

Abstract
The rapid development of the engineered nanomaterial (ENM) manufacturing industry has accelerated the incorporation of ENMs into a wide variety of consumer products across the globe. Unintentionally or not, some of these ENMs may be introduced into the environment or come into contact with humans or other organisms resulting in unexpected biological effects. It is thus prudent to have rapid and robust analytical metrology in place that can be used to critically assess and/or predict the cytotoxicity, as well as the potential genotoxicity of these ENMs. Many of the traditional genotoxicity test methods [e.g. unscheduled DNA synthesis assay, bacterial reverse mutation (Ames) test, etc.,] for determining the DNA damaging potential of chemical and biological compounds are not suitable for the evaluation of ENMs, due to a variety of methodological issues ranging from potential assay interferences to problems centered on low sample throughput. Recently, a number of sensitive, high-throughput genotoxicity assays/platforms (CometChip assay, flow cytometry/micronucleus assay, flow cytometry/γ-H2AX assay, automated ‘Fluorimetric Detection of Alkaline DNA Unwinding’ (FADU) assay, ToxTracker reporter assay) have been developed, based on substantial modifications and enhancements of traditional genotoxicity assays. These new assays have been used for the rapid measurement of DNA damage (strand breaks), chromosomal damage (micronuclei) and for detecting upregulated DNA damage signalling pathways resulting from ENM exposures. In this critical review, we describe and discuss the fundamental measurement principles and measurement endpoints of these new assays, as well as the modes of operation, analytical metrics and potential interferences, as applicable to ENM exposures. An unbiased discussion of the major technical advantages and limitations of each assay for evaluating and predicting the genotoxic potential of ENMs is also provided.
Introduction
The global value of engineered nanomaterials (ENMs), including nano-enabled devices and products is predicted to exceed $4.4 trillion by 2018 (1). The rapid expansion of the nanomaterial manufacturing industry and the increasing use of ENMs in a wide variety of industrial processes and consumer products may result in the intentional or inadvertent release of these novel materials into the environment. The unique physicochemical properties of ENMs (e.g. carbon nanotubes and metal/metal oxide nanoparticles, etc.) that make these new materials particularly attractive for use in biology, medicine, drug development, agriculture and in other fields, require that their potential biological effects (e.g. toxicity, alteration of cell signaling pathways, etc.) are fully characterised and/or predictively modelled in environmental and biological systems as accurately as possible (2). This vast research effort, known as nanotoxicology, necessitates the development and application of rapid, robust and reproducible protocols and assays applicable for ENM hazard and risk assessment. Both in vitro and in vivo quantitative assessments of potential cytotoxic and genotoxic responses generated from ENM exposures to cells and tissues are ultimately required in order to obtain the full compendium of data necessary to inform unbiased nanosafety risk assessments and regulatory decision making (3).
However, the current status of nanotoxicology, what we currently know and can confirm, cannot contend with the vast numbers of ENMs that make their way to the market (4). Moreover, solely using in vivo assessments, which are the gold standard in ENM hazard assessments, can have ethical and cost limitations; thus accurate and predictive in vitro assays are of great importance (5). Still, traditional in vitro toxicity evaluations consist of one parameter or treatment at a time, an approach which is highly ineffective and time consuming. High-throughput and high content nanotoxicity assays can overcome this limitation by providing more data and endpoints to understand potential outcomes and prioritise hazardous materials (6). Implementing high-throughput/content assays will enable assessment of structure–bioactivity relationships that may result in a deeper understanding of ENM toxicity and reduce the need for costly animal studies (3).
Due to the vast numbers, types and formulations of ENMs being manufactured on an industrial scale and in academic laboratories, there is a critical need to rapidly characterise these materials in terms of their physicochemical attributes and biological activities. There are very few assays that have been specifically designed for evaluating the cytotoxicity or genotoxicity of ENMs; in fact, the current paradigm is to adapt and modify existing assays that have been developed for chemical toxicity and apply those assays to the evaluation of ENMs (7,8). Unfortunately, this does not work seamlessly and often the resulting toxicity mechanism is obscured or the resulting data are flawed by artefacts due to ENM interference with components in the assay or the assay readout (9,10). However, one excellent example demonstrating the steps required for properly modifying a biological cytotoxicity assay for use with ENMs was recently reported (11). In this work, the authors described a successful approach for optimizing and modifying the control parameters of the highly utilised 3-(4,5-dimethylthiazol-2-yl)-5-(3-carboxy-methoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazolium (MTS) cell-viability assay to make it suitable for testing the cytotoxicity of ENMs in a 96-well plate format. However, assays similar to this in which the mechanism of toxicity is well characterised and understood, that are conducted on a high-throughput platform and that are capable of producing high-quality, reproducible and archivable data are challenging to develop and even more difficult to validate.
Genotoxicity hazard and risk assessment is an increasingly integral part of ENM safety evaluations. The mechanisms by which ENMs have the possibility of interacting with the genetic machinery and causing gene mutations, chromosomal aberrations and/or general damage to DNA are numerous and include both direct and indirect modes of action. Briefly, non-functionalised ENMs of a certain size (e.g. ≤ 9 nm in diameter) may directly interact with nuclear DNA following transportation through nuclear pore complexes (12–14) or with DNA and/or the spindle apparatus when the nuclear envelope breaks down during mitosis. ENMs that have entered the cell nucleus may also produce reactive oxygen species (ROS) that can directly damage DNA. In particular, ENMs composed of transition metals may catalyze (via Fenton reaction chemistry) the production of hydroxyl radicals (·OH) which are potent DNA, as well as protein and lipid, damaging agents (15,16). Probably the most established mechanism by which ENMs have the potential to produce genetic damage is via their capacity to induce oxidative stress (17). Exposure to ENMs may result in systemic inflammation and the resulting ‘oxidative burst’ in response to the inflammation may lead to the further production of DNA damaging ROS. The biological mechanisms through which ENMs have been shown or postulated to induce DNA damage have been thoroughly described in several recent reviews (9,18–21) and will not be further expanded upon here. ENM-induced genotoxicity can manifest as point mutations, clastogenicity, aneugenicity, aneuploidy, DNA adducts, modifications or strand breaks (SBs), DNA replication interference or in the inhibition of DNA repair, among other aberrations (20,22,23). Some of these endpoints are associated with increased cancer risk and other adverse health effects. The characterisation and measurement of these genotoxicological endpoints requires a battery of mechanism-based, high-throughput robust assays/methods (24,25). No single genotoxicity assay by itself can be used to compare, rank and predict the genotoxic potential or even determine and validate the biologically relevant results for all of the different types of ENMs that are currently available and forms of DNA damage that may arise.
The alkaline comet assay and the micronucleus (MN) assay are the methodologies most commonly utilised for measuring, characterizing and predicting the genotoxicity of ENMs (25–30). In their traditional or partially automated formats, the comet and MN assays can be considered low to medium throughput at best, in terms of sample capacity, as well as in terms of sample analysis. The γ-H2AX staining assay (31–33) and the ‘Fluorimetric Detection of Alkaline DNA Unwinding’ (FADU) assay (34–37) are also low to medium throughput assays which have commonly been used to detect and measure the DNA damaging potential of chemical agents, but are now being applied to the determination of the genotoxicity of ENMs (38–42). All these assays have been successfully applied in genetic toxicology (in vitro and in vivo), population biomonitoring, eco-toxicology, DNA damage and repair research and/or in cancer risk assessment for potential chemical and biological genotoxins. One of the most recent additions to the current suite of genotoxicity assays is the ToxTracker reporter assay (43). This assay provides a distinct, alternative measurement of genetic damage by identifying cell signalling pathways that are activated following chemical exposure.
Over the last decade, these established genotoxicity assays and associated instrumentation have been substantially modified and optimised so that the assays now incorporate standard 96-well plates (or 384-well plates) and/or flow cytometric sample processing/detection, along with fully-automated data processing and image analysis that give increased sample capacity and throughput, and are less labour-intensive. This review focuses on the five new high-throughput versions (CometChip, flow cytometry/MN, flow cytometry/γ-H2AX, automated FADU, ToxTracker reporter) of the established assays and their specific features and advancements that allow for efficient measurement of DNA damage, chromosomal damage or DNA damage signalling induced by ENMs. For each of the new high-throughput genotoxicity assays and platforms, a general overview of the established or traditional assay methodology is given initially, followed by a detailed description and discussion of the fundamental measurement principles and endpoints, general operating procedures, measurement sensitivity and potential measurement interferences as applied to ENMs. Finally, an unbiased discussion of the major technical advantages and limitations of the assays for evaluating and predicting the genotoxic potential of ENMs is also provided in each section.
The CometChip platform
The comet assay is one of the most widely used assays for evaluating DNA damage associated with ENM exposures with more than 100 published papers utilizing this technique (44). The core principle of the comet assay is based on damaged DNA migrating more readily in an agarose matrix than intact supercoiled DNA under electrophoretic conditions (45,46). After lysis of agarose-embedded cells, DNA remains connected in a supercoiled matrix called a nucleoid. During electrophoresis, supercoiled or undamaged DNA remain in the nucleoid core whereas broken or relaxed DNA strands migrate towards the anode creating a structure resembling a comet. The comet tail length and intensity are indicators of the level of injury incurred by cells either by direct or indirect insults to the DNA.
There are two types or versions of the comet assay that are commonly utilised—the alkaline and the neutral comet assay. The alkaline comet assay enables the detection and measurement of single strand breaks (SSBs), alkali sensitive sites (ALS) and abasic sites, whereas the neutral comet assay has been shown to be effective in detecting both single and double strand breaks (DSBs) (47,48). Modifications of the comet assay to detect various base lesions using lesion specific enzymes allows higher sensitivity, specificity and potentially the identification of the damage i.e. oxidative stress (49).
Most labs use image analysis software to score imaged comets individually using common parameters of DNA damage such as tail length, the percent DNA in tail (% tail DNA) and the tail moment (the product of the tail length and the fraction of total DNA in the tail). Although some researchers prefer the tail moment as a common descriptor, the percent tail DNA is the most common metric (50). Although widely accepted, the traditional comet assay tends to be laborious and can have high sample-to-sample variation due to use of separate glass slides and scoring differences between individuals (51). The low throughput and poor reproducibility of the traditional comet assay make it difficult to conduct comprehensive investigations that are needed for evaluating not only multiple ENMs, but also the potential effects from multiple physicochemical properties (e.g. size, charge and shape) (52) with this particular assay.
There have been many different approaches for enhancing the sample throughput of the traditional comet assay (53). Some of these approaches represent slight adaptations of the traditional assay while some approaches have required considerable assay modification and complex sample preparation steps (53). For example, the comet assay throughput can be increased by using 12 mini gels per glass side instead of the normal two gels per glass slide (54). The increase in the number of gels/slide requires fewer cells and smaller sample volumes to perform the assay. A recent trend is the use of mini gels placed onto polyester plastic films (Gelbond®) instead of onto glass slides (55–57). These films do not need precoating with agarose prior to sample application, the films do not break and after electrophoresis, the films can be fixed and stored for at least 1 year before rehydration and staining (57). Using these plastic films, 12-, 48- and 96 mini gels/film can be prepared and processed; 384 mini gels can be processed in total if four films containing 96 mini gels/film are placed into one large electrophoresis tank. A potentially more complex version of the 96 mini gels/film format based on multi-chamber microtitre plates has also been reported in the literature (58,59). In this instance, sample processing involved separating the walls of a multichamber plate from the bottom of the plate enabling fully automated comet scoring at greater efficiency (60).
In light of the need for nanotoxicity tools capable of providing high-throughput quantitative measurements of genotoxicity, this section describes the utility and versatility of the CometChip, a 96-well microfabricated high-throughput platform for determining ENM-mediated DNA SSBs in single cells (52) (Figure 1). The CometChip platform was developed at Massachusetts Institute of Technology (MIT) in the Engleward laboratory (61–63). Originally designed to assess chemical- and radiation-induced genotoxicity, the system now enables researchers to detect and measure abasic sites, DNA–DNA or DNA–protein crosslinks, SSBs and DSBs generated by ENM exposures. With slight modification to allow for ENM–cell interaction, the CometChip platform allows parallel processing of different types of ENM exposures in one experiment which greatly reduces work load, enhances productivity and reduces experimental variability (61). In a recent study examining the consistency and sensitivity of the CometChip system, the calculated inter-sample coefficient of variation (CV) was 5–10% (64). Conversely, the inter-sample CV can be as high as 15–30% using the traditional comet assay (61,65,66). Thus, the CometChip platform provides the high-throughput capacity and reproducibility required to determine and rank ENM genotoxic potential by investigating multiple ENM properties concurrently. The platform’s versatility also allows for the detection of other forms of DNA damage such as oxidatively modified bases using DNA repair glycosylases (67). The next section is an overview describing the application of the CometChip platform to assess the genotoxicity of ENMs.
Figure 1.

CometChip Platform Assembly and Protocol. (A) Using a microfabricated stamp, microwells are created in molten agarose. Once the agarose gel is set, the stamp is removed revealing patterned microwells. To create individual conditions or exposures, a bottomless 96-well plate is placed on the gel and secured. (B) ENM suspensions are prepared in distilled water by using sonication energy to reduce agglomerate size. (C) ENM exposure to either suspension or adherent cells occurs in a separate 96-well plate for 4 or 24 h. (D) The ENM-exposed cells are loaded into the microwells within the CometChip and electrophoresis is performed. The CometChip is then imaged using an automated imager or microscope. Images are then scored using MatLab software.
CometChip preparation and application
The CometChip enables the simultaneous evaluation of multiple ENM exposures by using a microwell array (Figure 1A). This feature is achieved by using a polydimethylsiloxane (PDMS) negative mold or stamp that has arrayed micropillars for patterning micro-wells within molten agarose. The PDMS stamp is placed into 1% (mass fraction) normal melting point agarose applied to a gel bond film positioned in a square petri dish and allowed to set for 30 min. The stamp is removed after the agarose polymerizes revealing a 300-μm thick gel containing the arrayed microwells. To create separate treatments or ENM exposures, a bottomless 96-well plate can be attached to the agarose gel/film using mechanical force.
An important aspect of ENM–cell interactions and experiment-to-experiment reproducibility is the standardisation of the colloidal preparation procedures. Sonication, which minimizes formed agglomerate size, is a well-known method for dispersing ENMs before diluting into the appropriate cell culture media for cell exposure (68). Likewise, colloidal characterization using dynamic light scattering (DLS) or other methods are equally important to determine the hydrodynamic diameter, zeta potential, conductivity and other parameters of the formed agglomerates. To ensure adequate ENM–cell interactions, with either adherent or suspension cell lines, exposures are performed in a separate 96-well plate at the desired exposure duration. A multichannel pipette is used to transfer the exposed cells into the CometChip, where one or more cells are captured in each microwell by gravitational settling. A thin layer of molten agarose is applied to the gel providing a protective coating for the nuclei during electrophoresis. The gels are neutralised and stained using common DNA fluorescent binding dyes such as SYBR Gold, then imaged using an automated fluorescent microscope. The collected images are then scored using MATLAB or another commercially available comet scoring software.
Comparison of the CometChip and the traditional comet assay—image acquisition and scoring
Imaging the CometChip involves acquiring images of each condition/well using a fluorescent microscope coupled to an automated stage. Fluorescent microscopes are standard instruments in most laboratories, however stage automation or fully automated imaging systems may not be available, which may reduce throughput and subsequently diminish the utility of the CometChip platform. However, upgrading or retrofitting a standard fluorescent microscope with an automated stage can be done at little cost. Given the format of the traditional comet assay, where individual mini gels exist on one slide and comets may lie on various focal planes, imaging can be tedious. A salient feature of the CometChip is the microfabricated design and arrayed format, which positions comets in one focal plane providing at least 50 comets when using the 4× objective. This aspect of the CometChip reduces imaging time and significantly improves data analysis by 100 fold (64).
Recently, automated scoring software such as OpenComet or CellProfiler, have significantly evolved, reducing imaging analysis time and potentially user bias associated with the traditional comet assay (69,70). Although automated scoring software is available now for the assessment of the traditional comet assay, overlapping and out of focus comets can decrease the number of measureable comets, as each comet requires its own image; therefore numerous replicates are required for statistical analysis. Images obtained from the CometChip are scored or analyzed using customised MATLAB software, which allows the identification of head/tail transitions, comets specifically within the arrayed area, and the quantification of DNA damage in each comet. For example, two images at the 4× objective provides 100 comets for scoring, whereas 50 images would be needed from individual slides of the traditional comet assay. As slide-to-slide variation due to different experimental conditions (i.e. buffer preparation) can be an issue, variability within the traditional comet assay has been reported (71).
Key advantages and limitations of the CometChip technology
A major advantage of the CometChip platform is the microfabrication/arrayed design, which allows for the concurrent evaluation of several types and concentrations of ENMs. The CometChip platform was successfully utilised to determine the genotoxic potential of five industrially relevant ENMs [zinc oxide nanoparticles (ZnO NPs), silver nanoparticles (AgNPs), cerium oxide nanoparticles (CeO2 NPs), iron oxide nanoparticles (Fe2O3 NPs), and amorphous silica nanoparticles (SiO2 NPs)] in two different cell lines: H9T3 Chinese hamster ovary adherent cells and TK6 human lymphoblastoid suspension cells (52). In agreement with previous studies (72), significant levels of SSBs (~80% DNA in tail) were found in the TK6 cells after 4 h exposure to ZnO NPs (20 μg/ml). At the same concentration, exposure to AgNPs resulted in less SSBs (~20% DNA in tail) in TK6 cells. The observed differences in the genotoxic profiles for ZnO NPs and AgNPs were presumably due to compensatory DNA repair mechanisms, impaired DNA repair or cell specific differences and dosimetry. Importantly, monitoring cytotoxicity is essential in genotoxicity assessments as high levels of toxicity can confound genotoxicity results. In both the ZnO NP and AgNP exposures, we observed a decline in cellular viability and proliferation of less than 30%, which is an acceptable level of toxicity when using the comet assay (73).
A potential limitation of the CometChip platform is the high cell density required for sufficient cell loading. For 100% cell loading, it is recommended to have a cell density of 104–106 cells, which is magnitudes higher than the traditional comet assay (74). This density may be difficult to achieve if sample amounts and volumes are low. Moreover, some cell lines that tend to aggregate can make cell loading difficult resulting in inefficient loading and empty wells in the CometChip. Thus, additional handling or processing of these cell types is necessary before loading into the CometChip platform. While using the CometChip to measure ENM mediated genotoxicity in adherent cell lines is mostly straightforward, its use with suspension cell lines can be problematic. Separating the ENMs from the cell suspension after exposure is impossible causing potential artifacts within the CometChip gel that may interfere with imaging. Gentle washing of the CometChip with warm phosphate buffered saline (PBS) after cell loading can help alleviate this problem by rinsing away excess cells, cellular debris and residual ENMs on the gel surface. This leads to a separate, but related issue concerning residual ENMs interacting with the naked DNA or nuclei after membrane dissolution during the lysis process; a problem that has been noted with the traditional comet assay (7). However, recent studies revealed that ENMs in close proximity to naked DNA do not influence the number of strand breaks in comparison to whole cell exposures using a range of concentrations (75). Finally, the cost of the CometChip system, complete with prefabricated gels and electrophoresis system, which is now commercially available from Trevigen may be cost prohibitive in some labs. However, when one accounts for the time saved, the amount of samples processed, and the information obtained by using high-throughput systems such as the CometChip, it may be a good investment for certain labs dealing with high sample volumes.
In totality, the CometChip platform is an important new high-throughput tool for nanogenotoxicity studies, which can help characterise the relationship between ENM physicochemical properties and potential biological responses in cells and tissues. While we have demonstrated the applicability of the CometChip system in nanogenotoxicity studies, other research areas such as nanomedicine, human biomonitoring, molecular epidemiology and drug safety fields could benefit from this robust system. High-throughput testing for potential DNA damage in human cells in clinical, epidemiological and drug development studies could provide diagnostic and/or prognostic insights into disease progression and related drug treatments.
The flow cytometry/MN assay
The in-vitro MN assay allows determination of gross chromosomal damage following exposure to an exogenous agent. The assay primarily involves scoring for micronuclei (MNi), which are manifested as miniature nuclei resulting from lagging acentric chromosomal fragments or lagging whole chromosomes, which fail to get incorporated into the main daughter nuclei during cell division. The type of chromosomal damage induced, can be identified by labelling the centromeric regions of chromosomes, whereby the centromere negative or positive MNi allows the determination of clastogenic (chromosome breakage) or aneugenic (chromosome loss) events, respectively (76).
The in vitro MN assay is a well-established, reliable, accurate and reproducible endpoint in nano(geno)toxicology, the use of which has steadily increased over the last few decades (16,77,78). However, the labour intensive and potentially subjective process of manually scoring micronuclei (MNi) has led to strong interest in developing automated systems to score MNi; a diagnostic marker for cancer in mammalian and human cells.
Out of various scoring approaches available for the assessment of the in vitro MN assay, the flow cytometry (FC) method provides the possibility of automation and high-throughput screening (79). Flow cytometry based MN (FCMN) analysis is based on the principle that lysis of the cell membrane and subsequent treatment with a DNA–dye enables discrimination between the nucleus and the MNi based on their differential fluorescence intensities (80).
The FCMN system enables scoring of the MNi frequency endpoint based on the analysis of ≥5000 cells (including those prepared in 96-well plates) providing rapid data acquisition thereby enhancing statistical power. This is proving to be advantageous and highly valuable not only for scoring weak genotoxins, but also for screening and determining the lowest observed genotoxic effects in non-linear dose-responses (79). Other advantages of the automated flow cytometric techniques, besides the generation of results in a short time interval, include simultaneous measurement of various parameters including frequency of MNi, DNA distribution of MNi, cell cycle, cytotoxicity and flow sorting for further analysis of MNi. Additionally, FC has the potential to determine the mode of action for a specific genotoxic agent; namely aneugenicity versus clastogenicity based on the ability of aneugens to increase the frequency of hypodiploid nuclei (79,81). However, the sensitivity of aneuploidy detection is still under evaluation.
Although FC allows simultaneous exploration of multiple endpoints and its speed of analysis represent significant advantages, the FCMN methodology presents several limitations (82) particularly when used for the genotoxicity evaluation of ENMs (83), which are discussed here in detail.
Confounding factors that interfere with MNi scoring
The use of FC for MNi analysis involves lysing the cells to release MN which can also result in the release of ENMs (sequestered within cells), nuclear fragments (from apoptotic cells/apoptotic bodies), chromosomes (in metaphase/anaphase) from cells or ENMs bound to DNA fragments, all of which could appear as false positives, thus confounding the MNi frequency results induced by a test ENM (84–87). Although there have been significant developments with regards to the incorporation of fluorescent dyes that discriminate between MNi and chromatin from apoptotic cells, contamination by fluorescent/auto-fluorescent ENMs particularly, fluorescent dye-loaded NPs, quantum dots, SWCNTs and gold clusters could potentially confound the FCMN frequency results (38,88,89). Thus plausible interference and/or cross-talk between different dyes/ENMs could pose a challenge in the interpretation of MNi data sets. However, one way to correct for potential interferences is to simultaneously combine cell cycle data with MNi data to overrule inaccuracies in the evaluation of the genotoxic effects of ENMs (90).
Influence of ENM agglomeration on dose–response data sets
The prepared suspension of nuclei and MNi used for FC measurements can be cross-contaminated with nonspecific debris and test ENMs (during centrifugation steps), which may skew the results if they cause scatter and overlap with the MNi population within a defined FC acquisition gate. Although nonspecific debris can often be gated out with ease, the degree of ENM agglomeration, which is a function of concentration, could vary between different doses of ENM exposure. This may have to be resolved by altering the FC settings to accommodate or eliminate a distinct population comprising those agglomerates, which could cause discrepancies in the analysis of results in dose–response data sets. Indeed, agglomerates comprised of small AgNPs have been shown to interact with laser light to emit a strong far red fluorescence signal due to their capacity to undergo surface plasmon resonance (83).
Flow cytometric versus microscopic evaluation of micronuclei frequency
Some discrepancies in the FCMN based genotoxic evaluation of ENMs may stem from the spectral properties from specific ENMs as observed by Zucker et al. (83). In this study, the authors noted that the MNi formation frequency measured using the FCMN assay was greater than the MNi formation frequency measured by the traditional microscopy-based MN assay. This increase was attributed to the far-red fluorescence increase in the 700–800 nm range (approximately 100 times the background) and the side-scattering effects of the citrate-coated AgNPs, which is particle size-dependent phenomenon (83).
Collins et al. (82) have also observed inconsistent results between FC and standard microscopy in mouse lymphoma cells. The higher MNi frequency observed in the FC-based evaluation as compared to other automated/microscopic systems is due to the flow cytometer segregating each MN encountered as an independent event. This can be problematic as (depending on the level of insult) multiple MNi can arise in a single cell. The flow cytometry system will score each of these MNi as a separate event, whilst microscopic evaluation will score a MN positive cell as one event irrespective of whether it presents with one or more MNi (as required by the OECD guidelines). Additionally, other related nuclear anomalies, e.g. nuclear budding, which may present as a consequence to the endocytotic uptake of ENMs could go unnoticed and cannot be scored using FCMN, which otherwise could be recorded along with the MNi during visual inspection (91).
Restriction to mononucleate cell scoring
Another challenge when using the FCMN assay is that it is not possible to discriminate between binucleate and mononucleate cells. This means that cells which present a MNi as a consequence of undergoing nuclear division will not be taken into account by the FCMN assay. This can have implications as the endocytosed ENMs whether sequestered in the cytoplasm or in the lysosomes can physically hinder or perturb cell division kinetics, which undoubtedly will go unnoticed as the FCMN assay is confined to the evaluation of MNi frequency in mononucleate cells only. Indeed, cell cycle arrest at higher concentrations of AgNP exposure as indicated by the decreased binucleate frequency have been observed, necessitating the need to scrutinise the sole reliance on FC-based genotoxic evaluation, which does not take binucleate cell scoring into consideration (92). However, scoring for mononucleated cells has its own advantage for biomonitoring purposes as it reflects in vivo damage present before the start of cell cultures. Finally, the FCMN assay does not provide the option of reanalysing the prepared sample, if desired at a later stage, as the processed sample cannot be preserved or archived.
Key advantages and limitations of the technology
The FCMN clearly presents significant advantages by enabling the high-throughput, rapid screening of ENMs and other potential genotoxins in conjunction with multiple end-point measurement with a 2- or 3-day turnaround time. Besides genotoxicity evaluations, FCMN is also a useful tool for biomonitoring studies and can be used for rapid assessment of base-line MNi frequency in human populations as increased baseline MNi frequency has been shown by various studies (including the International Human Micronucleus (HUMN) Project), to be predictive biomarker of cancer risk (93–95). Genomic instability arising as a result of environmental and occupational exposures to chemical and physical agents, genetic predisposition to spontaneous chromosomal damage, environment–gene interactions, adverse lifestyles factors and conditions, age, sex etc can be evaluated by means of population studies, utilising FCMN (93).
However, currently the technology is still embryonic in terms of application to ENM testing and presents certain challenges. One of the major difficulties for FCMN assay platforms lies in discriminating MNi induction as a result of genotoxic response as opposed to the formation of ENM/apoptotic entities (79). This can however be overcome by encompassing tests that pertain to the visual analysis of cells to generate robust, reliable and meaningful data sets and to avoid conflicting results. In conclusion, the emergence of new automated technologies such as the FCMN assay are presenting some significant advantages to the field. There remains a requirement for further modification and validation of the FCMN assay to enhance its robustness for the genotoxicity assessment of ENMs, but it is important to note that the limited data reported in the literature indicates it is a valuable advance to increase the throughput of the MN assay for screening purposes.
The flow cytometry/γ-H2AX assay
Phosphorylation of histone H2AX at serine 139 (γ-H2AX) occurs immediately after the formation of DNA DSBs in cells. When DSBs are formed, histone H2AX protein molecules, which extend over mega base-pair regions surrounding the break site, become phosphorylated within a few seconds (96). Phosphorylation is the first step in recruiting and localizing a variety of repair proteins to DNA damage sites (97). The γ-H2AX foci can be selectively detected via immunostaining, so that the number of γ-H2AX foci can be correlated to the number of DSBs present in the cell (98,99). The sensitivity and specificity of γ-H2AX immunostaining is high in comparison to other methods for measuring DSBs (98,100) and γ-H2AX immunostaining is considered one of the most reliable methodologies. DSBs are also indirectly formed by the collision of replication forks at sites for other types DNA damage (e.g. DNA adducts, DNA–DNA crosslinks, etc.,) and at DNA damage repair sites (101). The identification and measurement of γ-H2AX levels in cells is now considered a useful and important tool for characterizing the effects of potential genotoxins in biological and medical research.
Recently, some reports showed that ENMs increase γ-H2AX levels in in vitro experiments (102–105). Metal-oxide NPs in particular, such as CuO, ZnO and NiO, etc., induced the formation of γ-H2AX foci (102,104). Based on the nonspecific detection of increased intracellular ROS levels (via the 2′, 7′-dichlorodihydrofluorescein diacetate - DCFH-DA assay) following NP exposures (104,105), the authors surmised that the NP-induced ROS were contributory factors resulting in DNA damage and formation of γ-H2AX foci. Additional evidence to support this conclusion was obtained when the authors observed that the formation of DSBs and the number of γ-H2AX foci were attenuated by the inclusion of ROS scavengers, e.g. catalase and N-acetylcysteine into the system (104,105). ENM induced or generated ROS can also react with endogenous antioxidants, such as glutathione (GSH), resulting in an imbalance (oxidative stress) in the natural levels of antioxidants and ROS in cells (2,106). Metal ions released from metallic NPs can also react with –SH groups on antioxidant enzymes and further decrease anti-oxidant levels. Oxidative damage to DNA and its transformation to DSBs is a complex process. SSBs can form from direct ROS attack on DNA, from the process of repairing oxidatively induced DNA damage and/or from replication fork collapse at sites of oxidatively induced DNA damage. All of these processes could potentially produce DSBs, leading to the production of γ-H2AX.
It is advantageous to measure γ-H2AX levels for characterizing the potential genotoxicity of ENMs. One of the advantages is that the analytical detection of γ-H2AX is highly sensitive. As described previously, a few reports indicate that the γ-H2AX staining method is as sensitive as the comet assay and pulse-field gel electrophoresis analysis for the quantification of DSBs (98,100). It was recently reported that γ-H2AX generated by TiO2 NP could be detected more sensitively than DSBs detected by gel electrophoresis (107). Another advantage is that the number of γ-H2AX foci in immunostaining is directly correlated with the formation of DSBs (98,99). The simplicity of the γ-H2AX detection process is also worth mentioning. The γ-H2AX can be detected by simple immunostaining or western blotting due to the ready availability of well-characterised antibodies against γ-H2AX, and by commercial immunofluorescent stains and ELISA kits. It should be noted that γ-H2AX levels vary greatly depending on the cell line and the cell cycle. Thus, it is important to measure the background γ-H2AX levels before exposing cells to ENMs. After exposing the cells to ENMs, the background γ-H2AX levels should be subtracted from the measured γ-H2AX levels due to the ENM exposure; this correction is essential when comparing different cell lines. As the time points of γ-H2AX induction and disappearance are also different between different types of ENMs and cell lines, time course studies might also be required.
Principles of the flow cytometric γ-H2AX assay
Recently, a high-throughput version of γ-H2AX staining assay for the detection and measurement of DSBs was developed: flow cytometry γ-H2AX (FC γ-H2AX). This new assay format is especially useful for characterizing the DSB generation potential of ENMs. High-speed analysis is a fundamental characteristic of FC, e.g. tens of thousands of cells can be analysed in 1 s. Cells which are treated with ENMs are fixed and stained using either a primary γ-H2AX antibody plus a fluorescently labeled secondary antibody or a fluorescently-labeled primary γ-H2AX antibody. Preparation of a single cell suspension is required before fixation and statistically validated data on the γ-H2AX level (based on fluorescence intensity) can then be obtained quickly. One of the advantages of the FC γ-H2AX assay is that it is possible to obtain additional information regarding other cellular processes by using multiple fluorescent stains or fluorescent antibodies. For example, staining with propidium iodide (PI) can provide data regarding the cell cycle. This is important for characterizing the timing of the DNA damage response, e.g., ENM induced the formation of γ-H2AX (DSBs) during a specific phase of the cell cycle. PI intercalates into the major groove of dsDNA and produces a highly fluorescent adduct that can be excited at 488 nm with a broad emission centered around 610 nm. The fluorescence intensity reflects cellular DNA content; therefore, double-staining of γ-H2AX and DNA using fluorescein iso-thiocyanate (FITC; emission maximum 525 nm)-labeled antibody and PI, respectively, makes possible to show cell cycle-dependent generation of γ-H2AX (108). The enhancement of γ-H2AX after treatment with ENMs can be observed in all phases of the cell cycle, which demonstrates that DSB formation is independent of cell cycle phase and the level of cellular proliferation. On the other hand, γ-H2AX that are produced during S phase indicate that the DSBs are generated during DNA replication (due to replication fork collapse, etc.). Both ZnO NPs and CuO NPs were recently shown to produce γ-H2AX foci that were independent of the cell cycle phase, suggesting that DSB formation was due to ENM induced ROS attack on DNA and generation of excess SSBs (104). By gating the sub-G1 fraction of the cell cycle, it is possible to delete the apoptotic cells (DNA fragmented cells). This is a major advantage of the FCM γ-H2AX assay.
When applying the FC γ-H2AX assay system to ENM exposed cells, it is best to analyse the side-scattered (SS) light simultaneously with the fluorescence signal. In the FCM flow chamber, as cells flow in single file through the light beam (488 nm laser), the light is scattered in various directions. Light that is scattered in the forward direction (typically up to 20° offset from the laser beam’s axis) is called the forward-scattered light (FS). The FS intensity is correlated to the cell’s size. Scattered light measured approximately at a 90° angle to the excitation line is the SS light. The SS intensity provides information about the granular content within a cell. When metallic ENMs are taken-up into cells, SS light intensity consequently increases without a change in FS light intensity (104,109). Simultaneous detection of SS and γ-H2AX can provide information about the relationship between ENM incorporation and DSBs formation. It should be noted that γ-H2AX foci can also be generated when metal ions are released from metallic ENMs. ENMs composed of transition metals can lead to the release of transition metal ions, e.g. CuO NPs release Cu2+ ions. Transition metals as well as transition metal ions located in the cell nucleus can induce the formation of ·OH (due to Fenton chemistry), which subsequently attacks DNA to produce oxidatively modified DNA lesions, as well as SSBs (15,110). The DNA damage can transform into DSBs as described previously which induces the formation of γ-H2AX foci. Using only γ-H2AX detection, it is not possible to determine if DSBs are caused by metallic ENMs or their dissolved ions. Figure 2 shows hypothetical FCM images of dot plots indicating SS versus γ-H2AX responses after treatment with metallic ENMs. In particular, plot C illustrates the phenomenon where you might encounter low ENM uptake into cells, yet have high release of metallic ions which are able to induce DNA damage. Plot B represents the scenario where ENMs show high uptake but induce little DNA damage. It is critical to follow up with confirmatory genotoxicity assays when one observes a pattern indicating high genotoxicity as illustrated in plots A and C.
Figure 2.
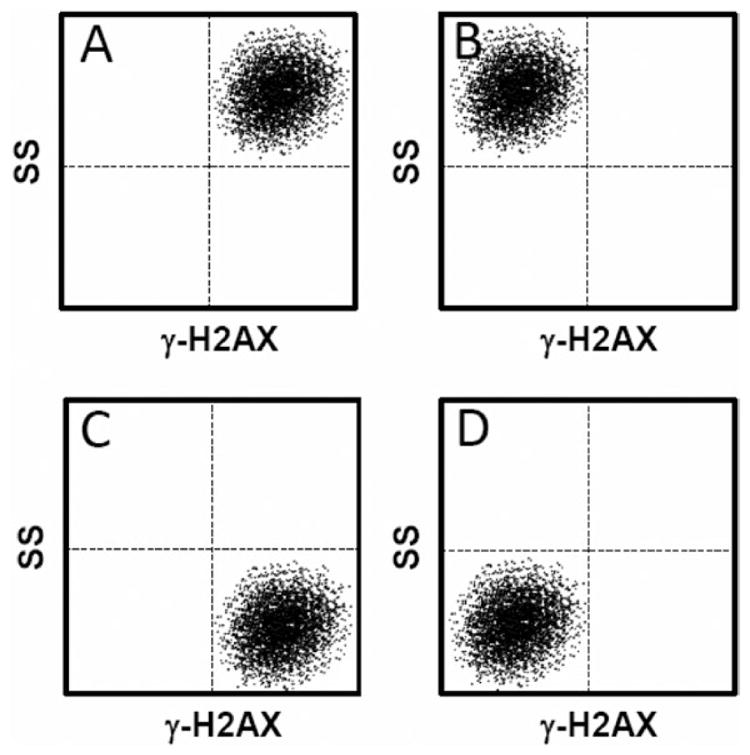
Hypothetical flow cytometry dot plot patterns for SS versus γ-H2AX responses after treatment with ENMs. The dot plot patterns are divided into four groups; (A) high uptake of ENMs and high genotoxicity (γ-H2AX), (B) high uptake and low genotoxicity, (C) low uptake and high genotoxicity, (D) low uptake and low genotoxicity.
Key advantages and limitations of the technology
The FC γ-H2AX assay combined with SS analysis is ideal for characterizing the genotoxicity (DSB formation) of ENMs under high-throughput conditions; however, there is a basic issue that needs attention before the assay can be applied in practice. The staining of γ-H2AX protein molecules with antibodies is necessary and this requires the cells to be fixed using formaldehyde or ethanol. One limitation of this sample preparation procedure is that ENMs inside the cells are sometimes arbitrarily released from the cells during the fixation step. It is now also possible to simultaneously measure SS light and ROS formation in living cells by incorporating the DCFH-DA reagent into the assay system (104). In order to enhance the capabilities of the FC γ-H2AX assay even further, the development of a specific γ-H2AX stain that is compatible with living cells is required. One possibility may involve the use of fluorescent fusion proteins. The green fluorescent protein (GFP)-fused mediator of DNA damage checkpoint 1 (MDC1) which directly binds to γ-H2AX in order to regulate cellular responses to DSBs, forms nuclear foci in the same location as γ-H2AX (111). Cells which are transfected with plasmids encoding fluorescent fusion proteins that are designed to increase their fluorescence in conjunction with increased γ-H2AX provide a potential path toward this goal. Alternatively, cells which are transfected with plasmids encoding fluorescent fusion proteins whose fluorescence intensity are enhanced by a change in intracellular distribution is another potential option. Either of these options, as well as others could lead to the FC γ-H2AX assay becoming a truly multimeasurement, high-throughput platform for characterizing the potential genotoxicity of ENMs in living cells.
The automated FADU assay
Quantification of DNA SSBs in living cells is an important endpoint in the assessment of genotoxicity. One of the available methods is the FADU assay. The original version (36) requires large numbers of cells and its manually operated steps make it technically very demanding. However, an automated version based on the use of a commercial laboratory robot with additional modifications of the assay protocol has been recently established (41,112). This automated version is more convenient and can be performed in a higher throughput. The main advantages are the following: agitation-free handling of the samples placed in 96-well microtiter plate to avoid shearing of the DNA; precise dispensing of solutions at a controlled rate without any mixing; accurate temperature control of the samples at all times; full protection from light at all times; reduction of the number of cells required by more than 100-fold; and finally automated performance of all the steps. The automated FADU assay essentially measures the number of DNA strand breaks in a cell population using high-throughput procedures. The assay can also measure DNA repair activity, the disappearance of DNA strand breaks, in living cells over controlled time periods. DNA repair can be evaluated in cells after the cells have been exposed to a potential DNA strand break inducer.
Principles of the automated FADU assay
The detection and measurement of DNA strand breaks (single and double) is based on progressive DNA unwinding (denaturation) under alkaline conditions and highly controlled pH, time and temperature. Denaturation begins at DNA ‘open sites’ like replication forks, chromosome ends, and at endogenously (metabolic ROS production) or exogenously (irradiation, chemical compounds) induced DNA strand breaks. Furthermore, DNA SSBs induced by DNA repair enzymes, such as formamidopyrimidine DNA glycosylase (Fpg), can be also detected in a cell-free system using plasmid DNA. In order to monitor DNA unwinding, a fluorescent dye (SybrGreen®), that binds to dsDNA is used. Low fluorescence intensity indicates high DNA unwinding and consequently a greater number of DNA strand breaks.
For suspension cells, treatment with DNA damaging agents can take place in 2 ml tubes. The number of tubes depends on the treatment points and necessary controls (Figure 3). After treatment, the tubes are transferred in a pre-cooled rack and placed in the robot workspace. The first automated step is to add ice-cold suspension buffer to the samples. The temperature of the samples decreases which mitigates (or stops) any further cellular reactions. The diluted cell suspensions are immediately transferred into pre-cooled 96-well plates (in replicate). For adherent cells, cells are incubated in a 96-well plate until they attach to the surface. After the treatment medium is removed, the plate is placed onto the pre-cooled plate-holder in the robot workspace. Next, lysis buffer is added to each well (containing suspension or adherent cells) and the plate is incubated for 12 min in order to break the cell membranes and expose DNA to the unwinding process. Lysis buffer also denatures and inactivates cell proteins, therefore the simultaneous addition of lysis buffer to all wells ensures the inhibition of any further cellular reactions in all of the samples at the same time. Next, ice-cold alkaline buffer is gently added onto the top of the lysates and incubated for 5 min, allowing the buffer to slowly diffuse into the cell lysate layer. Further incubation at 30°C begins the DNA unwinding process. After 60 min, the DNA denaturation process is stopped by addition of neutralisation buffer. Finally, SYBR Green dye is added into all wells and fluorometric detection is performed.
Figure 3.

Schematic representation of three different experimental layouts for analysis of DNA strand breaks using the automated FADU assay. (A) Cells attached to a 96-well plate are treated with two different compounds (C1 and C2) in six different concentrations (P1–P6). T and P0 are untreated controls; P0 represents the endogenous amount of DNA strand breaks and T the total DNA amount. In this example, three wells of T values are designated to assess the interference of compounds or solvents used with the fluorescence signal. For this purpose, cells are treated with the compound and/or solvents and compared to the untreated T values. This layout allows six replicates for conditions P0–P6 and three replicates for treated T and untreated T values, respectively. (B) Cells attached to a 96-well plate are treated with four different compounds (C1–C4) in one single concentration (P1). After damage occurs (P1t0), cells were incubated for different repair times (P1t1–Pt5). T and P0 are untreated controls; P0 represents the endogenous amount of DNA strand breaks and T the total DNA amount. This layout allows three replicates for each condition. (C) Lymphocytes from four different subjects (S1–S4) are irradiated in 2 ml tubes with doses ranging from 1 to 6 Gy. Cooled tube rack is placed in the liquid handling device (LHD) workspace and (D) samples are transferred to a pre-cooled 96-well plate. T and P0 represent non-radiated cells. This layout allows three replicates for each condition.
In order to determine the amount of intact dsDNA (DNA that has not been unwound) remaining in a cell lysate, neutralisation buffer is added before the alkaline unwinding solution. This control value is called the ‘T-value’ and set as 100% of DNA amount. Endogenous DNA damage can be assessed in samples, which are not exposed to external DNA damaging agents, resulting in the so called ‘P0 value’ (Figure 4) (113). All steps, beginning from the addition of suspension buffer until SybrGreen staining, are fully automated using a modified commercially available pipetting robot. Detailed information about the automated FADU assay for detecting DNA strand breaks has been published previously (112,114).
Figure 4.

Principle of the FADU assay (schematic representation). Left boxes represent cells without (T and P0) or with (Px and B) DNA damage. In the middle boxes the dsDNA with increasing levels of damage and increasing extent of unwinding is represented. Right boxes contain small circles representing the fluorescent dye SybrGreen®.T, P0 und B are controls. T values the total DNA amount (100 % fluorescence), P0 values represent endogenous DNA strand breaks and B values represent the background fluorescence. Px values (P1, P2, P3, P4 Px) are the different extents of damage to be measured. After treatment cells are lysed and DNA is exposed to alkaline unwinding (NaOH). Neutralisation buffer stops the unwinding and SybrGreen® stains the dsDNA. Reprinted with permission from Steinberg, P. (ed.), High-Throughput Methods inToxicityTesting. John Wiley and Sons, Hoboken, pp. 285–294).
Modifications of the automated FADU assay
Modifications to the original FADU assay have been incorporated in order to allow efficient measurement of DNA crosslinks (DNA–DNA crosslinks) in human cells (115), as well as oxidatively induced DNA base damage in plasmid and mitochondrial DNA (116,117).
Detection of crosslinks
The automated FADU assay can be applied to the detection of inter-strand crosslinks (ICL) based on the prevention of time-dependent alkaline unwinding of dsDNA. Interstrand crosslinks consist of a covalent bond between complementary DNA strands, which cannot be unwound under alkaline conditions. Interstrand crosslinks can be measured indirectly by inducing large numbers of DNA strand breaks, which causes high alkali unwinding. In the presence of crosslinks, the unwinding level decreases. Human peripheral blood mononuclear cells (PBMC) exposed to the model compounds mitomycin C, melphalan and sulphur mustard were used for detecting ICLs (115).
Detection of 8-oxodG
Free radical induced oxidation of 2′-deoxyguanosine (dG) to 8-oxo-2′-deoxyguanosine (8-oxodG) is a major type of oxidatively induced DNA base damage. The detection and measurement of 8-oxodG levels in genomic DNA is commonly pursued in the DNA damage and repair research community and 8-oxodG is considered an established biomarker for oxidative stress (110). Importantly, 8-oxodG can be converted to mutations in proliferating cells, therefore it may play an important role in carcinogenesis (118). High-performance liquid chromatography (HPLC) with electrochemical (EC) detection, gas chromatography-mass spectrometry (GC-MS) and LC tandem mass spectrometry (LC-MS/MS) have been used for the detection of 8-oxodG levels (119–121). The automated FADU assay has been further developed and modified for detecting oxidatively induced DNA lesions and for testing for the effects of antioxidants on lesion formation using plasmid DNA; results from this study demonstrated good correlation with 8-oxodG levels as measured by LC–MS/MS (117). Due to a decreased operation time and high throughput of an automated procedure, the FADU-Fpg method offers a potentially useful tool for screening compounds that are capable of inducing oxidatively induced DNA damage, as well as a tool for screening compounds with antioxidant properties.
Mathematical modeling of radiation-induced DNA strand breaks using the FADU assay
As mentioned previously, the FADU assay uses a fluorescence dye that determines the remaining dsDNA. The intensity of the fluorescence signal is inversely related to the number of DNA strand breaks: with increasing DNA strand breaks the fluorescence signal decreases. As reported in the literature, the number of strand breaks increases linearly with the dose of radiation applied (48,61). However, there is a non-linear relationship between fluorescence intensity and the radiation dose, especially at high radiation doses where the fluorescence intensity reaches a plateau. In order to quantify DNA strand breaks using the FADU assay, the percentage of dsDNA remaining after the alkaline unwinding or the decadic logarithm of the intensity has been used (41,112). However, in order to capture the non-linear relationship more precisely a mathematical model has been established (122).
Applications of the FADU assay
The automated FADU assay provides an accurate method for screening of genotoxic agents, including chemicals and ENMs. Numerous in vitro and in vivo studies have demonstrated that ENMs have the potential to induce DNA damage (19–21). For instance, zinc oxide nanoparticles (ZnO NPs) have been shown to exhibit genotoxic, clastogenic and cytotoxic effects (123). Zinc oxide nanoparticles are highly important due to their current and potential applications in biosensors, sunscreens, cosmetics, medicine and nutrition amongst others (124). The automated FADU assay has recently been shown to be a suitable method for detecting and measuring DNA strand breaks in cells exposed to ZnO NPs (41). In this preliminary study, A549 cells were exposed to increasing concentrations (0.4–160 μg/ml) of ZnO NPs up to 3 h and then the cells were analysed for levels of NP induced DNA damage. The key finding from this study was that the ZnO NPs induced significant levels of strand breaks only at the highest NP dose (3 h exposure period). Distinct formation of strand breaks from the assay positive controls demonstrated that the assay was performing as expected. The study demonstrates the potential of applying the high throughput capabilities of the FADU assay toward the future screening of large numbers of ENMs. There is an established concern that the use of animal models for genotoxicity testing is inappropriate because animal models do not mirror toxicity in humans very well. As a consequence, mammalian cell-based alternatives to typical animal-based clastogenicity assays are needed for early screening of mammalian genotoxicity (65). The automated FADU assay is a cell-based in vitro system, which is well suited for the measurement of DNA strand breaks in different types of human cell lines (125,126), as well as in primary human cells such as PBMCs, lymphocytes (126–129) and/or fibroblast (130) obtained from different subjects. The automated FADU offers an alternative method for reducing animal use during genotoxicity risk assessments. Furthermore the automated FADU assay has been applied successfully in human biomonitoring for such cases as: the assessment of DNA damage in Alzheimer’s patients (131), associations of subjective vitality with DNA damage, cardiovascular disease risk factors and physical performance (132), characterizing the influence of age, gender (133) and genetic and environmental factors (134) on DNA strand break generation and investigating the effects of psychotherapy on DNA strand breaks levels in post-traumatic stress disorders (135). Such strategies related directly to humans are needed rather than attempting to extrapolate results across species.
Key advantages and limitations of the technology
One of the main advantages of the recently automated FADU assay for ENM (and chemical) genotoxicity studies is its 96 well plate, high throughput sample preparation and sample processing capabilities. ENM sample throughput is further enhanced through the use of a dedicated robotic liquid handling device (LHD). The use of the LHD also reduces user bias because all of the pipetting is performed automatically. The 96-well plate format of the assay also allows one to efficiently perform DNA repair kinetic studies. As far as sensitive strand breaks detection is concerned, the liquid-phase fluorescent readout is fast and unbiased and the overall analytical limit of detection is 0.13 Gy based on the detection of ionizing radiation induced strand breaks (112), which compares well with the reported limit of detection (0.03 Gy) for ionizing radiation induced SSBs measured by the comet assay (136). Since DNA strand breaks are calculated based on the read out of fluorescence signal, this might become a problem for substances or reagents that quench or enhance fluorescence. However, this problem can be overcome by including additional controls. The effects of those factors on the fluorescence signal can be taken into consideration and values can be recalculated and corrected for artefactual fluorescence quenching or enhancement. One important limitation of the FADU assay for ENM genotoxicity applications is that it is not currently possible to archive ENM exposed samples after they have been processed with the FADU assay. Because the fluorescence detection readout is solution based, the samples cannot be stored (dried, refrigerated or frozen) for re-analysis at future time points due to quenching and/or degradation of the fluorescent signal. Furthermore, in the FADU assay, the unwinding process takes place starting from both SSBs and DSBs. Therefore, in order to determine the type of DNA strand breaks induced by a particular ENM or other strand break inducer, additional more selective assays will need to be performed. A good correlation in the detection of 8-oxodG levels measured by LC–MS/MS and FADU assay has been demonstrated in plasmid DNA (117). However, cell based detection of 8-oxodG using the FADU assay has challenges and has not been established yet. Therefore, another limitation of the assay is that potential cell based studies on DNA repair enzymes can not be initiated due to repair enzyme incompatibilities with critical components in the assay lysis buffer, i.e. urea. This limitation makes difficult to compare FADU assay results against comet assay results. For instance, the European Standards Committee on Oxidative DNA Damage (ESCODD) reported discrepancies between HPLC techniques and the enzymatic approach using the Fpg-comet assay (137). In this study, the DNA isolation procedure necessary for HPLC analysis might have induced additional DNA damage influencing the final measurement and could be responsible for the discrepancies between the HPLC and comet assay measurements.
However, in the future there may be ways to further modify the FADU assay to overcome the current technical limitations. In summary, the automated FADU assay greatly facilitates analyses of DNA damage in a variety of settings and provides a reliable, convenient and non-biased method for measuring endogenous DNA damage in both human biomonitoring and ENM genotoxicity studies. The FADU assay also finds application in the fields of (i) toxicology and pharmacology by assessing genotoxicity of chemicals and ENM; (ii) in occupational and environmental science by biomonitoring human populations for exposure to genotoxic agents; (iii) in nutritional research by investigating the protective effect of antioxidants and in (iv) medical science by monitoring DNA repair capacity in various diseases, particularly in cancer research.
TheToxTracker reporter assay
An attractive approach to enable high-throughput analyses of a range of ENMs is to use reporter cell lines that are constructed to fluoresce upon activation of certain signaling pathways. For genotoxicity, one of the most extensively validated reporter gene tests is the GreenScreen HC assay. This fluorescent reporter assay uses the GADD45α (growth arrest and DNA damage inducible alpha) gene promoter fused to a GFP marker for the detection of genotoxic agents in p53-competent human-derived TK6 cells (138). When testing the the European Centre for the Validation of Alternative Methods (ECVAM) recommended lists of genotoxic and non-genotoxic chemicals for assessment of new genotoxicity tests, the GreenScreen HC assays showed good sensitivity and specificity (139). More recently, the BlueScreen HC, a luciferase-based version of the GADD45α reporter assay, was described (140), and this assay has also been developed into a 384-well version (141). GADD45α is a member of the growth arrest and DNA damage (GADD) proteins that are induced upon various cellular stresses, including nutrient deprivation, oxidative stress and DNA damage (142).
To obtain more specific insight into different mechanisms of (geno)toxicity, a combination of various reporter cell lines will be required. In order to meet this need, the ToxTracker assay was developed by Hendriks and co-workers (43,143). The assay consists of a panel of six mouse embryonic stem (mES) cell lines that each contains a different GFP-tagged reporter for a distinct cellular signaling pathway. These reporters were constructed by first performing whole-genome transcription analysis of mES cells exposed to different classes of chemicals in order to identify specific biomarker genes. Next, GFP reporters were generated by bacterial artificial chromosome (BAC) recombineering. The assay was adapted to a 96-well plate format, thus enabling medium/high-throughput screening. The assay procedure is very straightforward; the reporter cells are exposed to the ENMs in the 96-well plates, typically for 24 h, and the fluorescence in live cells is examined by flow cytometry. Assessment of reporter activation by flow cytometry has the advantage that it allows detection of GFP reporter induction in individual viable cells.
The original ToxTracker assay consisted of two reporter cell lines. The DNA damage reporter Bscl2 (encodes a protein called Seipin) depends on the ATR (ataxia telangiectasia mutated and Rad3-related)-associated DNA damage signaling pathway, and is selectively activated after exposure to genotoxic agents and the subsequent interference with DNA replication. Furthermore, the Srxn1 (encodes the sulfiredoxin-1 protein) reporter is preferentially induced upon oxidative stress and is part of the Nrf2 (Nuclear Factor, Erythroid Derived 2, Like 2) antioxidant response pathway. By using these two reporters, it is thus possible to differentiate between compounds that are directly DNA-damaging and those that mainly act via oxidative stress.
Recently, the ToxTracker assay was expanded with four additional reporters. The Rtkn reporter is a second DNA damage reporter that is associated with NFκB signaling (143). As a second reporter for oxidative stress, Blvrb (encodes an oxidoreductase) was constructed as a reporter gene being independent of Nrf2 signaling. In addition to DNA damage and induction of ROS production, protein unfolding has been associated with carcinogenicity. Accumulation of misfolded proteins in the cytosol is recognised by heat shock proteins (HSP). As a reporter for protein unfolding, the Ddit3 reporter was constructed. Finally, the Btg2 reporter gene is controlled by p53 and is activated by various types of cellular stress. Thus, the ToxTracker assay now consist of six different reporter cell lines that detect DNA damage, oxidative stress, protein unfolding and general cellular stress, see Table 1 and Figure 5. In a recent study, an extensive validation using the compound library suggested by ECVAM was performed and the result showed that the ToxTracker assay had an outstanding sensitivity and specificity for the detection of human genotoxins (143).
Table 1.
The biological damage and cellular pathways that can be evaluated using the different reporter cell lines
| Biological damage | Cellular pathway | Biomarker gene |
|---|---|---|
| DNA damage | ATR/Chk1 DNA damage signalling | Bscl2 |
| NF-κB signalling | Rtkn | |
| Oxidative stress | Nrf2 antioxidant response | Srx1 |
| Nrf2-independent | Blvrb | |
| Protein damage | Unfolded protein response | Ddit3 |
| Cellular stress | P53 signalling | Btg2 |
Figure 5.

The ToxTracker assay applies a panel of six independent GFP-based reporter cell lines for detection of DNA damage, oxidative stress and protein unfolding. ToxTracker identifies activation of the ATR and Nf-KB-associated DNA damage responses by the Bscl2-GFP and Rtkn-GFP reporters, respectively. Induction of oxidative stress is established by the Srxn1-GFP and Blvrb-GFP reporters. The Ddit3-GFP reporter indicates activation of the unfolded protein response and the Btg2-GFP reporter detects activation of the p53 cellular stress response. Induction of the GFP reporters, as well as cytotoxicity, is determined by flow cytometry. Data analysis and visualisation of the results are performed by the Toxplot software platform.
Application of the technique to nanomaterial genotoxicity investigations
In published and unpublished studies, Karlsson and Hendriks have used the ToxTracker assay for testing 24 different ENMs. In the published study (102), the ability of the ToxTracker assay to identify the hazardous properties and underlying mechanisms of a panel of metal oxide- and AgNPs, as well as a selection of non-metallic materials (diesel, carbon nanotubes and quartz) using three of the reporter cell lines were evaluated. The reporter cells were able to take up NPs, and furthermore, exposure to CuO, NiO and ZnO NPs as well as to quartz (used as a benchmark particle) resulted in activation of the oxidative stress reporter (Srxn1), although only at high cytotoxicity for ZnO. Nickel oxide NPs also activated the p53-associated cellular stress response (Btg2), indicating additional reactive properties. Conventional assays for genotoxicity assessment (comet assay, γ-H2AX staining) confirmed the response observed in the ToxTracker assay. A comparison was also made between the NPs and the metals in ionic form, and it was concluded that for CuO NPs, the released Cu ions species contributed to the effects, whereas for NiO, the reporter response was related to the particles per se. The DNA replication stress-induced reporter (Bscl2), which is most strongly associated with carcinogenicity, was not activated by any of the tested NPs. The AgNPs included in the study showed no effect in the reporter cell lines. This was in contrast to a previous study on the same AgNPs using BEAS-2B cells in which an increase in DNA damage (comet assay) was observed with clear effects on viability for the smallest (10 nm) AgNPs (144).
In unpublished studies, Karlsson and colleagues have extended the toxicity screening of ENMs by investigating quantum dots (CdTe) of various sizes and found clear size-dependent effects in terms of oxidative stress reporter activation and cell viability. Of all ENMs tested so far, the Nrf2 dependent oxidative stress reporter (Srxn1) seems to be the one that is most sensitive following exposure to metal- and metal oxide ENMs. Furthermore, the ENMs with the lowest LC50 values were the smallest QDs (1.5 nm), as well as the metallic nickel ENMs, with LC50 values of approximately 1.5 μg/ml. As of now, none of the tested ENMs clearly induced the Bcl2 reporter indicating that, most likely, none of the tested ENMs could directly bind to DNA and cause stalled replication forks. Interestingly, cobalt (Co) NPs induced the Rtkn reporter suggesting DNA damage and NFκB signaling. Cobalt dust has been reported to cause lung cancer in 2-y inhalation studies in rats and mice (145).
Key advantages and limitations of the technology
There are several advantages with using the ToxTracker assay in ENM genotoxicity studies. The mES cells that are used in the ToxTracker assay are untransformed, proficient in all major DNA damage and cellular stress response pathways and have been shown to efficiently engulf ENMs (102). The assay has shown good sensitivity and specificity for detecting genotoxic compounds when evaluated using a wide range of genotoxic and non-genotoxic chemicals (143). The ENMs tested so far have shown various effects, some being highly toxic and efficiently inducing one or several of the reporters and others showing no effects at all. In all, this indicates that the ToxTracker reporter cell assay can be applied as a rapid mechanism-based tool for assessing the potential genotoxic effects of ENMs. The assay is adapted to a 96-well plate format thus enabling medium/high-throughput screening. As for all assays relying on fluorescence, possible assay-interactions may be found for fluorescent ENMs, but this can easily be tested for by using the mES cells without the reporter gene. Another possible limitation could be dark ENMs that may scatter the fluorescence. For most ENMs, however, an interaction that substantially may influence the results appears unlikely at this point. Since the method is rapid and no processing of the cells (DNA extraction, etc.) is needed, it seems plausible that no assay-interactions occur for most ENMs. In conclusion, the ToxTracker reporter assay seems to be very promising for rapidly assessing the genotoxicity potential of ENMs.
Future perspectives
As noted, the emerging uses of ENMs in biological and environmental applications, such as medicine and agriculture, necessitate the comprehensive characterisation of ENM interactions with specific molecular targets within these systems. While this remains a very complex objective, our understanding of the interactions between ENMs and biological and environmental systems has dramatically improved over recent years (146). Innovative physicochemical characterisation methods, development and optimisation of toxicity assays, and improved analytical tools have enabled these advances towards the overall goal of safely incorporating nanotechnology into our society.
However, there are many areas in which high-throughput nanotoxicity assays can be improved, particularly with respect to metrology. Currently, there has been no consensus regarding assays and protocols that should be used to analyse the in vitro toxicity of ENMs. Likewise, no clear standard and/or regulatory guidelines on testing and evaluation are available at present (147). High-throughput assays, while promising, should be validated against more conventional assays before their use in a regulatory setting. Of great concern, however, is the difficulty in validating the many different assays for the many different possible ENMs (148). A major concern here is the low throughput of ENM characterisation techniques, which reduces the overall efficiency of high-throughput screening assays for ENMs. The further development of hybrid instruments (149) or in-line techniques (150) would not only lead to more rapid ENM characterisation, but conceptually, could facilitate new instruments having the ability to simultaneously characterise and assess in a high-throughput fashion. For example, flow cytometry has been used to measure cell viability while simultaneously measuring whether ENMs interacted with the cell membrane or entered the cells (151,152); combining this technique with in-line ENM characterisation tools could enhance our ability for prompt assessments.
As previously noted, comprehensive characterisation of ENMs is critical, as the physicochemical properties of ENMs vary from material to material and with their environment (dry powder, water suspension, biological media, etc.); factors that can artificially influence assay results and hinder their validation (10). In addition, each assay operates on a different basis, which can make it difficult to cross-correlate outcomes between different assays, an important step in the validation process (147). To help facilitate these goals, standardised protocols (principally with respect to ENM handling) are needed, along with improved control and reference materials. An exemplary goal would be the development of a DNA or cell-based reference material with quantified amounts and types of damage (e.g. single/DSBs, etc.) for use as a positive control in toxicity assays. However, such materials would be quite challenging to produce, given the high unpredictability of biological entities. Perhaps more readily available would be ENM reference materials (with well-defined and consistent physicochemical properties) that have been validated for use as positive or negative controls in multiple toxicity assays; such materials are already being pursued by the research community (153). Validation of such materials is not trivial and would require rigorous comparative studies using robust assays that have been examined for ENM interference (10,11). Improved nanotoxicity assays will greatly benefit all realms of nanotechnology, one in particular being precision nanomedicine.
The primary conceptual goal of precision medicine can be briefly summarised as identifying and delivering a specific treatment to a patient based on their specific physiology, with minimal adverse effects (154). Due to their sizes on the scale of biological entities (DNA, proteins, etc.) and the vast potential for physicochemical property customisation, ENMs show much promise for precision medicine applications, including site-specific (i.e. cancer cell) cytotoxicity (155), localised drug-delivery vehicles (156), bioimaging agents (157) and diagnostic testing assays (158). While some ENM-based systems are already undergoing evaluation in clinical trials (159), there is still a need for developing more accurate, reproducible and high throughput methods for evaluating ENMs for precision medicine applications. This is especially true within the realm of precision oncology, where the high profile nature of the disease, the need for developing more effective and less damaging treatments, and the large degree of physiological variability among different types cancers (160) creates a scenario where the ability to quickly assess the biophysicochemical interactions of ENMs could have a significant impact on the field.
Towards the goal of understanding biophysicochemical interactions of ENMs, high-throughput nanotoxicity assays, as described previously, are typically used to evaluate the potential cytotoxic and genotoxic responses of ENM exposures. In this traditional sense, ENMs are being subjected to hazard assessment, to measure their levels of toxicity to biological and environmental entities. However, the underlying goal of cancer treatment is the elimination of cancer cells, one mechanism of which is the induction of cytotoxicity (potentially via genotoxicity). With this in mind, high-throughput genotoxicity assays could be adapted for use as both toxicity reporter measurement tools and as toxicity efficacy measurement tools. This concept is not foreign in the area of drug design, where high-throughput assays have been used as evaluative anticancer activity screens of molecular drugs for several years (161). Similar application of this ideology to ENMs could rapidly advance the use of ENMs for oncology applications, a simple change in perspective that could revolutionise cancer treatment. For example, the high-throughput imaging method described by Harris et al. (162) could be adapted for use with cancer cells. Likewise, the impedance-based spectroscopic method developed by Paget and Sergent et al. (151) does not require the use of dyes or markers and could also be used to identify critical time-points through its real-time monitoring of potential ENM toxic effects if used with cancer cells. By modifying assays in this fashion, hundreds, if not thousands of potential therapeutic ENMs could be rapidly assayed for their ability to induce toxic outcomes specifically in cancer cells and not in normal cells.
Acknowledgments
One of the authors, CMS, acknowledges funding and support from the National Academy of Sciences - National Research Council Postdoctoral Research Associateship Program and HLK acknowledges funding from the Swedish Research Council (VR, project 621-2014-4598.
Footnotes
NIST Disclaimer
Certain commercial equipment, instruments and materials are identified in this article to specify an experimental procedure as completely as possible. In no case does the identification of particular equipment or materials imply a recommendation or endorsement by the National Institute of Standards and Technology nor does it imply that the materials, instruments, or equipment are necessarily the best available for the purpose.
Conflict of interest statement: G.H. is the CEO of Toxys, a Dutch biotech company that offers the ToxTracker assay as a commercial service to industry.
References
- 1.Lux report. [accessed April 2016];Nanomaterials state of the market: Nanotechnology update: Corporations up their spending as revenues for nano-enabled products increase excerpt. https://portal.Luxresearchinc.Com/research/report_excerpt/16215.
- 2.Nel A, Xia T, Mädler L, Li N. Toxic potential of materials at the nanolevel. Science. 2006;311:622–627. doi: 10.1126/science.1114397. [DOI] [PubMed] [Google Scholar]
- 3.Nel A, Xia T, Meng H, Wang X, Lin S, Ji Z, Zhang H. Nanomaterial toxicity testing in the 21st century: use of a predictive toxicological approach and high-throughput screening. Acc Chem Res. 2013;46:607–621. doi: 10.1021/ar300022h. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Damoiseaux R, George S, Li M, et al. No time to lose–high throughput screening to assess nanomaterial safety. Nanoscale. 2011;3:1345–1360. doi: 10.1039/c0nr00618a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Cho WS, Duffin R, Bradley M, Megson IL, MacNee W, Lee JK, Jeong J, Donaldson K. Predictive value of in vitro assays depends on the mechanism of toxicity of metal oxide nanoparticles. Part Fibre Toxicol. 2013;10:55. doi: 10.1186/1743-8977-10-55. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Service RF. Nanotechnology. Can high-speed tests sort out which nanomaterials are safe? Science. 2008;321:1036–1037. doi: 10.1126/science.321.5892.1036. [DOI] [PubMed] [Google Scholar]
- 7.Stone V, Johnston H, Schins RP. Development of in vitro systems for nanotoxicology: methodological considerations. Crit Rev Toxicol. 2009;39:613–626. doi: 10.1080/10408440903120975. [DOI] [PubMed] [Google Scholar]
- 8.Warheit DB, Donner EM. Rationale of genotoxicity testing of nanomaterials: regulatory requirements and appropriateness of available OECD test guidelines. Nanotoxicology. 2010;4:409–413. doi: 10.3109/17435390.2010.485704. [DOI] [PubMed] [Google Scholar]
- 9.Doak SH, Griffiths SM, Manshian B, Singh N, Williams PM, Brown AP, Jenkins GJ. Confounding experimental considerations in nanogenotoxicology. Mutagenesis. 2009;24:285–293. doi: 10.1093/mutage/gep010. [DOI] [PubMed] [Google Scholar]
- 10.Petersen EJ, Henry TB, Zhao J, MacCuspie RI, Kirschling TL, Dobrovolskaia MA, Hackley V, Xing B, White JC. Identification and avoidance of potential artifacts and misinterpretations in nanomaterial ecotoxicity measurements. Environ Sci Technol. 2014;48:4226–4246. doi: 10.1021/es4052999. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Rösslein M, Elliott JT, Salit M, Petersen EJ, Hirsch C, Krug HF, Wick P. Use of cause-and-effect analysis to design a high-quality nanocytotoxicology assay. Chem Res Toxicol. 2015;28:21–30. doi: 10.1021/tx500327y. [DOI] [PubMed] [Google Scholar]
- 12.Nabiev I, Mitchell S, Davies A, et al. Nonfunctionalized nanocrystals can exploit a cell’s active transport machinery delivering them to specific nuclear and cytoplasmic compartments. Nano Lett. 2007;7:3452–3461. doi: 10.1021/nl0719832. [DOI] [PubMed] [Google Scholar]
- 13.Kodiha M, Wang YM, Hutter E, Maysinger D, Stochaj U. Off to the organelles - killing cancer cells with targeted gold nanoparticles. Theranostics. 2015;5:357–370. doi: 10.7150/thno.10657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Panté N, Kann M. Nuclear pore complex is able to transport macromolecules with diameters of about 39 nm. Mol Biol Cell. 2002;13:425–434. doi: 10.1091/mbc.01-06-0308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Atha DH, Wang H, Petersen EJ, Cleveland D, Holbrook RD, Jaruga P, Dizdaroglu M, Xing B, Nelson BC. Copper oxide nanoparticle mediated DNA damage in terrestrial plant models. Environ Sci Technol. 2012;46:1819–1827. doi: 10.1021/es202660k. [DOI] [PubMed] [Google Scholar]
- 16.Singh N, Jenkins GJ, Nelson BC, Marquis BJ, Maffeis TG, Brown AP, Williams PM, Wright CJ, Doak SH. The role of iron redox state in the genotoxicity of ultrafine superparamagnetic iron oxide nanoparticles. Biomaterials. 2012;33:163–170. doi: 10.1016/j.biomaterials.2011.09.087. [DOI] [PubMed] [Google Scholar]
- 17.Xia T, Kovochich M, Liong M, Mädler L, Gilbert B, Shi H, Yeh JI, Zink JI, Nel AE. Comparison of the mechanism of toxicity of zinc oxide and cerium oxide nanoparticles based on dissolution and oxidative stress properties. ACS Nano. 2008;2:2121–2134. doi: 10.1021/nn800511k. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Gonzalez L, Lison D, Kirsch-Volders M. Genotoxicity of engineered nanomaterials: A critical review. Nanotoxicology. 2008;2:252–273. [Google Scholar]
- 19.Magdolenova Z, Collins A, Kumar A, Dhawan A, Stone V, Dusinska M. Mechanisms of genotoxicity. A review of in vitro and in vivo studies with engineered nanoparticles. Nanotoxicology. 2014;8:233–278. doi: 10.3109/17435390.2013.773464. [DOI] [PubMed] [Google Scholar]
- 20.Petersen EJ, Nelson BC. Mechanisms and measurements of nanomaterial-induced oxidative damage to DNA. Anal Bioanal Chem. 2010;398:613–650. doi: 10.1007/s00216-010-3881-7. [DOI] [PubMed] [Google Scholar]
- 21.Singh N, Manshian B, Jenkins GJ, Griffiths SM, Williams PM, Maffeis TG, Wright CJ, Doak SH. NanoGenotoxicology: the DNA damaging potential of engineered nanomaterials. Biomaterials. 2009;30:3891–3914. doi: 10.1016/j.biomaterials.2009.04.009. [DOI] [PubMed] [Google Scholar]
- 22.Golbamaki N, Rasulev B, Cassano A, Marchese Robinson RL, Benfenati E, Leszczynski J, Cronin MT. Genotoxicity of metal oxide nanomaterials: review of recent data and discussion of possible mechanisms. Nanoscale. 2015;7:2154–2198. doi: 10.1039/c4nr06670g. [DOI] [PubMed] [Google Scholar]
- 23.Singh N, Asadi R, Griffiths SM, Jenkins GJS, Williams PM, Wright CJ, Doak SH. Understanding the in vitro genotoxicity induced by ultrafine superparamagnetic iron oxide nanoparticles (uspion) Mutagenesis. 2010;25:641–642. [Google Scholar]
- 24.Huk A, Collins AR, El Yamani N, Porredon C, Azqueta A, de Lapuente J, Dusinska M. Critical factors to be considered when testing nanomaterials for genotoxicity with the comet assay. Mutagenesis. 2015;30:85–88. doi: 10.1093/mutage/geu077. [DOI] [PubMed] [Google Scholar]
- 25.Doak SH, Manshian B, Jenkins GJS, Singh N. In vitro genotoxicity testing strategy for nanomaterials and the adaptation of current oecd guidelines. Mutat Res-Gen Tox En. 2012;745:104–111. doi: 10.1016/j.mrgentox.2011.09.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Gonzalez L, Sanderson BJ, Kirsch-Volders M. Adaptations of the in vitro MN assay for the genotoxicity assessment of nanomaterials. Mutagenesis. 2011;26:185–191. doi: 10.1093/mutage/geq088. [DOI] [PubMed] [Google Scholar]
- 27.Karlsson HL. The comet assay in nanotoxicology research. Anal Bioanal Chem. 2010;398:651–666. doi: 10.1007/s00216-010-3977-0. [DOI] [PubMed] [Google Scholar]
- 28.Karlsson HL, Di Bucchianico S, Collins AR, Dusinska M. Can the comet assay be used reliably to detect nanoparticle-induced genotoxicity? Environ Mol Mutagen. 2015;56:82–96. doi: 10.1002/em.21933. [DOI] [PubMed] [Google Scholar]
- 29.Magdolenova Z, Bilanicova D, Pojana G, Fjellsbo LM, Hudecova A, Hasplova K, Marcomini A, Dusinska M. Impact of agglomeration and different dispersions of titanium dioxide nanoparticles on the human related in vitro cytotoxicity and genotoxicity. J Environ Monitor. 2012;12:455–464. doi: 10.1039/c2em10746e. [DOI] [PubMed] [Google Scholar]
- 30.Møller P, Hemmingsen JG, Jensen DM, et al. Applications of the comet assay in particle toxicology: air pollution and engineered nanomaterials exposure. Mutagenesis. 2015;30:67–83. doi: 10.1093/mutage/geu035. [DOI] [PubMed] [Google Scholar]
- 31.Takahashi A, Ohnishi T. Does gammaH2AX foci formation depend on the presence of DNA double strand breaks? Cancer Lett. 2005;229:171–179. doi: 10.1016/j.canlet.2005.07.016. [DOI] [PubMed] [Google Scholar]
- 32.Geric M, Gajski G, Garaj-Vrhovac V. Gamma-h2ax as a biomarker for DNA double-strand breaks in ecotoxicology. Ecotox Environ Safe. 2014;105:13–21. doi: 10.1016/j.ecoenv.2014.03.035. [DOI] [PubMed] [Google Scholar]
- 33.Mah LJ, El-Osta A, Karagiannis TC. gammaH2AX: a sensitive molecular marker of DNA damage and repair. Leukemia. 2010;24:679–686. doi: 10.1038/leu.2010.6. [DOI] [PubMed] [Google Scholar]
- 34.Baumstark-Khan C, Hentschel U, Nikandrova Y, Krug J, Horneck G. Fluorometric analysis of DNA unwinding (FADU) as a method for detecting repair-induced DNA strand breaks in UV-irradiated mammalian cells. Photochem Photobiol. 2000;72:477–484. doi: 10.1562/0031-8655(2000)072<0477:faoduf>2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 35.Bhattacharya K, Dopp E, Kakkar P, Jaffery FN, Schiffmann D, Jaurand MC, Rahman I, Rahman Q. Biomarkers in risk assessment of asbestos exposure. Mutat Res Fund Mol M. 2005;579:6–21. doi: 10.1016/j.mrfmmm.2005.02.022. [DOI] [PubMed] [Google Scholar]
- 36.Birnboim HC, Jevcak JJ. Fluorometric method for rapid detection of DNA strand breaks in human white blood cells produced by low doses of radiation. Cancer Res. 1981;41:1889–1892. [PubMed] [Google Scholar]
- 37.Dusinská M, Slamenová D. Application of alkaline unwinding assay for detection of mutagen-induced DNA strand breaks. Cell Biol Toxicol. 1992;8:207–216. doi: 10.1007/BF00156731. [DOI] [PubMed] [Google Scholar]
- 38.Ahamed M, Karns M, Goodson M, Rowe J, Hussain SM, Schlager JJ, Hong Y. DNA damage response to different surface chemistry of silver nanoparticles in mammalian cells. Toxicol Appl Pharmacol. 2008;233:404–410. doi: 10.1016/j.taap.2008.09.015. [DOI] [PubMed] [Google Scholar]
- 39.Jugan ML, Barillet S, Simon-Deckers A, Herlin-Boime N, Sauvaigo S, Douki T, Carriere M. Titanium dioxide nanoparticles exhibit genotoxicity and impair DNA repair activity in A549 cells. Nanotoxicology. 2012;6:501–513. doi: 10.3109/17435390.2011.587903. [DOI] [PubMed] [Google Scholar]
- 40.Lim HK, Asharani PV, Hande MP. Enhanced genotoxicity of silver nanoparticles in DNA repair deficient Mammalian cells. Front Genet. 2012;3:104. doi: 10.3389/fgene.2012.00104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Moreno-Villanueva M, Eltze T, Dressler D, Bernhardt J, Hirsch C, Wick P, von Scheven G, Lex K, Buerkle A. The automated fadu-assay, a potential high-throughput in vitro method for early screening of DNA breakage. Altex-Altern Anim Ex. 2011;28:295–303. doi: 10.14573/altex.2011.4.295. [DOI] [PubMed] [Google Scholar]
- 42.Sergent JA, Paget V, Chevillard S. Toxicity and genotoxicity of nano-SiO2 on human epithelial intestinal HT-29 cell line. Ann Occup Hyg. 2012;56:622–630. doi: 10.1093/annhyg/mes005. [DOI] [PubMed] [Google Scholar]
- 43.Hendriks G, Atallah M, Morolli B, Calléja F, Ras-Verloop N, Huijskens I, Raamsman M, van de Water B, Vrieling H. The Tox-Tracker assay: novel GFP reporter systems that provide mechanistic insight into the genotoxic properties of chemicals. Toxicol Sci. 2012;125:285–298. doi: 10.1093/toxsci/kfr281. [DOI] [PubMed] [Google Scholar]
- 44.Karlsson HL. The comet assay in nanotoxicology research. Anal Bioanal Chem. 2010;398:651–666. doi: 10.1007/s00216-010-3977-0. [DOI] [PubMed] [Google Scholar]
- 45.Ostling O, Johanson KJ. Microelectrophoretic study of radiation-induced DNA damages in individual mammalian cells. Biochem Biophys Res Commun. 1984;123:291–298. doi: 10.1016/0006-291x(84)90411-x. [DOI] [PubMed] [Google Scholar]
- 46.Singh NP, McCoy MT, Tice RR, Schneider EL. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp Cell Res. 1988;175:184–191. doi: 10.1016/0014-4827(88)90265-0. [DOI] [PubMed] [Google Scholar]
- 47.Collins AR. The comet assay for DNA damage and repair: principles, applications, and limitations. Mol Biotechnol. 2004;26:249–261. doi: 10.1385/MB:26:3:249. [DOI] [PubMed] [Google Scholar]
- 48.Olive PL, Banáth JP. The comet assay: a method to measure DNA damage in individual cells. Nat Protoc. 2006;1:23–29. doi: 10.1038/nprot.2006.5. [DOI] [PubMed] [Google Scholar]
- 49.Smith CC, O’Donovan M, Martin EA. hOGG1 recognizes oxidative damage using the comet assay with greater specificity than Fpg or EndoIII. Mutagenesis. 2006;21:185–190. doi: 10.1093/mutage/gel019. [DOI] [PubMed] [Google Scholar]
- 50.Collins AR, Oscoz AA, Brunborg G, Gaivão I, Giovannelli L, Kruszewski M, Smith CC, Stetina R. The comet assay: topical issues. Mutagenesis. 2008;23:143–151. doi: 10.1093/mutage/gem051. [DOI] [PubMed] [Google Scholar]
- 51.García O, Mandina T, Lamadrid AI, Diaz A, Remigio A, Gonzalez Y, Piloto J, Gonzalez JE, Alvarez A. Sensitivity and variability of visual scoring in the comet assay. Results of an inter-laboratory scoring exercise with the use of silver staining. Mutat Res. 2004;556:25–34. doi: 10.1016/j.mrfmmm.2004.06.035. [DOI] [PubMed] [Google Scholar]
- 52.Watson C, Ge J, Cohen J, Pyrgiotakis G, Engelward BP, Demokritou P. High-throughput screening platform for engineered nanoparticle-mediated genotoxicity using CometChip technology. ACS Nano. 2014;8:2118–2133. doi: 10.1021/nn404871p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Brunborg G, Jackson P, Shaposhnikov S, Dahl H, Azqueta A, Collins AR, Gutzkow KB. High throughput sample processing and automated scoring. Frontiers Genet. 2014;5:373. doi: 10.3389/fgene.2014.00373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Shaposhnikov S, Azqueta A, Henriksson S, et al. Twelve-gel slide format optimised for comet assay and fluorescent in situ hybridisation. Toxicol Lett. 2010;195:31–34. doi: 10.1016/j.toxlet.2010.02.017. [DOI] [PubMed] [Google Scholar]
- 55.Sipinen V, Laubenthal J, Baumgartner A, Cemeli E, Linschooten JO, Godschalk RW, Van Schooten FJ, Anderson D, Brunborg G. In vitro evaluation of baseline and induced DNA damage in human sperm exposed to benzo[a]pyrene or its metabolite benzo[a] pyrene-7,8-diol-9,10-epoxide, using the comet assay. Mutagenesis. 2010;25:417–425. doi: 10.1093/mutage/geq024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Hansen SH, Olsen AK, Soderlund EJ, Brunborg G. In vitro investigations of glycidamide-induced DNA lesions in mouse male germ cells and in mouse and human lymphocytes. Mutat Res Gen Tox En. 2010;696:55–61. doi: 10.1016/j.mrgentox.2009.12.012. [DOI] [PubMed] [Google Scholar]
- 57.Gutzkow KB, Langleite TM, Meier S, Graupner A, Collins AR, Brunborg G. High-throughput comet assay using 96 minigels. Mutagenesis. 2013;28:333–340. doi: 10.1093/mutage/get012. [DOI] [PubMed] [Google Scholar]
- 58.Stang A, Witte I. Performance of the comet assay in a high-throughput version. Mutat Res Gen Tox En. 2009;675:5–10. doi: 10.1016/j.mrgentox.2009.01.007. [DOI] [PubMed] [Google Scholar]
- 59.Stang A, Witte I. The ability of the high-throughput comet assay to measure the sensitivity of five cell lines toward methyl methanesulfonate, hydrogen peroxide, and pentachlorophenol. Mutat Res. 2010;701:103–106. doi: 10.1016/j.mrgentox.2010.04.011. [DOI] [PubMed] [Google Scholar]
- 60.Stang A, Brendiamour M, Schunck C, Witte I. Automated analysis of DNA damage in the high-throughput version of the comet assay. Mutat Res Gen Tox En. 2010;698:1–5. doi: 10.1016/j.mrgentox.2010.02.014. [DOI] [PubMed] [Google Scholar]
- 61.Wood DK, Weingeist DM, Bhatia SN, Engelward BP. Single cell trapping and DNA damage analysis using microwell arrays. Proc Natl Acad Sci USA. 2010;107:10008–10013. doi: 10.1073/pnas.1004056107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Weingeist DM, Ge J, Wood DK, Mutamba JT, Huang Q, Rowland EA, Yaffe MB, Floyd S, Engelward BP. Single-cell microarray enables high-throughput evaluation of DNA double-strand breaks and DNA repair inhibitors. Cell Cycle. 2013;12:907–915. doi: 10.4161/cc.23880. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ge J, Prasongtanakij S, Wood DK, Weingeist DM, Fessler J, Navasummrit P, Ruchirawat M, Engelward BP. Cometchip: A high-throughput 96-well platform for measuring DNA damage in microarrayed human cells. Jove J Vis Exp. 2014;92:e50607. doi: 10.3791/50607. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Ge J, Chow DN, Fessler JL, Weingeist DM, Wood DK, Engelward BP. Micropatterned comet assay enables high throughput and sensitive DNA damage quantification. Mutagenesis. 2015;30:11–19. doi: 10.1093/mutage/geu063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Witte I, Plappert U, de Wall H, Hartmann A. Genetic toxicity assessment: employing the best science for human safety evaluation part III: the comet assay as an alternative to in vitro clastogenicity tests for early drug candidate selection. Toxicol Sci. 2007;97:21–26. doi: 10.1093/toxsci/kfl192. [DOI] [PubMed] [Google Scholar]
- 66.Azqueta A, Gutzkow KB, Priestley CC, Meier S, Walker JS, Brunborg G, Collins AR. A comparative performance test of standard, medium- and high-throughput comet assays. Toxicol In Vitro. 2013;27:768–773. doi: 10.1016/j.tiv.2012.12.006. [DOI] [PubMed] [Google Scholar]
- 67.Ge J, Wood DK, Weingeist DM, Prasongtanakij S, Navasumrit P, Ruchirawat M, Engelward BP. Standard fluorescent imaging of live cells is highly genotoxic. Cytometry, Part A. 2013;83:552–560. doi: 10.1002/cyto.a.22291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Cohen J, Deloid G, Pyrgiotakis G, Demokritou P. Interactions of engineered nanomaterials in physiological media and implications for in vitro dosimetry. Nanotoxicology. 2013;7:417–431. doi: 10.3109/17435390.2012.666576. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Gyori BM, Venkatachalam G, Thiagarajan PS, Hsu D, Clement MV. OpenComet: an automated tool for comet assay image analysis. Redox Biol. 2014;2:457–465. doi: 10.1016/j.redox.2013.12.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.González JE, Romero I, Barquinero JF, García O. Automatic analysis of silver-stained comets by CellProfiler software. Mutat Res. 2012;748:60–64. doi: 10.1016/j.mrgentox.2012.06.007. [DOI] [PubMed] [Google Scholar]
- 71.Collins A, El Yamani N, Lorenzo Y, Shaposhnikov S, Brunborg G, Azqueta A. Controlling variation in the comet assay. Front Genet. 2014:5. doi: 10.3389/fgene.2014.00359.. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Guan R, Kang T, Lu F, Zhang Z, Shen H, Liu M. Cytotoxicity, oxidative stress, and genotoxicity in human hepatocyte and embryonic kidney cells exposed to ZnO nanoparticles. Nanoscale Res Lett. 2012;7:602. doi: 10.1186/1556-276X-7-602. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.O’Donovan M. A critique of methods to measure cytotoxicity in mammalian cell genotoxicity assays. Mutagenesis. 2012;27:615–621. doi: 10.1093/mutage/ges045. [DOI] [PubMed] [Google Scholar]
- 74.Kumaravel TS, Vilhar B, Faux SP, Jha AN. Comet assay measurements: a perspective. Cell Biol Toxicol. 2009;25:53–64. doi: 10.1007/s10565-007-9043-9. [DOI] [PubMed] [Google Scholar]
- 75.Magdolenova Z, Lorenzo Y, Collins A, Dusinska M. Can standard genotoxicity tests be applied to nanoparticles? J Toxicol Environ Health A. 2012;75:800–806. doi: 10.1080/15287394.2012.690326. [DOI] [PubMed] [Google Scholar]
- 76.Manshian BB, Singh N, Doak SH. The in vitro micronucleus assay and kinetochore staining: methodology and criteria for the accurate assessment of genotoxicity and cytotoxicity. Methods Mol Biol. 2013;1044:269–289. doi: 10.1007/978-1-62703-529-3_14. [DOI] [PubMed] [Google Scholar]
- 77.AshaRani PV, Low Kah Mun G, Hande MP, Valiyaveettil S. Cytotoxicity and genotoxicity of silver nanoparticles in human cells. ACS Nano. 2009;3:279–290. doi: 10.1021/nn800596w. [DOI] [PubMed] [Google Scholar]
- 78.Papageorgiou I, Brown C, Schins R, Singh S, Newson R, Davis S, Fisher J, Ingham E, Case CP. The effect of nano- and micron-sized particles of cobalt-chromium alloy on human fibroblasts in vitro. Biomaterials. 2007;28:2946–2958. doi: 10.1016/j.biomaterials.2007.02.034. [DOI] [PubMed] [Google Scholar]
- 79.Avlasevich S, Bryce S, De Boeck M, Elhajouji A, Van Goethem F, Lynch A, Nicolette J, Shi J, Dertinger S. Flow cytometric analysis of micronuclei in mammalian cell cultures: past, present and future. Mutagenesis. 2011;26:147–152. doi: 10.1093/mutage/geq058. [DOI] [PubMed] [Google Scholar]
- 80.Nüsse M, Kramer J. Flow cytometric analysis of micronuclei found in cells after irradiation. Cytometry. 1984;5:20–25. doi: 10.1002/cyto.990050105. [DOI] [PubMed] [Google Scholar]
- 81.Bryce SM, Avlasevich SL, Bemis JC, Dertinger SD. Miniaturized flow cytometry-based CHO-K1 micronucleus assay discriminates aneugenic and clastogenic modes of action. Environ Mol Mutagen. 2011;52:280–286. doi: 10.1002/em.20618. [DOI] [PubMed] [Google Scholar]
- 82.Collins JE, Ellis PC, White AT, Booth AE, Moore CE, Burman M, Rees RW, Lynch AM. Evaluation of the Litron In Vitro MicroFlow Kit for the flow cytometric enumeration of micronuclei (MN) in mammalian cells. Mutat Res. 2008;654:76–81. doi: 10.1016/j.mrgentox.2008.05.003. [DOI] [PubMed] [Google Scholar]
- 83.Zucker RM, Daniel KM, Massaro EJ, Karafas SJ, Degn LL, Boyes WK. Detection of silver nanoparticles in cells by flow cytometry using light scatter and far-red fluorescence. Cytomet A. 2013;83:962–972. doi: 10.1002/cyto.a.22342. [DOI] [PubMed] [Google Scholar]
- 84.He X, Zhou L, He D, Wang K, Cao J. Rapid and ultra-sensitive E. coli O157:H7 quantitation by combination of ligandmagnetic nanoparticles enrichment with fluorescent nanoparticles based two-color flow cytometry. Analyst. 2011;136:4183–4191. doi: 10.1039/c1an15413c. [DOI] [PubMed] [Google Scholar]
- 85.Sahu SC, Njoroge J, Bryce SM, Yourick JJ, Sprando RL. Comparative genotoxicity of nanosilver in human liver HepG2 and colon Caco2 cells evaluated by a flow cytometric in vitro micronucleus assay. J Appl Toxicol. 2014;34:1226–1234. doi: 10.1002/jat.3065. [DOI] [PubMed] [Google Scholar]
- 86.Sahu SC, Njoroge J, Bryce SM, Zheng J, Ihrie J. Flow cytometric evaluation of the contribution of ionic silver to genotoxic potential of nanosilver in human liver HepG2 and colon Caco2 cells. J Appl Toxicol. 2016;36:521–531. doi: 10.1002/jat.3276. [DOI] [PubMed] [Google Scholar]
- 87.Shi J, Bezabhie R, Szkudlinska A. Further evaluation of a flow cytometric in vitro micronucleus assay in CHO-K1 cells: a reliable platform that detects micronuclei and discriminates apoptotic bodies. Mutagenesis. 2010;25:33–40. doi: 10.1093/mutage/gep040. [DOI] [PubMed] [Google Scholar]
- 88.Avlasevich SL, Bryce SM, Cairns SE, Dertinger SD. In vitro micronucleus scoring by flow cytometry: differential staining of micronuclei versus apoptotic and necrotic chromatin enhances assay reliability. Environ Mol Mutagen. 2006;47:56–66. doi: 10.1002/em.20170. [DOI] [PubMed] [Google Scholar]
- 89.Bryce SM, Bemis JC, Avlasevich SL, Dertinger SD. In vitro micronucleus assay scored by flow cytometry provides a comprehensive evaluation of cytogenetic damage and cytotoxicity. Mutat Res. 2007;630:78–91. doi: 10.1016/j.mrgentox.2007.03.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Gonzalez L, Lukamowicz-Rajska M, Thomassen LC, Kirschhock CE, Leyns L, Lison D, Martens JA, Elhajouji A, Kirsch-Volders M. Co-assessment of cell cycle and micronucleus frequencies demonstrates the influence of serum on the in vitro genotoxic response to amorphous monodisperse silica nanoparticles of varying sizes. Nanotoxicology. 2014;8:876–884. doi: 10.3109/17435390.2013.842266. [DOI] [PubMed] [Google Scholar]
- 91.Ruiz-Herrero T, Velasco E, Hagan MF. Mechanisms of budding of nanoscale particles through lipid bilayers. J Phys Chem B. 2012;116:9595–9603. doi: 10.1021/jp301601g. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Jiang X, Foldbjerg R, Miclaus T, Wang L, Singh R, Hayashi Y, Sutherland D, Chen C, Autrup H, Beer C. Multi-platform genotoxicity analysis of silver nanoparticles in the model cell line cho-k1. Toxicol Lett. 2013;222:55–63. doi: 10.1016/j.toxlet.2013.07.011. [DOI] [PubMed] [Google Scholar]
- 93.Sabharwal R, Verma P, Syed MA, Sharma T, Subudhi SK, Mohanty S, Gupta S. Emergence of micronuclei as a genomic biomarker. Indian J Med Paediatr Oncol. 2015;36:212–218. doi: 10.4103/0971-5851.171541. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Fenech M, Holland N, Zeiger E, et al. The HUMN and HUM-NxL international collaboration projects on human micronucleus assays in lymphocytes and buccal cells–past, present and future. Mutagenesis. 2011;26:239–245. doi: 10.1093/mutage/geq051. [DOI] [PubMed] [Google Scholar]
- 95.Bonassi S, Znaor A, Ceppi M, et al. An increased micronucleus frequency in peripheral blood lymphocytes predicts the risk of cancer in humans. Carcinogenesis. 2007;28:625–631. doi: 10.1093/carcin/bgl177. [DOI] [PubMed] [Google Scholar]
- 96.Rogakou EP, Boon C, Redon C, Bonner WM. Megabase chromatin domains involved in DNA double-strand breaks in vivo. J Cell Biol. 1999;146:905–916. doi: 10.1083/jcb.146.5.905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Scully R, Xie A. Double strand break repair functions of histone H2AX. Mutat Res. 2013;750:5–14. doi: 10.1016/j.mrfmmm.2013.07.007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Kuo LJ, Yang LX. Gamma-H2AX - a novel biomarker for DNA double-strand breaks. In Vivo. 2008;22:305–309. [PubMed] [Google Scholar]
- 99.Sedelnikova OA, Rogakou EP, Panyutin IG, Bonner WM. Quantitative detection of (125)IdU-induced DNA double-strand breaks with gamma-H2AX antibody. Radiat Res. 2002;158:486–492. doi: 10.1667/0033-7587(2002)158[0486:qdoiid]2.0.co;2. [DOI] [PubMed] [Google Scholar]
- 100.Ismail IH, Wadhra TI, Hammarsten O. An optimized method for detecting gamma-H2AX in blood cells reveals a significant interindividual variation in the gamma-H2AX response among humans. Nucleic Acids Res. 2007;35:e36. doi: 10.1093/nar/gkl1169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Bonner WM, Redon CE, Dickey JS, Nakamura AJ, Sedelnikova OA, Solier S, Pommier Y. GammaH2AX and cancer. Nat Rev Cancer. 2008;8:957–967. doi: 10.1038/nrc2523. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 102.Karlsson HL, Gliga AR, Calléja FM, Gonçalves CS, Wallinder IO, Vrieling H, Fadeel B, Hendriks G. Mechanism-based genotoxicity screening of metal oxide nanoparticles using the ToxTracker panel of reporter cell lines. Part Fibre Toxicol. 2014;11:41. doi: 10.1186/s12989-014-0041-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Kung ML, Hsieh SL, Wu CC, Chu TH, Lin YC, Yeh BW, Hsieh S. Enhanced reactive oxygen species overexpression by CuO nanoparticles in poorly differentiated hepatocellular carcinoma cells. Nanoscale. 2015;7:1820–1829. doi: 10.1039/c4nr05843g. [DOI] [PubMed] [Google Scholar]
- 104.Toduka Y, Toyooka T, Ibuki Y. Flow cytometric evaluation of nanoparticles using side-scattered light and reactive oxygen species-mediated fluorescence-correlation with genotoxicity. Environ Sci Technol. 2012;46:7629–7636. doi: 10.1021/es300433x. [DOI] [PubMed] [Google Scholar]
- 105.Wan R, Mo Y, Feng L, Chien S, Tollerud DJ, Zhang Q. DNA damage caused by metal nanoparticles: involvement of oxidative stress and activation of ATM. Chem Res Toxicol. 2012;25:1402–1411. doi: 10.1021/tx200513t. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Kermanizadeh A, Chauché C, Brown DM, Loft S, Møller P. The role of intracellular redox imbalance in nanomaterial induced cellular damage and genotoxicity: a review. Environ Mol Mutagen. 2015;56:111–124. doi: 10.1002/em.21926. [DOI] [PubMed] [Google Scholar]
- 107.Toyooka T, Amano T, Ibuki Y. Titanium dioxide particles phosphorylate histone h2ax independent of ros production. Mutat Res Gen Tox En. 2012;742:84–91. doi: 10.1016/j.mrgentox.2011.12.015. [DOI] [PubMed] [Google Scholar]
- 108.Tanaka T, Huang X, Halicka HD, Zhao H, Traganos F, Albino AP, Dai W, Darzynkiewicz Z. Cytometry of ATM activation and histone H2AX phosphorylation to estimate extent of DNA damage induced by exogenous agents. Cytomet A. 2007;71:648–661. doi: 10.1002/cyto.a.20426. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Suzuki H, Toyooka T, Ibuki Y. Simple and easy method to evaluate uptake potential of nanoparticles in mammalian cells using a flow cytometric light scatter analysis. Environ Sci Technol. 2007;41:3018–3024. doi: 10.1021/es0625632. [DOI] [PubMed] [Google Scholar]
- 110.Evans MD, Dizdaroglu M, Cooke MS. Oxidative DNA damage and disease: induction, repair and significance. Mutat Res. 2004;567:1–61. doi: 10.1016/j.mrrev.2003.11.001. [DOI] [PubMed] [Google Scholar]
- 111.Matsuda S, Matsuda R, Matsuda Y, Yanagisawa S, Ikura M, Ikura T, Matsuda T. An easy-to-use genotoxicity assay using egfp-mdc1-expressing human cells. Genes Environ. 2014;36:17–28. [Google Scholar]
- 112.Moreno-Villanueva M, Pfeiffer R, Sindlinger T, Leake A, Müller M, Kirkwood TB, Bürkle A. A modified and automated version of the ‘Fluorimetric Detection of Alkaline DNA Unwinding’ method to quantify formation and repair of DNA strand breaks. BMC Biotechnol. 2009;9:39. doi: 10.1186/1472-6750-9-39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Moreno-Villanueva M, Burkle A. High-throughput assays to quantify the formation of DNA strand breaks. In: Steinberg P, editor. High-throughput methods in toxicity testing. John Wiley and Sons; Hoboken: 2013. pp. 285–294. [Google Scholar]
- 114.Moreno-Villanueva M, Steinhaus R, Burkle TA. Quantification of DNA strand breaks by an automated version of fluorometric detection of alkaline unwinding (fadu) Exp Gerontol. 2009;44:131. [Google Scholar]
- 115.Dębiak M, Panas A, Steinritz D, Kehe K, Bürkle A. High throughput analysis of DNA interstrand crosslinks in human peripheral blood mononuclear cells by automated reverse FADU assay. Toxicology. 2011;280:53–60. doi: 10.1016/j.tox.2010.11.007. [DOI] [PubMed] [Google Scholar]
- 116.Wenzel P, Schuhmacher S, Kienhöfer J, et al. Manganese superoxide dismutase and aldehyde dehydrogenase deficiency increase mitochondrial oxidative stress and aggravate age-dependent vascular dysfunction. Cardiovasc Res. 2008;80:280–289. doi: 10.1093/cvr/cvn182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Müller N, Moreno-Villanueva M, Fischbach A, Kienhöfer J, Martello R, Dedon PC, Ullrich V, Bürkle A, Mangerich A. An automated Fpg-based FADU method for the detection of oxidative DNA lesions and screening of antioxidants. Toxicology. 2013;310:15–21. doi: 10.1016/j.tox.2013.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Cooke MS, Evans MD, Dizdaroglu M, Lunec J. Oxidative DNA damage: mechanisms, mutation, and disease. FASEB J. 2003;17:1195–1214. doi: 10.1096/fj.02-0752rev. [DOI] [PubMed] [Google Scholar]
- 119.Dizdaroglu M. Application of capillary gas chromatography-mass spectrometry to chemical characterization of radiation-induced base damage of DNA: implications for assessing DNA repair processes. Anal Biochem. 1985;144:593–603. doi: 10.1016/0003-2697(85)90158-7. [DOI] [PubMed] [Google Scholar]
- 120.Dizdaroglu M, Jaruga P, Rodriguez H. Measurement of 8-hydroxy-2′-deoxyguanosine in DNA by high-performance liquid chromatography-mass spectrometry: comparison with measurement by gas chromatography-mass spectrometry. Nucleic Acids Res. 2001;29:E12. doi: 10.1093/nar/29.3.e12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Valavanidis A, Vlachogianni T, Fiotakis C. 8-hydroxy-2′-deoxyguanosine (8-OHdG): A critical biomarker of oxidative stress and carcinogenesis. J Environ Sci Heal C. 2009;27:120–139. doi: 10.1080/10590500902885684. [DOI] [PubMed] [Google Scholar]
- 122.Junk M, Salzwedel J, Sindlinger T, Bürkle A, Moreno-Villanueva M. Mathematical modelling of the automated FADU assay for the quantification of DNA strand breaks and their repair in human peripheral mononuclear blood cells. BMC Biophys. 2014;7:9. doi: 10.1186/s13628-014-0009-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 123.Pati R, Das I, Mehta RK, Sahu R, Sonawane A. Zincoxide nanoparticles exhibit genotoxic, clastogenic, cytotoxic and actin depolymerization effects by inducing oxidative stress responses in macrophages and adult mice. Toxicol Sci. 2016;150:454–472. doi: 10.1093/toxsci/kfw010. [DOI] [PubMed] [Google Scholar]
- 124.Zhang Y, Nayak TR, Hong H, Cai W. Biomedical applications of zinc oxide nanomaterials. Curr Mol Med. 2013;13:1633–1645. doi: 10.2174/1566524013666131111130058. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 125.Kappes F, Fahrer J, Khodadoust MS, et al. DEK is a poly(ADP-ribose) acceptor in apoptosis and mediates resistance to genotoxic stress. Mol Cell Biol. 2008;28:3245–3257. doi: 10.1128/MCB.01921-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Korwek Z, Bielak-Zmijewska A, Mosieniak G, Alster O, Moreno-Villanueva M, Burkle A, Sikora E. DNA damage-independent apoptosis induced by curcumin in normal resting human T cells and leukaemic Jurkat cells. Mutagenesis. 2013;28:411–416. doi: 10.1093/mutage/get017. [DOI] [PubMed] [Google Scholar]
- 127.Korwek Z, Sewastianik T, Bielak-Zmijewska A, Mosieniak G, Alster O, Moreno-Villanueva M, Burkle A, Sikora E. Inhibition of atm blocks the etoposide-induced DNA damage response and apoptosis of human resting t cells. DNA Repair. 2013;12:872–872. doi: 10.1016/j.dnarep.2012.08.006. [DOI] [PubMed] [Google Scholar]
- 128.Alster O, Bielak-Zmijewska A, Mosieniak G, et al. The role of nibrin in doxorubicin-induced apoptosis and cell senescence in Nijmegen Breakage Syndrome patients lymphocytes. PLoS One. 2014;9:e104964. doi: 10.1371/journal.pone.0104964. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Brunner S, Herndler-Brandstetter D, Arnold CR, et al. Upregulation of miR-24 is associated with a decreased DNA damage response upon etoposide treatment in highly differentiated CD8(+) T cells sensitizing them to apoptotic cell death. Aging Cell. 2012;11:579–587. doi: 10.1111/j.1474-9726.2012.00819.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Kalfalah F, Seggewiß S, Walter R, et al. Structural chromosome abnormalities, increased DNA strand breaks and DNA strand break repair deficiency in dermal fibroblasts from old female human donors. Aging (Albany NY) 2015;7:110–122. doi: 10.18632/aging.100723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Maynard S, Hejl AM, Dinh TS, et al. Defective mitochondrial respiration, altered dNTP pools and reduced AP endonuclease 1 activity in peripheral blood mononuclear cells of Alzheimer’s disease patients. Aging (Albany NY) 2015;7:793–815. doi: 10.18632/aging.100810. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Maynard S, