

Abstract
Burkholderia cenocepacia is a versatile opportunistic pathogen that survives in a wide variety of environments, which can be limited in nutrients such as nitrogen. We have previously shown that the sigma factor σ54 is involved in the control of nitrogen assimilation and virulence in B. cenocepacia H111. In this work, we investigated the role of the σ54 enhancer binding protein NtrC in response to nitrogen limitation and in the pathogenicity of H111. Of 95 alternative nitrogen sources tested the ntrC showed defects in the utilisation of nitrate, urea, L-citrulline, acetamide, DL-lactamide, allantoin and parabanic acid. RNA-Seq and phenotypic analyses of an ntrC mutant strain showed that NtrC positively regulates two important phenotypic traits: exopolysaccharide (EPS) production and motility. However, the ntrC mutant was not attenuated in C. elegans virulence.
Introduction
The betaproteobacterium Burkholderia cenocepacia is an opportunistic pathogen that thrives in different environments, which can be limited in essential elements such as nitrogen [1, 2]. Bacterial adaptations to changes in nitrogen availability have been shown to be stringently regulated [3–7]. Enterobacteria respond to nitrogen starvation by activating the nitrogen regulatory response (Ntr) to facilitate N scavenging from alternative nitrogen sources. The Ntr system monitors the intracellular ratio of glutamine to α-ketoglutarate. Under nitrogen limiting conditions, the PII signal transduction proteins encoded by glnB and glnK are uridylylated and, by controlling the kinase and phosphatase activities of the regulator NtrB, regulate the transcription of nitrogen-regulated target genes [3, 6, 8, 9]. NtrB is a sensor kinase, and is part of the NtrBC two-component regulatory system. Under nitrogen limiting conditions, NtrB phosphorylates the response regulator NtrC, which then binds to DNA sequences in the promoter and together with the alternative sigma factor σ54 (or RpoN) activates transcription [10–12]. The sigma factor σ54 reversibly associates with the core RNA polymerase and recognizes its cognate promoter sequences via defined consensus sequences at positions –12 and –24 bp (relative to the transcription start site) [13]. The initiation of σ54-dependent transcription usually requires such an interaction with an enhancer binding protein (EBP). The specific protein involved varies depending on the respective environmental signals. NtrC is the EBP in the case of nitrogen starvation conditions [14, 15]. EBPs share a conserved modular structure, which consists of three domains: i) an amino terminal regulatory domain, ii) a central catalytic domain that belongs to the AAA+ superfamily of ATPases and is required for interaction with σ54, iii) a carboxy-terminal DNA-binding domain with a helix–turn–helix motif that is required for recognition of upstream activator sequences [16]. The EBP catalyzes ATP hydrolysis and thereby promotes conversion of the closed promoter into an open promoter complex from which transcription can proceed [16–20]. In enterobacteria, hexamers of phosphorylated NtrC bind to an upstream activator sequence (UAS), which is usually located 100–150 nucleotides upstream of the transcriptional start site [21, 22].
We have previously characterized the response of B. cenocepacia H111 to nitrogen starvation, a condition that is relevant during chronic lung infection [2, 23] and showed that nitrogen assimilation is largely dependent on the sigma factor σ54 [24]. Further phenotypic analysis of a σ54 mutant showed that this sigma factor is also important for other traits, which have been previously shown to play important roles during infection with B. cenocepacia [18, 25–30]: exopolysaccharide (EPS) production, biofilm formation and motility. The sigma factor σ54 is also required for H111 virulence in the Caenorhabditis elegans infection model [24].
Among the regulatory genes highly up-regulated under nitrogen starvation conditions was the two-component regulatory system NtrBC, suggesting that NtrC is the EBP of σ54 during nitrogen starvation [24]. In this study, the ntrC gene (I35_2149) was mutated in B. cenocepacia H111 and the derived NtrC regulon during nitrogen starvation was compared with the previously identified σ54 regulon. Phenotypic analysis of the ntrC mutant strain confirmed the transcriptomics data and showed that NtrC controls utilization of several nitrogen sources, EPS production and motility. While biofilm formation was partially compromised in the ntrC mutant, NtrC was not important for C. elegans virulence, suggesting that for this phenotype σ54 interacts with other EBPs or σ54 functions independently of EBPs.
Material and methods
Bacterial strains, media and growth conditions
The bacterial strains and plasmids used in this study are listed in S1 Table. E. coli and B. cenocepacia H111 cells were routinely grown in Luria broth (LB) [31] at 37°C using the following concentrations of antibiotics (in μg/ml): ampicillin (100 for E. coli), chloramphenicol (20 for E. coli and 80 for B. cenocepacia) and gentamycin (10 for both E. coli and B. cenocepacia). Cultures for RNA-Seq were grown first in AB minimal medium [32] using 10 mM sodium citrate as carbon source and 15 mM ammonium chloride (NH4Cl) as nitrogen source. Nitrogen starved conditions were created using AB minimal medium containing 0.3 mM NH4Cl. Cultures were grown in 500 ml Erlenmeyer flasks containing 100 ml medium on a shaker (200 rpm) at 37°C. For each strain or condition, the growth of three independent cultures was analysed.
Construction of B. cenocepacia H111 mutant strains
Plasmid DNA from E. coli strains was obtained by using the QIAprep Spin Miniprep Kit (QIAGEN). To generate H111-ntrC (an insertional mutant in I35_2149, the ortholog of B. cenocepacia J2315 BCAL2222), a 508 bp internal fragment of I35_2149 was amplified with Ex Taq polymerase (TaKaRa) using primers ntrC_mut_new_F and ntrC_mut_new_R (see S1 Table). The PCR product was cloned into pGEM®-T Easy Vector (Promega) and then sub-cloned into pSHAFT2 [33] as a NotI fragment to generate pSHAFT2-ntrC. The resulting plasmid was mobilized into B. cenocepacia H111 wild type by triparental mating. Correct genomic integration was verified by PCR using oligos ntrC_Comp_F and pSHAFT_R. To complement H111-ntrC, the complete I35_2149 ORF was amplified with Phusion High-Fidelity DNA polymerase (Thermo Fisher Scientific) using oligos ntrC_Comp_F and ntrC_Comp_R (S1 Table). The PCR product was first cloned into pGEM®-T Easy Vector, digested with EcoRI, and then cloned into EcoRI-digested pBBR1MSC-5 (GmR) to create the complementing plasmid pBBR1-ntrC. The sequence of the I35_2149 ORF in the complementation vector was verified by DNA sequencing. The complementing plasmid was mobilized into H111-ntrC by triparental mating to generate the complemented strain.
RNA-Seq and data analysis
H111 wild-type and mutant strains were grown to exponential phase in AB minimal medium containing 15 mM NH4Cl, and washed twice in AB minimal medium without NH4Cl. The cells were further incubated for 1 hour under nitrogen starvation conditions. Three independent RNA-Seq experiments were performed from three independent biological replicates. Total RNA was extracted using a modified hot acid phenol protocol [34]. The complete removal of genomic DNA using RQ1 RNase-Free DNase (Promega) was verified by a PCR reaction with 40 cycles. Samples were further purified using the RNeasy Mini Kit (QIAGEN) and RNA quality was checked using RNA Nano Chips (Agilent 2100 Bioanalyzer). A total of 150 ng RNA was used for cDNA synthesis and library preparation using the Ovation® Complete Prokaryotic RNA-Seq Library System from NuGEN. The cDNA libraries were analyzed and quantified by capillary electrophoresis using D1000 ScreenTape from Agilent (size range 100–800 bp). Illumina single-end sequencing was performed on a HiSeq2500 instrument. The sequence reads were processed and then mapped to the recently finished H111 genome sequence (accession no. HG938370, HG938371, and HG938372) [35] using CLC Genomics Workbench v7.0 (CLC bio) allowing up to 2 mismatches per read. The mapped reads were analysed using the DESeq software [36]. The RNA-Seq raw data files are accessible through the GEO Series accession number GSE95607.
Functional analysis was based on the EggNOG annotation that is available for B. cenocepacia J2315 [37] and was performed as previously described [24]. Distribution of categories was determined using a Fischer test with the online quick calculator of GraphPad (p-value < 0.01).
Phenotypic analysis of mutant strains
The utilization of nitrogen sources by H111 wild-type and ntrC mutant strains was assessed using Biolog PM3B plates according to the manufacturer’s instructions as described previously [38]. After inoculation, the plates were incubated at 37°C for 24 hours before the OD590 of each well was measured by a plate-reader (TECAN). The assay was performed on biological duplicate cultures for each strain. The OD590 of these was taken to calculate the average values and standard deviation. The 18 differentially utilized N sources were determined using the following criteria: i) ratio of average final OD590 (wild-type vs. mutant or vice-versa) > 2; ii) the higher OD590 average value > 0.3; iii) both standard deviations (of wild-type and mutant) < 0.1.
The assimilation of nitrogen sources was also assayed by observing growth in modified AB minimal medium, where 15 mM NH4Cl was replaced by other nitrogen sources at concentrations according to the number of N atoms in the molecule. The cells were incubated in 16 ml medium with a starting OD600 of 0.05 for 4 days at 37°C with shaking at 40 rpm. The final OD600 of the independent biological triplicate cultures for each strain was used to calculate the mean and the standard deviation. For each nitrogen source, all 3 strains (wild-type, ntrC mutant and the complemented mutant) were tested in parallel.
EPS production was assessed on modified YEM medium plates (0.4% mannitol, 0.05% yeast extract, 1.5% agar [39]). All 3 strains to be tested were streaked in parallel on the YEM plate to eliminate variations. Independent biological triplicate cultures for each strain were tested. Images were taken for evaluation after 24 hours incubation at 37°C.
