

Abstract
Background
The correct establishment of the human gut microbiota represents a crucial development that commences at birth. Different hypotheses propose that the infant gut microbiota is derived from, among other sources, the mother’s fecal/vaginal microbiota and human milk.
Results
The composition of bifidobacterial communities of 25 mother-infant pairs was investigated based on an internal transcribed spacer (ITS) approach, combined with cultivation-mediated and genomic analyses. We identified bifidobacterial strains/communities that are shared between mothers and their corresponding newborns. Notably, genomic analyses together with growth profiling assays revealed that bifidobacterial strains that had been isolated from human milk are genetically adapted to utilize human milk glycans. In addition, we identified particular bacteriophages specific of bifidobacterial species that are common in the viromes of mother and corresponding child.
Conclusions
This study highlights the transmission of bifidobacterial communities from the mother to her child and implies human milk as a potential vehicle to facilitate this acquisition. Furthermore, these data represent the first example of maternal inheritance of bifidobacterial phages, also known as bifidophages in infants following a vertical transmission route.
Electronic supplementary material
The online version of this article (doi:10.1186/s40168-017-0282-6) contains supplementary material, which is available to authorized users.
Keywords: Microbiota, Virome, Microbiome, Bifidobacteria, Vertical transmission
Background
The period immediately following birth is believed to be crucial for the correct establishment of the gut microbiota with possible temporary and long-lasting effects on host health [1]. During this infant stage of (human) life, microorganisms originating from the mother and from environmental microbial communities rapidly colonize the gastrointestinal tract (GIT) of the neonate to form the early infant microbiota whose establishment is also influenced by the feeding method [2–7]. Bifidobacteria are among the first colonizers of the intestine of newborns and are considered to play pivotal roles in terms of modulation of mucosal physiology and innate immunity of the host [6, 8, 9]. Genomically identical bifidobacterial strains have been isolated from fecal samples of mother and child combinations, as well as the corresponding human milk samples, indicative of a vertical transmission route from maternal GIT to (breastfed) infants [10–13]. This has given rise to the hypothesis that microbial colonization of the infant depends on the mother’s fecal/vaginal microbiota as well as on (providing) breast milk [14]. Bacteria may reach the mammary gland of the mother and may be transmitted directly to breastfed infants, although, inversely, bifidobacteria may be introduced into human milk from the infant’s oral cavity during suckling [14–17].
Notably, several studies have highlighted the possibility of vertical transmission of (components of) the gut microbiota from mother to child [12, 15, 18]. Furthermore, a recent pilot study involving four mother-child pairs revealed direct transmission of bifidobacterial strains from mothers to their newborns using a novel ITS-based approach [11]. Although intriguing, the biological relevance of such findings in this latter work is limited due to the small number of mother-infant pairs analysed [11].
Another important component of the gut microbiota is represented by virus-like particles in the GIT, which together constitute the gut virome [19, 20]. Currently, very little is known about the infant virome [21–23], although the existence and impact of phages specific for bifidobacteria, i.e., bifidophages, in the infant gut has recently been discribed [24].
In the current study, we analyzed the composition of the bifidobacterial communities of 25 mother-infant pairs by means of the ITS profiling approach mentioned above, combined with cultivation and genome investigations. Employing a next-generation sequencing (NGS) approach, we identified bifidobacterial strains that are shared between mothers and their corresponding newborns, while we furthermore obtained evidence for vertical, mother-child transmission of bifido(pro)phages.
Methods
Subject recruitment and sample collection
The study protocol was approved by the Ethical Committee of the “Azienda Provinciale per i Servizi Sanitari” in Trento, Italy, as well as by the Ethical Committee of the University of Parma, Italy, and informed written consent was obtained from all participants or their legal guardians. Twenty-five mother-infant pairs were enrolled in this study, which were selected based on the following criteria: the mother’s age (ranging in age from 18 to 40 years old) and those subjects who were not taking any probiotics or antibiotics. All subjects were considered to be healthy, as based on self-reporting. At the time of delivery, during birth or immediately after it, stool samples were collected of 21 mothers. Stool samples of infants were collected at two different time points, i.e., 7 days and 1 month following birth of the infant. Furthermore, 10 ml of breast milk sample of each mother was collected in sterile tubes (Table 1). For the sample set TVPR_03, we collected stool samples of infants and breast milk samples at 1 month following birth. Stool and milk samples were collected following the “Stool Packaging Instructions” of the “Core Microbiome Sampling Protocol A” reported in the “Manual of Procedures of the Human Microbiome Project” (http://www.hmpdacc.org/doc/HMP_MOP_Version12_0_072910.pdf) [25]. Samples were immediately frozen at −20 °C and shipped under frozen conditions to the laboratory, where they were immediately processed. DNA extractions of stool samples and human milk samples were performed as previously described [11].
Table 1.
List of samples included in this study
| CoupleID | Samples | |
|---|---|---|
| TVTR_10005 | Mum-T0 | 2062 |
| Milk-T3 | 2072 | |
| Infant-T3 | 2074 | |
| Milk-T4 | 2076 | |
| Infant-T4 | 2078 | |
| TVTR_10006 | Milk-T3 | 2090 |
| Infant-T3 | 2092 | |
| Milk-T4 | 2094 | |
| Infant-T4 | 2096 | |
| TVTR_10007 | Milk-T3 | 2108 |
| Infant-T3 | 2110 | |
| Milk-T4 | 2112 | |
| Infant-T4 | 2114 | |
| TVTR_10008 | Milk-T3 | 2126 |
| Infant-T3 | 2128 | |
| Milk-T4 | 2130 | |
| Infant-T4 | 2132 | |
| TVTR_10009 | Mum-T0 | 2134 |
| Milk-T3 | 2144 | |
| Infant-T3 | 2146 | |
| Milk-T4 | 2148 | |
| Infant-T4 | 2150 | |
| TVTR_10010 | Mum-T0 | 2152 |
| Milk-T3 | 2162 | |
| Infant-T3 | 2164 | |
| Milk-T4 | 2166 | |
| Infant-T4 | 2168 | |
| TVTR_10017 | Mum-T0 | 2278 |
| Milk-T3 | 2287 | |
| Infant-T3 | 2290 | |
| Milk-T4 | 2292 | |
| Infant-T4 | 2293 | |
| TVTR_10019 | Mum-T0 | 2314 |
| Milk-T3 | 2324 | |
| Infant-T3 | 2326 | |
| Milk-T4 | 2328 | |
| Infant-T4 | 2330 | |
| TVTR_10020 | Mum-T0 | 2332 |
| Milk-T3 | 2342 | |
| Infant-T3 | 2344 | |
| Milk-T4 | 2346 | |
| Infant-T4 | 2347 | |
| TVTR_10021 | Mum-T0 | 2350 |
| Milk-T3 | 2360 | |
| Infant-T3 | 2362 | |
| Milk-T4 | 2364 | |
| Infant-T4 | 2365 | |
| TVTR_10023 | Milk-T3 | 2396 |
| Infant-T3 | 2398 | |
| Milk-T4 | 2400 | |
| Infant-T4 | 2401 | |
| TVTR_10024 | Mum-T0 | 2404 |
| Milk-T3 | 2414 | |
| Infant-T3 | 2416 | |
| Milk-T4 | 2418 | |
| Infant-T4 | 2419 | |
| TVTR_10025 | Mum-T0 | 2422 |
| Milk-T3 | 2432 | |
| Infant-T3 | 2434 | |
| Milk-T4 | 2436 | |
| Infant-T4 | 2437 | |
| TVTR_10028 | Mum-T0 | 2476 |
| Milk-T3 | 2486 | |
| Infant-T3 | 2487 | |
| Milk-T4 | 2490 | |
| Infant-T4 | 2491 | |
| TVTR_10029 | Mum-T0 | 2492 |
| Milk-T3 | 2502 | |
| Infant-T3 | 2504 | |
| Milk-T4 | 2506 | |
| Infant-T4 | 2508 | |
| TVTR_10030 | Mum-T0 | 2509 |
| Milk-T3 | 2519 | |
| Infant-T3 | 2521 | |
| Milk-T4 | 2523 | |
| Infant-T4 | 2525 | |
| TVTR_10031 | Mum-T0 | 2526 |
| Milk-T3 | 2536 | |
| Infant-T3 | 2538 | |
| Milk-T4 | 2540 | |
| Infant-T4 | 2541 | |
| TVTR_10032 | Mum-T0 | 2544 |
| Milk-T3 | 2554 | |
| Infant-T3 | 2556 | |
| Milk-T4 | 2558 | |
| Infant-T4 | 2559 | |
| TVTR_10034 | Mum-T0 | 2580 |
| Milk-T3 | 2590 | |
| Infant-T3 | 2592 | |
| Milk-T4 | 2594 | |
| Infant-T4 | 2595 | |
| TVTR_10035 | Mum-T0 | 2598 |
| Milk-T3 | 2608 | |
| Infant-T3 | 2610 | |
| Milk-T4 | 2612 | |
| Infant-T4 | 2613 | |
| TVTR_10036 | Mum-T0 | 2616 |
| Milk-T3 | 2626 | |
| Infant-T3 | 2628 | |
| Milk-T4 | 2630 | |
| Infant-T4 | 2632 | |
| TVTR_10038 | Mum-T0 | 2652 |
| Milk-T3 | 2662 | |
| Infant-T3 | 2664 | |
| Milk-T4 | 2666 | |
| Infant-T4 | 2668 | |
| TVPR_01 | Mum-T0 | 3000 |
| Milk-T4 | 3001 | |
| Infant-T4 | 3002 | |
| TVPR_02 | Mum-T0 | 3006 |
| Milk-T3 | 3007 | |
| Infant-T3 | 3008 | |
| Milk-T4 | 3009 | |
| Infant-T4 | 3010 | |
| TVPR_03 | Mum-T0 | 3011 |
| Milk-T4 | 3012 | |
| Infant-T4 | 3013 | |
ITS gene amplification and MiSeq sequencing
Partial ITS sequences were amplified from extracted DNA using the primer pair Probio-bif_Uni/Probio-bif_Rev, which targets the spacer region between the 16S rRNA and the 23S rRNA genes within the ribosomal RNA (rRNA) locus [26]. Illumina adapter overhang nucleotide sequence was then added to the generated ITS amplicons of c. 200 bp. The library of ITS amplicons was prepared following the 16S Metagenomic Sequencing Library Preparation Protocol (Part No. 15044223 Rev. B—Illumina). Sequencing was performed using an Illumina MiSeq sequencer with MiSeq Reagent Kit v3 chemicals.
ITS microbial profiling analysis
Profiling of known bifidobacterial species was performed using the primer pair Probio_bif_uni/Probio_bif_rev, an improved bifidobacterial ITS database encompassing all publicly available bifidobacterial genomes and a custom bioinformatics script, as described previously [26].
Evaluation of the bifidobacterial cell density by qPCR
The presence of bifidobacteria in infant stool samples was evaluated using quantitative real-time PCR (qRT-PCR). The primers used in this study are Probio_bif_uni/Probio_bif_rev to determine numbers of bifidobacteria [26] and Probio_uni/Probio_rev to quantify total bacterial numbers [27]. The quantitative contribution of bifidobacteria to the overall infant gut microbiota of each sample was evaluated by a ratio of the genome copy number ITS gene/16S rRNA gene (the genes targeted were in the same copy per genome). qPCR was performed using SoFast EvaGreen Supermix (Bio-Rad) on a CFX96 system (BioRad, CA, USA) following previously described protocols [28].
Recovery of bifidobacteria on selective media
One gram of each fecal sample or 1 ml of human milk was mixed with 9 ml of phosphate-buffered saline (PBS), pH 6.5. Serial dilutions and subsequent plating were performed using the de Man-Rogosa-Sharpe (MRS) agar (Scharlau Chemie, Barcelona, Spain), supplemented with 0.05% (wt/col) l-cysteine hydrochloride and 50 μg/ml mupirocin (Delchimica, Italy). The agar plates were incubated in an anaerobic atmosphere (2.99% H2, 17.01% CO2, and 80% N2) in a chamber (Concept 400; Ruskin) at 37 °C for 48 h. Ten colonies were taken as a representation of the bacterial strains retrieved from the selective medium. DNA was extracted and subjected to (sub)-species identification as previously described [11].
De novo genome sequencing and bioinformatics analyses
The genome sequence of the new bifidobacterial isolates was determined by GenProbio srl (Parma, Italy) using a MiSeq platform (Illumina, UK). A genome library was generated following the TruSeq Nano DNA library Prep protocol (Part No. 15041110 Rev. D). Library samples were loaded into a Flow Cell V3 600 cycles (Illumina) according to the technical support guide, and generated reads were depleted of adapter sequences, quality-filtered, assembled and protein-encoding open reading frame (ORF) predicted through the MEGAnnotator pipeline [29].
Fermentation profiles on milk glycans
Prior to carrying out growth profile assays, Bifidobacterium cultures were grown from stock in Difco MRS (BD) and sub-cultured twice in the same medium. Subsequently, 5 ml of freshly prepared modified MRS (mMRS) medium [peptone from casein, 10 g/L; meat extract, 10 g/L, and yeast extract, 5 g/L (purchased from Difco); K2HPO4, 3 g/L; KH2PO4, 3 g/L; pyruvic acid, 0.2 g/L; polysorbate 80, 1 mL/L; tri-ammonium citrate, 2 g/L; MgSO4·7H2O, 0.575 g/L; MnSO4·4H2O, 0.12 g/L; cysteine-HCl, 0.3 g/L; and FeSO4·7H2O, 0.034 g/L], supplemented with 0.05% cysteine hydrochloride, and 1% of a particular carbohydrate was inoculated with 50 μl (1%) of a stationary phase culture. All carbohydrates (fucose, lactose, sialic acid, 3′-sialyllactose, 6′-sialyllactose, 2′-fucosyllactose, 3′-fucosyllactose, lacto-N-tetraose, or lacto-N-neotetraose) were dissolved in water and then sterilized by filtration (0.2-μ filter size) and then added to mMRS after sterilization by autoclaving. Uninoculated mMRS medium was used as a negative control. Cultures were incubated under anaerobic conditions in a modular atmosphere-controlled system (Davidson and Hardy, Belfast, Ireland) at 37 °C for 24 h, and the optical density at 600 nm (OD600) was determined manually, using a Shimadzu UV-1280 UV-399 VIS Spectrophotometer. Growth assays were carried out in triplicate, with standard deviation calculated for error values.
Shotgun metagenomic analyses
Fastq files obtained from metagenomic sequencing of infant and mother fecal samples, as well as milk samples, were used as input for the SPAdes assembler (version 3.9, using default parameters and enabling the metagenomic option—meta) for de novo metagenomic assemblies [30]. ORFs were predicted with Prodigal [31] and annotated using NCBI RefSeq databases through RAPSearch2 aligner (cut-off E value of 1 × 10−30) [32]. Each contig was taxonomically classified by means of the gene hit obtained through the NCBI RefSeq databases by the use of the Contig Classifier (CoCla) script [33, 34]. Species-specific contigs were subdivided in different files, obtaining reconstructed genomes of the most abundant (bifido)bacterial taxa within each analysed sample.
Bifidophage analyses
Contigs attributed to bifidobacterial (sub)species, by means of the CoCla script, were screened for the presence of bifidobacterial phages/prophages using a custom database composed of previously identified bifidophages [24]. Identified phage sequences were further manually curated in order to unveil overlapping extremities within the contig sequences. The presence of putatively identified bifidophages was screened in mother and infant fecal samples, and milk samples by PCR based on unique genes of that particular bifidophage.
