453:65–71. doi: 10.1038/nature06880. [DOI] [PubMed] [Google Scholar]
- 42.Rodrigues CP, Ferreira ACF, Pinho MP, de Moraes CJ, Bergami-Santos PC, Barbuto JAM. Tolerogenic IDO(+) dendritic cells are induced by PD-1-expressing mast cells. Front Immunol. 2016;7:9. doi: 10.3389/fimmu.2016.00009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Mellor AL, Munn DH. IDO expression by dendritic cells: tolerance and tryptophan catabolism. Nat Rev Immunol. 2004;4:762–774. doi: 10.1038/nri1457. [DOI] [PubMed] [Google Scholar]
- 44.Engin AB. Evaluation of tryptophan metabolism in chronic immune activation. In: Engin A, Engin AB, editors. Tryptophan Metabolism: Implications for Biological Processes, Health and Disease (Molecular and Integrative Toxicology) Cham, Switzerland: Springer International Publishing; 2015. pp. 121–145. [Google Scholar]
- 45.Opitz CA, Litzenburger UM, Sahm F, et al. An endogenous tumour-promoting ligand of the human aryl hydrocarbon receptor. Nature. 2011;478:197–203. doi: 10.1038/nature10491. [DOI] [PubMed] [Google Scholar]
- 46.Li L, Huang L, Lemos HP, Mautino M, Mellor AL. Altered tryptophan metabolism as a paradigm for good and bad aspects of immune privilege in chronic inflammatory diseases. Front Immunol. 2012;3:109. doi: 10.3389/fimmu.2012.00109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Puccetti P. On the non-redundant roles of TDO2 and IDO1. Front Immunol. 2014;5:522. doi: 10.3389/fimmu.2014.00522. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Poulain-Godefroy O, Eury E, Leloire A, et al. Induction of TDO2 and IDO2 in liver by high-fat feeding in mice: discrepancies with human obesity. Int J Tryptophan Res. 2013;6:29–37. doi: 10.4137/IJTR.S11717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.D’Amato NC, Rogers TJ, Gordon MA, et al. A TDO2-AhR signaling axis facilitates anoikis resistance and metastasis in triple-negative breast cancer. Cancer Res. 2015;75:4651–4664. doi: 10.1158/0008-5472.CAN-15-2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Novikov O, Wang Z, Stanford EA, et al. An aryl hydrocarbon receptor-mediated amplification loop that enforces cell migration in ER-/PR-/Her2- human breast cancer cells. Mol Pharmacol. 2016;90:674–688. doi: 10.1124/mol.116.105361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Hsu Y-L, Hung J-Y, Chiang S-Y, et al. Lung cancer-derived galectin-1 contributes to cancer associated fibroblast-mediated cancer progression and immune suppression through TDO2/kynurenine axis. Oncotarget. 2016;7:27584–27598. doi: 10.18632/oncotarget.8488. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Crosignani S, Driessens G, Detheux M, Van den Eynde B, Cauwenberghs S. Preclinical assessment of a novel small molecule inhibitor of tryptophan 2,3-di-oxygenase 2 (TDO2) J Immunother Cancer. 2014;2:196. [Google Scholar]
- 53.Hwang SL, Chung NP-Y, Chan JK-Y, Lin CL. Indoleamine 2,3-dioxygenase (IDO) is essential for dendritic cell activation and chemotactic responsiveness to chemokines. Cell Res. 2005;15:167–175. doi: 10.1038/sj.cr.7290282. [DOI] [PubMed] [Google Scholar]
- 54.Miwa N, Hayakawa S, Miyazaki S, et al. IDO expression on decidual and peripheral blood dendritic cells and monocytes/macrophages after treatment with CTLA-4 or interferon-gamma increase in normal pregnancy but decrease in spontaneous abortion. Mol Hum Reprod. 2005;11:865–870. doi: 10.1093/molehr/gah246. [DOI] [PubMed] [Google Scholar]