Biofilm formation was quantified in a microtiter-plate assay as described by Huber et al. [40]. For each condition, all 3 strains were tested in parallel in one plate and independent biological triplicate cultures for each strain were tested. Each culture was tested in at least 4 wells in one plate. The biofilm index was calculated for each well. The mean and the standard deviation of all the biofilm index data for each strain were used to make the histogram. A two-tailed unpaired Student’s t test was used to determine the significance between either mutant or complement and wild type.
Swarming and swimming activity were tested in the same way as previously described [41] with the following modifications: swarming and swimming assays were carried out on AB minimal medium plates containing 10 mM glucose as carbon source and 0.1% casamino acids, solidified with 0.4% and 0.2% agar, respectively. Independent biological triplicate cultures of each strain were tested. Swarming assays were carried out with minor modifications. In detail, swarming plates were freshly prepared and air-dried for half an hour under the laminar flow after solidification. Pre-culture cells were washed twice with AB minimal medium and adjusted to OD600 0.5. Then 3 μl of each culture was inoculated at the center of a plate. The plates were incubated in humid conditions at 37°C for 5 days. The colony swarming diameter for each plate was measured. For both swarming and swimming assays, at least independent biological triplicate cultures of each strain were tested and the 3 strains were always tested in parallel for each assay. To normalize the data, for each assay the average diameter of the wild type was set to 100% and used as a reference. The ntrC mutant and complement strain values were shown as a percentage of this reference value. Swimming and swarming motility were statistically analysed using a two-tailed unpaired Student’s t test, with either the mutant or complemented mutant compared to the wild-type strain.
Pathogenicity tests using Caenorhabditis elegans Bristol N2 strain and Galleria mellonella were performed as previously described [24] and [38], respectively. For C. elegans, the number of synchronized L1 larvae was recorded after seeding with the bacteria into each well of a 96-well plate to test. After 48 hours co-incubation at 20°C, the worms were scored according to their developmental stages and the reduction in the total number of surviving larvae was regarded as “dead”. The assays using both infection models were carried out in triplicate with all 3 strains tested in parallel.
qPCR analysis
The qPCR analysis was carried out using Brilliant III Ultra-Fast SYBR® Green QPCR Master Mix (Agilent, Switzerland) and an Mx3000P instrument (Agilent, Switzerland). The cDNA was prepared from an independent biological replicate (of both wild-type and ntrC mutant strains) as previously described [42]. Each PCR reaction was run in triplicate with 3 dilutions of cDNA (15, 7.5 and 3.75 ng) using 15 μl 2x Brilliant III Ultra-Fast SYBR® Green QPCR Master Mix, and 5 μM of individual primers in a total volume of 30 μl. Fold-changes in transcription and the standard deviation of 9 sample dilutions were calculated using the ΔΔ CT method [43]. The primary σ factor gene rpoD was used as a reference for normalization. All the primers used are listed in S1 Table.
Results
Construction and growth analysis of a B. cenocepacia H111 ntrC mutant and a complemented derivative
To study the role of the σ54 activator protein NtrC in response to nitrogen limitation, an H111 ntrC mutant and a complemented mutant were constructed (S1 Table). The gene I35_2149, an ortholog of B. cenocepacia strain J2315 gene BCAL2222, was chosen since it showed 55% amino acid identity and 67% amino acid similarity to NtrC of Escherichia coli. Furthermore, its transcription was found to be significantly up-regulated by nitrogen starvation and significantly down-regulated in the absence of the alternative sigma factor σ54, strongly suggesting that NtrC is the EBP of σ54 in nitrogen limited environments [44]. Similar to other bacteria, the gene is located in an operon downstream of the gene coding for its potential sensor kinase NtrB and the gene glnA which encodes a glutamine synthetase. While the ntrC mutant grew at a similar rate and to the same final optical density (i. e. OD600 of 4) as the wild type in LB, the mutant had a moderate growth delay in AB minimal medium containing citrate as carbon source (S1 Fig). The complemented ntrC mutant displayed a growth defect in both media, but was able to eventually reach the same final optical density (OD600) as the wild type (data not shown). In contrast, on LB or AB minimal medium agar plates, the growth of the wild type and the complemented ntrC mutant was indistinguishable after 24 and 48 hours of incubation, respectively. The growth defect of the complemented mutant may have been due to the over-expression of ntrC from the pBBR1MCS-5 plasmid. Our qPCR result showed that when the cells were grown in LB until stationary phase, ntrC transcription in the complemented strain was 162 fold higher than the wild-type strain (as determined by qPCR).
NtrC affects the utilization of alternative nitrogen sources
The ability of the wild-type strain and the ntrC mutant to utilize 95 different nitrogen sources was examined using Biolog plate assays (S2 Table). The utilization of 18 N sources was found to be affected by NtrC. In 7 cases (nitrate, urea, L-citrulline, acetamide, DL-lactamide, allantoin and parabanic acid), the ntrC mutant was compromised in substrate utilization. In contrast, with 11 N sources (biuret, L-cysteine, L-isoleucine, L-methionine, hydroxylamine, methylamine, ethylamine, ethylenediamine, N-acetyl-D-galactosamine, uric acid, ε-amino-N-caproic acid) the ntrC mutant showed enhanced growth compared to the wild-type. The utilization of selected N sources was also tested using a different experimental setup (Fig 1). When incubated at 37°C in AB minimal medium containing 15 mM ammonium as N source, the H111 wild-type, ntrC mutant and ntrC complemented strains grew at similar rates. Consistent with the Biolog results, the ntrC mutant was unable to use nitrate or urea as sole N source. However, in the case of L-citrulline and histamine, growth was only mildly affected in the mutant compared with the wild-type. Heterologous complementation with ntrC partially rescued growth of mutant with most of the N sources tested, except for histamine.
Fig 1. Nitrogen sources differentially utilised by the wild-type, ntrC mutant and the complemented strains.

Cells were grown at 37°C for 4 days with slow agitation in order to synchronize the growth. 3 independent cultures of each strain were tested with each N source. Columns and error bars indicate the average and the standard deviation of final OD600, respectively.
Mapping the NtrC regulon in B. cenocepacia H111
To identify genes controlled by NtrC, RNA-Seq was performed on the ntrC mutant and the wild-type strain grown under nitrogen limiting conditions. For this, RNA was extracted from independent biological triplicate cultures of wild-type and ntrC mutant cells grown first in AB minimal medium containing 15 mM NH4Cl until exponential phase (OD600 = 0.5, S1 Fig) and then shifted to nitrogen limiting conditions (0.3 mM NH4Cl) for one hour (shift experiment). Among the 150 top ranked differentially expressed genes (DESeq analysis p-value < 10−20, absolute log2(Fold Change) > 2.3) (Fig 2 and Table 1), 123 genes (82%) showed decreased transcription in the ntrC mutant. Genes involved in nitrogen assimilation such as the glutamine synthetase (glnA, I35_2151) and one of the two PII sensor proteins (glnB, I35_2936) showed 4.8- and 6.5- fold (log2 fold change) reduced transcription, respectively, in the ntrC mutant. In line with the inability of the ntrC mutant to grow with urea or nitrate as the sole nitrogen source, the genes required for urea (I35_0765–0775 and I35_7283–7286) and nitrate (I35_5545–5548) transport and utilization displayed down-regulated transcription in the ntrC mutant. Sixty-five percent of the genes found to be activated by NtrC have previously been shown to have increased transcription under nitrogen limited conditions [24], suggesting that B. cenocepacia NtrC plays a major role in the control of gene transcription during nitrogen deprivation. Among the top 150 genes significantly differentially transcribed in the ntrC mutant (Table 1), 95% were also regulated by σ54, suggesting that, as is the case in other bacteria, NtrC is the EBP of σ54 required for activating transcription of genes under nitrogen limiting conditions (see below). Among the common target genes of σ54 and NtrC, many were potentially involved in nitrogen metabolism: the urease and nitrate reductase gene clusters described above, a cluster coding for proteins containing a transglutaminase-like domain (I35_5080–83), a xanthine dehydrogenase (I35_0688–0689), a gene cluster containing a urate oxidase (I35_1963–64), several genes coding for components of ABC transporters (I35_0158, I35_4451, I35_4872–73, I35_5228–29, I35_5552, I35_5645, I35_6515, I35_7109–10, I35_7737–40), and an alanyl-alanine dipeptidase and its associated transporter (I35_5644–46). Moreover, other genes showed σ54 –NtrC–dependent transcription, including cidAB (I35_3288–89) coding for a putative holin and anti-holin system, I35_4672–73 encoding a poly-beta-hydroxyalkanoate (PHB) depolymerase and a glutathione-S-transferase, and I35_4766 (an ortholog of BCAM0853), which is part of the bce-I cluster that encodes the main B. cenocepacia EPS cepacian. We found several transcriptional regulators with decreased transcription in the ntrC mutant, including two genes coding for an ethanolamine operon regulatory protein (I35_0068 and I35_7815), I35_1967, ntrB, I35_4176, I35_4535, I35_4653, nasT (I35_5551), I35_5643, I35_5874 and I35_6218. An analysis of the categories associated with the top 150 NtrC-regulated genes, revealed that, beside category E (amino acid metabolism and transport), category N (motility) is over-represented among the genes positively regulated by NtrC (S2 Fig). In fact, Table 1 contains several genes involved in flagellar biosynthesis (I35_3089–90 and I35_3103) and rotation (motAB, I35_0133–34), and a gene (I35_0139) involved in chemotaxis.
Fig 2. Differential transcript expression in the ntrC mutant compared to the wild type.

