

Abstract
Neural elements of the intrinsic cardiac nervous system transduce sensory inputs from the heart, blood vessels and other organs to ensure adequate cardiac function on a beat-to-beat basis. This inter-organ crosstalk is critical for normal function of the heart and other organs; derangements within the nervous system hierarchy contribute to pathogenesis of organ dysfunction. The role of intact cardiac nerves in development of, as well as protection against, ischemic injury is of current interest since it may involve recruitment of intrinsic cardiac ganglia. For instance, ischemic conditioning, a novel protection strategy against organ injury, and in particular remote conditioning, is likely mediated by activation of neural pathways or by endogenous cytoprotective blood-borne substances that stimulate different signalling pathways. This discovery reinforces the concept that inter-organ communication, and maintenance thereof, is key. As such, greater understanding of mechanisms and elucidation of treatment strategies is imperative to improve clinical outcomes particularly in patients with comorbidities. For instance, autonomic imbalance between sympathetic and parasympathetic nervous system regulation can initiate cardiovascular autonomic neuropathy that compromises cardiac stability and function. Neuromodulation therapies that directly target the intrinsic cardiac nervous system or other elements of the nervous system hierarchy are currently being investigated for treatment of different maladies in animal and human studies.
Keywords: Intrinsic cardiac nervous system, Myocardial ischemia, Ischemic conditioning, Autonomic neuropathy, Coronary blood flow regulation
Core tip: Neural elements within the intrinsic cardiac nervous system are known to transduce sensory inputs from the heart, blood vessels and surrounding organs to ensure beat-to-beat regulation of cardiac function. Development of autonomic neurophathies in patients with comorbidities compromises clinical outcomes. Myocardial ischemia also significantly affects cardiocytes as well as cardiac neurons; post-ischemic remodelling might affect neuronal function and thereby contribute to cardiac instability. Different protection strategies including ischemic conditioning and neuromodulation interventions that limit neural injury and help maintain cardiovascular function are the subject of ongoing investigations.
INTRODUCTION
A dense network of parasympathetic, sympathetic and sensory neurons innervates the heart and cardiac conduction system; each population of neurons is distinct with respect to functional requirements of the heart. Increased attention is being focused on the complex anatomy and function of the cardiac neuroaxis and questions abound regarding the manner in which different neuronal populations communicate with each other and between different organ systems. Ardell et al[1] recently made the case that the cardiac neural hierarchy functions as a distributive processor with multiple nested feedback control loops that involve peripheral and central aspects of the autonomic nervous system. Remodeling of the cardiac nervous system at morphological and phenotypic levels during disease development is also under scrutiny[2-5]; neural remodeling can cause electrical instability that increases the incidence of arrhythmogenesis. Neuromodulation-based treatments for cardiovascular disease are being investigated as evidenced by the increasing use of diverse cardiac sympathetic decentralization and bioelectric interventions[6]. Herein, we briefly discuss experimental and clinical findings that highlight a role for the intrinsic cardiac nervous system on cardiodynamics. We also discuss mechanisms relevant to diverse protection stratagems. Finally, we focus on autonomic neurophathies that accompany comorbidities (Figure 1). For this review, clinical and basic science reports were searched using MEDLINE, PubMed and Google Scholar with the keywords intrinsic cardiac nervous system, myocardial ischemia-reperfusion injury, heart and kidney disease, cardioprotection, preconditioning and combinations thereof. Findings from our own studies on this, and related subjects were also consulted.
Figure 1.
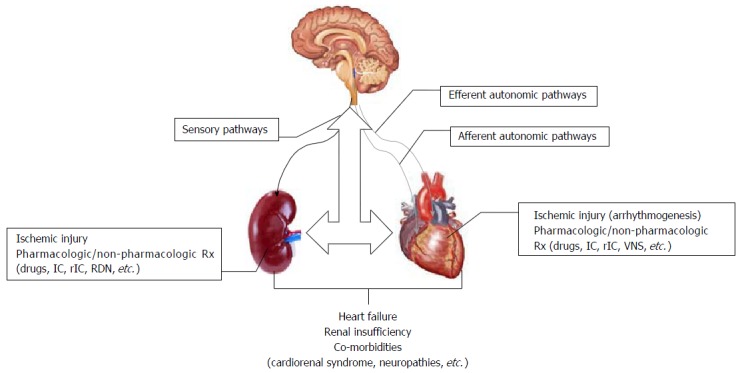
A schematic overview of efferent and afferent autonomic pathways on normal cardiac regulation, they also play a role in arrhythmogenesis caused by ischemic injury. Various pharmacologic/non-pharmacologic interventions that target autonomic pathways (IC: Ischemic conditioning; rIC: Remote IC; VNS: Vagus nerve stimulation) attenuate cardiac or renal symptoms. Sensory pathways are involved in renal regulation; injury (all cause) affects renal function that can be attenuated by different interventions (IC, rIC, RDN: Renal denervation). Inter-organ interactions also directly affect organ function; development of comorbidities is related to pathogenesis of disease in multiple organs (ex. heart-kidneys-brain, etc.). Pathology in one organ system can result in significant progression of disease in a distant organ; neuromodulation interventions may be beneficial to these patients.
Developmental aspects
Development of the nervous and cardiovascular systems is synchronized during embryogenesis; neural crest cells in the dorsal neural tube form the parasympathetic and sympathetic nervous systems that are important for cardiovascular function. Sympathetic interactions play a part in postnatal regulation of cardiocyte maturation; during life, cardiocytes remain quiescent and heart size increases by cellular hypertrophy[7].
Cardiac neural crest cells furnish mesenchymal cells to the heart and great arteries that are involved in vascular remodeling and development of the cardiac conduction system[8-10]. The sympathetic component of the autonomic nervous system promotes cardiac conduction while the parasympathetic selectively exerts an inhibitory influence[11,12]. The integration of information for neurocardiac regulation involves the neuraxis that comprises the cortex, amygdala and various subcortical structures with an ability to modulate lower-level neurons within the hierarchy (for a detailed explanation see ref.[12]). Principal contacts between preganglionic neurons and the heart occur via the vagus nerves[2,13]. Neurons of the autonomic nervous system are: (1) characterized by chemical phenotyping (cholinergic, adrenergic, etc.); (2) located within intrathoracic extracardiac ganglia and intrinsic cardiac ganglia[14,15]; and (3) found within atrial epicardium and ganglionated plexi along major vessels and in the ventricular wall[16,17] depending on species[18]. Sensory neurons, interneurons and sensory fibers that originate from the nucleus ambiguus are also located therein[19,20]. Sensory information from all of these peripheral structures is integrated with higher central nervous system centers to coordinate regulation of cardiovascular responses. For example, descending signals from higher brain centers as well as afferent sensory signals from systemic arteries, cardiopulmonary regions and viscera have their first synapse in the nucleus tractus solarius (NTS) found in the dorsomedial region of the medulla[21]. Transmission of afferent inputs from other sources such as skin and skeletal muscle to medullary vasomotor centers occur via the spinal cord. Vagal outflow to the heart is mediated by NTS neurons that synapse to preganglionic parasympathetic neurons located in the dorsal motor nucleus. All of these neural inputs to medullary vasomotor centers are involved in autonomic control of the cardiovascular system, for example, the arterial baroreceptor reflex plays a major role in blood pressure homeostasis on a beat-to-beat basis and involves stretch receptors that can be found in the carotid sinus and aortic arch. Accordingly, afferent baroreceptor discharge is relayed from the carotid sinus (via glossopharyngeal nerve) and aorta (via vagus nerve) to the NTS that stimulates afferent baroreceptor discharge and promotes efferent sympathetic and parasympathetic outflow to the heart and blood vessels, this enables adjustments of cardiac output and vessel resistance and ultimately facilitates return of blood pressure to steady state levels.
CORONARY BLOOD FLOW REGULATION AND MYOCARDIAL PERFUSION
Non-neural mechanisms (humoral, metabolic, mechanical, etc.) that contribute to control of vascular regulation act independently from autonomic neural mechanisms. For example, under normal physiological conditions myocardial perfusion across the ventricles is uniform as long as coronary artery pressure is maintained within the range of autoregulation[22]. Shifts in the lower pressure limit are produced by changes in left ventricular pressure and volume as well as biochemical modifications by a host of endogenous compounds that exercise their effects on myocytes, conduction tissues, vascular smooth muscle, etc. The scientific literature that has examined coronary vasoregulation with a focus on cardiac nerve status is relatively sparse. Most studies have concentrated on control of regional cardiodynamics by the intrinsic cardiac nervous system in either normal or pathological conditions.
In healthy individuals during exercise, activation of the sympathetic nervous system stimulates metabolic vasodilatation due to increases in heart rate, cardiac contractility and ventricular work. Direct sympathetic stimulation of coronary vessels induces either vasoconstriction or vasodilatation depending on activation of either α-, or β-adrenoreceptors, or vessel size. For example, large coronary vessels (> 100 μm) constrict when exposed to norepinephrine whereas small coronary vessels relax[23]; vasodilatation in arterioles permits coordination of oxygen delivery to myocardial oxygen demand[24]. On the other hand, simultaneous vasoconstriction in medium and large coronary arteries mediated by activation of α-adrenoreceptors helps to preserve subendocardial blood flow when oxygen demand increases. In a canine study, we examined myocardial perfusion following injection of select neuropeptides into active loci of the intrinsic cardiac nervous system and documented significant coronary vasodilatation secondary to increased myocardial metabolism and oxygen demand[25]. We also examined whether intact cardiac nerves were critical for coronary blood flow autoregulation; results confirmed a role for intrinsic cardiac neurons in autoregulatory control and myocardial perfusion even after ablation of extracardiac nerves from central nervous system control[26]. Ablation of external neuronal inputs to the heart also results in reduced myocardial efficiency that is consistent with impaired glucose utilization and depletion of cardiac catecholamine levels[27,28]; the latter directly affect myocardial oxygen demand[29-31]. Other animal studies reported that heterogeneity of myocardial perfusion is similar in innervated and denervated hearts[32-34]; possible explanations include: (1) the fact that regional denervation has little effect on vascular α-adrenergic receptors (in part due to circulating catecholamines); or (2) preserved neural modulation and autoregulation at different levels of the microcirculation across the ventricular wall[35,36].
Diverse central and peripheral elements within the cardiac nervous system act in sync to regulate cardiac function[20,37]; direct stimulation of intracardiac neurons occurs through central efferent neuronal inputs from the vagi or stellate ganglia[38]. G-protein coupled receptors are known to regulate cardiac function (see recent review by Capote et al[39] on structure, function and signalling pathways solicited by G-protein-coupled receptors in the heart). Control of heart rate requires intricate coordination between β-adrenergic and muscarinic cholinergic receptors found throughout the cardiac conduction system. Cardiac contraction controlled by β-adrenergic receptors are found in myocyte membranes while cardiac structure and morphology are coordinated by angiotensin II type 1 receptors in fibroblast and both endothelial cell and myocyte membranes[40,41]. Highly distinct processing capabilities of intracardiac neurons allow this complex network to respond to multiple inputs from all cardiac regions and major vessels near the heart. Disruption of these control networks by diverse cardiac pathologies ultimately increases the potential for sudden cardiac death[42-45].
