

Abstract
DNA relaxation is an important step in DNA replication. DNA topoisomerases play a major role in DNA relaxation. Hence these enzymes are important targets for cancer drugs. DNA topoisomerase inhibitors bind to the transient enzyme–DNA complex and inhibit DNA replication. Various inhibitors of topoisomerase I and II are prescribed as drugs. Topoisomerase II is considered as an important target for the development of anticancer drugs. In this study we have demonstrated molecular docking of thirteen fluoroquinolines with human DNA topoisomerase II alpha (a) and beta (b). Fluoroquinolines are broad spectrum antibacterial antibiotics and it is highly effective against various bacterial infections. Some of the fluoroquinolines like moxifloxacin exert antifungal as well as anti-cancer activity. It forms complexes with topoisomerase II a and are responsible for stoppage DNA replication. Molecular docking studies showed that fluoroquinolines has shown formation of hydrogen bond and good binding affinity with human Topo2a and Topo2b. Hence FQs may inhibit the activity of enzyme topoisomerase by binding at its active site. Ofloxacin, sparafloxacin, ciprofloxacin and moxifloxacin are predicted to be the most potent inhibitors among the thirteen FQs docked. GLN773, ASN770, LYS723 and TRP931 amino acid residues of Topo2a are involved in binding with FQs while ASP479, SER480, ARG820, ARG503, LYS456 and GLN778 amino acid residues of Topo2b are involved in binding with FQs. Our in silico study suggests that fluoroquinolines could be repositioned as DNA topoisomerase II inhibitors hence can be used as anticancer drugs. In vitro and in vivo experiments need to be done to confirm their efficacy.
Keywords: Human DNA topoisomerase II, Fluoroquinolines, Anticancer, Molecular docking, In silico, Drug repositioning
Introduction
DNA topological states depend upon the activity of topoisomerase I and II (Wang et al. 2002). Their activity is important for cellular activities like replication, recombination, transcription and DNA repair (Bertozzi et al. 2011, 2014; Marinello et al. 2013). Topoisomerase II is present in two isoforms Top2a and Topo2b in mammals (Chung et al. 1989; Drake et al. 1987). Top2a is actively involved in replicating cells while Top2b is constitutively expressed in specific cellular processes like neural development (Yang et al. 2000; Tsutsui et al. 2001). Top2a introduce double-strand nick into DNA during DNA replication. Top2 enzymes are important targets for several antitumour drugs (Nitiss 2009). Anticancer drugs inhibit the activity of DNA Topo2 enzyme which may responsible for cell death (Pommier 2013; Azarova et al. 2007). Although these drugs are having some side effects but still they are prescribed as anticancer drugs (Nitiss 2009). These drugs may act on Topo2b in normal cells in human body (Azarova et al. 2007; Bromberg et al. 2003; Zhang et al. 2012). Therefore screening of Topo2a inhibitors is an important task for drug developers. The exploration of differences in structure and function between the two Topo2 isoforms is important, and the identification Topo2a inhibitors from available and already approved drugs may be a beneficial strategy in the search for new cancer drugs with improved clinical safety. The combination of structure- and ligand-based computational methods has been successfully applied in many drug design projects (Drwal et al. 2013). Pharmacophores, representing the spatial arrangement of chemical features necessary for binding of a small molecule to its biological target, can be developed in different ways, depending on the available information.
The broad spectrum antibacterial antibiotics, fluoroquinolines are highly effective against various bacterial infections. Quinolone-induced inactivation of Topo II· at high concentrations is proposed to involve direct binding of quinolones to DNA (Bromberg et al. 2003). Moxifloxacin inhibited purified human topoisomerase II and acted synergistically with VP-16 at the purified enzyme level and enhanced its anti-proliferative effects in THP-1 and Jurkat cells (Fabian et al. 2006). In earlier studies we have reported that Moxifloxacin can inhibit growth and virulence factors of Candida albicans. Molecular docking of Candida DNA topoisomerase II with FQs was showed good binding affinity (Jadhav et al. 2017). In this study we are reporting molecular interactions of thirteen fluoroquinolines with human DNA topoisomerase II a and b by using molecular docking studies.
Materials and methods
Protein structure preparation
Human DNA topoisomerase II alpha (PDB ID: 4fm9) and beta (PDB ID: 3QX3) were retrieved from PDB (http://www.rcsb.org/pdb/). Autodock Tools version 1.5.6rc2 was used to generate the docking input files. Using AutoDock Tools (ADT) 1.5.6rc2 nonpolar hydrogens were merged, Gastegier charges were assigned saved in PDBQT file format. Grid and docking parameter files were prepared using ADT and molecular docking studies were performed with AutoDock 1.5.6rc2 considering all the bonds of ligands as rotatable and macromolecules as rigid (Morris et al. 1998). Grid box size of 60 × 60 × 60 Å with 0.375 Å spacing centered at the site of DNA cleavage of topo–DNA complexes was selected. Hetero-atoms and drug molecules were removed, Gasteiger charges were assigned and both the macromolecules were saved in PDBQT file format using Autodock tool (Kumar and Bora 2014).
