

ABSTRACT
The mouse t haplotype, a variant 20 cM genomic region on Chromosome 17, harbors 16 embryonic control genes identified by recessive lethal mutations isolated from wild mouse populations. Due to technical constraints so far only one of these, the tw5 lethal, has been cloned and molecularly characterized. Here we report the molecular isolation of the tw18 lethal. Embryos carrying the tw18 lethal die from major gastrulation defects commencing with primitive streak formation at E6.5. We have used transcriptome and marker gene analyses to describe the molecular etiology of the tw18 phenotype. We show that both WNT and Nodal signal transduction are impaired in the mutant epiblast, causing embryonic patterning defects and failure of primitive streak and mesoderm formation. By using a candidate gene approach, gene knockout by homologous recombination and genetic rescue, we have identified the gene causing the tw18 phenotype as Ppp2r1a, encoding the PP2A scaffolding subunit PR65alpha. Our work highlights the importance of phosphatase 2A in embryonic patterning, primitive streak formation, gastrulation, and mesoderm formation downstream of WNT and Nodal signaling.
KEY WORDS: t haplotype, tw18, Mesoderm, Primitive streak, Signal transduction, PP2A phosphatase
Summary: The tw18 lethal, a ‘classical’ mutation, affects gastrulation and embryo patterning. This article shows that phosphatase PP2A is dysfunctional in tw18 and essential for Nodal and WNT signaling in the epiblast.
INTRODUCTION
Between 6-25% of individuals in wild house mouse populations carry a t haplotype, a variant form of the t complex spanning over 20 cM (∼40 Mbp) of Chromosome 17 (Chr 17) (Carroll et al., 2004; reviewed in Silver, 1985; Sugimoto, 2014). This region comprises many important developmental regulators, among them Pou5f1 (Oct4), Nkx2-5, Axin and Brachyury (T), as well as 16 recessive lethal mutations, known as t lethals, which are encoded by different t haplotypes and affect early embryonic development (Klein et al., 1984; Bennett, 1975; Sugimoto, 2014). Due to particularities of the t haplotypes, to date only one t lethal, tw5, has been identified and characterized at the molecular level (Sugimoto et al., 2012). The t lethal tw18 and related t lethals of the same complementation group (t4, t9) were described about 50 years ago (Bennett and Dunn, 1960; Moser and Gluecksohn-Waelsch, 1967). They cause strong gastrulation defects with striking overgrowth of the primitive streak (PS) and bulging of cells into the amniotic cavity, commencing on the seventh and prominent on the eighth to ninth day of gestation, followed by embryonic death one day later. In contrast, normal development requires the ingression of epiblast cells at the PS, epithelial-to-mesenchymal transition (EMT) and migration of single mesodermal cells in between the epiblast and the visceral endoderm (VE) (reviewed in Arnold and Robertson, 2009). Thus, the failure of mesoderm formation characterizes the embryonic phenotype of these mutants. However, the molecular cause and consequences have remained obscure.
The tw18 haplotype originated from a rare recombination event between a t haplotype and the wild-type chromosome across a large inversion, causing a large deletion of at least 3.3 Mbp comprising many genes (Búcan et al., 1987; Barclay et al., 1996). Here we have determined the extent of the deletion and, by using a candidate gene approach, have identified Ppp2r1a as the gene causing the tw18 mutant phenotype. It encodes the scaffolding subunit Aα (PR65alpha) of protein phosphatase 2A (PP2A). The PP2A family of serine-threonine phosphatases is implicated in many cellular processes, such as the regulation of signaling pathways, cell cycle, DNA replication and apoptosis (Virshup, 2000; Janssens and Goris, 2001), and its deregulation is associated with a range of human diseases like Alzheimer's disease and different types of cancer (Eichhorn et al., 2009; Vafai and Stock, 2002). We describe the identification of the tw18 lethal and provide a detailed molecular characterization of the tw18 gastrulation defects.
RESULTS
Nodal and WNT signal transduction are impaired in homozygous tw18 embryos
Prior to this study, tw18/tw18 mutant embryos have not been characterized at the molecular level. We started the molecular characterization of tw18/tw18 mutants by analyzing the transcriptome of homozygous, heterozygous and wild-type embryos at embryonic day (E)6.5, when malformations start to develop, by RNA-sequencing (RNA-seq). Embryos were dissected in two parts: the embryo proper consisting of epiblast and surrounding visceral endoderm (VE), which was used for RNA extraction, and the extra-embryonic tissue, required for genotyping. The Pearson correlation coefficient (PCC) of the sequencing data showed a high similarity between wild-type and heterozygous tw18 samples (PCC 0.99, Fig. 1A). The tw18/tw18 sample differed from both the wild-type (PCC 0.77) and the heterozygous sample (PCC 0.82). After removing genes with FPKM (fragments per kilobase of exon per million fragments mapped) values <1 in the three samples, log2 of the fold change (FC)≥1.0 for any comparable pair was used as the threshold to define the affected genes, and we got 2539 deferentially expressed genes (Fig. 1B).
Fig. 1.

Transcriptome analysis reveals downregulation of WNT and Nodal targets in tw18 mutant embryos. (A) Pearson correlation coefficient of next generation sequencing data from wild-type, homozygous and heterozygous tw18 embryos. (B) Heatmap of 2539 genes differentially expressed genes in wild-type, tw18/+ and tw18/tw18 embryos with log2(FC)≥1. (C) GO term enrichment analyses of genes dysregulated between tw18/tw18 and wild-type log2(FC≥1). GO terms were analyzed for downregulated (upper panel) and upregulated (lower panel) genes. The diagrams shows selected GO terms with corresponding −Log10 of the P-value that were given by the Ontologizer tool. A background of genes expressed in the dataset with FPKM 2 was used. (D) Examples of Nodal and WNT targets downregulated in tw18 homozygotes; BMP targets were not affected (n.a.).
Among the most strongly downregulated genes in tw18/tw18 mutants is the pan-mesodermal marker Brachyury [T, log2(FC)=−4.3], indicating that mesoderm formation is severely compromised (Wilkinson et al., 1990; Arnold et al., 2000; Kubo et al., 2004). Other downregulated mesoderm control genes comprise Eomesodermin (Eomes, Russ et al., 2000), Mesp1 (Saga et al., 1996), Mesp2 (Saga et al., 1997), Mixl1 (Pearce and Evans, 1999), and the signal molecule Fgf8 (Crossley and Martin, 1995). A gene ontology (GO) term analysis revealed an enrichment of terms related to cellular signaling, mesoderm formation and cell fate specification among the downregulated genes, while biological adhesion was the most significant term related to upregulated genes (Fig. 1C). These data are consistent with the severe gastrulation defects observed previously in homozygous tw18 embryos (Bennett and Dunn, 1960).
Since the GO analysis pointed to impaired signaling, we searched the list of dysregulated genes for factors involved in signaling pathways. We found that targets of Nodal signaling in the epiblast, including Tdgf1 (Cripto, Brennan et al., 2001; Beck et al., 2002), Gsc (Toyama et al., 1995; Jones et al., 1995), and Lefty2 (Schier and Shen, 2000), are downregulated in tw18/tw18 embryos (Fig. 1D). Expression of Nodal itself, the Nodal convertases Furin (Spc1) and Pcsk6 (Spc4, Beck et al., 2002), and Smad2/3 (Kumar et al., 2001), as well as Smad4 (Lagna et al., 1996), the intracellular transmitters of Nodal signaling, are not affected (Table S1). However, Cer1 (Piccolo et al., 1999), Hesx1 (Thomas et al., 1995), and Hhex (Keng et al., 1998), markers of the anterior visceral endoderm (AVE) are upregulated, indicating that an anterior pole might be established. The fact that the latter markers are also dependent on Nodal signaling suggests that Nodal signaling is not generally impaired, but primarily in the epiblast.
Similarly, the WNT target genes T (Arnold et al., 2000), Sp5 (Weidinger et al., 2005), and Axin2 (Jho et al., 2002; Lustig et al., 2002) are downregulated, indicating that WNT signaling in the epiblast is also affected in the mutant. Expression of the Wnt3 morphogen itself and the final effector β-catenin, encoded by Ctnnb1, are not significantly dysregulated (Table S1).
Since Nodal and WNT signaling are required for pattern formation, formation of the PS, and mesoderm induction, the molecular data suggested that all these essential embryonic processes are impaired in tw18/tw18 embryos (Schier and Shen, 2000).
In contrast to WNT and Nodal signaling, dysregulation of bone morphogenetic protein (BMP) target genes, such as Wnt3 (Ben-Haim et al., 2006), Ifitm3 (Fragilis, Saitou et al., 2002), Id1/Id3 (Hollnagel et al., 1999), Smad6 (Ishida et al., 2000), or Smad7 (Benchabane and Wrana, 2003) was not apparent in the transcriptome data from mutant embryos (Fig. 1D). Moreover, Bmp2 is more strongly expressed in the mutant (Fig. 1B). BMP signaling is the major inducer of extra-embryonic and lateral mesoderm (Tonegawa et al., 1997; Arnold et al., 2006). Among the most highly upregulated mesodermal genes is the TGFβ co-receptor Endoglin (Eng), which is involved in extra-embryonic mesoderm formation and vasculogenesis (Arthur et al., 2000; Jonker and Arthur, 2001). Among the lateral mesoderm regulators several Gata factors (Gata2, Gata3), Hand1 and Hand2 are more highly expressed in the mutant transcriptome. These data indicate that mesoderm formation downstream of BMP signaling appears unaffected in the mutant.
We also observed that the pluripotency genes Nanog, Oct4 and Sox2, which are expressed in the epiblast (Nichols et al., 1998; Avilion et al., 2003; Mitsui et al., 2003), are expressed at a lower level in the mutants.
