

Abstract
Despite decades of basic research, biliary diseases remain prevalent, highly morbid, and notoriously difficult to treat. We have, however, dramatically increased our understanding of biliary developmental biology, cholangiocyte pathophysiology, and the endogenous mechanisms of biliary regeneration and repair. All of this complex and rapidly evolving knowledge coincides with an explosion of new technological advances in the area of regenerative medicine. New breakthroughs such as induced pluripotent stem cells and organoid culture are increasingly being applied to the biliary system and now it is only a matter of time until new regenerative therapeutics for the cholangiopathies are unveiled. In this review, we will integrate what is known about biliary development, regeneration, and repair and we will link these conceptual advances to the technological breakthroughs that are collectively driving the emergence of a new global field in biliary regenerative medicine.
Keywords: cholangiocytes, regenerative medicine, biliary tree, stem cells, organoids
Because of the unique regenerative ability of the liver, many decades of research into liver regeneration have given us unique insights into various modes of organ regeneration1 and therefore hepatology has historically been at the epicenter of the science of regeneration. While the underpinnings of modern regenerative medicine have been developing for much of that time, the worldwide pivot toward regenerative medicine, which now pervades all of modern medicine, can be most clearly landmarked by the Nobel Prize winning technology of induced pluripotent stem cells, reported a decade ago in 20062. While this technology was quickly adapted to the study of hepatocyte differentiation3–12, hepatocellular disease modeling13–16, and cell-based therapy17–19, there has been a subsequent expansion of similar advances in the realm of biliary disorders20–23. As we will see, the biliary tree has always been center stage in the quest to understand the regenerative capacities and limitations of the liver since it has a distinct developmental origin, is anatomically and functionally heterogeneous, harbors several putative stem cell niches, and is the target for a vast array of liver pathologies. Furthermore, in many ways, the biliary tree may be an even more attractive target for regenerative therapeutics than parenchymal hepatocytes, given its broad distribution throughout the liver and its endoscopic accessibility by endoscopic retrograde cholangiopancreatography (ERCP). Recently, several excellent recent review articles have been published on individual aspects that are relevant to the topic of biliary regenerative medicine24–27. However, a more comprehensive perspective has been lacking and may be useful to help further define this emerging field. In this review: we will revisit what is known about biliary development, regeneration, and repair; we will summarize current concepts related to biliary stem cells and cellular plasticity in the liver; we will review the current state of advances in biliary regenerative medicine; and we will provide a vision of where this field is most ideally poised for advances in basic discoveries and clinical applications in the future.
Cholangiocytes and Cholangiopathies
Liver, which is responsible for bile acid production, serum detoxification, synthesis of serum proteins, immune regulation, and metabolic activities, is composed of two types of epithelial cells: hepatocytes and cholangiocytes. While a majority of the essential functions are performed by hepatocytes, which make up 95% of the liver parenchyma, specialized cholangiocytes form the biliary tree. The biliary tree is composed of intrahepatic and extrahepatic bile ducts, lined by the mature epithelial cholangiocytes28. They facilitate secretion and modification of biliary constituents and serve as a conduit for bile transport to the intestine. These cells are now known to be the target of a diverse group of biliary disorders, known as cholangiopathies, many of which can lead to progressive peri-portal fibrosis, portal hypertension, biliary cirrhosis, and cholangiocarcinoma29. The cholangiopathies can be proliferative (e.g. polycystic liver disease) or fibro-obliterative (e.g. primary sclerosing cholangitis) in character and have heterogeneous etiopathogenesis (e.g. genetic, toxic, immune-mediated, vascular, etc.)30. A brief outline of five major cholangiopathies will demonstrate the breadth of clinical issues facing these patients.
Primary sclerosing cholangitis (PSC) is an idiopathic, fibro-obliterative cholangiopathy characterized by diffuse inflammation of intrahepatic and/or extrahepatic bile ducts31. This chronic process can progress to end-stage biliary cirrhosis with portal hypertension and hepatic failure. Ursodeoxycholic acid (UDCA) has been used for the treatment of PSC and, when administrated in low doses, shows improvements in serum liver biochemistries. Vancomycin has also been used, particularly in children with PSC.
Primary biliary cholangitis (PBC) is a complex, autoimmune-mediated cholangiopathy, characterized by progressive destruction of the intrahepatic bile ducts, leading to cholestasis and portal inflammation, which, when chronic, can progress to periportal fibrosis and cirrhosis32. Prognosis for PBC has been improved by the use of UDCA33, and recently, obeticholic acid also became an FDA approved treatment for PBC.
Autosomal-dominant polycystic kidney disease (ADPKD) is the most common inherited kidney disease, caused by mutations in the PKD1 and PKD2 genes34. In many cases, ADPKD is also associated with polycystic liver disease (PLD), a proliferative cholangiopathy associated with ciliary dysfunction in which multiple cysts develop within the liver parenchyma as a result of alterations in calcium homeostasis and cyclic adenosine monophosphate activity and subsequent effects on protein kinase mediated proliferation.
Biliary atresia (BA) is a rare childhood disease that affects the function and the anatomy along the canalicularbile duct continuum35. The obliteration or discontinuity of the extrahepatic biliary system, results in obstruction of bile flow, leading to cholestatic jaundice. BA is treated with the Kasai hepatoportoenterostomy and/or liver transplantation and the condition remains the most common childhood indication for liver transplantation.
Cholangiocarcinoma (CCA) is a group of rare, but devastating, hepatobiliary cancers that arise from the intrahepatic, perihilar, or distal biliary tree36. Chronic inflammation from liver fluke infestation, hepatitis B and C infections, and PSC are the main risk factors for CCA. Other etiologic factors include inflammatory bowel disease, hepatolithiasis, cirrhosis, alcohol, smoking, and fatty liver disease. A highly selected subgroup of patients with hilar cholangiocarcinoma can benefit from neoadjuvent chemoradiation followed by liver transplantation.
Overall, treatment of the cholangiopathies consists of pharmacotherapies (e.g. ursodeoxycholic acid, vancomycin, etc.) that are largely ineffective and surgical therapies (e.g. Kasai procedure, liver transplantation, etc.) that are limited in scope and availability37. As such, many times cholangiopathies remain essentially untreatable with high morbidity and mortality in both children and adults. Even after successful liver transplantation, patients can be stricken with devastating biliary complications such as ischemic cholangiopathy (in the case of donation after cardiac death or hepatic artery thrombosis). Living donor transplant (effectively a regenerative medicine therapy) has significantly expanded the pool of donor organs, but unfortunately, biliary complications such as strictures and leaks are notoriously prevalent following this procedure, effecting up to 20% of living donor recipients.
Biliary Regenerative Medicine and the R3 Paradigm
The unfortunate lack of effective therapies for biliary disease has prompted the aggressive evaluation of new therapeutic options in the realm of biliary regenerative medicine that have the potential to radically alter our management of these patients. An expanding understanding of biliary pathophysiology, the presence of endogenous stem cell niches within the biliary tree, and endoscopic access to the system make cholangiopathies attractive targets for regenerative medicine therapies. At the most simplistic level, biliary regenerative medicine is tasked with creating new cholangiocytes and building new bile ducts. While there is no definitive blueprint for the development of regenerative medicine therapies, one useful concept is the R3 paradigm, which incorporates three distinct regenerative tactics: replacement, regeneration, and rejuvenation38. Replacement strategy refers to transplantation of a cell-based product that reestablishes homeostasis (liver transplantation is an example of this approach)39,40. Regenerative strategy refers to engraftment of progenitor cells that require in vivo growth and differentiation (stem cell transplant is an example)17,41,42. Rejuvenation strategy refers to the induction of self-renewal of tissues by activation of endogenous stem cells43–45. In the context of biliary disease, replacement would include therapies designed to directly replace the damaged biliary epithelium (e.g. cholangiocyte-based cell therapies, bio-engineered tissue patches, etc.). Regeneration, in contrast, would encompass stem cell-based therapies (biodegradable stem cell-coated stents, for example). Lastly, rejuvenation therapies would be designed to activate a therapeutic subset of the endogenous biliary stem/progenitor cell systems (gene therapy, therapeutic exosome delivery, etc.).
Biliary Development
In order to properly envisage new regenerative therapeutics for biliary disorders, it is useful, if not mandatory, to understand, as clearly as possible, the normal embryological development of the biliary tree. Liver is formed from the ventral foregut endoderm, which also gives rise to the lung, ventral pancreas, and thyroid46. Transcription of liver specific genes, such as albumin, can be detected in the ventral foregut endoderm as early as embryonic day 8.5 (E8.5), specifying hepatic differentiation47. This hepatic induction is dependent on distinct, spatio-temporal regulation including signals of fibroblast growth factor (FGF) and bone morphogenetic protein (BMP) from cardiac mesoderm and septum transversum mesenchyme (STM) respectively (Figure 1)47–50. Subsequent to the FGF and BMP signaling cascades, Wnt signaling from the mesoderm is also required for liver specification51,52.
Figure 1. Spatio-temporal Regulation of Cholangiocyte Development.
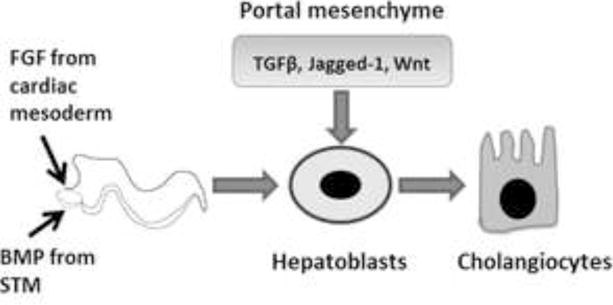
Schematic representation of key factors involved in biliary development from hepatoblasts. FGF: Fibroblast growth factor, BMP: Bone morphogenetic protein, STM: Septum tranversum mesenchyme, TGFβ: Transforming growth factor beta.