Ligand structure preparation
The structures Fluoroquinolines were retrieved from Pubchem (http://pubchem.ncbi.nlm.nih.gov/) in SDF format. Structure followed by 2D cleaning, 3D optimization and viewing is done by using MarwinView and saved in Mol2 file format. Mol2 files was then converted to PDBQT file format by in Autodock Tools version 1.5.6rc2 (Kumar and Bora 2014).
Molecular docking
Molecular docking study of Fluoroquinolines against Human Topoisomerase II was done using AutoDock1.5.6rc2® suite as molecular-docking tool (Kumar and Bora 2014; Morris et al. 1998). Empirical-free energy function and Lamarckian Genetic Algorithm, with an initial population of 150 randomly placed individuals, a maximum number of 2,500,000 energy evaluations, a mutation rate of 0.02 and a crossover rate of 0.80 were used. AutoDock1.5.6rc2® tools generated sixty possible binding conformations, i.e. sixty runs for each docking was done using genetic algorithm (GALS) searches. Autogrid was used to obtain pre calculated grid maps (Kumar and Bora 2014).
Molecular docking analysis
After completion of docking most suitable conformation was chosen to prepare Ligand–protein complex preparation using AutoDock 1.5.6rc2. Docked structures saved in the PDBQT format were analyzed for the amino acids involved in the ligand binding sites of the enzyme along with the type of interactions like hydrogen bonding etc. involved in the docking. This was performed using the Discovery Studio 4.1 Visualizer (http://accelrys.com/products/discovery-studio). 13 Docked structures showing the amino acid residues involved in the docking as well as the interactions between the residues and ligand were image captured using image save option in Discovery Studio 4.1 Visualizer (Kumar and Bora 2014).
Results
Fluoroquinolines docked with human DNA topoisomerase II alpha
Molecular docking of thirteen fluoroquinolines with human topoisomerase II a (4fm9) showed that each FQs formed hydrogen bond with the active site of topoisomerase II (Figs. 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13). Ciprofloxacin and enoxacin were found to bind to human topoisomerase II at GLN773 with binding energies −6.54 and −6.06 kcal/mol. Similarly gatifloxacin, lomefloxacin, moxifloxacin, norfloxacin and sarafloxacin formed hydrogen bonds with GLN773 with binding energies −6.86, −6.84, −7.7, −6.6, −7.23 kcal/mol respectively (Figs. 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13).
Fig. 1.
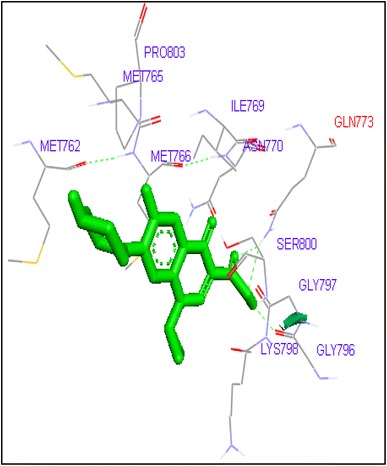
Binding of ciprofloxacin with human topoisomerase II a
Fig. 2.
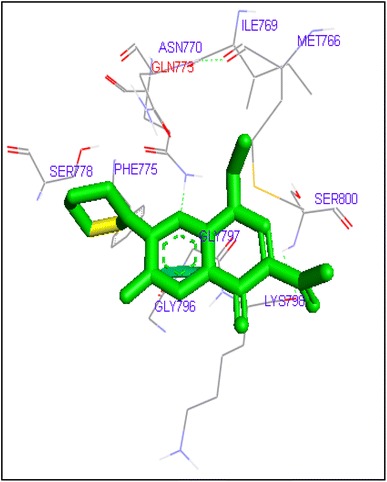
Binding of enoxacin with human topoisomerase II a
Fig. 3.
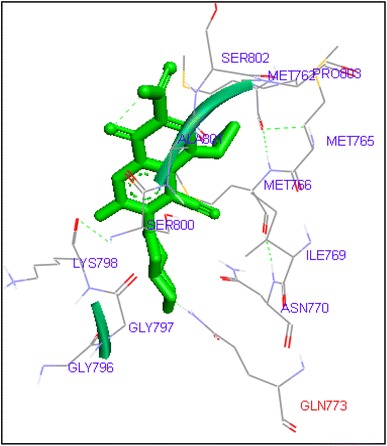
Binding of finafloxacin with human topoisomerase II a
Fig. 4.
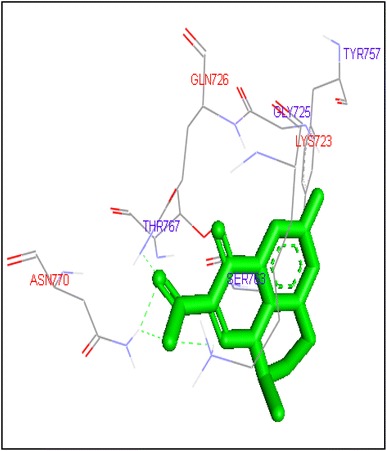
Binding of flumequine with human topoisomerase II a
Fig. 5.