In summary, the transcriptome data indicated that the tw18 mutants are characterized by impaired Nodal and WNT signaling downstream of the ligands.
Homozygous tw18 embryos are disorganized
Transcriptome data per se do not provide spatial resolution, and a reduction or increase of marker gene expression at the transcriptome level does not allow us to distinguish whether the expression per cell or the relative number of cells expressing the marker is altered. Thus, to characterize the etiology of the tw18 phenotype in embryos we performed whole-mount in situ hybridizations (WISHs) of marker genes on E6.5, E7.5, E8.5, and E9.5 embryos derived from inter-crosses of heterozygous tw18 animals.
At E6.5, PS formation is initiated by activities of Nodal and Wnt3 signaling in the proximal posterior epiblast in concert with BMP signaling from the extraembryonic ectoderm (reviewed in Tam and Loebel, 2007). In all embryos derived from inter-crosses, Nodal and Wnt3 mRNA showed the wild-type pattern according to the individual embryonic stage [mutant embryos were slightly younger (E6.0)] (Fig. 2A-B′). T expression in wild-type and heterozygous embryos overlapped with Wnt3, as expected (Fig. 2C). In contrast, T expression was undetectable (Fig. 2C′, n=3/4) or severely downregulated in tw18 homozygotes (Fig. 2C″, n=1/4). Thus, the WISH data show that the signal molecules Nodal and Wnt3, which are essential for gastrulation and mesoderm formation, are expressed at their correct position in the tw18 mutant. In contrast, activation of the downstream effector T is insufficient, indicating a deficiency in mesoderm formation.
Fig. 2.

Marker gene analyses by in situ hybridization highlight primitive streak and patterning defects in tw18 mutant embryos. (A-L″) WISH analysis of E6.5 (A-C″), E7.5 (D-H″), E8.5 (I-L′), and E9.5 (M-M″) tw18 heterozygous (tw18/+) and homozygous (tw18/tw18) mouse embryos showing the expression of Nodal (A-A′), Wnt3 (B-B′), T (C-C′, F-F″, J-J′), Foxa2 (D-D″, H-H′), Sox2 (E-E″, I-I′), Tbx6 (G-G″, L-L′), Cer1 (H-H″) and Shh (M-M″). Scale bars: 250 µM in A-G″ and I-M″, 200 µM in H-H″.
From E7.5 onwards, homozygous tw18 embryos can be identified morphologically by growth-retardation compared to wild-type and heterozygous littermates. At this stage, Foxa2 marks the AVE and the node (Monaghan et al., 1993; Ang and Rossant, 1994; Perea-Gomez et al., 1999) (Fig. 2D). In homozygous tw18 embryos Foxa2 expression was detectable only in superficial cells probably representing the VE (Fig. 2D′,D″). In general, Foxa2 transcripts appeared more widely expressed in homozygotes, with enrichment at either the distal tip or the presumed anterior pole. At E7.5, Sox2 marks the future brain and the chorion (Wood and Episkopou, 1999; Avilion et al., 2003) (Fig. 2E). In tw18 homozygotes, embryonic Sox2 expression was either not detectable or reduced and without a clear anterior restriction (Fig. 2E′,E″). Extraembryonic ectodermal expression was detectable in all mutants but appeared as a band-like domain at the border to the epiblast (n=5). T expression was still either undetectable (Fig. 2F′, n=2/3) or faint (Fig. 2F″, n=1/3). Tbx6, a T target and marker of the paraxial mesoderm (Chapman et al., 1996, 2003) (Fig. 2G) was also only faintly expressed at the posterior end (Fig. 2G′, n=5/6) or undetectable (Fig. 2G″, n=1/6). In wild-type embryos Cer1 was expressed in the AVE, as described (Fig. 2H) (Biben et al., 1998). Similarly, Cer1 was detectable in the AVE of tw18 homozygotes (Fig. 2H′,H″). In addition, in one of the mutants expression was detectable in migrating mesendoderm, as previously observed in early primitive streak-stage wild-type embryos (Fig. 2H′, n=1/2; see Fig. 4A in Biben et al., 1998).
Taken together, these data suggest that a functional AVE is present in the mutant embryos, but neuroectodermal patterning, as well as mesoderm and extraembryonic tissue development, is compromised in tw18 homozygotes at E7.5.
E8.5 and older tw18 mutants remain growth retarded, spherical and are disorganized. Organogenesis and axial elongation, as observed in wild-type and heterozygous littermates, is not taking place. Accordingly, the expression of marker genes lacks proper patterning. Foxa2 was either undetectable (n=1/2, data not shown) or widely expressed in homozygotes (Fig. 2I′, n=1/2), and a small separate domain had formed at the distal tip of the embryo (Fig. 2I′ inlet), while heterozygous and wild-type littermates showed expression in the notochord and posterior endoderm. The pan-neural marker Sox2 was diffusely and widespread expressed in tw18 homozygotes (Fig. 2J′). The pan-mesodermal marker T was detectable in a patch of cells in some homozygotes (Fig. 2K′, n=2/3). Tbx6 was hardly detectable in tw18/tw18 embryos (Fig. 2L′, n=2/2).
At E9.5 Shh marks axial mesoderm and definitive endoderm (Martí et al., 1995) (Fig. 2M). In tw18 mutants, a single domain of Shh expression at the distal tip of some embryos (Fig. 2M′, n=2/4) or no staining was observed (Fig. 2M″, n=2/4). The single distal domain of Shh expression might represent a node-like domain, given that a similar domain was observed for Foxa2 expression at E8.5.
The marker expression patterns confirm that mesoderm formation (T), in particular paraxial mesoderm (Tbx6) and notochord development (Foxa2, T), remains impaired in E8.5 tw18 homozygotes. Since the notochord and the mesoderm are essential for cellular organization and organ development, proper organ structures cannot form in the mutant.
To further characterize tw18 mutants at the protein level, we performed whole-mount immunofluorescence staining for markers of pluripotency and differentiation on E7.5 embryos. The pluripotency factor POU5F1 (OCT4) is strongly expressed in the entire epiblast and quickly downregulated in differentiating cells of wild-type and tw18 heterozygous embryos (Yeom et al., 1996; Yoshimizu et al., 1999; Hoffman et al., 2013) (Fig. 3A and data not shown). In tw18 mutant embryos, the epiblast was small and malformed, but also OCT4 positive (Fig. 3A′).
Fig. 3.

Marker gene analyses by immunofluorescence staining highlight primitive streak and patterning defects in tw18 mutant embryos. (A-D″) E7.5 tw18 heterozygous and homozygous mouse embryo whole-mount immunofluorescence staining for OCT4 (A-A′), FOXA2 and NANOG (B-B′), EOMES (C-C′), SOX2 and SOX3 (D-D″). Left big panels: lateral view of the embryo (3D volume renderings). Right small panels: optical transversal sections for the section planes indicated by white dashed lines in the left panels. Counterstaining was performed with DAPI. Scale bar: 100 µm.
At E7.5, FOXA2 protein marks the visceral endoderm, in particular the AVE, and the node; NANOG marks the posterior half of the epiblast, and EOMES the PS and nascent mesoderm (Fig. 3B,C). In homozygous tw18 embryos, FOXA2 protein expression occurred in the right tissue (Fig. 3B′). An accumulation of strongly FOXA2-positive cells was detectable at the distal tip, in line with the node-like structure described before (Fig. 3B′, arrowhead in section plane c). NANOG protein expression was patchy and disorganized, restricted to the proximal epiblast, but without a posterior restriction (Fig. 3B′, section plane a). EOMES was weakly detectable in a patch of cells located at the presumed posterior end of a tw18 mutant embryo, as part of a cell mass filling the amniotic cavity (Fig. 3C′).
With the onset of neurogenesis, the anterior epiblast expresses the neural control factors SOX2 and SOX3 (Wood and Episkopou, 1999) (Fig. 3C). Homozygous tw18 embryos either expressed SOX2 and SOX3 with an anterior restriction (Fig. 3C′, n=1/2), or SOX2 protein was hardly detectable, while SOX3 was detected throughout the epiblast (Fig. 3C″, n=1/2). Accordingly, in tw18 mutants neurogenesis appears to be initiated, while an anterior restriction does not occur in all cases.
Mesoderm formation involves epithelial to mesenchymal transition (EMT), a multistep process involving downregulation of E-cadherin (E-cad; Cdh1; Nakaya et al., 2008). In parallel, after uniform expression in the epiblast at E6.5, N-cadherin is up regulated (N-cad; Cdh2) in migrating mesodermal cells (Maeda et al., 2005; Wheelock et al., 2008; Bao et al., 2009) (Fig. 4A). In sections of tw18 homozygous embryos, N-cad expression was not detectable, while E-cad protein was observed throughout the thickened epithelium (Fig. 4A′).
Fig. 4.
