

Abstract
Dendritic cells (DCs) are important immune cells linking innate and adaptive immune responses. DCs encounter various self and non-self antigens present in the environment and induce different types of antigen specific adaptive immune responses. DCs can be classified into lymphoid tissue-resident DCs, migratory DCs, non-lymphoid resident DCs, and monocyte derived DCs (moDCs). Recent work has also established that DCs consist of developmentally and functionally distinct subsets that differentially regulate T lymphocyte function. The development of different DC subsets has been found to be regulated by a network of different cytokines and transcriptional factors. Moreover, the response of DC is tightly regulated to maintain the homeostasis of immune system. MicroRNAs (miRNAs) are an important class of cellular regulators that modulate gene expression and thereby influence cell fate and function. In the immune system, miRNAs act at checkpoints during hematopoietic development and cell subset differentiation, they modulate effector cell function, and are implicated in the maintenance of homeostasis. DCs are also regulated by miRNAs. In the past decade, much progress has been made to understand the role of miRNAs in regulating the development and function of DCs. In this review, we summarize the origin and distribution of different mouse DC subsets in both lymphoid and non-lymphoid tissues. The DC subsets identified in human are also described. Recent progress on the function of miRNAs in the development and activation of DCs and their functional relevance to autoimmune diseases are discussed.
Keywords: dendritic cell, mirna, activation, development, autoimmune disease
INTRODUCTION
DCs were discovered by Ralph Steinman and Zanvil Cohn in the late 1970s, while the notion that DCs have a unique role in the immune system was met with decades of skepticism (Steinman and Cohn, 1973; Steinman and Cohn, 1974). Forty years later, the exquisite ability of DCs to mount adaptive immune responses to foreign antigens is gradually recognized, and their contribution to the induction of tolerance to self-antigens is also becoming increasingly evident. Consequently, the potential therapeutic benefits of modulating the development and functions of DCs for vaccines against pathogens, tumors, and suppressive therapies for autoimmune diseases have attracted more attention in the research and clinical applications.
In the mid-1990s, the full scope and significance of DC diversity was first acknowledged. The finding that the DCs in murine spleen include two subsets defined by the presence or absence of CD8 expression and with distinct immune functions, substantially broadened our knowledge of the roles of DCs in the induction of adaptive immunity (Shortman and Heath, 2010). However, the existence of distinct DC subsets in non-lymphoid tissues was recognized much later. This was mainly due to the fact that although the functionally equivalent DC subsets shared several phenotypic features with their counterparts in lymphoid tissues, they did not, in fact, express CD8. Nevertheless, they were eventually characterized by the expression of the integrin CD103 (Helft et al., 2010).
Moreover, another major subset of the DC family was identified as a population of cells morphologically resembled plasma cells with weak antigen presentation ability, but, upon exposure to viral stimuli, they can produce large amounts of type-I interferon. In addition, upon viral stimulation these cells can also differentiate into immunogenic DCs that can prime T cells against viral antigens. These cells were named plasmacytoid DCs (pDCs) to distinguish them from the conventional DCs (cDCs) (Colonna et al., 2004).
MicroRNAs (miRNAs) are small regulatory non-coding RNAs that repress target transcripts post-transcriptionally. Within the past decade, the role of microRNAs (miRNAs) in immunology has been studied extensively. The importance of miRNAs in the control of differentiation and function of hematopoietic cells is clearly demonstrated by studies that genetically disrupt critical enzymes important for the biosynthesis of miRNAs (Xiao and Rajewsky, 2009). By gene targeting, it has been shown that genetic deficiency of Drosha results in the loss of mature miRNAs. In hematopoietic cells, T cell-specific Drosha and Dicer conditional knockout mice both spontaneously develop lymphoproliferative multi-organ inflammatory disease and die within a few weeks after birth (Chong et al., 2008). While the earliest studies of miRNA function in the immune system have demonstrated an essential role for miRNAs as a whole, later studies have focused on the contribution of specific miRNAs to specific immunologic processes. The important roles of miRNAs in hematopoietic development, cancer, immune homeostasis, inflammatory disease, and autoimmunity were gradually clarified. Such roles include their ability to negatively regulate signaling pathways and the expression of transcription factors essential in lineage commitment. It has been shown that development of specific cell lineages is often dependent on specific miRNAs. MiRNAs also play important roles in immune tolerance and many immune-related miRNAs regulate cell growth and apoptosis, leading to their high frequency of association with hematologic malignancy (Mehta and Baltimore, 2016). In this review, we will discuss the function of miRNAs in regulating DC differentiation and functions and their association with autoimmune diseases.
DC IN LYMPHOID TISSUES
DC in thymus
The thymus is a central immune organ and an important site for T cell differentiation, selection and generation of naive CD4+ and CD8+ T lymphocytes. Mouse thymus also contains CD11cintCD45RA+ plasmacytoid DCs (pDCs) and two CD11chiCD45RA− conventional DC (cDC) subsets that can be segregated on the basis of CD8α and the signal regulatory protein-α (Sirp-α) expression, as CD8α+Sirp-α− and CD8α−/loSirp-α+ cDC subsets (Fig. 1) (Lahoud et al., 2006; Donskoy and Goldschneider, 2006). The CD8α+Sirp-α− subset representing about 70% of thymic cDC is generated within the thymus from the earliest intrathymic progenitors, whereas the minor CD8α−/loSirp-α+ cDC subset originates from the peripheral migratory DCs (Donskoy and Goldschneider, 2006). Thymic cDC, although sharing many common features, differ from other peripheral DC subsets in that the majority of thymic cDCs in mouse is derived from an intrathymic precursor, and the former mostly present self-antigens (Ag) rather than foreign Ag (Wu et al., 1995; Wu et al., 1996). Thymic cDCs play important roles in the induction of central immune tolerance through a process called negative selection that deletes the developing thymocytes with self-reactivity and the generation of the naturally occurring CD4+CD25+ regulatory T cells (Goldschneider and Cone, 2003; Gallegos and Bevan, 2004; Watanabe et al., 2005; Bonasio et al., 2006; Proietto et al., 2008). Human thymus also contain DC subsets with similar functions as those found in the mouse (Bendriss-Vermare et al., 2001; Vandenabeele et al., 2001; Gurney et al., 2004; Keir et al., 2002).
Figure 1.
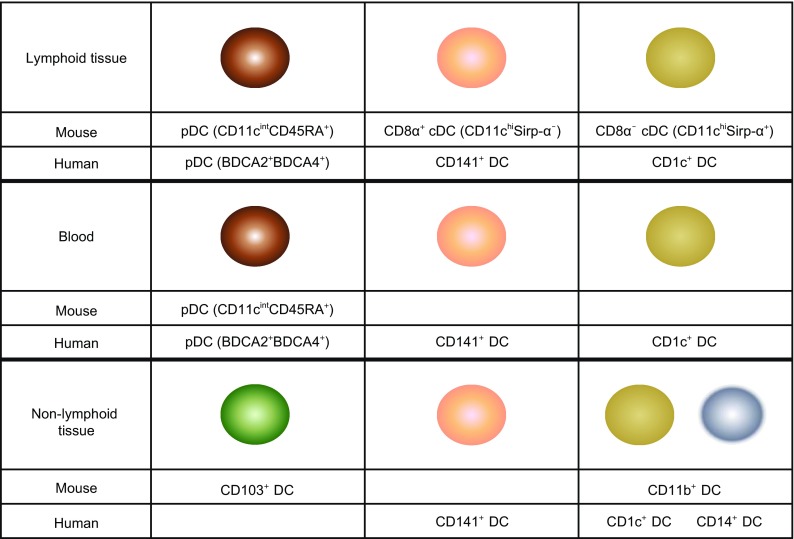
DC subsets in the lymphoid and non-lymphoid tissues of mouse and human. Lymphoid tissues in mouse contain pDC, CD8α+ cDC and CD4+ cDC, while lymphoid tissues in human contain pDC, CD141+ DC and CD1c+ DC. Mouse peripheral blood contains pDC while human blood contains pDC, CD141+ DC and CD1c+ DC. Non-lymphoid tissues in mouse contain CD103+ DC and CD11b+ DC, while non-lymphoid tissues in human contain CD141+ DC, CD1c+ DC and CD14+ DC
DC in spleen
The spleen is a rich source of lymphoid tissue-resident DCs. The DC populations in mouse spleen have been well characterized (Fig. 1). Three cDC subsets have been identified in the mouse spleen based on the surface expression of CD8α and CD4, in addition to high levels of CD11c expression on all cDCs. These cDC subsets are CD11chiCD4−CD8α+, CD11chiCD4−CD8α−, and CD11chiCD4+CD8α−. The CD4−CD8α+ cDCs also express CD205, but not Sirp-α. In contrast, both CD4−CD8α− and CD4+CD8α− cDC subsets do not express CD205−, but are Sirp-α+, and are sometimes considered as one CD8α− cDC subset (Lahoud et al., 2006; Vremec et al., 2000). The CD8α−CD205− cDCs are located in the marginal zone, whereas the CD8α+CD205+ cDCs are in T cell areas. Marginal-zone DC can rapidly migrate into the T cell zone upon activation (De Smedt et al., 1996). A significant number of splenic cDC can be generated in situ by the intrasplenic immediate cDC precursors, named pre-DCs (Naik et al., 2006; Diao et al., 2006).