Results and discussion
Evaluation of the bifidobacterial composition of the gut microbiota of infants
To characterize the bifidobacterial population in the infant gut, we employed a recently developed pipeline based on sequencing of a hypervariable ITS region, here referred to as ITS profiling [26]. We collected fecal samples of 25 infants at two different time points, i.e., 7 days and 1 month following birth (Table 1). ITS profiling of these 50 samples produced a total of 1,703,642 reads, ranging from 1224 to 113,414 reads per sample (Additional file 1: Table S1), which were grouped into clusters of identical sequences (OTUs or operational taxonomic units) and then taxonomically classified. OTU analysis revealed that the dominant bifidobacterial species detected in the investigated fecal samples were Bifidobacterium longum subsp. longum, Bifidobacterium breve, Bifidobacterium bifidum, Bifidobacterium longum subsp. infantis, Bifidobacterium adolescentis, and Bifidobacterium pseudocatenulatum. In addition, ITS-based OTUs display a similarity level below 93% to any of the ITS sequences of the current known bifidobacterial (sub)species (Fig. 1 and Additional file 2: Figure S1). As previously discussed by Milani et al. [26], the comparison between bifidobacterial-type strain ITS sequences reveal that different bifidobacterial (sub)species possess ITS identity above 98%; it is therefore a distinct possibility that identified ITS-based OTUs with a similarity level below 93% represent novel bifidobacterial (sub)species. Notably, and in contrast to what currently is known about the composition of the bifidobacterial communities of the infant gut, we identified that such populations are composed not only by typical infant-gut bifidobacterial species, such as B. breve, B. bifidum, and B. pseudocatenulatum, but also by adult-associated bifidobacterial taxa, like B. adolescentis and B. catenulatum [11, 15]. This finding suggests that an ecological division based on age is not valid or at least not absolute, thus underscoring a possible strain-sharing scenario between adult and infant microbiotas. Furthermore, in seven and six cases, we observed a high abundance of non-common infant gut bifidobacteria such as Bifidobacterium tissieri and Bifidobacterium dentium, respectively (Fig. 1 and Additional file 2: Figure S1). In order to assess the quantitative contribution of bifidobacteria to the overall infant gut microbiota of each sample, the total bacterial load as well as the bifidobacterial cell count were evaluated by qRT-PCR. These analyses indicate that bifidobacteria represent between 0.05 and 96.44% of the overall infant gut microbiota (average of 36.54%), ranging from 2.48E + 03 to 8.22E + 08 genome copy number per gram of feces. No significant differences between the two time points were detected. These findings strengthen the now well-established notion that bifidobacteria represent one of the dominant genera in the infant gut environment (Fig. 1) [10, 35–39].
Fig. 1.
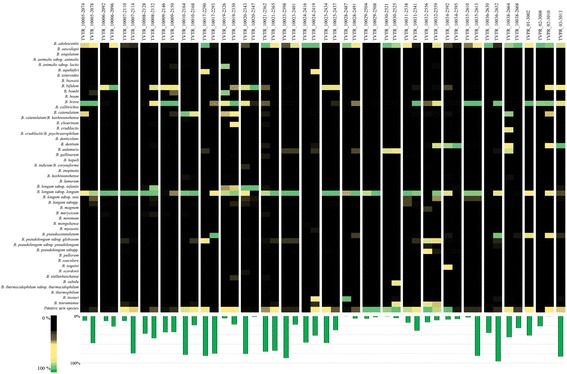
Profiling of the bifidobacterial communities of infant fecal samples. The heat map representation shows the relative abundance of Bifidobacterium species. On the bottom of the image, the bar plots show qPCR data regarding the proportion (in %) of bifidobacteria relative to other bacteria present in a given sample
Sharing of OTUs between mothers and children
Bacterial transmission from mother to child has been proposed to occur during birth and as a consequence of breastfeeding [40]. In order to explore if a similar scenario is applicable to the bifidobacterial population, we compared bifidobacterial ITS profiles of infants with those of their corresponding mother’s fecal (obtained during or soon after delivery) and milk samples obtained 7 and/or 30 days after delivery (Fig. 2 and Additional file 2: Figure S1). Interestingly, the number of ITS-based OTUs shared by a given sample set ranged from three in TVPR_01 to 273 in TVTR_10032 (Additional file 1: Table S2). These ITS-based OTUs encompass members of the species B. bifidum, B. adolescentis, B. dentium, B. breve, B. longum spp., B. pseudocatenulatum, and B. eulemuris and OTUs displaying a level of similarity below 93% to any of the current known bifidobacterial (sub)species (Fig. 2 and Additional file 2: Figure S1). Furthermore, we highlighted that the most prevalent bifidobacterial species are B. breve, B. longum spp., B. bifidum, B. adolescentis, B. dentium, and B. pseudocatenulatum, all being identified in infant feces. These taxa were also found in the corresponding fecal and milk samples of the mother (Fig. 2 and Additional file 2: Figure S1). Notably, comparison of the ITS-based OTUs identified in the data sets indicated the presence of identical ITS-based OTUs in different sample pairs, implying that these identical sequences correspond to very closely related strains that are present in non-corresponding mother-infant dyads. We therefore decided to verify these findings using a cultivation approach where we isolated such strains followed by de novo genome sequencing of such bifidobacterial isolates.
Fig. 2.

Profiling of the bifidobacterial communities of each CoupleID. The heat map represents the relative abundance of bifidobacterial species that were determined to be present in each sample. On the left side, all known and putative novel bifidobacterial species are reported. Sample origin and CoupleID codes are reported on the top side of the heat map
Isolation and genome sequencing of shared mother-child bifidobacterial strains
In order to validate the notion that the identical bifidobacterial ITS-based OTUs that seem to be shared between mother-newborn pairs are the result of vertical transmission, we used 25 infant and mother stool samples, as well as milk samples, to isolate bifidobacteria employing a selective cultivation medium (Table 1). Isolated colonies were identified by PCR amplification of the 16S rRNA gene followed by amplicon sequencing as previously described by Turroni et al. [41]. This cultivation effort resulted in the isolation of 14 bifidobacterial strains, whose taxonomical identity perfectly matched those identified by bifidobacterial ITS profiling described above for the same samples (Fig. 2 and Additional file 2: Figure S1). These 14 isolated strains, representing different bifidobacterial species (Table 2), were then subjected to whole-genome sequencing using an Illumina-MiSeq platform. As outlined in Table 2, the predicted general features of these genomes appear to be similar to those generally observed for bifidobacteria [42]. Moreover, to identify the presence of these strains in stool samples of the corresponding mother, we performed a comparative genome analysis to identify the occurrence of specific, unique genes for each strain. Such analyses allowed the design of strain-specific primer pairs to be used in a PCR-based detection method involving DNA isolated from (mother’s) stool samples as a template (Additional file 1: Table S3). Notably, these experiments resulted in the detection of each of the infant stool-isolated and genome-sequenced strains in the fecal samples of the corresponding mother. In addition, we used isolated DNA from a mother stool sample of a non-corresponding CoupleID to detect false positive results (Fig. 3). Altogether, these data clearly corroborate the notion that these bifidobacterial isolates were obtained by infants via a maternal vertical transmission route (Fig. 3).
Table 2.
General genome features of vertical transmitted bifidobacterial strains
| Couple ID | Biological origin | Contig | Genome length | % GC | Predicted ORFs | tRNA | rRNA | |
|---|---|---|---|---|---|---|---|---|
| Bifidobacterium longum subsp. longum 1886B | TVTR_10006 | Human milk | 47 | 2473746 bp | 61.14% | 2137 | 62 | 3 |
| Bifidobacterium bifidum 1887B | TVTR_10006 | Human milk | 21 | 2255543 bp | 59.49% | 1851 | 53 | 3 |
| Bifidobacterium longum subsp. infantis 1888B | TVTR_10020 | Infant stool | 26 | 2579732 bp | 59.40% | 2212 | 55 | 3 |
| Bifidobacterium breve 1889B | TVTR_10005 | Infant stool | 22 | 2344818 bp | 59.66% | 2037 | 53 | 2 |
| Bifidobacterium longum subsp. longum 1890B | TVTR_10023 | Infant stool | 109 | 2341670 bp | 59.55% | 1955 | 60 | 2 |
| Bifidobacterium breve 1891B | TVTR_10008 | Infant stool | 38 | 2089648 bp | 59.70% | 2115 | 54 | 2 |
| Bifidobacterium adolescentis 1892B | TVTR_10024 | Infant stool | 11 | 2150850 bp | 58.65% | 1720 | 55 | 4 |
| Bifidobacterium dentium 1893B | TVTR_10034 | Infant stool | 24 | 2271880 bp | 58.74% | 2070 | 56 | 4 |
| Bifidobacterium breve 1895B | TVPR_01 | Infant stool | 17 | 20270860 bp | 58.58% | 1871 | 53 | 2 |
| Bifidobacterium pseudocatenulatum1896B | TVPR_01 | Human milk | 15 | 2197471 bp | 56.15% | 1733 | 54 | 5 |
| Bifidobacterium longum subsp. longum 1897B | TVTR_10030 | Human milk | 55 | 2453605 bp | 59.50% | 2143 | 55 | 2 |
| Bifidobacterium longum subsp. longum 1898B | TVTR_10008 | Infant stool | 41 | 2474386 bp | 59.47% | 2049 | 57 | 3 |
| Bifidobacterium catenulatum 1899B | TVTR_10029 | Infant stool | 14 | 2124599 bp | 58.90% | 1749 | 55 | 5 |
| Bifidobacterium breve 1900B | TVTR_10031 | Infant stool | 22 | 2287865 bp | 59.50% | 1910 | 53 | 2 |
Fig. 3.
