

Abstract
Canine distemper virus (CDV) and rabies virus (RABV) occur worldwide in wild carnivore and domestic dog populations and pose threats to wildlife conservation and public health. In Etosha National Park (ENP), Namibia, anthrax is endemic and generates carcasses frequently fed on by an unusually dense population of black-backed jackals (Canis mesomelas). Using serology and phylogenetic analyses (on samples obtained from February, 2009 to July, 2010), and historical mortality records (1975–2011), we assessed jackal exposure to Bacillus anthracis (BA; the causal bacterial agent of anthrax), CDV, and RABV. Seroprevalence to all three pathogens was relatively high with 95% (n = 86), 73% (n = 86), and 9% (n = 81) of jackals exhibiting antibodies to BA, CDV, and RABV, respectively. Exposure to BA, as assessed with an anti-Protective Antigen ELISA test, increased significantly with age and all animals >1 yr old tested positive. Seroprevalence of exposure to CDV also increased significantly with age, with similar age-specific trends during both years of the study. No significant effect of age was found on RABV seroprevalence. Three of the seven animals exhibiting immunity to RABV were monitored for more than one year after sampling and did not succumb to the disease. Mortality records revealed that rabid animals are destroyed nearly every year inside the ENP tourist camps. Phylogenetic analyses demonstrated that jackal RABV in ENP is part of the same transmission cycle as other dog-jackal RABV cycles in Namibia.
Keywords: anthrax, Canis mesomelas, Bacillus anthracis, black-backed jackal, canine distemper virus, infectious disease, rabies virus, reservoir
INTRODUCTION
Canine distemper virus (CDV) and rabies virus (RABV) infect wild carnivore and domestic dog populations globally (Harder and Osterhaus, 1997; Rupprecht et al., 2002). As multihost pathogens, their dynamics are particularly complicated by the need to understand both intraspecific and interspecific transmission (Dobson, 2004). Moreover, because they are multihost pathogens both CDV and RABV threaten endangered species conservation (Laurenson et al., 1998; Goller et al., 2010) with the latter being a disease of major public health concern (Knobel et al., 2005). Effective management of these diseases requires our understanding of their dynamics in both domestic animals and wildlife. Some species of wildlife may play greater epidemiological roles than others and thus warrant extra attention. We conducted a serosurvey of CDV, RABV, and Bacillus anthracis (BA; the causal bacterial agent of anthrax) in one species with such potential, the black-backed jackal (Canis mesomelas), in Etosha National Park (ENP), Namibia.
The black-backed jackal (hereafter jackal) is a common canid species found across southern and eastern Africa (MacDonald et al., 2004). As opportunistic generalists, jackals are found in a variety of habitat types, but prefer open grassland (Loveridge and MacDonald, 2003). Relatively high density, widespread geographic range, and long dispersal distances make jackals an epidemiologically important species for a variety of diseases of public health, domestic animal, or conservation concern (Loveridge and MacDonald, 2001). Yet the role of jackal in the transmission of multispecies carnivore pathogens remains unclear, particularly with respect to whether they independently maintain particular pathogens as well as the degree to which they are responsible for transmission of disease to wildlife, livestock, and humans.
Using mathematical models Rhodes et al. (1998) argued that jackals (C. mesomelas and C. adustus) occur at densities too low to maintain RABV without repeated introduction of the pathogen from domestic dogs. Bingham (2005) responded that jackals are as capable as domestic dogs of maintaining RABV and that the above conclusion resulted from ignoring how spatial scale and metapopulation structure affect pathogen persistence. Recent molecular analyses demonstrated that jackal in northern South Africa are capable of maintaining RABV strains independently of domestic dogs (Zulu et al., 2009). Other evidence also highlights the important roles jackal play in RABV dynamics. For instance, in Zimbabwe jackals were the second greatest cause of RABV transmission to humans (Pfukenyi et al., 2007). In the central stock-ranching area of Namibia where settlements and towns are sparse, reported cases of rabies were greater in jackal than domestic dogs and the former appear to drive the epidemic dynamics (Courtin et al., 2000). Thus, in southern Africa the jackal is viewed as a maintenance host, implying that jackals maintain the virus by continuous but low-grade transmission of the virus. Given such evidence and the proposed elimination of canine rabies in Africa via vaccination of domestic dogs (Hampson et al., 2009), it is especially important to understand the conditions under which jackal populations can independently facilitate RABV persistence.
Jackals may also play an important role in CDV dynamics. The spread of CDV between domestic dog populations during an outbreak along the Namibian coast appears to have been due jackals (Gowtage-Sequeira et al., 2009). Craft et al. (2009) demonstrated that to explain the properties of the 1994 CDV epidemic that killed one third of the Serengeti lions, the virus must have been repeatedly introduced into the lion population from sympatric carnivores such as jackals and hyenas who may have been infected by domestic dogs (Craft et al., 2009). As jackals generally occur at greater densities than threatened carnivore species and are also found in human altered habitat, they may frequently provide an epidemiological link between threatened wildlife with domestic dogs.
While jackals may maintain epidemics of rabies at large spatial scales, the question remains under which conditions do they drive epidemic processes or facilitate long-term persistence of pathogens such as RABV and CDV. Host density is a key determinant of transmission dynamics, with host populations at low enough densities unable to maintain a pathogen (Bartlett, 1957). However, density is merely a proxy for host contact network structure (Keeling, 2005), with higher density populations corresponding to a greater number of average infectious contacts per individual. Yet host density, contact structures, and demography for a given species may vary greatly with habitat, resource availability and sympatric species. As jackal easily adapt to different environments and resource types (MacDonald et al., 2004), their transmission dynamics undoubtedly vary with density, movement, and social contact networks. We assessed jackal exposure to RABV and CDV in an unusually dense and mobile population of jackals that frequently scavenges on carrion produced by anthrax outbreaks in herbivores.
Anthrax, an environmentally-transmitted and highly fatal disease is seasonally endemic in the plains ungulates and elephants of ENP (Lindeque and Turnbull, 1994). Avian and mammalian scavenger species scavenge upon disease-generated carcasses. Jackals are the most frequently observed mammalian scavenger species at anthrax confirmed carcasses (Bellan, unpublished data). While all mammals are susceptible to anthrax, carnivores are generally less so than herbivores and frequently scavenge on BA contaminated carrion without apparent morbidity or mortality. Still, the timescale on which animals develop and lose immunity to BA remains unknown. Depending on the duration of immunity, scavengers may be useful biosentinels for anthrax surveillance, particularly in wildlife systems where carcasses are rarely detected. Correlates of scavenger exposure to anthrax can be used to assess trends in anthrax incidence (Lembo et al., 2011). Anthrax may also affect jackals in other ways. For instance, the apparently high density of jackals in ENP may be due to the seasonal pulse in carrion resources generated by anthrax outbreaks (Getz, 2011). Large aggregations of jackals at carcasses may also facilitate transmission of RABV or CDV, especially when jackals travel long distances to carcasses.