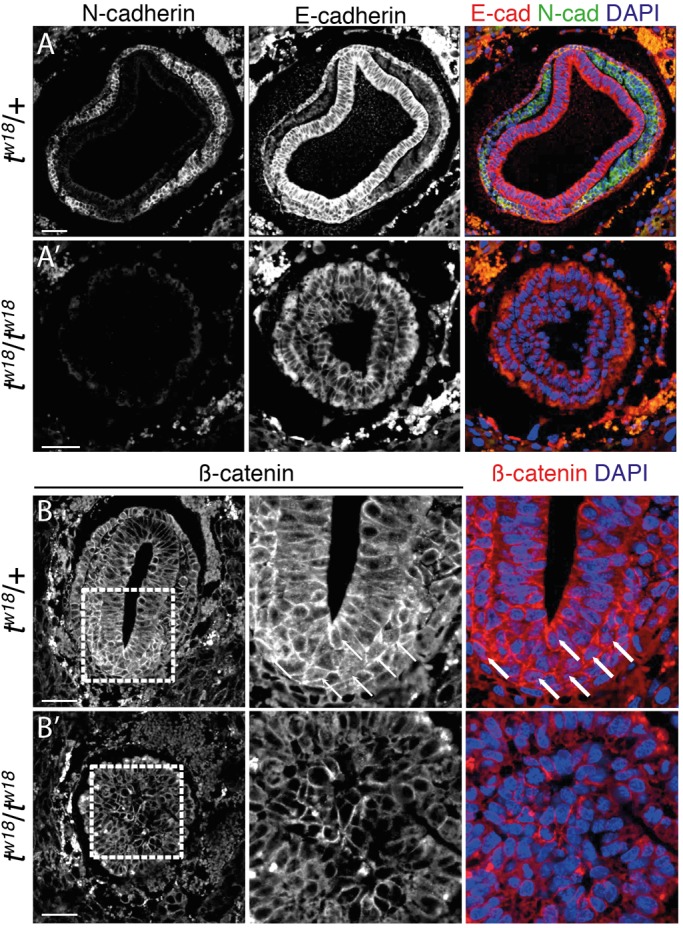
tw18 mutant embryos are unable to undergo EMT. (A-B′) Immunofluorescence staining on transversal paraffin sections of E7.5 tw18 heterozygous (tw18/+) (A,B) and homozygous (tw18/tw18) (A′,B′) mouse embryos within the deciduum. Sections were probed for either E-Cadherin and N-Cadherin (A-A′) or β-Catenin (B,B′), and counterstained with DAPI. Dashed rectangles in B and B′ indicate regions magnified in the neighboring panels. Arrows in B point to some exemplary cells with nuclear β-Catenin staining. Scale bar: 50 µm.
Another fundamental component of the adherens junctional complex is β-catenin (Ctnnb1; β-cat), which provides the intracellular link to the actin cytoskeleton, but also functions in the canonical WNT signaling pathway (Hulsken et al., 1994; reviewed in Kemler, 1993; Peifer, 1995). Accordingly, in sections of wild-type (not shown) and tw18 heterozygous embryos, we observed β-cat at the membrane and also in nuclei at the PS (Fig. 4B). In sections of tw18 homozygotes, β-cat was detected only at the membrane of epiblast cells and of cells filling the amniotic cavity (Fig. 4B′). These findings suggest that mutant cells retain epithelial characteristics, are unable to undergo EMT, and therefore accumulate at the site of the PS and bulge into the amniotic cavity.
The combined transcriptome, WISH, and immunofluorescence data provide a sufficiently detailed picture of the events leading to the tw18 mutant phenotype. While the Nodal and Wnt3 signal molecules are properly expressed at the future posterior pole, the Nodal and WNT signal pathways in the epiblast are impaired and thus their targets are not or insufficiently expressed. The AVE is established in the mutants allowing neural induction at the anterior pole. PS induction by BMP signaling is likely to be initiated, and at E8.5 a node-like domain is present at the distal tip of some mutant embryos. However, notochord formation does not occur, mesoderm formation is impaired, and the embryo is disorganized. Since EMT does not occur either, proliferating cells in the posterior epiblast eventually fill the (pro)amniotic cavity in some of the embryos. However, some mesoderm formation downstream of BMP signaling appears to take place. There is also considerable variation in the penetrance of the mutant phenotype, which might be due to inhomogeneous (non-inbred) genetic backgrounds between embryos causing variable expression of the phenotype in individual embryos.
The gastrulation defects of homozygous tw18 mutants are caused by loss of Ppp2r1a
Next we attempted to identify the gene responsible for the tw18 mutant phenotype. The tw18 haplotype is characterized by a large deletion of at least 3.3 Mb of the t complex on Chr17 (Búcan et al., 1987; Barclay et al., 1996). Since all protein-coding genes have been identified by genome sequencing, we decided to take a candidate gene approach to identify the tw18 lethal. This required us to determine the breakpoints and exact extent of the deletion, and thus all genes deleted. We attempted to generate tw18 homozygous embryonic stem cell (ESC) lines, which failed. Therefore we sequenced the genomic DNA of heterozygous tw18/+ by massive parallel sequencing. Read mapping to Chr17 revealed a sharp increase of the read density at position 23.6 Mb marking the distal deletion breakpoint (Fig. 5A, asterisk). On the centromere-close side only an approximate interval of the deletion breakpoint could be demarcated between Chr 17:18.0-19.7 Mb (Fig. 5A, red line). To position the breakpoint more precisely, we performed polymerase chain reaction (PCR) analyses on genomic DNA derived from heterozygous tw18/+ and homozygous tw18/tw18 embryos. Amplicons could be generated until position Chr17:19190492 in both DNA samples (Fig. 5B, 18.48-19.19 Mb), whereas fragments from Chr17:19330513 on could not be amplified from tw18/tw18 genomic DNA (Fig. 5B, 19.33 and 19.41 Mb, lower panel). Thus, we defined the region between Chr17:19190492-19330513 and 23.6 Mb, an interval of 4.3 Mb, as the deletion of the tw18 haplotype carrying the tw18 lethal.
Fig. 5.

The tw18 deletion extends over 4.3 Mbp of Chromosome 17. (A) Number of reads obtained by next generation sequencing of tw18/+ derived genomic DNA mapped to position Chr 17:17000000-25090000. The 3′ end of the deletion is found approximately at position Chr 17:23600000 (asterisk) whereas the 5′ end was roughly positioned between Chr 17: 18.4-19.4 Mb. (B) The presence of DNA elements around the region determined to contain the proximal breakpoint were analyzed by PCR. DNA fragments obtained by PCR amplification and visualized by gel electrophoresis; homozygous tw18 genomic DNA (lower panel) was amplified up to position Chr 17:19.19 Mb, whereas amplicons were absent from position Chr 17:19.33 Mb, indicating that the deletion starts between position 19.19 and 19.33 Mb. (C) Rough functional assignment of the 74 genes deleted in tw18.
The entire deletion contains 74 genes (Fig. 5C; Table S2). We excluded the 34 vomeronasal receptor genes and the non-coding RNAs as likely candidates, and searched the databases and literature for information on the remaining 25 candidates. For most of the 20 zinc finger genes the available data provided no information on expression or function.
The gene encoding the PP2A scaffolding subunit Aα, Ppp2r1a (Mayer-Jaekel and Hemmings, 1994) was selected as the most likely candidate, since PP2A plays an important role in signal transduction. In addition, homozygous loss of the catalytic subunit Cα (Ppp2ca) of PP2A has been shown to cause mesodermal defects and embryonic lethality (Götz et al., 1998). However, PPP2R1A can be substituted in the PP2A complex by PPP2R1B, and it was uncertain if loss of Ppp2r1a would be complemented by Ppp2r1b in the gastrulating embryo. To test this we generated a Ppp2r1a knockout in G4 ESCs (George et al., 2007) using a construct provided by the KOMP consortium (Fig. 6A). We screened for homologous recombination by using Southern blot analysis (Fig. 6B). We generated heterozygous mice and established a multiplex PCR approach to facilitate genotyping of mutants (Fig. 6C).
Fig. 6.

Targeting of the Ppp2r1a locus by homologous recombination. (A) Ppp2r1a targeting construct integrated into the wild-type Ppp2r1a locus (upper panel) by homologous recombination (lower panel). The stop cassette in intron 2 leads to abrogation of transcription. (B) Southern blot analysis of a successfully targeted ESC clone; a BamHI digest reveals a wild-type fragment of 13 kb, the targeted allele shows a 7 kb fragment with the 5′ and a 6.5 kb fragment with the 3′ external Southern probe. (C) PCR genotyping using three primers (P1, P2, P3) distinguishes between wild-type mice and mice carrying the targeted allele. (D) WISH analysis of embryos at E6.5 resulting from Ppp2r1a+/− inter-crosses shows absence of Ppp2r1a expression in homozygous Ppp2r1a−/− embryos. Scale bar: 200 µm.
To determine the expression pattern of Ppp2r1a in embryos and to validate the loss of function mutation, we performed WISH of Ppp2r1a RNA on E6.5 embryos obtained from Ppp2r1a+/− inter-crosses. Ppp2r1a mRNA is ubiquitously expressed in wild-type and heterozygous embryos (Fig. 6D). Homozygous littermates lacked the Ppp2r1a transcript, demonstrating that the knockout allele is a true null allele.
Next we tested if the Ppp2r1a knockout allele would complement the tw18 phenotype or cause a phenotype in embryos carrying both mutant alleles (Fig. 7A). Ppp2r1a+/− mice were mated with tw18/+ mice and embryos were dissected at E8.5. Embryos carrying tw18 and the Ppp2r1a knockout allele on the wild-type chromosome (tw18/+; Ppp2r1a−/−) showed growth retardation and no organ structures, while heterozygous littermates of either genotype had developed normally. These data showed that the Ppp2r1a knockout uncovers a developmental lethal within the tw18 deletion and does not complement the tw18 lethal phenotype. Next we asked if the mutant phenotype of the Ppp2r1a knockout and of tw18 homozygous embryos is similar (Fig. 7B). Histological sections of homozygous embryos showed that the amniotic cavity was filled with cells in both mutants at E7.5. This similarity of the phenotype suggested that the tw18 lethal phenotype indeed may be due to loss of Ppp2r1a.
Fig. 7.

