

Abstract
Pine wilt disease (PWD) is a devastating forest disease present worldwide. In this study we analyzed the effects of the invasion of the pinewood nematode Bursaphelenchus xylophilus, the major pathogen causing PWD, on the endophytic microbiome of adult P. pinaster trees. Wood samples from trees with different degrees of PWD disease were collected at two sites (A and M) in Portugal. Endophytic bacteria were characterized based on directly extracted DNA by fingerprinting and barcoding using the 16S rRNA gene as marker. Furthermore, cultivation-based approaches were used to obtain isolates of the major taxa to study their ecophysiology. The endophytic microbiome from P. pinaster trees differed significantly between the two sampling sites. Main bacterial OTUs belonged to the Proteobacteria (39% (site M) - 97% (site A)), and Firmicutes (0.70% (site A) - 44% (site M)). However, consequences of the invasion with the pathogen were comparable. Interestingly diversity of wood endophytic bacteria increased with the severity of the diseases, with highest diversity levels observed in in the most affected trees. Our results suggest that in the first stages of the disease, the defence mechanisms of plants are repressed by the pathogen, resulting in a colonization of the wood interior by soil microorganisms.
Introduction
Native to North America (the USA and Canada), pine wilt disease (PWD) is one of the most devastating forest diseases in the world. The disease has spread to Asia and to Europe, mainly affecting Japan, China, Korea, Taiwan, Portugal and Spain1. In Portugal, PWD was reported first in 1999 by Mota and coworkers2, in an area south of the Tagus river; after 2008, PWD has spread to the center and north part of the country3. Portuguese forests are mainly composed of several species of pine, eucalyptus and oak trees. The most abundant species of pine, P. pinaster, making up 62.5% of total trees, is a PWD-susceptible tree species3.
Bursaphelenchus xylophilus, the pinewood nematode, has been considered for a long time to be the only known causal agent of the disease4. Once having entered the plant inoculated by the insect vector when feeding on young shoots, the nematode destroys the vascular system of plants5. The nematodes can reproduce quickly in the sapwood under favourable conditions within susceptible pine species, causing wilting and death, sometimes in only a few weeks. In order to make phytosanitary decisions, a classification based on the aspect of the crown and the colour of the leaves was created6. This classification system includes six classes from trees without symptoms (s0) to trees without leaves (sV).
A relationship of mutuality has been suggested between bacteria and B. xylophilus 7 in the development of the disease. The origin of these bacteria, their functional traits as well as which plant compartments they colonize is still under discussion6, 8–12. In this study we addressed the question how the diversity of endophytic bacteria colonizing the wood interior changes as a result of PWD.
Endophytic bacteria have been shown to provide different services to the plants. Besides acting as plant growth promoters13, they can exhibit strong anti-fungal activity, antagonise bacterial pathogens and control plant parasitic nematodes14–17. Consequently changes in the microbiome structure of endophytes might have pronnounced effects for the plant phenotype. Several studies have confirmed the presence of endophytic microbes as part of the Pinus spp. holobiont. In Scots pine, P. sylvestris, the major genera of endophytic bacteria found were Methylobacterium, Pseudomonas 18–20, Bacillus and Paenibacillus 21. Paenibacillus and Bacillus were also found as endophytes of P. contorta 22, 23, while the genera Acetobacter and Gluconacetobacter were part of the foliar endophytic community of P. flexilis 24.
Taking in consideration that the pinewood nematode produces effector proteins, as cellulases and peroxiredoxin4, that modulate the plant response and might reduce the plant defenses, we postulated that the infection with B. xylophilus of pine trees will change the structure of bacterial endophytes. We hypothesize that the diversity of endophytes will increase mainly in heavily infected trees, due to reduced plant defense mechanisms against entering the plant interior. This might overall accelerate the course of the disease. To study these hypotheses, we extracted microbes from the woody part of P. pinaster trees from two regions in Portugal. In both regions, trees with different degree of severity of the disease were present. We used molecular fingerprinting and barcoding methods to describe bacterial diversity based on extracted community DNA and amplified 16S rRNA genes, as well as isolation based approaches to assess the phenotypes of major (cultivable) wood colonizing endophytes.
Results
Bacterial community fingerprinting
The overall diversity of wood colonizing bacterial endophytes was assessed from 43 P. pinaster trees, with differing severity of PWD as well as unaffected healthy trees from two regions in Portugal (Table S1). The DNA used was extracted directly from a microbial fraction, obtained from the core of the woody material obtained from the trunk respectively crown, and amplified 16S rRNA genes were subjected to DGGE based fingerprinting.
Overall, between 30 to 44 bands were obtained per sample, when the bacterial community fingerprints were analyzed (data not shown). After excluding bands related to chloroplast sequences, no significant differences in diversity according to Shannon-Weaver and Simpson 1-D indexes were found when both sites, Avô and Malhada, were compared (H′ = 2.86 and 3.45; D = 0.94 and 0.96, respectively); at both sites for diseased trees of the highest symptom class (sV), a higher diversity was visible (H′ = 2.92 and 3.27; D = 0.94 and 0.96) compared to healthy trees (s0) (H′ = 2.71 and 3.13; D = 0.93 and 0.95) (Table S2). Comparing the presence and absence of individual bands, the clearest differences for bacterial communities were obtained when comparing trees from both sites (paired samples t-test, t 43df = 5.50, p < 0.001) (Fig. 1). Whereas at site A no clear clustering was observed according to the disease symptoms, at site M, mainly the trees with the highest degree of disease outbreak differed from the other trees. Overall DGGE pattern did not change when samples from crown and trunk from the same tree were compared.
Figure 1.