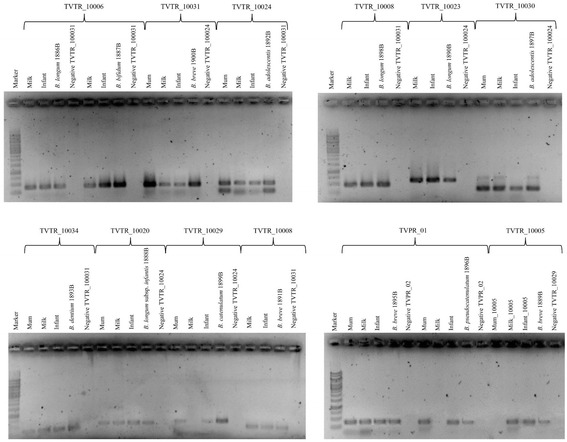
Evaluation of persistence of putative vertically transmitted strains in fecal samples of the infants and mothers as well as in milk samples by PCR assays. Marker lane is the Thermo Scientific GeneRuler 1 kb DNA Ladder. The reference strains used as positive controls for each CoupleID respectively are B. longum 1886B, B. bifidum 1887B, B. breve 1900B, B. adolescentis 1892B, B. longum 1898B, B. longum 1897B, B. dentium 1893B, B. longum subsp. infantis 1888B, B. catenulatum 1899B, B. breve 1891B, B. breve 1895B, B. pseudocatenulatum 1896B, and B. breve 1889B. Primer sequences are reported in Additional file 1: Table S3
Genetic adaptation of early bifidobacterial colonizers to the infant gut
Transmission of (bifido)bacteria from mother to her child may be possible through ingestion of mother’s milk [38, 39, 43, 44]. This human fluid represents a bacterial transmission medium that is ideal not only for microbiota dispersal but also for the provision of nutrients to the initial colonizers of the infant gut [10, 38, 39, 44, 45]. Human milk is a very rich source of glycans, including lactose and human milk oligosaccharides (HMOs) [46], which represent an important carbon and energy source for pioneering saccharolytic members of the infant gut microbiota, in particular bifidobacteria [47–49]. However, not all bifidobacteria are genetically adapted to utilize such milk-derived glycans [47, 50].
In order to identify the genomic repertoire involved in the utilization of HMO, lactose, and/or HMO-derived carbohydrates, such as fucose, lactose, sialic acid, fucosyllactose (2-FL and 3-FL), sialyllactose (3-SL and 6-SL), lacto-N-tetraose (LNT), and lacto-N-neotetraose (LNnT), we performed a comparative genomic analysis of all genomes decoded in this study. Our analysis revealed that six strains belonging to B. longum, B. breve, or B. bifidum possess a variety of genes allowing them to metabolize various HMO and HMO-derived glycans [47, 49–51] (Fig. 4a). In contrast, these genes are absent in the genomes of B. adolescentis 1892B and B. dentium 1893B, except for the genes coding for β-galactosidase (Fig. 4a). Moreover, this analysis revealed that putative fucosidase- and sialidase-encoding genes are absent in the examined B. longum genomes, except for that of B. longum 1886B, which encompasses an ORF (B1886_0565) specifying a predicted sialidase. Furthermore, no predicted sialidase-encoding genes were identified in the genome of B. pseudocatenulatum 1896B, while no homologs of the gene encoding lacto-N-biose phosphorylase were detected in the chromosomes of B. pseudocatenulatum 1896B, B. longum 1897B, or B. catenulatum 1899B.
Fig. 4.

Human milk oligosaccharide degradation capabilities of vertically transmitted bifidobacterial strains. a Heat map representation of the presence/absence of genes encoding enzymes for degradation of milk oligosaccharides and transporters for uptake of the released compounds. Red and black squares represent the presence and absence of genes, respectively. b Growth profile on HMO-derived carbohydrates. Black color indicates that the final OD600 nm is <0.3, yellow indicates the range of final OD600 nm from 0.3 to 0.5, light green indicates the range between >0.5 and 0.8, and green indicates the final OD600 nm >0.8
In order to validate these in silico data, we performed growth experiments involving the bifidobacterial strains that we isolated in this study using an mMRS medium containing various HMOs and HMO-derived glycans, i.e., LNT, LNnT, 3-SL, 6-SL, 2-FL, 3-FL, fucose, sialic acid, and lactose, as the sole carbon source (Fig. 4b). Based on these culturing experiments, we observed that the B. longum strains, B. bifidum 1887B, and B. breve strains were able to grow (OD values greater than 0.3) on LNT, LNnT, and lactose, while B. adolescentis 1892B and B. dentium 1893B were unable to grow on these carbohydrates, except for lactose. Moreover, only B. breve 1889B and B. breve 1895B were able to utilize sialic acid and fucose, while B. bifidum 1887B and B. longum subsp. infantis 1888B were able to utilize 3-SL, 6-SL, and 3-FL. Finally, only B. bifidum 1887B was able to degrade 2-FL. Notably, these data are consistent with current knowledge on bifidobacterial HMO/HMO-derived glycan utilization abilities [47, 49, 51–55].
Interestingly, the results of these fermentation profiling experiments, correspond very well with those predicted from the in silico analyses of the genomes decoded in this study.
Genome reconstruction of shared bifidobacteria based on in silico analyses of microbiomes
In order to verify the occurrence of identical bifidobacterial strains in the microbiota of an infant’s fecal sample and in that of the corresponding milk and/or fecal sample of the mother, a shotgun metagenomic analysis was performed of three infant samples (TVPR-01, TVPR-02, and TVPR-03) for which we observed a high number of potentially vertically transmitted bifidobacterial strains. Taxonomic assignments (based on publicly available bifidobacterial genomic data) of the obtained microbiome data (i.e., shotgun metagenomic reads) corresponding to the TVPR-01, TVPR-02, and TVPR-03 infant fecal samples showed that 2.09, 9.02, and 12.16% of these shotgun metagenomic reads were classified as bifidobacterial DNA, respectively (Additional file 1: Table S4). Furthermore, taxonomic classification of these data sets determined that in sample TVPR-01, 1.56% of reads was taxonomically assigned to B. breve, while for TVPR-02, 6.16% of reads were annotated as B. dentium sequences, and finally, in the case of TVPR-03, 8.21% of reads were shown to correspond to B. dentium (Additional file 1: Table S4). Moreover, we reconstructed the complete genome sequences of each of the abovementioned bifidobacterial strains, by an in silico analysis of these three shotgun metagenomics datasets using a previously described, in-house developed pipeline [11, 34].
Genomic comparison of the isolated B. breve 1895B and the reconstructed B. breve TVPR-01 clearly showed the isogenic nature of both genomes, exhibiting an ANI value of 99.99%. Thus, using three different approaches, i.e., ITS profiling, strain isolation from fecal samples, and genome reconstruction from shotgun metagenomic data, we were able to confirm the essentially identical genetic composition of these bifidobacterial strains. Furthermore, we verified, through PCR-amplification of unique gene (Additional file 1: Table S3), that B. breve 1895B/TVPR-01 was shared between mother and child (Fig. 3). Unfortunately, despite several attempts, we were not able to isolate the B. dentium TVPR-02 and B. dentium TVPR-03 strains, most likely because of exigent nutritional requirements of these strains. Nonetheless, PCR amplification attempts targeting specific genes of TVPR-02 and TVPR-03, which were identified by means of comparative genomic analyses of the TVPR-02 and TVPR-03 microbiome-reconstructed genomes against the nine publicly available B. dentium strains, and confirmed the presence of these strains in corresponding mothers and infants (Additional file 3: Figure S2 and Additional file 1: Table S3). In order to avoid false positive results, we used isolated DNA from a mother stool sample of a non-corresponding dataset (Additional file 3: Figure S2). Consequently, these data clearly support the assumption of a vertical maternal inheritance of the most abundant bifidobacterial strains identified in the gut microbiota of children, as previously described in other studies [11, 15].