Estimating incidence of infection, disease and mortality is difficult in wildlife because they are elusive and difficult to sample. When sampling blood from animals is feasible, the presence of antibodies to a pathogen can help yield insight about its epidemiological dynamics. For instance, consistent detection of antibodies in all age groups over several years suggests that a pathogen may persist endemically in a population, though frequent reintroduction from adjacent populations or other species must be considered (Haydon et al., 2002). In contrast, in populations experiencing a recent epidemic, all animals born after the epidemic burn out should be seronegative (but only once older than the 12 wk duration of the colostral antibodies). We used such serological indicators of exposure to assess jackal exposure to RABV, CDV, and BA. To further illuminate the dynamics of these pathogens, we compared serological results with mortality data and conducted a phylogenetic analysis of RABV isolates.
MATERIALS AND METHODS
Study area
This study was undertaken in Etosha National Park, a 22,915 km2 fenced national park in northern Namibia between 18°30’S – 19°30’S and 14°15′ – 17°10′E. The vegetation is classified as arid savanna (Huntley, 1982) and exhibits a single wet and dry season each year with rain falling mainly between November and April (Engert, 1997). The park contains a 4,760 km2 salt pan, a dominant landscape feature remnant of a palaeolake (Hipondoka et al., 2006). Mopane (Colophospermum mopane) shrubveld or treeveld cover much of ENP, but extensive sweet grassveld (the Okaukuejo plains) lie around the Etosha pan (le Roux et al., 1988). Boreholes and artesian or contact springs supply the only perennial water (Auer, 1997).
This study was conducted on the Okaukuejo plains that feed migratory herds of plains ungulates (Burchell’s zebra, Equus quagga; springbok, Antidorcas marsupialis; gemsbok, Oryx gazella; blue wildebeest, Connochaetes taurinus). Jackal are opportunistic, monogamous, territorial carnivores whose young of both sexes often aid in rearing the next cohort of pups (their siblings) prior to dispersing (Moehlman, 1979). On the Okaukuejo plains, jackal are abundant and feed on insects, fruit, and human food waste (near tourist camps). They also hunt small mammals and ungulates, and scavenge on lion (Panthera leo) and spotted hyena (Crocuta crocuta) kills as well as on disease-generated carcasses. Anthrax is responsible for generating a large proportion of available carcasses consumed by jackals and other scavengers (Lindeque and Turnbull, 1994). Anthrax carcasses are generally found during the end of the wet season, in the preferred wet season habitat of plains ungulates (northwest of the Okaukuejo tourist camp: Leeubron study area); during the dry seasons, the ungulate herds spend more time southeast of Okaukuejo (Gemsbokvlakte study area), and anthrax carcasses are found less frequently in this location and at this time (Figure 1).
Figure 1.
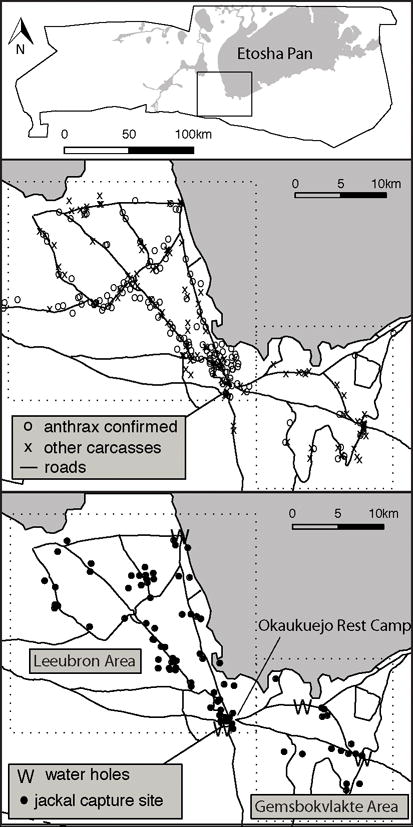
Top panel displays map of Etosha National Park, Namibia, boundaries with rectangle displaying the Okaukuejo plains where the research was conducted. Middle panel shows distribution of plains ungulate carcasses observed during opportunistic road-based surveillance over the duration of the study (January 2009 to July 2010). Bottom panel shows distribution of captured jackals. Dotted boxes show the division of the region into the Leeubron and Gemsbokvlatke study areas based on anthrax carcass distribution.
Jackal capture and sampling
Serum samples were obtained between January 2009 and July 2010 from 80 live-trapped jackals, 5 destroyed rabid jackals, and 1 jackal euthanized after a motor vehicle collision. All trapped jackals were captured within the two study areas defined in Figure 1 except for one animal captured 12km west of the Leeubron study area and one animal captured 115km east on the eastern edge of the park. All captured animals were unique except for CM02 (three captures) and CM26 (two captures). Twenty-two jackals were fitted with GPS VHF collars recording GPS locations hourly for 1–2 yr. All animals were captured and released safely. This animal handling protocol was approved under University of California, Berkeley ACUC R217-0509B and by the Namibian Ministry of Environment and Tourism Game Capture Veterinarian.
Captured jackals were aged and assigned to one of five age groups (<1, 1–2, 2–3, 3–4, >4 yr old) based on incisor wear (Lombaard, 1971). Ages of animals with missing teeth were considered unknown due to less predictable tooth wear. Jackals breed synchronously with pups first emerging from dens 8 wk after parturition (Bingham and Purchase, 2002). In ENP pups are first sighted during late November and early December each year. Thus, we estimated approximate age in months by assuming all jackals were born in October. Animals categorized in the >4 yr old age class were assumed to be 4–5 yr old when estimating age in months. Sample year was divided into jackal reproductive seasons with sample years hereafter referred to as 2008 and 2009 corresponding to October 2008 – September 2009 and October 2009 – September 2010, respectively. Sample sizes were too small to assess within year temporal patterns. Sera were stored at −20°C. Not all serum samples could be tested for all agents because of poor quality or insufficient quantity of serum.