MYOCARDIAL ISCHEMIA
Myocardial ischemia significantly influences cardiocytes as well as local and remote neurons that are involved in regulation of cardiac function[1,46]; the survival threshold of intra-/extra-cardiac sympathetic/parasympathetic neurons during development of coronary artery disease is not well established. However, viable nerves that course over an infarcted region tend to remain so oxygen and energy needs are fulfilled by an independent blood supply from extracardiac sources[47]. Reorganization of cardiocytes and nerves during development of diverse cardiac pathologies could occur in response to shifts of cardiac demand and function[3,48]. Mechanisms involved in the pathogenesis of cardiac dysfunction are multifactorial; a short list of possible factors include cardiac substrates, neural/cardiocyte interface, hormonal influences, inflammation and reflex responses between intra- and extra-cardiac nervous systems and their interactions with higher center neurons. Cardiocytes and cardiac neurons conceivably share common pathways for survival but this remains to be proven.
In the setting of transient ischemia, intact cardiac nerves are believed to play a key role on post-ischemic restoration of cardiac function[49]. Direct ischemic effects include progressive neuronal dysfunction and regional nerve terminal sprouting which ultimately diminishes local sensory and motor neurite function[50,51]. Indirect effects that modulate local neurite function are caused by local release of a host of endogenous chemicals (purinergic agents, peptides, hydroxyl radicals, etc.) that also affect neuronal function. Post-ischemic remodeling of cardiac neural networks could promote conflicts between central and peripheral reflexes that increases the risk of autonomic imbalances, arrhythmogenesis and sudden cardiac death[3,15,37,52]. A recent position paper by Ardell et al[1] discussed the significance of remodeling of the cardiac neuronal hierarchy to cardiac arrhythmia induction. In addition, inotropic stimulation is deleterious to myocyte survival as it occasions an imbalance between oxygen demand and supply (i.e., increased oxygen demand with limited coronary vascular reserve)[49,53].
Acute occlusion of a coronary artery produces distinct alterations of myocyte pathology that lead to cell death unless blood flow is restored to the affected myocardium, a transmural gradient of cell death occurs in relation to the duration of ischemia and degree of blood perfusion via coronary collateral vessels to the underperfused myocardium[54]. In animal models, necrosis is generally fully developed by 6 h after which tissue salvage is not possible (this time frame may not be the same for human myocardium) with currently available interventions. In addition, early restoration of blood flow to an infarct-related coronary vessel could cause “reperfusion injury” in already damaged or otherwise affected myocytes[55]. The physiopathology of ischemic, or reperfusion injury has been reviewed and discussed over the past several decades[56-59]; however, less attention has focused on the ability of the cardiac nervous system to accommodate the stress of ischemic, or reperfusion injury. Post-ischemic changes in peptide expression due to release of inflammatory cytokines combined with nerve damage could affect neuropeptide production in sympathetic cardiac neurons. In one study, Habecker et al[60] documented extensive axon damage after infarction; they also reported a significant increase of galanin (promotes regeneration of sensory neurons[61]) in cardiac sympathetic neurons in the left ventricle. These findings indicate that cardiac sympathetic neurons retain a certain capability to respond to nerve growth factor which is increased during ischemia-reperfusion[62].
While sympathetic dysinnervation has been reported secondary to myocardial infarction, the injury threshold of sympathetic and parasympathetic cardiac neurons within the ischemic region has not been established[63,64]. Several studies have documented that sympathetic impairment could exceed the area of underperfusion and necrosis[65,66]. Ischemic stress stimulates release of autocoids such as adenosine and bradykinin, along with nitric oxide and reactive oxygen species that can trigger cellular signal transduction pathways. These compounds can initiate responses in somata and axons within the intrinsic cardiac nervous system[37]. Indeed, oxidative stress, changes in growth factor expression and inflammatory cytokines released within the heart and vasculature contribute to neuronal remodelling[2,3,5,67]. As mentioned earlier, the regenerative capacity of cardiocytes is limited[68]; cardiocytes withdraw from the cell cycle early after birth and subsequently remain quiescent. Transition from proliferative to hypertrophic growth corresponds to the period of sympathetic growth into the heart tissues; in vitro studies with neonatal cardiocytes cultivated in the presence of innervating sympathetic fibers showed significant cellular proliferation[69] thereby confirming that early sympathetic signalling plays a role. In earlier in vitro studies, Horackova et al[70] reported that adult ventricular myocytes co-cultured with intrathoracic neurons retained similar structural properties to those observed in vivo; cardiocytes and intrinsic cardiac neurons that were cultured alone displayed a variety of morphologies (unipolar, bipolar, multipolar).
Sympathetic regulation might also be involved in myocyte regeneration following ischemia, or reperfusion, injury; however, disruption of peripheral nerves inhibits regeneration[71,72]. Chemical sympathectomy blocks early regeneration of damaged myocytes and increases tissue scarring[73]. Though additional studies are necessary, available data support the role of the intact cardiac nervous system on cardiocyte development and proliferation. On the other hand, post-ischemic regeneration and remodeling of the cardiac nervous system also merits further consideration and investigation. Rajendran et al[46] recently evaluated post-ischemic changes in neural signalling in a porcine model; they presented a “cardiac electroneurogram” between injured and adjacent non-injured myocardial tissue and reported: (1) that different intra-cardiac ganglia undergo morphological and phenotypic remodeling depending on the site of injury; (2) attenuation of afferent neural signals from the infarcted region to intra-cardiac neurons (activity in border and remote regions is apparently preserved); (3) maintenance of autonomic efferent inputs to the intrinsic cardiac nervous system; (4) augmented transduction capacity of convergent intrinsic cardiac local circuit neurons; and (5) reduced network connectivity within the intrinsic cardiac nervous system. The heterogeneity of afferent neural signals probably results from the presence of a “neural sensory border zone” (i.e., analogous to the so-called myocardial border zone) caused by scar formation during post-ischemic myocardial healing. This infarct-induced asymmetry of afferent inputs probably contributes to reflex activation of the autonomic nervous system; recent findings from Wang et al[74] using resiniferatoxin (a potent agonist of transient receptor potential vanilloid 1) showed reductions in cardiac afferent nociceptive signalling, and sympatho-excitation along with preserved cardiac function in rat hearts.
The role of intact cardiac nerves in modulating responses to ischemia and post-ischemic ventricular function has been studied in a variety of experimental models. In a cardiac decentralized porcine model subject to acute coronary artery stenosis Huang et al[49] reported significant ventricular dysfunction accompanied by patchy subendocardial necrosis; they proposed that the impaired recovery of left ventricular function is mediated by nitric oxide (NO) and reactive oxygen species (ROS). Cardiac nerves may help to attenuate production of ROS and/or prevent conversion of NO to peroxynitrite (via release of still unknown mediators/scavengers); neurotransmitters from cardiac nerves could stimulate or upregulate different isoforms of nitric oxide synthase (i.e., endothelial, neural)[75]. Myocardial perfusion-function relations are not altered by cardiac denervation[49]; this can be partly explained by the similarity between intact innervated and denervated hearts with regard to determinants of myocardial oxygen demand. In a recent study, we reported no significant change in coronary vascular reserve (intact cardiac nerves vs acute decentralized) in a canine model of ischemia-reperfusion injury[76]; these findings concur with most[77,78], but not all, earlier studies[79]. Of particular note is that protection against ischemic injury occurred even when affected myocardium was disconnected from central command; this suggests that local intrinsic cardiac neurons share common protection pathways to delay progression of cellular necrosis. Neurotransmitters that originate from cardiac nerves or intrinsic cardiac neurons might stimulate release of endogenous compounds that activate intracellular signalling pathways involved in cytoprotection; they could also inhibit peroxynitrite formation by modulating activation of various nitric oxide synthase isoforms. Indeed, many questions remain regarding the role of intact cardiac nerves within the context of cardioprotection against ischemia-reperfusion injury.
Myocardial ischemia also results in excessive activation of extracardiac cholinergic and adrenergic inputs of local circuit neurons within the intrinsic cardiac nervous system[38,80] that initiate cardiac arrhythmias[81]. A novel treatment for suppression of ventricular arrhythmias and treatment of refractory angina pectoris in current use in preclinical and clinical studies is spinal cord stimulation[80,82-84]; this intervention alters peripheral ganglia neural processing along the neural end-organ interface[85,86] and transduces neural signals to higher centers via the spinal cord[1,87,88]. Spinal cord stimulation influences autonomic reflexes within the neuroaxis and stimulates discharge of neuromodulators that limit release of select neurotransmitters and alter basal activity of sympathetic preganglionic neurons[89,90]. Intermittent spinal cord stimulation is suggested to stimulate neural memory and may be used for management of cardiac control and angina[91]; this could be akin to “electrical conditioning” and may be useful to limit cellular injury caused by ischemia. Vagus nerve stimulation is also being used to protect against ischemic injury and its consequences[92]; vagus nerve stimulation activates a host of signalling pathways and inhibits release of pro-inflammatory cytokines (see Ardell et al[1] for an up-to-date review). Vagus nerve stimulation might also affect myocardial energetics and maintain the equilibrium between energy supply and demand in the failing heart[93,94]. Interventions using vagus nerve stimulation favourably modulate cardiac disease as well as arrhythmogenesis; in several clinical studies this non-pharmacologic treatment is safe and well tolerated and is documented to improve cardiodynamics in patients with compromised ventricular function[95,96].
MYOCARDIAL PROTECTION
Sympathetic and parasympathetic nerves located near cardiocytes permit rapid crosstalk between cell types that may, or may not, activate cytoprotective pathways. Ischemic conditioning was first described by Murry et al[97] in 1986 in barbiturate-anesthetized dogs subjected to repeated episodes of sublethal coronary occlusion/reperfusion in advance of a prolonged period of acute ischemia. To date, ischemic conditioning has been reported to delay development of cellular necrosis in all organs examined in animals and in humans[98]; two distinct windows of cellular protection have been described but the causative mechanism(s) remain unanswered. The reader is referred to a recent review that summarizes research into this cytoprotective intervention over the past 30 years[99]. Interestingly, Kudej et al[100] showed that intact cardiac nerves were not required for first window protection in a porcine ischemia-reperfusion injury model; however, the presence of functional cardiac nerves was considered essential for development of second window protection. This delayed protection could occur through α1-adrenergic receptor pathways mediated by iNOS and COX-2[101].