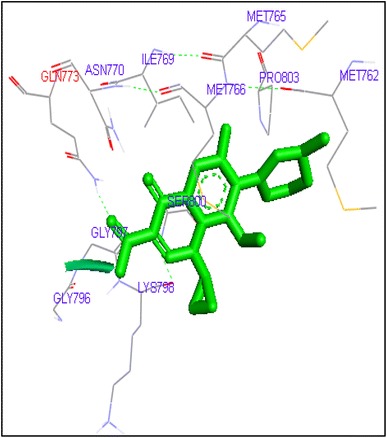
Binding of gatifloxacin with human topoisomerase II a
Fig. 6.
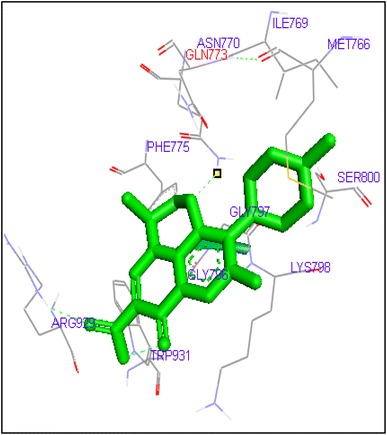
Binding of levofloxacin with human topoisomerase II a
Fig. 7.
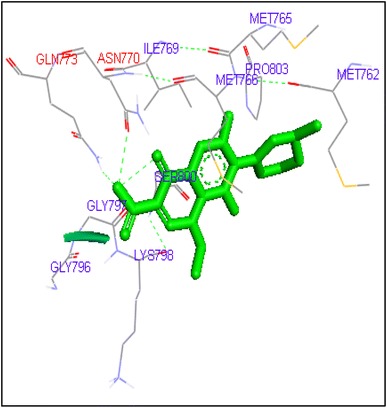
Binding of lomefloxacin with human topoisomerase II a
Fig. 8.
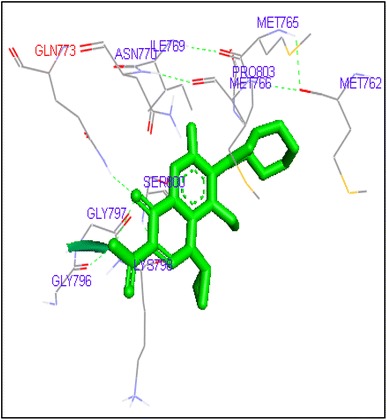
Binding of moxifloxacin with human topoisomerase II a
Fig. 9.
Binding of norfloxacin with human topoisomerase II a
Fig. 10.
Binding of ofloxacin with human topoisomerase II a
Fig. 11.
Binding of prulifloxacin with human topoisomerase II a
Fig. 12.
Binding of sarafloxacin with human topoisomerase II a
Fig. 13.
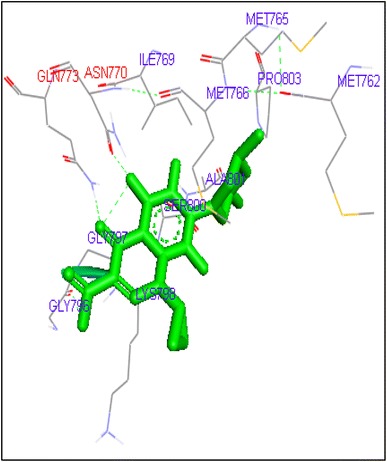
Binding of sparafloxacin with human topoisomerase II a
Ciprofloxacin exhibited binding at the active site of human topoisomerase II b at ASP479 amino acid residue with −9.62 and −10.78 kcal/mol binding affinities. Finafloxacin and flumequine formed hydrogen bond with ARG820 amino acid residue with binding energies −8.92 and −8.79 kcal/mol respectively (Figs. 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13).
Gatifloxacin and moxifloxacin formed H bond with ARG503 and LYS 456 amino acid residue and binding energies were −10.73 and −9.95 kcal/mol respectively. Prulifloxacin was formed H and with SER480 residue and binding energy was −10.77 kcal/mol while sparafloxacin bound at GLN778 and ARG820 of Topo2b with −11.43 kcal/mol binding score (Table 1).
Table 1.