Bacillus anthracis Anti-Protective Antigen Enzyme-Linked Immunosorbent Assay (ELISA)
The ELISA procedure used to measure anti-protective antigen (PA) antibody titers in was adapted from previous studies (Turnbull et al., 2008). Wildtype PA was provided by Bryan Krantz (University of California, Berkeley), at a concentration of 8.5mg PA/ml of phosphate buffered saline (PBS). Each well of a 96 well ELISA plate (Nalgene Nunc, USA) was coated with PA at a concentration of 0.375μl per well, covered to prevent drying, and incubated at 20°C for 1–48 hours. Plates were washed with PBS containing 10% Tween-20 (PBST), and then blocked for 30 minutes with 200μl per well of PBS, 0.5ml/l Tween-20, and 10% (w/v) skim milk powder (Oxoid Laboratory Preparations, United Kingdom) (PBSTM). After washing with PBST, serial two-fold dilutions to the ends of rows were made (with PBSTM) in duplicate for all samples, ranging from dilutions of 1:32 to 1:32,768. Plates were incubated at room temperature for an hour before washing with PBST. Commercially-available goat-anti-dog IgG-heavy and light chain horseradish peroxidase conjugate (Bethyl Laboratories, USA) was used as the secondary antibody, at the suggested dilution of 1:60,000. After further incubation and washing, TMB substrate was added (Kirkegaard & Perry Laboratories, USA) and the reaction was stopped after 30 minutes with 2N sulfuric acid. Well absorbance was read at 450nm on a SpectraMax M2 Microplate Reader using SoftMax Pro software v5.3 (Molecular Devices, USA).
We obtained 20 serum samples to use as negative controls from jackal in the Laikipia region of Kenya where anthrax is relatively uncommon (Prager, 2011). Negative controls were first analyzed individually using the above ELISA procedure. After it was determined that none of the samples had significant anti-PA titers, all negative control samples were then pooled equally into a single mixed negative control. Endpoint titers were defined as the last titer before the mean optical density (of duplicate serial dilutions) of a sample fell below the mean optical density (of duplicate serial dilutions) of the pooled negative control on the same plate.
Canine Distemper Virus Serum Neutralization Test
The test sera were diluted 1:5 in PBS+ and inactivated in a waterbath for 30min at 56°C prior to testing. Two-fold dilutions of the sera were made in duplicate (using MEM containing 5% foetal calf serum) in 96-well microtiter plates using a volume of 100μl. The stock virus (CDV Boekarest strain titre 103.85 TCID50/ml) was diluted in MEM containing 5% foetal calf serum to obtain 100TCID50/100 μl. One hundred microliters of the 100TCID50 antigen was added to all the wells containing the diluted test sera. A series of four, ten-fold dilutions was made from the 100TCID50 antigen, to be used as the virus control/back titration. The virus control was set up over three rows and six columns, and the rest of the reagents as follows: (1) 100μl MEM containing 5% foetal calf serum was added to all the wells; (2) 100μl of the 100TCID50 virus was added to the first two columns; (3) 100μl of the four dilutions (10−1 – 10−4) for back titration was added to the remaining four columns, starting with the highest virus dilution. The cell control was set up in duplicate rows, adding only 200μl MEM containing 5% foetal calf serum. The plates were then incubated for one hour at 37°C in a humid atmosphere of 5% CO2 in air. Vero cells were harvested, counted and adjusted to 480 000 cells per ml. 80μl of the cell suspension was added to all the wells. The plates were then incubated at 37°C in a humid atmosphere of 5% CO2 in air for 4–7 days, until the back titration (virus control) showed 50% CPE at the 10−2 dilution. A sample was deemed positive when there was a reduction of more than 50% of the cytopathic effect. Results were analyzed with cutoff titers of both ≥1:10 and ≥1:20. Two samples from ENP spotted hyena were made available by collaborators and also tested.
Rabies Fluorescent Antibody Virus Neutralization Test
Levels of neutralisation activity in the serum samples were assessed by the standard Fluorescent Antibody Virus Neutralisation test (FAVN) (Cliquet et al., 1998). Briefly, three-fold dilutions of test serum were incubated with a laboratory strain of rabies, Challenge Virus Standard (CVS-11, ATCC number VR959) at 100TCID50/ml (Anses, France), and incubated for 1 hr at 37°C in a humidified incubator at 5% CO2. Thereafter, 4×105 cells of 2–3 day culture of baby hamster kidney (BHK)-21 cells were added to each well and incubated for a further 48 hours. The plates were acetone-fixed and stained with an anti-rabies conjugate (produced by Onderstepoort Veterinary Institute, South Africa) and read under a fluorescence microscope to detect infected cells. Neutralisation antibody titres were calculated using the Spearman-Karber formula compared against an OIE dog reference serum (Anses, France).
Rabies virus phylogenetic analysis
Total viral RNA was extracted from approximately 100μg of original brain tissues from destroyed rabid animals using Tri Reagent according to the supplier’s instructions (Sigma, USA). The RNA pellet was solubilised in 50 μl of RNase-free water, quantified spectrophotometrically and stored at −70°C until required for RT-PCR. Approximately 1 μg of the total viral RNA was included in a reaction mixture that consisted of Lys001 (+) (20 pmoles), 200 units of Murine Moloney Leukemia Virus Reverse Transcriptase (M-MLV, USB™), 20 units of RNasin® ribonuclease inhibitor (Promega), 20 mM of dNTP mixture, 0.2M DTT and 5X M-MLV reaction buffers as described previously (Sabeta et al., 2007). The cDNA synthesis mixture was inactivated at 70°C for 10 minutes, diluted two-fold with sterile nuclease free water and stored at −20°C until further use.
Amplification of the partial N region of each of the virus isolates was achieved by using 40 pmoles each of lys (+) and 550B (reverse primer) in a previously described protocol (Sabeta et al., 2007). The amplified DNA products were visualized under UV transillumination and purified using spin columns. The amplicons were then cycle sequenced using the Big Dye Terminator chemistry (Applied Biosystems, USA). Nucleotide sequences were edited and a consensus sequence was obtained after alignment of the forward and reverse sequences. About 400 bp of the N region was obtained region. Multiple sequence alignment of the nucleotide sequences were performed using ClustalX version 1.82 package (Thompson et al., 1997).