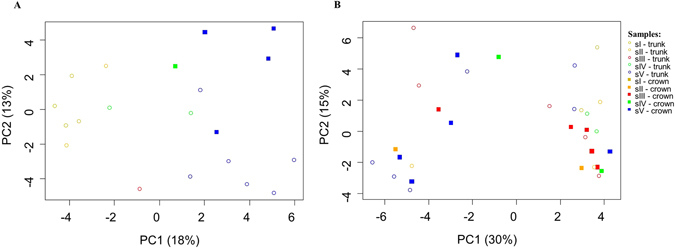
Principal component analysis (PCA) of DGGE fingerprinting of endophytic, wood colonizing bacterial communities from all pine wood tree samples from the two sampling sites obtained after PCR amplification of 16S rRNA gene fragments. The samples grouped according to the different symptomatic physiological classes in Malhada site (A); no clear clustering was observed in Avô site (B).
Bacterial community barcoding
A total of 2,791,770 high quality reads were generated by sequencing 16S rRNA gene based amplicons derived from community DNA from 12 samples covering trees of six different symptomatic physiological classes from the two sampling areas (Table S3). The total number of reads ranged from 55,806 (A-sI) to 208,182 (A-sIV). After filtering and removal of 16S rRNA gene sequences derived from chloroplasts, reads ranged from 24,069 to 176,403 for M-s0 and A-sV, respectively. Thus, for further analysis all reads from all sequenced samples were subsampled to 24,069 reads. The difference observed in read numbers for the individual samples were mainly a result of sequences which were identified as chloroplasts from Pine trees (0.3–62%), which were obviously co-extracted during the preparation procedure. However, even after subsampling, the analysis of rarefaction curves showed for all samples sufficient sequencing depth to assess OTU diversity of early classes of disease (s0-sII) at a level of 97% similarity, while a large increase of OTU diversity, at classes sIV and sV, was observed mainly from trees at site M (Figure S2). Representative sequences from each OTU were phylogenetically classified using the Greengenes database. The number of detected OTUs ranged from 454 (A-sI) to 2,260 (M-sIV).
Interestingly, the highest diversity was found in the wood samples of the most diseased trees both for site A and site M, which confirms the data of the fingerprint analysis of the individual samples. Overall, diversity of OTUs was higher at site M compared to site A (H′ = 5.72 and 4.29; D = 0.97 and 0.95, respectively) but no statistical differences were found between these two sites, (Table S2). Also the alpha diversity measured based on Chao1 (richness) indicated higher diversity at site M compared to site A (Fig. 2). Chao1 based analysis also indicated that trees in different symptomatic stages presented different endophytic community richness, with lower diversity in the first stages of the disease and higher diversity at later stages (sIV and sV).
Figure 2.
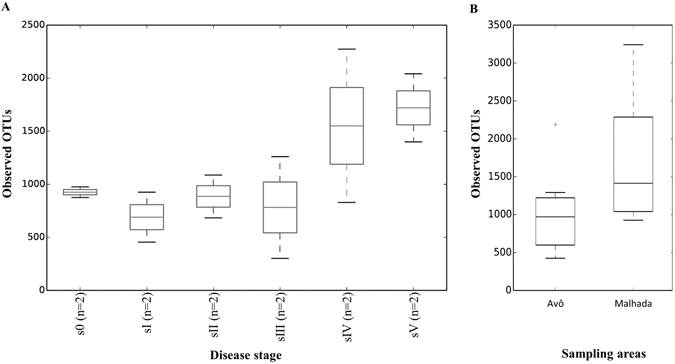
α-Diversity analysis of endophytic, wood colonizing bacterial communities from pine trees of six different symptomatic classes from the two sampling areas based on sequencing of PCR amplified 16S rRNA gene fragments (Chao1). (A) Comparison of different symptomatic stages (B) Comparison of sites.
Like expected from the fingerprint analysis, our results demonstrated that the composition of the endophytic communities from pine trees differed in response to the investigated site but also to the symptomatic stages (p < 0.001) (Fig. 3). Proteobacteria, varying between 39% (M-sII) and 97% (A-sIII), and Firmicutes ranging from 0.70% (A-sIV) and 44% (M-sII), were the predominant endophyte bacterial phyla in the pine trees at both sites, followed by Actinobacteria, Bacteroidetes, Acidobacteria at site A, and Fusobacteria, Acidobacteria and Thermi at site M. Members of Proteobacteria included mainly Gammaproteobacteria (1% (M-sI) - 94% (A-sIII)), Alphaproteobacteria (5.71% (M-s0) - 40% (M-s4)) and Betaproteobacteria (10% (M-sIII) - 34% (M-s0)). Bacilli (<1% (A-sIV) - 44% (M-sII)) and Clostridia (<1% (A-sIII - 11% (M-s0)) were the predominant Firmicutes.
Figure 3.
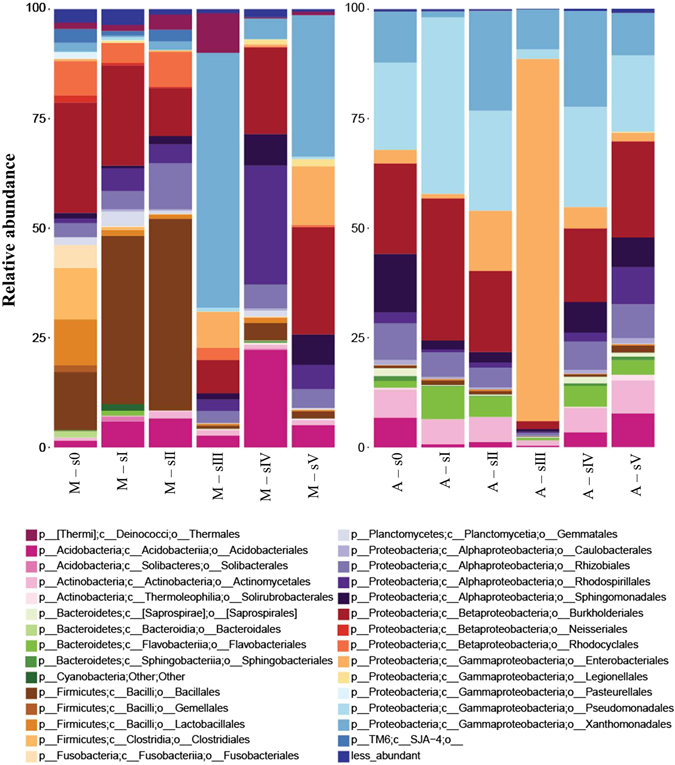
Diversity of endophytic, wood colonizing bacterial communities at the level of classes from pine trees of six different symptomatic classes from the two sampling areas (A and M) based on sequencing of PCR amplified 16S rRNA gene fragments. Only bacterial classes higher in abundance than 0.5% were included.