In addition to cDCs, pDCs are also found in mouse spleen. They are defined as CD11cintCD45RA+B220+SiglecH+. Similar to the blood pDC, the freshly isolated splenic pDC do not have the phenotypic and functional features of the antigen-presenting cDC, but can assume a cDC morphology and upregulate the cDC markers CD11c and MHC class II after activation with microbial stimuli. They represent the major cell type that produce large amounts of type-I interferon, a cytokine involved in innate immunity to virus. The pDCs in spleen migrate from the peripheral blood, because cells with the characteristics of pDC can be found in mouse blood, and the intrasplenic pre-DC do not differentiate into pDC (Asselin-Paturel et al., 2001; Nakano et al., 2001; O’Keeffe et al., 2002; O’Keeffe et al., 2003). Human spleen also contains pDCs, displaying plasma cell morphology, that selectively express Toll-like receptor (TLR)-7 and TLR9, and are specialized to rapidly secret massive amounts of type 1 interferon following viral stimulation. These are the CD4+CD11c−Lin−BDCA-2+BDCA-4+ cells (Siegal et al., 1999; Kadowaki et al., 2001; Liu, 2005; Mittag et al., 2011).
DC in lymph node
The DC populations found in mouse LNs are more complex (Fig. 1). In addition to the three phenotypically and functionally equivalent cDC populations found in mouse spleen, two additional subpopulations have been described in the skin draining LNs. These correspond to the mature CD8αloCD205int and CD8αloCD205hi cDC that migrate from the epidermis and dermis, respectively, to the LNs. Subcutaneous LNs contain a higher percentage of the CD8αloCD205hi Langerhans cell (LC)-like cells than mesenteric LNs. The DCs derived from the migratory LC are responsible for carrying antigens picked up from skin to the draining LNs (Henri et al., 2001; Hochrein et al., 2001). In human LN, HLA−DR+CD11c−BDCA4+ cells have been identified as pDCs. HLA−DR+CD11c+ cells were separated into CD14+ and CD1a+ cells, which can be further divided into EpCAM+ LCs and CD1a+ DCs. CD1a−CD14− cells can be further fractionated into Clec9A+ and BDCA1+ populations. Finally, BDCA1+ cells are comprised two subsets which either do or do not express CD206. Similar analysis of lymphoid organs that do not drain the skin showed that three of these DC subsets (LCs, CD1a+, and CD206+ DCs) were absent from cervical LNs draining the oropharynx, iliac LNs, tonsils, and spleen, suggesting that these DCs in skin-draining LNs are unique to and derived from the skin (Segura et al., 2012).
ORIGINS OF LYMPHOID TISSUE DC
DCs, like all other leukocytes, develop from bone marrow-derived hematopoietic stem cells. Both cDC and pDC can be generated from the Flt3 expressing early myeloid or lymphoid progenitors, and Flt3L is essential for the development of steady-state DC populations (Fig. 2). When common lymphoid precursors (CLPs) and common myeloid precursors (CMPs) were purified from mouse bone marrow (BM) and adoptively transferred intravenously into irradiated recipient mice, they both showed the potential to give rise to splenic cDCs and pDCs. However, CMPs are 10-fold more abundant than CLPs; therefore, most spleen cDCs originate from CMPs. pDC are also derived from CMP, CLP, and DC restricted precursors CDP (common DC precursors) when these precursors are transferred into irradiated recipients (Wu et al., 2001; Manz et al., 2001; D’Amico and Wu, 2003; Martín et al., 2000).
Figure 2.

The development of different DC subsets. All DC subsets are derived from Flt3+ LMPP. CD8α+ cDC, CD8α− cDC and CD103+ DC originate from CMP and CDP. pDC are differentiated from CLP, CMP and CDP. Langerhans cells and moDC are from monocytes
DC IN PERIPHERAL TISSUES
Non-lymphoid tissue-resident DCs are present in most tissues in the steady state (Fig. 1). Phenotypically, these DCs lack the expression of other hematopoietic cell lineage markers, but express the hematopoietic specific marker CD45, the integrin CD11c, and constitutively the major histocompatibility complex class II (MHCII). Several studies have now established that this definition is too broad and includes distinct DC populations with different origins and functions. Studies of non-lymphoid tissue DCs so far have largely involved tissues that are in contact with the body surfaces, such as the skin, lung, and intestine. We therefore will focus our review on the DCs from these specific tissues.
DC in intestine
The intestine harbors a complex system of organized lymphoid tissues as primary sites for the induction of adaptive mucosal immune responses, including ileal Peyer’s patches, colonic isolated lymphoid follicles, and mesenteric lymph nodes (MLNs). Due to the difficulty of obtaining normal human gut tissues, most of the studies were done using mouse models. Like other tissue cDCs, mouse intestinal non-lymphoid tissue cDCs are Flt3L dependent and pre-DC derived (Bogunovic et al., 2009; Varol et al., 2009; Schulz et al., 2009; Edelson et al., 2010). Under steady-state, all intestinal cDCs are migratory cells, arising from classic DC precursors, without monocytic intermediates.
Intestinal DCs were originally defined as CD11c+ cells which were then subfractionated into intestinal macrophages (now best defined as CD11c+CD64+ cells) and DC (MHCII+CD11c+CD64−) subsets. MHCII+CD11c+ DCs included subpopulations best characterized as CD103+CD11b− and CD103+CD11b+ (Bogunovic et al., 2009; Varol et al., 2009). The generation of intestinal CD103+CD11b− cDCs depends on BATF3, Id2, and IRF8 and these cells thus represent the gut equivalent of splenic CD8α+ cDCs (Edelson et al., 2010; Klebanoff et al., 2013). Intestinal CD103+CD11b+ cDCs were subfractionated via the surface markers CD24 and CD64, revealing their composition of bona fide Flt3-dependent CD24+CD64− cells and contaminating CD24−CD64+ cells that represent intestinal macrophages.
The development of CD24+CD103+CD11b+ DCs depends on transcription factor IRF4 (Persson et al., 2013), and the presence of granulocyte-macrophage colony stimulating factor (GM-CSF) is also required for the generation of CD103+CD11b+ lamina propria (LP) DCs (Farache et al., 2013). These CD103+CD11b+ LP DCs have been implicated in both the maintenance of tolerance towards the commensal microflora, and the generation of protective immune responses against pathogens (Jaensson et al., 2008; Farache et al., 2013). In the steady state, the functional properties of DCs appear to vary according to their anatomical locations. Indeed, CD103+CD11b+ DCs are required in vivo for efficient generation of Treg cells (Scott et al., 2014; Spadoni et al., 2012; Goto et al., 2014).
DC in lung
The lung, like the intestine, is vulnerable to pathogenic insult and is constantly exposed to potentially harmful substances. Studies have highlighted the abundance of DCs in lung tissue and their dynamic accumulation upon stimulation. The mouse lung is grossly divided into large conducting airways and lung interstitium containing alveolar septa and capillaries where gas exchange is taking place. Under steady-state conditions, the conducting airways of all species studied are lined with an intraepithelial highly dendritic network of MHCIIhiCD11chi cells that are mostly CD11b− and express langerin and the mucosal integrin CD103 (αEβ7) (Lambrecht and Hammad, 2009).
In addition the network has the propensity of extending dendrites into the airway lumen through formation of tight junctions with bronchial epithelial cells. Immediately below, the lamina propria of the conducting airways contains MHCIIhiCD11chi cells that also express high levels of CD11b, but not CD103, and are a rich source of proinflammatory chemokines (Lambrecht and Hammad, 2009; Guilliams et al., 2013; Grayson, 2006). The CD11b+CD103− subset also expresses the signal regulatory protein-α (SIRPα) molecule. The exact location of the CD11cint Siglec-H+ BSA-1+ pDCs in the lung is unclear although they can be found to line alveolar septa in situ and have been recovered from digests of large conducting airways. The alveolar space also contains CD11chiMHCIIhi DCs and is easily accessible by broncho alveolar lavage.
The development of lung CD103+CD11b− cDCs depends on Id2 and IRF8, aligning these cells with the classical CD8α+ and CD103+ DC lineage (Edelson et al., 2010; Ginhoux et al., 2009; Schlitzer et al., 2013). Consistent with this, lung CD103+CD11b− cDCs are specialized in cross-presentation of antigens to CD8+ T cells. Notably, lung CD103+CD11b− cDCs prominently express RALDH after Ag inhalation and facilitate de novo Treg cell induction (Plantinga et al., 2010). The lung CD103−CD11b+ cDCs are outnumbered by the cross-presenting CD103+ cDCs. More recently, CD11b+ cDCs, further defined as being CD24+, were shown to be dependent on IRF4. The CD11b+CD24+ cDCs were found to direct a Th17 cell response after challenge possibly due to their production of IL-23 (Schlitzer et al., 2013; Kim and Lee, 2014). Fate-mapping systems have also shown that while almost all lung CD103+ cDCs are CDP derived, this is not the case for CD11b+ cDCs, for which only about 50% of cells have been found to be of CDP origin (Liu and Nussenzweig, 2010). The additional developmental pathway for lung CD11b+ cDCs in the steady state remains to be defined.