Between E9.0 and E9.5, hepatic endoderm cells called hepatoblasts delaminate from the epithelium and expand into the adjacent STM to form the liver bud, coordinated by signals from endothelial cells and a series of transcriptional events53–55. Sonic hedgehog (SHH) is expressed in the ventral foregut endoderm during development, but at the onset of liver bud formation, its expression is down regulated. At E11.5, hepatoblasts show expression of SHH and its downstream transcription factor, Gli-1, which are then later attenuated. Thus, a temporally restricted activation of Hh signaling appears to be required to promote hepatoblast proliferation, a signal which is then shut off for normal hepatic differentiation of the hepatoblasts56.
The hepatoblasts are bipotent and differentiate into both cholangiocytes and hepatocytes beginning around E13. Liver bud hepatoblasts residing adjacent to the portal tracts, upon influence of the portal mesenchyme, adopt a cholangiocyte fate and form the lumen of the intrahepatic bile ducts (IHBD), while the hepatoblasts in the parenchyma continue to differentiate toward hepatocytes. The maturation and specification of these cells are regulated by diverse growth factors, cytokines, and transcription factors, which have been reviewed in detail elsewhere24,57.
Parenchymal hepatocyte differentiation requires exposure to Oncostatin M secreted from the hematopoietic cells in the liver in combination with HGF and Wnt hormones58,59. The activity of these factors is further balanced by TNFα, which maintains the proliferation of fetal hepatocytes for appropriate liver growth. These signals together regulate a network of liver enriched transcription factors that control hepatocyte gene expression.
The biliary fate of the periportal hepatoblasts is orchestrated through temporally coordinated TGFβ, Notch, Wnt, and FGF signaling (Figure1)60–65. Jagged-1 (Jag-1), a notch ligand, is a key signaling molecule for biliary development, and is thought to be derived from the portal mesenchyme. Deletion of the Jag-1 gene in the portal mesenchyme results in profound defects in bile duct formation66. In humans, mutations in Jag-1 or Notch2 lead to bile duct paucity in Alagille syndrome (AGS) patients67–70. Furthermore, biliary differentiation is prevented by inhibiting notch signaling, whereas, ectopic notch signaling promotes parenchymal hepatoblasts to adopt a biliary fate62–64. Signaling through the Jag-1/Notch2 ligand-receptor pair, essential for biliary morphogenesis, is evolutionarily conserved in vertebrate liver development66,71,72.
Another important signaling pathway essential for biliary development is the TGFβ/Activin pathway. A gradient of TGFβ signaling exists in fetal liver, with high levels in the periportal region and low levels in the pericentral region, which controls the induction of the biliary fate. Wnt signals also regulate biliary differentiation from the hepatoblasts. Taken together, a combination of spatially restricted signaling factors collectively allow for biliary differentiation in the periportal region.
Biliary Regeneration and Repair
Among the solid organs, liver is distinguished in its unique and remarkable capacity to regenerate upon injury (surgical resection or toxic insults). Humans can tolerate a 70% hepatectomy, rodents can tolerate a 90% hepatectomy, and zebrafish liver can regenerate after near total obliteration of parenchymal hepatocytes74,75. While the bulk of historical work on liver regeneration focuses on hepatocyte regeneration, restoration of the biliary tree is also essential for proper organ function. Additionally, the peri-portal location of putative liver stem cell niches and the expansion of biliary progenitors seen in the context of chronic liver diseases puts cholangiocytes at center stage in discussions of liver regeneration and repair mechanisms76. In contrast to biliary development, which occurs through differentiation of hepatoblasts, it is useful to classify biliary regeneration and repair into three distinct domains: 1) homeostatic self-replication of cholangiocytes in normal liver; 2) accelerated biliary regeneration after liver resection; and 3) repair of the biliary tree upon injury (Figure 2).
Figure 2. Biliary Development, Regeneration, and Repair.
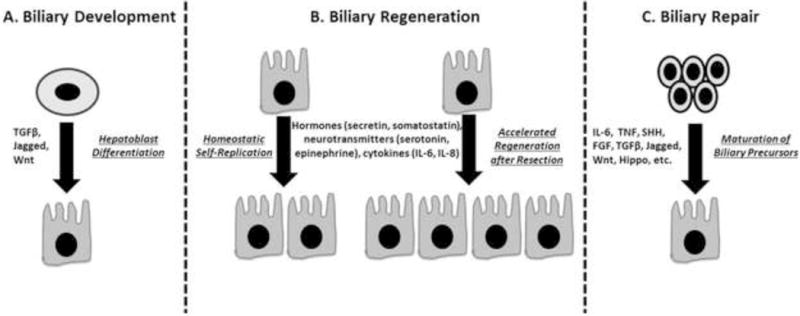
A) Biliary development involving differentiation of hepatoblasts. B) Biliary regeneration involving homeostatic self-replication of cholangiocytes in normal liver or accelerated self-replication following partial hepatectomy. C) Biliary repair involving activation and maturation of biliary precursors.
One of the early proposed models for liver cell replacement was the so-called “streaming liver hypothesis”, where a continuous flow of new hepatocytes would emerge from the periportal stem cell niche and progress toward the central vein region for eventual apoptosis, analogous to the crypt-to-villi flow of intestinal epithelial cells77. Although elegant, lineage tracing evidence eventually accumulated and argued against such a model78,79 and it is now fairly well-established that normal liver tissue maintenance is achieved via homeostatic self-replication of preexisting hepatocytes and cholangiocytes. Occasional and apparently random apoptosis of adult cells is counterbalanced by occasional and apparently random mitotic events.
The partial hepatectomy (PHx) model in rat was the first classical model of liver regeneration, described by Higgins and Anderson in 193180. They surgically excised two anterior lobes of the rat liver equating to 70% reduction in liver size. The cells of the remaining lobes proliferated to regain the lost mass over the course of 1–2 weeks. In this process, liver cells are able to overcome cell cycle checkpoints and re-enter the cell cycle. This is followed by waves of DNA synthesis, cellular hypertrophy, and proliferation. Organ mass and function is eventually restored through compensatory hypertrophy of the remnant lobes as opposed to regrowth of resected lobes. There is apparently little activation of the peri-portal stem cell niche in this circumstance, arguing that normal liver regeneration can be achieved by simple proliferation of adult parenchymal cells without invoking any stem/progenitor cell population.
However, when the liver suffers from severe and/or chronic damage, cellular proliferation and regenerative capacity are thought to be attenuated. In this circumstance, historical paradigms have suggested that there is emergence of a facultative bipotent liver stem/progenitor cell compartment to contribute to the process of liver regeneration. These stem/progenitor cells are also referred to as oval cells in rodent models, due to their stem cell-like morphology81,82, and have been extensively evaluated as a potential source of new hepatocytes during liver regeneration25,83. Even with suppressed hepatocyte proliferation in rats by administration of the chemical acetylaminofluorene (AAF), followed by PHx, oval cells can appear and expand in the periportal regions of the lobule within a few days and reach a peak at ~7–9 days after PHx. Administration of certain diets such as 3,5-diethoxycarbonyl-1,4-dihidro-collidine (DDC)84 feeding or the choline-deficient, ethanolamine supplemented (CDE)85 diet are well known to produce robust oval cell responses in both mice and rats. Cells bearing a strong resemblance to the rodent oval cells are also observed in human liver diseases. In humans, however, these cells are usually termed “hepatic progenitor cells” or “intermediate hepatobiliary cells”86. The hematopoietic and epidermal systems as well as the small intestine have defined stem cell populations responsible for normal cell turnover that have been isolated and anatomically localized. These stem cells exhibit self-renewal properties and differentiate into mature cell types under normal physiological conditions to replace cell losses in those organ systems, thereby maintaining normal tissue homeostasis. In the liver, normal hepatocyte and cholangiocyte turnover is slow and the concept of such a stem cell niche to support homeostatic self-renewal or tissue repair after injury has long been contemplated. The lack of a well-organized and universally accepted system of nomenclature for ductular cells and biliary precursors has led to a proliferation of poorly understood and loosely applied terminology (e.g. liver progenitor cells, biliary tree stem cells, atypical ductular cells, oval cells, biliary tree stem/progenitors, ductular hepatocytes, intermediate hepatocytes, reactive cholangiocytes). While each term is meant to strictly refer to a specific subpopulation of a heterogeneous group of cells along the biliary lineage, the terms are often used synonymously and interchangeably, which has led to some confusion in the literature. Perhaps the simplest term to apply to the diverse expansion of the biliary compartment seen in chronic liver disease is the descriptive term, ductular reactive cells (DRCs), which describes the emergence of a histologic lesion, known as the ductular reaction. The ductular reaction consists of activation and expansion of immature cholangiocytic cells that coalesce into primitive duct-like structures. The ductular reaction is, in fact, a collective signaling response to the surrounding hepatic stellate cells, macrophages, mature cholangiocytes, hepatocytes, portal myofibrobalsts, and endothelial cells, which form a niche that regulates the formation, expansion, and differentiation of DRCs87. This reaction is apparently an attempt to activate endogenous repair mechanisms, but can also be viewed as an abnormal regenerative response since it is accompanied by excessive ECM deposition and promotes the progression of fibrosis88–90.