- 55.Takikawa O. Biochemical and medical aspects of the indoleamine 2,3-dioxygenase-initiated L-tryptophan metabolism. Biochem Bioph Res Co. 2005;338:12–19. doi: 10.1016/j.bbrc.2005.09.032. [DOI] [PubMed] [Google Scholar]
- 56.Fujigaki S, Saito K, Takemura M, et al. Species differences in L-tryptophan-kynurenine pathway metabolism: quantification of anthranilic acid and its related enzymes. Arch Biochem Biophys. 1998;358:329–335. doi: 10.1006/abbi.1998.0861. [DOI] [PubMed] [Google Scholar]
- 57.Werner-Felmayer G, Werner ER, Fuchs D, Hausen A, Reibnegger G, Wachter H. Characteristics of interferon induced tryptophan metabolism in human cells in vitro. Biochim Biophys Acta. 1989;1012:140–147. doi: 10.1016/0167-4889(89)90087-6. [DOI] [PubMed] [Google Scholar]
- 58.Dai W, Gupta SL. Regulation of indoleamine 2,3-dioxygenase gene expression in human fibroblasts by interferon-gamma. Upstream control region discriminates between interferon-gamma and interferon-alpha. J Biol Chem. 1990;265:19871–19877. [PubMed] [Google Scholar]
- 59.Hill M, Tanguy Royer S, Royer P, et al. IDO expands human CD4+CD25high regulatory T-cells by promoting maturation of LPS-treated dendritic cells. Eur J Immunol. 2007;37:3054–3062. doi: 10.1002/eji.200636704. [DOI] [PubMed] [Google Scholar]
- 60.Sioud M, Nyakas M, Sæbøe-Larssen S, Mobergslien A, Aamdal S, Kvalheim G. Diversification of antitumour immunity in a patient with metastatic melanoma treated with ipilimumab and an IDO-silenced dendritic cell vaccine. Case Rep Med. 2016;2016:9639585. doi: 10.1155/2016/9639585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Zheng X, Koropatnick J, Chen D, et al. Silencing IDO in dendritic cells: a novel approach to enhance cancer immunotherapy in a murine breast cancer model. Int J Cancer. 2013;132:967–977. doi: 10.1002/ijc.27710. [DOI] [PubMed] [Google Scholar]
- 62.Yamahira A, Narita M, Iwabuchi M, et al. Activation of the leukemia plasmacytoid dendritic cell line PMDC05 by Toho-1, a novel IDO inhibitor. Anticancer Res. 2014;34:4021–4028. [PubMed] [Google Scholar]
- 63.Wang X-F, Wang H-S, Wang H, et al. The role of indoleamine 2,3-dioxygenase (IDO) in immune tolerance: focus on macrophage polarization of THP-1 cells. Cell Immunol. 2014;289:42–48. doi: 10.1016/j.cellimm.2014.02.005. [DOI] [PubMed] [Google Scholar]
- 64.Prendergast GC, Metz R, Muller AJ. IDO recruits Tregs in melanoma. Cell Cycle. 2009;8:1818–1819. doi: 10.4161/cc.8.12.8887. [DOI] [PubMed] [Google Scholar]
- 65.Wainwright DA, Dey M, Chang A, Lesniak MS. Targeting Tregs in malignant brain cancer: overcoming IDO. Front Immunol. 2013;4:116. doi: 10.3389/fimmu.2013.00116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Davies NW. Tryptophan, neurodegeneration and HIV-associated neurocognitive disorder. Int J Tryptophan Res. 2010;3:121–140. doi: 10.4137/ijtr.s4321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Abel J, Haarmann-Stemmann T. An introduction to the molecular basics of aryl hydrocarbon receptor biology. Biol Chem. 2010;391:1235–1248. doi: 10.1515/BC.2010.128. [DOI] [PubMed] [Google Scholar]
- 68.Mimura J, Yamashita K, Nakamura K, et al. Loss of teratogenic response to 2,3,7,8-tetrachlorodibenzo-p-dioxin (TCDD) in mice lacking the Ah (dioxin) receptor. Genes Cells. 1997;2:645–654. doi: 10.1046/j.1365-2443.1997.1490345.x. [DOI] [PubMed] [Google Scholar]