Mortality Data
Since 1975 the Etosha Ecological Institute and visiting researchers have routinely recorded opportunistically observed mortalities. Samples for diagnosis of anthrax are taken by swabbing the nasal turbinate’s of ungulate carcasses (or other body parts when the skull is not available) with a sterile throat swab. Swabs are sent to the Central Veterinary Laboratory (Windhoek, Namibia) and the University of Hohenheim (Stuttgart, Germany) for diagnosis via selective culture and PCR, respectively (Turnbull, 2008). Brain samples from suspected rabid animals and fresh carnivore carcasses are routinely submitted to the Central Veterinary Laboratory for rabies diagnostics via the fluorescent antibody test or histopathology (Dean and Abelseth, 1973; Tierkel, 1973).
Statistical Analyses
Generalized linear models (McCulloch and Searle, 2001) were used to examine the effects of age, study area, and sample year on the three serological outcomes. All statistical analyses were conducted in R (R Development Core Team, 2010). For the RABV FAVN and CDV SNT tests, we fit binary outcomes (positive or negative) with logistic regression. All CDV SNT models were performed with seropositivity titer cutoffs of both ≥1:10 and ≥1:20. Because nearly all jackals were seropositive for the anti-PA ELISA, we modeled log endpoint titer as an ordered multinomial response variable using a proportional odds logistic regression model (Faraway, 2006) (using the R function polr in the MASS package). All independent variables were first fitted in a univariate model followed by a multivariate model including all three independent variables. Odds ratios and confidence intervals for univariate analyses between binary outcomes and binary explanatory variables were performed using Fisher’s exact test. All other confidence intervals are profile likelihood confidence intervals (using the R function confint).
RESULTS
B. anthracis Anti-Protective Antigen Elisa
The anti-PA ELISA curves for the 20 negative control samples were clustered at much lower optical densities than the ENP samples, suggesting that these animals were indeed unlikely to have been exposed to BA (Figure 2). The ELISA curves of 82/86 samples were above those of the negative controls for at least the first dilution plated. All 4 of the remaining animals were considered to be negative by our criteria and were < 1 yr (Figure 2). Age was the only significant explanatory variable in both univariate and multivariate analyses (Table 1) and titer increased with age in both seasons (Figure 2). Two animals were tested from outside the study area, one 12km west of the Okaukuejo area and one animal near the Namutoni tourist camp on the eastern edge of the park, approximately 115km north-east. Both had titers of >1:32768.
Figure 2.
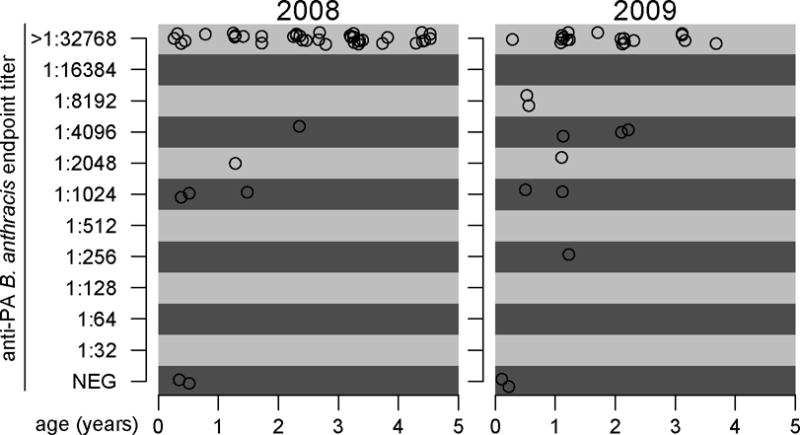
Endpoint titers in black-backed jackals (Canis mesomela) sampled in Etosha National Park, Namibia to the anti-protective antigen (anti-PA) Enzyme-Linked Immunosorbent Assay for exposure to Bacillus anthracis by age. Points have been jittered to facilitate display of number of samples when titers and ages are similar.
Table 1.
Distribution of number of laboratory-confirmed rabid carnivores destroyed per year in the Okaukuejo tourist camp between 1975–2006.
| confirmed cases | # years |
|---|---|
| 0 | 10 |
| 1 | 6 |
| 2 | 7 |
| 3 | 3 |
| 4 | 1 |
| 5 | 2 |
| 6 | 1 |
| 7 | 2 |
| 8 | 2 |
| 9 | 2 |
|
| |
| 45 | 36 |
Canine Distemper Virus Serum Neutralization Test
All results were qualitatively similar with CDV SNT cutoff titers of either ≥1:10 or ≥1:20. Of 86 animals tested, 73% or 65% were seropositive for the CDV SNT with cutoff titers of ≥1:10 or ≥1:20, respectively. Hereafter we consider titer cutoffs of ≥1:10 unless otherwise specified. Age was the only significant explanatory variable in both the univariate and multivariate analyses, with a steep climb in seroprevalence during the first year (Figure 3). Of recaptured animals, CM02 tested negative on the first two captures (January and July, 2009) but had seroconverted by the third capture (January 2010) and CM26 increased in titer from 1:40 (March, 2009) to >1:320 (June, 2010) between captures. Of the two spotted hyenas tested, one exhibited antibodies to CDV.
Figure 3.
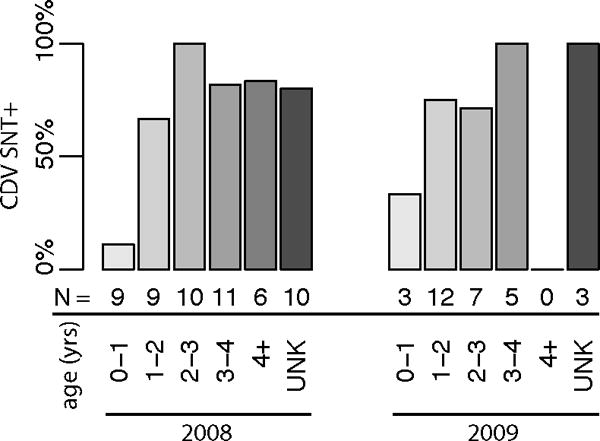
Seroprevalence of positivity to the canine distemper virus serum neutralization test for black-backed jackals (Canis mesomelas) by sample year and age class sampled in Etosha National Park, Namibia. Sample sizes for each category are given below each bar.
Rabies Mortalities
During the 2008 sample year, six jackals and one honey badger (Mellivora capensis) were suspected rabid and destroyed and one opportunistically found fresh lion carcass was also sampled; all eight samples were laboratory-confirmed positive for RABV. During the 2009 sample year four jackals were destroyed and laboratory-confirmed positive for RABV. Historical records of laboratory-confirmed rabies demonstrate consistent circulation of RABV over the past several decades (Table 2).