DC in skin
The skin can be divided into two anatomical sites: the epidermis and the dermis. The epidermis composed of keratinocytes is a tightly packed stratified epithelium that forms the water-impermeable stratum corneum. A basement membrane separates the epidermis from the underlying dermis, which is made up of fibroblasts and of intertwined collagen and elastin fibers (Merad et al., 2008; Malissen et al., 2014). The mononuclear phagocyte system is composed of DCs, monocytes, and macrophages. In the skin, DCs may include pDC, cDC, and moDC. Under steady-state conditions, pDCs are absent from the skin. They have only been identified in inflamed skin where they promote wound repair and mediate the systemic pro-inflammatory response that is seen after stimulation with Toll-like receptor 7 (TLR7) agonists (Gregorio et al., 2010).
Mouse cDCs in the dermis are generally categorized using the expression of CD11c and CD11b (also known as ITGAM). Dermal CD11c+CD11b− cDCs express the C-type lectin langerin (also known as CD207 and CLEC4K) and include CD103+ and CD103− cells (Poulin et al., 2007; Crozat et al., 2011). They are developmentally related to the CD8α+ cDCs that reside in secondary lymphoid tissues (Crozat et al., 2011; Hildner et al., 2008). Dermal CD11b+ cDCs are the most abundant type of DC in healthy dermis and, until recently, it has been difficult to distinguish them from CD11b+ monocyte- derived DCs and macrophages. The cDCs that are found in healthy non-lymphoid tissue, such as the skin, are not ‘at rest’. A small proportion of these cDCs undergoes a maturation process that is referred to as homeostatic maturation and that involves morphological and phenotypical changes that lead to their migration to the draining lymph nodes (Wilson et al., 2008). Such homeostatic maturation occurs continuously in germ-free mice and does not require TLR signals (Wilson et al., 2008).
Following homeostatic maturation, non-lymphoid tissue cDCs upregulate their expression of MHCII molecules and can transport cutaneous self-antigens to the T cell zones of the draining lymph nodes. If they encounter T cells that have escaped central tolerance, these cDCs trigger an abortive programme of activation in autoreactive T cells. In response to activation by protozoal, fungal, viral or bacterial stimuli, cDCs that reside in skin undergo a terminal differentiation program that differs from homeostatic maturation in that it additionally results in upregulation of the expression of co-stimulatory molecules. Such maturation leads to the migration of cDCs to draining lymph nodes, where they promote the clonal expansion of naïve antigen-specific T cells and the acquisition of T cell effector functions (Probst et al., 2003; Spörri and Reis e Sousa, 2005).
In human dermis, during steady-state conditions, DCs are classically divided into CD14+ and CD1a+ DCs (Nestle et al., 2009). CD14+ dermal dendritic cells (DDC) show variable expression of CD1a, CD1c, and CD163 (Haniffa et al., 2009). CD14+ DDC express low levels of CD80 and CD86 and are relatively poor inducers of naïve T-cell proliferation. They do however efficiently take up antigen, possibly due to expression of c-type lectins like CD206 and CD209/DC-SIGN (Haniffa et al., 2012; Klechevsky et al., 2008; Angel et al., 2007; Angel et al., 2006; Nestle et al., 1994). They also have the ability to induce Treg through high production of IL-10 (Chu et al., 2012). Skin-derived CD1a+ DCs in contrast express high levels of CD80 and CD86 and strongly induce allogeneic naïve CD4+ T cells and CD8+ T-cell proliferation (Haniffa et al., 2012; Klechevsky et al., 2008; Angel et al., 2007; Angel et al., 2006; Nestle et al., 1994). CD1a+ DDC isolated from skin-draining lymph nodes were found to preferentially induce TH2 polarization similar to LC (Segura et al., 2012).
HUMAN DC SUBSETS IN PERIPHERAL BLOOD
In addition to the DC subsets described in different human tissues, human blood also contains different DC subsets. All blood DCs express high levels of HLA-DR and lack typical lineage markers CD3 (T cell), CD19/20 (B cell), and CD56 (natural killer cell). Myeloid DCs (mDCs) express typical myeloid antigens CD11c, CD13, CD33, and CD11b, corresponding to mouse CD11c ‘classical’ or ‘conventional’ DCs. In humans, both monocytes and mDCs express CD11c, but DCs lack CD14 or CD16 and can be split into CD1c+ and CD141+ subsets (Mittag et al., 2011; Jongbloed et al., 2010; Poulin et al., 2010; Crozat et al., 2010; Bachem et al., 2010), functionally corresponding to the mouse CD11b+ and CD8+/CD103+ cDC, respectively (Mittag et al., 2011; Jongbloed et al., 2010; Poulin et al., 2010; Crozat et al., 2010; Bachem et al., 2010). pDCs were first identified in humans. They typically lack myeloid antigens and can be distinguished by the expression of CD123, CD303, and CD304 (Cella et al., 1999; Mathan et al., 2013).
INFLAMMATORY DC
During inflammation or infection, lymphoid and nonlymphoid organs can harbor DCs that originate from monocyte infiltrates and have been termed “monocyte-derived DCs” (moDCs) or “inflammatory DCs”. MoDCs are phenotypically difficult to discern from cDCs because they share similar expression patterns of MHC-II, CD11b, and CD11c. However, as an indicative of their monocyte precursor, moDCs express CD64, the Fc-gamma receptor 1 (FcγRI) (Mildner et al., 2013). Monocytes have long been known to give rise to DC-like cells that can efficiently stimulate T cells when cultured in vitro in the presence of GM-CSF and IL-4 (Plantinga et al., 2013). Gene expression profiles of cDCs and BM culture-derived DCs differ considerably. A relation of moDCs and cDCs is supported by the fact that culture-derived moDCs express the transcription factor zbtb46 that is restricted to cDCs present in the lymphoid tissues (Satpathy et al., 2012). MoDCs have since been described during pathogenic inflammation, experimental sterile inflammation, and in models of inflammatory diseases, such as colitis, rheumatoid arthritis (RA), systemic lupus erythematosus (SLE), and experimental autoimmune encephalomyelitis (EAE) (Ganguly et al., 2013).
THE FUNCTION OF miRNA IN THE DEVELOPMENT OF DC
In DCs, CD11c-specific deletion of Dicer caused reduction in Langerhans cells without obvious perturbation of other DC subsets (Kuipers et al., 2010). However, within hematopoietic cells, miRNAs are differentially expressed by different lineages of hematopoietic cells and their precursors. Amongst DCs, the miRNA expression patterns are also distinct amongst different DC subsets (Kuipers et al., 2010).
It has been found that miR-22 is highly expressed in mouse CD11c+CD11b+B220− cDCs compared to pDCs, and is induced in DC progenitor cell cultures with GM-CSF, which stimulates CD11c+CD11b+B220− cDCs differentiation. Overexpression of miR-22 during DC development enhanced CD11c+CD11b+B220− cDC generation at the expense of pDCs, while miR-22 knockdown demonstrated an opposite effect (Li et al., 2012). Overexpression and knockdown of miR-22 showed significant effects on the mRNA abundance of IRF8, a transcription factor essential for pDC and CD8α+ cDC development. These studies demonstrated that miR-22 was important in regulating the differentiation of DC subsets (Li et al., 2012).
Another important miRNA involved in DC homeostasis is miR-142. By analyzing miRNA expression profiles for distinct myeloid populations, including BM-resident progenitors, monocytes, and mature splenic DC subsets, miR-142 was found to be highly expressed in classic FLT3-L–dependent CD4+ cDCs, whereas reduced expression was observed in CD8α+ or CD4−CD8α− cDCs (Mildner et al., 2013). Moreover, mice deficient for miR-142 displayed an impairment of CD4+ cDC homeostasis both in vitro and in vivo. Comparison of the expression profiles of WT and miR-142−/−CD4+ cDC equivalents using ingenuity pathway analysis revealed an up-regulation of the transcription factors HoxA9, IRF8, and Meis1 in miR-142−/−CD4+ cDCs. The up-regulation of IRF8 may suggest a function for miR-142 in the specification of CD4+ versus CD8α+ cDCs through regulation of the IRF8 pathway (Mildner et al., 2013).
THE ROLE OF miRNA IN THE ACTIVATION OF DC
In addition to the role of miRNA in DC development, miRNA also functions in DC activation. When human moDCs were stimulated with LPS, miR-155 and several other miRNAs were highly up-regulated. Further in vitro studies showed that miR-155 acts as a positive regulator for the production of several pro-inflammatory cytokines including IL-6, IL-23, IL-12, and TNF-α by mouse moDCs stimulated using LPS. MiR-155 modulates cytokine production by targeting the negative regulators of signaling, such as SOCS1 and SHIP1 (Ceppi et al., 2009; O’Connell et al., 2009; O’Connell et al., 2010).