- 69.Chen X, Oppenheim JJ. Th17 cells and Tregs: unlikely allies. J Leukoc Biol. 2014;95:723–731. doi: 10.1189/jlb.1213633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Pan F, Fan H, Lu L, Liu Z, Jiang S. The yin and yang of signaling in Tregs and TH17 cells. Sci Signal. 2011;4:mr4. doi: 10.1126/scisignal.2001709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Kipnis E, Dessein R. Bacterial modulation of Tregs/Th17 in intestinal disease: a balancing act? Inflamm Bowel Dis. 2012;18:1389–1390. doi: 10.1002/ibd.21941. [DOI] [PubMed] [Google Scholar]
- 72.Swain SL, McKinstry KK, Strutt TM. Expanding roles for CD4+ T-cells in immunity to viruses. Nat Rev Immunol. 2012;12:136–148. doi: 10.1038/nri3152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Jiang S, Dong C. A complex issue on CD4(+) T-cell subsets. Immunol Rev. 2013;252:5–11. doi: 10.1111/imr.12041. [DOI] [PubMed] [Google Scholar]
- 74.Loser K, Mehling A, Loeser S, et al. Epidermal RANKL controls regulatory T-cell numbers via activation of dendritic cells. Nat Med. 2006;12:1372–1379. doi: 10.1038/nm1518. [DOI] [PubMed] [Google Scholar]
- 75.Fritsche E, Schäfer C, Calles C, et al. Lightening up the UV response by identification of the arylhydrocarbon receptor as a cytoplasmatic target for ultraviolet B radiation. Proc Natl Acad Sci USA. 2007;104:8851–8856. doi: 10.1073/pnas.0701764104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Park BV, Pan F. The role of nuclear receptors in regulation of Th17/Treg biology and its implications for diseases. Cell Mol Immunol. 2015;12:533–542. doi: 10.1038/cmi.2015.21. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Duarte JH, Di Meglio P, Hirota K, Ahlfors H, Stockinger B. Differential influences of the aryl hydrocarbon receptor on Th17 mediated responses in vitro and in vivo. PLoS ONE. 2013;8:e79819. doi: 10.1371/journal.pone.0079819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Voo KS, Wang Y-H, Santori FR, et al. Identification of IL-17-producing FOXP3+ regulatory T-cells in humans. Proc Natl Acad Sci USA. 2009;106:4793–4798. doi: 10.1073/pnas.0900408106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Komatsu N, Okamoto K, Sawa S, et al. Pathogenic conversion of Foxp3+ T-cells into TH17 cells in autoimmune arthritis. Nat Med. 2014;20:62–68. doi: 10.1038/nm.3432. [DOI] [PubMed] [Google Scholar]
- 80.Roncarolo MG, Gregori S, Battaglia M, Bacchetta R, Fleischhauer K, Levings MK. Interleukin-10-secreting type 1 regulatory T-cells in rodents and humans. Immunol Rev. 2006;212:28–50. doi: 10.1111/j.0105-2896.2006.00420.x. [DOI] [PubMed] [Google Scholar]
- 81.Pot C, Jin H, Awasthi A, et al. Cutting edge: IL-27 induces the transcription factor c-Maf, cytokine IL-21, and the costimulatory receptor ICOS that coordinately act together to promote differentiation of IL-10-producing Tr1 cells. J Immunol. 2009;183:797–801. doi: 10.4049/jimmunol.0901233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Schmitt V, Rink L, Uciechowski P. The Th17/Treg balance is disturbed during aging. Exp Gerontol. 2013;48:1379–1386. doi: 10.1016/j.exger.2013.09.003. [DOI] [PubMed] [Google Scholar]
- 83.Scheller J, Chalaris A, Schmidt-Arras D, Rose-John S. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim Biophys Acta. 2011;1813:878–888. doi: 10.1016/j.bbamcr.2011.01.034. [DOI] [PubMed] [Google Scholar]