Table 2.
Regression coefficients and their confidence intervals from univariate and multivariate models of how age, sample year, and study area affect Bacillus anthracis anti-protective antigen enzyme-linked immunosorbent assay (anti-PA ELISA) endpoint titers (proportional odds logistic regression), canine distemper virus serum neutralization test (CDV SNT) and rabies fluorescent antibody virus neutralization test (RABV FAVN).
| response | parameter | odds ratio (95% confidence interval) | |||
|---|---|---|---|---|---|
| univariate model | multivariate model | ||||
| B. anthracis anti-PA | age | 4.7* | (2.2, 12) | 4.7* | (2.2, 12) |
| sample yeara | 0.34* | (0.11, 0.96) | 0.51 | (0.13, 1.8) | |
| study areab | 0.54 | (0.079, 2.2) | 1.9 | (0.20, 15) | |
|
| |||||
| CDV SNT | Age | 2.6* | (1.5, 4.8) | 2.8* | (1.6, 5.4) |
| sample year | 1.4 | (0.46, 4.6) | 1.8 | (0.53, 6.6) | |
| study area | 2.2 | (0.60, 7.6) | 2.3 | (0.45, 12) | |
|
| |||||
| RABV FAVN | Age | 1.6 | (0.77, 3.8) | 1.4c | (0.65, 3.4)c |
| sample year | 0 | (0,1.1) | c | c | |
| study area | 1.6 | (0.17, 77) | 5.8×107c | (0, Inf)c | |
Odds ratio is significantly different from 1 (p < .05).
Sample year odds ratios are OR2009/ OR2008.
Study area odds are ORLeeubron/ ORGemsbokvlakte.
Multivariate models for RABV FAVN were only fitted to study area and age for 2008 sample year data because all samples were negative for this assay in 2009.
Rabies Fluorescent Antibody Virus Neutralization Test
None of the explanatory variables exhibited a significant effect on seropositivity to RABV. However, only 7/81 (9%) samples from live-captured animals were positive, limiting statistical power. Three of 7 seropositive live-captured jackals were fitted with GPS collars and known to have remained healthy for at least a year after testing. Neither of the two recaptured jackals seroconverted between captures. One of two tested serum samples from laboratory-confirmed rabid animals was seropositive.
Rabies Virus Phylogenetic Analysis
Amplicons of the expected size were obtained from each virus isolate included in the study. The virus isolates (n = 4) were closely related to previously characterized viruses from dogs, jackals, and greater kudu (Tragelaphus strepsiceros) from the same geographical region, demonstrating that these viruses belong to the same dog-jackal transmission cycle.
DISCUSSION
Anthrax
Laikipia jackals and four juvenile ENP jackals exhibited ELISA optical density curves quite distinct from the rest of the ENP jackals. While we do not know the exposure history of Laikipia jackals, these results strongly suggest that both they and the four juvenile jackals in ENP had not been exposed to BA. The estimated ages of the four seronegative juveniles were 2, 3, 4 and 6 mo, though our aging method is very rough at the monthly resolution. The youngest seropositive animal (>1:32768) was estimated to be 3 mo old. Colostral antibodies are assumed to be absent by 12 wk. Thus, while it remains unclear whether jackal pups acquire colostral antibodies against BA, colostral antibodies clearly disappear before juveniles develop their own antibodies. As such naïve juvenile jackals must survive an initial challenge. Seropositivity cannot distinguish between a cleared infection or just an immune response to PA toxin consumed in the carcass (Turnbull et al., 2008). However, viable BA spores have been found in ENP scavenger feces, indicating they do ingest the infectious life phase of the bacterium (Lindeque and Turnbull, 1994). The rapid increase in titers within the first year of life suggests that juvenile jackals readily consume BA-laden carrion early in life and that continual consumption of anthrax carrion acts as a booster vaccine and maintains high levels of immunity to BA. Jackals are small and so their carcasses are difficult to sight. Thus, if juveniles occasionally died from anthrax before developing immunity we would be unlikely to observe it. While herbivores are generally more susceptible to anthrax than carnivores, anthrax confirmed carnivore deaths have historically been recorded in ENP (since 1975: 1 jackal, 3 lion, 9 cheetah) and elsewhere (Hugh-Jones and de Vos, 2002; Clegg et al., 2007).
Generally high titers to BA in ENP jackals were not surprising given the frequency of anthrax carcasses in the region sampled (Figure 1). That seroprevalence did not differ between study areas can be explained by ENP jackal mobility. Movement data from GPS collared jackals indicated that animals residing over territories sized 6–10km2 frequently traveled tens of kilometers to carcasses (Bellan, unpublished data). Thus, the spatial scale of this study was too small to adequately compare jackals affected by anthrax-generated carrion with those who were not. The absence of immunity in the Laikipia jackals, however, suggests that anthrax has been absent from that region in recent history. Jackal serology may thus be useful as an indicator for anthrax presence at wider spatial scales, though data concerning the duration of immunity would help to clarify how recent an exposure event could have occurred.
Canine Distemper Virus
CDV seroprevalence in Etosha jackals was high in both years and study areas. Several pieces of information suggest that transmission was ongoing for at least two years: in each sample year a juvenile (< 1 yr) was seropositive for CDV; of the two recaptured animals, one seroconverted between the two samples years and the titer of the other increased between the two sample years. Seroprevalence significantly increased with age in both sample years. This pattern could be due to increased hazard of exposure with age, decreased case fatality rate with age, or simply from the accumulation of hazard with age. During the recent CDV epidemic on the Namibian coast, Gowtage-Sequeira et al. (2009) also found that seroprevalence increased with age in the Skeleton Coast Park jackal population. However, the inter-annual patterns they found contrast starkly with our own. Prior to the 2002 six month-long outbreak (as determined by observation of clinical signs an carcass surveillance) in that jackal population, no animals exhibited antibodies to CDV. During the year of the outbreak, seropositive animals occurred in all age classes but seroprevalence increased with age. The year after the outbreak, the juvenile age class was entirely seronegative and the older classes had lower seroprevalence levels than the outbreak year. This is clearly indicative of epidemic dynamics in an immunologically naïve population. Our contrasting results—relatively consistent age-seroprevalence trends along with seroconversion of both recaptured animals and seropositive juveniles in both years—are consistent with endemic transmission of CDV in Etosha, though without longer term data this remains cautious speculation. However, the only historical serological data available on CDV in Etosha is also consistent with CDV endemicity in jackals; CDV seroprevalence from 1991–1996 in ENP was found to be 53%, 24%, and 13% in 15 jackal, 25 spotted hyena, and 145 lion, respectively (Alexander et al., 2008).