Knockout of Ppp2r1a reconstitutes and Ppp2r1a expression rescues the tw18 homozygous phenotype. (A) Morphology of E8.5 embryos derived by crossing tw18/+ and Ppp2r1a+/− mice; embryos were genotyped by PCR following imaging. tw18/+;Ppp2r1a−/− embryos failed to gastrulate and resemble tw18 homozygotes. (B) Histological H&E-stained paraffin sections of E7.5 Ppp2r1a knockout and tw18/tw18 embryos. The amniotic cavities of both embryos are filled with cells. (C) Rescue construct containing the Ppp2r1a locus comprising 3 kb upstream and 20 kb downstream of the transcription start site (TSS). (D) Test for functioning of the Ppp2r1a rescue construct in ESCs. Ppp2r1a RNA expression analyzed by qPCR and normalized to Pmm2 and Hmbs (upper panel), PPP2R1A protein levels analyzed by western blotting (lower panel) followed by quantification (upper panel) relative to H3 protein. Expression of the rescue transgene increased Ppp2r1a mRNA and protein levels in the transgenic ESC clone. Error bars indicate standard deviation of technical replicates. (E) Ppp2r1a expression rescues the tw18 homozygous phenotype. Embryos from breedings of tw18/+ females with tw18/+, Tg(Ppp2r1a) males were analyzed at E10.5 and E18.5; homozygous tw18 embryos carrying the transgene Tg(Ppp2r1a) developed normally at least until E18.5. (F) qPCR analyses of E10.5 embryos from (E). Ppp2r1a expression levels were restored to 67% of the single wild-type allele of tw18 heterozygotes by the rescue construct. Error bars indicate standard deviation of technical replicates. (G-I″) WISH analysis of E7.5 Ppp2r1a+/− and Ppp2r1a−/− mouse embryos for the expression of Foxa2, Sox2, and T. Scale bar: 100 µm in A,G-I″ and 500 µm in E.
The morphological data, however, did not exclude that the tw18 deletion might contain another gene showing a similar phenotype upon loss of function, and that the tw18 homozygous phenotype may be the result of several gene losses. To exclude the latter possibility we performed a rescue experiment. We subcloned the Ppp2r1a gene including 3 kb upstream and 20 kb downstream of the TSS comprising the gene promoter and poly(A) signal via gap repair recombineering from a bacterial artificial chromosome (BAC) clone and introduced the linearized clone as a transgene randomly into tw18/+ ESCs (Fig. 7C). We analyzed Ppp2r1a mRNA levels by using quantitative PCR (qPCR) and analyzed protein levels by using western blotting in wild-type (F1G4), tw18/+ and transgenic ESCs (Fig. 7D). As expected, Ppp2r1a expression was around twofold higher in the wild-type cells compared to tw18/+ cells, which express only one allele. The transgenic construct increased the mRNA and protein expression of Ppp2r1a in transgenic ESCs to approximately the wild-type level, suggesting that it functions properly (Fig. 7D).
We established a mouse line from transgenic ESCs by morula aggregation (Eakin and Hadjantonakis, 2006). We mated transgenic mice with tw18 heterozygotes and isolated embryos at E10.5. The tw18 homozygous embryos, which had survived to this stage, did not show recognizable organ structures, while homozygotes carrying the transgene [tw18/tw18; Tg (Ppp2r1a)] were morphologically indistinguishable from heterozygotes (Fig. 7E). The rescue was also complete at E18.5 suggesting that the tw18 deletion contains no other genes besides Ppp2r1a, which are essential for embryogenesis (Fig. 7E). qPCR data confirmed the expression of Ppp2r1a mRNA in transgenic embryos homozygous for tw18 (Fig. 7F).
Thus, loss of Ppp2r1a is solely responsible for the embryonic lethal phenotype caused by the tw18 deletion.
To further confirm that the molecular phenotype of Ppp2r1a−/− embryos showed the same expression patterns of marker genes as tw18 homozygotes, we performed WISH on E7.5 embryos. The expression of Foxa2, Sox2, and T showed very similar dysregulation in Ppp2r1a−/− as in tw18 homozygotes at this stage, though the severity of the malformation was variable probably due to mixed genetic background of the mutants (compare Fig. 7G-I″ to Fig. 2D-F″).
In summary, we were able to demonstrate beyond doubt that the tw18 lethal is due to loss of Ppp2r1a.
DISCUSSION
The Brachyury (T) mutation was the first ‘classical’ lethal mutation identified, which affects mesoderm formation and axial development (Chesley and Dunn, 1936). Studying its properties led to the discovery of the mouse t haplotype and, over decades, to the identification of 16 recessive lethal mutations, the ‘t lethals’, causing early embryonic death (Ruvinsky et al., 1991; Bennett, 1975). Therefore, some four decades ago, the t complex (the wild-type form of the t haplotype) has been speculated to represent the prime genomic region encoding control genes for early embryonic development in the mouse. However, this proposition did not hold true. Attempts made in the following decades to isolate t lethals have been hampered by the difficulties to precisely localize the responsible genes within the t haplotype, and despite recent technical advances, prior to this report only a single t lethal, tw5, had been identified and molecularly characterized (Sugimoto et al., 2012).
Here we have shown that the t lethal tw18 is a loss-of-function mutation of the gene encoding the PP2A scaffold protein PPP2R1A, which is located within a large deletion of approximately 4.3 Mbp in the tw18 haplotype. Via genetic rescue of the tw18 deletion by a Ppp2r1a transgene we show that none of the other 73 genes including 20 zinc finger genes located in this deletion is essential for embryonic development. We provide a molecular characterization of the dysfunction leading to the gastrulation defects described for homozygous embryos of the t9 complementation group (t4, t9, tw18) about five decades ago (reviewed in Bennett, 1975).
The WNT and Nodal signaling pathways play essential roles in embryonic pattern formation, gastrulation and differentiation. Nodal and Wnt3 expression at the early streak stage (E6.0-E6.5) appeared largely unaffected in homozygous tw18 embryos, whereas their target genes were downregulated (Fig. 5D), suggesting defects in signal transduction of these pathways. Therefore, Ppp2r1a functions at some components of these signaling cascades.
PP2A activity is mediated by assembly of the core enzyme, consisting of the structural A subunit (PPP2R1A or PPP2R1B) and the catalytic C subunit (PPP2CA, PPP2CB), with a variable regulatory B subunit mediating substrate specificity (Sents et al., 2013). There are several lines of evidence for a role of PP2A in WNT signaling, both as a negative and a positive regulator of the cascade (reviewed in Eichhorn et al., 2009; Seeling et al., 1999; Li et al., 2001; Yamamoto et al., 2001). The downregulation of WNT targets in tw18 mutants provides good evidence that in the context of gastrulation of mouse embryos PP2A is required as an activator in the WNT signaling pathway.
Bajpai et al. (2004) showed that in Drosophila, loss of the homologue of a B subunit of PP2A, PR55, results in downregulation of WNT targets, suggesting a role of PP2A:PR55α in stabilizing cytoplasmic β-catenin. This role is supported by the finding that PP2A is required for β-catenin dephosphorylation thereby inhibiting its degradation in vivo (Zhang et al., 2009; Park et al., 2016). Next to the B regulatory subunits, also the scaffolding subunit itself might contribute to the regulation of the canonical WNT pathway. In malignant colon cancer cell lines, which display a highly activated WNT pathway, PPP2R1A is almost exclusively used to assemble the PP2A holoenzyme, while in normal colon cell lines, both the α and β scaffolds are present (Carmen Figueroa-Aldariz et al., 2015). Thus, it appears that PPP2R1A is preferred over PPP2R1B in regulating WNT signaling. In tw18 homozygous embryos, the lack of PPP2R1A might be partially complemented by the β scaffolding subunit, providing a possible explanation for the variable severity of the mutant phenotype.
The second developmentally important signaling cascade found to be dysregulated in tw18 homozygous embryos is the Nodal pathway. It was shown that the PR55α B regulatory subunit (PPP2R2A) can interact with type I TGFβ receptors (Griswold-Prenner et al., 1998). A later study showed that PR55α (PPP2R2A) and the closely related B regulatory subunit PR55δ (PPP2R2D) exert opposing effects on the TGFβ/Activin/Nodal pathway by regulating type I receptor stability and activity (Batut et al., 2008). Knockdown of PR55α or overexpression of PR55δ in Xenopus embryos results in anteroposterior axis truncation, resembling phenotypes of mutants with reduced Nodal signaling. Indeed, expression levels of the Activin/Nodal target genes, such as Gsc were strongly reduced in these embryos. However, the exact molecular mechanisms behind these effects have so far not been elucidated. Since a reduced expression of Nodal targets was observed in mouse embryos lacking Ppp2r1a, it is possible that PPP2R1A in complex with PR55α is required for positive regulation of Nodal signaling by stabilizing type I TGFβ receptors, as has been shown in Xenopus. Another line of evidence comes from the knockout phenotype of the Activin receptor type IIa in combination with a heterozygous type IIb or a heterozygous Nodal mutation, which have been shown to cause growth arrest at the egg cylinder stage and to affect PS and mesoderm formation, reminiscent of the tw18 lethal phenotype (Song et al., 1999). Thus, the growth retardation observed in tw18 homozygous embryos might relate to impaired Nodal signaling in the mutant epiblast. The combined data provide compelling evidence for an essential role of Ppp2r1a in Nodal signal transduction during gastrulation of the mouse embryo. The spatiotemporal expression of the different PP2A subunits during early embryogenesis and how the type I receptor activity is affected in tw18 embryos requires further investigation.