- 84.Jordan SC, Choi J, Kim I, et al. Interleukin 6 (IL-6) a cytokine critical to mediation of inflammation, autoimmunity and allograft rejection: therapeutic implications of IL-6 receptor blockade. Transplantation. 2017;101:32–44. doi: 10.1097/TP.0000000000001452. [DOI] [PubMed] [Google Scholar]
- 85.Carpenter LL, Gawuga CE, Tyrka AR, Lee JK, Anderson GM, Price LH. Association between plasma IL-6 response to acute stress and early-life adversity in healthy adults. Neuropsychopharmacology. 2010;35:2617–2623. doi: 10.1038/npp.2010.159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Bessede A, Gargaro M, Pallotta MT, et al. Aryl hydrocarbon receptor control of a disease tolerance defence pathway. Nature. 2014;511:184–190. doi: 10.1038/nature13323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Cantó C, Menzies KJ, Auwerx J. NAD(+) metabolism and the control of energy homeostasis: a balancing act between mitochondria and the nucleus. Cell Metab. 2015;22:31–53. doi: 10.1016/j.cmet.2015.05.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Harden A, Young WJ. The alcoholic ferment of yeast-juice. P Roy Soc B: Biol Sci. 1906;77:405–420. [Google Scholar]
- 89.Essa MM, Subash S, Braidy N, et al. Role of NAD+, oxidative stress, and tryptophan metabolism in autism spectrum disorders. Int J Tryptophan Res. 2013:15–14. doi: 10.4137/IJTR.S11355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Ying W. NAD+ and NADH in ischemic brain injury. Front Biosci. 2008;13:1141–1151. doi: 10.2741/2751. [DOI] [PubMed] [Google Scholar]
- 91.Bruzzone S, Fruscione F, Morando S, et al. Catastrophic NAD+ depletion in activated T lymphocytes through Nampt inhibition reduces demyelination and disability in EAE. PLoS ONE. 2009;4:e7897. doi: 10.1371/journal.pone.0007897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Van Gool F, Galli M, Gueydan C, et al. Intracellular NAD levels regulate tumor necrosis factor protein synthesis in a sirtuin-dependent manner. Nat Med. 2009;15:206–210. doi: 10.1038/nm.1906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Braidy N, Grant R, Brew BJ, Adams S, Jayasena T, Guillemin GJ. Effects of kynurenine pathway metabolites on intracellular NAD synthesis and cell death in human primary astrocytes and neurons. Int J Tryptophan Res. 2009;2:61–69. doi: 10.4137/ijtr.s2318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Kurnasov O, Goral V, Colabroy K, et al. NAD biosynthesis: identification of the tryptophan to quinolinate pathway in bacteria. Chem Biol. 2002;10:1195–1204. doi: 10.1016/j.chembiol.2003.11.011. [DOI] [PubMed] [Google Scholar]
- 95.Houtkooper RH, Cantó C, Wanders RJ, Auwerx J. The secret life of NAD+: an old metabolite controlling new metabolic signaling pathways. Endocr Rev. 2010;31:194–223. doi: 10.1210/er.2009-0026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Schwarcz R, Whetsell WO, Mangano RM. Quinolinic acid: an endogenous metabolite that produces axon-sparing lesions in rat brain. Science. 1983;219:316–318. doi: 10.1126/science.6849138. [DOI] [PubMed] [Google Scholar]
- 97.Sahm F, Oezen I, Opitz CA, et al. The endogenous tryptophan metabolite and NAD+ precursor quinolinic acid confers resistance of gliomas to oxidative Stress. Cancer Res. 2013;73:3225–3234. doi: 10.1158/0008-5472.CAN-12-3831. [DOI] [PubMed] [Google Scholar]