MiR-146a, which can also be induced by LPS stimulation, was found to negatively regulate DC activation. Knockdown of miR-146a caused an increase in NF-κB signaling through direct targeting TRAF6 and IRAK1 (Taganov et al., 2006). MiR-142-3p was also identified as a key negative regulator of IL-6. In contrast to miR-155, which is strongly up-regulated after LPS stimulation, miR-142-3p is constitutively and highly expressed in resting moDCs but down-regulated after LPS stimulation. MiR-142-3p directly targets IL-6 mRNA and thus specifically affects IL-6 expression (Sun et al., 2011).
Several miRNAs have been reported to modulate IL-12 production. MiR-21 was identified to directly inhibit IL-12p35 expression. Compared to wild type mice, in vitro derived moDCs from miR-21 deficient mice had enhanced production of IL-12, but not other cytokines (including TNF-α, IL-6, and IL-23) upon stimulation by LPS (Lu et al., 2011). MiR-10a also directly targets the IL-12 gene. Unlike miR-21, miR-10a negatively regulates the production of IL-12/IL-23p40. Ectopic expression of miR-10a in moDCs suppressed both production of IL-12 and IL-23 (Xue et al., 2011). Compared to miR-21 and miR-10a that directly target IL-12 genes, some other miRNAs target the signaling components that will affect multiple downstream targets. For example, miR-148/152 suppressed IL-12 as well as IL-6 production; miR-23b suppressed IL-12 production while enhancing IL-10 production (Liu et al., 2010).
Maturation of moDCs from human monocytes is accompanied by upregulation of DC-SIGN and downregulation of CD14. Based on DC-SIGN/CD14 expression ratios, miR-21, miR-34a, and their cognate targets WNT1 and JAG1 were found to negatively influence moDC differentiation (Hashimi et al., 2009). Similarly, inhibition of miR-511 and miR-99b resulted in reduced DC-SIGN levels (Tserel et al., 2011).
Although the main body of current research on the role of miRNAs in regulating DC differentiation and function used in vitro GM-CSF differentiated DCs, several studies have investigated other type of DCs. MiR-223 was found highly expressed in freshly isolated epidermal LCs, and lack of miR-223 significantly increased LCs-mediated antigen-specific CD8+ T cell proliferation in vivo and in vitro (Mi et al., 2013). pDCs are specialized cells that produce type-I IFN and express high levels of TLR7 and TLR9. Studies with human pDCs have revealed a role for miR-155 in fine tuning the TLR7-stimulated IFN-α production. Although TLR7 induced up-regulation of both miR-155* and miR-155 through the JNK pathway in pDCs, miR-155* induced during early phase of pDC activation enhanced IFN-α production by suppressing IRAKM, whereas miR-155 induced during later phase of pDC activation inhibited IFN-α expression by targeting TAB2. Thus, these two miRNAs cooperatively regulated the production of type-I IFN by human pDCs (Zhou et al., 2010).
THE ROLE OF DC INTRINSIC miRNA IN AUTOIMMUNE DISEASE
The involvement of DCs in tolerance and autoimmunity is complex and bidirectional. Indeed, DCs may promote tolerance through multiple mechanisms, such as through central and peripheral tolerance induction, the generation and maintenance of Treg cells, as well as the induction of T cell unresponsiveness. Conversely, the powerful antigen presentation capacity of DCs may promote the activation and/or differentiation of self-reactive effector T cells, either because of the defective processes of central and peripheral tolerance induction or inappropriate activation of the self-reactive T cells due to impaired/ineffective negative regulation.
Increasing evidence has demonstrated that miRNAs are important regulators for normal immune responses and contribute to the prevention of autoimmunity (Table 1). They exert their effects either through regulating the differentiation and function of DCs or through direct regulation of immune responses.
Table 1.
The role of miRNAs in autoimmune diseases
| miRNA | Predicted/Identified targets | Function | Related diseases |
|---|---|---|---|
| miR-22 | IRF8 | Enhances CD11c+CD11b+B220− cDC generation at the expense of pDCs | |
| miR-142 | IRF8 | Plays a pivotal role in the maintenance of CD4+ DCs | |
| miR-142-3p | IL-6 | Specifically inhibits IL-6 expression by moDC | MS |
| miR-21 | IL-12p35, Wnt1 | Negatively regulates the production of IL-12 by moDC; negatively regulate the development of moDC | SLE, IBD, UC, MS |
| miR-10a | IL-12/IL-23p40 | Suppress the production of IL-12 and IL-23 by moDC | SLE |
| miR-148/152 | Calcium/Calmodulin- dependent protein kinase IIa | Suppress the production of IL-12 and IL-6 | SLE |
| miR-23b | Notch1, NF-κB | Inhibits the production of IL-12 while promotes IL-10 production | UC |
| miR-155 | SOCS1, SHIP1, TAB2 | Positively regulates the production of several pro-inflammatory cytokines including IL-6, IL-23, IL-12, and TNF-α | RA, IBD |
| miR-146a | IRAK1, TRAF6 | Negatively regulates TLR4-NF-κB pathway in monocytes | RA, SLE, IBD |
| miR-34a | JAG1 | Negatively regulates the development of moDC | MS |
| miR-223 | C/EBPβ | Negatively regulates LCs-mediated antigen-specific CD8+ T cell proliferation, production of inflammatory cytokine TNFα, IL-1β, and IL-23 by intestinal DCs. Positively regulates the differentiation of intestinal CX3CR1+ macrophages | UC |
| miR-29 | IL-12p40, ATF2 | Negatively regulates the production of IL-23 production by moDC | UC |
| miR-155* | IRAKM | Positively regulates the production of IFN-α by human pDC |
Rheumatoid arthritis (RA) is a systemic autoimmune disorder characterized by chronic inflammation of synovial tissue that results in irreversible joint damage. Inflammatory cytokines, especially TNF-α, IL-1β, and IL-6 are known to play an important role in RA pathogenesis, as inhibition of these cytokines can ameliorate disease in some patients (Smolen et al., 2007; Bresnihan et al., 1998). Stanczyk et al. reported increased miR-155 and miR-146a expression in RA synovial fibroblasts compared to those in osteoarthritis (OA) patients (Stanczyk et al., 2008). Furthermore, miR-155 expression was higher in RA synovial tissue compared to that of OA. MiR-155 expression was also higher in monocytes from RA synovial fluid compared to those from RA peripheral blood. Enhanced expression of miR-155 in RA synovial fibroblasts revealed matrix metalloproteinase 3 (MMP-3) as a potential target of miR-155, suggesting that miR-155 may modulate downstream tissue damage (Stanczyk et al., 2008). Since miR-155 is a positive regulator for moDC secreted IL-12p40, IL-12p35, and TNF-α (Dunand-Sauthier et al., 2011), the increased expression of miR-155 probably facilitates the excessive inflammatory response.
SLE is another systemic inflammatory autoimmune disease characterized by the presence of autoantibodies against numerous self-antigens including chromatin, ribonucleoproteins, and phospholipids. Seven miRNAs (miR-31, miR-95, miR-99a, miR-130b, miR-10a, miR-134, and miR-146a) were expressed at 6-fold lower level in SLE patients than that of healthy controls (Tang et al., 2009). Among these miRNAs, miR-146a in particular has been reported to negatively regulate the innate immune response by targeting interleukin-1 receptor–associated kinase 1 (IRAK1) and tumor necrosis factor receptor–associated factor 6 (TRAF6) in monocyte and moDC. Further analysis showed that under-expression of miR-146a negatively correlated with clinical disease activity and the interferon (IFN) scores in patients with SLE. Over-expression of miR-146a reduced, whereas inhibition of endogenous miR-146a increased, the expression of type I IFNs in peripheral blood mononuclear cells (PBMCs) (Steinman and Cohn, 1973). Besides, as miR-10a has also been demonstrated to negatively regulate DC function by direct targeting IL-12/IL-23p40 (Xue et al., 2011), its lower expression in SLE patients may also promote the autoimmune responses.
MiRNAs are also likely involved in the pathogenesis of psoriasis, a disease that may occur in association with inflammatory bowel disease (IBD). In a study comparing psoriasis patients to healthy controls, miR-203, miR-21, and miR-146a were significantly higher, while miR-123b was decreased in psoriatic skin compared to patients with atopic dermatitis and healthy controls. MiR-203 is expressed at the highest level in skin keratinocytes. One potential target of miR-203, SOCS-3, is upregulated in psoriatic lesions. Suppression of SOCS-3 activity leads to activation of STAT3, which when activated in keratinocytes in transgenic mice leads to the development of psoriasis (Sonkoly et al., 2007). Interestingly, a significant accumulation of FLT3+CD11c+ DCs in human psoriatic lesions and in the skin of experimental preclinical K14-VEGF transgenic homozygous mice, a mouse model for psoriasis. Targeted inhibition of FLT3 almost completely cured the psoriasis-like disease (Yan et al., 2014). As STAT3 is the key transcription factor for FLT3 induced DC production, the increased miR-203 expression may lead to activation of STAT3, and then facilitate pDC and cDC differentiation.