Besides dysregulation of the WNT and Nodal signaling cascades, the gene expression profile of E6.5 homozygous tw18 embryos showed an upregulation of genes involved in cell adhesion. Mutant cells move towards the streak but then fail to delaminate and become stalled in that region. Under wild-type conditions, cells ingressing through the PS undergo EMT, which includes a switch from E-cad to N-cad expression that provides cells with a migratory behavior (Maeda et al., 2005; Wheelock et al., 2008). In our transcriptome data Eomes expression was downregulated in tw18 mutants. Arnold et al. (2008) showed that mutants lacking Eomes specifically in the epiblast display defects in EMT and maintenance of E-cad expression. We identified a similar persistence of E-cad, accompanied by a lack of N-cad in the embryo proper of E7.5 tw18 homozygotes (Fig. 4A-B″). Thus, the defects in EMT observed in tw18 mutant embryos might be secondary to the downregulation of Eomes.
The exact molecular mechanisms by which PPP2R1A contributes to the regulation of the WNT and Nodal signaling pathways, and in particular the protein targets of PP2A in these signaling pathways, remain to be investigated.
MATERIAL AND METHODS
Mice and embryos
Mice were maintained in the animal facility of the Max Planck Institute for Molecular Genetics, Berlin, in accordance with international standards and protocols. Animal maintenance and all procedures performed on mice described here were performed in accordance with the German Animal Welfare Act (Tierschutzgesetz) and had prior approval from local authorities (LAGeSo).
Generation of ESC lines and transgenic mouse lines
We generated a tw18/+ embryonic stem cell line according to standard procedures (Behringer et al., 2014). Blastocysts were obtained from matings of tw18/+ males kept on a 129/SvEv genetic background to C57BL/6 females. To knock out Ppp2r1a G4 ESCs (George et al., 2007) were targeted by homologous recombination using a construct (PRPGS00108_A_B03) obtained from the KOMP repository. Clones were screened for successful homologous recombination by Southern blot analysis using external 5′ and 3′ probes. For the rescue experiment the transgene construct Tg(Ppp2r1a) containing only the Ppp2r1a locus was generated from a BAC (RPCI-23-209P21, Osoegawa et al., 2000) by Red/ET recombineering (Muyrers et al., 1999) and randomly integrated in tw18/+ ESCs. Successful integration was verified by PCR (see genotyping). Mouse lines were generated from transgenic ESCs by morula aggregation (Eakin and Hadjantonakis, 2006).
Genotyping
Mouse ear biopsies, embryos and sections were genotyped by PCR under standard conditions using the following primer pairs: tw18: Vil2s 5′-tcatggaccaacacaagctc-3′, Vil2as 5′-cacaaaactgaaatctccctc-3′(wt: 180 bp, tw18: 230 bp); Ppp2r1a KO alleles: fwd1 5′-gcaggagctggagcatgcca-3′, rev1 5′-tgcagcacaggatgttcagtgga-3′, rev2 5′-ggagagggacctggctcctatg-3′ (wt: 258 bp. KO: 436 bp); Tg(Ppp2r1a): fwd 5′-ttttgctgctgtaatggggacc-3′, rev 5′-gctaaagcgcatgctccaga-3′ (293 bp).
WISH
For WISH including synthesis of DIG-labeled antisense probes the protocol provided by the MAMEP database (http://mamep.molgen.mpg.de) was used. The probe for Ppp2r1a corresponds to nucleotides 930-1858 of NM_016891.3. Probe templates were amplified using standard PCR procedure with a reverse primer containing a T7 site for antisense transcription. In situ hybridizations for Foxa2 (clone name: UP001), T (clone name: 9226), Tbx6 (clone name: 9310), Sox2 (clone name: UL027), and Cer1 (clone name: CE232) were performed using probes from MAMEP. The Nodal probe template was kindly provided by the Robertson laboratory and linearized using BamHI. The Wnt3 probe template was kindly provided by the McMahon laboratory.
Histology
Whole deciduae were fixed in 4% paraformaldehyde (PFA)/phosphate-buffered saline (PBS) overnight, processed into paraffin and sectioned at 5 µm. Hematoxylin and eosin (H&E) staining was performed according to standard procedures. For immunofluorescence staining, sections were de-paraffinized using xylene and rehydrated in an ethanol serious. For antigen retrieval, sections were boiled in 0.01 M citrate buffer pH 6.0 for 10 min. Blocking was performed with 2% bovine serum albumin/2% lamb serum/PBS. The following antibodies were diluted in blocking solution and incubated overnight at 4°C: anti-E-cadherin (610181, BD Biosciences, 1/200), anti-N-cadherin (13116, CST, 1/100), and anti-β-catenin (610153, BD Biosciences, 1/200). Sections were incubated with the secondary antibodies for 1 h at room temperature (RT): anti-rabbit IgG-Alexa488 conjugated (A-11034, Thermo Fisher Scientific, 1/500) and anti-mouse IgG-Alexa546 conjugated (A-11030, Thermo Fisher Scientific, 1/500). Sections were mounted with VECTASHIELD HardSet Mounting Medium with DAPI (Vector Laboratories).
Whole-mount immunofluorescence staining and clearing
For whole-mount immunofluorescence, E7.5 embryos were dissected in ice-cold PBS and processed according to the protocol provided by Abcam (www.abcam.com). Primary antibodies were applied for 48-72 h in blocking solution: anti-OCT4 (sc-8628, Santa-Cruz Biotechnology, 1:200), anti-Sox2 (AF2018, R&D systems, 1:200), anti-Sox3 (provided by L. Gunhaga, Umeå Centre for Molecular Medicine, Umeå University, Sweden, 1:200) anti-Nanog (ab80892, Abcam, 1/100) anti-FoxA2 (sc-6554, Santa Cruz Biotechnology, 1/200), and anti-Eomes (ab23345, Abcam, 1/200). Secondary antibodies were applied in blocking solution overnight: anti-rabbit IgG-Alexa488 conjugated (A-21206, Thermo Fisher Scientific, 1/200) and anti-goat IgG-Alexa594 conjugated (A-11058, Thermo Fisher Scientific, 1/500). For clearing, specimens were postfixed in PBS/4% PFA for 20 min at 4°C, rinsed twice in 0.02 M phosphate buffer (PB) pH 7.4 and incubated in refraction index matching medium (RIMS, Yang et al., 2014) until transparency was reached. Before lightsheet imaging, the samples were embedded in PBS/1.5% low-melting point agarose and incubated in RIMS for another 24 h.
Deletion mapping by next-generation DNA sequencing and PCR
Genomic DNA was extracted from wild-type and tw18/+ ESCs. Sequencing was performed using the Illumina HiSeq 2000 system (shotgun sequencing) for 36 bp single reads that were mapped to the mouse genome mm10. PCR according to standard procedures was performed using the following primers: 18.5 Mb fwd 5′-agtccgaggccacgtccttga-3′, rev 5′-gctgcagttgctgtgctggc-3′; 18.6 Mb fwd 5′-ggccttccattcggtcctggg-3′, rev 5′-aggagtgctgtgctggacgc-3′; 18.7 Mb fwd 5′-tcttccccagaggacccggc-3′, rev 5′-gcagctgcagtggtgggcag-3′; 19.1 Mb fwd 5′-gccaacctgagaacgcgctg-3′, rev 5′-agggaaatgcctggggcagga-3′; 19.2 Mb fwd 5′-gccggccagcagctcacata-3′, rev 5′-ggcagcgtgccccactgaat-3′; 19.3 Mb fwd 5′-ccccggcatatcctgcgtagatga-3′, rev 5′-gggtgatgggcgagcaacaaatca-3′; 19.4 Mb fwd 5′-agccccatgcccagcgacta-3′, rev 5′-agtggctgtctggcccacca-3′.
RNA extraction, qRT-PCR and RNA-seq
RNA was extracted from cells or embryo tissues using the RNeasy Mini or Micro Kits (Qiagen). cDNA was synthesized from total RNA using the QuantiTect® Reverse Transcription Kit (Qiagen). qPCR was performed using the GoTaq® qPCR Master Mix (Promega) on the StepOnePlus Real-Time PCR System (Life Technologies). The following primer pairs were used: Ppp2r1a: fwd 5′-ccaccaagcacatgctgccc-3′, rev 5′-tccacatcctggtcctgggtca-3′; Pmm2: fwd 5′-agggaaaggcctcacgttct-3′; rev 5′-aataccgcttatcccatccttca-3′.
For RNA-seq the TotalScript™ RNA-Seq Kit (epicenter) was used for cDNA synthesis and library preparation. The sequencing was performed using the Illumina HiSeq 2500 (High Output) system for 50 bp paired end reads. Reads were mapped to the mouse genome mm10 (GRCm38) with TopHat (Trapnell et al., 2009) (v2.0.8b) and Cuffdiff (Trapnell et al., 2010) was used to calculate normalized FPKM values. Heat maps of differentially expressed genes and PCC were generated using the R program (http://www.r-project.org).
Immunoblotting
Whole-cell lysates were prepared using Western lysis buffer (1% SDS, 2 mM ethylenediaminetetraacetic acid, 10 mM Tris; pH 8.0). Immunoblotting was performed following standard protocols with anti-Ppp2r1a (6F9, Abcam, 1:1000) and anti-H3 (ab1791, Abcam, 1:10,000) antibodies. Horseradish peroxidase-linked secondary antibodies were obtained from Cell Signaling Technologies. Signals were imaged and quantified using the Fusion SL Vilber Lourmat system (Peqlab).
Imaging
For imaging of WISH-stained embryos, a MZ16A dissection microscope (Leica) fitted with an AxioCam MRc5 (Carl Zeiss MicroImaging) in combination with AxioVision software (Carl Zeiss MicroImaging), and a Zeiss Axio Zoom.V16 in combination with an Axiocam 512 color and ZEN software (Carl Zeiss MicroImaging), were used. H&E-stained paraffin sections were imaged using a Zeiss Observer.Z1 microscope fitted with an AxioCam MRc (Carl Zeiss MicroImaging) and AxioVision software. Immunofluorescence stainings on paraffin sections were imaged using an LSM 710NLO (Carl Zeiss MicroImaging) and ZEN software (Carl Zeiss MicroImaging). Immunofluorescent embryos were imaged in RIMS (Yang et al., 2014) using a Lightsheet Z.1 (Carl Zeiss MicroImaging) and ZEN software.