Bifidoprophages transmission from mother to child
Bacteriophages may also be transmitted from mother to child during the early phases of life, provided that the corresponding hosts for such phages are also passed across. Thus, we searched shotgun metagenomics data, obtained from stool samples of various mother and corresponding child and milk sample sets, for sequences that matched with known bacteriophages infecting bifidobacteria, i.e., bifidophages [24], focusing on datasets in which we had already identified bifidobacterial transmission. In order to identify the presence of both bifidophages as well as bifidoprophages, we scanned all obtained metagenomic datasets using a previously developed bioinformatics pipeline [11, 34]. Putative bifidobacterial prophage sequences were identified by manual examination of all genes belonging to the contigs attributed to bifidobacterial (sub)species [24, 56, 57]. The bifidobacterial phage/prophage screening revealed the presence of 21 putative bifido(pro)phages within nine samples that exhibit integrated or circular phage genome sequences (Table 3). A manually curated screening of the reconstructed phages unveiled the circular status of their genomes for eight bifidophages, highlighting overlapping sequence ends within the assembled contigs (Additional file 4: Figure S3). Interestingly, taxonomical gene classification based on NCBI RefSeq database matches reveal that these phages were predicted to belong to several bifidobacterial (sub)species, i.e., B. adolescentis, B. breve, B. dentium, B. longum, and B. pseudocatenulatum (Additional file 4: Figure S3). Since it was not possible to retrieve sequences of these identified bifidophages from each of the three components of a given metagenomic dataset (i.e., a corresponding mother and infant fecal samples and associated milk sample), we decided to utilize a PCR-based approach involving a PCR primer pair that was designed on unique sequences of a given phage (Additional file 1: Table S5). Furthermore, a second PCR primer pair was designed on the contig edges, in order to determine if the phage DNA was circular/concatameric or integrated into the host chromosome (Additional file 1: Table S5). As shown in Fig. 5, B. longum phage 10029 was identified in the fecal samples T3 and T4 of the same child as well as in the milk samples of the corresponding mother. While the use of the primer pair (P19-P20) results in the amplification of phage DNA in infant and milk samples, the second primer pair (P17-P18) unveiled the presence of a circular phage, yet only in sample T3 of the child. These data clearly indicates that this bifido(pro)phage was vertically transmitted by the mother through breastfeeding as part of the bifidobacterial host and was then apparently induced in the gut of the newborn. Furthermore, for samples Mum_2598, Infant_T3_2610, and Infant_T4_2613, the PCR amplification of both primer sets of the B. longum phage 10035 produced an amplicon (Fig. 5). This finding suggests the presence of the same bifidophage in both mother and infant microbiome, expanding the previous view of bifidobacterial phages multiplying in the infant gut thereby perhaps limiting overgrowth by their bifidobacterial host [24].
Table 3.
Identified bifidophage/bifidoprophages within metagenomes
| Samples | Species assignment | Genome status |
|---|---|---|
| TVPR3B | B. adolescentis | Integrated |
| B. adolescentis | Integrated | |
| B. adolescentis | Circular | |
| C10009IS2149FEt5 | B. bifidum | Integrated |
| B. bifidum | Unknown | |
| B. breve | Integrated | |
| B. breve | Integrated | |
| C10009MS2134FEt0 | B. adolescentis | Circular |
| C10019IS2329FEt5 | B. breve | Circular |
| C10029IS2503FEt3 | B. longum subsp. longum | Unknown |
| B. longum subsp. longum | Unknown | |
| B. longum subsp. longum | Unknown | |
| B. longum subsp. longum | Unknown | |
| B. longum subsp. longum | Circular | |
| n.d. | Circular | |
| C10029MS2492FEt0 | B. pseudolongum | Circular |
| C10035IS2609FEt3 | B. longum subsp. longum | Integrated |
| B. longum subsp. longum a | Circular | |
| C10035IS2913FEt4 | B. breve | Circular |
| B. longum subsp. longum a | Circular | |
| C10035MS2598FEt0 | B. longum subsp. longum | Unknown |
n.d. Bifidobacterial species not detected, Unknown genome-status unknown
a B. longum phages that share the same genomic sequences between samples
Fig. 5.
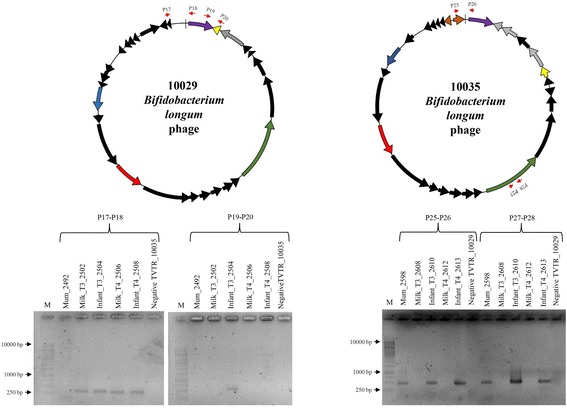
Vertically transmitted bifidophages from mothers to newborns. Indicated at the top of the figure are the genomic maps of the identified B. longum phage 10029 and 10035. The modular genomic structure is indicated by different patterns, which specifies their predicted function (violet, lysogeny module; blue, DNA replication; red, DNA packaging and head; green, tail and tail fiber; yellow, lysis module; black arrows, hypothetical protein; grey arrows, similar to bacterial protein). On the bottom are reported the presence of the phage within the mother’s and infant’s metagenomic samples through PCR analysis. Primer sequences are reported in Additional file 1: Table S5
Conclusions
Passage through the birth canal during delivery and subsequent breastfeeding are considered important events that affect microbial colonization of the gastrointestinal tract of the newborn [15, 45, 58, 59]. In this study, an NGS approach was employed to confirm the existence of a vertical transmission route of bifidobacterial communities in 25 mother and newborn pairs. Furthermore, our analyses highlight how bifidobacteria, being one of the dominant members of the infant gut microbiota, are inherited from the mother through a vertical transmission route, and imply human milk as a potential vehicle to facilitate this acquisition. Milk-mediated transmission of certain bifidobacterial strains may be supported by their ability to utilize HMOs and/or HMO-derived glycans. It is plausible that the first microbial colonizers of the human gut include commensals that are able to metabolize such milk glycans, such as the bifidobacterial strains belonging to B. bifidum species. However, these initial bifidobacterial colonizers may be important to promote the establishment of bifidobacterial colonizers that are not by themselves able to degrade and thus utilize milk carbohydrates. These bifidobacterial colonizers can access HMO degradation products, such as sialic acid and fucose, employing a cross-feeding behavior, which has been recently described [54, 55, 60, 61].
Another crucial microbial player involved in the establishment and maintenance of a so-called climax gut microbiota encompasses bacteriophages [19, 62]. Very recently, bifidophages have been discovered and their phage particles have been morphologically characterized [24]. However, their inheritance in the infant gut from the gut virome of the respective mother was only hypothesized but never experimentally demonstrated. Our data clearly highlight the existence of a vertical transfer route for bifidophages, thereby facilitating their spread from mother to newborn.
Additional files
Table S1. Taxonomy and filtering report of samples included in this study. Table S2 ITS-based OTUs shared by an entire sample sets. Table S3 List of bifidobacterial strain-specific primers. Table S4 Metagenomic reads classification of samples TVPR-01, TVPR-02, and TVPR-03. Table S5 List of specific primers for each bifidophage/bifidoprophages identified. (DOCX 37 kb)
{kind=link}
Figure S1. ITS-based OTUs shared between mother-milk-infant sample sets. The bar plot represents the percentage of the total bifidobacterial population found in mother-milk, mother-infant, milk-infant and mother-milk-infant samples. (JPG 1306 kb)
{kind=link}
Figure S2. Evaluation of presence of TVPR-02 and TVPR-03 microbiome-reconstructed genomes in fecal samples of the infants and mothers as well as in milk samples. Primer sequences are reported in Additional file 1: Table S2. (JPG 235 kb)
{kind=link}
Figure S3. Identified bifidophage/bifidoprophage within the mother’s and infant’s metagenomic samples. Genomic maps of phages recall their modular genomic structure indicated by different patterns, which specifies their predicted function (violet: lysogeny module; blue: DNA replication; red: DNA packaging and head; green: tail and tail fiber; yellow: lysis module; black arrows: hypothetical protein; grey arrows: similar to bacterial protein). For each phage is reported the presence within the mother’s and infant’s metagenomic samples through PCR analysis. Primer sequences are reported in Additional file 1: Table S5. (JPG 1173 kb)
Acknowledgements
We thank the operative units of Obstetrics and Gynecology and of Neonatology of the S. Chiara Hospital (Trento, Italy). Part of this research is conducted using the High Performance Computing (HPC) facility of the University of Parma.