The origin and fate of the ductular reactive cells is a subject of active debate in the literature and a topic on which concepts are rapidly evolving. What is clear is that a subpopulation of immature biliary-like cells is highly expanded in certain forms of chronic liver injury and that these cells have a bipotent differentiation capacity when isolated in vitro. There are varying theories on the origin of DRCs including a purely biliary origin or, alternatively, transdifferentiation of mature hepatocytes. Once established however, the ductular reaction appears to be composed of cells of a biliary phenotype, based on the following: 1) Many known oval cell markers are also markers for cholangiocytes91,92; 2) The arrangement of these cells histologically is often in a ductular pattern93; and 3) They typically emanate from and cluster near the portal tracts94. Historically, the canals of Hering, terminal structures where the hepatocyte canaliculi and the interlobular bile ducts interconnect, have been proposed as the site of origin for DRCs. Given the ideal anatomic location of this structure between the two epithelial cells, it was reasonable to postulate that it could serve as a niche to supply the putative stem cells for both the hepatocytes and cholangiocytes. Direct proof of this model is hampered due to lack of specific marker proteins and the inexact nature of lineage tracing studies. Additional stem cell niches are being described including the peri-biliary glands and a maturational lineage of stem and progenitor cells along the length of the biliary tree95. Recent studies also suggest that a self-renewing population of Axin2+ hepatocytes can regenerate damaged liver from a pericentral location through a reverse streaming mechanism96.
Stem Cells and Cellular Plasticity in Liver
In addition to the complexities surrounding the origin of DRCs, an even more relevant question is their ultimate fate (i.e. do ductular progenitors give rise to mature cholangiocytes to facilitate biliary repair and/or do they also contribute to parenchymal hepatocyte regeneration?). Pluripotent stem cell technology has revealed that adult cells, previously thought to be “terminally differentiated” retain remarkable cellular plasticity. This seems to be particularly true in the liver where complex transdifferentiation events are being documented, raising questions about whether or not cellular plasticity and reprograming events may obviate the need for stem cell-based liver regeneration97–99. The idea of biliary-derived DRCs being bipotent in vivo has been supported by lineage tracing studies utilizing Cre/Lox technology with various Cre drivers and injury models; however, there are also a number of publications that do not support or show minimal contribution of biliary cells to regenerating hepatocytes79,100–109 (Table 1). Studies done in zebrafish suggest that only in severe, but not in moderate hepatocyte ablation, that biliary cells can contribute to hepatocyte restoration74,75. Another recent study in mice also showed that following deletion of 98% of hepatocytes, transplanted cells of biliary origin contributed significantly to restoration of liver parenchyma, regenerating both hepatocytes and cholangiocytes110. The key limitations in definitively resolving the apparent contradictory findings lie in the imperfect and variable models of liver disease available (none of which accurately represent human chronic liver disease) and some technical limitations of lineage tracing studies. Genetic lineage tracing is a powerful strategy for in vivo fate tracing experiments because it allows for cell type specificity (using cell-specific Cre drivers) and Cre expression can be controlled temporally (with tamoxifen inducible systems). While lineage tracing remains the “Gold Standard” to trace the origin of new cells, a number of important caveats need to be kept in mind111,112. Leaky or unexpected Cre expression in different cell types, even in small amounts, can bias results. Injury states may alter the cell-specificity of Cre. Furthermore, tamoxifen can persist within the animal so that temporal precision of labelling is not always possible. Tamoxifen is also excreted in feces and can lead to Cre activation in untreated animals if co-housed. Despite these known limitations, genetic lineage tracing is likely to continue to provide novel and important insights into liver cell plasticity in the future.
Table 1.
Linage tracing studies utilizing various Cre drivers and injury models to study the fate of ductular reactive cells.
| Reference | Genetic Mice for Lineage Tracing | Injury Model |
|---|---|---|
| Sackett 2009 | Foxl1-Cre | BDL, DDC, CDE |
| Furuyama 2011 | Sox9 Cre | CCl4, PHx, CDE, BDL, DDC, APAP |
| Malato 2011 | Ttr-Cre | CCl4, PHx, BDL, DDC |
| Espanol-Suner 2012 | OPN-Cre | PHx, CCl4, CDE, DDC |
| Huch 2013 | LGR5-Cre | CCl4, MCDE, DDC |
| Rodrigo-Torres 2014 | HNF1β-Cre | CCl4, PHx, CDE, DDC, APAP |
| Schaub 2014 | Ttr-Cre, CK19-Cre, PDGFR-β-Cre | CDE |
| Tarlow 2014 | Sox9-Cre | PHx, CCl4, CDE, DDC |
| Yanger 2014 | KRT-19-Cre, TBG-Cre | CCl4, CDE, DDC, ANIT |
| Lu 2015 | KRT-19-Cre | CDE, ΔMdm2 |
| Kamimoto 2016 | Prom1-Cre, AAV8-iCre | TAA, DDC |
Sox9: SRY-related HMG-box transcription factor 9; OPN: Osteopontin; LGR5: Leucine-rich repeat-containing G-protein coupled receptor 5; KRT19/CK19: Cytokeratin 19; HNF1β: Hepatocyte Nuclear Factor 1 Homeobox B; Ttr: Transthyretin; AAV8: Adeno-Associated Virus 8; Foxl1: Forkhead Box l1; TBG: thyroid hormone-binding globulin; Prom-1: Prominin-1; PHx: Partial Hepatectomy; CCl4: Carbon tetrachloride; CDE: choline-deficient ethionine supplemented, DDC: 3,5-diethoxycarbonyl-1,4-dihiro-collidine; BDL: Bile-duct ligation; ANIT: Alpha napthylisothiocyanate; APAP: Acute acetaminophen; TAA: Thioacetamide.
Stem Cell-Derived Cholangiocytes
Human embryonic stem cells (hESC) and induced pluripotent stem cells (iPSC) have self-renewal capacity, are pluripotent, and have the ability to differentiate into cells of all three primary germ layers. Soon after the development of iPSC, a number of in vitro hepatocyte differentiation strategies quickly emerged. While these protocols produced cells with many features of adult hepatocytes, it has been notoriously difficult to achieve a fully mature adult hepatocyte phenotype and, because of this, the differentiated cells have typically been referred to as hepatocyte-like cells. Nonetheless, these protocols provide powerful tools for studying liver developmental biology, recapitulating specific disease phenotypes, and could potentially provide unlimited resources for drug testing applications and cell-based therapies.
The natural and logical evolution of the application of iPSC technology to liver disease was to develop iPSC-derived cholangiocytes for modeling biliary disease. Indeed, several groups have now published unique protocols for the differentiation of iPSC into cholangiocytes. The protocols available for the differentiation of stem cells to cholangiocytes were mainly designed to mimic the patterns and stages observed during biliary development in utero. All were designed with a stepwise approach in which the cells were exposed to soluble factors as they proceed through various phases of endodermal differentiation. In principle, these phases should mimic normal embryologic development through phases including a definitive endoderm phase, a hepatic specification phase, a hepatoblast-like phase, and a differentiated cholangiocyte phase (Table 2).
Table 2.
Growth factors employed by different groups for differentiation of stem cells to biliary cells.
| Reference | Definitive Endoderm | Hepatic Specification | Hepatic Progenitor | Cholangiocyte |
|---|---|---|---|---|
| Dianat 2014 | Wnt3A, ActivinA, FGF2 | ActivinA, FGF2, BMP4 | FGF4, HGF, EGF, RA | HG, EGF, IL-6, ST |
| De Assuncao 2015 | Wnt3A, ActivinA | SHH, BMP4, FGF2 | Jagged1, SHH | TGFβ |
| Ogawa 2015 | ActivinA | FGF2, BMP4 | HGF, Dex., OSM | HGF, EGF, TGFβ, OP9 co-culture |
| Sampaziotis 2015 | ActivinA,FGF2, BMP4 | BMP4 | FGF10, ActivinA, RA | EGF |
| Takayama 2016 | ActivinA | FGF4, BMP4 | Dex., HGF, EGF | EGF, IL-6, ST |
SHH: Sonic Hedgehog, TGFβ: Transforming growth factor beta, FGF: Fibroblast growth factor, BMP: Bone morphogenetic protein, HGF: Hepatocyte growth factor, EGF: Epidermal growth factor, Dex: Dexamethasone, OSM: Oncostatin M, RA: retinoic acid, IL-6: Interleukin-6, ST: Sodium Taurocholate, HG: Human Growth Hormone.
In 2014, Dianat et al., showed that embryonic stem cells could be differentiated to cholangiocytes and also subsequently applied the technique to iPSC and HepaRG cells. They generated hepatoblasts using methods previously described in the generation of hepatocytes from pluripotent cells. In a monolayer, cells in the hepatoblast stage were matured to cholangiocytes with exposure to human growth hormone (HG), epidermal growth factor (EGF), interleukin 6 (IL-6) and sodium taurocholate (ST). Cells from the final stage of differentiation expressed high levels of cholangiocyte markers such as CK7, CFTR, TGR5, HNF6, SOX9 and AQP1. The cells also formed cilia and when cultured in a 3D matrix, they developed epithelial/apicobasal polarity, and they formed functional cysts and biliary ducts20.
Our group utilized defined media and feeder-free culture conditions along with temporal exposure to key biliary morphogens to achieve cholangiocyte differentiation from patient-derived iPSC. We reported the use of temporally-restricted Hh signaling during the differentiation of iPSC to cholangiocytes. Hepatic specification was achieved using a combination of SHH, BMP4, and FGF2. To induce hepatic progenitor cells, SHH was also used in combination with Jag-1 (to activate NOTCH signaling). For the cholangiocyte maturation, TGFβ was used in conjunction with a collagen-1 matrix. The resulting cholangiocytes showed expression of cholangiocyte markers (CK19, CK7, PKD2, CFTR, AE2), the presence of primary cilia, intact calcium signaling, and were able to form duct-like structures in 3D culture. This protocol also showed that iPS-derived cholangiocytes were able to engraft within mouse liver in vivo, following retrograde intrabiliary infusion21.
Ogawa et al., used a combination of HGF, Dexamethasone, and oncostatin to obtain hepatic progenitors and a subsequent combination of HGF, EGF, and TGFβ to obtain mature cholangiocytes. Notch signaling was activated using co-culture with OP9 cells, a stromal cell line that expresses Notch ligands. The resulting cells, when grown in 3D culture, formed epithelialized cystic and/or ductular structures that expressed markers found in mature cholangiocytes. This study also showed that cholangiocytes generated from the iPSC of cystic fibrosis patients had impaired cyst swelling and that CFTR chemical correctors increased the levels of CFTR on the apical side of the lumen and augmented cyst swelling22.
Sampaziotis et al., showed that in their protocol FGF10, retinoic acid, and TGFβ were key factors for biliary specification. Their iPSC derived cholangiocytes were characterized using a range of assays and they resembled primary common bile duct cholangiocytes at the transcriptional level. Patient derived iPSC in 3D culture conditions were used to model polycystic liver disease and cystic fibrosis23. 3D cultures of iPS-derived cholangiocytes generated from polycystic patients were responsive to secretin and somatostatin and this response was blunted by the use of octreotide. They also showed that the cholangiocytes derived from cystic fibrosis patients had minimal CFTR protein expression and were unable to modify intracellular chloride. They corrected the disease phenotype with an experimental drug, VX809.
Takayama et al., used an approach similar to the Dianat protocol to promote cholangiocyte differentiation. The maturation of their hepatoblast like cells was done with HG, EGF and IL-6 along with extracellular matrix molecules. The gene expression levels of the cholangiocyte markers, AQP1, SOX9, CFTR, G protein-coupled bile acid receptor 1, Jag-1, secretin receptor, and GGT were all increased by using laminin 411 or laminin 511 as a matrix113.
Overall, the collective efforts at generating cholangiocytes derived from stem cells have been remarkably successful. These cells are already proving to be promising tools that can be used in many applications such as cell transplantation studies, biliary disease modeling, deciphering biliary development, and small molecule screening. While each of the various differentiation protocols have some similarities and some unique aspects, it is quite fascinating that the disparate protocols all seem to result in a mature, adult cholangiocyte phenotype, insofar as these features have been evaluated. This may suggest that, in vitro, the default differentiation pathway of liver progenitors, is biased toward cholangiocyte differentiation, especially given the difficulties noted in generating fully mature stem cell-derived hepatocytes. This is in contradistinction to the conceptualized sequence of events in liver development where it is thought that intervening signals from the portal mesenchyme are required for cholangiocyte differentiation while the “default” fate for most hepatoblasts is thought to be hepatocellular. Most likely, additional fine-tuning of the various cholangiocyte differentiation approaches will be needed to achieve the most robust and efficient cholangiocyte differentiation possible. It may also be that certain protocols will have advantages or disadvantages in certain applications. Regardless, this powerful new technology has provided unique opportunities to study cholangiocyte development, pathogenesis, and treatment strategies in ways that were previously unthinkable. When combined with large scale efforts that are underway to generate robust biorepositories of iPSCs, iPSC-derived cholangiocytes, and biliary organoids from subjects with cholangiopathies, this technology will become an even more powerful tool for individualized medicine applications.
3-Dimensional Culture Systems and Biliary Organoids
Organoids are collections of organ-specific cell types that develop from stem cells or organ progenitors114. Organoid formation recapitulates the major processes of self-organization during development including cell sorting and spatially-restricted lineage commitment in a manner similar to the in vivo environment115. Grouped together and spatially organized similar to an organ, these structures are capable of recapitulating specific functions of the adult organs (e.g. secretion / absorption, filtration, neural activity, contraction, etc.). An important distinction to bear in mind is the difference between 3-dimentional (3D) cultures of a single cell type and true organoids, which consist of multiple self-organizing cell types. An example of this distinction from the intestine is the difference between an enteroid (which consists of a single cell type in 3D culture) and a true intestinal organoid (a stem cell-derived and self-organizing multicellular cluster). Organoid research has tremendous potential, across multiple organ systems, to facilitate disease modeling, pharmacologic testing, and therapeutic regenerative medicine applications (Figure 3). Indeed, organoids have been generated to model diverse organs including kidney, lung, brain, thyroid, and others114. Within gastroenterology, organoid systems have been generated to represent esophagus, stomach, pancreas, small intestine, large intestine, and pancreas116. Liver organoids have been generated from both tissue derived progenitor cells103,117 and from iPSC118. Organoid research is a logical extension of the existing iPSC technology that been leveraged toward the biliary system. Furthermore, the infrastructure for overseeing biorepositories of biliary organoids likely already exists within existing iPSC biorepositories.
Figure 3. iPSC-Derived Cholangiocytes and Applications.
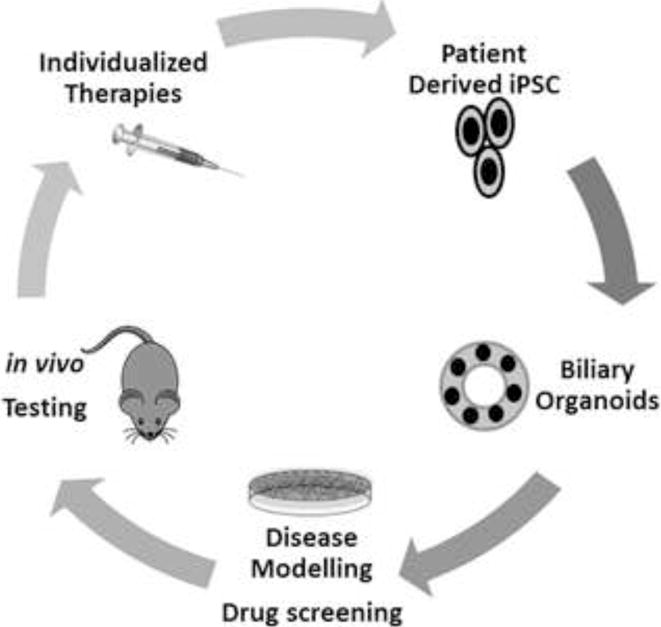
Schematic depicting patient-derived iPSC that could potentially be expanded as organoids, utilized for disease modelling and drug screening, tested in vivo, and eventually returned to the patient as an individualized therapy.
Several groups have utilized 3D culture systems to facilitate research in primary liver stem cells. As early as 2001, it was shown that primary liver cells cultured in 3D conditions could be maintained in vitro in the presence of EGF, HGF, and dexamethasone119. Mouse Foxl1+ hepatic progenitor cells, when cultured in type 1 collagen gels, formed CK19 positive branches120. In 2007, Tanimizu et al., showed that HPPL, a mouse liver progenitor cell line, when cultured in 3D, formed cysts with a luminal space and apicobasal polarity121. Kido et al., showed that CPM+ cells, sorted and matured to cholangiocytes in 3D culture, formed cysts with a luminal structure and proper apicobasal polarity122. Recently, it was also shown that rat liver stem cells can be isolated and grown as cystic structures when cultured in high levels of Wnt3a and noggin (a BMP signaling inhibitor). Gallbladder stem/progenitor cells from non-injured livers can form 3D structures that express stem cell markers in the presence of R-spondin 1, noggin, and nicotinamide123. Yu et al. published a protocol for direct reprograming of fibroblasts to induced hepatic stem cells. These cells expressed hepatocyte and cholangiocyte markers in vitro and were able to form cysts and branching structures that were positive for CK19 and CK7 under 3D culture124.
In 2013, an elegant study generated true liver organoids by combining human-iPSC-derived hepatic cells with mesenchymal stem cells and endothelial cells. Although the cells were cultured in two-dimensional conditions, the cells self-organized into three-dimensional clusters, which resembled embryonic liver118. When transplanted in mice these organ buds developed into hepatic tissue with features of adult liver. Notably, however, these organoids lacked biliary elements. Huch et al. have demonstrated that LGR5+ adult liver stem cells can be isolated and cultured as liver organoids in a 3D culture method involving matrigel with HGF, EGF, FGF, and Rspondin1103. The expanded cells self-organized into 3D structures with a ductal (CK19+) single-layered epithelium compartment and a pseudo-stratified compartment expressing both ductal (CK7, CK19) and hepatocyte (E-cadherin, Hnf4a) markers. When cultured in 3D, a single LGR5+ cell can be expanded as organoids and differentiated into hepatocytes and cholangiocytes. In 2015, the same group used a similar approach to expand liver biopsy tissue and single EPCAM+ cells as liver organoids117. As powerful as these emerging systems are, organoid technology does have some notable limitations. Since they are generated in vitro, organoids in culture may not fully recapitulate all aspects of liver development that occur in the in vivo environment. In particular, they lack several essential components of the intact liver such as a vascular system, interaction with other cell types, specific extracellular matrix interactions, and immune surveillance (although organoid transplantation studies may help to overcome some of these issues). Despite these known limitations, 3D culture systems and liver organoid technology are rapidly advancing and are likely to be increasingly utilized for individualized disease modelling and regenerative medicine applications for the biliary system.
Summary, Future Applications, and Regenerative Therapeutics
In the past several decades, we have witnessed significant advances in our understanding of biliary development, basic physiology of cholangiocytes, and pathogenesis of the cholangiopathies. Despite these advances, effective treatment modalities remain elusive. For this reason, the recent explosion of work in biliary regenerative medicine is particularly encouraging and provides great hope for future regenerative therapeutics for biliary disease. In this review, we outlined some of the primary clinical challenges associated with the biliary system and we identified a standard regenerative medicine paradigm involving replacement, regeneration, and rejuvenation that may help to categorize the development of future regenerative therapies. We also highlighted both historical and more recent basic science advances in biliary development, regeneration, and repair, and we reviewed new regenerative technologies involving iPS-derived cholangiocytes and biliary organoids. All of these conceptual and technical advances now set the stage for future translation and application in order to ultimately develop new regenerative service lines for patients with biliary disease. When and how these new therapies will emerge is unknowable, but development and translation will likely require multidisciplinary transformational teams consisting of basic scientists, hepatologists, biliary and transplant surgeons, as well as interventional radiologists and advanced endoscopists. It is likely that the field will take advantage of its existing endoscopic access to the biliary tree by building new platforms in regenerative endoscopy. ERCP-based delivery of cell-based (e.g. stem cells or stem-cell derived cholangiocytes) or cell-free regenerative therapeutics (e.g. biodegradable stents or therapeutic exosome delivery) are particularly appealing since advanced endoscopy is available at academic centers throughout the world. Other technological applications being contemplated include 3D bioprinting of ductular tissue, biliary biostents incorporating cellular elements, and tissue engineering approaches, such as recellularization of decellularized bile duct units. Our embrace of regenerative medicine also involves some challenges. We will need to face down the issues of inexact technologies, inefficient or incomplete differentiation from stem cells, epigenetic memory, and malignancy potential, as well as highly complex tissue engineering and regulatory challenges. In the end, however, we are left with great hope for clinical advances. Clearly, the future is bright in terms of regenerative medicine and the biliary tree. We are, indeed, awakening to the dawn of a new golden age in regenerative hepatology that promises development and application of new regenerative therapeutics for previously untreatable liver diseases.
Acknowledgments
Financial Support: This work was supported by grants DK100575, DK084567, and the Loan Repayment Program from the National Institutes of Health; by Regenerative Medicine Minnesota; by Gilead Sciences; and by the Mayo Clinic Center for Regenerative Medicine.
List of Abbreviations
- AAF
acetylaminofluorene
- AQP
Aquaporin
- BMP
bone morphogenetic protein
- CCL4
carbon tetrachloride
- CDE
choline-deficientethione
- CFTR
cystic fibrosis transmembrane conductance regulator
- CK
cytokeratin
- DDC
3,5-diethoxycarbonyl-1,4-dhidro-collidine
- DRCs
ductular reactive cells
- ECM
extracellular matrix
- EGF
epidermal growth factor
- ERCP
endoscopic retrograde cholangiopancreatography
- FGF
fibroblast growth factor
- Hh
Hedgehog
- HG
human growth hormone
- hESC
human embryonic stem cells
- HNF
Hepatocyte Nuclear Factor
- IL-6
interleukin 6
- iPSC
induced pluripotent stem cells
- Jag-1
Jagged-1
- OPN
Osteopontin
- PKD
Polycystic kidney disease
- PHx
partial hepatectomy
- SHH
sonic hedgehog
- Sox9
HMG-box transcription factor 9
- STM
septum transversum mesenchyme
- TGFβ
transforming growth factor beta
Contributor Information
Thiago M. De Assuncao, Email: deassuncao.thiago@mayo.edu.
Nidhi Jalan-Sakrikar, Email: sakrikar.nidhi@mayo.edu.
Robert C. Huebert, Email: huebert.robert@mayo.edu.
References
- 1.Forbes SJ, Newsome PN. Liver regeneration - mechanisms and models to clinical application. Nat Rev Gastroenterol Hepatol. 2016 Aug;13(8):473–485. doi: 10.1038/nrgastro.2016.97. [DOI] [PubMed] [Google Scholar]
- 2.Takahashi K, Yamanaka S. Induction of pluripotent stem cells from mouse embryonic and adult fibroblast cultures by defined factors. Cell. 2006 Aug 25;126(4):663–676. doi: 10.1016/j.cell.2006.07.024. [DOI] [PubMed] [Google Scholar]
- 3.Si-Tayeb K, Noto FK, Nagaoka M, et al. Highly efficient generation of human hepatocyte-like cells from induced pluripotent stem cells. Hepatology. 2010 Jan;51(1):297–305. doi: 10.1002/hep.23354. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Sancho-Bru P, Roelandt P, Narain N, et al. Directed differentiation of murine-induced pluripotent stem cells to functional hepatocyte-like cells. J Hepatol. 2011 Jan;54(1):98–107. doi: 10.1016/j.jhep.2010.06.014. [DOI] [PubMed] [Google Scholar]
- 5.Choi SM, Kim Y, Liu H, Chaudhari P, Ye Z, Jang YY. Liver engraftment potential of hepatic cells derived from patient-specific induced pluripotent stem cells. Cell cycle. 2011 Aug 1;10(15):2423–2427. doi: 10.4161/cc.10.15.16869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Chen YF, Tseng CY, Wang HW, Kuo HC, Yang VW, Lee OK. Rapid generation of mature hepatocyte-like cells from human induced pluripotent stem cells by an efficient three-step protocol. Hepatology. 2012 Apr;55(4):1193–1203. doi: 10.1002/hep.24790. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Yu Y, Liu H, Ikeda Y, et al. Hepatocyte-like cells differentiated from human induced pluripotent stem cells: relevance to cellular therapies. Stem Cell Res. 2012 Nov;9(3):196–207. doi: 10.1016/j.scr.2012.06.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Aravalli RN, Cressman EN, Steer CJ. Hepatic differentiation of porcine induced pluripotent stem cells in vitro. Veterinary journal. 2012 Dec;194(3):369–374. doi: 10.1016/j.tvjl.2012.05.013. [DOI] [PubMed] [Google Scholar]
- 9.Zhu S, Rezvani M, Harbell J, et al. Mouse liver repopulation with hepatocytes generated from human fibroblasts. Nature. 2014 Apr 3;508(7494):93–97. doi: 10.1038/nature13020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Szkolnicka D, Farnworth SL, Lucendo-Villarin B, Hay DC. Deriving functional hepatocytes from pluripotent stem cells. Curr Protoc Stem Cell Biol. 2014;30:1G 5 1–1G 5 12. doi: 10.1002/9780470151808.sc01g05s30. [DOI] [PubMed] [Google Scholar]
- 11.Takayama K, Inamura M, Kawabata K, et al. Generation of metabolically functioning hepatocytes from human pluripotent stem cells by FOXA2 and HNF1alpha transduction. J Hepatol. 2012 Sep;57(3):628–636. doi: 10.1016/j.jhep.2012.04.038. [DOI] [PubMed] [Google Scholar]
- 12.Schwartz RE, Fleming HE, Khetani SR, Bhatia SN. Pluripotent stem cell-derived hepatocyte-like cells. Biotechnol Adv. 2014 Mar-Apr;32(2):504–513. doi: 10.1016/j.biotechadv.2014.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Cayo MA, Cai J, DeLaForest A, et al. JD induced pluripotent stem cell-derived hepatocytes faithfully recapitulate the pathophysiology of familial hypercholesterolemia. Hepatology. 2012 Dec;56(6):2163–2171. doi: 10.1002/hep.25871. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Xu D, Alipio Z, Fink LM, et al. Phenotypic correction of murine hemophilia A using an iPS cell-based therapy. Proc Natl Acad Sci U S A. 2009 Jan 20;106(3):808–813. doi: 10.1073/pnas.0812090106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Yusa K, Rashid ST, Strick-Marchand H, et al. Targeted gene correction of alpha1-antitrypsin deficiency in induced pluripotent stem cells. Nature. 2011 Oct 20;478(7369):391–394. doi: 10.1038/nature10424. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Rashid ST, Corbineau S, Hannan N, et al. Modeling inherited metabolic disorders of the liver using human induced pluripotent stem cells. J Clin Invest. 2010 Sep;120(9):3127–3136. doi: 10.1172/JCI43122. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Huebert RC, Rakela J. Cellular therapy for liver disease. Mayo Clinic proceedings. 2014 Mar;89(3):414–424. doi: 10.1016/j.mayocp.2013.10.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Yu Y, Fisher JE, Lillegard JB, Rodysill B, Amiot B, Nyberg SL. Cell therapies for liver diseases. Liver transplantation : official publication of the American Association for the Study of Liver Diseases and the International Liver Transplantation Society. 2012 Jan;18(1):9–21. doi: 10.1002/lt.22467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Subba Rao M, Sasikala M, Nageshwar Reddy D. Thinking outside the liver: induced pluripotent stem cells for hepatic applications. World J Gastroenterol. 2013 Jun 14;19(22):3385–3396. doi: 10.3748/wjg.v19.i22.3385. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Dianat N, Dubois-Pot-Schneider H, Steichen C, et al. Generation of functional cholangiocyte-like cells from human pluripotent stem cells and HepaRG cells. Hepatology. 2014 Aug;60(2):700–714. doi: 10.1002/hep.27165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.De Assuncao TM, Sun Y, Jalan-Sakrikar N, et al. Development and characterization of human-induced pluripotent stem cell-derived cholangiocytes. Lab Invest. 2015 Oct;95(10):1218. doi: 10.1038/labinvest.2015.99. [DOI] [PubMed] [Google Scholar]
- 22.Ogawa M, Ogawa S, Bear CE, et al. Directed differentiation of cholangiocytes from human pluripotent stem cells. Nat Biotechnol. 2015 Aug;33(8):853–861. doi: 10.1038/nbt.3294. [DOI] [PubMed] [Google Scholar]
- 23.Sampaziotis F, Cardoso de Brito M, Madrigal P, et al. Cholangiocytes derived from human induced pluripotent stem cells for disease modeling and drug validation. Nat Biotechnol. 2015 Aug;33(8):845–852. doi: 10.1038/nbt.3275. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Strazzabosco M, Fabris L. Development of the bile ducts: essentials for the clinical hepatologist. J Hepatol. 2012 May;56(5):1159–1170. doi: 10.1016/j.jhep.2011.09.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Itoh T. Stem/progenitor cells in liver regeneration. Hepatology. 2016 Aug;64(2):663–668. doi: 10.1002/hep.28661. [DOI] [PubMed] [Google Scholar]
- 26.Ghanekar A, Kamath BM. Cholangiocytes derived from induced pluripotent stem cells for disease modeling. Curr Opin Gastroenterol. 2016 May;32(3):210–215. doi: 10.1097/MOG.0000000000000260. [DOI] [PubMed] [Google Scholar]
- 27.Hindley CJ, Cordero-Espinoza L, Huch M. Organoids from adult liver and pancreas: Stem cell biology and biomedical utility. Developmental biology. 2016 Jun 27; doi: 10.1016/j.ydbio.2016.06.039. [DOI] [PubMed] [Google Scholar]
- 28.Tabibian JH, Masyuk AI, Masyuk TV, O’Hara SP, LaRusso NF. Physiology of cholangiocytes. Compr Physiol. 2013 Jan;3(1):541–565. doi: 10.1002/cphy.c120019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.O’Hara SP, Tabibian JH, Splinter PL, LaRusso NF. The dynamic biliary epithelia: molecules, pathways, and disease. J Hepatol. 2013 Mar;58(3):575–582. doi: 10.1016/j.jhep.2012.10.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Lazaridis KN, Strazzabosco M, Larusso NF. The cholangiopathies: disorders of biliary epithelia. Gastroenterology. 2004 Nov;127(5):1565–1577. doi: 10.1053/j.gastro.2004.08.006. [DOI] [PubMed] [Google Scholar]
- 31.Halilbasic E, Fuchs C, Hofer H, Paumgartner G, Trauner M. Therapy of Primary Sclerosing Cholangitis–Today and Tomorrow. Dig Dis. 2015;33(Suppl 2):149–163. doi: 10.1159/000440827. [DOI] [PubMed] [Google Scholar]
- 32.Carbone M, Ronca V, Bruno S, Invernizzi P, Mells GF. Toward precision medicine in primary biliary cholangitis. Dig Liver Dis. 2016 Aug;48(8):843–850. doi: 10.1016/j.dld.2016.05.023. [DOI] [PubMed] [Google Scholar]
- 33.Mousa HS, Carbone M, Malinverno F, Ronca V, Gershwin ME, Invernizzi P. Novel therapeutics for primary biliary cholangitis: Toward a disease-stage-based approach. Autoimmun Rev. 2016 Sep;15(9):870–876. doi: 10.1016/j.autrev.2016.07.003. [DOI] [PubMed] [Google Scholar]
- 34.Mikolajczyk AE, Te HS, Chapman AB. Gastrointestinal Manifestations of Autosomal-Dominant Polycystic Kidney Disease. Clin Gastroenterol Hepatol. 2016 Jun 29; doi: 10.1016/j.cgh.2016.06.017. [DOI] [PubMed] [Google Scholar]
- 35.Verkade HJ, Bezerra JA, Davenport M, et al. Biliary atresia and other cholestatic childhood diseases: Advances and future challenges. J Hepatol. 2016 Sep;65(3):631–642. doi: 10.1016/j.jhep.2016.04.032. [DOI] [PubMed] [Google Scholar]
- 36.Rizvi S, Gores GJ. Pathogenesis, diagnosis, and management of cholangiocarcinoma. Gastroenterology. 2013 Dec;145(6):1215–1229. doi: 10.1053/j.gastro.2013.10.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lazaridis KN, LaRusso NF. The Cholangiopathies. Mayo Clin Proc. 2015 Jun;90(6):791–800. doi: 10.1016/j.mayocp.2015.03.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Nelson TJ, Behfar A, Terzic A. Strategies for therapeutic repair: The “R(3)” regenerative medicine paradigm. Clin Transl Sci. 2008 Sep;1(2):168–171. doi: 10.1111/j.1752-8062.2008.00039.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Atala A. Advances in tissue and organ replacement. Curr Stem Cell Res Ther. 2008 Jan;3(1):21–31. doi: 10.2174/157488808783489435. [DOI] [PubMed] [Google Scholar]
- 40.Mavila N, Trecartin A, Spurrier R, et al. Functional Human and Murine Tissue-Engineered Liver Is Generated From Adult Stem/Progenitor Cells. Stem Cells Transl Med. 2016 Aug 30; doi: 10.5966/sctm.2016-0205. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Korbling M, Estrov Z. Adult stem cells for tissue repair - a new therapeutic concept? N Engl J Med. 2003 Aug 7;349(6):570–582. doi: 10.1056/NEJMra022361. [DOI] [PubMed] [Google Scholar]
- 42.Nicolas CT, Wang Y, Nyberg SL. Cell therapy in chronic liver disease. Curr Opin Gastroenterol. 2016 May;32(3):189–194. doi: 10.1097/MOG.0000000000000262. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Surani MA, McLaren A. Stem cells: a new route to rejuvenation. Nature. 2006 Sep 21;443(7109):284–285. doi: 10.1038/443284a. [DOI] [PubMed] [Google Scholar]
- 44.Basu J, Ludlow JW. Exosomes for repair, regeneration and rejuvenation. Expert Opin Biol Ther. 2016;16(4):489–506. doi: 10.1517/14712598.2016.1131976. [DOI] [PubMed] [Google Scholar]
- 45.Conboy IM, Conboy MJ, Rebo J. Systemic Problems: A perspective on stem cell aging and rejuvenation. Aging (Albany NY) 2015 Oct;7(10):754–765. doi: 10.18632/aging.100819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Tremblay KD, Zaret KS. Distinct populations of endoderm cells converge to generate the embryonic liver bud and ventral foregut tissues. Dev Biol. 2005 Apr 1;280(1):87–99. doi: 10.1016/j.ydbio.2005.01.003. [DOI] [PubMed] [Google Scholar]
- 47.Gualdi R, Bossard P, Zheng M, Hamada Y, Coleman JR, Zaret KS. Hepatic specification of the gut endoderm in vitro: cell signaling and transcriptional control. Genes Dev. 1996 Jul 1;10(13):1670–1682. doi: 10.1101/gad.10.13.1670. [DOI] [PubMed] [Google Scholar]
- 48.Jung J, Zheng M, Goldfarb M, Zaret KS. Initiation of mammalian liver development from endoderm by fibroblast growth factors. Science. 1999 Jun 18;284(5422):1998–2003. doi: 10.1126/science.284.5422.1998. [DOI] [PubMed] [Google Scholar]
- 49.Rossi JM, Dunn NR, Hogan BL, Zaret KS. Distinct mesodermal signals, including BMPs from the septum transversum mesenchyme, are required in combination for hepatogenesis from the endoderm. Genes Dev. 2001 Aug 1;15(15):1998–2009. doi: 10.1101/gad.904601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Serls AE, Doherty S, Parvatiyar P, Wells JM, Deutsch GH. Different thresholds of fibroblast growth factors pattern the ventral foregut into liver and lung. Development. 2005 Jan;132(1):35–47. doi: 10.1242/dev.01570. [DOI] [PubMed] [Google Scholar]
- 51.Goessling W, North TE, Lord AM, et al. APC mutant zebrafish uncover a changing temporal requirement for wnt signaling in liver development. Dev Biol. 2008 Aug 1;320(1):161–174. doi: 10.1016/j.ydbio.2008.05.526. [DOI] [PubMed] [Google Scholar]
- 52.Ober EA, Verkade H, Field HA, Stainier DY. Mesodermal Wnt2b signalling positively regulates liver specification. Nature. 2006 Aug 10;442(7103):688–691. doi: 10.1038/nature04888. [DOI] [PubMed] [Google Scholar]
- 53.Bort R, Signore M, Tremblay K, Martinez Barbera JP, Zaret KS. Hex homeobox gene controls the transition of the endoderm to a pseudostratified, cell emergent epithelium for liver bud development. Dev Biol. 2006 Feb 1;290(1):44–56. doi: 10.1016/j.ydbio.2005.11.006. [DOI] [PubMed] [Google Scholar]
- 54.Margagliotti S, Clotman F, Pierreux CE, et al. The Onecut transcription factors HNF-6/OC-1 and OC-2 regulate early liver expansion by controlling hepatoblast migration. Dev Biol. 2007 Nov 15;311(2):579–589. doi: 10.1016/j.ydbio.2007.09.013. [DOI] [PubMed] [Google Scholar]
- 55.Sosa-Pineda B, Wigle JT, Oliver G. Hepatocyte migration during liver development requires Prox1. Nat Genet. 2000 Jul;25(3):254–255. doi: 10.1038/76996. [DOI] [PubMed] [Google Scholar]
- 56.Hirose Y, Itoh T, Miyajima A. Hedgehog signal activation coordinates proliferation and differentiation of fetal liver progenitor cells. Exp Cell Res. 2009 Sep 10;315(15):2648–2657. doi: 10.1016/j.yexcr.2009.06.018. [DOI] [PubMed] [Google Scholar]
- 57.Zong Y, Stanger BZ. Molecular mechanisms of liver and bile duct development. Wiley Interdiscip Rev Dev Biol. 2012 Sep-Oct;1(5):643–655. doi: 10.1002/wdev.47. [DOI] [PubMed] [Google Scholar]
- 58.Kamiya A, Gonzalez FJ. TNF-alpha regulates mouse fetal hepatic maturation induced by oncostatin M and extracellular matrices. Hepatology. 2004 Sep;40(3):527–536. doi: 10.1002/hep.20362. [DOI] [PubMed] [Google Scholar]
- 59.Schmidt C, Bladt F, Goedecke S, et al. Scatter factor/hepatocyte growth factor is essential for liver development. Nature. 1995 Feb 23;373(6516):699–702. doi: 10.1038/373699a0. [DOI] [PubMed] [Google Scholar]
- 60.Decaens T, Godard C, de Reynies A, et al. Stabilization of beta-catenin affects mouse embryonic liver growth and hepatoblast fate. Hepatology. 2008 Jan;47(1):247–258. doi: 10.1002/hep.21952. [DOI] [PubMed] [Google Scholar]
- 61.Clotman F, Jacquemin P, Plumb-Rudewiez N, et al. Control of liver cell fate decision by a gradient of TGF beta signaling modulated by Onecut transcription factors. Genes Dev. 2005 Aug 15;19(16):1849–1854. doi: 10.1101/gad.340305. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Zong Y, Panikkar A, Xu J, et al. Notch signaling controls liver development by regulating biliary differentiation. Development. 2009 May;136(10):1727–1739. doi: 10.1242/dev.029140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Tchorz JS, Kinter J, Muller M, Tornillo L, Heim MH, Bettler B. Notch2 signaling promotes biliary epithelial cell fate specification and tubulogenesis during bile duct development in mice. Hepatology. 2009 Sep;50(3):871–879. doi: 10.1002/hep.23048. [DOI] [PubMed] [Google Scholar]
- 64.Tanimizu N, Miyajima A. Notch signaling controls hepatoblast differentiation by altering the expression of liver-enriched transcription factors. J Cell Sci. 2004 Jul 1;117(Pt 15):3165–3174. doi: 10.1242/jcs.01169. [DOI] [PubMed] [Google Scholar]
- 65.Yanai M, Tatsumi N, Hasunuma N, Katsu K, Endo F, Yokouchi Y. FGF signaling segregates biliary cell-lineage from chick hepatoblasts cooperatively with BMP4 and ECM components in vitro. Dev Dyn. 2008 May;237(5):1268–1283. doi: 10.1002/dvdy.21520. [DOI] [PubMed] [Google Scholar]
- 66.Hofmann JJ, Zovein AC, Koh H, Radtke F, Weinmaster G, Iruela-Arispe ML. Jagged1 in the portal vein mesenchyme regulates intrahepatic bile duct development: insights into Alagille syndrome. Development. 2010 Dec;137(23):4061–4072. doi: 10.1242/dev.052118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.McDaniell R, Warthen DM, Sanchez-Lara PA, et al. NOTCH2 mutations cause Alagille syndrome, a heterogeneous disorder of the notch signaling pathway. Am J Hum Genet. 2006 Jul;79(1):169–173. doi: 10.1086/505332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.McCright B, Lozier J, Gridley T. A mouse model of Alagille syndrome: Notch2 as a genetic modifier of Jag1 haploinsufficiency. Development. 2002 Feb;129(4):1075–1082. doi: 10.1242/dev.129.4.1075. [DOI] [PubMed] [Google Scholar]
- 69.Li L, Krantz ID, Deng Y, et al. Alagille syndrome is caused by mutations in human Jagged1, which encodes a ligand for Notch1. Nat Genet. 1997 Jul;16(3):243–251. doi: 10.1038/ng0797-243. [DOI] [PubMed] [Google Scholar]
- 70.Oda T, Elkahloun AG, Pike BL, et al. Mutations in the human Jagged1 gene are responsible for Alagille syndrome. Nat Genet. 1997 Jul;16(3):235–242. doi: 10.1038/ng0797-235. [DOI] [PubMed] [Google Scholar]
- 71.Geisler F, Nagl F, Mazur PK, et al. Liver-specific inactivation of Notch2, but not Notch1, compromises intrahepatic bile duct development in mice. Hepatology. 2008 Aug;48(2):607–616. doi: 10.1002/hep.22381. [DOI] [PubMed] [Google Scholar]
- 72.Lorent K, Yeo SY, Oda T, et al. Inhibition of Jagged-mediated Notch signaling disrupts zebrafish biliary development and generates multi-organ defects compatible with an Alagille syndrome phenocopy. Development. 2004 Nov;131(22):5753–5766. doi: 10.1242/dev.01411. [DOI] [PubMed] [Google Scholar]
- 73.Hussain SZ, Sneddon T, Tan X, Micsenyi A, Michalopoulos GK, Monga SP. Wnt impacts growth and differentiation in ex vivo liver development. Exp Cell Res. 2004 Jan 1;292(1):157–169. doi: 10.1016/j.yexcr.2003.08.020. [DOI] [PubMed] [Google Scholar]
- 74.Choi TY, Ninov N, Stainier DY, Shin D. Extensive conversion of hepatic biliary epithelial cells to hepatocytes after near total loss of hepatocytes in zebrafish. Gastroenterology. 2014 Mar;146(3):776–788. doi: 10.1053/j.gastro.2013.10.019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.He J, Lu H, Zou Q, Luo L. Regeneration of liver after extreme hepatocyte loss occurs mainly via biliary transdifferentiation in zebrafish. Gastroenterology. 2014 Mar;146(3):789–800. doi: 10.1053/j.gastro.2013.11.045. e788. [DOI] [PubMed] [Google Scholar]
- 76.Itoh T. Stem/progenitor cells in liver regeneration. Hepatology. 2016 Aug;64(2):663–668. doi: 10.1002/hep.28661. [DOI] [PubMed] [Google Scholar]
- 77.Zajicek G, Oren R, Weinreb M., Jr The streaming liver. Liver. 1985 Dec;5(6):293–300. doi: 10.1111/j.1600-0676.1985.tb00252.x. [DOI] [PubMed] [Google Scholar]
- 78.Malato Y, Naqvi S, Schurmann N, et al. Fate tracing of mature hepatocytes in mouse liver homeostasis and regeneration. J Clin Invest. 2011 Dec;121(12):4850–4860. doi: 10.1172/JCI59261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Espanol-Suner R, Carpentier R, Van Hul N, et al. Liver progenitor cells yield functional hepatocytes in response to chronic liver injury in mice. Gastroenterology. 2012 Dec;143(6):1564–1575 e1567. doi: 10.1053/j.gastro.2012.08.024. [DOI] [PubMed] [Google Scholar]
- 80.Higgins GMARM. Experimental pathology of the liver I. Restoration of the liver of the white rat following partial surgical removal. Arch Pathol. 1931;12:186–202. [Google Scholar]
- 81.Farber E. Similarities in the sequence of early histological changes induced in the liver of the rat by ethionine, 2-acetylamino-fluorene, and 3′-methyl-4-dimethylaminoazobenzene. Cancer Res. 1956 Feb;16(2):142–148. [PubMed] [Google Scholar]
- 82.Fausto N. Liver regeneration and repair: hepatocytes, progenitor cells, and stem cells. Hepatology. 2004 Jun;39(6):1477–1487. doi: 10.1002/hep.20214. [DOI] [PubMed] [Google Scholar]
- 83.Duncan AW, Dorrell C, Grompe M. Stem cells and liver regeneration. Gastroenterology. 2009 Aug;137(2):466–481. doi: 10.1053/j.gastro.2009.05.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Preisegger KH, Factor VM, Fuchsbichler A, Stumptner C, Denk H, Thorgeirsson SS. Atypical ductular proliferation and its inhibition by transforming growth factor beta1 in the 3,5-diethoxycarbonyl-1,4-dihydrocollidine mouse model for chronic alcoholic liver disease. Lab Invest. 1999 Feb;79(2):103–109. [PubMed] [Google Scholar]
- 85.Akhurst B, Croager EJ, Farley-Roche CA, et al. A modified choline-deficient, ethionine-supplemented diet protocol effectively induces oval cells in mouse liver. Hepatology. 2001 Sep;34(3):519–522. doi: 10.1053/jhep.2001.26751. [DOI] [PubMed] [Google Scholar]
- 86.Roskams TA, Theise ND, Balabaud C, et al. Nomenclature of the finer branches of the biliary tree: canals, ductules, and ductular reactions in human livers. Hepatology. 2004 Jun;39(6):1739–1745. doi: 10.1002/hep.20130. [DOI] [PubMed] [Google Scholar]
- 87.Miyajima A, Tanaka M, Itoh T. Stem/progenitor cells in liver development, homeostasis, regeneration, and reprogramming. Cell Stem Cell. 2014 May 1;14(5):561–574. doi: 10.1016/j.stem.2014.04.010. [DOI] [PubMed] [Google Scholar]
- 88.Van Hul NK, Abarca-Quinones J, Sempoux C, Horsmans Y, Leclercq IA. Relation between liver progenitor cell expansion and extracellular matrix deposition in a CDE-induced murine model of chronic liver injury. Hepatology. 2009 May;49(5):1625–1635. doi: 10.1002/hep.22820. [DOI] [PubMed] [Google Scholar]