A host of conditioning strategies have been described in animal and clinical studies; however, the potential to translate conditioning-mediated protection in patients remains controversial[102,103]. Remote conditioning was first described in dogs subject to acute coronary occlusion and was referred to as “preconditioning at a distance”[104]. In that study, animals were subject to repetitive periods of non-lethal ischemia of the left circumflex artery vascular bed before exposure to a prolonged occlusion of the left anterior descending coronary artery; results demonstrated that a cytoprotective factor could be activated, produced, or transported from the heart or elsewhere to affected tissues to afford protection. Since the publication of these key findings numerous studies using remote conditioning either before, during or after coronary occlusion have been reported[105-109] but the mechanisms involved have not been established. An important but unanswered question that persists is how the protective signals are transferred from distant tissues to the target organ. Various hypotheses (not mutually exclusive) including: (1) communication via blood or perfusate borne humoral factors; (2) communication by neuronal stimulation and transmission; and (3) communication by systemic alteration of circulating immune cells have been proposed[106,110,111]. Intrinsic neural loops in the heart process sensory information from the myocardium that modulate efferent autonomic output from the intrinsic cardiac ganglia even in the absence of input from the central nervous system[37,38,93,112]. Transmission of sensory messages within intrinsic cardiac ganglia is regulated by release of acetylcholine into the synaptic cleft; nerve impulses are initiated by acetylcholine that activates specific receptors in post-ganglionic nerves[112-114]. The risk of injury or remodeling of these neural loops escalates during myocardial ischemia; studies with pharmacologic ganglionic blockade document abolition of remote conditioning-mediated cytoprotection and suggest that protective signals could transfer between organs via neural pathways[112,115-117]. Early preclinical studies in different experimental models (including heart failure) reported positive results with vagal nerve stimulation (VNS) with respect to ventricular remodeling, ejection fraction and biomarker levels[118-120]. In patients with advanced heart failure, VNS reportedly attenuates left ventricular contractile dysfunction[121] and may reduce ischemic injury[122-124]. Clinical studies show that diminished heart-rate responses and depressed sensitivity of vagal reflexes are associated with poor cardiovascular outcomes and cardiac-related mortality[125-127]. Smith et al[127] recently reviewed efficacy of VNS for hypertension and heart failure in several small, randomized clinical trials (ANTHEM-HF, NECTAR-HF, INOVATE-HF, etc.) and concluded that further studies are required; VNS titration studies are also needed to validate potential clinical benefits of these interventions[128]. Stimulation of vagal nerves activate a host of signalling pathways via increased release of acetylcholine that activates downstream receptors (cholinergic, muscarinic, etc.) to impact cardiodynamics and could also promote myocyte resistance to stress by improving myocyte energetics[93]. Cross-talk between humoral mediators and neural pathways could also produce cytoprotection by stimulation of local afferent nerves[129,130]; but it remains unclear whether intact, functional nerves are required to assure conditioning-mediated cytoprotection[131,132]. On the basis of data showing that intact sensory innervation of peripheral ischemic tissue is essential to remote conditioning protection, Mastitskaya et al[133] proposed a “remote preconditioning reflex” that requires sensory input from remote ischemic tissue; recruitment of vagal pre-ganglionic neurons within the dorsal motor nucleus of the vagus nerve was considered to be critical for cytoprotection. While this data does not negate the concept that humoral factors are required for protection by remote conditioning, they strongly suggest that functional neurons within the parasympathetic nervous system are critical[134,135]. Bilateral vagotomy reportedly abolished protection afforded by remote conditioning[136]. On the other hand, findings from our laboratory (summarized in Figure 2) documented significant protection against ischemic injury independent of intact extrinsic cardiac nerves (note the similarity between groups with respect to reduction in infarct size) regardless of the conditioning protocol[76,137]. Briefly, in those studies isoflurane anesthetized dogs underwent remote conditioning (4 × 5-min renal artery occlusion/reperfusion) combined with/without treatment with the autonomic ganglionic blocker, hexamethonium (HEXA; 20 mg/kg, IV) or acute cardiac decentralization (DCN). Additional experiments were performed in dogs subject to classical preconditioning either before or after DCN. Based on these findings we suggested that neural pathways might not directly influence ischemic conditioning (either classical or remote) mediated cardioprotection. Moreover, others have brought forward the view that intact connections between the heart and central nervous system are not necessary for remote conditioning-mediated cardioprotection as long as recruitable parasympathetic neurons within a target organ can be activated. Use of remote conditioning as a potential therapeutic intervention for organ protection in man continues to merit investigation because it is non-invasive, cost-effective and easily applicable; however, the period for successful application of this intervention has yet to be determined and clinical strategies aimed at reducing myocardial damage by ischemic conditioning have been unsuccessful. While cellular protection by ischemic conditioning is possible in the presence of comorbidities, a stronger triggering stimulus appears necessary to assure cytoprotection[138].
Figure 2.
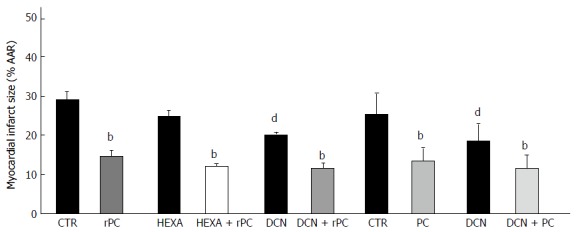
Myocardial infarct size (% anatomic area at risk: AAR) is shown for different study groups subject to ischemia-reperfusion injury. Data are means ± 1SD; bP ≤ 0.01 vs respective control (CTR), HEXA (hexmethonium; 20 mg/kg, IV), or DCN (acute cardiac decentralized) group; dP ≤ 0.01 vs CTR groups. Group differences determined by ANOVA. PC: Ischemic preconditioning; rPC: Remote preconditioning. Data reported in earlier studies from our laboratory[76,137].
Understanding bidirectional interactions between elements of the nervous system and its remodeling during evolution of different comorbidities (senescence, kidney dysfunction, diabetes, etc.) is essential to help in the development of strategies to delay progression of disease not only in the heart but also in other organs. For instance, autonomic neuropathies defined by abnormalities of the sympathetic and parasympathetic nervous systems could be responsible for significant morbidity and mortality in patients; cardiovascular events are considered a primary risk factor for mortality. Cardiovascular autonomic dysfunction is the result of complex interplay between vascular, neural, cardiac, paracrine and endocrine entities; the outcome is tissue injury that compromises integrity of cardiac reflexes.
HEART FAILURE
Heart failure subsequent to cardiac injury or chronic stress causes significant loss of contractile efficacy. Investigations into the role of autonomic imbalance between sympathetic and parasympathetic nervous systems and its contribution to pathogenesis of heart failure is ongoing for more than 25 years. Altered autonomic function also plays a role in other cardiac interrelated conditions such as hypertension, myocardial ischemia, cardiac arrhythmogenesis and sudden cardiac death[48], see recent review by Florea and Cohn[139]. Dynamic interactions between cardiocytes and compensatory neurohumoral mechanisms allow the heart to maintain cardiac output; stimulation of the adrenergic nervous and renin-angiotensin-aldosterone systems along with activation of cytokines play a critical role to prevent progressive worsening of cardiac function associated with heart failure[140,141]. Lymperopoulos et al[141] recently reviewed: (1) the actions of neurotransmitters on cell surface adrenergic and G-protein-coupled receptors; and (2) adrenergic receptor polymorphisms in the physiopathology of heart failure. They concluded that activation of the autonomic nervous system plays a critical role in compensatory responses to progressive cardiac dysfunction; however, excessive activation of these compensatory pathways could accelerate development of heart failure. In addition, they examined various therapeutic approaches (i.e., sympathomimetic drugs, activation of cardiac parasympathetic nervous system, increasing β-adrenergic receptor function using novel G-protein-coupled receptor blockade, etc.).
CHRONIC KIDNEY DISEASE AND NEUROPATHY
Physiopathology of chronic kidney disease (CKD) is complex and results either from a primary renal disorder or from multisystem disorders related to various comorbidities such as diabetes. Indeed, diabetes is considered to be the most common cause of CKD in patients. Neurological derangements are a common occurrence in CKD[142]. The spectrum of CKD ranges from mild kidney damage (largely asymptomatic) to end-stage renal disease (potentially fatal); neurological complications that include cognitive dysfunction, stroke, as well as peripheral and autonomic neuropathy can markedly affect clinical outcomes[143]. Accumulation of urea, creatinine, parathyroid hormone in high concentrations provide a biochemical milieu that rapidly produces neurological dysfunction; however, most symptoms can be reversed with treatments such as hemodialysis[144]. Mechanisms responsible for increased cardiovascular risk in patients with CKD are multifactorial and include hypertension and diabetes[145], along with increased oxidative stress, decreased bioavailability of nitric oxide, inflammation, abnormal calcium and phosphorous metabolism, overstimulation of the sympathetic nervous system, etc.[146-148]. Anemia is another major complication associated with both CKD and diabetes[149]; the latter may be present before overt evidence of symptoms of renal impairment[150].
Essential structures of the kidneys (renal vessels, tubules, juxtaglomerular apparatus, etc.) are richly innervated. Renal afferent nerves transmit sensory information via chemo- and mechano-receptors to higher centers within the brain[151,152], to maintain water retention, sodium reabsorption and blood flow. These nerves might also play a role in renal inflammation and injury; suggested mechanisms include β-adrenergic receptor activation, release of neuropeptides (neuropeptide Y, vasoactive intestinal polypeptide, substance P, etc.), renin release from juxtaglomerular cells (increases plasma angiotensin II levels) and other pro-inflammatory cytokines (tumor necrosis factor, IL-1β, etc.).
Autonomic dysfunction is prevalent (> 60%) in CKD patients and is associated with vascular calcification, cardiac arrhythmias and sudden cardiac death[153]. Reduced sensitivity to baroreceptors in the vessel wall caused by autonomic dysfunction can modulate cardiac regulation and contribute to intradialytic hypotension (i.e., no increase in heart rate to compensate the decrease in arterial pressure)[154]; these symptoms can be corrected with pharmaceuticals or, if necessary, renal transplantation.
DIABETIC AUTONOMIC NEUROPATHY
Autonomic dysfunction is a recognized complication of diabetes mellitus; diverse contributory mechanisms to increased mortality includes medial hyperplasia at baroreceptor sites, impaired cardiac vagal function, left ventricular hypertrophy and endothelial dysfunction[155] due in part to oxidative stress and reduced availability of nitric oxide which can affect sympathetic nerve activity[156]. Endothelial nitric oxide synthesis is known to be defective in insulin resistant states and is a central factor to neuronal abnormalities during metabolic syndrome (increases cardiovascular risk to some extent due to sympathetic activation)[155]. Insulin also plays a key role in nitric oxide and autonomic nervous system interactions and is involved in regulation of peripheral vascular tone and arterial blood pressure. Significant evidence shows that nitric oxide is critical to the vasodilator actions of insulin[157]; sympathectomy and autonomic failure can severely limit insulin-induced vasodilatation in patients[158]. Vulnerability to lethal arrhythmias in diabetic patients with autonomic dysfunction is also elevated[159]. Cardiac autonomic dysfunction may occur more frequently when diabetes is coupled with micro albuminuria caused by microvascular damage and endothelial dysfunction[160-162]; however, it was reported in the Hoorn Study that cardiovascular autonomic dysfunction and microalbuminuria were independently associated with mortality[163]. Additionally, in that study the presence of cardiovascular autonomic dysfunction doubled the 9-year mortality risk[155,164]; the ACCORD study also confirmed a significantly higher rate of mortality in patients with autonomic dysfunction[165].