Funding
This work was financially supported by Fondazione Caritro, Trento, Italy; GenProbio srl for the financial support of the Laboratory of Probiogenomics; and EU Joint Programming Initiative—A Healthy Diet for a Healthy Life (JPI HDHL, http://www.healthydietforhealthylife.eu/) to MV and DvS (Grant No. 15/JP/HDHL/3280) and MIUR to MV. SD is supported by Fondazione Caritro, Trento, Italy. LM is supported by Fondazione Cariparma, Parma, Italy. DvS is a member of The APC Microbiome Institute funded by Science Foundation Ireland (SFI), through the Irish Government’s National Development Plan (Grant No. SFI/12/RC/2273).
Availability of data and materials
Bifidobacterial ITS profiling data for all the sample sets are accessible through the accession number SRP102222. The bifidobacterial sequences reported in this article have been deposited in the GenBank database under accession numbers NAPZ00000000, NAQA00000000, NAQB00000000, NAQC00000000, NAQD00000000, NAQE00000000, NAQF00000000, NAQG00000000, NAQH00000000, NAQI00000000, NAQJ00000000, NAQK00000000, and NAQL00000000. The versions described in this paper are NAPZ01000000, NAQA01000000, NAQB01000000, NAQC01000000, NAQD01000000, NAQE01000000, NAQF01000000, NAQG01000000, NAQH01000000, NAQI01000000, NAQJ01000000, NAQK01000000, and NAQL01000000. Shotgun metagenomics data have been deposited in GenBank under the following accession numbers SRP102273.
Authors’ contributions
SD and GAL designed and performed the experiments and wrote the manuscript. LM, GAL, CM, EP, and MZ performed the bioinformatics analyses. FT, KJ, CF, MM, RS, and AV performed the experiments. FA, PF, VG, IL, RC, RC, MC, TC, GC, SF, SG, CI, CM, DP, DS, MS, ST, and AP provided the samples. MV, FT, and DVS conceived the study and revised and approved the manuscript. All authors reviewed the manuscript. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
Stool and milk samples were collected in accordance with the protocols approved by the Ethical Committee of the “Azienda Provinciale per i Servizi Sanitari” in Trento, Italy, and Ethical Committee of the University of Parma, Italy.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- 2-FL
2-Fucosyllactose
- 3-FL
3-Fucosyllactose
- 3-SL
3-Sialyllactose
- 6-SL
6-Sialyllactose
- GIT
Gastrointestinal tract
- HMO
Human milk oligosaccharides
- ITS
Internal transcribed spacer
- LNnT
Lacto-N-neotetraose
- LNT
Lacto-N-tetraose
- MRS
de Man-Rogosa-Sharpe medium
- OTUs
Operational taxonomic units
- PBS
Phosphate-buffered saline
- qRT-PCR
Quantitative real-time PCR
Footnotes
Electronic supplementary material
The online version of this article (doi:10.1186/s40168-017-0282-6) contains supplementary material, which is available to authorized users.
References
- 1.Sim K, Powell E, Shaw AG, McClure Z, Bangham M, Kroll JS. The neonatal gastrointestinal microbiota: the foundation of future health? Arch Dis Child Fetal Neonatal Ed. 2013;98:F362–4. doi: 10.1136/archdischild-2012-302872. [DOI] [PubMed] [Google Scholar]
- 2.Azad MB, Konya T, Maughan H, Guttman DS, Field CJ, Chari RS, Sears MR, Becker AB, Scott JA, Kozyrskyj AL, Investigators CS. Gut microbiota of healthy Canadian infants: profiles by mode of delivery and infant diet at 4 months. Can Med Assoc J. 2013;185:385–94. doi: 10.1503/cmaj.121189. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.La Rosa PS, Warner BB, Zhou Y, Weinstock GM, Sodergren E, Hall-Moore CM, Stevens HJ, Bennett WE, Jr, Shaikh N, Linneman LA, et al. Patterned progression of bacterial populations in the premature infant gut. Proc Natl Acad Sci U S A. 2014;111:12522–7. doi: 10.1073/pnas.1409497111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Dominguez-Bello MG, Costello EK, Contreras M, Magris M, Hidalgo G, Fierer N, Knight R. Delivery mode shapes the acquisition and structure of the initial microbiota across multiple body habitats in newborns. Proc Natl Acad Sci U S A. 2010;107:11971–5. doi: 10.1073/pnas.1002601107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Backhed F, Ley RE, Sonnenburg JL, Peterson DA, Gordon JI. Host-bacterial mutualism in the human intestine. Science. 2005;307:1915–20. doi: 10.1126/science.1104816. [DOI] [PubMed] [Google Scholar]
- 6.Turroni F, Taverniti V, Ruas-Madiedo P, Duranti S, Guglielmetti S, Lugli GA, Gioiosa L, Palanza P, Margolles A, van Sinderen D, Ventura M. Bifidobacterium bifidum PRL2010 modulates the host innate immune response. Appl Environ Microbiol. 2014;80(2):730–40. doi:10.1128/AEM.03313-13. Epub 2013 Nov 15. [DOI] [PMC free article] [PubMed]
- 7.Zhang CH, Derrien M, Levenez F, Brazeilles R, Ballal SA, Kim J, Degivry MC, Quere G, Garault P, Vlieg JETV, et al. Ecological robustness of the gut microbiota in response to ingestion of transient food-borne microbes. ISME J. 2016;10:2235–45. doi: 10.1038/ismej.2016.13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Duranti S, Gaiani F, Mancabelli L, Milani C, Grandi A, Bolchi A, Santoni A, Lugli GA, Ferrario C, Mangifesta M, et al. Elucidating the gut microbiome of ulcerative colitis: bifidobacteria as novel microbial biomarkers. FEMS Microbiol Ecol. 2016;92(fiw191):1–12. [DOI] [PubMed]
- 9.Round JL, Mazmanian SK. The gut microbiota shapes intestinal immune responses during health and disease. Nat Rev Immunol. 2009;9:313–23. doi: 10.1038/nri2515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Martin R, Jimenez E, Heilig H, Fernandez L, Marin ML, Zoetendal EG, Rodriguez JM. Isolation of bifidobacteria from breast milk and assessment of the bifidobacterial population by PCR-denaturing gradient gel electrophoresis and quantitative real-time PCR. Appl Environ Microbiol. 2009;75:965–9. doi: 10.1128/AEM.02063-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Milani C, Mancabelli L, Lugli GA, Duranti S, Turroni F, Ferrario C, Mangifesta M, Viappiani A, Ferretti P, Gorfer V, et al. Exploring vertical transmission of bifidobacteria from mother to child. Appl Environ Microbiol. 2015;81:7078–87. doi: 10.1128/AEM.02037-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Makino H, Kushiro A, Ishikawa E, Muylaert D, Kubota H, Sakai T, Oishi K, Martin R, Ben Amor K, Oozeer R, et al. Transmission of intestinal Bifidobacterium longum subsp. longum strains from mother to infant, determined by multilocus sequencing typing and amplified fragment length polymorphism. Appl Environ Microbiol. 2011;77:6788–93. doi: 10.1128/AEM.05346-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Jost T, Lacroix C, Braegger CP, Rochat F, Chassard C. Vertical mother-neonate transfer of maternal gut bacteria via breastfeeding. Environ Microbiol. 2014;16:2891–904. doi: 10.1111/1462-2920.12238. [DOI] [PubMed] [Google Scholar]
- 14.O’Sullivan A, Farver M, Smilowitz JT. The influence of early infant-feeding practices on the intestinal microbiome and body composition in infants. Nutr Metab Insights. 2015;8(Suppl 1):1–9. [DOI] [PMC free article] [PubMed]
- 15.Makino H, Kushiro A, Ishikawa E, Kubota H, Gawad A, Sakai T, Oishi K, Martin R, Ben-Amor K, Knol J, Tanaka R. Mother-to-infant transmission of intestinal bifidobacterial strains has an impact on the early development of vaginally delivered infant’s microbiota. PLoS One. 2013;8:e78331. doi: 10.1371/journal.pone.0078331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Scarpellini E, Ianiro G, Attili F, Bassanelli C, De Santis A, Gasbarrini A. The human gut microbiota and virome: Potential therapeutic implications. Dig Liver Dis. 2015;47(12):1007–12. doi:10.1016/j.dld.2015.07.008. [DOI] [PMC free article] [PubMed]