At site M healthy (s0-sII) trees harbored a large number of OTUs which could be related to Bacillus and Streptococcus; this number was significantly reduced at later symptom stages (sIII-sV). At site A, however, Bacilli related OTUs only played a minor role and did not change in response to the different symptom stages. Xanthomonadaceae, in the case of site M and Enterobacteriaceae at site A; were the dominant endophytic bacterial group in the trees of symptomatic stage III, which had the most distinct microbiome according to Chao1 analysis at both sites (67% at M - 94% at A), with high abundance of Serratia and especially Erwinia found at site A (8% at M - 82% at A) while Dyella and Pseudoxanthomonas were highly enriched at site M (57% M - 9% A). These results were also confirmed by a PCA analysis (Fig. 4).
Figure 4.
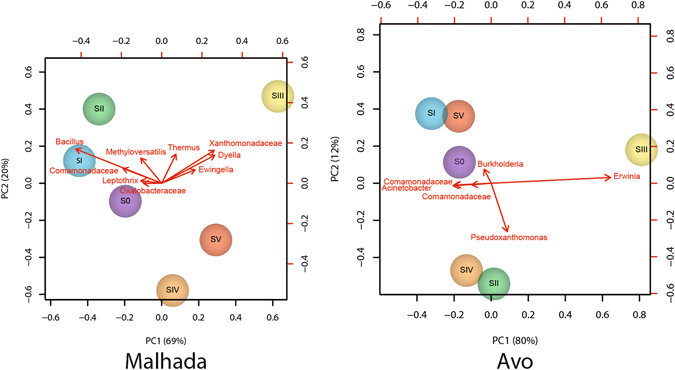
Principal component of endophytic, wood colonizing bacterial communities at different phylogenetic levels from pine trees of six different symptomatic classes from the two sampling areas (A: Malhada B: Avo) based on sequencing of PCR amplified 16S rRNA gene fragments.
Isolation and characterization of endophytic bacteria
Colony-Forming Units per gram of wood (CFU/g) varied from 1.5 × 103 ± 7.0 × 102 to 1.5 × 106 ± 6.3 × 105 in area M and from 7.1 × 101 ± 3.7 × 101 to 2.3 × 107 ± 7.0 × 106 in area A. In general, the number of bacteria was higher in site A than in site M as found by community barcoding. Furthermore, symptomatic trees showed more CFU per gram of wood than healthy trees or trees with low level symptoms. In 11 trees of area M and 7 trees of area A, no bacterial isolates were obtained, most of them in healthy condition (data not shown).
In total, 313 endophytic bacterial isolates were grouped in 277 RAPD group types (data not shown). Overall, 16S rRNA genes from 144 isolates and 133 isolates from site M and A, respectively, were sequenced, representing 162 bacterial isolates (M) and 151 bacterial isolates (A). As expected, not all of the taxa detected by sequencing were cultivatable under the conditions used.
Diversity of isolates was comparable at site A and site M (H′ = 2.90 and 2.92; D = 0.91 and 0.92, respectively). As found by barcoding, the highest diversity was found in the samples of the most diseased trees in both sites (Table S2). In accordance to the fingerprint and barcoding data, redundancy analysis (RDA) indicated that the majority of the trees clustered according to the sampling areas and according to the symptomatic stage of the trees inside each sampling site (data not shown). Also one-way ANOVA showed statistical differences between healthy (s0-sIII) and symptomatic trees (sV) in both areas (p < 0.05, respectively), when isolates were analyzed on the genus level (Figure S1). Differences were not observed between symptomatic trees (sV) of site A and site M (p > 0.05).
Proteobacteria were the most abundant isolates at both sites (80–85%), independently of the degree of PWD (Fig. 5). Whereas at site A 59% of the isolates were classified as Gammaproteobacteria (Enterobacteriaceae, Pseudomonadaceae, Xantomonadaceae), followed by Alphaproteobacteria (20%) and Betaproteobacteria (4%), at site M Gammaproteobacteria were less abundant (35%) but the abundance of Betaproteobacteria was much higher (28%); the number of isolates for Alphaproteobacteria was comparable between both sites. These differences in the abundance of Gammaproteobacteria were mainly related to different numbers of isolates from Xanthomonadaceae (31% and 17% of the isolates for site A and M, respectively). For Betaproteobacteria, diverging numbers of Burkholderiaceae isolates were responsible for the differences observed for site A (4%) and M (24% with the additional families Commomonadaceae and Alcaligenaceae). Overall, besides Proteobacteria also Acidobacteria, Actinobacteria, Bacilli, Flavobacteriia and Sphingobacteria were isolated but with low frequency.
Figure 5.