CONCLUSION
Impaired sympathetic and parasympathetic nervous system regulation contributes to organ dysfunction and leads to significant morbidity and mortality particularly in patients with comorbidities. Early detection and management of these patients could markedly reduce adverse effects and thereby affect clinical outcomes. Prospectively, autonomic dysfunction develops because of damage at multiple sites within organs but pathogenesis remains to be clarified. Cardiovascular autonomic dysfunction, for instance, reflects compromised interactions between vascular, neural, cardiac, inflammatory, paracrine and endocrine mechanisms. Restoration of autonomic equilibrium in animal and clinical studies using either pharmacologic or non-pharmacologic interventions is currently possible. Further investigations in neurocardiology should continue to provide important findings apropos connections between cardiac and neurohumoral control systems and thereby allow continued development of clinically relevant opportunities for neuroscience-based treatments.
Footnotes
Manuscript source: Invited manuscript
Specialty type: Cardiac and cardiovascular systems
Country of origin: Canada
Peer-review report classification
Grade A (Excellent): A, A
Grade B (Very good): B
Grade C (Good): C
Grade D (Fair): 0
Grade E (Poor): 0
Conflict-of-interest statement: The authors declare no conflict of interest with regard to research, authorship or publication for this article.
Peer-review started: February 10, 2017
First decision: March 6, 2017
Article in press: April 24, 2017
P- Reviewer: Ilgenli TF, Lymperopoulos A, Peteiro J, Ueda H S- Editor: Ji FF L- Editor: A E- Editor: Li D
References
- 1.Ardell JL, Andresen MC, Armour JA, Billman GE, Chen PS, Foreman RD, Herring N, O’Leary DS, Sabbah HN, Schultz HD, et al. Translational neurocardiology: preclinical models and cardioneural integrative aspects. J Physiol. 2016;594:3877–3909. doi: 10.1113/JP271869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Kimura K, Ieda M, Fukuda K. Development, maturation, and transdifferentiation of cardiac sympathetic nerves. Circ Res. 2012;110:325–336. doi: 10.1161/CIRCRESAHA.111.257253. [DOI] [PubMed] [Google Scholar]
- 3.Fukuda K, Kanazawa H, Aizawa Y, Ardell JL, Shivkumar K. Cardiac innervation and sudden cardiac death. Circ Res. 2015;116:2005–2019. doi: 10.1161/CIRCRESAHA.116.304679. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Habecker BA, Anderson ME, Birren SJ, Fukuda K, Herring N, Hoover DB, Kanazawa H, Paterson DJ, Ripplinger CM. Molecular and cellular neurocardiology: development, and cellular and molecular adaptations to heart disease. J Physiol. 2016;594:3853–3875. doi: 10.1113/JP271840. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Gardner RT, Ripplinger CM, Myles RC, Habecker BA. Molecular Mechanisms of Sympathetic Remodeling and Arrhythmias. Circ Arrhythm Electrophysiol. 2016;9:e001359. doi: 10.1161/CIRCEP.115.001359. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Shivkumar K, Ardell JL. Cardiac autonomic control in health and disease. J Physiol. 2016;594:3851–3852. doi: 10.1113/JP272580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Ahuja P, Perriard E, Pedrazzini T, Satoh S, Perriard JC, Ehler E. Re-expression of proteins involved in cytokinesis during cardiac hypertrophy. Exp Cell Res. 2007;313:1270–1283. doi: 10.1016/j.yexcr.2007.01.009. [DOI] [PubMed] [Google Scholar]
- 8.Stoller JZ, Epstein JA. Cardiac neural crest. Semin Cell Dev Biol. 2005;16:704–715. doi: 10.1016/j.semcdb.2005.06.004. [DOI] [PubMed] [Google Scholar]
- 9.Creazzo TL, Godt RE, Leatherbury L, Conway SJ, Kirby ML. Role of cardiac neural crest cells in cardiovascular development. Annu Rev Physiol. 1998;60:267–286. doi: 10.1146/annurev.physiol.60.1.267. [DOI] [PubMed] [Google Scholar]
- 10.Coskun V, Lombardo DM. Studying the pathophysiologic connection between cardiovascular and nervous systems using stem cells. J Neurosci Res. 2016;94:1499–1510. doi: 10.1002/jnr.23924. [DOI] [PubMed] [Google Scholar]
- 11.Pardini BJ, Lund DD, Schmid PG. Organization of the sympathetic postganglionic innervation of the rat heart. J Auton Nerv Syst. 1989;28:193–201. doi: 10.1016/0165-1838(89)90146-x. [DOI] [PubMed] [Google Scholar]
- 12.Wallis D, Watson AH, Mo N. Cardiac neurones of autonomic ganglia. Microsc Res Tech. 1996;35:69–79. doi: 10.1002/(SICI)1097-0029(19960901)35:1<69::AID-JEMT6>3.0.CO;2-N. [DOI] [PubMed] [Google Scholar]
- 13.Hasan W. Autonomic cardiac innervation: development and adult plasticity. Organogenesis. 2013;9:176–193. doi: 10.4161/org.24892. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Cardinal R, Pagé P, Vermeulen M, Ardell JL, Armour JA. Spatially divergent cardiac responses to nicotinic stimulation of ganglionated plexus neurons in the canine heart. Auton Neurosci. 2009;145:55–62. doi: 10.1016/j.autneu.2008.11.007. [DOI] [PubMed] [Google Scholar]
- 15.Kember G, Armour JA, Zamir M. Neural control hierarchy of the heart has not evolved to deal with myocardial ischemia. Physiol Genomics. 2013;45:638–644. doi: 10.1152/physiolgenomics.00027.2013. [DOI] [PubMed] [Google Scholar]
- 16.Randall WC, Milosavljevic M, Wurster RD, Geis GS, Ardell JL. Selective vagal innervation of the heart. Ann Clin Lab Sci. 1986;16:198–208. [PubMed] [Google Scholar]
- 17.Singh S, Johnson PI, Lee RE, Orfei E, Lonchyna VA, Sullivan HJ, Montoya A, Tran H, Wehrmacher WH, Wurster RD. Topography of cardiac ganglia in the adult human heart. J Thorac Cardiovasc Surg. 1996;112:943–953. doi: 10.1016/S0022-5223(96)70094-6. [DOI] [PubMed] [Google Scholar]
- 18.Kuder T, Nowak E, Szczurkowski A, Kuchinka J. A comparative study on cardiac ganglia in midday gerbil, Egyptian spiny mouse, chinchilla laniger and pigeon. Anat Histol Embryol. 2003;32:134–140. doi: 10.1046/j.1439-0264.2003.00445.x. [DOI] [PubMed] [Google Scholar]
- 19.Ai J, Epstein PN, Gozal D, Yang B, Wurster R, Cheng ZJ. Morphology and topography of nucleus ambiguus projections to cardiac ganglia in rats and mice. Neuroscience. 2007;149:845–860. doi: 10.1016/j.neuroscience.2007.07.062. [DOI] [PubMed] [Google Scholar]
- 20.McAllen RM, Salo LM, Paton JF, Pickering AE. Processing of central and reflex vagal drives by rat cardiac ganglion neurones: an intracellular analysis. J Physiol. 2011;589:5801–5818. doi: 10.1113/jphysiol.2011.214320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Thomas GD. Neural control of the circulation. Adv Physiol Educ. 2011;35:28–32. doi: 10.1152/advan.00114.2010. [DOI] [PubMed] [Google Scholar]
- 22.Rouleau J, Boerboom LE, Surjadhana A, Hoffman JI. The role of autoregulation and tissue diastolic pressures in the transmural distribution of left ventricular blood flow in anesthetized dogs. Circ Res. 1979;45:804–815. doi: 10.1161/01.res.45.6.804. [DOI] [PubMed] [Google Scholar]
- 23.Chilian WM, Layne SM, Eastham CL, Marcus ML. Heterogeneous microvascular coronary alpha-adrenergic vasoconstriction. Circ Res. 1989;64:376–388. doi: 10.1161/01.res.64.2.376. [DOI] [PubMed] [Google Scholar]
- 24.Tune JD, Gorman MW, Feigl EO. Matching coronary blood flow to myocardial oxygen consumption. J Appl Physiol (1985) 2004;97:404–415. doi: 10.1152/japplphysiol.01345.2003. [DOI] [PubMed] [Google Scholar]
- 25.Kingma JG, Armour JA, Rouleau JR. Chemical modulation of in situ intrinsic cardiac neurones influences myocardial blood flow in the anaesthetised dog. Cardiovasc Res. 1994;28:1403–1406. doi: 10.1093/cvr/28.9.1403. [DOI] [PubMed] [Google Scholar]
- 26.Rouleau JR, Simard D, Rodrigue N, Blouin A, Kingma JG. Myocardial blood flow after chronic cardiac decentralization in anesthetized dogs: effects of ACE-inhibition. Auton Neurosci. 2002;97:12–18. doi: 10.1016/s1566-0702(02)00002-4. [DOI] [PubMed] [Google Scholar]
- 27.Drake AJ, Papadoyannis DE, Butcher RG, Stubbs J, Noble MI. Inhibition of glycolysis in the denervated dog heart. Circ Res. 1980;47:338–345. doi: 10.1161/01.res.47.3.338. [DOI] [PubMed] [Google Scholar]
- 28.van der Vusse GJ, Dubelaar ML, Coumans WA, Seymour AM, Clarke SB, Bonen A, Drake-Holland AJ, Noble MI. Metabolic alterations in the chronically denervated dog heart. Cardiovasc Res. 1998;37:160–170. doi: 10.1016/s0008-6363(97)00220-4. [DOI] [PubMed] [Google Scholar]
- 29.Butcher RG, Drake-Holland AJ, Wanless RB, Papadoyannis DE, Noble MI. Effect of lack of noradrenaline on myocardial oxygen consumption in denervated dog hearts. Cardiovasc Res. 1987;21:515–520. doi: 10.1093/cvr/21.7.515. [DOI] [PubMed] [Google Scholar]
- 30.Vatner DE, Lavallee M, Amano J, Finizola A, Homcy CJ, Vatner SF. Mechanisms of supersensitivity to sympathomimetic amines in the chronically denervated heart of the conscious dog. Circ Res. 1985;57:55–64. doi: 10.1161/01.res.57.1.55. [DOI] [PubMed] [Google Scholar]
- 31.Vergroesen I, Merkus D, van Teeffelen JW, Dankelman J, Spaan JA, van Wezel HB, Noble MI, Drake-Holland AJ. Chronic cardiac denervation affects the speed of coronary vascular regulation. Cardiovasc Res. 1999;44:615–622. doi: 10.1016/s0008-6363(99)00257-6. [DOI] [PubMed] [Google Scholar]
- 32.Rimoldi OE, Drake-Holland AJ, Noble MI, Camici PG. Basal and hyperaemic myocardial blood flow in regionally denervated canine hearts: an in vivo study with positron emission tomography. Eur J Nucl Med Mol Imaging. 2007;34:197–205. doi: 10.1007/s00259-006-0233-0. [DOI] [PubMed] [Google Scholar]
- 33.Alders DJ, Cornelussen RN, Prinzen FW, Specht PA, Noble MI, Drake-Holland AJ, de Kanter FJ, van Beek JH. Regional sympathetic denervation affects the relation between canine local myocardial blood flow and oxygen consumption. Exp Physiol. 2007;92:541–548. doi: 10.1113/expphysiol.2006.036228. [DOI] [PubMed] [Google Scholar]
- 34.Chilian WM, Ackell PH. Transmural differences in sympathetic coronary constriction during exercise in the presence of coronary stenosis. Circ Res. 1988;62:216–225. doi: 10.1161/01.res.62.2.216. [DOI] [PubMed] [Google Scholar]
- 35.Marcus ML, Chilian WM, Kanatsuka H, Dellsperger KC, Eastham CL, Lamping KG. Understanding the coronary circulation through studies at the microvascular level. Circulation. 1990;82:1–7. doi: 10.1161/01.cir.82.1.1. [DOI] [PubMed] [Google Scholar]
- 36.Chilian WM, Harrison DG, Haws CW, Snyder WD, Marcus ML. Adrenergic coronary tone during submaximal exercise in the dog is produced by circulating catecholamines. Evidence for adrenergic denervation supersensitivity in the myocardium but not in coronary vessels. Circ Res. 1986;58:68–82. doi: 10.1161/01.res.58.1.68. [DOI] [PubMed] [Google Scholar]
- 37.Armour JA. Potential clinical relevance of the ‘little brain’ on the mammalian heart. Exp Physiol. 2008;93:165–176. doi: 10.1113/expphysiol.2007.041178. [DOI] [PubMed] [Google Scholar]
- 38.Beaumont E, Salavatian S, Southerland EM, Vinet A, Jacquemet V, Armour JA, Ardell JL. Network interactions within the canine intrinsic cardiac nervous system: implications for reflex control of regional cardiac function. J Physiol. 2013;591:4515–4533. doi: 10.1113/jphysiol.2013.259382. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Capote LA, Mendez Perez R, Lymperopoulos A. GPCR signaling and cardiac function. Eur J Pharmacol. 2015;763:143–148. doi: 10.1016/j.ejphar.2015.05.019. [DOI] [PubMed] [Google Scholar]
- 40.Lymperopoulos A, Bathgate A. Arrestins in the cardiovascular system. Prog Mol Biol Transl Sci. 2013;118:297–334. doi: 10.1016/B978-0-12-394440-5.00012-7. [DOI] [PubMed] [Google Scholar]
- 41.Siryk-Bathgate A, Dabul S, Lymperopoulos A. Current and future G protein-coupled receptor signaling targets for heart failure therapy. Drug Des Devel Ther. 2013;7:1209–1222. doi: 10.2147/DDDT.S35905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Arora RC, Cardinal R, Smith FM, Ardell JL, Dell’Italia LJ, Armour JA. Intrinsic cardiac nervous system in tachycardia induced heart failure. Am J Physiol Regul Integr Comp Physiol. 2003;285:R1212–R1223. doi: 10.1152/ajpregu.00131.2003. [DOI] [PubMed] [Google Scholar]
- 43.Hardwick JC, Southerland EM, Girasole AE, Ryan SE, Negrotto S, Ardell JL. Remodeling of intrinsic cardiac neurons: effects of β-adrenergic receptor blockade in guinea pig models of chronic heart disease. Am J Physiol Regul Integr Comp Physiol. 2012;303:R950–R958. doi: 10.1152/ajpregu.00223.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Nguyen BL, Li H, Fishbein MC, Lin SF, Gaudio C, Chen PS, Chen LS. Acute myocardial infarction induces bilateral stellate ganglia neural remodeling in rabbits. Cardiovasc Pathol. 2012;21:143–148. doi: 10.1016/j.carpath.2011.08.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Zucker IH, Patel KP, Schultz HD. Neurohumoral stimulation. Heart Fail Clin. 2012;8:87–99. doi: 10.1016/j.hfc.2011.08.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Rajendran PS, Nakamura K, Ajijola OA, Vaseghi M, Armour JA, Ardell JL, Shivkumar K. Myocardial infarction induces structural and functional remodelling of the intrinsic cardiac nervous system. J Physiol. 2016;594:321–341. doi: 10.1113/JP271165. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Janes RD, Johnstone DE, Armour JA. Functional integrity of intrinsic cardiac nerves located over an acute transmural myocardial infarction. Can J Physiol Pharmacol. 1987;65:64–69. doi: 10.1139/y87-012. [DOI] [PubMed] [Google Scholar]
- 48.Vaseghi M, Shivkumar K. The role of the autonomic nervous system in sudden cardiac death. Prog Cardiovasc Dis. 2008;50:404–419. doi: 10.1016/j.pcad.2008.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Huang CH, Vatner SF, Peppas AP, Yang G, Kudej RK. Cardiac nerves affect myocardial stunning through reactive oxygen and nitric oxide mechanisms. Circ Res. 2003;93:866–873. doi: 10.1161/01.RES.0000097762.64561.D2. [DOI] [PubMed] [Google Scholar]
- 50.Armour JA. Cardiac neuronal hierarchy in health and disease. Am J Physiol Regul Integr Comp Physiol. 2004;287:R262–R271. doi: 10.1152/ajpregu.00183.2004. [DOI] [PubMed] [Google Scholar]
- 51.Chen PS, Chen LS, Cao JM, Sharifi B, Karagueuzian HS, Fishbein MC. Sympathetic nerve sprouting, electrical remodeling and the mechanisms of sudden cardiac death. Cardiovasc Res. 2001;50:409–416. doi: 10.1016/s0008-6363(00)00308-4. [DOI] [PubMed] [Google Scholar]
- 52.Nakamura K, Ajijola OA, Aliotta E, Armour JA, Ardell JL, Shivkumar K. Pathological effects of chronic myocardial infarction on peripheral neurons mediating cardiac neurotransmission. Auton Neurosci. 2016;197:34–40. doi: 10.1016/j.autneu.2016.05.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Schulz R, Rose J, Martin C, Brodde OE, Heusch G. Development of short-term myocardial hibernation. Its limitation by the severity of ischemia and inotropic stimulation. Circulation. 1993;88:684–695. doi: 10.1161/01.cir.88.2.684. [DOI] [PubMed] [Google Scholar]
- 54.Reimer KA, Lowe JE, Rasmussen MM, Jennings RB. The wavefront phenomenon of ischemic cell death. 1. Myocardial infarct size vs duration of coronary occlusion in dogs. Circulation. 1977;56:786–794. doi: 10.1161/01.cir.56.5.786. [DOI] [PubMed] [Google Scholar]
- 55.Braunwald E, Kloner RA. Myocardial reperfusion: a double-edged sword? J Clin Invest. 1985;76:1713–1719. doi: 10.1172/JCI112160. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Jennings RB. Historical perspective on the pathology of myocardial ischemia/reperfusion injury. Circ Res. 2013;113:428–438. doi: 10.1161/CIRCRESAHA.113.300987. [DOI] [PubMed] [Google Scholar]
- 57.Kloner RA, Jennings RB. Consequences of brief ischemia: stunning, preconditioning, and their clinical implications: part 1. Circulation. 2001;104:2981–2989. doi: 10.1161/hc4801.100038. [DOI] [PubMed] [Google Scholar]
- 58.Kloner RA, Jennings RB. Consequences of brief ischemia: stunning, preconditioning, and their clinical implications: part 2. Circulation. 2001;104:3158–3167. doi: 10.1161/hc5001.100039. [DOI] [PubMed] [Google Scholar]
- 59.Yellon DM, Hausenloy DJ. Myocardial reperfusion injury. N Engl J Med. 2007;357:1121–1135. doi: 10.1056/NEJMra071667. [DOI] [PubMed] [Google Scholar]
- 60.Habecker BA, Gritman KR, Willison BD, Van Winkle DM. Myocardial infarction stimulates galanin expression in cardiac sympathetic neurons. Neuropeptides. 2005;39:89–95. doi: 10.1016/j.npep.2004.11.003. [DOI] [PubMed] [Google Scholar]
- 61.Mahoney SA, Hosking R, Farrant S, Holmes FE, Jacoby AS, Shine J, Iismaa TP, Scott MK, Schmidt R, Wynick D. The second galanin receptor GalR2 plays a key role in neurite outgrowth from adult sensory neurons. J Neurosci. 2003;23:416–421. doi: 10.1523/JNEUROSCI.23-02-00416.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Abe T, Morgan DA, Gutterman DD. Protective role of nerve growth factor against postischemic dysfunction of sympathetic coronary innervation. Circulation. 1997;95:213–220. doi: 10.1161/01.cir.95.1.213. [DOI] [PubMed] [Google Scholar]
- 63.Fallavollita JA, Canty JM. Dysinnervated but viable myocardium in ischemic heart disease. J Nucl Cardiol. 2010;17:1107–1115. doi: 10.1007/s12350-010-9292-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Bulow HP, Stahl F, Lauer B, Nekolla SG, Schuler G, Schwaiger M, Bengel FM. Alterations of myocardial presynaptic sympathetic innervation in patients with multi-vessel coronary artery disease but without history of myocardial infarction. Nucl Med Commun. 2003;24:233–239. doi: 10.1097/00006231-200303000-00002. [DOI] [PubMed] [Google Scholar]
- 65.Barber MJ, Mueller TM, Henry DP, Felten SY, Zipes DP. Transmural myocardial infarction in the dog produces sympathectomy in noninfarcted myocardium. Circulation. 1983;67:787–796. doi: 10.1161/01.cir.67.4.787. [DOI] [PubMed] [Google Scholar]
- 66.Dae MW, Herre JM, O’Connell JW, Botvinick EH, Newman D, Munoz L. Scintigraphic assessment of sympathetic innervation after transmural versus nontransmural myocardial infarction. J Am Coll Cardiol. 1991;17:1416–1423. doi: 10.1016/s0735-1097(10)80156-1. [DOI] [PubMed] [Google Scholar]
- 67.Olivas A, Gardner RT, Wang L, Ripplinger CM, Woodward WR, Habecker BA. Myocardial Infarction Causes Transient Cholinergic Transdifferentiation of Cardiac Sympathetic Nerves via gp130. J Neurosci. 2016;36:479–488. doi: 10.1523/JNEUROSCI.3556-15.2016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Zak R. Cell proliferation during cardiac growth. Am J Cardiol. 1973;31:211–219. doi: 10.1016/0002-9149(73)91034-5. [DOI] [PubMed] [Google Scholar]
- 69.Kreipke RE, Birren SJ. Innervating sympathetic neurons regulate heart size and the timing of cardiomyocyte cell cycle withdrawal. J Physiol. 2015;593:5057–5073. doi: 10.1113/JP270917. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Horackova M, Croll RP, Hopkins DA, Losier AM, Armour JA. Morphological and immunohistochemical properties of primary long-term cultures of adult guinea-pig ventricular cardiomyocytes with peripheral cardiac neurons. Tissue Cell. 1996;28:411–425. doi: 10.1016/s0040-8166(96)80027-9. [DOI] [PubMed] [Google Scholar]
- 71.Mahmoud AI, O’Meara CC, Gemberling M, Zhao L, Bryant DM, Zheng R, Gannon JB, Cai L, Choi WY, Egnaczyk GF, et al. Nerves Regulate Cardiomyocyte Proliferation and Heart Regeneration. Dev Cell. 2015;34:387–399. doi: 10.1016/j.devcel.2015.06.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Bryant DM, O’Meara CC, Ho NN, Gannon J, Cai L, Lee RT. A systematic analysis of neonatal mouse heart regeneration after apical resection. J Mol Cell Cardiol. 2015;79:315–318. doi: 10.1016/j.yjmcc.2014.12.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.White IA, Gordon J, Balkan W, Hare JM. Sympathetic Reinnervation Is Required for Mammalian Cardiac Regeneration. Circ Res. 2015;117:990–994. doi: 10.1161/CIRCRESAHA.115.307465. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Wang HJ, Wang W, Cornish KG, Rozanski GJ, Zucker IH. Cardiac sympathetic afferent denervation attenuates cardiac remodeling and improves cardiovascular dysfunction in rats with heart failure. Hypertension. 2014;64:745–755. doi: 10.1161/HYPERTENSIONAHA.114.03699. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Sears CE, Bryant SM, Ashley EA, Lygate CA, Rakovic S, Wallis HL, Neubauer S, Terrar DA, Casadei B. Cardiac neuronal nitric oxide synthase isoform regulates myocardial contraction and calcium handling. Circ Res. 2003;92:e52–e59. doi: 10.1161/01.RES.0000064585.95749.6D. [DOI] [PubMed] [Google Scholar]
- 76.Kingma JG, Simard D, Voisine P, Rouleau JR. Influence of cardiac decentralization on cardioprotection. PLoS One. 2013;8:e79190. doi: 10.1371/journal.pone.0079190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Jones CE, Devous MD, Thomas JX, DuPont E. The effect of chronic cardiac denervation on infarct size following acute coronary occlusion. Am Heart J. 1978;95:738–746. doi: 10.1016/0002-8703(78)90504-5. [DOI] [PubMed] [Google Scholar]
- 78.Jones CE, Beck LY, DuPont E, Barnes GE. Effects of coronary ligation of the chromically sympathectomized dog ventricle. Am J Physiol. 1978;235:H429–H434. doi: 10.1152/ajpheart.1978.235.4.H429. [DOI] [PubMed] [Google Scholar]
- 79.Lavallee M, Amano J, Vatner SF, Manders WT, Randall WC, Thomas JX. Adverse effects of chronic cardiac denervation in conscious dogs with myocardial ischemia. Circ Res. 1985;57:383–392. doi: 10.1161/01.res.57.3.383. [DOI] [PubMed] [Google Scholar]
- 80.Gibbons DD, Southerland EM, Hoover DB, Beaumont E, Armour JA, Ardell JL. Neuromodulation targets intrinsic cardiac neurons to attenuate neuronally mediated atrial arrhythmias. Am J Physiol Regul Integr Comp Physiol. 2012;302:R357–R364. doi: 10.1152/ajpregu.00535.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Armour JA, Richer LP, Pagé P, Vinet A, Kus T, Vermeulen M, Nadeau R, Cardinal R. Origin and pharmacological response of atrial tachyarrhythmias induced by activation of mediastinal nerves in canines. Auton Neurosci. 2005;118:68–78. doi: 10.1016/j.autneu.2005.01.006. [DOI] [PubMed] [Google Scholar]
- 82.Cardinal R, Pagé P, Vermeulen M, Bouchard C, Ardell JL, Foreman RD, Armour JA. Spinal cord stimulation suppresses bradycardias and atrial tachyarrhythmias induced by mediastinal nerve stimulation in dogs. Am J Physiol Regul Integr Comp Physiol. 2006;291:R1369–R1375. doi: 10.1152/ajpregu.00056.2006. [DOI] [PubMed] [Google Scholar]
- 83.Armour JA, Linderoth B, Arora RC, DeJongste MJ, Ardell JL, Kingma JG, Hill M, Foreman RD. Long-term modulation of the intrinsic cardiac nervous system by spinal cord neurons in normal and ischaemic hearts. Auton Neurosci. 2002;95:71–79. doi: 10.1016/s1566-0702(01)00377-0. [DOI] [PubMed] [Google Scholar]
- 84.Kingma JG, Linderoth B, Ardell JL, Armour JA, DeJongste MJ, Foreman RD. Neuromodulation therapy does not influence blood flow distribution or left-ventricular dynamics during acute myocardial ischemia. Auton Neurosci. 2001;91:47–54. doi: 10.1016/S1566-0702(01)00285-5. [DOI] [PubMed] [Google Scholar]
- 85.Ardell JL, Cardinal R, Vermeulen M, Armour JA. Dorsal spinal cord stimulation obtunds the capacity of intrathoracic extracardiac neurons to transduce myocardial ischemia. Am J Physiol Regul Integr Comp Physiol. 2009;297:R470–R477. doi: 10.1152/ajpregu.90821.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Southerland EM, Milhorn DM, Foreman RD, Linderoth B, DeJongste MJ, Armour JA, Subramanian V, Singh M, Singh K, Ardell JL. Preemptive, but not reactive, spinal cord stimulation mitigates transient ischemia-induced myocardial infarction via cardiac adrenergic neurons. Am J Physiol Heart Circ Physiol. 2007;292:H311–H317. doi: 10.1152/ajpheart.00087.2006. [DOI] [PubMed] [Google Scholar]
- 87.Linderoth B, Foreman RD. Physiology of spinal cord stimulation: review and update. Neuromodulation. 1999;2:150–164. doi: 10.1046/j.1525-1403.1999.00150.x. [DOI] [PubMed] [Google Scholar]
- 88.Yamakawa K, Howard-Quijano K, Zhou W, Rajendran P, Yagishita D, Vaseghi M, Ajijola OA, Armour JA, Shivkumar K, Ardell JL, et al. Central vs. peripheral neuraxial sympathetic control of porcine ventricular electrophysiology. Am J Physiol Regul Integr Comp Physiol. 2016;310:R414–R421. doi: 10.1152/ajpregu.00252.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Ding X, Ardell JL, Hua F, McAuley RJ, Sutherly K, Daniel JJ, Williams CA. Modulation of cardiac ischemia-sensitive afferent neuron signaling by preemptive C2 spinal cord stimulation: effect on substance P release from rat spinal cord. Am J Physiol Regul Integr Comp Physiol. 2008;294:R93–101. doi: 10.1152/ajpregu.00544.2007. [DOI] [PubMed] [Google Scholar]
- 90.Ding X, Hua F, Sutherly K, Ardell JL, Williams CA. C2 spinal cord stimulation induces dynorphin release from rat T4 spinal cord: potential modulation of myocardial ischemia-sensitive neurons. Am J Physiol Regul Integr Comp Physiol. 2008;295:R1519–R1528. doi: 10.1152/ajpregu.00899.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Foreman RD, Linderoth B. Neural mechanisms of spinal cord stimulation. Int Rev Neurobiol. 2012;107:87–119. doi: 10.1016/B978-0-12-404706-8.00006-1. [DOI] [PubMed] [Google Scholar]
- 92.Jänig W. Autonomic nervous system and inflammation. Auton Neurosci. 2014;182:1–3. doi: 10.1016/j.autneu.2014.02.002. [DOI] [PubMed] [Google Scholar]
- 93.Beaumont E, Southerland EM, Hardwick JC, Wright GL, Ryan S, Li Y, KenKnight BH, Armour JA, Ardell JL. Vagus nerve stimulation mitigates intrinsic cardiac neuronal and adverse myocyte remodeling postmyocardial infarction. Am J Physiol Heart Circ Physiol. 2015;309:H1198–H1206. doi: 10.1152/ajpheart.00393.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.De Ferrari GM, Vanoli E, Stramba-Badiale M, Hull SS, Foreman RD, Schwartz PJ. Vagal reflexes and survival during acute myocardial ischemia in conscious dogs with healed myocardial infarction. Am J Physiol. 1991;261:H63–H69. doi: 10.1152/ajpheart.1991.261.1.H63. [DOI] [PubMed] [Google Scholar]
- 95.Premchand RK, Sharma K, Mittal S, Monteiro R, Dixit S, Libbus I, DiCarlo LA, Ardell JL, Rector TS, Amurthur B, et al. Autonomic regulation therapy via left or right cervical vagus nerve stimulation in patients with chronic heart failure: results of the ANTHEM-HF trial. J Card Fail. 2014;20:808–816. doi: 10.1016/j.cardfail.2014.08.009. [DOI] [PubMed] [Google Scholar]
- 96.De Ferrari GM, Tuinenburg AE, Ruble S, Brugada J, Klein H, Butter C, Wright DJ, Schubert B, Solomon S, Meyer S, et al. Rationale and study design of the NEuroCardiac TherApy foR Heart Failure Study: NECTAR-HF. Eur J Heart Fail. 2014;16:692–699. doi: 10.1002/ejhf.80. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Murry CE, Jennings RB, Reimer KA. Preconditioning with ischemia: a delay of lethal cell injury in ischemic myocardium. Circulation. 1986;74:1124–1136. doi: 10.1161/01.cir.74.5.1124. [DOI] [PubMed] [Google Scholar]
- 98.Yellon DM, Downey JM. Preconditioning the myocardium: from cellular physiology to clinical cardiology. Physiol Rev. 2003;83:1113–1151. doi: 10.1152/physrev.00009.2003. [DOI] [PubMed] [Google Scholar]
- 99.Hausenloy DJ, Barrabes JA, Bøtker HE, Davidson SM, Di Lisa F, Downey J, Engstrom T, Ferdinandy P, Carbrera-Fuentes HA, Heusch G, et al. Ischaemic conditioning and targeting reperfusion injury: a 30 year voyage of discovery. Basic Res Cardiol. 2016;111:70. doi: 10.1007/s00395-016-0588-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Kudej RK, Shen YT, Peppas AP, Huang CH, Chen W, Yan L, Vatner DE, Vatner SF. Obligatory role of cardiac nerves and alpha1-adrenergic receptors for the second window of ischemic preconditioning in conscious pigs. Circ Res. 2006;99:1270–1276. doi: 10.1161/01.RES.0000251282.79411.44. [DOI] [PubMed] [Google Scholar]
- 101.Shinmura K, Tang XL, Wang Y, Xuan YT, Liu SQ, Takano H, Bhatnagar A, Bolli R. Cyclooxygenase-2 mediates the cardioprotective effects of the late phase of ischemic preconditioning in conscious rabbits. Proc Natl Acad Sci USA. 2000;97:10197–10202. doi: 10.1073/pnas.97.18.10197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Ovize M, Thibault H, Przyklenk K. Myocardial conditioning: opportunities for clinical translation. Circ Res. 2013;113:439–450. doi: 10.1161/CIRCRESAHA.113.300764. [DOI] [PubMed] [Google Scholar]
- 103.Vander Heide RS, Steenbergen C. Cardioprotection and myocardial reperfusion: pitfalls to clinical application. Circ Res. 2013;113:464–477. doi: 10.1161/CIRCRESAHA.113.300765. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Przyklenk K, Bauer B, Ovize M, Kloner RA, Whittaker P. Regional ischemic ‘preconditioning’ protects remote virgin myocardium from subsequent sustained coronary occlusion. Circulation. 1993;87:893–899. doi: 10.1161/01.cir.87.3.893. [DOI] [PubMed] [Google Scholar]
- 105.Konstantinov IE, Arab S, Kharbanda RK, Li J, Cheung MM, Cherepanov V, Downey GP, Liu PP, Cukerman E, Coles JG, et al. The remote ischemic preconditioning stimulus modifies inflammatory gene expression in humans. Physiol Genomics. 2004;19:143–150. doi: 10.1152/physiolgenomics.00046.2004. [DOI] [PubMed] [Google Scholar]
- 106.Saxena P, Newman MA, Shehatha JS, Redington AN, Konstantinov IE. Remote ischemic conditioning: evolution of the concept, mechanisms, and clinical application. J Card Surg. 2010;25:127–134. doi: 10.1111/j.1540-8191.2009.00820.x. [DOI] [PubMed] [Google Scholar]
- 107.Kharbanda RK, Mortensen UM, White PA, Kristiansen SB, Schmidt MR, Hoschtitzky JA, Vogel M, Sorensen K, Redington AN, MacAllister R. Transient limb ischemia induces remote ischemic preconditioning in vivo. Circulation. 2002;106:2881–2883. doi: 10.1161/01.cir.0000043806.51912.9b. [DOI] [PubMed] [Google Scholar]
- 108.Vinten-Johansen J, Shi W. Perconditioning and postconditioning: current knowledge, knowledge gaps, barriers to adoption, and future directions. J Cardiovasc Pharmacol Ther. 2011;16:260–266. doi: 10.1177/1074248411415270. [DOI] [PubMed] [Google Scholar]
- 109.Lim SY, Hausenloy DJ. Remote ischemic conditioning: from bench to bedside. Front Physiol. 2012;3:27. doi: 10.3389/fphys.2012.00027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Przyklenk K. Reduction of myocardial infarct size with ischemic “conditioning”: physiologic and technical considerations. Anesth Analg. 2013;117:891–901. doi: 10.1213/ANE.0b013e318294fc63. [DOI] [PubMed] [Google Scholar]
- 111.Weber C. Far from the heart: Receptor cross-talk in remote conditioning. Nat Med. 2010;16:760–762. doi: 10.1038/nm0710-760. [DOI] [PubMed] [Google Scholar]
- 112.Pickard JM, Davidson SM, Hausenloy DJ, Yellon DM. Co-dependence of the neural and humoral pathways in the mechanism of remote ischemic conditioning. Basic Res Cardiol. 2016;111:50. doi: 10.1007/s00395-016-0568-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Gotti C, Clementi F. Neuronal nicotinic receptors: from structure to pathology. Prog Neurobiol. 2004;74:363–396. doi: 10.1016/j.pneurobio.2004.09.006. [DOI] [PubMed] [Google Scholar]
- 114.Bibevski S, Zhou Y, McIntosh JM, Zigmond RE, Dunlap ME. Functional nicotinic acetylcholine receptors that mediate ganglionic transmission in cardiac parasympathetic neurons. J Neurosci. 2000;20:5076–5082. doi: 10.1523/JNEUROSCI.20-13-05076.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Gho BC, Schoemaker RG, van den Doel MA, Duncker DJ, Verdouw PD. Myocardial protection by brief ischemia in noncardiac tissue. Circulation. 1996;94:2193–2200. doi: 10.1161/01.cir.94.9.2193. [DOI] [PubMed] [Google Scholar]
- 116.Weinbrenner C, Nelles M, Herzog N, Sárváry L, Strasser RH. Remote preconditioning by infrarenal occlusion of the aorta protects the heart from infarction: a newly identified non-neuronal but PKC-dependent pathway. Cardiovasc Res. 2002;55:590–601. doi: 10.1016/s0008-6363(02)00446-7. [DOI] [PubMed] [Google Scholar]
- 117.Loukogeorgakis SP, Panagiotidou AT, Broadhead MW, Donald A, Deanfield JE, MacAllister RJ. Remote ischemic preconditioning provides early and late protection against endothelial ischemia-reperfusion injury in humans: role of the autonomic nervous system. J Am Coll Cardiol. 2005;46:450–456. doi: 10.1016/j.jacc.2005.04.044. [DOI] [PubMed] [Google Scholar]
- 118.Sabbah HN, Ilsar I, Zaretsky A, Rastogi S, Wang M, Gupta RC. Vagus nerve stimulation in experimental heart failure. Heart Fail Rev. 2011;16:171–178. doi: 10.1007/s10741-010-9209-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Zhang Y, Popovic ZB, Bibevski S, Fakhry I, Sica DA, Van Wagoner DR, Mazgalev TN. Chronic vagus nerve stimulation improves autonomic control and attenuates systemic inflammation and heart failure progression in a canine high-rate pacing model. Circ Heart Fail. 2009;2:692–699. doi: 10.1161/CIRCHEARTFAILURE.109.873968. [DOI] [PubMed] [Google Scholar]
- 120.Hamann JJ, Ruble SB, Stolen C, Wang M, Gupta RC, Rastogi S, Sabbah HN. Vagus nerve stimulation improves left ventricular function in a canine model of chronic heart failure. Eur J Heart Fail. 2013;15:1319–1326. doi: 10.1093/eurjhf/hft118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.De Ferrari GM, Crijns HJ, Borggrefe M, Milasinovic G, Smid J, Zabel M, Gavazzi A, Sanzo A, Dennert R, Kuschyk J, et al. Chronic vagus nerve stimulation: a new and promising therapeutic approach for chronic heart failure. Eur Heart J. 2011;32:847–855. doi: 10.1093/eurheartj/ehq391. [DOI] [PubMed] [Google Scholar]
- 122.Katare RG, Ando M, Kakinuma Y, Arikawa M, Handa T, Yamasaki F, Sato T. Vagal nerve stimulation prevents reperfusion injury through inhibition of opening of mitochondrial permeability transition pore independent of the bradycardiac effect. J Thorac Cardiovasc Surg. 2009;137:223–231. doi: 10.1016/j.jtcvs.2008.08.020. [DOI] [PubMed] [Google Scholar]
- 123.Calvillo L, Vanoli E, Andreoli E, Besana A, Omodeo E, Gnecchi M, Zerbi P, Vago G, Busca G, Schwartz PJ. Vagal stimulation, through its nicotinic action, limits infarct size and the inflammatory response to myocardial ischemia and reperfusion. J Cardiovasc Pharmacol. 2011;58:500–507. doi: 10.1097/FJC.0b013e31822b7204. [DOI] [PubMed] [Google Scholar]
- 124.Mioni C, Bazzani C, Giuliani D, Altavilla D, Leone S, Ferrari A, Minutoli L, Bitto A, Marini H, Zaffe D, et al. Activation of an efferent cholinergic pathway produces strong protection against myocardial ischemia/reperfusion injury in rats. Crit Care Med. 2005;33:2621–2628. doi: 10.1097/01.ccm.0000186762.05301.13. [DOI] [PubMed] [Google Scholar]
- 125.Mortara A, La Rovere MT, Pinna GD, Prpa A, Maestri R, Febo O, Pozzoli M, Opasich C, Tavazzi L. Arterial baroreflex modulation of heart rate in chronic heart failure: clinical and hemodynamic correlates and prognostic implications. Circulation. 1997;96:3450–3458. doi: 10.1161/01.cir.96.10.3450. [DOI] [PubMed] [Google Scholar]
- 126.Zoccali C, Mallamaci F, Parlongo S, Cutrupi S, Benedetto FA, Tripepi G, Bonanno G, Rapisarda F, Fatuzzo P, Seminara G, et al. Plasma norepinephrine predicts survival and incident cardiovascular events in patients with end-stage renal disease. Circulation. 2002;105:1354–1359. doi: 10.1161/hc1102.105261. [DOI] [PubMed] [Google Scholar]
- 127.Smith S, Rossignol P, Willis S, Zannad F, Mentz R, Pocock S, Bisognano J, Nadim Y, Geller N, Ruble S, et al. Neural modulation for hypertension and heart failure. Int J Cardiol. 2016;214:320–330. doi: 10.1016/j.ijcard.2016.03.078. [DOI] [PubMed] [Google Scholar]
- 128.Kishi T. Deep and future insights into neuromodulation therapies for heart failure. J Cardiol. 2016;68:368–372. doi: 10.1016/j.jjcc.2016.05.010. [DOI] [PubMed] [Google Scholar]
- 129.Manintveld OC, te Lintel Hekkert M, Keijzer E, Verdouw PD, Duncker DJ. Intravenous adenosine protects the myocardium primarily by activation of a neurogenic pathway. Br J Pharmacol. 2005;145:703–711. doi: 10.1038/sj.bjp.0706258. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Ding YF, Zhang MM, He RR. Role of renal nerve in cardioprotection provided by renal ischemic preconditioning in anesthetized rabbits. Sheng Li Xue Bao. 2001;53:7–12. [PubMed] [Google Scholar]