Molecular interactions of fluoroquinolines with human DNA topoisomerase II (4fm9)
| Ligand | Run no. | Interacting residue in human Topo II a | Interacting atoms (amino acid……ligand) | H bonds formed | Binding energy | Electrostatic energy |
|---|---|---|---|---|---|---|
| Ciprofloxacin | 51 | GLN773 | HE22……O3 | 1 | −6.54 | −0.07 |
| Enoxacin | 37 | GLN773 | HE22……O4 | 1 | −6.06 | 0.05 |
| Finafloxacin | 43 | ASN770 | OD1……O4 | 1 | −6.41 | −0.02 |
| GLN773 | HE22……O4 | |||||
| Flumequine | 4 | GLN726 | HE21……O2 | 3 | −6.15 | −0.55 |
| ASN770 | HD21……O2 | |||||
| LYS723 | HZ2……O3 | |||||
| Gatifloxacin | 58 | GLN773 | HE22……O4 | 1 | −6.86 | −0.04 |
| Levofloxacin | 46 | TRP931 | HE1……O3 | 2 | −8.06 | −0.38 |
| Lomefloxacin | 1 | GLN773 | HE22……O2 | 1 | −6.84 | −0.0 |
| Moxifloxacin | 12 | GLN773 | HE22……O2 | 1 | −7.7 | −0.09 |
| Norfloxacin | 22 | GLN773 | HE22……O2 | 2 | −6.6 | −0.01 |
| Ofloxacin | 48 | GLN773 | HE22……O1 | 2 | −8.05 | −0.36 |
| TRP931 | HE1……O3 | |||||
| Prulifloxacin | 22 | TRP931 | HE1……O4 | 1 | −8.0 | 0.3 |
| Sarafloxacin | 38 | GLN773 | HE22……O2 | 1 | −7.23 | 0.01 |
| Sparafloxacin | 49 | GLN773 | HE22……O1 | 2 | −8.17 | −0.05 |
| ASN770 | HHOD1……N4 | |||||
| Etoposide | 56 | SER800 | O……H26 | 3 | −7.9 | −0.15 |
| GLY796 | O……H20 | |||||
| GLN773 | HE22……O8 |
Fluoroquinolines showed efficient binding with human DNA topoisomerase II beta
Molecular docking of thirteen fluoroquinolines with human topoisomerase II b (3Q3X) revealed that there was significant docking between ligand and active site of topoisomerase II b protein (Figs. 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26). Enoxacin and levofloxacin were found to bind with human topoisomerase II b at ASP479 and SER480 residues with binding energy −9.96 and −11.78 kcal/mol. Similarly norfloxacin, ofloxacin, lomefloxacin formed hydrogen bonds with ASP479 and SER480 with binding energies −10.43, −11.8 and −10.35 kcal/mol respectively (Figs. 14, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26).
Fig. 14.
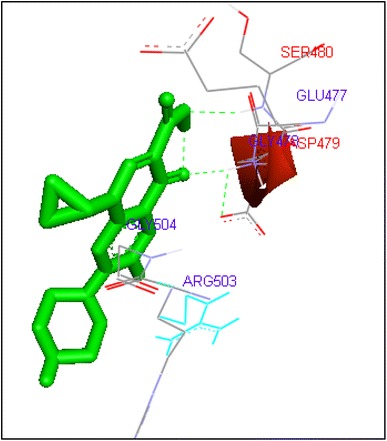
Binding of ciprofloxacin with human topoisomerase II b
Fig. 15.
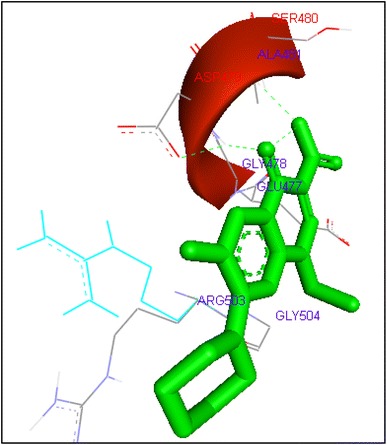
Binding of enoxacin with human topoisomerase II b
Fig. 16.
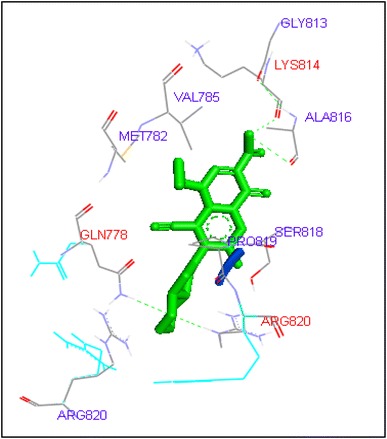
Binding of finafloxacin with human topoisomerase II b
Fig. 17.
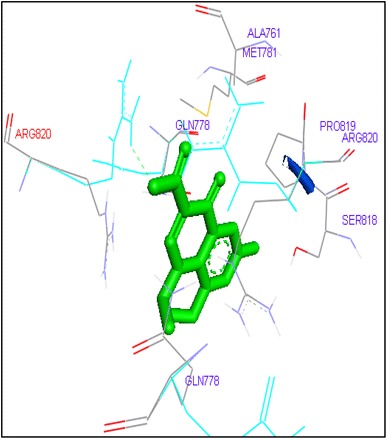
Binding of flumefoxacin with human topoisomerase II b
Fig. 18.