Acknowledgements
The vector used for this research project was generated by the trans-National Institutes of Health (NIH) Knockout Mouse Project (KOMP) and obtained from the KOMP Repository (www.komp.org). NIH grants to Velocigene at Regeneron Inc (U01HG004085) and the CSD Consortium (U01HG004080) funded the generation of gene-targeted ESCs for 8500 genes in the KOMP Program and archived and distributed by the KOMP Repository at UC Davis and CHORI (U42RR024244). We thank Andras Nagy for G4 ESCs. We further thank Anna Dorothea Senft and Elizabeth J. Robertson for providing the template for the mouse Nodal in situ probe, Jill and Andrew McMahon for the Wnt3 probe, and Cedric Patthey and Lena Gunhaga for SOX3 antibody. We thank Martin Werber for mapping of sequencing reads for the deletion analysis, Frederic Koch for help with RNA-seq library construction and GO analyses, Manuela Scholze for support in embryo dissection, Judith Fiedler and Karol Macura for generation of chimeric embryos, all members of the Sequencing Core facility, and Carolin Willke and Sonja Banko for animal husbandry.
Footnotes
Competing interests
The authors declare no competing or financial interests.
Author contributions
Conceptualization: L.L., M.M., L.W., H.B., B.G.H.; Methodology: B.G.H.; Formal analysis: L.L., M.M., J.L., L.W., H.B., B.G.H.; Investigation: L.L., M.M., J.L., L.W., H.B., S.P., G.B., B.T., B.G.H.; Writing - original draft: L.L., M.M., B.G.H.; Writing - review & editing: L.L., M.M., L.W., B.G.H.; Supervision: B.G.H.
Funding
This work was funded by the Max-Planck Society (Max-Planck-Gesellschaft).
Data availability
The RNA-seq data are accessible at GEO (GSE89734).
Supplementary information
Supplementary information available online at http://bio.biologists.org/lookup/doi/10.1242/bio.023200.supplemental
References
- Ang S.-L. and Rossant J. (1994). HNF-3β is essential for node and notochord formation in mouse development. Cell 78, 561-574. 10.1016/0092-8674(94)90522-3 [DOI] [PubMed] [Google Scholar]
- Arnold S. J. and Robertson E. J. (2009). Making a commitment: cell lineage allocation and axis patterning in the early mouse embryo. Nat. Rev. Mol. Cell Biol. 10, 91-103. 10.1038/nrm2618 [DOI] [PubMed] [Google Scholar]
- Arnold S. J., Stappert J., Bauer A., Kispert A., Herrmann B. G. and Kemler R. (2000). Brachyury is a target gene of the Wnt/β-catenin signaling pathway. Mech. Dev. 91, 249-258. 10.1016/S0925-4773(99)00309-3 [DOI] [PubMed] [Google Scholar]
- Arnold S. J., Maretto S., Islam A., Bikoff E. K. and Robertson E. J. (2006). Dose-dependent Smad1, Smad5 and Smad8 signaling in the early mouse embryo. Dev. Biol. 296, 104-118. 10.1016/j.ydbio.2006.04.442 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arnold S. J., Hofmann U. K., Bikoff E. K. and Robertson E. J. (2008). Pivotal roles for eomesodermin during axis formation, epithelium-to-mesenchyme transition and endoderm specification in the mouse. Development 135, 501-511. 10.1242/dev.014357 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Arthur H. M., Ure J., Smith A. J. H., Renforth G., Wilson D. I., Torsney E., Charlton R., Parums D. V., Jowett T., Marchuk D. A. et al. (2000). Endoglin, an ancillary TGFβ receptor, is required for extraembryonic angiogenesis and plays a key role in heart development. Dev. Biol. 217, 42-53. 10.1006/dbio.1999.9534 [DOI] [PubMed] [Google Scholar]
- Avilion A. A., Nicolis S. K., Pevny L. H., Perez L., Vivian N. and Lovell-Badge R. (2003). Multipotent cell lineages in early mouse development depend on SOX2 function. Genes Dev. 17, 126-140. 10.1101/gad.224503 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bajpai R., Makhijani K., Rao P. R. and Shashidhara L. S. (2004). Drosophila twins regulates armadillo levels in response to Wg/Wnt signal. Development 131, 1007-1016. 10.1242/dev.00980 [DOI] [PubMed] [Google Scholar]
- Bao S., Tang F., Li X., Hayashi K., Gillich A., Lao K. and Surani M. A. (2009). Epigenetic reversion of post-implantation epiblast to pluripotent embryonic stem cells. Nature 461, 1292-1295. 10.1038/nature08534 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barclay J., King T. F., Crossley P. H., Barnard R. C., Larin Z., Lehrach H. and Little P. F. R. (1996). Physical analysis of the region deleted in the tw18 allele of the mouse tcl-4 complementation group. Genomics 36, 39-46. 10.1006/geno.1996.0423 [DOI] [PubMed] [Google Scholar]
- Batut J., Schmierer B., Cao J., Raftery L. A., Hill C. S. and Howell M. (2008). Two highly related regulatory subunits of PP2A exert opposite effects on TGF-beta/Activin/Nodal signalling. Development 135, 2927-2937. 10.1242/dev.020842 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beck S., Le Good J. A., Guzman M., Ben Haim N., Roy K., Beermann F. and Constam D. B. (2002). Extraembryonic proteases regulate Nodal signalling during gastrulation. Nat. Cell Biol. 4, 981-985. 10.1038/ncb890 [DOI] [PubMed] [Google Scholar]
- Behringer R., Gertsenstein M., Vintersten Nagy K. and Nagy A. (2014). Manipulating the Mouse Embryo: A Laboratory Manual, 4th edn. New York: Cold Spring Habor Laboratory Press. [Google Scholar]
- Ben-Haim N., Lu C., Guzman-Ayala M., Pescatore L., Mesnard D., Bischofberger M., Naef F., Robertson E. J. and Constam D. B. (2006). The nodal precursor acting via activin receptors induces mesoderm by maintaining a source of its convertases and BMP4. Dev. Cell 11, 313-323. 10.1016/j.devcel.2006.07.005 [DOI] [PubMed] [Google Scholar]
- Benchabane H. and Wrana J. L. (2003). GATA- and Smad1-dependent enhancers in the Smad7 gene differentially interpret bone morphogenetic protein concentrations. Mol. Cell. Biol. 23, 6646-6661. 10.1128/MCB.23.18.6646-6661.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bennett D. (1975). The T-locus of the mouse. Cell 6, 441-454. 10.1016/0092-8674(75)90033-1 [DOI] [Google Scholar]
- Bennett D. and Dunn L. C. (1960). A lethal mutant (tw18) in the house mouse showing partial duplications. J. Exp. Zool. 143, 203-219. 10.1002/jez.1401430206 [DOI] [Google Scholar]
- Biben C., Stanley E., Fabri L., Kotecha S., Rhinn M., Drinkwater C., Lah M., Wang C.-C., Nash A., Hilton D. et al. (1998). Murine cerberus homologue mCer-1: a candidate anterior patterning molecule. Dev. Biol. 194, 135-151. 10.1006/dbio.1997.8812 [DOI] [PubMed] [Google Scholar]
- Brennan J., Lu C. C., Norris D. P., Rodriguez T. A., Beddington R. S. P. and Robertson E. J. (2001). Nodal signalling in the epiblast patterns the early mouse embryo. Nature 411, 965-969. 10.1038/35082103 [DOI] [PubMed] [Google Scholar]
- Búcan M., Herrmann B. G., Frischauf A. M., Bautch V. L., Bode V., Silver L. M., Martin G. R. and Lehrach H. (1987). Deletion and duplication of DNA sequences is associated with the embryonic lethal phenotype of the t9 complementation group of the mouse t complex. Genes Dev. 1, 376-385. 10.1101/gad.1.4.376 [DOI] [PubMed] [Google Scholar]
- Carmen Figueroa-Aldariz M., Castañeda-Patlán M. C., Santoyo-Ramos P., Zentella A. and Robles-Flores M. (2015). Protein phosphatase 2A is essential to maintain active Wnt signaling and its Aβ tumor suppressor subunit is not expressed in colon cancer cells. Mol. Carcinog. 54, 1430-1441. 10.1002/mc.22217 [DOI] [PubMed] [Google Scholar]
- Carroll L. S., Meagher S., Morrison L., Penn D. J. and Potts W. K. (2004). Fitness effects of a selfish gene (the Mus t complex) are revealed in an ecological context. Evolution (N Y) 58, 1318-1328. [DOI] [PubMed] [Google Scholar]
- Chapman D. L., Agulnik I., Hancock S., Silver L. M. and Papaioannou V. E. (1996). Tbx6, a mouse T-Box gene implicated in paraxial mesoderm formation at gastrulation. Dev. Biol. 180, 534-542. 10.1006/dbio.1996.0326 [DOI] [PubMed] [Google Scholar]
- Chapman D. L., Cooper-Morgan A., Harrelson Z. and Papaioannou V. E. (2003). Critical role for Tbx6 in mesoderm specification in the mouse embryo. Mech. Dev. 120, 837-847. 10.1016/S0925-4773(03)00066-2 [DOI] [PubMed] [Google Scholar]
