

Abstract
Homocysteine (Hcy) and glutathione (GSH) are crucial reduction-oxidation mediators. The underlying mechanisms governing the effects of Hcy on GSH generation in the progression of alcoholic liver disease has so far received little attention. The present study hypothesized that the antioxidant transcriptional factor nuclear factor (erythroid-derived 2)-like 2 (Nrf2) may participate in Hcy-mediated regulation of GSH production in HepG2 human liver cancer cells. MTT assay was used to study the cytotoxicity of homocysteine, western blot analysis and immunofluorescence staining were used to determine the effect of Hcy on Nrf2 expression. Our data demonstrated that HepG2 cells exposed to exogenous levels of Hcy (0–100 µM) exhibited elevated GSH levels in a concentration-dependent manner. Furthermore, 4-hydroxynonenal (4-HNE)-induced cell injury was attenuated by Hcy; however, this protective effect was blocked by the GSH-production inhibitor buthionine sulfoximine. Hcy treatment was able to induce Nrf2 protein expression in HepG2 cells. Treatment with the Nrf2 activator tert-butylhydroquinone (0–100 µM) increased GSH expression in a concentration-dependent manner; however, Nrf2-siRNA abolished the Hcy-induced increase in GSH expression and cellular protection in 4-HNE-stressed HepG2 cells. In conclusion, the antioxidant transcriptional factor Nrf2 was demonstrated to mediate the Hcy-induced increase in GSH expression levels and cellular protection in HepG2 cells.
Keywords: nuclear factor (erythroid-derived 2)-like 2, homocysteine, glutathione, 4-hydroxynonenal, oxidative stress
Introduction
Homocysteine (Hcy) is a sulfur-containing, non-protein amino acid that is derived from methionine metabolism (1). An imbalance in Hcy metabolism is considered an independent risk factor for many disorders, such as physical impairment (2), atherosclerosis (3,4), neurodegenerative diseases (5) and alcoholic liver disease (ALD) (6,7). Alcoholic hyperhomocysteinemia (HHcy) is often seen in rodents and humans with during the development of alcoholic steatohepatitis (8). In contrast with the adverse effects of Hcy in the progression of ALD, Hcy may serve a positive role in cellular antioxidant reactions as an important producer of cellular total glutathione (GSH) (9,10). This metabolic link between Hcy and GSH occurs by the transsulfuration of Hcy to cysteine, a precursor for GSH (11,12). GSH performs a crucial role in maintaining hepatic reduction-oxidation (redox) balance in biological systems (13). However, the underlying molecular mechanism of the positive role that Hcy serves in the progression of ALD has not been clearly elucidated.
The nuclear factor (erythroid-derived 2)-like 2 (Nrf2) functions as a master regulator of the antioxidant response by regulating the expression of cytoprotective genes, including genes involved in the antioxidant GSH pathway (14–16). Notably, a Hcy-activated Nrf2-antioxidant reaction has been demonstrated in mouse macrophage and human hepatoma cell lines (6,9,10). Therefore, the present study hypothesized that Nrf2 may function to mediate Hcy-regulated GSH production in hepatocytes, and that the hepatic Hcy-Nrf2-GSH pathway may serve a protective role in the progression of ALD. Cellular viability and intracellular GSH expression levels were assayed to investigate the antioxidant effects of Hcy and Nrf2 on HepG2 cell injury induced by the hepatic lipid-peroxidation product 4-hydroxynonenal (4-HNE). Hcy-activated Nrf2 expression was demonstrated by western blot analysis and immunofluorescence staining assay. Further research into this area will provide deeper insights into the association between Hcy and its potentially protective roles in the progression of ALD.
Materials and methods
Cell culture conditions and MTT assay
The HepG2 human hepatocarcinoma cell line was purchased from the American Type Culture Collection (Manassas, VA, USA) and maintained in Dulbecco's modified Eagle's medium (DMEM) containing 10% fetal bovine serum (FBS; both from HyClone; GE Healthcare Life Sciences, Logan, UT, USA) at 37°C in a 5% CO2 atmosphere.
Immunofluorescence staining
HepG2 cells were grown on 6-well cover slips at a density of 3×103 cells/well, and treated with Hcy (0, 10, 20, 50 or 100 µM) for 24 h at 37°C. Adherent cells were fixed to slides with paraformaldehyde for 15 min at 37°C and incubated with 0.5% Trion-X-100 for 10 min at 37°C. Nonspecific binding was blocked by 3% BSA for 30 min at 37°C as previously described (17). Immunostaining was performed with an anti-Nrf2 antibody overnight at 4°C (1:300; bs-1074R, Rabbit polyclonal IgG, BIOSS, Beijing, China), followed by incubation with a cyanine-3 (Cy3)-conjugated secondary antibody for 2 h at 37°C (1:200. P0183, Goat Anti-Rabbit IgG, Beyotime Institute of Biotechnology, Shanghai, China). Nuclear staining was performed by incubating with DAPI (1:100) for 10 min at 37°C. Increased Nrf2 expression was observed under an inverted microscope (Nikon Corporation, Tokyo, Japan). Images were processed using Image ProPlus 6.0 (Media Cybernetics, Inc., Rockville, MD, USA).