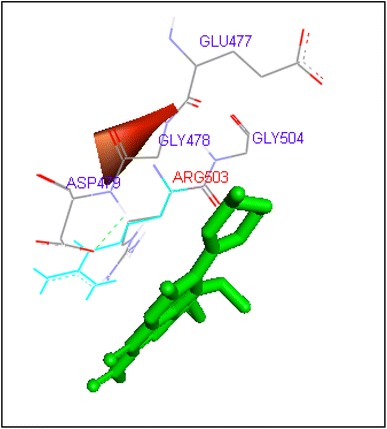
Binding of gatifloxacin with human topoisomerase II b
Fig. 19.
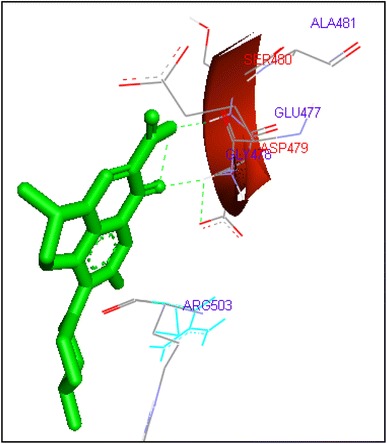
Binding of levofloxacin with human topoisomerase II b
Fig. 20.
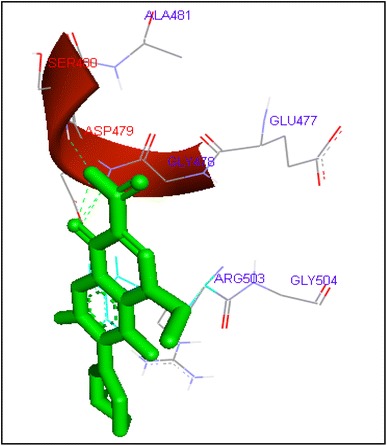
Binding of lomefloxacin with human topoisomerase II b
Fig. 21.
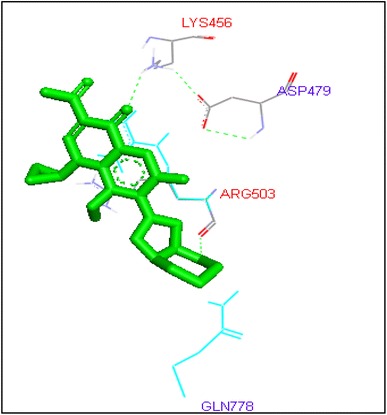
Binding of moxifloxacin with human topoisomerase II b
Fig. 22.
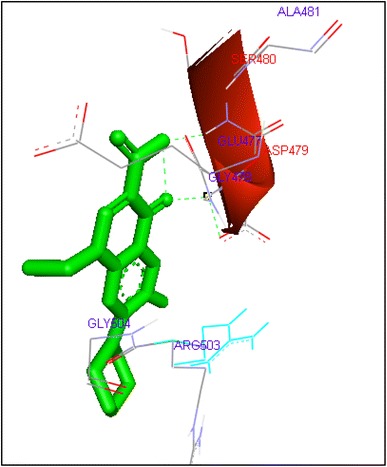
Binding of norfloxacin with human topoisomerase II b
Fig. 23.
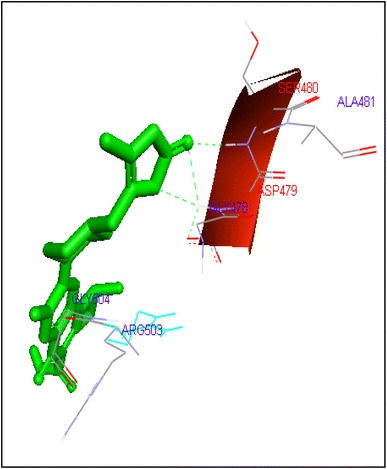
Binding of prulifloxacin with human topoisomerase II b
Fig. 24.
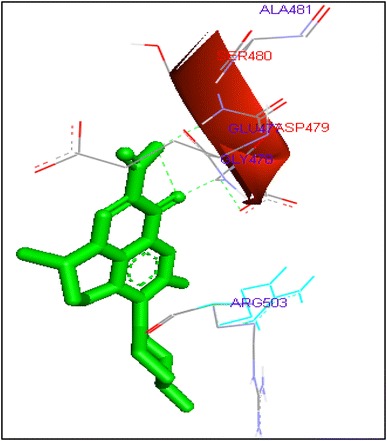
Binding of ofloxacin with human topoisomerase II b
Fig. 25.
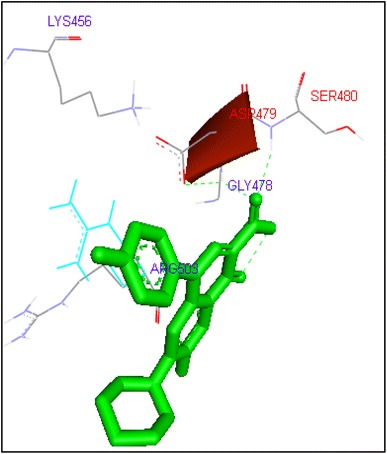
Binding of sarafloxacin with human topoisomerase II b
Fig. 26.