- Chesley P. and Dunn L. C. (1936). The Inheritance of Taillessness (Anury) in the House Mouse. Genetics 21, 525-536. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crossley P. and Martin G. (1995). The mouse Fgf8 gene encodes a family of polypeptides and is expressed in regions that direct outgrowth and patterning in the developing embryo. Development 121, 439-451. [DOI] [PubMed] [Google Scholar]
- Eakin G. S. and Hadjantonakis A.-K. (2006). Production of chimeras by aggregation of embryonic stem cells with diploid or tetraploid mouse embryos. Nat. Protoc. 1, 1145-1153. 10.1038/nprot.2006.173 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Eichhorn P. J. A., Creyghton M. P. and Bernards R. (2009). Protein phosphatase 2A regulatory subunits and cancer. Biochim. Biophys. Acta Rev. Cancer 1795, 1-15. 10.1016/j.bbcan.2008.05.005 [DOI] [PubMed] [Google Scholar]
- George S. H. L., Gertsenstein M., Vintersten K., Korets-Smith E., Murphy J., Stevens M. E., Haigh J. J. and Nagy A. (2007). Developmental and adult phenotyping directly from mutant embryonic stem cells. Proc. Natl. Acad. Sci. USA 104, 4455-4460. 10.1073/pnas.0609277104 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Götz J., Probst A., Ehler E., Hemmings B., Kues W. (1998). Delayed embryonic lethality in mice lacking protein phosphatase 2A catalytic subunit Cα. PNAS 95, 12370-12375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griswold-Prenner I., Kamibayashi C., Maruoka E. M., Mumby M. C. and Derynck R. (1998). Physical and functional interactions between type I transforming growth factor beta receptors and Balpha, a WD-40 repeat subunit of phosphatase 2A. Mol. Cell. Biol. 18, 6595-6604. 10.1128/MCB.18.11.6595 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hoffman J. A., Wu C.-I. and Merrill B. J. (2013). Tcf7l1 prepares epiblast cells in the gastrulating mouse embryo for lineage specification. Development 140, 1665-1675. 10.1242/dev.087387 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hollnagel A., Oehlmann V., Heymer J., Rüther U. and Nordheim A. (1999). Id genes are direct targets of bone morphogenetic protein induction in embryonic stem cells. J. Biol. Chem. 274, 19838-19845. 10.1074/jbc.274.28.19838 [DOI] [PubMed] [Google Scholar]
- Hulsken J., Birchmeier W. and Behrens J. (1994). E-cadherin and APC compete for the interaction with beta-catenin and the cytoskeleton. J. Cell Biol. 127, 2061-2069. 10.1083/jcb.127.6.2061 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ishida W., Hamamoto T., Kusanagi K., Yagi K., Kawabata M., Takehara K., Sampath T. K., Kato M. and Miyazono K. (2000). Smad6 is a Smad1/5-induced Smad inhibitor. Characterization of bone morphogenetic protein-responsive element in the mouse Smad6 promoter. J. Biol. Chem. 275, 6075-6079. 10.1074/jbc.275.9.6075 [DOI] [PubMed] [Google Scholar]
- Janssens V. and Goris J. (2001). Protein phosphatase 2A: a highly regulated family of serine/threonine phosphatases implicated in cell growth and signalling. Biochem. J. 353, 417-439. 10.1042/bj3530417 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jho E., Zhang T., Domon C., Joo C.-K., Freund J.-N. and Costantini F. (2002). Wnt/beta-catenin/Tcf signaling induces the transcription of Axin2, a negative regulator of the signaling pathway. Mol. Cell. Biol. 22, 1172-1183. 10.1128/MCB.22.4.1172-1183.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jones C., Kuehn M., Hogan B., Smith J. and Wright C. (1995). Nodal-related signals induce axial mesoderm and dorsalize mesoderm during gastrulation. Development 121, 3651-3662. [DOI] [PubMed] [Google Scholar]
- Jonker L. and Arthur H. M. (2001). Endoglin expression in early development is associated with vasculogenesis and angiogenesis. Mech. Dev. 110, 193-196. 10.1016/S0925-4773(01)00562-7 [DOI] [PubMed] [Google Scholar]
- Kemler R. (1993). From cadherins to catenins: cytoplasmic protein interactions and regulation of cell adhesion. Trends Genet. 9, 317-321. 10.1016/0168-9525(93)90250-L [DOI] [PubMed] [Google Scholar]
- Keng V. W., Fujimori K. E., Myint Z., Tamamaki N., Nojyo Y. and Noguchi T. (1998). Expression of Hex mRNA in early murine postimplantation embryo development. FEBS Lett. 426, 183-186. 10.1016/S0014-5793(98)00342-1 [DOI] [PubMed] [Google Scholar]
- Klein J., Sipos P. and Figueroa F. (1984). Polymorphism of t-complex genes in European wild mice. Genet. Res. 44, 39-46. 10.1017/S0016672300026239 [DOI] [Google Scholar]
- Kubo A., Shinozaki K., Shannon J. M., Kouskoff V., Kennedy M., Woo S., Fehling H. J. and Keller G. (2004). Development of definitive endoderm from embryonic stem cells in culture. Development 131, 1651-1662. 10.1242/dev.01044 [DOI] [PubMed] [Google Scholar]
- Kumar A., Novoselov V., Celeste A. J., Wolfman N. M., ten Dijke P. and Kuehn M. R. (2001). Nodal signaling uses activin and transforming growth factor-beta receptor-regulated Smads. J. Biol. Chem. 276, 656-661. 10.1074/jbc.M004649200 [DOI] [PubMed] [Google Scholar]
- Lagna G., Hata A., Hemmati-Brivanlou A. and Massagué J. (1996). Partnership between DPC4 and SMAD proteins in TGF-beta signalling pathways. Nature 383, 832-836. 10.1038/383832a0 [DOI] [PubMed] [Google Scholar]
- Li X., Yost H. J., Virshup D. M. and Seeling J. M. (2001). Protein phosphatase 2A and its B56 regulatory subunit inhibit Wnt signaling in Xenopus. EMBO J. 20, 4122-4131. 10.1093/emboj/20.15.4122 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lustig B., Jerchow B., Sachs M., Weiler S., Pietsch T., Karsten U., van de Wetering M., Clevers H., Schlag P. M., Birchmeier W. et al. (2002). Negative feedback loop of Wnt signaling through upregulation of conductin/axin2 in colorectal and liver tumors. Mol. Cell. Biol. 22, 1184-1193. 10.1128/MCB.22.4.1184-1193.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Maeda M., Johnson K. R. and Wheelock M. J. (2005). Cadherin switching: essential for behavioral but not morphological changes during an epithelium-to-mesenchyme transition. J. Cell Sci. 118, 873-887. 10.1242/jcs.01634 [DOI] [PubMed] [Google Scholar]
- Marti E., Takada R., Bumcrot D. A., Sasaki H., McMahon A. P. (1995). Distribution of Sonic hedgehog peptides in the developing chick and mouse embryo. Development 121, 2537-2547. [DOI] [PubMed] [Google Scholar]
- Mayer-Jaekel R. E. and Hemmings B. A. (1994). Protein phosphatase 2A — a “ménage à trois”. Trends Cell Biol. 4, 287-291. 10.1016/0962-8924(94)90219-4 [DOI] [PubMed] [Google Scholar]
- Mitsui K., Tokuzawa Y., Itoh H., Segawa K., Murakami M., Takahashi K., Maruyama M., Maeda M., Yamanaka S. (2003). The Homeoprotein Nanog Is Required for Maintenance of Pluripotency in Mouse Epiblast and ES Cells. Cell 113, 631-642. [DOI] [PubMed] [Google Scholar]
- Monaghan A., Kaestner K., Grau E. and Schutz G. (1993). Postimplantation expression patterns indicate a role for the mouse forkhead/HNF-3 alpha, beta and gamma genes in determination of the definitive endoderm, chordamesoderm and neuroectoderm. Development 119, 567-578. [DOI] [PubMed] [Google Scholar]
- Moser G. C. and Gluecksohn-Waelsch S. (1967). Developmental genetics of a recessive allele at the complex T-locus in the mouse. Developmental Biology 16, 564-576. [DOI] [PubMed] [Google Scholar]
- Muyrers J. P. P., Zhang Y., Testa G. and Stewart A. F. (1999). Rapid modification of bacterial artificial chromosomes by ET-recombination. Nucleic Acids Res. 27, 1555-1557. 10.1093/nar/27.6.1555 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakaya Y., Sukowati E. W., Wu Y. and Sheng G. (2008). RhoA and microtubule dynamics control cell-basement membrane interaction in EMT during gastrulation. Nat. Cell Biol. 10, 765-775. 10.1038/ncb1739 [DOI] [PubMed] [Google Scholar]
- Nichols J., Zevnik B., Anastassiadis K., Niwa H., Klewe-Nibus D., Chambers I., Schöler H., Smith A. (1998). Formation of Pluripotent Stem Cells in the Mammalian Embryo Depends on the POU Transcription Factor Oct4. Cell 95, 379-391. [DOI] [PubMed] [Google Scholar]
- Osoegawa K., Tateno M., Woon P. Y., Frengen E., Mammoser A. G., Catanese J. J., Hayashizaki Y. and de Jong P. J. (2000). Bacterial artificial chromosome libraries for mouse sequencing and functional analysis. Genome Res. 10, 116-128. [PMC free article] [PubMed] [Google Scholar]