Knockdown of Nrf2 by siRNA
Nrf2 expression was silenced by Nrf2-siRNA (Suzhou GenePharma LLC). A non-targeted control siRNA (NC) was used as a negative control. The siRNA sense sequences were as follows: siNrf2, 5′-GCACCUUAUAUCUCGAAGUTT-3′; NC, 5′-UUCUCCGAACGUGUCACGUTT-3′. Briefly, HepG2 cells were grown to 50–70% confluence. A total of 0.27 µg (1 µl) siRNA, 10 µl INTERFERin Gene siRNA Transfection Reagent (Suzhou GenePharma LLC) and 400 µl serum-free Opti-MEM-1 medium (Gibco; Thermo Fisher Scientific, Inc., Waltham, MD, USA) were combined and added to the cells, which were incubated at room temperature for 10 min, followed by the subsequent addition of the siRNA/Transfection Reagent mixture directly onto the cells with 4 ml serum-free Opti-MEM-1 medium (final siRNA concentration, 1 nM). Following incubation for 4 h at 37°C, the media was replaced with DMEM containing 10% FBS. Total RNA was extracted and reverse transcribed to cDNA using a Golden 1st cDNA Synthesis kit (HaiGene Bio Inc., Harbin, China). cDNA was stored at −80°C and transfection efficiency was later validated by reverse transcription-quantitative polymerase chain reaction (RT-qPCR).
MTT assay
HepG2 cells were seeded at a density of 2×103 cells/well in two 96-well plates; one plate was treated with 70 µM 4-HNE (Cayman Chemical Company, Ann Arbor, MI, USA), 200 µM buthionine sulfoximine (BSO; Shanghai Aladdin Bio-Chem Technology Co., Ltd., Shanghai, China), 100 µM Hcy (Sigma-Aldrich; Merck KGaA, Darmstadt, Germany), 4-HNE+Hcy, BSO+Hcy, BSO+4-HNE and BSO+4-HNE+Hcy for 24 h at 37°C; the second plate was transfected with 1 nM negative control and Nrf2-targeted small interfering RNA (siRNA; Suzhou GenePharma LLC, Suzhou, China), followed by addition of 100 µM Hcy, 70 µM 4-HNE or 4-HNE+Hcy respectively, for 24 h at 37°C. The following day, 20 µl MTT (5 mg/ml), the yellow mitochondrial dye, was added to the cells, and the plates were incubated at 37°C for an additional 4 h. The culture medium was discarded, and any formazan crystals that may have formed were dissolved in 150 µl DMSO. Absorbance was measured at 570 nm using a SPECTRA Max 190 (Molecular Devices, LLC, Sunnyvale, CA, USA).
Measurement of reduced GSH and oxidized GSH levels in HepG2 cells
HepG2 cells (3×105) were treated with increasing concentrations of Hcy (0–500 µM) and tert-butylhydroquinone (t-BHQ; 0–100 µM, Sigma-Aldrich; Merck KGaA) and 200 µM BSO for 24 h, respectively, or transfected with 1 nM Nrf2- or NC-siRNA for 4 h at 37°C. The media was subsequently replaced with DMEM containing 10% FBS with or without Hcy for 9 h. Cells were trypsinized and collected by centrifugation at 170 × g for 5 min at room temperature, re-suspended in protein removal reagent M (Beyotime Institute of Biotechnology) and immediately vortex mixed for 10s. The samples were frozen rapidly with liquid nitrogen and thawed at 37°C, this was repeated twice, and then the samples were incubated in an ice-bath for 5 min. The resulting supernatant was collected by centrifugation at 10,000 × g for 10 min, and analyzed using the sulfhydryl reagent 5,5′-dithiobis (2-nitrobenzoic acid), which forms the yellow derivative 5′-thio-2-nitrobenzoic acid (TNB), and is measured at 412 nm using a SPECTRA Max 190 (Molecular Devices, LLC). Levels of reduced and oxidized GSH were determined using a GSH Assay kit (Beyotime Institute of Biotechnology), according to the manufacturers protocol (18).