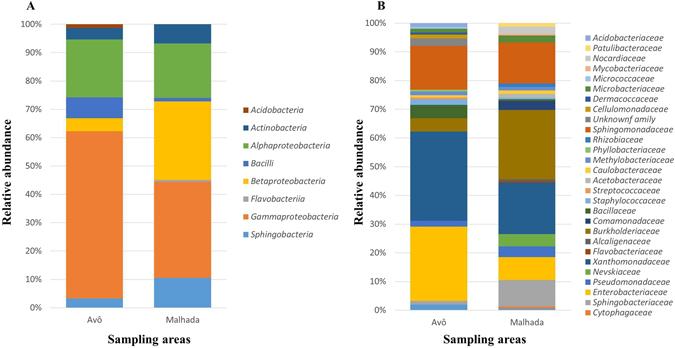
Phylogenetic characterization of endophytic, wood colonizing bacteria from pine trees of the two study sites (A and M) obtained by cultivation based approaches at the level of phyla (A) and families (B).
At site M, the genera Pseudoxanthomonas, Ewingella and Brevundimonas were the most abundant at sIII, Burkholderia and Sphingomonas at sIV, and Ralstonia, Sphingomonas, and Pseudoxanthomonas at sV. Moreover, Variovorax and Sphingomonas were common in all the symptomatic classes. While the most abundant genera at site A were Streptococcus, Staphylococcus in s0, isolates belonging to the genera Erwinia in sI, Pseudoxanthomonas, Erwinia, Methylobacterium and Pantoea in sII, Pseudoxanthomonas, Erwinia, Serratia, Klebsiella in sIII, Pseudoxanthomonas, Enterobacter and Pantoea in sIV, and Sphingomonas, Pseudoxanthomonas and Burkholderia in sV.
One hundred and eighty-five isolates representatives of the OTU detected with more than 1% abundance were phenotypically characterized. Interesting, most of the 29 strains of Erwinia, which genus includes 81% of the OTU of sIII trees at site A, were positive for all tested characteristics except chitin degradation. In site M, six strains of Dyella (28.2% of the OTU in sIII), were positive for lipases, proteases, siderophores and zinc solubilization (Table S4).
Isolates from site A tend to have more proteolytic and cellulolytic activity when isolated from trees in symptomatic sV, whereas at site M, no clear pattern was visible and the functional activities of the isolates from trees in the different symptomatic stages were similar (Figure S3).
Discussion
Endophytic bacteria exist within the living tissues of probably all plants, but compared with herbaceous plants, remarkably little is known about their ecology in trees. In this study, we assessed for the first time the diversity of endophytic microbial community of Portuguese P. pinaster trees, by cultivation and molecular methods, focusing on trees infected and non-infected with B. xylophilus.
All sampled trees were colonized by a diverse endophytic community, although in some cases no isolates were obtained and the presence of these microbes was only described by 16S rRNA gene barcoding. Interestingly when considering the composition of the endophytic microbiome, pine trees grouped by sampling areas mainly. The main components of the community were Proteobacteria of the class Gammaproteobacteria and, depending on the sampling area, also Firmicutes. Six families were the most abundant found in all symptomatic stages from both sampling areas – Bacillaceae, Burkholderiaceae, Enterobacteriaceae, Comamonadaceae, Sphingomonadaceae and Xanthomonadaceae. However, the relative abundance of bacterial families differed among the two sampling areas. These pronounced differences between the two sites that are geographically close, might be due to differences in environmental conditions, probably related to water availability, pH and soil composition25.
As expected, the pathogen impacted the microbiome structure of wood colonizing endophytes. Whereas at early stages of the disease only slight shifts in the bacterial diversity were observed, from sIII on drastic shifts were visible. Overall, an increase in diversity with more severe symptomatic stage was visible. This may be a result of a loss in the ability of the plants to control microbial invasion of the plant interior from outside, or of a loss in the control of the growth of endophytes as a result of the disease. An increase in the endophytic microbial diversity related with the decline of the tree was previously also observed when studying the endophytic fungi of pine trees26. The sudden modification of the microbial community of the trees at sIII might be related to the fact, that mostly at this stage the pathogen is detected in higher numbers for the first time after tree infection at the trunk level27, which could lead to significant reduction of plant immunity as well as the creating new conditions inside the tree28. However, it is also likely that bacteria which are carried by or able to adhere to the pathogen at this stage are present in high abundance simply as a result of the increased number of B. xylophilus. This assumption is confirmed by the observation that mainly an increase in OTUs linked to Xanthomonadaceae and Enterobacteriaceae, including Dyella and Ewingella strains, respectively, which were previously shown to be carried by with the pathogen12, increased in number up to 58% of the community at sIII was observed at site M. A phylogenetic analysis of the sequences obtained in this study, which the strains described previously as carried by the nematode4 confirmed this assumption (Figure S4). At site A comparable pattern was observed. Here OTUs related to Erwinia, again a previously described species associated to the pathogen12, increased at sIII. Interestingly the relative abundance of typical plant endophytes for example OTUs related to Bacillus, Comamonadaceae or Methyloversatilis at both sites decreased at the same time.
Although some comparable developments were seen at both sites, the present study indicated mainly at later stages of the disease site specific effects. At site A, the microbiome at symptomatic stage sV resembles community composition similar to early stages of the disease which was not the case in site M. Thus it is quite likely that both the original composition of the microbiome as well as site specific conditions determine the course of the disease in terms of microbial community composition.
Interestingly none of the ten most important plant pathogenic bacteria29 were isolated or detected by 16S rRNA gene sequencing in this study, and a pathogenic role for the endophytic community was previously disregarded. However, it is quite likely that most of the new endophytes are capable to act as saprotrophs and degrade important structural elements of plants. This assumption is confirmed by the analysis of the phenotype of the isolates mainly at site A, where higher symptomatic stages lead to an increase in strains with proteolytic and cellulolytic activity. Cellulases (β-1,4-endoglucanses) were also described in the genome and in the microbiome of B. xylophilus and may help the nematode in the invading the plant tissue30, 31. However, for root associated microbes cellulolytic activities have been described as a part of their ability to enter the root32. In our study typical soil borne microbes like Chitinophaga and Ewingella have been isolated carrying those functional traits. Thus it stays unclear if the increase in cellulolytic activities is more related to microbes introduced by the nematode, or microbes that entered the plant interior via the soil.