- Park D.-S., Yoon G.-H., Lee H.-S. and Choi S.-C. (2016). Capsaicin inhibits the Wnt/β-catenin signaling pathway by down-regulating PP2A. Biochem. Biophys. Res. Commun. 478, 455-461. [DOI] [PubMed] [Google Scholar]
- Pearce J. J. H. and Evans M. J. (1999). Mml, a mouse Mix-like gene expressed in the primitive streak. Mech. Dev. 87, 189-192. 10.1016/S0925-4773(99)00135-5 [DOI] [PubMed] [Google Scholar]
- Peifer M. (1995). Cell adhesion and signal transduction: the Armadillo connection. Trends Cell Biol. 5, 224-229. 10.1016/S0962-8924(00)89015-7 [DOI] [PubMed] [Google Scholar]
- Perea-Gomez A., Shawlot W., Sasaki H., Behringer R. and Ang S. (1999). HNF3beta and Lim1 interact in the visceral endoderm to regulate primitive streak formation and anterior-posterior polarity in the mouse embryo. Development 126, 4499-4511. [DOI] [PubMed] [Google Scholar]
- Piccolo S., Agius E., Leyns L., Bhattacharyya S., Grunz H., Bouwmeester T. and De Robertis E. M. (1999). The head inducer Cerberus is a multifunctional antagonist of Nodal, BMP and Wnt signals. Nature 397, 707-710. 10.1038/17820 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russ A. P., Wattler S., Colledge W. H., Aparicio S. A., Carlton M. B. L., Pearce J. J., Barton S. C., Surani M. A., Ryan K., Nehls M. C. et al. (2000). Eomesodermin is required for mouse trophoblast development and mesoderm formation. Nature 404, 95-99. 10.1038/35003601 [DOI] [PubMed] [Google Scholar]
- Ruvinsky A., Polyakov A., Agulnik A., Tichy H., Figueroa F. and Klein J. (1991). Low diversity of t haplotypes in the eastern form of the house mouse, Mus musculus L. Genetics 127, 161-168. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Saga Y., Hata N., Kobayashi S., Magnuson T., Seldin M. and Taketo M. (1996). MesP1: a novel basic helix-loop-helix protein expressed in the nascent mesodermal cells during mouse gastrulation. Development 122, 2769-2778. [DOI] [PubMed] [Google Scholar]
- Saga Y., Hata N., Koseki H. and Taketo M. M. (1997). Mesp2: a novel mouse gene expressed in the presegmented mesoderm and essential for segmentation initiation. Genes Dev. 11, 1827-1839. 10.1101/gad.11.14.1827 [DOI] [PubMed] [Google Scholar]
- Saitou M., Barton S. C. and Surani M. A. (2002). A molecular programme for the specification of germ cell fate in mice. Nature 418, 293-300. 10.1038/nature00927 [DOI] [PubMed] [Google Scholar]
- Schier A. F. and Shen M. M. (2000). Nodal signalling in vertebrate development. Nature 403, 385-389. 10.1038/35000126 [DOI] [PubMed] [Google Scholar]
- Seeling J. M., Miller J. R., Gil R., Moon R. T., White R., Virshup D. M., Cadigan K. M., Nusse R., Moon R. T., Miller J. R. et al. (1999). Regulation of beta-catenin signaling by the B56 subunit of protein phosphatase 2A. Science 283, 2089-2091. 10.1126/science.283.5410.2089 [DOI] [PubMed] [Google Scholar]
- Sents W., Ivanova E., Lambrecht C., Haesen D. and Janssens V. (2013). The biogenesis of active protein phosphatase 2A holoenzymes: a tightly regulated process creating phosphatase specificity. FEBS J. 280, 644-661. 10.1111/j.1742-4658.2012.08579.x [DOI] [PubMed] [Google Scholar]
- Silver L. M. (1985). Mouse tau haplotypes. Annu. Rev. Genet. 19, 179-208. 10.1146/annurev.ge.19.120185.001143 [DOI] [PubMed] [Google Scholar]
- Song J., Oh S. P., Schrewe H., Nomura M., Lei H., Okano M., Gridley T. and Li E. (1999). The type II activin receptors are essential for egg cylinder growth, gastrulation, and rostral head development in mice. Dev. Biol. 213, 157-169. 10.1006/dbio.1999.9370 [DOI] [PubMed] [Google Scholar]
- Sugimoto M. (2014). Developmental genetics of the mouse t-complex. Genes Genet. Syst. 89, 109-120. 10.1266/ggs.89.109 [DOI] [PubMed] [Google Scholar]
- Sugimoto M., Kondo M., Hirose M., Suzuki M., Mekada K., Abe T., Kiyonari H., Ogura A., Takagi N., Artzt K. et al. (2012). Molecular identification of tw5: Vps52 promotes pluripotential cell differentiation through cell-cell interactions. Cell Rep. 2, 1363-1374. 10.1016/j.celrep.2012.10.004 [DOI] [PubMed] [Google Scholar]
- Tam P. P. L. and Loebel D. A. F. (2007). Gene function in mouse embryogenesis: get set for gastrulation. Nat. Rev. Genet. 8, 368-381. 10.1038/nrg2084 [DOI] [PubMed] [Google Scholar]
- Thomas P. Q., Johnson B. V., Rathjen J. and Rathjen P. D. (1995). Sequence, genomic organization, and expression of the novel homeobox gene Hesx1. J. Biol. Chem. 270, 3869-3875. 10.1074/jbc.270.8.3869 [DOI] [PubMed] [Google Scholar]
- Tonegawa A., Funayama N., Ueno N., Takahashi Y., Aoyama H., Charlebois T. S., Henry J. J., Grainger R. M., Charlebois T. S., Spencer D. H. et al. (1997). Mesodermal subdivision along the mediolateral axis in chicken controlled by different concentrations of BMP-4. Development 124, 1975-1984. [DOI] [PubMed] [Google Scholar]
- Toyama R., O'Connell M. L., Wright C. V., Kuehn M. R. and Dawid I. B. (1995). Nodal induces ectopic goosecoid and lim1 expression and axis duplication in zebrafish. Development 121, 383-391. [DOI] [PubMed] [Google Scholar]
- Trapnell C., Pachter L., Salzberg S. L. (2009). TopHat: Discovering splice junctions with RNA-Seq. Bioinformatics 25, 1105-1111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Trapnell C., Williams B. A., Pertea G. et al. (2010). Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat Biotechnol 28, 511-515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vafai S. B. and Stock J. B. (2002). Protein phosphatase 2A methylation: a link between elevated plasma homocysteine and Alzheimer's Disease. FEBS Lett. 518, 1-4. 10.1016/S0014-5793(02)02702-3 [DOI] [PubMed] [Google Scholar]
- Virshup D. M. (2000). Protein phosphatase 2A: a panoply of enzymes. Curr. Opin. Cell Biol. 12, 180-185. 10.1016/S0955-0674(99)00074-5 [DOI] [PubMed] [Google Scholar]
- Weidinger G., Thorpe C. J., Wuennenberg-Stapleton K., Ngai J. and Moon R. T. (2005). The Sp1-related transcription factors sp5 and sp5-like act downstream of Wnt/beta-catenin signaling in mesoderm and neuroectoderm patterning. Curr. Biol. 15, 489-500. 10.1016/j.cub.2005.01.041 [DOI] [PubMed] [Google Scholar]
- Wheelock M. J., Shintani Y., Maeda M., Fukumoto Y. and Johnson K. R. (2008). Cadherin switching. J. Cell Sci. 121, 727-735. 10.1242/jcs.000455 [DOI] [PubMed] [Google Scholar]
- Wilkinson D. G., Bhatt S. and Herrmann B. G. (1990). Expression pattern of the mouse T gene and its role in mesoderm formation. Nature 343, 657-659. 10.1038/343657a0 [DOI] [PubMed] [Google Scholar]
- Wood H. B. and Episkopou V. (1999). Comparative expression of the mouse Sox1, Sox2 and Sox3 genes from pre-gastrulation to early somite stages. Mech. Dev. 86, 197-201. 10.1016/S0925-4773(99)00116-1 [DOI] [PubMed] [Google Scholar]
- Yamamoto H., Hinoi T., Michiue T., Fukui A., Usui H., Janssens V., Van Hoof C., Goris J., Asashima M. and Kikuchi A. (2001). Inhibition of the Wnt signaling pathway by the PR61 subunit of protein phosphatase 2A. J. Biol. Chem. 276, 26875-26882. 10.1074/jbc.M100443200 [DOI] [PubMed] [Google Scholar]
- Yang B., Treweek J. B., Kulkarni R. P., Deverman B. E., Chen C.-K., Lubeck E., Shah S., Cai L. and Gradinaru V. (2014). Single-Cell Phenotyping within Transparent Intact Tissue through Whole-Body Clearing. Cell 158, 945-958. 10.1016/j.cell.2014.07.017 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yeom Y. I., Fuhrmann G., Ovitt C. E., Brehm A., Ohbo K., Gross M., Hubner K. and Scholer H. R. (1996). Germline regulatory element of Oct-4 specific for the totipotent cycle of embryonal cells. Development 122, 881-894. [DOI] [PubMed] [Google Scholar]
- Yoshimizu T., Sugiyama N., De Felice M., Yeom Y. I., Ohbo K., Masuko K., Obinata M., Abe K., Scholer H. R. and Matsui Y. (1999). Germline-specific expression of the Oct-4/green fluorescent protein (GFP) transgene in mice. Dev. Growth Differ. 41, 675-684. 10.1046/j.1440-169x.1999.00474.x [DOI] [PubMed] [Google Scholar]
- Zhang W., Yang J., Liu Y., Chen X., Yu T., Jia J. and Liu C. (2009). PR55 alpha, a regulatory subunit of PP2A, specifically regulates PP2A-mediated beta-catenin dephosphorylation. J. Biol. Chem. 284, 22649-22656. 10.1074/jbc.M109.013698 [DOI] [PMC free article] [PubMed] [Google Scholar]