No evidence of clinical CDV has been observed in any ENP carnivores with the exception of 6 jackals and 4 captive wild dogs (Lycaon pictus) suspected to have died of distemper in the 1980s, though no laboratory-confirmation was performed. Despite the frequency with which suspected rabid jackals are destroyed in the Okaukejo tourist camp and the similarity between the signs of clinical distemper and rabies, only 4 of 71 destroyed suspected rabid jackals have tested negative for RABV since 1975. Thus, we believe that CDV is endemic but not clinically apparent in ENP jackals. This contrasts with the morbidity and mortality observed during the recent CDV epidemic in the jackals along the Namibian coast (Gowtage-Sequeira et al., 2009) suggesting differences in either CDV strain or immunological status of the jackal populations. In general, however, CDV is rarely associated with clinical disease or population declines in southern African carnivores (Alexander et al., 2008). CDV may still pose a risk to sympatric carnivores such as lions and spotted hyenas as evidenced by the Serengeti lion and wild dog outbreaks, (Roelke-Parker et al., 1996; Goller et al., 2010), particularly when animals are immunocompromised due to other infections (Munson et al., 2008).
Rabies Virus
Rabies epidemics generally occur at multiyear periods due to the relatively low R0 and rate of susceptible replenishment in carnivores (Hampson et al., 2007). Nearly all rabies cases recorded in ENP are inside or near tourist camps and thus represent a small proportion of total cases. Nevertheless, rabid animals are destroyed in Okaukuejo nearly every year. The 9% seroprevalence we found amongst ENP jackals is amongst the highest seroprevalence recorded in African wildlife (Alexander et al., 1994; Cleaveland and Dye, 1995; Creel et al., 1997; Prager, 2011). All seven seropositive results were from the first sample year, potentially indicating that an epidemic burned through the population in that year, though similar numbers of rabid animals were also destroyed the following year. Given recent proposals for the necessity and feasibility of eliminating canine rabies in Africa (Hampson et al., 2009), the ability for wildlife populations to facilitate RABV persistence is an important question. We note that there are no domestic dogs in ENP and that there are no towns within 100km of Okaukuejo. The phylogenetic results indicated that the recent virus isolates are part of the same dog-jackal cycles as previously described (Mansfield et al., 2006).
RABV dynamics in ENP may have particular implications for the reintroduction of wild dogs into the park. Three failed attempts were made to reintroduce wild dogs to the park between 1978 and 1990, the last of which failed when several animals succumbed to RABV (Scheepers and Venzke, 1995). This, in addition to their susceptibility to anthrax (Clegg et al., 2007), suggests that future reintroductions will have to take into account the obstacles posed by disease.
Summary
Due to their widespread distribution and opportunistic scavenging, jackals may potentially be useful as biosentinels for anthrax. Without knowing the exposure history of jackals sampled in Laikipia, Kenya, the grouping of their results with juvenile jackals from ENP suggests that these animals had not been exposed to anthrax-generated carrion. In contrast, jackals sampled in the Okaukuejo area of ENP have all been exposed to anthrax carrion, without succumbing to fatal infection, within their first year of age. Increasing titers with age suggests that consumption of carrion throughout life continually boosts their immunity to BA. Increasing seroprevalence to CDV with age in both years, and relatively high levels of seropositivity in general suggest that this pathogen may be endemic in ENP jackals, though no clinical disease or mortality is evident. Rabies cases are found almost every year in central ENP and a relatively large proportion of live-captured jackals exhibited immunity to RABV. These results suggest that the ENP jackal population may be capable of facilitating the persistence of both CDV and RABV, though future sampling and mathematical modeling could clarify these transmission dynamics.
Supplementary Material
Acknowledgments
We thank the Namibian Ministry of Environment and Tourism for permission to do this research, the Directorate of Parks, Wildlife and Management for permission to work throughout Etosha, and the staff in the Directorate of Scientific Services at the Etosha Ecological Institute for logistic support and assistance. We give a special thanks to Zepee Havarua, Werner Kilian, Shayne Kötting, Wilferd Versfeld, Marthin Kasaona, Gabriel Shatumbu, Birgit Kötting, Ortwin Aschenborn, and Mark Jago for all their help keeping our research program running smoothly. We also thank the Central Veterinary Laboratory in Windhoek for conducting rabies and anthrax diagnostics and Wolfgang Beyer for molecular anthrax diagnostics. Finally, we thank Wendy Turner and Alan Hubbard for feedback on the manuscript. This research was supported by the Chang-Lin Tien Environmental Fellowship, Andrew and Mary Thompson Rocca Scholarships, the Edna and Yoshinori Tanada Fellowship to S.E.B., and a James S. McDonnell grant and NIH grant GM83863 to W.M.G.