Overall the presented data gives clear evidence for significant shifts both in the structure and the function of endophytic bacteria with ongoing disease development, which might lead to an additional decline of the tree fitness. Such data might be of great importance when mitigation strategies are developed for example by introducing biocontrol active antagonist like recognized nematicidal bacteria33 into the wood material. If such approaches are successful needs to be studied in future experiments.
Experimental Procedures
Field sites and sample collection
Samples were collected from Malhada Velha, Arganil (M) and Avô, Oliveira do Hospital (A)6. These are two different areas affected by PWD since 2008 from Coimbra District, Central Portugal. The areas include mainly P. pinaster, with a sparse number of P. radiata, Quercus faginea and Eucalyptus globulus trees (Table S1). The sampled trees were classified into six different PWD symptom classes.
Sampling was performed from April to August 2009 according to Proença et al.6. Briefly, each sample consisted of pinewood cross-sections from cut trees or wood obtained by drilling a 5 mm diameter hole to a depth of 10 to 15 cm with a sterilized hand brace drill (Haglof, Mora, Sweden). Forty-seven trees were sampled at the trunk (1.5 m height). For those pines cut for phytosanitary purposes, an additional sample was obtained from the crown of the tree. From area M, 18 P. pinaster trees were sampled. From area A, 25 P. pinaster trees were sampled (Table S1). Sampled trees had an average trunk diameter at breast height of 19.9 ± 8.03 cm in Malhada and 24.7 ± 10.8 cm in Avô. All samples were stored at 4 °C after sampling and further processed as described below latest one day after sampling.
The presence of nematodes was screened in all sampled trees using modified Baermann funnels34. B. xylophilus was detected in three symptomatic P. pinaster trees area M (M24, M47, M67) and 14 symptomatic P. pinaster trees from area A (A12, A25, A36, A37, A41, A52, A60, A72, A88, A96, A103, AB20, AB23, AB29). B. xylophilus was not detected in healthy trees.
The bark and sapwood of each sample were removed under sterile conditions and the wood cut in pieces of the size of about 1 × 0.5 × 0.5 cm. Microbes were released from the wood material by shaking 5 g of wood pieces at 25 °C, for 2 h in 0.9% NaCl. The resulting suspensions were either stored at −20 °C for molecular analysis or directly processed for isolation based approaches.
Extraction of community DNA
Total DNA of microbes extracted from each tree wood sample was obtained, following the method of Nielsen et al.35 with an extra incubation step with 1% CTAB (cetyl trimethyl ammonium bromide)/0.7 M NaCl at 65 °C for 20 min.
Fingerprinting of endophytic bacteria
The extracted community DNA was used as template for fingerprinting of the 16S rRNA gene from Bacteria using denaturating gradient gel electrophoresis (DGGE). The 16S rRNA amplicons were generated using primer pairs 27 F and 1525R36 for the first PCR. The obtained amplicons were subsequently used as DNA template for a Nested-PCR by using primer pair 341F-GC clamp and 907R37.
Fingerprints were obtained by DGGE analysis using a DCode™ Universal Mutation Detection System (Bio-Rad Laboratories, Hercules, California, USA). PCR samples were loaded into 8% polyacrylamide gels with a denaturing gradient ranging from 30–70% (100% denaturant is defined as 7 M urea and 40% formamide). Gels were run at 70 V, for 17 h, at 60 °C and stained with ethidium bromide.
Barcoding of endophytic bacteria
The extracted community DNA was further used as template for 16S rRNA amplicon sequencing using MiSeq Reagent Kit v3 (Illumina, San Diego, USA). Therefore the extracted community DNA of each symptomatic state from each sampling area was pooled together, resulting in 12 samples (six classes, from two sampling areas; M-s0 through M-sV; A-s0 through A-sV) and amplified using primers S-D-Bact-0008-a-S-16 and S-D-Bact-0343-a-A-1538. Amplicons were verified for their correct size by gel electrophoresis; bands with the expected size were excised and purified by PCR Purification Kit (Macherey-Nagel, Düren, Germany). Each PCR was performed in triplicates, which were pooled and analyzed by Bioanalyzer DNA 7500 Chip (Agilent, Santa Clara, USA). DNA quantification was performed by Picogreen (Thermo Fisher, Waltham, USA). Sequencing was performed using the paired end methods on a Illumina MiSeq (Illumina, Berlin, Germany).
Sequences were trimmed and merged using Adapterremoval39 using default values, except ambiguous bases at 5′/3′ termini were trimmed and bases at 5′/3′ termini with quality scores equal or less than 15 were trimmed. Contaminating PhiX sequences were removed using DeconSeq v.0.4.340. Chimeric sequences and plastid sequences were removed by using QIIME v.1.9.141. OTU picking was done using Open-reference OTU picking against the 97% similarity clustering of Greengenes as of May 201342. Basic diversity analysis was likewise done using Qiime v.1.9.1 at a subsampling of 24,069 reads. Stacked barplots of the taxonomical distribution, filtering away all OTUs with less than 1% of reads per sample, was done using R v.3.2.4 (http://r-project.org). The PCA biplot was done on Hellinger transformed data filtering away all OTUs with less than 5% of reads per sample, also using R v.3.2.4 (http://r-project.org). The following R packages were used: vegan43, gplots, gplots2, sciplot, plyr, MASS and gridExtra.
Isolation and phylotyping of endophytic bacteria
To isolate bacteria from the woody part of pine trees, dilutions of the suspensions obtained as described above were plated on R2A agar. Plates were incubated up to five days, in the dark, at 25 °C. The number of colonies per plate was counted using a stereomicroscope, and the number of colonies per gram of sampled wood was assessed. Colonies from each tree sample with different morphologies were randomly isolated, and preserved in LB medium (Difco Laboratories, Detroit, USA) with 15% glycerol stocks at −80 °C after sub-cultivation and purification.