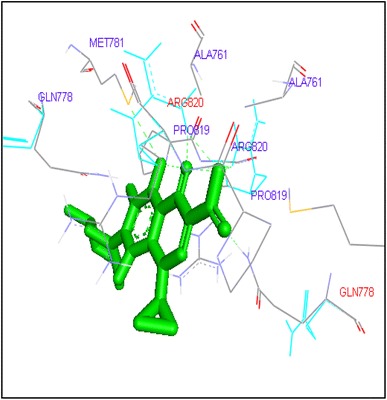
Binding of sparafloxacin with human topoisomerase II b
Ciprofloxacin and sarafloxacin exhibited binding at the active site of human Topoisomerase II b at ASP479 amino acid residue with −9.62 and −10.78 kcal/mol binding affinities. Finafloxacin and flumequine were observed to bind with ARG820 residue and binding energy was −8.92 and −8.79 kcal/mol (Figs. 14, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26).
Gatifloxacin and moxifloxacin docked with ARG503 and LYS 456 amino acid residue and binding free energies were −10.73 and −9.95 kcal/mol respectively. Prulifloxacin was formed H and at SER480 residue and −10.77 kcal/mol binding energy while sparafloxacin binds at GLN778 and ARG820 of Topo2b with −11.43 kcal/mol binding score (Table 2).
Table 2.
Molecular interactions of fluoroquinolines with human DNA topoisomerase II b (3QX3)
| Ligand | Run no. | Interacting residue in human Topo II b | Interacting atoms (amino acid……ligand) | H bonds formed | Binding energy | Electrostatic energy |
|---|---|---|---|---|---|---|
| Ciprofloxacin | 54 | ASP479 | HN……O1 | 1 | −9.62 | 0.11 |
| Enoxacin | 32 | ASP479 | HN……O1 | 2 | −9.96 | 0.1 |
| SER480 | HN……O2 | |||||
| Finafloxacin | 44 | ARG 820 | HE……O4 | 1 | −8.92 | −0.15 |
| Flumequine | 41 | ARG820 | HH12……O3 | 2 | −8.79 | −0.76 |
| ARG820 | HH22……O3 | |||||
| Gatifloxacin | 15 | ARG503 | HH12……O3 | 1 | −10.73 | −0.3 |
| Levofloxacin | 59 | ASP479 | HN……O2 | 2 | −11.78 | −0.18 |
| SER480 | HN……O3 | |||||
| Lomefloxacin | 54 | ASP479 | HN……O1 | 1 | −10.35 | −0.1 |
| SER80 | HN……O2 | 1 | ||||
| Moxifloxacin | 17 | LYS456 | HZ1……O2 | 1 | −9.95 | −0.24 |
| Norfloxacin | 29 | ASP479 | HN……O1 | 2 | −10.43 | −0.16 |
| SER480 | HN……O2 | |||||
| Ofloxacin | 16 | ASP479 | HN……O2 | 2 | −11.8 | 0.18 |
| SER480 | HN……O3 | |||||
| Prulifloxacin | 47 | SER480 | HN……O6 | 1 | −10.77 | 0.07 |
| Sarafloxacin | 56 | ASP479 | HN……O2 | 1 | −10.78 | 0.01 |
| Sparafloxacin | 39 | GLN778 | HE22……O3 | 2 | −11.43 | −0.83 |
| ARG820 | HH12……O2 | |||||
| Etoposide | 37 | DG10 | H19……OP2 | −9.38 | −0.51 | |
| DA12 | H12……O11 | |||||
| DG10 | H10……O5 | |||||
| DC11 | H11……O5,O2 |
Discussion
Human topo2 is the target of several anticancer agents (Nitiss 2009) which includes doxorubicin, amascrine, mitoxantrone and etoposide (Shen et al. 1989). Our molecular docking studies revealed that fluoroquinoline antibiotics are showing molecular interactions with human topoisomerase II (Figs. 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26).
FQs are the broad spectrum antibiotics and inhibit the activity of topoisomerase II (Shen et al. 1989; Aldred et al. 2014). Fluoroquinolones and several classes of anti-cancer agents affect type II a topoisomerase activity and function (Aldred et al. 2014; Chen et al. 2013; Chan et al. 2013; Robinson et al. 1991).
Docking of Etoposide showed interactions with GLN778, SER800 and GLY796 residues in Top2a and DG10, DG11, DG12 in Topo2b complex (Figs. 27, 28). In this study we found that FQs majorly formed hydrogen bond with GLN773, ASN770, LYS723 and TRP931 in human DNA Topo2a and ASP479, SER480, ARG820, ARG503, LYS456 and GLN778 in binding pocket of human DNA Topo2b. Out of thirteen FQs sparafloxacin has shown lowest binding energy −8.17 kcal/mol with Topo2a. Ofloxacin showed binding energy of −11.8 kcal/mol with Topo2b. Anticancer activity of norfloxacin, ofloxacin, prulifloxacin, sarafloxacin, finafloxacin and flumequine are not reported. These FQs are showing good binding score in molecular docking studies with Topo2 enzyme and there is need for in vitro and in vivo studies of these FQs. There is a possibility for repositioning of these antibiotics as anticancer agents.