Western blot analysis
HepG2 cells were treated with Hcy (0–100 µM) for 24 h at 37°C. Cells were homogenized in RIPA lysis buffer (Beyotime Institute of Biotechnology) containing leupeptin. Protein concentrations were quantified using a Bicinchoninic Acid Protein Assay kit (Beyotime Institute of Biotechnology), according to the manufacturer's protocol. Cellular protein samples (30 µg) were separated by 10% SDS-PAGE as previously described (19). Proteins were transferred onto nitrocellulose membranes and the membranes were incubated with an anti-Nrf2 polyclonal antibody (1:500; bs-1074R, BIOSS, Beijing, China) or an anti-β-actin monoclonal antibody (1:3,000; TA-09; Zhongshan Golden Bridge Biotechnology, Beijing, China) overnight at 4°C, followed by horseradish peroxidase-conjugated goat anti-rabbit immunoglobulin G secondary antibody (1: 5,000; ZB-2301, Zhongshan Golden Bridge Biotechnology). Protein bands were visualized by enhanced chemiluminescence (HaiGene Bio Inc.). Densitometric analysis of band intensity was calculated using Quantity One software, version v4.62 (Bio-Rad Laboratories, Inc., Hercules, CA, USA).
Relative mRNA expression quantification by RT-qPCR
Total RNA was extracted from cultured HepG2 cells (2×105 cells) using TRIzol Reagent (HaiGene Bio Inc.), according to the manufacturer's protocol (9), and subsequently reverse transcribed to cDNA using a Golden 1st cDNA Synthesis Kit (HaiGene Bio Inc.). cDNA samples were amplified using a DNA thermal cycler (Thermo Fisher Scientific, Inc.). qPCR reactions were set up by mixing 10X SYBR Green PCR Master Mix (Roche Diagnostics GmbH, Mannheim, Germany) with 2 µl 100 fold diluted cDNA template, 0.6 µl each of forward and reverse primers (0.3 µM) and 6.8 µl PCR-grade water in a 20 µl reaction volume. The reaction mixtures were amplified using an ABI 7300 Sequence Detection System (Applied Biosystems; Thermo Fisher Scientific, Inc.) with an initial denaturation step at 95°C for 10 min, followed by 40 cycles of: denaturation at 95°C for 15 sec; annealing at 58°C for 20 sec; extension at 72°C for 30 sec; and melting curve analysis. The primer sequences were as follows: 18S, forward 5′-AACTTTCGATGGTAGTCGCCG−3′, reverse 5′-CCTTGGATGTGGTAGCCGTTT-3′; Nrf2, forward 5′-GCACCUUAUAUCUCGAAGUTT-3′, reverse 5′-ACUUCGAGAUAUAAGGUGCTT-3′; GCLc, forward 5′-ATGTGGACACCCGATGCAGTATT-3′; reverse 5′-TGTCTTGCTTGTAGTCAGGATGGTTT-3′. Primers were designed using the GenBank database (https://www.ncbi.nlm.nih.gov/genbank). Nrf2 mRNA and GCLc mRNA expression were measured by RT-qPCR and normalized to 18s. Data was analyzed using Applied Biosystems SDS V1.4 software (Thermo Fisher Scientific, Inc.) using the 2−ΔΔCq method, where ΔΔCq=ΔCq target-ΔCq internal control (20).
Statistical analysis
Statistical analysis was performed using one-way analysis of variance followed by Newman-keuls test. Analyses were performed using OriginPro7.5 software (OriginLab, Northampton, MA, USA) and Graph Pad Prism version 5 (Graph Pad Software Inc., La Jolla, CA, USA). Data were presented as the mean ± standard error of the mean. Values of P<0.05 were considered to indicate a statistically significant difference. All experiments were performed at least three times and representative results were presented.
Results
Hcy treatment increases total GSH expression levels in HepG2 cells
Intracellular levels of the antioxidant thiol GSH were evaluated using the GSH Assay kit. In response to exogenous Hcy (10, 20, 100 or 500 µM) treatment, the levels of intracellular total GSH (Fig. 1A), oxidized GSH (Fig. 1B) and reduced GSH (Fig. 1C) were increased in a concentration-dependent manner, with a peak at 100 µM Hcy. Therefore, exogenous Hcy was administered at a concentration of 100 µM for the following tests.
Figure 1.