- 98.Adriouch S, Haag F, Boyer O, Seman M, Koch-Nolte F. Extracellular NAD+: a danger signal hindering regulatory T-cells. Microbes Infect. 2012;14:1284–1292. doi: 10.1016/j.micinf.2012.05.011. [DOI] [PubMed] [Google Scholar]
- 99.Adriouch S, Hubert S, Pechberty S, Koch-Nolte F, Haag F, Seman M. NAD+ released during inflammation participates in T-cell homeostasis by inducing ART2-mediated death of naive T-cells in vivo. J Immunol. 2007;179:186–194. doi: 10.4049/jimmunol.179.1.186. [DOI] [PubMed] [Google Scholar]
- 100.Bandeira LG, Bortolot BS, Cecatto MJ, Monte-Alto-Costa A, Romana-Souza B. Exogenous tryptophan promotes cutaneous wound healing of chronically stressed mice through inhibition of TNF-α and IDO activation. PLoS ONE. 2015;10:e0128439. doi: 10.1371/journal.pone.0128439. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Swanson KA, Zheng Y, Heidler KM, Mizobuchi T, Wilkes DS. CDIIc+ cells modulate pulmonary immune responses by production of indoleamine 2,3-di-oxygenase. Am J Resp Cell Mol. 2004;30:311–318. doi: 10.1165/rcmb.2003-0268OC. [DOI] [PubMed] [Google Scholar]
- 102.Zinkernagel RM, Doherty PC. The discovery of MHC restriction. Immunol Today. 1997;18:14–17. doi: 10.1016/s0167-5699(97)80008-4. [DOI] [PubMed] [Google Scholar]
- 103.Haskins K, Kubo R, White J, Pigeon M, Kappler J, Marrack P. The major histocompatibility complex-restricted antigen receptor on T-cells. I. Isolation with a monoclonal antibody. J Exp Med. 1983;157:1149–1169. doi: 10.1084/jem.157.4.1149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Exley M, Terhorst C, Wileman T. Structure, assembly and intracellular transport of the T-cell receptor for antigen. Semin Immunol. 1991;3:283–297. [PubMed] [Google Scholar]
- 105.Lemos MP, Esquivel F, Scott P, Laufer TM. MHC class II expression restricted to CD8alpha+ and CD11b+ dendritic cells is sufficient for control of Leishmania major. J Exp Med. 2004;199:725–730. doi: 10.1084/jem.20030795. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Kambayashi T, Laufer TM. Atypical MHC class II-expressing antigen-presenting cells: can anything replace a dendritic cell? Nat Rev Immunol. 2014;14:719–730. doi: 10.1038/nri3754. [DOI] [PubMed] [Google Scholar]
- 107.Iwasaki A, Medzhitov R. Control of adaptive immunity by the innate immune system. Nat Immunol. 2015;16:343–353. doi: 10.1038/ni.3123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Suidan GL, Duerschmied D, Dillon GM, et al. Lack of tryptophan hydroxylase-1 in mice results in gait abnormalities. PLoS ONE. 2013;8:e59032. doi: 10.1371/journal.pone.0059032. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Kane MJ, Angoa-Peréz M, Briggs DI, et al. Mice genetically depleted of brain serotonin display social impairments, communication deficits and repetitive behaviors: possible relevance to autism. PLoS ONE. 2012;7:e48975. doi: 10.1371/journal.pone.0048975. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Abrahams BS, Geschwind DH. Advances in autism genetics: on the threshold of a new neurobiology. Nat Rev Genet. 2008;9:341–355. doi: 10.1038/nrg2346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Silverman JL, Yang M, Lord C, Crawley JN. Behavioural phenotyping assays for mouse models of autism. Nat Rev Neurosci. 2010;11:490–502. doi: 10.1038/nrn2851. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Pardo CA, Vargas DL, Zimmerman AW. Immunity, neuroglia and neuroin-flammation in autism. Int Rev Psychiatry. 2005;17:485–495. doi: 10.1080/02646830500381930. [DOI] [PubMed] [Google Scholar]