MA plot showing the log2 fold change in transcript expression of B. cenocepacia H111 wild type and ntrC mutant strains grown under nitrogen limited conditions. The top regulated genes (p-value < 0.001, absolute log2 (Fold Change) > 1.5) are shown in color: genes with increased transcription in the ntrC mutant compared to the wild type are indicated in orange, down-regulated genes in green. The names of the genes of particular interest are labelled.
Table 1. List of the 150 genes with statistically significant differential expression, comparing the expression profile of the ntrC mutant with the profile of the wild type (DE-Seq analysis, p-value < 10−20, absolute log2(Fold Change) > 2.3).
| Class | Locus IDa | Orthologs J2315b | Descriptiona | Gene name | Log2 ntrC vs wtc |
|---|---|---|---|---|---|
| Amino acid metabolism and transport | I35_0158 | BCAL0151 | Leucine-, isoleucine-, valine-, threonine-, and alanine-binding protein | -3.6 | |
| I35_0767 | BCAL3106 | Urease alpha subunit | ureC | -5.2 | |
| I35_0769 | BCAL3104 | Urease gamma subunit | ureA | -4.4 | |
| I35_0772 | BCAL3102 | Urea ABC transporter, ATPase protein | urtD | -4.2 | |
| I35_0774 | BCAL3099 | Urea ABC transporter, permease protein | urtB | -5.9 | |
| I35_0775 | BCAL3098 | Urea ABC transporter, urea binding protein | -4.8 | ||
| I35_1846 | BCAL1926 | Homoserine dehydrogenase | hom | 2.8 | |
| I35_2151 | BCAL2224 | Glutamine synthetase type I | glnA | -4.8 | |
| I35_2936 | BCAL0729 | Nitrogen regulatory protein P-II | glnB | -6.5 | |
| I35_4872 | BCAM0952 | Putrescine transport ATP-binding protein | potA | -3.1 | |
| I35_4873 | BCAM0953 | ABC transporter, periplasmic spermidine putrescine-binding protein | potD | -3.1 | |
| I35_5082 | BCAM1235 | Protein containing transglutaminase-like domain | -9.4 | ||
| I35_5083 | BCAM1236 | Large protein containing transglutaminase-like domain | -6.0 | ||
| I35_5229 | BCAM1377 | ABC-type spermidine/putrescine transport systems,ATPase components |
4.0 | ||
| I35_7109 | BCAS0112 | ABC-type arginine/histidine transport system,permease component |
-5.2 | ||
| I35_7110 | BCAS0113 | Histidine ABC transporter, ATP-binding protein | hisP | -3.8 | |
| I35_7286 | BCAS0272 | Urea carboxylase | -6.0 | ||
| I35_7447 | BCAS0409 | Zinc metalloprotease | zmpA | 2.4 | |
| I35_7737 | BCAS0574 | Glutamate transport ATP-binding protein | -4.3 | ||
| I35_7739 | BCAS0576 | amino acid ABC transporter, permease protein | na(m) | ||
| I35_7740 | BCAS0577 | ABC-type amino acid transport, periplasmic component | -10.1 | ||
| I35_7857 | BCAS0734 | Pyridine nucleotide-disulphide oxidoreductase | -6.8 | ||
| I35_7858 | BCAS0735 | Beta-ureidopropionase | -6.9 | ||
| I35_7874 | BCAS0751 | Gamma-glutamyltranspeptidase | -4.8 | ||
| Carbohydrate metabolism and transport | I35_1964 | BCAL2040 |
Chitooligosaccharide deacetylase; putative uricase |
-3.2 | |
| I35_2891 | BCAL0782 | Beta-hexosaminidase | 4.7 | ||
| Cell wall, membrane, envelope biogenesis | I35_0402 | BCAL3473 | Outer membrane protein (porin) | -4.2 | |
| I35_0812 | BCAL3057 | Soluble lytic murein transglycosylase | 3.2 | ||
| I35_3042 | BCAL0624 | Outer membrane protein (porin) | -6.4 | ||
| I35_3288 | BCAL3508 | CidA-associated membrane protein | cidB | -6.0 | |
| I35_4371 | BCAM0478 | Glucosamine—fructose-6-phosphate aminotransferase [isomerizing] |
glmS2 | -5.0 | |
| I35_4934 | BCAM1015 | Outer membrane protein (porin) | -4.0 | ||
| I35_5228 | BCAM1376 | Porin, Gram-negative type | 5.6 | ||
| I35_5644 | BCAM1769 | D-alanyl-D-alanine dipeptidase | ddpX | -4.5 | |
| I35_5658 | BCAM1780 | Lipoprotein | nlpD | -3.5 | |
| I35_7271 | BCAS0256 | Outer membrane protein (porin) | -3.2 | ||
| Cell motility | I35_0133 | BCAL0126 | Flagellar motor rotation protein | motA | -2.7 |
| I35_0134 | BCAL0127 | Flagellar motor rotation protein | motB | -3.1 | |
| I35_0139 | BCAL0132 | Chemotaxis protein methyltransferase | cheR | -2.5 | |
| I35_3089 | BCAL0577 | Flagellar hook-associated protein | flgL | -2.9 | |
| I35_3090 | BCAL0576 | Flagellar hook-associated protein | flgK | -3.2 | |
| I35_3103 | BCAL0561 | Flagellar biosynthesis protein | flgN | -3.3 | |
| Coenzyme metabolism | I35_1039 | BCAL1047 | Pyridoxal kinase | pdxY | -2.4 |
| I35_1623 | BCAL1711 | Component of cobalt chelatase involved in B12 biosynthesis | cobN | 3.5 | |
| I35_5550 | BCAM1687 | Uroporphyrinogen-III methyltransferase | -11.4 | ||
| Defense mechanism | I35_3251 | BCAL0420 | Type I restriction-modification system, restriction subunit R | 2.9 | |
| Energy production and conversion | I35_1414 | BCAL1516 | Dihydrolipoamide succinyltransferase component | sucB | 2.9 |
| I35_1735 | BCAL1819 | Oxidoreductase (flavoprotein) | -5.4 | ||
| I35_2213 | BCAL2287 | Fumarate hydratase class I | 3.2 | ||
| I35_2268 | BCAL2344 | NADH ubiquinone oxidoreductase chain A | nuoA | 2.4 | |
| I35_2713 | BCAL0957 | Succinyl-CoA ligase [ADP-forming] alpha chain | sucD | 2.4 | |
| I35_3124 | BCAL0541 | Aminobutyraldehyde dehydrogenase | -2.7 | ||
| I35_4890 | BCAM0970 | Succinate dehydrogenase iron-sulfur protein | sdhB | 2.3 | |
| I35_4892 | BCAM0972 | Citrate synthase | gltA | 3.1 | |
| I35_5545 | BCAM1683 | Assimilatory nitrate reductase large subunit | -6.7 | ||
| I35_5546 | BCAM1684 | Nitrite reductase [NAD(P)H] small subunit | -6.8 | ||
| I35_5547 | BCAM1685 | Nitrite reductase [NAD(P)H] large subunit | nirB | -6.5 | |
| I35_7856 | BCAS0733 | Dihydropyrimidine dehydrogenase [NADP+] | -5.1 | ||
| Inorganic ion transport and metabolism | I35_2600 | BCAL2740 | HoxN/HupN/NixA family nickel/cobalt transporter | hoxN | -4.7 |
| I35_4451 | BCAM0556 | Dibenzothiophene desulfurization enzyme B | na(m) | ||
| I35_5230 | BCAM1378 |
ABC Fe3+ siderophore transporter inner membrane subunit |
3.8 | ||
| I35_5548 | BCAM1686 | Nitrate/nitrite transporter | -6.6 | ||
| I35_5552 | BCAM1689 | Nitrate ABC transporter, nitrate-binding protein | -4.2 | ||
| I35_5646 | BCAM1771 | Dipeptide transport system permease protein | dppB | -3.2 | |
| I35_5693 | BCAM1814 | Cyclohexanone monooxygenase | -2.8 | ||
| I35_7283 | BCAS0269 |
Urea carboxylase-related ABC transporter, periplasmic protein |
na(m) | ||
| Lipid metabolism | I35_4672 | BCAM0774 | Poly-beta-hydroxyalkanoate depolymerase | -6.6 | |
| Nucleotide metabolism and transport | I35_0495 | BCAL3380 | Allantoicase | -3.5 | |
| I35_0689 | BCAL3172 | Xanthine dehydrogenase, molybdenum binding subunit |
xdhB | -3.1 | |
| I35_7854 | BCAS0731 | Dihydropyrimidinase | dhT | -3.7 | |
| I35_7855 | BCAS0732 | Putative pyrimidine permease in reductive pathway | -3.8 | ||
| Post translational modifications | I35_0765 | BCAL3108 | Urease accessory protein | ureF | -7.1 |
| I35_0766 | BCAL3107 | Urease accessory protein | ureE | -7.8 | |
| I35_2591 | BCAL2731 | ATP-dependent Clp protease adaptor protein | clpS | -2.6 | |
| I35_2821 | BCAL0849 | Putative lipoprotein | 3.1 | ||
| I35_3125 | BCAL0540 | ATP-dependent protease domain protein | -4.1 | ||
| I35_4673 | BCAM0775 | Glutathione S-transferase | -5.3 | ||
| I35_5615 | BCAM1744 | Extracellular protease precursor | -2.3 | ||
| Secondary structures | I35_2997 | BCAL0668 | Dienelactone hydrolase and related enzymes | -2.6 | |
| Transcription and signal transduction | I35_0068 | BCAL0066 | Ethanolamine operon regulatory protein | -5.7 | |
| I35_0200 | BCAL0209 | Histone acetyltransferase HPA2 | -5.1 | ||
| I35_1574 | BCAL1663 | Serine protein kinase | prkA | -4.0 | |
| I35_1967 | BCAL2043 | Transcriptional regulator, GntR family | -5.9 | ||