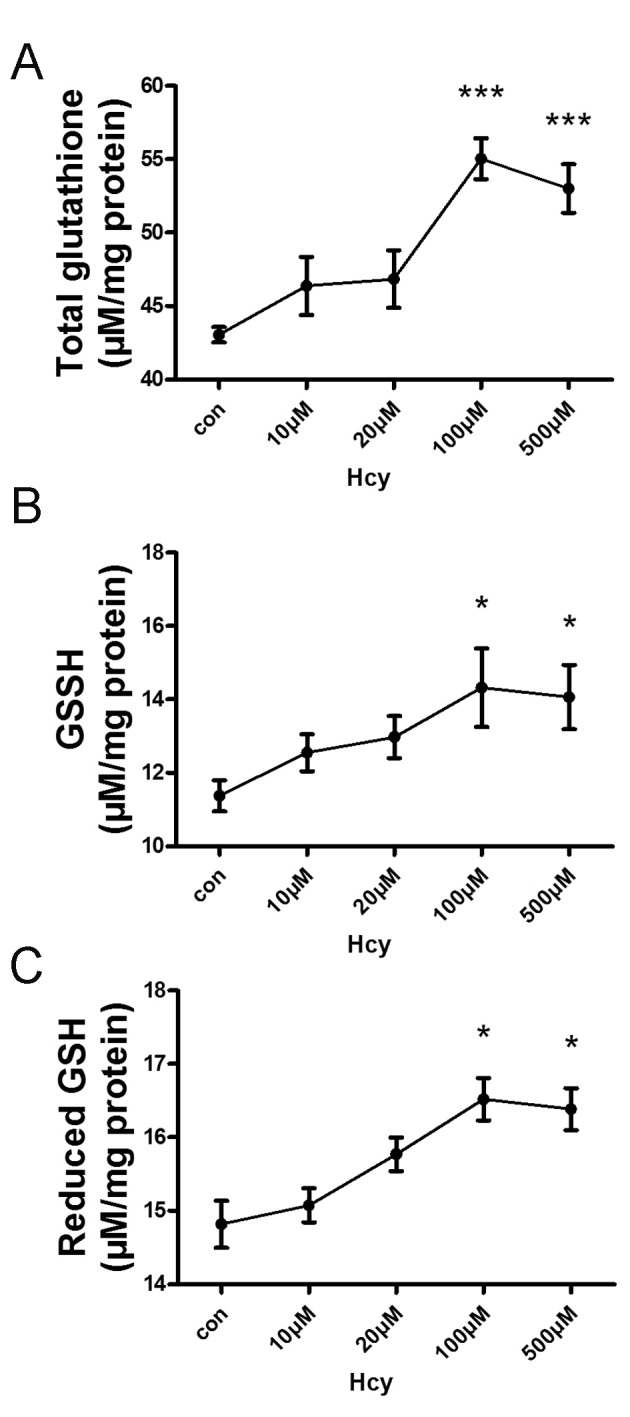
GSH levels in response to Hcy treatment in HepG2 cells. HepG2 cells were treated with Hcy (0–500 µM) for 24 h. The levels of (A) total GSH, (B) oxidized GSH and (C) reduced GSH were determined using a GSH Assay kit. Values are presented as the mean ± standard error of the mean (n=3). *P<0.05, ***P<0.001; GSH, total glutathione; Hcy, homocysteine.
Hcy protects against oxidative stress-induced cell injury in HepG2 cells
HepG2 cells were pretreated with the specific GSH production inhibitor BSO (200 µM) for 2 h, and subsequently treated or co-treated with Hcy (100 µM) for 24 h. Hcy treatment led to elevated GSH content compared with the untreated control group (n=6, P<0.05). However, BSO co-treatment significantly abolished the Hcy-induced GSH expression in HepG2 cells, compared with the Hcy-treated group (Fig. 2A; n=6; P<0.05). HepG2 cells were treated with several combinations, as follows: 4-HNE (70 µM)+Hcy (100 µM); BSO (200 µM)+4-HNE; BSO+Hcy; BSO+4-HNE+Hcy all together. MTT assay indicated that treatment with the lipid peroxidation product 4-HNE decreased cell viability, and no cellular toxicity was observed in HepG2 cells at 10 µM Hcy. However, Hcy co-treatment attenuated 4-HNE-induced cell injury, compared with the 4-HNE-treated group (Fig. 2B; n=6; P<0.05). Notably, BSO pretreatment significantly weakened the protective effect of Hcy on 4-HNE-induced cell injury in HepG2 cells, compared with the 4-HNE+Hcy group (n=6; P<0.05). Conversely, BSO pretreatment had no effect on viability in Hcy- or 4-HNE-treated cells (Fig. 2B; n=6; P>0.05).
Figure 2.