As seen in RA, SLE, and psoriasis, there is increasing data that patients with ulcerative colitis (UC) and Crohn’s disease (CD) have altered miRNA profiles in involved tissues compared to controls. The first study examined miRNA expression in IBD compared sigmoid colon biopsies from patients with active UC, inactive UC, chronic active CD, irritable bowel syndrome, and microscopic colitis with healthy control subjects. Eight miRNAs (miR-16, miR-21, miR-23a, miR-24, miR-29a, miR-126, miR-195, and Let-7f) were significantly increased in active UC tissues and three miRNAs (miR-192, miR-375, and miR-422b) were significantly decreased in the UC tissues compared to healthy controls (Wu et al., 2008). MiR-192 and miR-21 were the most highly expressed miRNAs associated with active UC in human colon tissues. MiR-21 has been demonstrated positively regulating the progression of DSS induced colitis and miR-21 deficient mice showed improved survival rate during DSS induction (Shi et al., 2013). A second study confirmed that miR-21 was elevated in inflamed tissue from 12 UC patients compared to 12 healthy controls. This study also identified upregulated miR-155 in inflamed tissues (Takagi et al., 2010). Similar to miR-21, miR-155 knockout mice showed alleviated symptoms in DSS induced colitis model (Singh et al., 2014). A third analysis of miRNAs in UC tissue examined eight UC patients and 10 healthy controls. Biopsies were obtained from inflamed and non-inflamed tissues of UC patients. In this study, five miRNAs (miR-29a, miR-29b, miR-126*, miR-127-3p, miR324-3p) were found upregulated and four (miR-188-5p, miR-25, miR-320a, miR-346) downregulated in both quiescent and active UC compared to healthy controls (Fasseu et al., 2010). As DCs are important regulators for IBD, we examined miRNA expression specifically in intestinal DC subsets and found that the expression of miR-223 by intestinal DCs decreased continuously during the progression of colitis. By using the DSS-induced colitis mouse model, we demonstrated that miR-223 deficiency resulted in more severe symptoms. Intestinal DCs in miR-223 deficient mice showed a more pro-inflammatory phenotype and a decreased number of CX3CR1+ regulatory macrophages was also observed in miR-223 deficient mice. Mechanistic study revealed that miR-223 play an important role in maintaining the homeostasis of intestinal macrophages and DCs by directly targeting C/EBPβ (Zhou et al., 2015). Although different colitis related miRNAs have been reported, most of the expression profiles of these miRNAs were obtained form the tissue or a mixture of different cell types. In order to get a better understanding of the functions of these miRNAs, further analysis of their changes in different cell types is required. As one example, miR-29a was shown to be a negative regulator for colitis by directly inhibiting the production of IL-23 by moDCs (Brain et al., 2013), it was also found upregulated in active UC tissues (Fasseu et al., 2010).
CONCLUSIONS
Rapid progress has been made in recent years in our understanding of the roles of miRNAs in regulating differentiation and function of DCs. These studies have helped us to better understand the biological significance of these miRNAs. Meanwhile, increasing numbers of studies have suggested that the dysregulated expression of miRNAs in DCs may be closely associated with different immune disorders. However, most of the current available evidence was mainly derived from in vitro studies with GM-CSF-differentiated DCs, the equivalents of moDCs that become abundant during inflammation. It remains to be confirmed whether these findings from in vitro studies can be applied to in vivo settings when very dynamic interaction between multiple types of immune cells and multiple components of microbes occur. Furthermore, complex DC networks consist of many DC subsets with shared and distinct functions. It remains largely unknown how miRNAs regulate the development and function of different DC subsets. Further studies are required to clarify the contribution of miRNAs to the diversity of DC subsets in lymphoid and non-lymphoid tissues, and to the development of DC related immune disorders.
Acknowledgments
Abbreviations
BM, bone marrow; CD, Crohn’s disease; cDCs, conventional DCs; CLPs, common lymphoid precursors; CMPs, common myeloid precursors; DCs, dendritic cells; EAE, experimental autoimmune encephalomyelitis; FcγRI, Fc-gamma receptor 1; GM-CSF, granulocyte-macrophage colony stimulating factor; IBD, inflammatory bowel disease; IFN, interferon; IRAK1, interleukin-1 receptor-associated kinase 1; LC, Langerhans cell; LP, lamina propria; mDCs, myeloid DCs; MHCII, major histocompatibility complex class II; miRNAs, microRNAs; MLNs, mesenteric lymph nodes; MMP-3, matrix metalloproteinase 3; moDCs, monocyte derived DCs; OA, osteoarthritis; pDCs, plasmacytoid DCs; RA, rheumatoid arthritis; SLE, systemic lupus erythematosus; TLR, Toll-like receptor; TRAF6, tumor necrosis factor receptor-associated factor 6; UC, ulcerative colitis.
Compliance with ethics guidelines
Li Wu and Haibo Zhou declare that they have no conflict of interest.
References
- Angel CE, George E, Brooks AE, Ostrovsky LL, Brown TL, Dunbar PR. Cutting edge: CD1a+ antigen-presenting cells in human dermis respond rapidly to CCR7 ligands. J Immunol. 2006;176:5730–5734. doi: 10.4049/jimmunol.176.10.5730. [DOI] [PubMed] [Google Scholar]
- Angel CE, Lala A, Chen CJ, Edgar SG, Ostrovsky LL, Dunbar PR. CD14+ antigen-presenting cells in human dermis are less mature than their CD1a+ counterparts. Int Immunol. 2007;19:1271–1279. doi: 10.1093/intimm/dxm096. [DOI] [PubMed] [Google Scholar]
- Asselin-Paturel C, Boonstra A, Dalod M, et al. Mouse type I IFN-producing cells are immature APCs with plasmacytoid morphology. Nat Immunol. 2001;2:1144–1150. doi: 10.1038/ni736. [DOI] [PubMed] [Google Scholar]
- Bachem A, Güttler S, Hartung E, et al. Superior antigen cross-presentation and XCR1 expression define human CD11c+CD141+ cells as homologues of mouse CD8+ dendritic cells. J Exp Med. 2010;207:1273–1281. doi: 10.1084/jem.20100348. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bendriss-Vermare N, Barthélémy C, Durand I, et al. Human thymus contains IFN-alpha-producing CD11c(-), myeloid CD11c(+), and mature interdigitating dendritic cells. J Clin Invest. 2001;107:835–844. doi: 10.1172/JCI11734. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bogunovic M, Ginhoux F, Helft J, et al. Origin of the lamina propria dendritic cell network. Immunity. 2009;31:513–525. doi: 10.1016/j.immuni.2009.08.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bonasio R, Scimone ML, Schaerli P, Grabie N, Lichtman AH, von Andrian UH. Clonal deletion of thymocytes by circulating dendritic cells homing to the thymus. Nat Immunol. 2006;7:1092–1100. doi: 10.1038/ni1385. [DOI] [PubMed] [Google Scholar]
- Brain O, Owens BM, Pichulik T, et al. The intracellular sensor NOD2 induces microRNA-29 expression in human dendritic cells to limit IL-23 release. Immunity. 2013;39:521–536. doi: 10.1016/j.immuni.2013.08.035. [DOI] [PubMed] [Google Scholar]
- Bresnihan B, Alvaro-Gracia JM, Cobby M, et al. Treatment of rheumatoid arthritis with recombinant human interleukin-1 receptor antagonist. Arthritis Rheum. 1998;41:2196–2204. doi: 10.1002/1529-0131(199812)41:12<2196::AID-ART15>3.0.CO;2-2. [DOI] [PubMed] [Google Scholar]
- Cella M, Jarrossay D, Facchetti F, et al. Plasmacytoid monocytes migrate to inflamed lymph nodes and produce large amounts of type I interferon. Nat Med. 1999;5:919–923. doi: 10.1038/11360. [DOI] [PubMed] [Google Scholar]
- Ceppi M, Pereira PM, Dunand-Sauthier I, et al. MicroRNA-155 modulates the interleukin-1 signaling pathway in activated human monocyte-derived dendritic cells. Proc Natl Acad Sci USA. 2009;106:2735–2740. doi: 10.1073/pnas.0811073106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chong MM, Rasmussen JP, Rudensky AY, Littman DR. The RNAseIII enzyme Drosha is critical in T cells for preventing lethal inflammatory disease. J Exp Med. 2008;205:2005–2017. doi: 10.1084/jem.20081219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chu CC, Ali N, Karagiannis P, et al. Resident CD141 (BDCA3)+ dendritic cells in human skin produce IL-10 and induce regulatory T cells that suppress skin inflammation. J Exp Med. 2012;209:935–945. doi: 10.1084/jem.20112583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Colonna M, Trinchieri G, Liu YJ. Plasmacytoid dendritic cells in immunity. Nat Immunol. 2004;5:1219–1226. doi: 10.1038/ni1141. [DOI] [PubMed] [Google Scholar]