- 113.Ganz ML. The lifetime distribution of the incremental societal costs of autism. Arch Pediatr Adolesc Med. 2007;161:343–349. doi: 10.1001/archpedi.161.4.343. [DOI] [PubMed] [Google Scholar]
- 114.Tipton LA, Blacher J. Brief report: autism awareness: views from a campus community. J Autism Dev Disord. 2014;44:477–483. doi: 10.1007/s10803-013-1893-9. [DOI] [PubMed] [Google Scholar]
- 115.McDougle CJ, Naylor ST, Cohen DJ, Aghajanian GK, Heninger GR, Price LH. Effects of tryptophan depletion in drug-free adults with autistic disorder. Arch Gen Psychiatry. 1996;53:993–1000. doi: 10.1001/archpsyc.1996.01830110029004. [DOI] [PubMed] [Google Scholar]
- 116.Leboyer M, Philippe A, Bouvard M, et al. Whole blood serotonin and plasma beta-endorphin in autistic probands and their first-degree relatives. Biol Psychiatry. 1999;45:158–163. doi: 10.1016/s0006-3223(97)00532-5. [DOI] [PubMed] [Google Scholar]
- 117.Chugani DC, Muzik O, Rothermel R, et al. Altered serotonin synthesis in the dentatothalamocortical pathway in autistic boys. Ann Neurol. 1997;42:666–669. doi: 10.1002/ana.410420420. [DOI] [PubMed] [Google Scholar]
- 118.Chugani DC, Muzik O, Behen M, et al. Developmental changes in brain serotonin synthesis capacity in autistic and nonautistic children. Ann Neurol. 1999;45:287–295. doi: 10.1002/1531-8249(199903)45:3<287::aid-ana3>3.0.co;2-9. [DOI] [PubMed] [Google Scholar]
- 119.Adamsen D, Ramaekers V, Ho HT, et al. Autism spectrum disorder associated with low serotonin in CSF and mutations in the SLC29A4 plasma membrane monoamine transporter (PMAT) gene. Mol Autism. 2014;5:43. doi: 10.1186/2040-2392-5-43. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Zimmerman AW, Jyonouchi H, Comi AM, et al. Cerebrospinal fluid and serum markers of inflammation in autism. Pediatr Neurol. 2005;33:195–201. doi: 10.1016/j.pediatrneurol.2005.03.014. [DOI] [PubMed] [Google Scholar]
- 121.Adams JB, Audhya T, McDonough-Means S, et al. Nutritional and metabolic status of children with autism vs neurotypical children, and the association with autism severity. Nutr Metab (Lond) 2011;8:34. doi: 10.1186/1743-7075-8-34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Essa MM, Braidy N, Waly MI, et al. Impaired antioxidant status and reduced energy metabolism in autistic children. Res Autism Spect Dis. 2013;7:557–565. [Google Scholar]
- 123.Kałuzna-Czaplinska J, Michalska M, Rynkowski J. Determination of tryptophan in urine of autistic and healthy children by gas chromatography/mass spectrometry. Med Sci Monit. 2010;16:CR488–CR492. [PubMed] [Google Scholar]
- 124.Munn DH, Shafizadeh E, Attwood JT, Bondarev I, Pashine A, Mellor AL. Inhibition of T-cell proliferation by macrophage tryptophan catabolism. J Exp Med. 1999;189:1363–1372. doi: 10.1084/jem.189.9.1363. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.El-Zaatari M, Chang Y-M, Zhang M, et al. Tryptophan catabolism restricts IFN-γ-expressing neutrophils and Clostridium difficile immunopathology. J Immunol. 2014;193:807–816. doi: 10.4049/jimmunol.1302913. [DOI] [PMC free article] [PubMed] [Google Scholar]