Hcy treatment protects HepG2 cells from 4-HNE-induced cell injury by increasing GSH expression levels. (A) Treatment with the GSH biosynthesis inhibitor BSO abolished Hcy-induced GSH expression in HepG2 cells. HepG2 cells were pretreated with or without 200 µM BSO for 2 h, followed by treatment with 100 µM Hcy for 24 h. Total GSH levels were determined using a GSH Assay kit. (B) Hcy treatment reduced 4-HNE-induced cell injury, and co-treatment with BSO weakened the protective effect of Hcy on 4-HNE-induced cell injury in HepG2 cells. HepG2 cells were pretreated with or without BSO for 2 h, followed by treatment with 100 µM Hcy and/or 70 µM 4-HNE for 24 h. The viability of HepG2 was detected by MTT assay. Values are presented as the mean ± standard error of the mean (n=6). *P<0.05 and ***P<0.001 vs. control or Hcy group; ###P<0.001 vs. 4-HNE group; &&&P<0.001 vs. 4-HNE+Hcy group. 4-HNE, 4-hydroxynonenal; BSO, buthionine sulfoximine; Con, control; GSH, total glutathione; Hcy, homocysteine.
Hcy treatment induces Nrf2 protein expression and promotes nuclear accumulation in HepG2 cells
HepG2 cells were treated with Hcy (0–100 µM) for 24 h. Western blot analysis revealed that Hcy exposure induced Nrf2 protein expression in a concentration-dependent manner (Fig. 3A). Immunofluorescence analysis with an anti-Nrf2 antibody revealed that the protein expression of Nrf2 increased with the Hcy concentration (Fig. 3B). In addition, Hcy (0–100 µM) treatment promoted the expression of glutamate-cysteine ligase catalytic subunit (GCLc), a downstream target gene of Nrf2 (Fig. 3C).
Figure 3.

Hcy induces Nrf2 protein expression. HepG2 cells were exposed to Hcy (0–100 µM) for 24 h. (A) Hcy treatment induced Nrf2 protein expression in a concentration-dependent manner. Nrf2 expression was assessed by western blot analysis; a representative western blot and semi-quantification are presented. (B) Hcy treatment promoted Nrf2 protein nuclear accumulation in HepG2 cells. Scale bar, 50 µm. (C) Hcy (0–100 µM) treatment increased GCLc expression levels in a concentration-dependent manner. Values are presented as the mean ± standard error of the mean (n=3). *P<0.05 and **P<0.01 vs. untreated (0 µM) control. GCLc, glutamate cysteine ligase catalytic subunit; Hcy, homocysteine; Nrf2, nuclear factor erythroid-derived 2-like 2.
Nrf2 mediates Hcy-induced GSH expression and cell protection
To explore whether Nrf2 participates in the Hcy-induced increase in GSH expression in HepG2 cells, cells were treated with the Nrf2 activator t-BHQ (0, 10, 20, 50 or 100 µM) for 24 h. Cells exposed to t-BHQ exhibited an increase in GSH expression levels in a concentration-dependent manner (Fig. 4A). Additionally, Nrf2 gene expression was knocked down in HepG2 cells by siRNA, which resulted in the decreased expression of Nrf2 mRNA levels by ~50% (Fig. 4B; n=3; P<0.001 vs. NC treated cells). Hcy+NC treatment for 24 h slightly elevated the intracellular GSH content in comparison with the NC group. This small increase may be attributed to invasion of the exogenous siRNA into the serum-starved cells, which may have resulted in decreasing the cell sensitivity to the following Hcy treatment. Nrf2-siRNA exposure resulted in decreased GSH expression levels, compared with the NC-treated HepG2 cells. Furthermore, Hcy-induced GSH expression in HepG2 cells was also suppressed by Nrf2 siRNA, compared with the NC+Hcy group (Fig. 4C; n=3; P<0.05). Analysis of cell viability with MTT indicated that Nrf2-siRNA pretreatment reduced the protective role of Hcy in limiting 4-HNE-induced cell injury in HepG2 cells (Fig. 4D; n=3; P<0.05). In brief, Nrf2 was observed to mediate Hcy-induced GSH expression and the protective effect of Hcy on cell viability.
Figure 4.