- Crozat K, Guiton R, Contreras V, et al. The XC chemokine receptor 1 is a conserved selective marker of mammalian cells homologous to mouse CD8alpha+ dendritic cells. J Exp Med. 2010;207:1283–1292. doi: 10.1084/jem.20100223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crozat K, Tamoutounour S, Vu Manh TP, et al. Cutting edge: expression of XCR1 defines mouse lymphoid-tissue resident and migratory dendritic cells of the CD8α+ type. J Immunol. 2011;187:4411–4415. doi: 10.4049/jimmunol.1101717. [DOI] [PubMed] [Google Scholar]
- D’Amico A, Wu L. The early progenitors of mouse dendritic cells and plasmacytoid predendritic cells are within the bone marrow hemopoietic precursors expressing Flt3. J Exp Med. 2003;198:293–303. doi: 10.1084/jem.20030107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- De Smedt T, Pajak B, Muraille E, et al. Regulation of dendritic cell numbers and maturation by lipopolysaccharide in vivo. J Exp Med. 1996;184:1413–1424. doi: 10.1084/jem.184.4.1413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Diao J, Winter E, Cantin C, et al. In situ replication of immediate dendritic cell (DC) precursors contributes to conventional DC homeostasis in lymphoid tissue. J Immunol. 2006;176:7196–7206. doi: 10.4049/jimmunol.176.12.7196. [DOI] [PubMed] [Google Scholar]
- Donskoy E, Goldschneider I. Two developmentally distinct populations of dendritic cells inhabit the adult mouse thymus: demonstration by differential importation of hematogenous precursors under steady state conditions. J Immunol. 2006;170:3514–3521. doi: 10.4049/jimmunol.170.7.3514. [DOI] [PubMed] [Google Scholar]
- Dunand-Sauthier I, Santiago-Raber ML, Capponi L, et al. Silencing of c-Fos expression by microRNA-155 is critical for dendritic cell maturation and function. Blood. 2011;117:4490–4500. doi: 10.1182/blood-2010-09-308064. [DOI] [PubMed] [Google Scholar]
- Edelson BT, Kc W, Juang R, Kohyama M, et al. Peripheral CD103+ dendritic cells form a unified subset developmentally related to CD8alpha+ conventional dendritic cells. J Exp Med. 2010;207:823–836. doi: 10.1084/jem.20091627. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farache J, Zigmond E, Shakhar G, Jung S. Contributions of dendritic cells and macrophages to intestinal homeostasis and immune defense. Immunol Cell Biol. 2013;91:232–239. doi: 10.1038/icb.2012.79. [DOI] [PubMed] [Google Scholar]
- Farache J, Koren I, Milo I, et al. Luminal bacteria recruit CD103+ dendritic cells into the intestinal epithelium to sample bacterial antigens for presentation. Immunity. 2013;38:581–595. doi: 10.1016/j.immuni.2013.01.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fasseu M, Tréton X, Guichard C et al (2010) Identification of restricted subsets of mature microRNA abnormally expressed in inactive colonic mucosa of patients with inflammatory bowel disease. PLoS One 5. pii: e13160 [DOI] [PMC free article] [PubMed]
- Gallegos AM, Bevan MJ. Central tolerance to tissue-specific antigens mediated by direct and indirect antigen presentation. J Exp Med. 2004;200:1039–1049. doi: 10.1084/jem.20041457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ganguly D, Haak S, Sisirak V, Reizis B. The role of dendritic cells in autoimmunity. Nat Rev Immunol. 2013;13:566–577. doi: 10.1038/nri3477. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ginhoux F, Liu K, Helft J, et al. The origin and development of nonlymphoid tissue CD103+ DCs. J Exp Med. 2009;206:3115–3130. doi: 10.1084/jem.20091756. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goldschneider I, Cone RE. A central role for peripheral dendritic cells in the induction of acquired thymic tolerance. Trends Immunol. 2003;4:77–81. doi: 10.1016/S1471-4906(02)00038-8. [DOI] [PubMed] [Google Scholar]
- Goto Y, Panea C, Nakato G, et al. Segmented filamentous bacteria antigens presented by intestinal dendritic cells drive mucosal Th17 cell differentiation. Immunity. 2014;40:594–607. doi: 10.1016/j.immuni.2014.03.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grayson MH. Lung dendritic cells and the inflammatory response. Ann Allergy Asthma Immunol. 2006;96:643–651. doi: 10.1016/S1081-1206(10)61061-7. [DOI] [PubMed] [Google Scholar]
- Gregorio J, Meller S, Conrad C. Plasmacytoid dendritic cells sense skin injury and promote wound healing through type I interferons. J Exp Med. 2010;207:2921–2930. doi: 10.1084/jem.20101102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guilliams M, Lambrecht BN, Hammad H. Division of labor between lung dendritic cells and macrophages in the defense against pulmonary infections. Mucosal Immunol. 2013;6:464–473. doi: 10.1038/mi.2013.14. [DOI] [PubMed] [Google Scholar]
- Gurney KB, Colantonio AD, Blom B, Spits H, Uittenbogaart CH. Endogenous IFN-alpha production by plasmacytoid dendritic cells exerts an antiviral effect on thymic HIV-1 infection. J Immunol. 2004;73:7269–7276. doi: 10.4049/jimmunol.173.12.7269. [DOI] [PubMed] [Google Scholar]
- Haniffa M, Ginhoux F, Wang XN, et al. Differential rates of replacement of human dermal dendritic cells and macrophages during hematopoietic stem cell transplantation. J Exp Med. 2009;206:371–385. doi: 10.1084/jem.20081633. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haniffa M, Shin A, Bigley V, et al. Human tissues contain CD141hi cross-presenting dendritic cells with functional homology to mouse CD103+ nonlymphoid dendritic cells. Immunity. 2012;37:60–73. doi: 10.1016/j.immuni.2012.04.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hashimi ST, Fulcher JA, Chang MH, Gov L, Wang S, Lee B. MicroRNA profiling identifies miR-34a and miR-21 and their target genes JAG1 and WNT1 in the coordinate regulation of dendritic cell differentiation. Blood. 2009;114:404–414. doi: 10.1182/blood-2008-09-179150. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Helft J, Ginhoux F, Bogunovic M, Merad M. Origin and functional heterogeneity of non-lymphoid tissue dendritic cells in mice. Immunol Rev. 2010;234:55–75. doi: 10.1111/j.0105-2896.2009.00885.x. [DOI] [PubMed] [Google Scholar]
- Henri S, Vremec D, Kamath A, et al. The dendritic cell populations of mouse lymph nodes. J Immunol. 2001;167:741–748. doi: 10.4049/jimmunol.167.2.741. [DOI] [PubMed] [Google Scholar]
- Hildner K, Edelson BT, Purtha WE, et al. Batf3 deficiency reveals a critical role for CD8alpha+ dendritic cells in cytotoxic T cell immunity. Science. 2008;322:1097–1100. doi: 10.1126/science.1164206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hochrein H, Shortman K, Vremec D, Scott B, Hertzog P, O’Keeffe M. Differential production of IL-12, IFN-alpha, and IFN-gamma by mouse dendritic cell subsets. J Immunol. 2001;166:5448–5455. doi: 10.4049/jimmunol.166.9.5448. [DOI] [PubMed] [Google Scholar]
- Jaensson E, Uronen-Hansson H, Pabst O, et al. Small intestinal CD103+ dendritic cells display unique functional properties that are conserved between mice and humans. J Exp Med. 2008;205:2139–2149. doi: 10.1084/jem.20080414. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jongbloed SL, Kassianos AJ, McDonald KJ, et al. Human CD141+ (BDCA-3)+ dendritic cells (DCs) represent a unique myeloid DC subset that cross-presents necrotic cell antigens. J Exp Med. 2010;207:1247–1260. doi: 10.1084/jem.20092140. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kadowaki N, Ho S, Antonenko S, Malefyt RW, et al. Subsets of human dendritic cell precursors express different Toll-like receptors and respond to different microbial antigens. J Exp Med. 2001;194:863–869. doi: 10.1084/jem.194.6.863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Keir ME, Stoddart CA, Linquist-Stepps V, Moreno ME, McCune JM. IFN-alpha secretion by type 2 predendritic cells up-regulates MHC class I in the HIV-1-infected thymus. J Immunol. 2002;168:325–331. doi: 10.4049/jimmunol.168.1.325. [DOI] [PubMed] [Google Scholar]
- Kim TH, Lee HK. Differential roles of lung dendritic cell subsets against respiratory virus infection. Immune Netw. 2014;14:128–137. doi: 10.4110/in.2014.14.3.128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klebanoff CA, Spencer SP, Torabi-Parizi P, et al. Retinoic acid controls the homeostasis of pre-cDC-derived splenic and intestinal dendritic cells. J Exp Med. 2013;210:1961–1976. doi: 10.1084/jem.20122508. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Klechevsky E, Morita R, Liu M, et al. Functional specializations of human epidermal Langerhans cells and CD14+ dermal dendritic cells. Immunity. 2008;29:497–510. doi: 10.1016/j.immuni.2008.07.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuipers H, Schnorfeil FM, Fehling HJ, Bartels H, Brocker T. Dicer-dependent microRNAs control maturation, function, and maintenance of Langerhans cells in vivo. J Immunol. 2010;185:400–409. doi: 10.4049/jimmunol.0903912. [DOI] [PubMed] [Google Scholar]