| I35_2150 | BCAL2223 | Nitrogen regulation protein NR(II) | ntrB | -5.6 | |
| I35_3013 | BCAL0652 | EAL domain protein | -7.2 | ||
| I35_4176 | BCAM0176 | Transcriptional regulator, AsnC family | -3.6 | ||
| I35_4382 | BCAM0489 | Mercuric resistance operon regulatory protein | -3.6 | ||
| I35_4535 | BCAM0639 | Two-component response regulator | -6.5 | ||
| I35_4613 | BCAM0715 | Signal transduction histidine kinase | 3.5 | ||
| I35_4653 | BCAM0754 | Transcriptional regulator, TetR family | -3.6 | ||
| I35_5551 | BCAM1688 | Response regulator | nasT | -8.1 | |
| I35_5643 | BCAM1768 | Transcriptional regulators | -6.1 | ||
| I35_5806 | BCAM1975 | Ethanolamine operon regulatory protein | -4.8 | ||
| I35_5874 | BCAM2039 | Two-component response regulator | -5.0 | ||
| I35_6218 | BCAM2327 | Transcriptional regulator | -6.7 | ||
| Translation | I35_2322 | BCAL2395 | Cytoplasmic axial filament protein CafA and Ribonuclease G | cafA | -2.5 |
| I35_2859 | BCAL0812 | Ribosome hibernation protein | yhbH | -3.9 | |
| Others | I35_0339 | BCAL0348 | Uncharacterized protein | impA | 3.4 |
| I35_0341 | BCAL0350 | Hypothetical protein | 2.6 | ||
| I35_0688 | BCAL3173 | Xanthine dehydrogenase, iron-sulfur cluster and FAD-binding subunit A | xdhA | -3.0 | |
| I35_0743 | Hypothetical protein | 2.9 | |||
| I35_0963 | BCAL2904 | Hypothetical protein | -2.7 | ||
| I35_1398 | BCAL1500 | Transposase and inactivated derivatives | -5.2 | ||
| I35_1575 | BCAL1664 | Hypothetical protein | -3.4 | ||
| I35_1576 | BCAL1665 | SpoVR-like protein | -3.0 | ||
| I35_1588 | BCAL1677 | Type 1 fimbriae major subunit | fimA | 5.2 | |
| I35_1734 | BCAL1818 | Zn-dependent hydrolases, including glyoxylases | -6.6 | ||
| I35_1736 | Oxidoreductase (flavoprotein) | -5.2 | |||
| I35_1739 | BCAL1822 | Putrescine transport system permease protein | potH | 3.9 | |
| I35_1872 | BCAL1952 | Hypothetical protein | 4.4 | ||
| I35_1881 | BCAL1961 | Ankyrin repeat protein | -2.5 | ||
| I35_1963 | BCAL2039 | Urate oxidase | -3.4 | ||
| I35_2376 | BCAL2448 | Phenazine biosynthesis protein PhzF like | 3.1 | ||
| I35_2564 | BCAL2703 | Branched-chain amino acid transport ATP-binding protein | -2.4 | ||
| I35_3012 | BCAL0653 | Hypothetical protein | -4.3 | ||
| I35_3082 | BCAL0584 | Outer membrane porin | -3.3 | ||
| I35_3289 | BCAL3509 | Holin-like protein | cidA | -8.0 | |
| I35_4192 | BCAM0193 | Hypothetical protein | 6.0 | ||
| I35_4193 | BCAM0194 | Hypothetical protein | 5.9 | ||
| I35_4195 | BCAM0196 | Hypothetical protein | 4.7 | ||
| I35_4401 | BCAM0507 | Uncharacterized protein conserved in bacteria | -3.5 | ||
| I35_4471 | BCAM0576 | Hypothetical protein | -3.4 | ||
| I35_4651 | BCAM0752 | Hydrolase-related protein | -3.6 | ||
| I35_4652 | BCAM0753 | Hypothetical protein | -3.9 | ||
| I35_4669 | BCAM0770 | Hypothetical protein | -7.4 | ||
| I35_4766 | BCAM0853 | Transposase and inactivated derivatives | -4.1 | ||
| I35_4943 | BCAM1098 | MutT/nudix family protein | -2.7 | ||
| I35_5080 | BCAM1233 | Protein containing domains DUF404, DUF407 | -8.2 | ||
| I35_5081 | BCAM1234 | Protein containing domains DUF403 | -9.4 | ||
| I35_5153 | BCAM1304 | Phage-related protein | -2.4 | ||
| I35_5345 | BCAM1491 | Hypothetical protein | -4.8 | ||
| I35_5549 | Hypothetical protein | -11.1 | |||
| I35_5645 | BCAM1770 | Dipeptide-binding ABC transporter, periplasmic component | -4.8 | ||
| I35_5655 | BCAM1777A | Hypothetical protein | -3.6 | ||
| I35_5695 | BCAM1816 | Hypothetical protein | -2.7 | ||
| I35_5683 | BCAM1804 | Methyl-accepting chemotaxis protein | -2.9 | ||
| I35_5753 | BCAM1927 | Membrane-fusion protein | -3.1 | ||
| I35_6093 | BCAM2207 | Hypothetical protein | -2.5 | ||
| I35_6098 | BCAM2209 | Hypothetical protein | -3.0 | ||
| I35_6344 | BCAM2444 | Conserved domain protein | -4.1 | ||
| I35_6461 | BCAM2564 | Aerotaxis sensor receptor protein | -4.9 | ||
| I35_6515 | BCAM2618 | Histidine ABC transporter, histidine-binding periplasmic protein | hisJ | -4.4 | |
| I35_6573 | BCAM2679 | Hypothetical protein | -9.0 | ||
| I35_6574 | BCAM2680 | Putative exported protein | -3.6 | ||
| I35_6579 | Hypothetical protein | -4.6 | |||
| I35_7149 | Hypothetical protein | -3.7 | |||
| I35_7282 | BCAS0267a | 3',5'-cyclic-nucleotide phosphodiesterase | -4.7 | ||
| I35_7285 | BCAS0271 | Urea carboxylase-related aminomethyltransferase | na(m) | ||
| I35_7735 | BCAS0571 | Salicylate hydroxylase | -4.5 | ||
| I35_7815 | Ethanolamine operon regulatory protein | -4.2 | |||
aNomenclature and description according to GenBank file (accession no. HG938370, HG938371, and HG9383729).
bOrthologs were identified as described in the Material and Methods section.
cFold change (FC) of transcription, comparing ntrC mutant with wild type grown under nitrogen limited conditions.
na, not applicable because the read number in the mutant is equal to 0.
all the genes with an rpoN box in the promoter region are indicated in bold.
Among the 27 genes with increased transcription in the ntrC mutant (Table 1) we found fimA (I35_1588 encoding a type I fimbria), zmpA (I35_7447 coding for a zinc metalloprotease), two genes in a type VI secretion system cluster (I35_0339 and I35_0341), I35_3251 coding for a subunit of a type I restriction-modification system and several genes involved in the Tricarboxylic Acid Cycle (TCA) cycle (sucB, sucD, sdhB, gltA and I35_2213 coding for a fumarase). Using a less stringent p-value threshold (DESeq analysis p-value < 10−13, absolute log2(Fold Change) > 1.5; S3 Table) additional genes involved in nitrogen metabolism and motility were identified as NtrC-regulated. Additionally, two genes in the cepacian clusters bceI (I35_4767 and I35_4772), the gene rhlA (I35_6233) coding for an enzyme catalysing the first step in biosynthesis of rhamnolipids, I35_1797 coding for the RNA binding protein Hfq and several protease-encoding genes (clpS, clpX, lon, ybbK) showed decreased transcription in the ntrC mutant. The NtrC-dependent transcription of genes involved in cepacian, rhamnolipid biosynthesis and cell motility was validated by an independent qPCR analysis (Table 2).
Table 2. Validation of selected RNA-Seq results by qPCR.
| Locus IDa | J2315 orthologsb | Descriptiona | Gene name | Log2 FC MT vs WTc | Log2 FC MT vs WTd |
|---|---|---|---|---|---|
| I35_0767 | BCAL3106 | Urease alpha subunit | ureC | -5.0 | -5.2 |
| I35_2151 | BCAL2223 | Glutamine synthetase | glnA | -3.6 | -4.8 |
| I35_4771 | BCAM0858 | Polysaccharide export lipoprotein | bceE | -1.2 | -2.6 |
| I35_4928 | BCAM1009 | O-antigen acetylase | -1.1 | -4 | |
| I35_6233 | BCAM2340 | 3-(3-hydroxyalkanoyloxy) alkanoic acids synthase | rhlA | -1.6 | -2.9 |
| I35_0133 | BCAL0126 | Flagellar motor rotation protein | motA | -0.9 | -2.7 |
| I35_3089 | BCAL0577 | Flagellar hook-associated protein | flgL | -2.4 | -2.9 |
| I35_3103 | BCAL0561 | Flagellar biosynthesis protein | flgN | -1.8 | -3.3 |
aNomenclature and description according to GenBank file (accession no. HG938370, HG938371, and HG938372).
bOrthologs were identified as described in the Material and Methods section.
cFold change (FC) of transcription determined by qPCR, comparing the ntrC mutant (MT) with the wild type (WT) grown in AB minimal medium with a shift experiment. The standard deviation is less than 10% of the fold change.
dFold change (FC) of transcription determined by RNA-Seq, comparing the ntrC mutant (MT) with the wild type (WT) grown in AB minimal medium (shift experiment).
NtrC regulates EPS production but has only a slight effect on biofilm formation
The results from this comparative transcription analysis (Table 1 and S3 Table) suggest that NtrC is required for activation of transcription of the cepacian clusters (bceI and bceII). Cepacian is the main EPS produced by B. cenocepacia H111 [29, 45–47] and the expression of these two clusters has previously been shown to be induced under nitrogen limited conditions and controlled by σ54 [24]. Indeed, EPS production was reduced in the ntrC mutant to a similar extent as that of the σ54 mutant. EPS production was restored to the level of the wild-type in the complemented ntrC mutant (Fig 3). Biofilm formation, which was previously shown to be dependent on the alternative sigma factor (σ54) [24], was only slightly affected in the ntrC mutant. However, the complemented strain produced significantly more biofilm than the wild-type strain (Fig 4).
Fig 3. NtrC–dependent EPS production.

EPS production in the wild type (1), the ntrC mutant (2) and the complemented strain (3) was tested on YEM plates. Three independent biological replicates were tested; the result of one is shown here.
Fig 4. Biofilm formation is only slightly dependent on NtrC.

Biofilm production was assessed for the wild type, ntrC mutant and complemented strain in 96-well plates. The columns represent the mean biofilm index generated from independent biological triplicate cultures of each strain. The error bars indicate the standard deviation. The increased biofilm formation in the complemented mutant compared to the wild type was statistically significant (*** p<0.0001). However, the slight reduction in the mutant was not statistically significant (ns).
NtrC regulates swarming and swimming motility
The positive effect of NtrC on the transcription of genes encoding the flagellar motor (motAB) and flagellar biosynthesis proteins, prompted us to test motility of the strains. We first compared the swarming ability of the wild type strain with those of the ntrC mutant and the complemented strain (Fig 5A). The ntrC mutant was able to swarm, but to a lesser extent than wild type H111. Complementation increased swarming to above the level of the wild type. Swimming motility was also compromised in the ntrC mutant, but this defect was only partly restored in the complemented strain (Fig 5B). The results clearly show that NtrC plays an important role in the control of motility. The stronger effect on swarming is in line with the decreased transcription of rhlA (coding for a rhamnolipid) and motility-controlling flagellar-associated genes, such as flgN and flgL, in the ntrC mutant (Table 2).
Fig 5. NtrC–dependent motility.
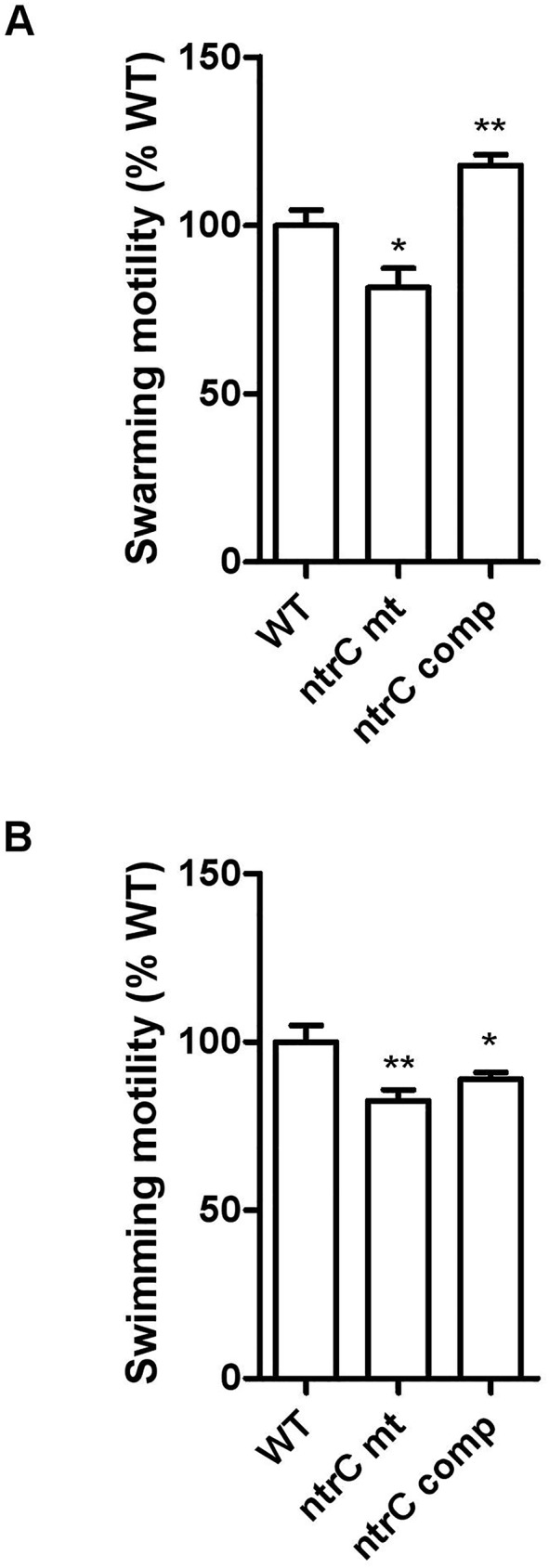
The histograms show the swarming (A) and swimming (B) motility of the ntrC mutant and the complemented mutant relative to the wild-type strain H111. Both assays were performed in triplicate. Significance was calculated by comparing the mutant or the complemented mutant with the wild type (* p<0.05 and ** p<0.01).
Virulence of B. cenocepacia H111 in the C. elegans infection model is not regulated by NtrC
Since σ54 was previously shown to be important for virulence in the C. elegans model, we set out to elucidate if this phenotype is also dependent on NtrC. For this, we fed nematode larvae with the wild type, the ntrC mutant and the complemented mutant. As a positive control, the σ54 mutant was included. While the σ54 mutant showed reduced virulence [24], worms infected with the ntrC mutant developed only to the L1-L2 larval stage, similarly to worms fed with the wild-type strain. This result is in line with our RNA-Seq data, which showed that the transcription of the nematocidal gene aidA (I35_7308), which was shown to be dependent on σ54 [24], was not controlled by NtrC (S3 Fig). The role of NtrC in the pathogenicity of H111 was also assessed in a different infection model, the larvae of the greater wax moth Galleria mellonella. In this model, we again did not observe significant attenuation of pathogenicity of the ntrC mutant compared with the wild type (data not shown).
Discussion
We previously identified the B. cenocepacia genes responding to nitrogen starvation conditions and found that the alternative sigma factor σ54 (or RpoN) plays a major role in the control of nitrogen metabolism. That study furthermore uncovered that σ54 controls other important cellular processes such as EPS production, biofilm formation and C. elegans virulence [24]. Sigma 54 is an alternative sigma factor since it binds to a characteristic -24/-12 binding sequence (GGcacg-N4-ttGC) in the promoter of target genes and requires an additional ATP-dependent activation event to initiate transcription. This step is provided by transcriptional activators with an AAA (ATPase Associated with various cellular Activities) protein domain, which bind as inactive dimers to a consensus sequence upstream of the promoter, assemble as hexameric rings (in their active form) and interact through DNA looping with the σ54 promoter complex to activate transcription [20]. This requirement for an activator protein (or EBP) allows σ54 to tightly and rapidly control gene expression in response to cellular and extracellular signals that regulate the activity of a specific AAA-domain containing protein. Bacteria usually encode several AAA-family activator proteins and each one is needed for a specific and precise response to an environmental change. By looking for all B. cenocepacia proteins containing a σ54 activation AAA domain (PFAM family: PF00158), we were able to identify 22 proteins that could potentially serve as σ54 activator proteins (S4 Table). Among them we identified the regulatory protein NtrC, which is known to be the master regulator of nitrogen control in other bacteria. The ntrC gene had previously been shown to have increased expression under nitrogen limiting conditions [24]. The ntrC gene is usually located downstream of ntrB, which encodes a sensor kinase that phosphorylates the response regulator NtrC in nitrogen limited environments. In this study, we first performed a comprehensive growth analysis of an ntrC mutant on 95 different nitrogen sources including all common amino acids, the nitrogenous bases, several di-peptides and other compounds (Biolog Phenotype MicroArray). The ntrC mutant was revealed to be affected in the utilisation of 7 N sources i. e. nitrate, urea, L-citrulline, acetamide, DL-lactamide, allantoin and parabanic acid suggesting that utilisation of these sources in B. cenocepacia is dependent on the presence of a functional NtrC. We next used RNA-Seq analysis to elucidate the role of NtrC in the control of transcription during nitrogen starvation conditions. The results clearly show that NtrC is a major regulator under nitrogen limiting conditions and that the large majority of the NtrC-regulated genes are co-regulated by σ54. At the phenotypic level, the ntrC mutant strain behaved like a σ54 mutant strain for the utilization of alternative nitrogen sources such as nitrate and urea. Both urea and nitrate were not utilised as nitrogen sources by the mutant strain (Fig 1 and S2 Table). Accordingly, the transcription of genes involved in urea and nitrate assimilation (urease and nitrate/nitrite reductase, respectively) was significantly down-regulated in the ntrC mutant strain (Table 1). In addition to, and in line with, a poor utilization of allantoin in the ntrC mutant (S2 Fig and S2 Table), two genes coding for allantoicases (I35_0495 and I35_1962, Table 2 and S3 Table) showed significantly decreased expression in the ntrC mutant. Inspection of the NtrC regulon revealed two clusters of genes (bceI and bceII) involved in the production of cepacian, the main EPS in B. cenocepacia [29, 45–47]. Accordingly, EPS production was clearly reduced in an ntrC mutant and this defect could be rescued by genetic complementation. Control of EPS synthesis by NtrC has been demonstrated in several bacteria including the human pathogen Vibrio vulnificus [48], Agrobacterium sp. ATCC 31749 [49] and Sinorhizobium meliloti [50]. However, and in contrast to our previous results obtained with the σ54 mutant [24], biofilm formation was only slightly reduced in the ntrC mutant, suggesting that σ54 is using another EBP for controlling biofilm formation. This result also suggested that the reduced transcription of the bceI and bceII clusters does not drastically affect biofilm formation in microtiter plates.
We show here that the ntrC mutant is affected in motility and for the first time that the ability of B. cenocepacia H111 to swarm is controlled by NtrC and by σ54 (Fig 5). Many factors have been shown to regulate swarming [51, 52]. For example, in Serratia liquefaciens and in other bacteria, the flhD master operon needed for flagellar biosynthesis is essential for swarming motility [41, 53]. In B. cenocepacia H111 this social motility is under the control of the CepRI and RpfRF quorum sensing systems [40, 54] and in B. glumae it has recently been shown that quorum sensing controls swarming through the regulation of rhamnolipid biosynthesis under nutrient-limited conditions [55, 56]. In Pseudomonas aeruginosa swarming has been shown to be controlled at the post-transcriptional level by the RsmAB system and by c-di-GMP levels [57–59]. While transcription of cepRI and rpfRF was not altered in a ntrC mutant, we noted in our RNA-Seq data that NtrC positively regulates transcription of several genes which could be involved in the control of swarming motility: i) motA and motB, encoding the flagellar motor as well as several genes involved in flagellar biosynthesis; ii) genes involved in regulation of c-di-GMP levels by coding for proteins with a diguanylate cyclase phosphodiesterase (EAL) domain and iii) the gene rhlA, which encodes a 3-(3-hydroxyalkanoyloxy) alkanoic acid synthase, probably involved in rhamnolipid biosynthesis. Additional work will be required to test these possibilities.
The RNA-Seq approach allowed us to verify genes expected to be under NtrC control [60, 61] and to identify new potential NtrC targets such as the cepacian clusters and genes involved in motility (motA, motB, flgL, flgK, flgN, rhlA) (Table 1). Moreover, transcription of the holin and anti-holin gene pair cidA and cidB was significantly down-regulated in a ntrC mutant suggesting that this system, which is known to control peptidoglycan hydrolase activity and penicillin tolerance in Staphylococccus aureus [62] and has recently been proposed to be a key player in the regulation of the stress response [63], is under NtrC control in B. cenocepacia H111. However, our ntrC mutant was neither affected in antibiotic resistance nor in resistance to oxidative stress (data not shown).
Several genes coding for proteins involved in the TCA cycle were found among the genes showing increased transcription in the absence of NtrC under nitrogen limiting conditions, strengthening the proposal made by Hervas and co-workers that in Pseudomonas putida, NtrC represses carbon metabolism [61]. The fact that NtrC plays an important role in maintaining the balance between nitrogen and carbon metabolism under nitrogen limiting conditions may explain why constantly higher ntrC expression in the complemented mutant (driven by the promoter of the expression vector pBBR1MCS-5) leads to a growth defect in this strain.
This work demonstrates that NtrC is an activator of σ54-dependent gene transcription, that controls not only nitrogen metabolism but also various other functions, including EPS production and motility. However, as yet unidentified σ54 activators are required to control other phenotypes such as biofilm production and virulence to C. elegans. Further studies will be required to identify these activators.
Supporting information
Wild-type and ntrC mutant strains were grown in AB minimal medium from a starting OD600 of 0.05. Optical density was monitored over about 20 hours. The dotted line shows OD600 = 0.5, after which point the samples were subjected to nitrogen starvation and then harvested for RNA-Seq. The experiment was done in triplicate. Error bars indicate standard deviation.
(DOCX)
Percentages of induced and repressed genes are given for the comparison of ntrC mutant vs. wild-type cells grown under nitrogen limiting conditions. Percentages were calculated by dividing the number of significantly induced or repressed genes (Table 1) in each category by the total number of retained genes in the corresponding category. Asterisks (*) indicate statistical significance for overexpressed genes in a particular category (p-value < 0.01). C, energy production and conversion; E, amino acid transport and metabolism; F nucleotide transport and metabolism; G carbohydrate transport and metabolism; H coenzyme transport and metabolism; I lipid transport and metabolism; J translation, ribosomal structure and biogenesis; K transcription; L replication, recombination and repair; M cell wall/membrane/ envelope biogenesis; N cell motility; O post-translational modification, protein turnover and chaperon; P inorganic ion transport and metabolism; Q secondary metabolites biosynthesis, transport and catabolism; R general function prediction only; S function unknown; T signal transduction mechanisms; U intracellular trafficking, secretion and vesicular transport; V defense mechanisms.
(DOCX)
Pathogenicity assay of bacterial strains to C. elegans N2 strain was carried out as described in the material and methods. The number of L1 larvae in each well of a 96-well plate was counted after seeding with the bacterial strains to be tested. After 48 hours co-incubation at 20°C, the developmental stages of the worms were evaluated and the numbers were counted. Error bars represent standard deviation of the means (n = 3).
(DOCX)
(DOCX)
(XLSX)
(XLSX)
(XLSX)
Acknowledgments
We are thankful to Kirsty Agnoli for proofreading the manuscript. Claudio Aguilar and Elisabeth Steiner are acknowledged for initial help in the phenotypic characterization of the ntrC mutant strain. We are grateful to Catherine Aquino Fournier and Hubert Rehrauer from the Functional Genomics Center Zürich (FGCZ) for support in RNA-Seq experiments.
Data Availability
The RNA-Seq raw data files are accessible through the GEO Series accession number GSE95607.
Funding Statement
This work was supported by the Swiss National Science Foundation (31003A-169307/1 to LE and 31003A_153374 to GP).
References
- 1.Coenye T, Vandamme P. Diversity and significance of Burkholderia species occupying diverse ecological niches. Environ Microbiol. 2003;5(9):719–29. Epub 2003/08/16. . [DOI] [PubMed] [Google Scholar]
- 2.Mira NP, Madeira A, Moreira AS, Coutinho CP, Sá-Correia I. Genomic expression analysis reveals strategies of Burkholderia cenocepacia to adapt to cystic fibrosis patients' airways and antimicrobial therapy. PLoS One. 2011;6(12):e28831 Epub 2012/01/05. doi: 10.1371/journal.pone.0028831 ; PubMed Central PMCID: PMC3244429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Reitzer L. Nitrogen assimilation and global regulation in Escherichia coli. Annu Rev Microbiol. 2003;57:155–76. Epub 2003/05/06. doi: 10.1146/annurev.micro.57.030502.090820. . [DOI] [PubMed] [Google Scholar]
- 4.Huergo LF, Chandra G, Merrick M. P(II) signal transduction proteins: nitrogen regulation and beyond. FEMS Microbiol Rev. 2013;37(2):251–83. Epub 2012/08/07. doi: 10.1111/j.1574-6976.2012.00351.x. . [DOI] [PubMed] [Google Scholar]
- 5.Van Heeswijk WC, Westerhoff HV, Boogerd FC. Nitrogen assimilation in Escherichia coli: putting molecular data into a systems perspective. Microbiol Mol Biol Rev. 2013;77(4):628–95. Epub 2013/12/04. doi: 10.1128/MMBR.00025-13. ; PubMed Central PMCID: PMC3973380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Merrick MJ, Edwards RA. Nitrogen control in bacteria. Microbiol Rev. 1995;59(4):604–22. Epub 1995/12/01. ; PubMed Central PMCID: PMC239390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Hirschman J, Wong PK, Sei K, Keener J, Kustu S. Products of nitrogen regulatory genes ntrA and ntrC of enteric bacteria activate glnA transcription in vitro: evidence that the ntrA product is a sigma factor. Proc Natl Acad Sci U S A. 1985;82(22):7525–9. Epub 1985/11/01. ; PubMed Central PMCID: PMC390849. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Ninfa AJ, Atkinson MR. PII signal transduction proteins. Trends Microbiol. 2000;8(4):172–9. Epub 2001/02/07. . [DOI] [PubMed] [Google Scholar]
- 9.Pioszak AA, Jiang P, Ninfa AJ. The Escherichia coli PII signal transduction protein regulates the activities of the two-component system transmitter protein NRII by direct interaction with the kinase domain of the transmitter module. Biochemistry. 2000;39(44):13450–61. Epub 2000/11/07. . [DOI] [PubMed] [Google Scholar]
- 10.Wyman C, Rombel I, North AK, Bustamante C, Kustu S. Unusual oligomerization required for activity of NtrC, a bacterial enhancer-binding protein. Science. 1997;275(5306):1658–61. Epub 1997/03/14. . [DOI] [PubMed] [Google Scholar]
- 11.Wigneshweraraj S, Bose D, Burrows PC, Joly N, Schumacher J, Rappas M, et al. Modus operandi of the bacterial RNA polymerase containing the s54 promoter-specificity factor. Mol Microbiol. 2008;68(3):538–46. Epub 2008/03/12. doi: 10.1111/j.1365-2958.2008.06181.x. . [DOI] [PubMed] [Google Scholar]
- 12.Yang Y, Darbari VC, Zhang N, Lu D, Glyde R, Wang YP, et al. Structures of the RNA polymerase-σ54 reveal new and conserved regulatory strategies. Science. 2015;349(6250):882–5. Epub 2015/08/22. doi: 10.1126/science.aab1478. ; PubMed Central PMCID: PMC4681505. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Morett E, Buck M. In vivo studies on the interaction of RNA polymerase-σ54 with the Klebsiella pneumoniae and Rhizobium meliloti nifH promoters. The role of NifA in the formation of an open promoter complex. J Mol Biol. 1989;210(1):65–77. Epub 1989/11/05. . [DOI] [PubMed] [Google Scholar]
- 14.Francke C, Groot Kormelink T, Hagemeijer Y, Overmars L, Sluijter V, Moezelaar R, et al. Comparative analyses imply that the enigmatic sigma factor 54 is a central controller of the bacterial exterior. BMC Genomics. 2011;12:385 Epub 2011/08/03. doi: 10.1186/1471-2164-12-385. ; PubMed Central PMCID: PMC3162934. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bush M, Dixon R. The role of bacterial enhancer binding proteins as specialized activators of σ54-dependent transcription. Microbiol Mol Biol Rev. 2012;76(3):497–529. Epub 2012/08/31. doi: 10.1128/MMBR.00006-12. ; PubMed Central PMCID: PMC3429621. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Studholme DJ, Dixon R. Domain architectures of σ54-dependent transcriptional activators. J Bacteriol. 2003;185(6):1757–67. Epub 2003/03/06. ; PubMed Central PMCID: PMC150144. doi: 10.1128/JB.185.6.1757-1767.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Buck M, Gallegos MT, Studholme DJ, Guo Y, Gralla JD. The bacterial enhancer-dependent σ54 (σN) transcription factor. J Bacteriol. 2000;182(15):4129–36. Epub 2000/07/14. ; PubMed Central PMCID: PMC101881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Lee SY, De La Torre A, Yan D, Kustu S, Nixon BT, Wemmer DE. Regulation of the transcriptional activator NtrC1: structural studies of the regulatory and AAA+ ATPase domains. Genes Dev. 2003;17(20):2552–63. Epub 2003/10/17. doi: 10.1101/gad.1125603. ; PubMed Central PMCID: PMC218149. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Vale RD. AAA proteins. Lords of the ring. J Cell Biol. 2000;150(1):F13–9. Epub 2000/07/13. ; PubMed Central PMCID: PMC2185557. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Morris L, Cannon W, Claverie-Martin F, Austin S, Buck M. DNA distortion and nucleation of local DNA unwinding within σ54 (σN) holoenzyme closed promoter complexes. J Biol Chem. 1994;269(15):11563–71. Epub 1994/04/15. . [PubMed] [Google Scholar]
- 21.Ferro-Luzzi Ames G, Nikaido K. Nitrogen regulation in Salmonella typhimurium. Identification of an NtrC protein-binding site and definition of a consensus binding sequence. EMBO J. 1985;4(2):539–47. Epub 1985/02/01. ; PubMed Central PMCID: PMC554219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Reitzer LJ, Magasanik B. Transcription of glnA in E. coli is stimulated by activator bound to sites far from the promoter. Cell. 1986;45(6):785–92. Epub 1986/06/20. . [DOI] [PubMed] [Google Scholar]
- 23.Saldías MS, Lamothe J, Wu R, Valvano MA. Burkholderia cenocepacia requires the RpoN sigma factor for biofilm formation and intracellular trafficking within macrophages. Infect Immun. 2008;76(3):1059–67. Epub 2008/01/16. doi: 10.1128/IAI.01167-07. ; PubMed Central PMCID: PMC2258854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Lardi M, Aguilar C, Pedrioli A, Omasits U, Suppiger A, Carcamo-Oyarce G, et al. σ54-dependent response to nitrogen limitation and virulence in Burkholderia cenocepacia strain H111. Appl Environ Microbiol. 2015;81(12):4077–89. Epub 2015/04/05. doi: 10.1128/AEM.00694-15. ; PubMed Central PMCID: PMC4524130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Tomich M, Herfst CA, Golden JW, Mohr CD. Role of flagella in host cell invasion by Burkholderia cepacia. Infect Immun. 2002;70(4):1799–806. Epub 2002/03/16. ; PubMed Central PMCID: PMC127839. doi: 10.1128/IAI.70.4.1799-1806.2002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Urban TA, Griffith A, Torok AM, Smolkin ME, Burns JL, Goldberg JB. Contribution of Burkholderia cenocepacia flagella to infectivity and inflammation. Infect Immun. 2004;72(9):5126–34. Epub 2004/08/24. doi: 10.1128/IAI.72.9.5126-5134.2004. ; PubMed Central PMCID: PMC517433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Cunha MV, Sousa SA, Leitão JH, Moreira LM, Videira PA, Sá-Correia I. Studies on the involvement of the exopolysaccharide produced by cystic fibrosis-associated isolates of the Burkholderia cepacia complex in biofilm formation and in persistence of respiratory infections. J Clin Microbiol. 2004;42(7):3052–8. Epub 2004/07/10. doi: 10.1128/JCM.42.7.3052-3058.2004. ; PubMed Central PMCID: PMC446245. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Bylund J, Burgess LA, Cescutti P, Ernst RK, Speert DP. Exopolysaccharides from Burkholderia cenocepacia inhibit neutrophil chemotaxis and scavenge reactive oxygen species. J Biol Chem. 2006;281(5):2526–32. Epub 2005/12/01. doi: 10.1074/jbc.M510692200. . [DOI] [PubMed] [Google Scholar]
- 29.Ferreira AS, Silva IN, Oliveira VH, Cunha R, Moreira LM. Insights into the role of extracellular polysaccharides in Burkholderia adaptation to different environments. Front Cell Infect Microbiol. 2011;1:16 Epub 2011/01/01. doi: 10.3389/fcimb.2011.00016. ; PubMed Central PMCID: PMC3417362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Cuzzi B, Cescutti P, Furlanis L, Lagatolla C, Sturiale L, Garozzo D, et al. Investigation of bacterial resistance to the immune system response: cepacian depolymerisation by reactive oxygen species. Innate Immun. 2012;18(4):661–71. Epub 2012/01/27. doi: 10.1177/1753425911435954. . [DOI] [PubMed] [Google Scholar]
- 31.Miller J. Experiments in molecular genetics Press CSHL, editor. Cold Spring Harbor Laboratory Press; N. Y. USA1972. [Google Scholar]
- 32.Clark DJ, Maaloe O. DNA replication and division cycle in Escherichia coli. J Mol Biol. 1967;23(1):99–112. doi: 10.1016/S0022-2836(67)80070-6. . [Google Scholar]
- 33.Shastri S, Spiewak HL, Sofoluwe A, Eidsvaag VA, Asghar AH, Pereira T, et al. An efficient system for the generation of marked genetic mutants in members of the genus Burkholderia. Plasmid. 2017;89:49–56. Epub 2016/11/09. doi: 10.1016/j.plasmid.2016.11.002. ; PubMed Central PMCID: PMC5312678. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Pessi G, Ahrens CH, Rehrauer H, Lindemann A, Hauser F, Fischer HM, et al. Genome-wide transcript analysis of Bradyrhizobium japonicum bacteroids in soybean root nodules. Mol Plant Microbe Interact. 2007;20(11):1353–63. Epub 2007/11/06. doi: 10.1094/MPMI-20-11-1353. . [DOI] [PubMed] [Google Scholar]
- 35.Carlier A, Agnoli K, Pessi G, Suppiger A, Jenul C, Schmid N, et al. Genome sequence of Burkholderia cenocepacia H111, a cystic fibrosis airway isolate. Genome Announc. 2014;10(2):pii: e00298-14. Epub 2014/04/12. doi: 10.1128/genomeA.00298-14. ; PubMed Central PMCID: PMC3983312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Anders S, Huber W. Differential expression analysis for sequence count data. Genome Biol. 2010;11(10):R106 Epub 2010/10/29. doi: 10.1186/gb-2010-11-10-r106. ; PubMed Central PMCID: PMC3218662. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Powell S, Forslund K, Szklarczyk D, Trachana K, Roth A, Huerta-Cepas J, et al. eggNOG v4.0: nested orthology inference across 3686 organisms. Nucleic Acids Res. 2014;42(Database issue):D231–9. Epub 2013/12/04. doi: 10.1093/nar/gkt1253. ; PubMed Central PMCID: PMC3964997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Agnoli K, Schwager S, Uehlinger S, Vergunst A, Viteri DF, Nguyen DT, et al. Exposing the third chromosome of Burkholderia cepacia complex strains as a virulence plasmid. Mol Microbiol. 2012;83(2):362–78. Epub 2011/12/17. doi: 10.1111/j.1365-2958.2011.07937.x. . [DOI] [PubMed] [Google Scholar]
- 39.Richau JA, Leitão JH, Correia M, Lito L, Salgado MJ, Barreto C, et al. Molecular typing and exopolysaccharide biosynthesis of Burkholderia cepacia isolates from a Portuguese cystic fibrosis center. J Clin Microbiol. 2000;38(4):1651–5. Epub 2000/04/04. ; PubMed Central PMCID: PMC86514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Huber B, Riedel K, Hentzer M, Heydorn A, Götschlich A, Givskov M, et al. The cep quorum-sensing system of Burkholderia cepacia H111 controls biofilm formation and swarming motility. Microbiology. 2001;147(Pt 9):2517–28. Epub 2001/09/06. doi: 10.1099/00221287-147-9-2517 [DOI] [PubMed] [Google Scholar]