- 17.Rodriguez JM. The origin of human milk bacteria: is there a bacterial entero-mammary pathway during late pregnancy and lactation? Adv Nutr. 2014;5:779–84. doi: 10.3945/an.114.007229. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Vaishampayan PA, Kuehl JV, Froula JL, Morgan JL, Ochman H, Francino MP. Comparative metagenomics and population dynamics of the gut microbiota in mother and infant. Genome Biol Evol. 2010;2:53–66. doi: 10.1093/gbe/evp057. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ogilvie LA, Jones BV. The human gut virome: a multifaceted majority. Front Microbiol. 2015;6(918):1-12. [DOI] [PMC free article] [PubMed]
- 20.Minot S, Sinha R, Chen J, Li HZ, Keilbaugh SA, Wu GD, Lewis JD, Bushman FD. The human gut virome: inter-individual variation and dynamic response to diet. Genome Res. 2011;21:1616–25. doi: 10.1101/gr.122705.111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Lim ES, Zhou YJ, Zhao GY, Bauer IK, Droit L, Ndao IM, Warner BB, Tarr PI, Wang D, Holtz LR. Early life dynamics of the human gut virome and bacterial microbiome in infants. Nat Med. 2015;21:1228-+. doi: 10.1038/nm.3950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Lim ES, Wang D, Holtz LR. The bacterial microbiome and virome milestones of infant development. Trends Microbiol. 2016;24:801–10. doi: 10.1016/j.tim.2016.06.001. [DOI] [PubMed] [Google Scholar]
- 23.Kramna L, Kolarova K, Oikarinen S, Pursiheimo JP, Ilonen J, Simell O, Knip M, Veijola R, Hyoty H, Cinek O. Gut virome sequencing in children with early islet autoimmunity. Diabetes Care. 2015;38:930–3. doi: 10.2337/dc14-2490. [DOI] [PubMed] [Google Scholar]
- 24.Lugli GA, Milani C, Turroni F, Tremblay D, Ferrario C, Mancabelli L, Duranti S, Ward DV, Ossiprandi MC, Moineau S, et al. Prophages of the genus Bifidobacterium as modulating agents of the infant gut microbiota. Environ Microbiol. 2016;18:2196–213. doi: 10.1111/1462-2920.13154. [DOI] [PubMed] [Google Scholar]
- 25.Human Microbiome Project C Structure, function and diversity of the healthy human microbiome. Nature. 2012;486:207–14. doi: 10.1038/nature11234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Milani C, Lugli GA, Turroni F, Mancabelli L, Duranti S, Viappiani A, Mangifesta M, Segata N, van Sinderen D, Ventura M. Evaluation of bifidobacterial community composition in the human gut by means of a targeted amplicon sequencing (ITS) protocol. FEMS Microbiol Ecol. 2014;90:493–503. doi: 10.1111/1574-6941.12410. [DOI] [PubMed] [Google Scholar]
- 27.Milani C, Hevia A, Foroni E, Duranti S, Turroni F, Lugli GA, Sanchez B, Martin R, Gueimonde M, van Sinderen D, et al. Assessing the fecal microbiota: an optimized ion torrent 16S rRNA gene-based analysis protocol. PLoS One. 2013;8:e68739. doi: 10.1371/journal.pone.0068739. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Milani C, Lugli GA, Duranti S, Turroni F, Mancabelli L, Ferrario C, Mangifesta M, Hevia A, Viappiani A, Scholz M, Arioli S, Sanchez B, Lane J, Ward DV, Hickey R, Mora D, Segata N, Margolles A, van Sinderen D, Ventura M. Bifidobacteria exhibit social behavior through carbohydrate resource sharing in the gut. Sci Rep. 2015;5:15782. doi: 10.1038/srep15782. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Lugli GA, Milani C, Mancabelli L, van Sinderen D, Ventura M. MEGAnnotator: a user-friendly pipeline for microbial genomes assembly and annotation. FEMS Microbiol Lett. 2016;363(fnw049):1–8. [DOI] [PubMed]
- 30.Bankevich A, Nurk S, Antipov D, Gurevich AA, Dvorkin M, Kulikov AS, Lesin VM, Nikolenko SI, Pham S, Prjibelski AD, et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J Comput Biol. 2012;19:455–77. doi: 10.1089/cmb.2012.0021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Hyatt D, Chen GL, LoCascio PF, Land ML, Larimer FW, Hauser LJ. Prodigal: prokaryotic gene recognition and translation initiation site identification. Bmc Bioinformatics. 2010;11(119):1–11. [DOI] [PMC free article] [PubMed]
- 32.Zhao Y, Tang H, Ye Y. RAPSearch2: a fast and memory-efficient protein similarity search tool for next-generation sequencing data. Bioinformatics. 2012;28:125–6. doi: 10.1093/bioinformatics/btr595. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Lugli GA, Milani C, Mancabelli L, Turroni F, Ferrario C, Duranti S, van Sinderen D, Ventura M. Ancient bacteria of the Ötzi’s microbiome: a genomic tale from the Copper Age. Microbiome. 2017;5:5. doi: 10.1186/s40168-016-0221-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Lugli GA, Milani C, Mancabelli L, Turroni F, Ferrario C, Duranti S, van Sinderen D, Ventura M. Erratum to: ancient bacteria of the Otzi’s microbiome: a genomic tale from the Copper Age. Microbiome. 2017;5:23. doi: 10.1186/s40168-017-0243-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Turroni F, Peano C, Pass DA, Foroni E, Severgnini M, Claesson MJ, Kerr C, Hourihane J, Murray D, Fuligni F, et al. Diversity of Bifidobacteria within the infant gut microbiota. PLoS One. 2012;7(5/e36957):1–12. [DOI] [PMC free article] [PubMed]
- 36.Arboleya S, Watkins C, Stanton C, Ross RP. Gut Bifidobacteria populations in human health and aging. Front Microbiol. 2016;7(1204):1–9. [DOI] [PMC free article] [PubMed]
- 37.Heavey MP, Rowland RI. The gut microflora of the developing infant: microbiology and metabolism. Microb Ecol Health Dis. 1999;11:75–83. doi: 10.1080/089106099435808. [DOI] [Google Scholar]
- 38.Makino H, Martin R, Ishikawa E, Gawad A, Kubota H, Sakai T, Oishi K, Tanaka R, Ben-Amor K, Knol J, Kushiro A. Multilocus sequence typing of bifidobacterial strains from infant’s faeces and human milk: are bifidobacteria being sustainably shared during breastfeeding? Benef Microbes. 2015;6:563–72. doi: 10.3920/BM2014.0082. [DOI] [PubMed] [Google Scholar]
- 39.Jost T, Lacroix C, Braegger C, Chassard C. Assessment of bacterial diversity in breast milk using culture-dependent and culture-independent approaches. Br J Nutr. 2013;110:1253–62. doi: 10.1017/S0007114513000597. [DOI] [PubMed] [Google Scholar]
- 40.Jiménez E, María Marín L, Martín R, Odriozola JM, Olivares M, Xaus J, Fernández L, Rodríguez JM. Is meconium from healthy newborns actually sterile? Res Microbiol. 2008;159(3):187–93. [DOI] [PubMed]
- 41.Turroni F, Foroni E, Pizzetti P, Giubellini V, Ribbera A, Merusi P, Cagnasso P, Bizzarri B, de’Angelis GL, Shanahan F, et al. Exploring the diversity of the bifidobacterial population in the human intestinal tract. Appl Environ Microbiol. 2009;75:1534–45. doi: 10.1128/AEM.02216-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Milani C, Lugli GA, Duranti S, Turroni F, Bottacini F, Mangifesta M, Sanchez B, Viappiani A, Mancabelli L, Taminiau B, et al. Genomic encyclopedia of type strains of the genus Bifidobacterium. Appl Environ Microbiol. 2014;80:6290–302. doi: 10.1128/AEM.02308-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Rodriguez JM, Murphy K, Stanton C, Ross RP, Kober OI, Juge N, Avershina E, Rudi K, Narbad A, Jenmalm MC, et al. The composition of the gut microbiota throughout life, with an emphasis on early life. Microb Ecol Health Dis. 2015;26:26050. doi: 10.3402/mehd.v26.26050. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Fernandez L, Langa S, Martin V, Maldonado A, Jimenez E, Martin R, Rodriguez JM. The human milk microbiota: origin and potential roles in health and disease. Pharmacol Res. 2013;69:1–10. doi: 10.1016/j.phrs.2012.09.001. [DOI] [PubMed] [Google Scholar]
- 45.Funkhouser LJ, Bordenstein SR. Mom knows best: the universality of maternal microbial transmission. PLoS Biol. 2013;11(8):1–9. e1001631. [DOI] [PMC free article] [PubMed]