- Kuipers H, Schnorfeil FM, Brocker T. Differentially expressed microRNAs regulate plasmacytoid vs. conventional dendritic cell development. Mol Immunol. 2010;48:333–340. doi: 10.1016/j.molimm.2010.07.007. [DOI] [PubMed] [Google Scholar]
- Lahoud MH, Proietto AI, Gartlan KH, et al. Signal regulatory protein molecules are differentially expressed by CD8- dendritic cells. J Immunol. 2006;177:372–382. doi: 10.4049/jimmunol.177.1.372. [DOI] [PubMed] [Google Scholar]
- Lambrecht BN, Hammad H. Biology of lung dendritic cells at the origin of asthma. Immunity. 2009;31:412–424. doi: 10.1016/j.immuni.2009.08.008. [DOI] [PubMed] [Google Scholar]
- Li HS, Greeley N, Sugimoto N, Liu YJ, Watowich SS. miR-22 controls Irf8 mRNA abundance and murine dendritic cell development. PLoS One. 2012;7:e52341. doi: 10.1371/journal.pone.0052341. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu YJ. IPC: professional type 1 interferon-producing cells and plasmacytoid dendritic cell precursors. Annu Rev Immunol. 2005;23:275–306. doi: 10.1146/annurev.immunol.23.021704.115633. [DOI] [PubMed] [Google Scholar]
- Liu K, Nussenzweig MC. Origin and development of dendritic cells. Immunol Rev. 2010;234(1):45–54. doi: 10.1111/j.0105-2896.2009.00879.x. [DOI] [PubMed] [Google Scholar]
- Liu X, Zhan Z, Xu L, et al. MicroRNA-148/152 impair innate response and antigen presentation of TLR-triggered dendritic cells by targeting CaMKIIα. J Immunol. 2010;185:7244–7251. doi: 10.4049/jimmunol.1001573. [DOI] [PubMed] [Google Scholar]
- Lu TX, Hartner J, Lim EJ, et al. MicroRNA-21 limits in vivo immune response-mediated activation of the IL-12/IFN-gamma pathway, Th1 polarization, and the severity of delayed-type hypersensitivity. J Immunol. 2011;187(6):3362–3373. doi: 10.4049/jimmunol.1101235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Malissen B, Tamoutounour S, Henri S. The origins and functions of dendritic cells and macrophages in the skin. Nat Rev Immunol. 2014;14:417–428. doi: 10.1038/nri3683. [DOI] [PubMed] [Google Scholar]
- Manz MG, Traver D, Akashi K, et al. Dendritic cell development from common myeloid progenitors. Ann N Y Acad Sci. 2001;938:167–173. doi: 10.1111/j.1749-6632.2001.tb03586.x. [DOI] [PubMed] [Google Scholar]
- Martín P, del Hoyo GM, Anjuère F, et al. Concept of lymphoid versus myeloid dendritic cell lineages revisited: both CD8alpha(-) and CD8alpha(+) dendritic cells are generated from CD4(low) lymphoid-committed precursors. Blood. 2000;96:2511–2519. [PubMed] [Google Scholar]
- Mathan TS, Figdor CG, Buschow SI. Human plasmacytoid dendritic cells: from molecules to intercellular communication network. Front Immunol. 2013;4:372. doi: 10.3389/fimmu.2013.00372. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mehta A, Baltimore D. MicroRNAs as regulatory elements in immune system logic. Nat Rev Immunol. 2016;16:279–294. doi: 10.1038/nri.2016.40. [DOI] [PubMed] [Google Scholar]
- Merad M, Ginhoux F, Collin M. Origin, homeostasis and function of Langerhans cells and other langerin-expressing dendritic cells. Nat Rev Immunol. 2008;8:935–947. doi: 10.1038/nri2455. [DOI] [PubMed] [Google Scholar]
- Mi QS, Xu YP, Wang H, Qi RQ, Dong Z, Zhou L. Deletion of microRNA miR-223 increases Langerhans cell cross-presentation. Int J Biochem Cell Biol. 2013;45:395–400. doi: 10.1016/j.biocel.2012.11.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mildner A, Yona S, Jung S. A close encounter of the third kind: monocyte-derived cells. Adv. Immunol. 2013;120:69–103. doi: 10.1016/B978-0-12-417028-5.00003-X. [DOI] [PubMed] [Google Scholar]
- Mildner A, Chapnik E, Manor O. Mononuclear phagocyte miRNome analysis identifies miR-142 as critical regulator of murine dendritic cell homeostasis. Blood. 2013;121:1016–1027. doi: 10.1182/blood-2012-07-445999. [DOI] [PubMed] [Google Scholar]
- Mittag D, Proietto AI, Loudovaris T, et al. Human dendritic cell subsets from spleen and blood are similar in phenotype and function but modified by donor health status. J Immunol. 2011;186:6207–6217. doi: 10.4049/jimmunol.1002632. [DOI] [PubMed] [Google Scholar]
- Naik SH, Metcalf D, van Nieuwenhuijze A, et al. Intrasplenic steady-state dendritic cell precursors that are distinct from monocytes. Nat Immunol. 2006;7:663–671. doi: 10.1038/ni1340. [DOI] [PubMed] [Google Scholar]
- Nakano H, Yanagita M, Gunn MD. CD11c(+)B220(+)Gr-1(+) cells in mouse lymph nodes and spleen display characteristics of plasmacytoid dendritic cells. J Exp Med. 2001;194:1171–1178. doi: 10.1084/jem.194.8.1171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nestle FO, Turka LA, Nickoloff BJ. Characterization of dermal dendritic cells in psoriasis. Autostimulation of T lymphocytes and induction of Th1 type cytokines. J Clin Invest. 1994;94:202–209. doi: 10.1172/JCI117308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nestle FO, Di Meglio P, Qin JZ, Nickoloff BJ. Skin immune sentinels in health and disease. Nat Rev Immunol. 2009;9:679–691. doi: 10.1038/nri2622. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Connell RM, Chaudhuri AA, Rao DS, Baltimore D. Inositol phosphatase SHIP1 is a primary target of miR-155. Proc Natl Acad Sci USA. 2009;106:7113–7118. doi: 10.1073/pnas.0902636106. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Connell RM, Kahn D, Gibson WS, et al. MicroRNA-155 promotes autoimmune inflammation by enhancing inflammatory T cell development. Immunity. 2010;33:607–619. doi: 10.1016/j.immuni.2010.09.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Keeffe M, Hochrein H, Vremec D, et al. Mouse plasmacytoid cells: long-lived cells, heterogeneous in surface phenotype and function, that differentiate into CD8(+) dendritic cells only after microbial stimulus. J Exp Med. 2002;196:1307–1319. doi: 10.1084/jem.20021031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- O’Keeffe M, Hochrein H, Vremec D, et al. Dendritic cell precursor populations of mouse blood: identification of the murine homologues of human blood plasmacytoid pre-DC2 and CD11c+ DC1 precursors. Blood. 2003;101:1453–1459. doi: 10.1182/blood-2002-03-0974. [DOI] [PubMed] [Google Scholar]
- Persson EK, Uronen-Hansson H, Semmrich M, et al. IRF4 transcription-factor-dependent CD103(+)CD11b(+) dendritic cells drive mucosal T helper 17 cell differentiation. Immunity. 2013;38:958–969. doi: 10.1016/j.immuni.2013.03.009. [DOI] [PubMed] [Google Scholar]
- Plantinga M, Hammad H, Lambrecht BN. Origin and functional specializations of DC subsets in the lung. Eur J Immunol. 2010;40:2112–2118. doi: 10.1002/eji.201040562. [DOI] [PubMed] [Google Scholar]
- Plantinga M, Guilliams M, Vanheerswynghels M, et al. Conventional and monocyte-derived CD11b(+) dendritic cells initiate and maintain T helper 2 cell-mediated immunity to house dust mite allergen. Immunity. 2013;38:322–335. doi: 10.1016/j.immuni.2012.10.016. [DOI] [PubMed] [Google Scholar]
- Poulin LF, Henri S, de Bovis B, Devilard E, Kissenpfennig A, Malissen B. The dermis contains langerin+ dendritic cells that develop and function independently of epidermal Langerhans cells. J Exp Med. 2007;204:3119–3131. doi: 10.1084/jem.20071724. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poulin LF, Salio M, Griessinger E, et al. Characterization of human DNGR-1+ BDCA3+ leukocytes as putative equivalents of mouse CD8alpha+ dendritic cells. J Exp Med. 2010;207:1261–1271. doi: 10.1084/jem.20092618. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Probst HC, Lagnel J, Kollias G, van den Broek M. Inducible transgenic mice reveal resting dendritic cells as potent inducers of CD8+ T cell tolerance. Immunity. 2003;18:713–720. doi: 10.1016/S1074-7613(03)00120-1. [DOI] [PubMed] [Google Scholar]
- Proietto AI, van Dommelen S, Zhou P et al (2008) Dendritic cells in the thymus contribute to T-regulatory cell induction. Proc Natl Acad Sci 105:19869–19874 [DOI] [PMC free article] [PubMed]