DNA from each bacterial isolate was extracted according to Nielsen et al.35 and isolates were grouped by RAPD typing. RAPD fragments were amplified by PCR, using primer OPA-03 as previously described6. DNA profiles for isolates were grouped on basis of visual similarities of the fragments analyzed by electrophoresis in a 2% agarose gel stained with ethidium bromide. Representative strains from each RAPD group were phylogenetically identified by 16S rRNA gene sequencing using primers 27 F and 1525 R as described36.
All obtained sequences from bacterial isolates were compared with sequences available in the EMBL/GenBank database using BLASTN network services44 and with sequences available on the Eztaxon-e server (http://eztaxon-e.ezbiocloud.net/)45. Sequences were aligned within the SINA alignment service46. Sequences were also checked for chimeric artefacts by using Mallard software47. Sequences were included in 16S rRNA-based Living Tree Project (LTP) release 115 database (http://www.arb-silva.de/projects/living-tree/) by parsimony implemented in the ARB software package version 5.548. Phylogenetic dendrograms of sequences from this study and closest reference sequences were constructed by the Randomized Axelerated Maximum Likelihood (RAxML) method with GTRGAMMA model49 included in the ARB software48.
The phenotype of strains belonging to the genera which were also detected by sequencing of extracted community DNA (>1%) was analyzed focusing on the solubilization of phosphate and zinc, production of siderophores; and hydrolyzing activities including protease (casein), lipase (Tween 20, 40, 60 and 80), cellulase and chitinase activities50.
Statistical analysis
PAST 2.08 software51 was used to calculate the Shannon–Weaver index (H′) and Simpson indices, one-way ANOVA, paired samples t-test and similarity percentage (SIMPER). Indices were calculated for isolates based on number of strains belonging to each phylotype (genera) and for DGGE profiles were based on the presence of each DGGE band.
Based on the results of OTUs obtained by sequencing of the obtained amplicons, alpha diversity was calculated, including Chao1 (QIIME package), as well as Shannon and the Simpson indices (PAST 2.08 software). To estimate Beta diversity, Bray-Curtis dissimilarity matrix were used to examine the similarity of the membership and structure found in the various samples.
Redundancy analysis (RDA) was performed in order to reveal relationships between pine trees, bacterial species and the environmental variables (two sampling areas, PWD symptom class6 and presence of B. xylophilus) by using the software package CANOCO (version 4.5.1). RDA was accompanied by Monte Carlo permutation tests to evaluate the statistical significance of the effects of the explanatory variables on the species composition of the samples52.
PCoA based on OTUs was performed in order to understand the pine’s endophytic microbiomes from two sampling areas belonging to six PWD symptom classes.
Nucleotide sequence accession numbers
The 16S rRNA gene sequences of the isolates, have been deposited in the NCBI Genbank database under the accession numbers according KJ654634 - KJ654916, KT730168 and KT730169. The sequences obtained from the amplification of the 16S rRNA gene using the extracted community DNA as template have been deposited GenBank under the SRA Accession number SRP077519.
Electronic supplementary material
Acknowledgements
We thank to CAULE association (Associação Florestal da Beira Serra) for their help and sampling authorization. We thank Professor Isabel Abrantes for help on B. xylophilus identification and to Dr. Gregor Grass for his critical revision. This research was partially supported by Direcção Regional de Florestas, Fundo Florestal Permanente and Autoridade Florestal Nacional, through a national project ‘O nemátode-da-madeira-do-pinheiro (NMP), Bursaphelenchus xylophilus’, by FEDER funds through the Programa Operacional Factores de Competitividade – COMPETE and by national funds through the Fundação para a Ciência e a Tecnologia (FCT), Portugal, under the project PTDC/AGR-CFL/115373/2009. This work was further supported by COST Action FP1305 - Biolink: Linking belowground biodiversity and ecosystem function in European forests. D.N.P. was supported by FCT, postdoctoral fellowship SFRH/BPD/100721/2014.
Author Contributions
P.V.M. and D.N.P. conceived the project. D.N.P., P.V.M. and M.S. designed the study. D.N.P. and R.F. collected the samples. D.N.P., P.V.M., R.F., S.K., A.S. and G.V. performed the analytical work including data evaluation. D.N.P., P.V.M., G.V., A.S. and M.S. wrote the manuscript. All authors approved the manuscript.
Competing Interests
The authors declare that they have no competing interests.
Footnotes
Michael Schloter and Paula V. Morais contributed equally to this work.
Electronic supplementary material
Supplementary information accompanies this paper at doi:10.1038/s41598-017-04141-6
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
References
- 1.Proença DN, Grass G, Morais PV. Understanding pine wilt disease: roles of the pine endophytic bacteria and of the bacteria carried by the disease-causing pinewood nematode. Microbiologyopen. 2016;0:1–20. doi: 10.1002/mbo3.415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Mota M, et al. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology. 1999;1:727–734. doi: 10.1163/156854199508757. [DOI] [Google Scholar]
- 3.Rodrigues, J. In Pine wilt disease: a worldwide threat to forest ecosystems. (eds Vieira, P. & Mota, M.) 5–14 (Springer, 2008).
- 4.Nickle WR, Golden AM, Mamiya Y, Wergin WP. On the taxonomy and morphology of the pine wood nematode, Bursaphelenchus xylophilus (Steiner & Buhrer 1934) Nickle 1970. J. Nematol. 1981;13:385–392. [PMC free article] [PubMed] [Google Scholar]
- 5.EPPO/OEPP. Diagnostic protocols for regulated pests: Bursaphelenchus xylophilus. Bull. OEPP/EPPO39, 344–353 (2009).