- 41.Eberl L, Christiansen G, Molin S, Givskov M. Differentiation of Serratia liquefaciens into swarm cells is controlled by the expression of the flhD master operon. J Bacteriol. 1996;178(2):554–9. Epub 1996/01/01. ; PubMed Central PMCID: PMC177693. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Pessi G, Braunwalder R, Grunau A, Omasits U, Ahrens CH, Eberl L. Response of Burkholderia cenocepacia H111 to micro-oxia. PLoS One. 2013;8(9):e72939 Epub 2013/09/12. doi: 10.1371/journal.pone.0072939. ; PubMed Central PMCID: PMC3759415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Pfaffl MW. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001;29(9):e45 Epub 2001/05/09. ; PubMed Central PMCID: PMC55695. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Rombel I, North A, Hwang I, Wyman C, Kustu S. The bacterial enhancer-binding protein NtrC as a molecular machine. Cold Spring Harb Sym. 1998;63:157–66. doi: 10.1101/sqb.1998.63.157 [DOI] [PubMed] [Google Scholar]
- 45.Moreira LM, Videira PA, Sousa SA, Leitão JH, Cunha MV, Sá-Correia I. Identification and physical organization of the gene cluster involved in the biosynthesis of Burkholderia cepacia complex exopolysaccharide. Biochem Biophys Res Commun. 2003;312(2):323–33. Epub 2003/11/26. . [DOI] [PubMed] [Google Scholar]
- 46.Ferreira AS, Leitão JH, Sousa SA, Cosme AM, Sá-Correia I, Moreira LM. Functional analysis of Burkholderia cepacia genes bceD and bceF, encoding a phosphotyrosine phosphatase and a tyrosine autokinase, respectively: role in exopolysaccharide biosynthesis and biofilm formation. Appl Environ Microbiol. 2007;73(2):524–34. Epub 2006/11/23. doi: 10.1128/AEM.01450-06 ; PubMed Central PMCID: PMC1796985. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Ferreira AS, Leitão JH, Silva IN, Pinheiro PF, Sousa SA, Ramos CG, et al. Distribution of cepacian biosynthesis genes among environmental and clinical Burkholderia strains and role of cepacian exopolysaccharide in resistance to stress conditions. Appl Environ Microbiol. 2010;76(2):441–50. Epub 2009/12/02. doi: 10.1128/AEM.01828-09. ; PubMed Central PMCID: PMC2805225. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Kim HS, Park SJ, Lee KH. Role of NtrC-regulated exopolysaccharides in the biofilm formation and pathogenic interaction of Vibrio vulnificus. Mol Microbiol. 2009;74(2):436–53. Epub 2009/09/10. doi: 10.1111/j.1365-2958.2009.06875.x. . [DOI] [PubMed] [Google Scholar]
- 49.Ruffing AM, Chen RR. Transcriptome profiling of a curdlan-producing Agrobacterium reveals conserved regulatory mechanisms of exopolysaccharide biosynthesis. Microb Cell Fact. 2012;11:17 Epub 2012/02/07. doi: 10.1186/1475-2859-11-17. ; PubMed Central PMCID: PMC3293034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Wang D, Xue H, Wang Y, Yin R, Xie F, Luo L. The Sinorhizobium meliloti ntrX gene is involved in succinoglycan production, motility, and symbiotic nodulation on alfalfa. Appl Environ Microbiol. 2013;79(23):7150–9. Epub 2013/09/17. doi: 10.1128/AEM.02225-13. ; PubMed Central PMCID: PMC3837732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Verstraeten N, Braeken K, Debkumari B, Fauvart M, Fransaer J, Vermant J, et al. Living on a surface: swarming and biofilm formation. Trends Microbiol. 2008;16(10):496–506. Epub 2008/09/09. doi: 10.1016/j.tim.2008.07.004 . [DOI] [PubMed] [Google Scholar]
- 52.Daniels R, Vanderleyden J, Michiels J. Quorum sensing and swarming migration in bacteria. FEMS Microbiol Rev. 2004;28(3):261–89. Epub 2004/09/29. . [DOI] [PubMed] [Google Scholar]
- 53.Patrick JE, Kearns DB. Swarming motility and the control of master regulators of flagellar biosynthesis. Mol Microbiol. 2012;83(1):14–23. Epub 2011/11/19. doi: 10.1111/j.1365-2958.2011.07917.x. ; PubMed Central PMCID: PMC3245337. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Deng Y, Schmid N, Wang C, Wang J, Pessi G, Wu D, et al. Cis-2-dodecenoic acid receptor RpfR links quorum-sensing signal perception with regulation of virulence through cyclic dimeric guanosine monophosphate turnover. Proc Natl Acad Sci U S A. 2012;109(38):15479–84. Epub 2012/09/06. doi: 10.1073/pnas.1205037109. ; PubMed Central PMCID: PMC3458384. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Nickzad A, Lepine F, Deziel E. Quorum Sensing controls swarming motility of Burkholderia glumae through regulation of rhamnolipids. PLoS One. 2015;10(6):e0128509 Epub 2015/06/06. doi: 10.1371/journal.pone.0128509. ; PubMed Central PMCID: PMC4457897. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Nickzad A, Deziel E. Adaptive significance of quorum sensing-dependent regulation of rhamnolipids by integration of growth rate in Burkholderia glumae: a trade-off between survival and efficiency. Front Microbiol. 2016;7:1215 Epub 2016/08/20. doi: 10.3389/fmicb.2016.01215. ; PubMed Central PMCID: PMC4972832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Heurlier K, Williams F, Heeb S, Dormond C, Pessi G, Singer D, et al. Positive control of swarming, rhamnolipid synthesis, and lipase production by the posttranscriptional RsmA/RsmZ system in Pseudomonas aeruginosa PAO1. J Bacteriol. 2004;186(10):2936–45. Epub 2004/05/06. ; PubMed Central PMCID: PMC400603. doi: 10.1128/JB.186.10.2936-2945.2004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Baker AE, Diepold A, Kuchma SL, Scott JE, Ha DG, Orazi G, et al. PilZ domain protein FlgZ mediates cyclic di-GMP-dependent swarming motility control in Pseudomonas aeruginosa. J Bacteriol. 2016;198(13):1837–46. Epub 2016/04/27. doi: 10.1128/JB.00196-16. ; PubMed Central PMCID: PMC4907108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Kuchma SL, Delalez NJ, Filkins LM, Snavely EA, Armitage JP, O'Toole GA. Cyclic di-GMP-mediated repression of swarming motility by Pseudomonas aeruginosa PA14 requires the MotAB stator. J Bacteriol. 2015;197(3):420–30. Epub 2014/10/29. doi: 10.1128/JB.02130-14. ; PubMed Central PMCID: PMC4285984. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Zimmer DP, Soupene E, Lee HL, Wendisch VF, Khodursky AB, Peter BJ, et al. Nitrogen regulatory protein C-controlled genes of Escherichia coli: scavenging as a defense against nitrogen limitation. Proc Natl Acad Sci U S A. 2000;97(26):14674–9. Epub 2000/12/20. doi: 10.1073/pnas.97.26.14674. ; PubMed Central PMCID: PMC18977. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Hervas AB, Canosa I, Santero E. Transcriptome analysis of Pseudomonas putida in response to nitrogen availability. J Bacteriol. 2008;190(1):416–20. Epub 2007/10/30. doi: 10.1128/JB.01230-07 ; PubMed Central PMCID: PMC2223721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Rice KC, Firek BA, Nelson JB, Yang SJ, Patton TG, Bayles KW. The Staphylococcus aureus cidAB operon: Evaluation of its role in regulation of murein hydrolase activity and penicillin tolerance. Journal of Bacteriology. 2003;185(8):2635–43. doi: 10.1128/JB.185.8.2635-2643.2003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ahn SJ, Rice KC. Understanding the Streptococcus mutans Cid/Lrg System through CidB Function. Appl Environ Microb. 2016;82(20):6189–203. doi: 10.1128/Aem.01499-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Wild-type and ntrC mutant strains were grown in AB minimal medium from a starting OD600 of 0.05. Optical density was monitored over about 20 hours. The dotted line shows OD600 = 0.5, after which point the samples were subjected to nitrogen starvation and then harvested for RNA-Seq. The experiment was done in triplicate. Error bars indicate standard deviation.
(DOCX)
Percentages of induced and repressed genes are given for the comparison of ntrC mutant vs. wild-type cells grown under nitrogen limiting conditions. Percentages were calculated by dividing the number of significantly induced or repressed genes (Table 1) in each category by the total number of retained genes in the corresponding category. Asterisks (*) indicate statistical significance for overexpressed genes in a particular category (p-value < 0.01). C, energy production and conversion; E, amino acid transport and metabolism; F nucleotide transport and metabolism; G carbohydrate transport and metabolism; H coenzyme transport and metabolism; I lipid transport and metabolism; J translation, ribosomal structure and biogenesis; K transcription; L replication, recombination and repair; M cell wall/membrane/ envelope biogenesis; N cell motility; O post-translational modification, protein turnover and chaperon; P inorganic ion transport and metabolism; Q secondary metabolites biosynthesis, transport and catabolism; R general function prediction only; S function unknown; T signal transduction mechanisms; U intracellular trafficking, secretion and vesicular transport; V defense mechanisms.
(DOCX)
Pathogenicity assay of bacterial strains to C. elegans N2 strain was carried out as described in the material and methods. The number of L1 larvae in each well of a 96-well plate was counted after seeding with the bacterial strains to be tested. After 48 hours co-incubation at 20°C, the developmental stages of the worms were evaluated and the numbers were counted. Error bars represent standard deviation of the means (n = 3).
(DOCX)
(DOCX)
(XLSX)
(XLSX)
(XLSX)
Data Availability Statement
The RNA-Seq raw data files are accessible through the GEO Series accession number GSE95607.