LITERATURE CITED
- 1.R: A language and environment for statistical computing. R Foundation for Statistical Computing, R Development Core Team; Vienna, Austria: 2010. [Google Scholar]
- 2.ALEXANDER KA, KAT PW, WAYNE RK, FULLER TK. Serologic Survey of Selected Canine Pathogens among Free-Ranging Jackals in Kenya. Journal of Wildlife Diseases. 1994;30:486–491. doi: 10.7589/0090-3558-30.4.486. [DOI] [PubMed] [Google Scholar]
- 3.ALEXANDER KA, MCNUTT JW, BRIGGS MB, STANDERS PE, FUNSTON P, HEMSON G, KEET D, VAN VUUREN M. Multi-host pathogens and carnivore management in southern Africa. Comparative Immunology Microbiology and Infectious Diseases. 2008 doi: 10.1016/j.cimid.2008.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.AUER C. Chemical quality of water at waterholes in the Etosha National Park. Madoqua. 1997;20:121–128. [Google Scholar]
- 5.BARTLETT MS. Measles Periodicity and Community Size. Journal of the Royal Statistical Society. Series A (General) 1957;120:48–70. [Google Scholar]
- 6.BINGHAM J, PURCHASE GK. Reproduction in the jackals Canis adustus Sundevall, 1846, and Canis mesomelas Schreber, 1778 (Carnivora: Canidae), in Zimbabwe. African Zoology. 2002;37:21–26. [Google Scholar]
- 7.CLEAVELAND S, DYE C. Maintenance of a microparasite infecting several host species: Rabies in the Serengeti. Parasitology. 1995;111:S33–S47. doi: 10.1017/s0031182000075806. [DOI] [PubMed] [Google Scholar]
- 8.CLEGG SB, TURNBULL PC, FOGGIN CM, LINDEQUE PM. Massive outbreak of anthrax in wildlife in the Malilangwe Wildlife Reserve, Zimbabwe. Veterinary Record. 2007;160:113–118. doi: 10.1136/vr.160.4.113. [DOI] [PubMed] [Google Scholar]
- 9.CLIQUET F, AUBERT M, SAGNE L. Development of a fluorescent antibody virus neutralisation test (FAVN test) for the quantitation of rabies-neutralising antibody. Journal of Immunological Methods. 1998;212:79–87. doi: 10.1016/s0022-1759(97)00212-3. [DOI] [PubMed] [Google Scholar]
- 10.COURTIN F, CARPENTER TE, PASKIN RD, CHOMEL BB. Temporal patterns of domestic and wildlife rabies in central Namibia stock-ranching area, 1986–1996. Preventive Veterinary Medicine. 2000;43:13–28. doi: 10.1016/s0167-5877(99)00083-5. [DOI] [PubMed] [Google Scholar]
- 11.CRAFT ME, VOLZ E, PACKER C, MEYERS LA. Distinguishing epidemic waves from disease spillover in a wildlife population. Proceedings of the Royal Society B-Biological Sciences. 2009;276:1777–1785. doi: 10.1098/rspb.2008.1636. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.CREEL S, CREEL NM, MUNSON L, SANDERLIN D, APPEL MJ. Serosurvey for selected viral diseases and demography of African wild dogs in Tanzania. Journal of Wildlife Diseases. 1997;33:823–832. doi: 10.7589/0090-3558-33.4.823. [DOI] [PubMed] [Google Scholar]
- 13.DEAN DJ, ABELSETH MK. Laboratory techniques in rabies: the fluorescent antibody test. Monograph Series of the World Health Organization. 1973:73–84. [PubMed] [Google Scholar]
- 14.DOBSON A. Population Dynamics of Pathogens with Multiple Host Species. The American Naturalist. 2004;164:S64–S78. doi: 10.1086/424681. [DOI] [PubMed] [Google Scholar]
- 15.ENGERT S. Spatial variability and temporal periodicity of rainfall in the Etosha National Park and surrounding areas in northern Namibia. Madoqua. 1997;20:115–120. [Google Scholar]
- 16.FARAWAY JJ. Extending the linear model with R generalized linear, mixed effects and nonparametric regression models Texts in statistical science. Chapman & Hall/CRC; Boca Raton, FL: 2006. p. ix, 301. [Google Scholar]
- 17.GETZ WM. Biomass transformation webs provide a unified approach to consumer–resource modelling. Ecology Letters. 2011;14:113–124. doi: 10.1111/j.1461-0248.2010.01566.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.GOLLER KV, FYUMAGWA RD, NIKOLIN V, EAST ML, KILEWO M, SPECK S, M¸LLER T, MATZKE M, WIBBELT G. Fatal canine distemper infection in a pack of African wild dogs in the Serengeti ecosystem, Tanzania. Veterinary Microbiology. 2010;146:245–252. doi: 10.1016/j.vetmic.2010.05.018. [DOI] [PubMed] [Google Scholar]
- 19.GOWTAGE-SEQUEIRA S, BANYARD AC, BARRETT T, BUCZKOWSKI H, FUNK SM, CLEAVELAND S. Epidemiology, pathology, and genetic analysis of a canine distemper epidemic in Namibia. Journal of Wildlife Diseases. 2009;45:1008–1020. doi: 10.7589/0090-3558-45.4.1008. [DOI] [PubMed] [Google Scholar]
- 20.HAMPSON K, DUSHOFF J, BINGHAM J, BRUCKNER G, ALI YH, DOBSON A. Synchronous cycles of domestic dog rabies in sub-Saharan Africa and the impact of control efforts. Proceedings of the National Academy of Sciences of the United States of America. 2007;104:7717–7722. doi: 10.1073/pnas.0609122104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.HAMPSON K, DUSHOFF J, CLEAVELAND S, HAYDON DT, KAARE M, PACKER C, DOBSON A. Transmission dynamics and prospects for the elimination of canine rabies. Plos Biology. 2009;7:e53. doi: 10.1371/journal.pbio.1000053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.HARDER TC, OSTERHAUS ADME. Canine distemper virus - A morbillivirus in search of new hosts? Trends in Microbiology. 1997;5:120–124. doi: 10.1016/S0966-842X(97)01010-X. [DOI] [PubMed] [Google Scholar]
- 23.HAYDON DT, CLEAVELAND S, TAYLOR LH, LAURENSON MK. Identifying reservoirs of infection: A conceptual and practical challenge. Emerging Infectious Diseases. 2002;8:1468–1473. doi: 10.3201/eid0812.010317. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.HIPONDOKA MHT, JOUSSE H, KEMPF J, BUSCHE D. Fossil evidence for perennial lake conditions during the Holocene at Etosha Pan, Namibia. South African Journal of Science. 2006;102:93–95. [Google Scholar]
- 25.HUGH-JONES ME, DE VOS V. Anthrax and wildlife. Revue Scientifique Et Technique De L Office International Des Epizooties. 2002;21:359–383. doi: 10.20506/rst.21.2.1336. [DOI] [PubMed] [Google Scholar]
- 26.HUNTLEY BJ. Southern African savannas. In: WALKER BH, editor. Ecology of tropical savannas. Springer; Berlin, Germany: 1982. pp. 101–119. [Google Scholar]