Fig. 27.
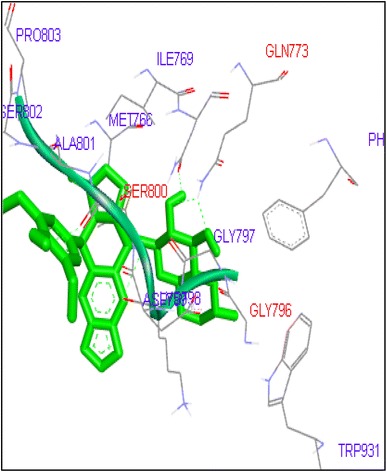
Binding of etoposide with human topoisomerase II a
Fig. 28.
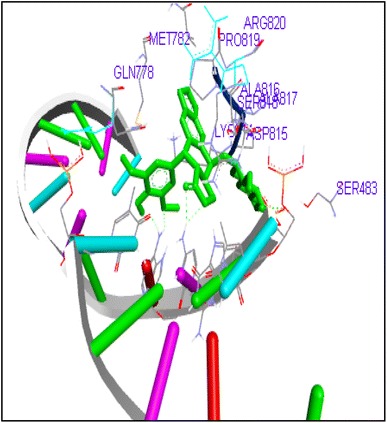
Binding of etoposide to human topoisomerase II b
Conclusion
In this study we have explored topoisomerase II as an effective target for FQs by in silico studies. All thirteen FQs have shown significant binding affinity with both human topoisomerase II a and b. FQs formed significant molecular interactions with topoisomerase II at active sites. FQs may form a complex with DNA and topoisomerase II, and cause inhibition of the activity of Topoisomerase leading to the inhibition of growth. As FQS are showing promising anticancer activity against various cancer cell lines, in vitro and in vivo experiments are necessary to put forward therapeutic repositioning of FQs as anticancer agents.
Acknowledgements
AKJ and SMK are thankful to Prof. Pandit Vidyasagar, Vice Chancellor, SRTM University, Nanded for his kind support. SMK is also thankful to UGC, New Delhi for SAP DRS II Program for infrastructural support.
References
- Aldred KJ, Kerns RJ, Osheroff N. Mechanism of quinolone action and resistance. Biochemistry. 2014;53:1565–1574. doi: 10.1021/bi5000564. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Azarova AM, Lyu YL, Lin CP, Tsai YC, Lau JYN, Wang JC, Liu LF. Roles of DNA topoisomerase II isozymes in chemotherapy and secondary malignancies. Proc Natal Acad Sci USA. 2007;104(26):11014–11019. doi: 10.1073/pnas.0704002104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bertozzi D, Iurlaro R, Sordet O, Marinello J, Zaffaroni N, Capranico G. Characterization of novel antisense HIF-1α transcripts in human cancers. Cell Cycle. 2011;10(18):3189–3197. doi: 10.4161/cc.10.18.17183. [DOI] [PubMed] [Google Scholar]
- Bertozzi D, Marinello J, Manzo SG, Fornari F, Gramantieri L, Capranico G. The natural inhibitor of DNA topoisomerase I, camptothecin, modulates HIF-1α activity by changing miR expression patterns in human cancer cells. Mol Cancer Ther. 2014;13(1):239–248. doi: 10.1158/1535-7163.MCT-13-0729. [DOI] [PubMed] [Google Scholar]
- Bromberg KD, Burgin AB, Osheroff N. Quinolone action against human topoisomerase II alpha: stimulation of enzyme mediated double-stranded DNA cleavage. Biochemistry. 2003;42:3393–3398. doi: 10.1021/bi027383t. [DOI] [PubMed] [Google Scholar]
- Bromberg KD, Burgin AB, Osheroff N. Quinolone action against human topoisomerase II alpha: stimulation of enzyme mediated double-stranded DNA cleavage. Biochemistry. 2003;42:3393–3398. doi: 10.1021/bi027383t. [DOI] [PubMed] [Google Scholar]
- Chan PF, Huang J, Bax BD, Gwynn MN. Recent developments in inhibitors of bacterial type IIA topoisomerases. In: Gualerzi VO, Brandi L, Fabbretti A, Pon CL, editors. Antibiotics: targets, mechanisms and resistance. Weinheim: Wiley; 2013. pp. 263–297. [Google Scholar]
- Chen SH, Chan NL, Hsieh TS. New mechanistic and functional insights into DNA topoisomerases. Annu Rev Biochem. 2013;82:139–170. doi: 10.1146/annurev-biochem-061809-100002. [DOI] [PubMed] [Google Scholar]
- Chung TD, Drake FH, Tan KB, Per SR, Crooke ST, Mirabelli CK. Characterization and immunological identification of cDNA clones encoding two human DNA topoisomerase II isozymes. Proc Natl Acad Sci. 1989;86(23):9431–9435. doi: 10.1073/pnas.86.23.9431. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Drake FH, Zimmerman JP, McCabe FL, Bartus HF, Per SR, Sullivan DM, Crooke ST. Purification of topoisomerase II from amsacrine-resistant P388 leukemia cells. Evidence for two forms of the enzyme. J Biol Chem. 1987;262(34):16739–16747. [PubMed] [Google Scholar]