- 131.Ardell JL, Butler CK, Smith FM, Hopkins DA, Armour JA. Activity of in vivo atrial and ventricular neurons in chronically decentralized canine hearts. Am J Physiol. 1991;260:H713–H721. doi: 10.1152/ajpheart.1991.260.3.H713. [DOI] [PubMed] [Google Scholar]
- 132.Vander Heide RS, Schwartz LM, Jennings RB, Reimer KA. Effect of catecholamine depletion on myocardial infarct size in dogs: role of catecholamines in ischemic preconditioning. Cardiovasc Res. 1995;30:656–662. doi: 10.1016/0008-6363(95)00074-7. [DOI] [PubMed] [Google Scholar]
- 133.Mastitskaya S, Marina N, Gourine A, Gilbey MP, Spyer KM, Teschemacher AG, Kasparov S, Trapp S, Ackland GL, Gourine AV. Cardioprotection evoked by remote ischaemic preconditioning is critically dependent on the activity of vagal pre-ganglionic neurones. Cardiovasc Res. 2012;95:487–494. doi: 10.1093/cvr/cvs212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Lim SY, Yellon DM, Hausenloy DJ. The neural and humoral pathways in remote limb ischemic preconditioning. Basic Res Cardiol. 2010;105:651–655. doi: 10.1007/s00395-010-0099-y. [DOI] [PubMed] [Google Scholar]
- 135.Redington KL, Disenhouse T, Strantzas SC, Gladstone R, Wei C, Tropak MB, Dai X, Manlhiot C, Li J, Redington AN. Remote cardioprotection by direct peripheral nerve stimulation and topical capsaicin is mediated by circulating humoral factors. Basic Res Cardiol. 2012;107:241. doi: 10.1007/s00395-011-0241-5. [DOI] [PubMed] [Google Scholar]
- 136.Basalay M, Barsukevich V, Mastitskaya S, Mrochek A, Pernow J, Sjöquist PO, Ackland GL, Gourine AV, Gourine A. Remote ischaemic pre- and delayed postconditioning - similar degree of cardioprotection but distinct mechanisms. Exp Physiol. 2012;97:908–917. doi: 10.1113/expphysiol.2012.064923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Kingma JG, Simard D, Voisine P, Rouleau JR. Role of the autonomic nervous system in cardioprotection by remote preconditioning in isoflurane-anaesthetized dogs. Cardiovasc Res. 2011;89:384–391. doi: 10.1093/cvr/cvq306. [DOI] [PubMed] [Google Scholar]
- 138.Tsang A, Hausenloy DJ, Mocanu MM, Carr RD, Yellon DM. Preconditioning the diabetic heart: the importance of Akt phosphorylation. Diabetes. 2005;54:2360–2364. doi: 10.2337/diabetes.54.8.2360. [DOI] [PubMed] [Google Scholar]
- 139.Florea VG, Cohn JN. The autonomic nervous system and heart failure. Circ Res. 2014;114:1815–1826. doi: 10.1161/CIRCRESAHA.114.302589. [DOI] [PubMed] [Google Scholar]
- 140.Mudd JO, Kass DA. Tackling heart failure in the twenty-first century. Nature. 2008;451:919–928. doi: 10.1038/nature06798. [DOI] [PubMed] [Google Scholar]
- 141.Lymperopoulos A, Rengo G, Koch WJ. Adrenergic nervous system in heart failure: pathophysiology and therapy. Circ Res. 2013;113:739–753. doi: 10.1161/CIRCRESAHA.113.300308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 142.Arnold R, Issar T, Krishnan AV, Pussell BA. Neurological complications in chronic kidney disease. JRSM Cardiovasc Dis. 2016;5:2048004016677687. doi: 10.1177/2048004016677687. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Krishnan AV, Kiernan MC. Neurological complications of chronic kidney disease. Nat Rev Neurol. 2009;5:542–551. doi: 10.1038/nrneurol.2009.138. [DOI] [PubMed] [Google Scholar]
- 144.Kiernan MC, Walters RJ, Andersen KV, Taube D, Murray NM, Bostock H. Nerve excitability changes in chronic renal failure indicate membrane depolarization due to hyperkalaemia. Brain. 2002;125:1366–1378. doi: 10.1093/brain/awf123. [DOI] [PubMed] [Google Scholar]
- 145.Park J. Cardiovascular risk in chronic kidney disease: role of the sympathetic nervous system. Cardiol Res Pract. 2012;2012:319432. doi: 10.1155/2012/319432. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Grassi G. Sympathetic neural activity in hypertension and related diseases. Am J Hypertens. 2010;23:1052–1060. doi: 10.1038/ajh.2010.154. [DOI] [PubMed] [Google Scholar]
- 147.Grassi G, Quarti-Trevano F, Seravalle G, Arenare F, Volpe M, Furiani S, Dell’Oro R, Mancia G. Early sympathetic activation in the initial clinical stages of chronic renal failure. Hypertension. 2011;57:846–851. doi: 10.1161/HYPERTENSIONAHA.110.164780. [DOI] [PubMed] [Google Scholar]
- 148.Converse RL, Jacobsen TN, Toto RD, Jost CM, Cosentino F, Fouad-Tarazi F, Victor RG. Sympathetic overactivity in patients with chronic renal failure. N Engl J Med. 1992;327:1912–1918. doi: 10.1056/NEJM199212313272704. [DOI] [PubMed] [Google Scholar]
- 149.McClellan W, Aronoff SL, Bolton WK, Hood S, Lorber DL, Tang KL, Tse TF, Wasserman B, Leiserowitz M. The prevalence of anemia in patients with chronic kidney disease. Curr Med Res Opin. 2004;20:1501–1510. doi: 10.1185/030079904X2763. [DOI] [PubMed] [Google Scholar]
- 150.Bosman DR, Winkler AS, Marsden JT, Macdougall IC, Watkins PJ. Anemia with erythropoietin deficiency occurs early in diabetic nephropathy. Diabetes Care. 2001;24:495–499. doi: 10.2337/diacare.24.3.495. [DOI] [PubMed] [Google Scholar]
- 151.DiBona GF, Kopp UC. Neural control of renal function. Physiol Rev. 1997;77:75–197. doi: 10.1152/physrev.1997.77.1.75. [DOI] [PubMed] [Google Scholar]
- 152.DiBona GF. Nervous kidney. Interaction between renal sympathetic nerves and the renin-angiotensin system in the control of renal function. Hypertension. 2000;36:1083–1088. doi: 10.1161/01.hyp.36.6.1083. [DOI] [PubMed] [Google Scholar]
- 153.Jassal SV, Coulshed SJ, Douglas JF, Stout RW. Autonomic neuropathy predisposing to arrhythmias in hemodialysis patients. Am J Kidney Dis. 1997;30:219–223. doi: 10.1016/s0272-6386(97)90055-x. [DOI] [PubMed] [Google Scholar]
- 154.Henderson LW. Symptomatic hypotension during hemodialysis. Kidney Int. 1980;17:571–576. doi: 10.1038/ki.1980.67. [DOI] [PubMed] [Google Scholar]
- 155.Lefrandt JD, Smit AJ, Zeebregts CJ, Gans RO, Hoogenberg KH. Autonomic dysfunction in diabetes: a consequence of cardiovascular damage. Curr Diabetes Rev. 2010;6:348–358. doi: 10.2174/157339910793499128. [DOI] [PubMed] [Google Scholar]
- 156.Sartori C, Scherrer U. Insulin, nitric oxide and the sympathetic nervous system: at the crossroads of metabolic and cardiovascular regulation. J Hypertens. 1999;17:1517–1525. doi: 10.1097/00004872-199917110-00003. [DOI] [PubMed] [Google Scholar]
- 157.Steinberg HO, Brechtel G, Johnson A, Fineberg N, Baron AD. Insulin-mediated skeletal muscle vasodilation is nitric oxide dependent. A novel action of insulin to increase nitric oxide release. J Clin Invest. 1994;94:1172–1179. doi: 10.1172/JCI117433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 158.Sartori C, Trueb L, Nicod P, Scherrer U. Effects of sympathectomy and nitric oxide synthase inhibition on vascular actions of insulin in humans. Hypertension. 1999;34:586–589. doi: 10.1161/01.hyp.34.4.586. [DOI] [PubMed] [Google Scholar]
- 159.Gill GV, Woodward A, Casson IF, Weston PJ. Cardiac arrhythmia and nocturnal hypoglycaemia in type 1 diabetes--the ‘dead in bed’ syndrome revisited. Diabetologia. 2009;52:42–45. doi: 10.1007/s00125-008-1177-7. [DOI] [PubMed] [Google Scholar]
- 160.Deckert T, Feldt-Rasmussen B, Borch-Johnsen K, Jensen T, Kofoed-Enevoldsen A. Albuminuria reflects widespread vascular damage. The Steno hypothesis. Diabetologia. 1989;32:219–226. doi: 10.1007/BF00285287. [DOI] [PubMed] [Google Scholar]
- 161.Jensen T, Bjerre-Knudsen J, Feldt-Rasmussen B, Deckert T. Features of endothelial dysfunction in early diabetic nephropathy. Lancet. 1989;1:461–463. doi: 10.1016/s0140-6736(89)91365-2. [DOI] [PubMed] [Google Scholar]
- 162.Pavy-Le Traon A, Fontaine S, Tap G, Guidolin B, Senard JM, Hanaire H. Cardiovascular autonomic neuropathy and other complications in type 1 diabetes. Clin Auton Res. 2010;20:153–160. doi: 10.1007/s10286-010-0062-x. [DOI] [PubMed] [Google Scholar]
- 163.Beijers HJ, Ferreira I, Bravenboer B, Dekker JM, Nijpels G, Heine RJ, Stehouwer CD. Microalbuminuria and cardiovascular autonomic dysfunction are independently associated with cardiovascular mortality: evidence for distinct pathways: the Hoorn Study. Diabetes Care. 2009;32:1698–1703. doi: 10.2337/dc08-1544. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Gerritsen J, Dekker JM, TenVoorde BJ, Kostense PJ, Heine RJ, Bouter LM, Heethaar RM, Stehouwer CD. Impaired autonomic function is associated with increased mortality, especially in subjects with diabetes, hypertension, or a history of cardiovascular disease: the Hoorn Study. Diabetes Care. 2001;24:1793–1798. doi: 10.2337/diacare.24.10.1793. [DOI] [PubMed] [Google Scholar]
- 165.Pop-Busui R, Evans GW, Gerstein HC, Fonseca V, Fleg JL, Hoogwerf BJ, Genuth S, Grimm RH, Corson MA, Prineas R. Effects of cardiac autonomic dysfunction on mortality risk in the Action to Control Cardiovascular Risk in Diabetes (ACCORD) trial. Diabetes Care. 2010;33:1578–1584. doi: 10.2337/dc10-0125. [DOI] [PMC free article] [PubMed] [Google Scholar]