- 27.KEELING M. The implications of network structure for epidemic dynamics. Theoretical Population Biology. 2005;67:1–8. doi: 10.1016/j.tpb.2004.08.002. [DOI] [PubMed] [Google Scholar]
- 28.KNOBEL DL, CLEAVELAND S, COLEMAN PG, FEVRE EM, MELTZER MI, MIRANDA MEG, SHAW A, ZINSSTAG J, MESLIN FX. Re-evaluating the burden of rabies in Africa and Asia. Bulletin of the World Health Organization. 2005;83:360–368. [PMC free article] [PubMed] [Google Scholar]
- 29.LAURENSON K, SILLERO-ZUBIRI C, THOMPSON H, SHIFERAW F, THIRGOOD S, MALCOLM J. Disease as a threat to endangered species: Ethiopian wolves, domestic dogs and canine pathogens. Animal Conservation. 1998;1:273–280. [Google Scholar]
- 30.LE ROUX CJG, GRUNOW JO, MORRIS JW, BREDENKAMP GJ, SCHEEPERS JC. A classification of the vegetation of the Etosha National Park. South AFrican Journal of Botany. 1988;54:1–10. [Google Scholar]
- 31.LEMBO T, HAMPSON K, AUTY H, BEESLEY CA, BESSELL P, PACKER C, HALLIDAY J, FYUMAGWA R, HOARE R, ERNEST E, MENTZEL C, MLENGEYA T, STAMEY K, WILKINS PP, CLEAVELAND S. Serologic Surveillance of Anthrax in the Serengeti Ecosystem, Tanzania, 1996–2009. Emerging Infectious Diseases. 2011;17:387–394. doi: 10.3201/eid1703.101290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.LINDEQUE PM, TURNBULL PCB. Ecology and epidemiology of anthrax in the Etosha National Park, Namibia. Onderstepoort Journal of Veterinary Research. 1994;61:71–83. [PubMed] [Google Scholar]
- 33.LOMBAARD LJ. Age determination and growth curves in the black-backed jackal, Canis mesomelas Schreber, 1775 (Carnivora: Canidae) Annals of the Transvaal Museum. 1971;27:135–169. [Google Scholar]
- 34.LOVERIDGE AJ, MACDONALD DW. Seasonality in spatial organization and dispersal of sympatric jackals (Canis mesomelas and C. adustus): implications for rabies management. Journal of Zoology, London. 2001;253:101–111. [Google Scholar]
- 35.LOVERIDGE AJ, MACDONALD DW. Niche separation in sympatric jackals (Canis mesomelas and Canis adustus) Journal of Zoology, London. 2003;259:143–153. [Google Scholar]
- 36.MACDONALD DW, LOVERIDGE AJ, ATKINSON ANDRPD. Jackals. In: MACDONALD DW, SILLERO-ZUBIRI ANDC, editors. The Biology and Conservation of Wild Canids. 2004. pp. 255–270. [Google Scholar]
- 37.MANSFIELD KL, MCELHINNEY LM, HUBSCHLE O, METTLER F, SABETA C, NEL LH, FOOKS AR. A molecular epidemiological study of rabies epizootics in kudu (Tragelaphus strepsiceros) in Namibia. BMC Veterinary Research. 2006;2:2. doi: 10.1186/1746-6148-2-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.MCCULLOCH CE, SEARLE SR. Generalized, linear, and mixed models Wiley series in probability and statistics Texts, references, and pocketbooks section. John Wiley & Sons; New York; Chichester: 2001. p. xxi, 325. [Google Scholar]
- 39.MOEHLMAN P. Jackal helpers and pup survival. Nature. 1979;277:382–383. [Google Scholar]
- 40.MUNSON L, TERIO KA, KOCK R, MLENGEYA T, ROELKE ME, DUBOVI E, SUMMERS B, SINCLAIR ARE, PACKER C. Climate Extremes Promote Fatal Co-Infections during Canine Distemper Epidemics in African Lions. PLoS ONE. 2008;3:e2545. doi: 10.1371/journal.pone.0002545. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.PFUKENYI DM, PAWANDIWA D, MAKAYA PV, USHEWOKUNZE-OBATOLU U. A retrospective study of rabies in humans in Zimbabwe, between 1992 and 2003. Acta Tropica. 2007;102:190–196. doi: 10.1016/j.actatropica.2007.04.013. [DOI] [PubMed] [Google Scholar]
- 42.PRAGER KC. An investigation of infectious disease dynamcs in African carnivores: identifying reservoirs and risk factors and investigating control strategies. University of California, Davis; Davis: 2011. [Google Scholar]
- 43.ROELKE-PARKER ME, MUNSON L, PACKER C, KOCK R, CLEAVELAND S, CARPENTER M, OBRIEN SJ, POSPISCHIL A, HOFMANN-LEHMANN R, LUTZ H, MWAMENGELE GLM, MGASA MN, MACHANGE GA, SUMMERS BA, APPEL MJG. A canine distemper virus epidemic in Serengeti lions (Panthera leo) Nature. 1996;379:441–445. doi: 10.1038/379441a0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.RUPPRECHT CE, HANLON CA, HEMACHUDHA T. Rabies re-examined. Lancet Infectious Diseases. 2002;2:327–343. doi: 10.1016/s1473-3099(02)00287-6. [DOI] [PubMed] [Google Scholar]
- 45.SABETA CT, MARKOTTER W, MOHALE DK, SHUMBA W, WANDELER AI, NEL LH. Mokola virus in domestic mammals, South Africa. Emerging Infectious Diseases. 2007;13:1371–1373. doi: 10.3201/eid1309.070466. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.SCHEEPERS JL, VENZKE KAE. Attempts to reintroduce African wild dogs Lycaon pictus into Etosha National Park, Namibia. South African Journal of Wildlife Research. 1995;25:138–140. [Google Scholar]
- 47.THOMPSON JD, GIBSON TJ, PLEWNIAK F, JEANMOUGIN F, HIGGINS DG. The CLUSTAL-X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research. 1997;25:4876–4882. doi: 10.1093/nar/25.24.4876. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.TIERKEL ES. (Monograph Series of the World Health Organization).Laboratory techniques in rabies: rapid microscopic examination for Negri bodies and preparation of specimens for biological test. 1973:41–55. [PubMed] [Google Scholar]
- 49.TURNBULL PCB. Guidelines for the Surveillance and Control of Anthrax in Humans and Animals. World Health Organization World Organisation for Animal Health, Food and Agriculture, United Nations; Geneva, Switzerland: 2008. [Google Scholar]
- 50.TURNBULL PCB, DIEKMANN M, KILIAN JW, VERSFELD W, DE VOS V, ARNTZEN L, WOLTER K, BARTELS P, KOTZE A. Naturally acquired antibodies to Bacillus anthracis protective antigen in vultures of southern Africa. Onderstepoort Journal of Veterinary Research. 2008;75:95–102. doi: 10.4102/ojvr.v75i2.6. [DOI] [PubMed] [Google Scholar]
- 51.ZULU GC, SABETA CT, NEL LH. Molecular epidemiology of rabies: Focus on domestic dogs (Canis familiaris) and black-backed jackals (Canis mesomelas) from northern South Africa. Virus Research. 2009;140:71–78. doi: 10.1016/j.virusres.2008.11.004. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.