- Satpathy AT, Wumesh KC, Albring JC, et al. Zbtb46 expression distinguishes classical dendritic cells and their committed progenitors from other immune lineages. J Exp Med. 2012;209:1135–1152. doi: 10.1084/jem.20120030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schlitzer A, McGovern N, Teo P, et al. IRF4 transcription factor-dependent CD11b+ dendritic cells in human and mouse control mucosal IL-17 cytokine responses. Immunity. 2013;38:970–983. doi: 10.1016/j.immuni.2013.04.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schulz O, Jaensson E, Persson EK, et al. Intestinal CD103+, but not CX3CR1+, antigen sampling cells migrate in lymph and serve classical dendritic cell functions. J Exp Med. 2009;206:3101–3114. doi: 10.1084/jem.20091925. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scott CL, Bain CC, Wright PB, et al. CCR2CD103 intestinal dendritic cells develop from DC-committed precursors and induce interleukin-17 production by T cells. Mucosal Immunol. 2014;8:327–339. doi: 10.1038/mi.2014.70. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Segura E, Valladeau-Guilemond J, Donnadieu MH, Sastre-Garau X, Soumelis V, Amigorena S. Characterization of resident and migratory dendritic cells in human lymph nodes. J Exp Med. 2012;209:653–660. doi: 10.1084/jem.20111457. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shi C, Liang Y, Yang J, et al. MicroRNA-21 knockout improve the survival rate in DSS induced fatal colitis through protecting against inflammation and tissue injury. PLoS One. 2013;8:e66814. doi: 10.1371/journal.pone.0066814. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shortman K, Heath WR. The CD8+ dendritic cell subset. Immunol Rev. 2010;234:18–31. doi: 10.1111/j.0105-2896.2009.00870.x. [DOI] [PubMed] [Google Scholar]
- Siegal FP, Kadowaki N, Shodell M, et al. The nature of the principal type 1 interferon-producing cells in human blood. Science. 1999;284:1835–1837. doi: 10.1126/science.284.5421.1835. [DOI] [PubMed] [Google Scholar]
- Singh UP, Murphy AE, Enos RT, et al. miR-155 deficiency protects mice from experimental colitis by reducing T helper type 1/type 17 responses. Immunology. 2014;143:478–489. doi: 10.1111/imm.12328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smolen JS, Aletaha D, Koeller M, Weisman MH, Emery P. New therapies for treatment of rheumatoid arthritis. Lancet. 2007;370:1861–1874. doi: 10.1016/S0140-6736(07)60784-3. [DOI] [PubMed] [Google Scholar]
- Sonkoly E, Wei T, Janson PC, et al. MicroRNAs: novel regulators involved in the pathogenesis of psoriasis? PLoS One. 2007;2:e610. doi: 10.1371/journal.pone.0000610. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Spadoni I, Iliev ID, Rossi G, Rescigno M. Dendritic cells produce TSLP that limits the differentiation of Th17 cells, fosters Treg development, and protects against colitis. Mucosal Immunol. 2012;5:184–193. doi: 10.1038/mi.2011.64. [DOI] [PubMed] [Google Scholar]
- Spörri R, Reis e Sousa C. Inflammatory mediators are insufficient for full dendritic cell activation and promote expansion of CD4+ T cell populations lacking helper function. Nat Immunol. 2005;6:163–170. doi: 10.1038/ni1162. [DOI] [PubMed] [Google Scholar]
- Stanczyk J, Pedrioli DM, Brentano F, et al. Altered expression of MicroRNA in synovial fibroblasts and synovial tissue in rheumatoid arthritis. Arthritis Rheum. 2008;58:1001–1009. doi: 10.1002/art.23386. [DOI] [PubMed] [Google Scholar]
- Steinman RM, Cohn ZA. Identification of a novel cell type in peripheral lymphoid organs of mice. I. Morphology, quantitation, tissue distribution. J Exp Med. 1973;137:1142–1162. doi: 10.1084/jem.137.5.1142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Steinman RM, Cohn ZA. Identification of a novel cell type in peripheral lymphoid organs of mice. II. Functional properties in vitro. J Exp Med. 1974;139:380–397. doi: 10.1084/jem.139.2.380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sun Y, Varambally S, Maher CA. Targeting of microRNA-142-3p in dendritic cells regulates endotoxin-induced mortality. Blood. 2011;117:6172–6183. doi: 10.1182/blood-2010-12-325647. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Taganov KD, Boldin MP, Chang KJ, Baltimore D. NF-kappaB-dependent induction of microRNA miR-146, an inhibitor targeted to signaling proteins of innate immune responses. Proc Natl Acad Sci USA. 2006;103:12481–12486. doi: 10.1073/pnas.0605298103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Takagi T, Naito Y, Mizushima K, et al. Increased expression of microRNA in the inflamed colonic mucosa of patients with active ulcerative colitis. J Gastroenterol Hepatol. 2010;25(Suppl 1):S129–S133. doi: 10.1111/j.1440-1746.2009.06216.x. [DOI] [PubMed] [Google Scholar]
- Tang Y, Luo X, Cui H, et al. MicroRNA-146A contributes to abnormal activation of the type I interferon pathway in human lupus by targeting the key signaling proteins. Arthritis Rheum. 2009;60:1065–1075. doi: 10.1002/art.24436. [DOI] [PubMed] [Google Scholar]
- Tserel L, Runnel T, Kisand K, et al. MicroRNA expression profiles of human blood monocyte-derived dendritic cells and macrophages reveal miR-511 as putative positive regulator of Toll-like receptor 4. J Biol Chem. 2011;286:26487–26495. doi: 10.1074/jbc.M110.213561. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vandenabeele S, Hochrein H, Mavaddat N, Winkel K, Shortman K. Human thymus contains 2 distinct dendritic cell populations. Blood. 2001;97:1733–1741. doi: 10.1182/blood.V97.6.1733. [DOI] [PubMed] [Google Scholar]
- Varol C, Vallon-Eberhard A, Elinav E, et al. Intestinal lamina propria dendritic cell subsets have different origin and functions. Immunity. 2009;31:502–512. doi: 10.1016/j.immuni.2009.06.025. [DOI] [PubMed] [Google Scholar]
- Vremec D, Pooley J, Hochrein H, Wu L, Shortman K. CD4 and CD8 expression by dendritic cell subtypes in mouse thymus and spleen. J Immunol. 2000;164:2978–2986. doi: 10.4049/jimmunol.164.6.2978. [DOI] [PubMed] [Google Scholar]
- Watanabe N, Wang YH, Lee HK, et al. Hassall’s corpuscles instruct dendritic cells to induce CD4+CD25+ regulatory T cells in human thymus. Nature. 2005;436:1181–1185. doi: 10.1038/nature03886. [DOI] [PubMed] [Google Scholar]
- Wilson NS, Young LJ, Kupresanin F, et al. Normal proportion and expression of maturation markers in migratory dendritic cells in the absence of germs or Toll-like receptor signaling. Immunol Cell Biol. 2008;86:200–205. doi: 10.1038/sj.icb.7100125. [DOI] [PubMed] [Google Scholar]
- Wu L, Vremec D, Ardavin C, et al. Mouse thymus dendritic cells: kinetics of development and changes in surface markers during maturation. Eur J Immunol. 1995;25:418–425. doi: 10.1002/eji.1830250217. [DOI] [PubMed] [Google Scholar]
- Wu L, Li CL, Shortman K. Thymic dendritic cell precursors: relationship to the T lymphocyte lineage and phenotype of the dendritic cell progeny. J Exp Med. 1996;184:903–911. doi: 10.1084/jem.184.3.903. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wu L, D’Amico A, Hochrein H, O’Keeffe M, Shortman K, Lucas K. Development of thymic and splenic dendritic cell populations from different hemopoietic precursors. Blood. 2001;98:3376–3382. doi: 10.1182/blood.V98.12.3376. [DOI] [PubMed] [Google Scholar]
- Wu F, Zikusoka M, Trindade A, et al. MicroRNAs are differentially expressed in ulcerative colitis and alter expression of macrophage inflammatory peptide-2 alpha. Gastroenterology. 2008;135:1624–1635. doi: 10.1053/j.gastro.2008.07.068. [DOI] [PubMed] [Google Scholar]
- Xiao C, Rajewsky K. MicroRNA control in the immune system: basic principles. Cell. 2009;136:26–36. doi: 10.1016/j.cell.2008.12.027. [DOI] [PubMed] [Google Scholar]
- Xue X, Feng T, Yao S, et al. Microbiota downregulates dendritic cell expression of miR-10a, which targets IL-12/IL-23p40. J Immunol. 2011;187:5879–5886. doi: 10.4049/jimmunol.1100535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yan HX, Li WW, Zhang Y, et al. Accumulation of FLT3(+) CD11c (+) dendritic cells in psoriatic lesions and the anti-psoriatic effect of a selective FLT3 inhibitor. Immunol Res. 2014;60:112–126. doi: 10.1007/s12026-014-8521-4. [DOI] [PubMed] [Google Scholar]
- Zhou H, Huang X, Cui H, et al. miR-155 and its star-form partner miR-155* cooperatively regulate type I interferon production by human plasmacytoid dendritic cells. Blood. 2010;116:5885–5894. doi: 10.1182/blood-2010-04-280156. [DOI] [PubMed] [Google Scholar]
- Zhou H, Xiao J, Wu N, et al. MicroRNA-223 regulates the differentiation and function of intestinal dendritic cells and macrophages by targeting C/EBPβ. Cell Rep. 2015;13:1149–1160. doi: 10.1016/j.celrep.2015.09.073. [DOI] [PubMed] [Google Scholar]