Nrf2 mediates Hcy-induced GSH expression and cellular protection in 4-HNE-stressed HepG2 cells. (A) t-BHQ treatment increased total GSH expression levels in a concentration-dependent manner in HepG2 cells. HepG2 cells were treated with t-BHQ (0–100 µM) for 24 h (n=3). (B) Nrf2 mRNA expression was downregulated by ~50% following transfection with Nrf2-siRNA. Nrf2 siRNA transfection was performed for 4 h; the transfection media was subsequently removed and replaced with medium containing 10% fetal bovine serum for 9 h (n=3). (C) Nrf2-siRNA transfection reduced Hcy-induced total GSH elevation. HepG2 cells were transfected with NC-siRNA or Nrf2-siRNA for 4 h, then co-treated with Hcy for 9 h (n=3). (D) Nrf2 siRNA attenuated the protective role of Hcy in 4-HNE-induced cell injury in HepG2 cells. HepG2 cells were pretreated with 1 nM NC siRNA or 1 nM Nrf2 siRNA for 4 h, followed by Hcy (100 µM) and/or 4-HNE (70 µM) for 24 h. Cellular viability was assessed by MTT Assay (n=6). Values are presented as the mean ± standard error of the mean. *P<0.05, **P<0.01 and ***P<0.001 vs. control group; $$$P<0.001 vs. NC group; ###P<0.001 vs. NC+4-HNE-treated cells. 4-HNE, 4-hydroxynonenal; Con, control; GSH, glutathione; Hcy, homocysteine; NC, non-targeted control siRNA; Nrf2, nuclear factor erythroid-derived 2-like 2; siRNA, small interfering RNA; t-BHQ, tert-butylhydroquinone
Discussion
The present study is the first, to the best of our knowledge, to demonstrate the protective role of exogenous Hcy in 4-HNE-induced cell injury in HepG2 cells. Furthermore, the present study investigated the underlying molecular mechanism of this process, which involves an increase in GSH expression mediated by the antioxidant transcriptional factor Nrf2.
Previous studies indicated that circulating and hepatic Hcy levels were elevated in alcohol-fed mice, and aberrant Hcy metabolism elicited oxidative stress and hepatic GSH depletion, which has been demonstrated in both experimental animals and in humans (21,22). One study indicated that Hcy treatment was able to trigger acute GSH depletion in HepG2 cells within 45 min (6). By contrast, increased GSH expression that was induced by Hcy has also been demonstrated in HepG2 cells (9) and macrophages (10); this may be due to increased activity via the transsulfuration pathway or de novo synthesis of GSH. Consistent with these reports, the present study demonstrated that 24 h Hcy treatment increased the levels of intracellular GSH expression in a concentration-dependent manner. GSH is a crucial mediator in various antioxidant responses; therefore, the data suggested that the GSH metabolism-related amino acid Hcy may be involved in the antioxidant process, and may protect against oxidative stress-induced cell injury in HepG2 cells.
HHcy is a risk factor for cardiovascular disease and ALD, and patients with HHcy may develop hepatic steatosis and fibrosis (1,3,23,24). However, the balance of Hcy metabolism and its role in the progression of these diseases is not clearly understood. In contrast to the adverse effects of elevated Hcy in these pathological conditions, the present study indicated that exogenous Hcy treatment may protect HepG2 cells against cell injury induced by the lipid peroxidation product 4-HNE. Notably, a previous in vitro investigation indicated that exogenous Hcy exposure had no effect on HepG2 cell viability (6). Consistent with this observation, the present study demonstrated that Hcy exposure did not affect HepG2 cell viability in non-stress conditions. Also of note, the GSH biosynthesis inhibitor BSO significantly lowered the Hcy-induced increase in GSH expression levels and the Hcy-mediated cellular protection against 4-HNE. Collectively, these data indicated that Hcy protects against 4-HNE-indued cell injury in HepG2 cells, via increased expression of intracellular GSH levels.
Nrf2 is critical for maintaining the GSH redox state and protecting cells against oxidative stress (19). High levels of Hcy treatment suppressed Nrf2-dependent antioxidant protection, but lower levels of Hcy (100 µM) upregulated the expression of Nrf2 in U373 astroglial cells (25). In addition, HepG2 cells exposed to 50 µM Hcy exhibited an increase in Nrf2 protein expression in a time-dependent manner (9). In the present study, HepG2 cells exposed to Hcy (0–100 µM) for 24 h exhibited increased Nrf2 protein expression in a concentration-dependent manner. These results indicated that nontoxic concentrations of Hcy may activate the antioxidant transcriptional factor Nrf2 in HepG2 cells.
Previous studies have reported that Nrf2 affects GSH homeostasis by regulating de novo synthesis and salvage pathways, to protect cells against oxidative stress (15,16,25,26). Consistent with these reports, the present study demonstrated that the Nrf2 activator t-BHQ increased total intracellular GSH levels in a concentration-dependent manner. By contrast, Nrf2-siRNA treatment inhibited the Hcy-induced GSH elevation. Furthermore, Nrf2-siRNA treatment reduced the ability of Hcy to protect against 4-HNE-induced cell injury. Collectively, these data suggested that Nrf2 mediates Hcy-induced GSH expression and cellular protection in 4-HNE-treated HepG2 cells.
In conclusion, the present study demonstrated that Hcy treatment may protect HepG2 cells against cell injury induced by the lipid peroxidation product 4-HNE, by inducing the expression of GSH, and this was mediated by the antioxidant transcriptional factor Nrf2. Hcy has been regarded as an independent risk factor for the progression of arteriosclerosis, hypertension or ALD (3,24,27,28). The concentration of Hcy (<100 µM) applied in the present study represented mild - moderate HHcy, which is usually a characteristic symptom in the early stages of these diseases; an initial elevation of Hcy may serve a compensatory protective effect on vulnerable cells. Therefore, the results indicated that hepatic Hcy elevation may be involved in an antioxidant mechanism, particularly in the early stage of these pathological conditions.
Acknowledgements
This work was supported by The National Natural Science Foundation (grant no. 81370523), The Postdoctoral Science Foundation Special Project (grant no. 201104420), The China Postdoctoral Science Foundation General Project (grant no. 20100471022) and The Heilongjiang Young Key Academic Staff support program (grant no. 1251G039).
References
- 1.Ganguly P, Alam SF. Role of homocysteine in the development of cardiovascular disease. Nutr J. 2015;14:6. doi: 10.1186/1475-2891-14-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Veeranki S, Tyagi SC. Defective homocysteine metabolism: Potential implications for skeletal muscle malfunction. Int J Mol Sci. 2013;14:15074–15091. doi: 10.3390/ijms140715074. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Zhang D, Fang P, Jiang X, Nelson J, Moore JK, Kruger WD, Berretta RM, Houser SR, Yang X, Wang H. Severe hyperhomocysteinemia promotes bone marrow-derived and resident inflammatory monocyte differentiation and atherosclerosis in LDLr/CBS-deficient mice. Circ Res. 2012;111:37–49. doi: 10.1161/CIRCRESAHA.112.269472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kamat PK, Vacek JC, Kalani A, Tyagi N. Homocysteine induced cerebrovascular dysfunction: A link to alzheimer's disease etiology. Open Neurol J. 2015;9:9–14. doi: 10.2174/1874205X01509010009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sharma M, Tiwari M, Tiwari RK. Hyperhomocysteinemia: Impact on neurodegenerative diseases. Basic Clin Pharmacol Toxicol. 2015;117:287–296. doi: 10.1111/bcpt.12424. [DOI] [PubMed] [Google Scholar]
- 6.Mani M, Golmohammadi T, Khaghani S, Zamani Z, Azadmanesh K, Meshkani R, Pasalar P. Homocysteine induces heme oxygenase-1 expression via transcription factor Nrf2 activation in HepG2 cell. Iran Biomed J. 2013;17:93–100. doi: 10.6091/ibj.1158.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ji C. Mechanisms of alcohol-induced endoplasmic reticulum stress and organ injuries. Biochem Res Int. 2012;2012:216450. doi: 10.1155/2012/216450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Esfandiari F, Medici V, Wong DH, Jose S, Dolatshahi M, Quinlivan E, Dayal S, Lentz SR, Tsukamoto H, Zhang YH, et al. Epigenetic regulation of hepatic endoplasmic reticulum stress pathways in the ethanol-fed cystathionine beta synthase-deficient mouse. Hepatology. 2010;51:932–941. doi: 10.1002/hep.23382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Mani M, Khaghani S, Mohammadi Gol T, Zamani Z, Azadmanesh K, Meshkani R, Pasalar P, Mostafavi E. Activation of Nrf2-antioxidant response element mediated glutamate cysteine ligase expression in hepatoma cell line by homocysteine. Hepat Mon. 2013;13:e8394. doi: 10.5812/hepatmon.8394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Bea F, Hudson FN, Neff-Laford H, White CC, Kavanagh TJ, Kreuzer J, Preusch MR, Blessing E, Katus HA, Rosenfeld ME. Homocysteine stimulates antioxidant response element-mediated expression of glutamate-cysteine ligase in mouse macrophages. Atherosclerosis. 2009;203:105–111. doi: 10.1016/j.atherosclerosis.2008.06.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Lu SC. Regulation of glutathione synthesis. Mol Aspects Med. 2009;30:42–59. doi: 10.1016/j.mam.2008.05.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Lu SC. Glutathione synthesis. Biochim Biophys Acta. 2013;1830:3143–3153. doi: 10.1016/j.bbagen.2012.09.008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Niu LY, Guan YS, Chen YZ, Wu LZ, Tung CH, Yang QZ. A turn-on fluorescent sensor for the discrimination of cystein from homocystein and glutathione. Chem Commun (Camb) 2013;49:1294–1296. doi: 10.1039/c2cc38429a. [DOI] [PubMed] [Google Scholar]