- 46.Smilowitz JT, Lebrilla CB, Mills DA, German JB, Freeman SL. Breast milk oligosaccharides: structure-function relationships in the neonate. Annu Rev Nutr. 2014;34:143–69. doi: 10.1146/annurev-nutr-071813-105721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Sela DA, Chapman J, Adeuya A, Kim JH, Chen F, Whitehead TR, Lapidus A, Rokhsar DS, Lebrilla CB, German JB, et al. The genome sequence of Bifidobacterium longum subsp. infantis reveals adaptations for milk utilization within the infant microbiome. Proc Natl Acad Sci U S A. 2008;105:18964–9. doi: 10.1073/pnas.0809584105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Turroni F, Foroni E, Serafini F, Viappiani A, Montanini B, Bottacini F, Ferrarini A, Bacchini PL, Rota C, Delledonne M, et al. Ability of Bifidobacterium breve to grow on different types of milk: exploring the metabolism of milk through genome analysis. Appl Environ Microbiol. 2011;77:7408–17. doi: 10.1128/AEM.05336-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Asakuma S, Hatakeyama E, Urashima T, Yoshida E, Katayama T, Yamamoto K, Kumagai H, Ashida H, Hirose J, Kitaoka M. Physiology of consumption of human milk oligosaccharides by infant gut-associated bifidobacteria. J Biol Chem. 2011;286:34583–92. doi: 10.1074/jbc.M111.248138. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sela DA. Bifidobacterial utilization of human milk oligosaccharides. Int J Food Microbiol. 2011;149:58–64. doi: 10.1016/j.ijfoodmicro.2011.01.025. [DOI] [PubMed] [Google Scholar]
- 51.Katayama T. Host-derived glycans serve as selected nutrients for the gut microbe: human milk oligosaccharides and bifidobacteria. Biosci Biotechnol Biochem. 2016;80:621–32. doi: 10.1080/09168451.2015.1132153. [DOI] [PubMed] [Google Scholar]
- 52.Turroni F, Bottacini F, Foroni E, Mulder I, Kim JH, Zomer A, Sanchez B, Bidossi A, Ferrarini A, Giubellini V, et al. Genome analysis of Bifidobacterium bifidum PRL2010 reveals metabolic pathways for host-derived glycan foraging. Proc Natl Acad Sci U S A. 2010;107:19514–9. doi: 10.1073/pnas.1011100107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Turroni F, Strati F, Foroni E, Serafini F, Duranti S, van Sinderen D, Ventura M. Analysis of predicted carbohydrate transport systems encoded by Bifidobacterium bifidum PRL2010. Appl Environ Microbiol. 2012;78:5002–12. doi: 10.1128/AEM.00629-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Egan M, Motherway MO, Kilcoyne M, Kane M, Joshi L, Ventura M, van Sinderen D. Cross-feeding by Bifidobacterium breve UCC2003 during co-cultivation with Bifidobacterium bifidum PRL2010 in a mucin-based medium. BMC Microbiol. 2014;14:282. doi: 10.1186/s12866-014-0282-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Egan M, O’Connell Motherway M, Ventura M, van Sinderen D. Metabolism of sialic acid by Bifidobacterium breve UCC2003. Appl Environ Microbiol. 2014;80:4414–26. doi: 10.1128/AEM.01114-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ventura M, Turroni F, Lima-Mendez G, Foroni E, Zomer A, Duranti S, Giubellini V, Bottacini F, Horvath P, Barrangou R, et al. Comparative analyses of prophage-like elements present in bifidobacterial genomes. Appl Environ Microbiol. 2009;75:6929–36. doi: 10.1128/AEM.01112-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Ventura M, Lee JH, Canchaya C, Zink R, Leahy S, Moreno-Munoz JA, O’Connell-Motherway M, Higgins D, Fitzgerald GF, O’Sullivan DJ, van Sinderen D. Prophage-like elements in bifidobacteria: insights from genomics, transcription, integration, distribution, and phylogenetic analysis. Appl Environ Microbiol. 2005;71:8692–705. doi: 10.1128/AEM.71.12.8692-8705.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Tsuji H, Oozeer R, Matsuda K, Matsuki T, Ohta T, Nomoto K, Tanaka R, Kawashima M, Kawashima K, Nagata S, Yamashiro Y. Molecular monitoring of the development of intestinal microbiota in Japanese infants. Benef Microbes. 2012;3:113–25. doi: 10.3920/BM2011.0038. [DOI] [PubMed] [Google Scholar]
- 59.Martin R, Makino H, Cetinyurek Yavuz A, Ben-Amor K, Roelofs M, Ishikawa E, Kubota H, Swinkels S, Sakai T, Oishi K, et al. Early-Life events, including mode of delivery and type of feeding, siblings and gender, shape the developing gut microbiota. PLoS One. 2016;11:e0158498. doi: 10.1371/journal.pone.0158498. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Turroni F, Ozcan E, Milani C, Mancabelli L, Viappiani A, van Sinderen D, Sela DA, Ventura M. Glycan cross-feeding activities between bifidobacteria under in vitro conditions. Front Microbiol. 2015;6(1030):1–8. [DOI] [PMC free article] [PubMed]
- 61.Turroni F, Milani C, Duranti S, Mancabelli L, Mangifesta M, Viappiani A, Lugli GA, Ferrario C, Gioiosa L, Ferrarini A, et al. Deciphering bifidobacterial-mediated metabolic interactions and their impact on gut microbiota by a multi-omics approach. ISME J. 2016;10:1656–68. doi: 10.1038/ismej.2015.236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Lozupone CA, Stombaugh JI, Gordon JI, Jansson JK, Knight R. Diversity, stability and resilience of the human gut microbiota. Nature. 2012;489:220–30. doi: 10.1038/nature11550. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table S1. Taxonomy and filtering report of samples included in this study. Table S2 ITS-based OTUs shared by an entire sample sets. Table S3 List of bifidobacterial strain-specific primers. Table S4 Metagenomic reads classification of samples TVPR-01, TVPR-02, and TVPR-03. Table S5 List of specific primers for each bifidophage/bifidoprophages identified. (DOCX 37 kb)
Figure S1. ITS-based OTUs shared between mother-milk-infant sample sets. The bar plot represents the percentage of the total bifidobacterial population found in mother-milk, mother-infant, milk-infant and mother-milk-infant samples. (JPG 1306 kb)
Figure S2. Evaluation of presence of TVPR-02 and TVPR-03 microbiome-reconstructed genomes in fecal samples of the infants and mothers as well as in milk samples. Primer sequences are reported in Additional file 1: Table S2. (JPG 235 kb)
Figure S3. Identified bifidophage/bifidoprophage within the mother’s and infant’s metagenomic samples. Genomic maps of phages recall their modular genomic structure indicated by different patterns, which specifies their predicted function (violet: lysogeny module; blue: DNA replication; red: DNA packaging and head; green: tail and tail fiber; yellow: lysis module; black arrows: hypothetical protein; grey arrows: similar to bacterial protein). For each phage is reported the presence within the mother’s and infant’s metagenomic samples through PCR analysis. Primer sequences are reported in Additional file 1: Table S5. (JPG 1173 kb)
Data Availability Statement
Bifidobacterial ITS profiling data for all the sample sets are accessible through the accession number SRP102222. The bifidobacterial sequences reported in this article have been deposited in the GenBank database under accession numbers NAPZ00000000, NAQA00000000, NAQB00000000, NAQC00000000, NAQD00000000, NAQE00000000, NAQF00000000, NAQG00000000, NAQH00000000, NAQI00000000, NAQJ00000000, NAQK00000000, and NAQL00000000. The versions described in this paper are NAPZ01000000, NAQA01000000, NAQB01000000, NAQC01000000, NAQD01000000, NAQE01000000, NAQF01000000, NAQG01000000, NAQH01000000, NAQI01000000, NAQJ01000000, NAQK01000000, and NAQL01000000. Shotgun metagenomics data have been deposited in GenBank under the following accession numbers SRP102273.