- Drwal MN, Agama K, Pommier Y, Griffith R. Development of purely structure-based pharmacophores for the topoisomerase I-DNA-ligand binding pocket. J Comput Aided Mol Des. 2013;27(12):1037–1049. doi: 10.1007/s10822-013-9695-x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fabian I, Reuveni D, Levitov A, Halperin D, Priel E, Shalit I. Moxifloxacin enhances antiproliferative and apoptotic effects of etoposide but inhibits its proinflammatory effects in THP-1 and Jurkat cells. Br J Cancer. 2006;95(8):1038–1046. doi: 10.1038/sj.bjc.6603355. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jadhav A, Bansode B, Phule D, Shelar A, Patil R, Gade W, Karuppayil SM. The antibacterial agent, moxifloxacin inhibits virulence factors of Candida albicans through multitargeting. World J Microbiol Biotechnol. 2017;33(5):96. doi: 10.1007/s11274-017-2264-z. [DOI] [PubMed] [Google Scholar]
- Kumar A, Bora U. Molecular docking studies of curcumin natural derivatives with DNA topoisomerase I and II-DNA complexes. Interdiscip Sci Comput Life Sci. 2014;6(4):285–291. doi: 10.1007/s12539-012-0048-6. [DOI] [PubMed] [Google Scholar]
- Marinello J, Chillemi G, Bueno S, Manzo SG, Capranico G (2013) Antisense transcripts and R-loops caused by DNA topoisomerase I inhibition by camptothecin at human active CpG island promoters. Cancer Res 73(8 Supplement). doi:10.1158/1538-7445.AM2013-637 [DOI] [PMC free article] [PubMed]
- Morris GM, Goodsell DS, Halliday RS, Huey R, Hart WE, Belew RK, Olson AJ. Automated docking using a Lamarckian genetic algorithm and an empirical binding free energy function. J Comput Chem. 1998;19(14):1639–1662. doi: 10.1002/(SICI)1096-987X(19981115)19:14<1639::AID-JCC10>3.0.CO;2-B. [DOI] [Google Scholar]
- Nitiss JL. Targeting DNA topoisomerase II in cancer chemotherapy. Nat Rev Cancer. 2009;9:338–350. doi: 10.1038/nrc2607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pommier Y. Drugging topoisomerases: lessons and challenges. ACS Chem Biol. 2013;8:82–95. doi: 10.1021/cb300648v. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Robinson MJ, Martin BA, Gootz TD, et al. Effects of quinolone derivatives on eukaryotic topoisomerase-II. A novel mechanism for enhancement of enzyme-mediated DNA cleavage. J Biol Chem. 1991;266:14585–14592. [PubMed] [Google Scholar]
- Shen LL, Baranowski J, Pernet AG. Mechanism of inhibition of DNA gyrase by quinolone antibacterials: specificity and cooperativity of drug binding to DNA. Biochemistry. 1989;28:3879–3885. doi: 10.1021/bi00435a038. [DOI] [PubMed] [Google Scholar]
- Tsutsui K, Tsutsui K, Sano K, Kikuchi A, Tokunaga A. Involvement of DNA topoisomerase IIβ in neuronal differentiation. J Biol Chem. 2001;276(8):5769–5778. doi: 10.1074/jbc.M008517200. [DOI] [PubMed] [Google Scholar]
- Wang S, Miller W, Milton J, Vicker N, Stewart A, Charlton P, Denny WA. Structure–activity relationships for analogues of the phenazine-based dual topoisomerase I/II inhibitor XR11576. Bioorg Med Chem Lett. 2002;12(3):415–418. doi: 10.1016/S0960-894X(01)00770-3. [DOI] [PubMed] [Google Scholar]
- Yang X, Li W, Prescott ED, Burden SJ, Wang JC. DNA topoisomerase IIβ and neural development. Science. 2000;287(5450):131–134. doi: 10.1126/science.287.5450.131. [DOI] [PubMed] [Google Scholar]
- Zhang S, Liu X, Bawa-Khalfe T, Lu LS, Lyu YL, Liu LF, Yeh ET. Identification of the molecular basis of doxorubicin-induced cardiotoxicity. Nat Med. 2012;18(11):1639–1642. doi: 10.1038/nm.2919. [DOI] [PubMed] [Google Scholar]