- 14.Soeur J, Eilstein J, Léreaux G, Jones C, Marrot L. Skin resistance to oxidative stress induced by resveratrol: From Nrf2 activation to GSH biosynthesis. Free Radic Biol Med. 2015;78:213–223. doi: 10.1016/j.freeradbiomed.2014.10.510. [DOI] [PubMed] [Google Scholar]
- 15.Bell KF, Fowler JH, Al-Mubarak B, Horsburgh K, Hardingham GE. Activation of Nrf2-regulated glutathione pathway genes by ischemic preconditioning. Oxid Med Cell Longev. 2011;2011:689524. doi: 10.1155/2011/689524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Harvey CJ, Thimmulappa RK, Singh A, Blake DJ, Ling G, Wakabayashi N, Fujii J, Myers A, Biswal S. Nrf2-regulated glutathione recycling independent of biosynthesis is critical for cell survival during oxidative stress. Free Radic Biol Med. 2009;46:443–453. doi: 10.1016/j.freeradbiomed.2008.10.040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ho HK, White CC, Fernandez C, Fausto N, Kavanagh TJ, Nelson SD, Bruschi SA. Nrf2 activation involves an oxidative-stress independent pathway in tetrafluoroethylcysteine-induced cytotoxicity. Toxicol Sci. 2005;86:354–364. doi: 10.1093/toxsci/kfi205. [DOI] [PubMed] [Google Scholar]
- 18.Jayakumar S, Kunwar A, Sandur SK, Pandey BN, Chaubey RC. Differential response of DU145 and PC3 prostate cancer cells to ionizing radiation: Role of reactive oxygen species, GSH and Nrf2 in radiosensitivity. Biochim Biophys Acta. 2014;1840:485–494. doi: 10.1016/j.bbagen.2013.10.006. [DOI] [PubMed] [Google Scholar]
- 19.Chen CC, Chen HL, Hsieh CW, Yang YL, Wung BS. Upregulation of NF-E2-related factor-2-dependent glutathione by carnosol provokes a cytoprotective response and enhances cell survival. Acta Pharmacol Sin. 2011;32:62–69. doi: 10.1038/aps.2010.181. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) Method. Methods. 2001;25:402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 21.Gong Z, Yan S, Zhang P, Huang Y, Wang L. Effects of S-adenosylmethionine on liver methionine metabolism and steatosis with ethanol-induced liver injury in rats. Hepatol Int. 2008;2:346–352. doi: 10.1007/s12072-008-9082-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Paul R, Borah A. The potential physiological crosstalk and interrelationship between two sovereign endogenous amines, melatonin and homocysteine. Life Sci. 2015;139:97–107. doi: 10.1016/j.lfs.2015.07.031. [DOI] [PubMed] [Google Scholar]
- 23.Schalinske KL, Smazal AL. Homocysteine imbalance: A pathological metabolic marker. Adv Nutr. 2012;3:755–762. doi: 10.3945/an.112.002758. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Medici V, Peerson JM, Stabler SP, French SW, Gregory JF, III, Virata MC, Albanese A, Bowlus CL, Devaraj S, Panacek EA, et al. Impaired homocysteine transsulfuration is an indicator of alcoholic liver disease. J Hepatol. 2010;53:551–557. doi: 10.1016/j.jhep.2010.03.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Steele ML, Fuller S, Patel M, Kersaitis C, Ooi L, Münch G. Effect of Nrf2 activators on release of glutathione, cysteinylglycine and homocysteine by human U373 astroglial cells. Redox Biol. 2013;1:441–445. doi: 10.1016/j.redox.2013.08.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gong P, Cederbaum AI. Transcription factor Nrf2 protects HepG2 cells against CYP2E1 plus arachidonic acid-dependent toxicity. J Biol Chem. 2006;281:14573–14579. doi: 10.1074/jbc.M600613200. [DOI] [PubMed] [Google Scholar]
- 27.Pacana T, Cazanave S, Verdianelli A, Patel V, Min HK, Mirshahi F, Quinlivan E, Sanyal AJ. Dysregulated hepatic methionine metabolism drives homocysteine elevation in diet-induced nonalcoholic fatty liver disease. PLoS One. 2015;10:e0136822. doi: 10.1371/journal.pone.0136822. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Zhou Y, Zhao L, Zhang Z, Lu X. Protective effect of enalapril against methionine-enriched diet-induced hypertension: Role of endoplasmic reticulum and oxidative stress. Biomed Res Int. 2015;2015:724876. doi: 10.1155/2015/724876. [DOI] [PMC free article] [PubMed] [Google Scholar]