- 89.Lorenzini S, Bird TG, Boulter L, et al. Characterisation of a stereotypical cellular and extracellular adult liver progenitor cell niche in rodents and diseased human liver. Gut. 2010 May;59(5):645–654. doi: 10.1136/gut.2009.182345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Kuramitsu K, Sverdlov DY, Liu SB, et al. Failure of fibrotic liver regeneration in mice is linked to a severe fibrogenic response driven by hepatic progenitor cell activation. Am J Pathol. 2013 Jul;183(1):182–194. doi: 10.1016/j.ajpath.2013.03.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Hayner NT, Braun L, Yaswen P, Brooks M, Fausto N. Isozyme profiles of oval cells, parenchymal cells, and biliary cells isolated by centrifugal elutriation from normal and preneoplastic livers. Cancer Res. 1984 Jan;44(1):332–338. [PubMed] [Google Scholar]
- 92.Yaswen P, Hayner NT, Fausto N. Isolation of oval cells by centrifugal elutriation and comparison with other cell types purified from normal and preneoplastic livers. Cancer Res. 1984 Jan;44(1):324–331. [PubMed] [Google Scholar]
- 93.Sirica AE, Mathis GA, Sano N, Elmore LW. Isolation, culture, and transplantation of intrahepatic biliary epithelial cells and oval cells. Pathobiology. 1990;58(1):44–64. doi: 10.1159/000163564. [DOI] [PubMed] [Google Scholar]
- 94.Theise ND, Saxena R, Portmann BC, et al. The canals of Hering and hepatic stem cells in humans. Hepatology. 1999 Dec;30(6):1425–1433. doi: 10.1002/hep.510300614. [DOI] [PubMed] [Google Scholar]
- 95.Lanzoni G, Cardinale V, Carpino G. The Hepatic, Biliary, and Pancreatic Network of Stem/Progenitor Cell Niches in Humans: A New Reference Frame for Disease and Regeneration. Hepatology. 2016 Jul;64(1):277–286. doi: 10.1002/hep.28326. [DOI] [PubMed] [Google Scholar]
- 96.Wang B, Zhao LD, Fish M, Logan CY, Nusse R. Self-renewing diploid Axin2(+) cells fuel homeostatic renewal of the liver. Nature. 2015 Aug 13;524(7564):180–+. doi: 10.1038/nature14863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Kopp JL, Grompe M, Sander M. Stem cells versus plasticity in liver and pancreas regeneration. Nature Cell Biology. 2016 Mar;18(3):238–245. doi: 10.1038/ncb3309. [DOI] [PubMed] [Google Scholar]
- 98.Reid LM. Stem/Progenitor Cells and Reprogramming (Plasticity) Mechanisms in Liver, Biliary Tree, and Pancreas. Hepatology. 2016 Jul;64(1):4–7. doi: 10.1002/hep.28606. [DOI] [PubMed] [Google Scholar]
- 99.Huch M, Dolle L. The plastic cellular states of liver cells: Are EpCAM and Lgr5 fit for purpose? Hepatology. 2016 Aug;64(2):652–662. doi: 10.1002/hep.28469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Sackett SD, Li Z, Hurtt R, et al. Foxl1 is a marker of bipotential hepatic progenitor cells in mice. Hepatology. 2009 Mar;49(3):920–929. doi: 10.1002/hep.22705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Furuyama K, Kawaguchi Y, Akiyama H, et al. Continuous cell supply from a Sox9-expressing progenitor zone in adult liver, exocrine pancreas and intestine. Nat Genet. 2011 Jan;43(1):34–41. doi: 10.1038/ng.722. [DOI] [PubMed] [Google Scholar]
- 102.Malato Y, Naqvi S, Schurmann N, et al. Fate tracing of mature hepatocytes in mouse liver homeostasis and regeneration. J Clin Invest. 2011 Dec;121(12):4850–4860. doi: 10.1172/JCI59261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Huch M, Dorrell C, Boj SF, et al. In vitro expansion of single Lgr5+ liver stem cells induced by Wnt-driven regeneration. Nature. 2013 Feb 14;494(7436):247–250. doi: 10.1038/nature11826. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Rodrigo-Torres D, Affo S, Coll M, et al. The biliary epithelium gives rise to liver progenitor cells. Hepatology. 2014 Oct;60(4):1367–1377. doi: 10.1002/hep.27078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Schaub JR, Malato Y, Gormond C, Willenbring H. Evidence against a stem cell origin of new hepatocytes in a common mouse model of chronic liver injury. Cell Rep. 2014 Aug 21;8(4):933–939. doi: 10.1016/j.celrep.2014.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Tarlow BD, Finegold MJ, Grompe M. Clonal tracing of Sox9+ liver progenitors in mouse oval cell injury. Hepatology. 2014 Jul;60(1):278–289. doi: 10.1002/hep.27084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Yanger K, Knigin D, Zong Y, et al. Adult hepatocytes are generated by self-duplication rather than stem cell differentiation. Cell Stem Cell. 2014 Sep 4;15(3):340–349. doi: 10.1016/j.stem.2014.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Lu WY, Bird TG, Boulter L, et al. Hepatic progenitor cells of biliary origin with liver repopulation capacity. Nat Cell Biol. 2015 Aug;17(8):971–983. doi: 10.1038/ncb3203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Kamimoto K, Kaneko K, Kok CY, Okada H, Miyajima A, Itoh T. Heterogeneity and stochastic growth regulation of biliary epithelial cells dictate dynamic epithelial tissue remodeling. Elife. 2016;5 doi: 10.7554/eLife.15034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Lu WY, Bird TG, Boulter L, et al. Hepatic progenitor cells of biliary origin with liver repopulation capacity. Nature Cell Biology. 2015 Aug;17(8):971–U943. doi: 10.1038/ncb3203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 111.Lemaigre FP. Determining the fate of hepatic cells by lineage tracing: Facts and pitfalls. Hepatology. 2015 Jun;61(6):2100–2103. doi: 10.1002/hep.27659. [DOI] [PubMed] [Google Scholar]
- 112.Reid LM. Paradoxes in studies of liver regeneration: Relevance of the parable of the blind men and the elephant. Hepatology. 2015 Aug;62(2):330–333. doi: 10.1002/hep.27917. [DOI] [PubMed] [Google Scholar]
- 113.Takayama K, Mitani S, Nagamoto Y, et al. Laminin 411 and 511 promote the cholangiocyte differentiation of human induced pluripotent stem cells. Biochem Biophys Res Commun. 2016 May 20;474(1):91–96. doi: 10.1016/j.bbrc.2016.04.075. [DOI] [PubMed] [Google Scholar]
- 114.Clevers H. Modeling Development and Disease with Organoids. Cell. 2016 Jun 16;165(7):1586–1597. doi: 10.1016/j.cell.2016.05.082. [DOI] [PubMed] [Google Scholar]
- 115.Lancaster MA, Knoblich JA. Organogenesis in a dish: modeling development and disease using organoid technologies. Science. 2014 Jul 18;345(6194):1247125. doi: 10.1126/science.1247125. [DOI] [PubMed] [Google Scholar]
- 116.Dedhia PH, Bertaux-Skeirik N, Zavros Y, Spence JR. Organoid Models of Human Gastrointestinal Development and Disease. Gastroenterology. 2016 May;150(5):1098–1112. doi: 10.1053/j.gastro.2015.12.042. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Huch M, Gehart H, van Boxtel R, et al. Long-term culture of genome-stable bipotent stem cells from adult human liver. Cell. 2015 Jan 15;160(1–2):299–312. doi: 10.1016/j.cell.2014.11.050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Takebe T, Sekine K, Enomura M, et al. Vascularized and functional human liver from an iPSC-derived organ bud transplant. Nature. 2013 Jul 25;499(7459):481–484. doi: 10.1038/nature12271. [DOI] [PubMed] [Google Scholar]
- 119.Michalopoulos GK, Bowen WC, Mule K, Stolz DB. Histological organization in hepatocyte organoid cultures. Am J Pathol. 2001 Nov;159(5):1877–1887. doi: 10.1016/S0002-9440(10)63034-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Shin S, Walton G, Aoki R, et al. Foxl1-Cre-marked adult hepatic progenitors have clonogenic and bilineage differentiation potential. Genes Dev. 2011 Jun 1;25(11):1185–1192. doi: 10.1101/gad.2027811. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Tanimizu N, Miyajima A, Mostov KE. Liver progenitor cells develop cholangiocyte-type epithelial polarity in three-dimensional culture. Mol Biol Cell. 2007 Apr;18(4):1472–1479. doi: 10.1091/mbc.E06-09-0848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 122.Kido T, Koui Y, Suzuki K, et al. CPM Is a Useful Cell Surface Marker to Isolate Expandable Bi-Potential Liver Progenitor Cells Derived from Human iPS Cells. Stem Cell Reports. 2015 Oct 13;5(4):508–515. doi: 10.1016/j.stemcr.2015.08.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Lugli N, Kamileri I, Keogh A, et al. R-spondin 1 and noggin facilitate expansion of resident stem cells from non-damaged gallbladders. EMBO Rep. 2016 May;17(5):769–779. doi: 10.15252/embr.201642169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Yu B, He ZY, You P, et al. Reprogramming fibroblasts into bipotential hepatic stem cells by defined factors. Cell stem cell. 2013 Sep 5;13(3):328–340. doi: 10.1016/j.stem.2013.06.017. [DOI] [PubMed] [Google Scholar]