- 6.Proença DN, et al. Diversity of bacteria associated with Bursaphelenchus xylophilus and other nematodes isolated from Pinus pinaster trees with pine wilt disease. PLoS One. 2010;5:e15191. doi: 10.1371/journal.pone.0015191. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Zhao BG, Lin F. Mutualistic symbiosis between Bursaphelenchus xylophilus and bacteria of the genus Pseudomonas. For. Pathol. 2005;35:339–345. doi: 10.1111/j.1439-0329.2005.00417.x. [DOI] [Google Scholar]
- 8.Higgins, D., Harmey, M. & Jones, D. Pathogenicity related gene expression in Bursaphelenchus xylophilus. In Sustainability of pine forests in relation to pine wilt and decline (eds Kazuyoshi, F., Togashi, K. & Ikedu, T.) 23–28 (Proceedings of international symposium, Tokyo, 1999).
- 9.Han ZM, Hong YD, Zhao BG. A study on pathogenicity of bacteria carried by pine wood nematodes. J. Phytopathol. 2003;151:683–689. doi: 10.1046/j.1439-0434.2003.00790.x. [DOI] [Google Scholar]
- 10.Guo Q, Guo D, Zhao B, Xu J, Li R. Two cyclic dipeptides from Pseudomonas fluorescens GcM5-1A carried by the pine wood nematode and their toxicities to japanese black pine suspension cells and seedlings in vitro. J. Nematol. 2007;39:243–247. [PMC free article] [PubMed] [Google Scholar]
- 11.Zhao BG, et al. Pathogenic roles of the bacteria carried by Bursaphelenchus mucronatus. J. Nematol. 2009;41:11–16. [PMC free article] [PubMed] [Google Scholar]
- 12.Proença DN, Fonseca L, Powers TO, Abrantes IMO, Morais PV. Diversity of bacteria carried by pinewood nematode in USA and phylogenetic comparison with isolates from other countries. PLoS One. 2014;9:e105190. doi: 10.1371/journal.pone.0105190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Hardoim PR, et al. The hidden world within plants: ecological and evolutionary considerations for defining functioning of microbial endophytes. Microbiol. Mol. Biol. Rev. 2015;79:293–320. doi: 10.1128/MMBR.00050-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.van Buren AM, Andre C, Ishmaru CA. Biological control of the bacterial ring rot pathogen by endophytic bacteria isolated from potato. Phytopathology. 1993;83:1406. [Google Scholar]
- 15.Brooks DS, Gonzalez CF, Appel DN, Filer TH. Evaluation of endophytic bacteria as potential biocontrol agents for oak wilt. Biol. Con. 1994;4:373–381. doi: 10.1006/bcon.1994.1047. [DOI] [Google Scholar]
- 16.Hinton DM, Bacon CW. Enterobacter cloacae is an endophytic symbiont of corn. Mycopathologia. 1995;129:117–125. doi: 10.1007/BF01103471. [DOI] [PubMed] [Google Scholar]
- 17.Mukhopadhyay NK, et al. Ultrastructural studies on the colonisation of banana tissue and Fusarium oxysporum f. sp. cubense race 4 by the endophytic bacterium Burkholderia cepacia. J Phytopathol. 1997;145:479–486. doi: 10.1111/j.1439-0434.1997.tb00354.x. [DOI] [Google Scholar]
- 18.Pirttilä AM, Laukkanen H, Pospiech H, Myllylä R, Hohtola A. Detection of intracellular bacteria in the buds of Scotch pine (Pinus sylvestris L.) by in situ hybridization. Appl. Environ. Microbiol. 2000;66:3073–3077. doi: 10.1128/AEM.66.7.3073-3077.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Pirttilä AM, Podolich O, Koskimäki JJ, Hohtola E, Hohtola A. Role of origin and endophyte infection in browning of bud-derived tissue cultures of Scots pine (Pinus sylvestris L.) Plant Cell. Tissue Organ Cult. 2008;95:47–55. doi: 10.1007/s11240-008-9413-x. [DOI] [Google Scholar]
- 20.Strzelczyk E, Li CY. Bacterial endobionts in the big non-mycorrhizal roots of Scots pine (Pinus sylvestris L.) Microbiol. Res. 2000;155:229–232. doi: 10.1016/S0944-5013(00)80037-3. [DOI] [PubMed] [Google Scholar]
- 21.Izumi H, Anderson IC, Killham K, Moore ERB. Diversity of predominant endophytic bacteria in European deciduous and coniferous trees. Can. J. Microbiol. 2008;54:173–179. doi: 10.1139/W07-134. [DOI] [PubMed] [Google Scholar]
- 22.Shishido M, Loeb BM, Chanway CP. External and internal root colonization of lodgepole pine seedlings by two growth-promoting Bacillus strains originated from different root microsites. Can. J. Microbiol. 1995;41:707–713. doi: 10.1139/m95-097. [DOI] [Google Scholar]
- 23.Bal, A. S. Isolation of endophytic bacteria from Lodgepole Pine (Pinus contorta var. latifolia (Dougl.) Engelm.) and Western Red Cedar (Thuja plicata Donn.) and determination of their nitrogen fixing ability. (The University of British Columbia, 2003).
- 24.Carrell AA, Frank AC. Pinus flexilis and Picea engelmannii share a simple and consistent needle endophyte microbiota with a potential role in nitrogen fixation. Front. Microbiol. 2014;5:1–11. doi: 10.3389/fmicb.2014.00333. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Eschen R, Hunt S, Mykura C, Gange AC, Sutton BC. The foliar endophytic fungal community composition in Cirsium arvense is affected by mycorrhizal colonization and soil nutrient content. Fungal Biol. 2010;114:991–998. doi: 10.1016/j.funbio.2010.09.009. [DOI] [PubMed] [Google Scholar]
- 26.Giordano L, Gonthier P, Varese GC, Miserere L, Nicolotti G. Mycobiota inhabiting sapwood of healthy and declining Scots pine (Pinus sylvestris L.) trees in the Alps. Fungal Divers. 2009;38:69–83. [Google Scholar]
- 27.Reva V, Fonseca L, Lousada JL, Abrantes I, Viegas DX. Impact of the pinewood nematode, Bursaphelenchus xylophilus, on gross calorific value and chemical composition of Pinus pinaster woody biomass. Eur. J. For. Res. 2012;131:1025–1033. doi: 10.1007/s10342-011-0574-5. [DOI] [Google Scholar]
- 28.Lygis V, Vasiliauskas R, Stenlid J, Vasiliauskas A. Silvicultural and pathological evaluation of Scots pine afforestations mixed with deciduous trees to reduce the infections by Heterobasidion annosum s.s. For. Ecol. Manage. 2004;201:275–285. doi: 10.1016/j.foreco.2004.07.013. [DOI] [Google Scholar]
- 29.Mansfield J, et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012;13:614–629. doi: 10.1111/j.1364-3703.2012.00804.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kikuchi T, et al. Genomic insights into the origin of parasitism in the emerging plant pathogen Bursaphelenchus xylophilus. PLoS Pathog. 2011;7:e1002219. doi: 10.1371/journal.ppat.1002219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Vicente CSL, et al. Characterization of bacteria associated with pinewood nematode Bursaphelenchus xylophilus. PLoS One. 2012;7:e46661. doi: 10.1371/journal.pone.0046661. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Quecine MC, et al. Sugarcane growth promotion by the endophytic bacterium Pantoea agglomerans 33.1. Appl. Environ. Microbiol. 2012;78:7511–7518. doi: 10.1128/AEM.00836-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Paiva G, et al. Nematicidal bacteria associated to pinewood nematode produce extracellular proteases. PLoS One. 2013;8:e79705. doi: 10.1371/journal.pone.0079705. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Abrantes IdeO, de Morais M, Paiva IdeF, Santos MdeA, et al. Análise nematológica de solos e plantas. Ciência Biológica. 1976;1:139–155. [Google Scholar]
- 35.Nielsen P, Fritze D, Priest FG. Phenetic diversity of alkaliphilic Bacillus strains: proposal for nine new species. Microbiology. 1995;141:1745–1761. doi: 10.1099/13500872-141-7-1745. [DOI] [Google Scholar]
- 36.Rainey FA, Dorsch M, Morgan HW, Stackebrandt E. 16S rDNA analysis of Spirochaeta thermophila: its phylogenetic position and implications for the systematics of the order. Spirochaetales. Syst. Appl. Microbiol. 1992;15:197–202. doi: 10.1016/S0723-2020(11)80092-5. [DOI] [Google Scholar]
- 37.Santos SS, et al. Diversity of cloacal microbial community in migratory shorebirds that uses tagus estuary as stopover habitats and their potential to harbor and disperse pathogenic microorganisms. FEMS Microbiol. Ecol. 2012;82:63–74. doi: 10.1111/j.1574-6941.2012.01407.x. [DOI] [PubMed] [Google Scholar]
- 38.Klindworth A, et al. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013;41:1–11. doi: 10.1093/nar/gks808. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lindgreen S. AdapterRemoval: easy cleaning of next-generation sequencing reads. BMC Res. Notes. 2012;51:337. doi: 10.1186/1756-0500-5-337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Schmieder R, Edwards R. Fast identification and removal of sequence contamination from genomic and metagenomic datasets. PLoS One. 2011;6:e17288. doi: 10.1371/journal.pone.0017288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Caporaso JG, et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods. 2010;7:335–336. doi: 10.1038/nmeth.f.303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.DeSantis TZ, et al. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006;72:5069–5072. doi: 10.1128/AEM.03006-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Oksanen, J. et al. The vegan Package. 285 at http://vegan.r-forge.r-project.org/ (2016).
- 44.Altschul SF, et al. Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 1997;25:3389–3402. doi: 10.1093/nar/25.17.3389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Kim O-S, et al. Introducing EzTaxon-e: a prokaryotic 16S rRNA gene sequence database with phylotypes that represent uncultured species. Int. J. Syst. Evol. Microbiol. 2012;62:716–721. doi: 10.1099/ijs.0.038075-0. [DOI] [PubMed] [Google Scholar]
- 46.Pruesse E, Peplies J, Glöckner FO. SINA: accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics. 2012;28:1823–1829. doi: 10.1093/bioinformatics/bts252. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ashelford KE, Chuzhanova NA, Fry JC, Jones AJ, Weightman AJ. New screening software shows that most recent large 16S rRNA gene clone libraries contain chimeras. Appl. Environ. Microbiol. 2006;72:5734–5741. doi: 10.1128/AEM.00556-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Ludwig W, et al. ARB: a software environment for sequence data. Nucleic Acids Res. 2004;32:1363–1371. doi: 10.1093/nar/gkh293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Stamatakis A. RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics. 2006;22:2688–2690. doi: 10.1093/bioinformatics/btl446. [DOI] [PubMed] [Google Scholar]
- 50.Naveed M, et al. The endophyte Enterobacter sp. FD17: a maize growth enhancer selected based on rigorous testing of plant beneficial traits and colonization characteristics. Biol. Fertil. Soils. 2014;50:249–262. doi: 10.1007/s00374-013-0854-y. [DOI] [Google Scholar]
- 51.Hammer Ø, Harper DAT, Ryan PD. Past: Paleontological Statistics Software Package for Education and Data Analysis. Palaeontol. Electron. 2001;4:1–9. [Google Scholar]
- 52.van den Brink PJ, Braak CJFT. Principal response curves: analysis of time-dependent multivariate responses of biological community to stress. Environ. Toxicol. Chem. 1999;18:138–148. doi: 10.1002/etc.5620180207. [DOI] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.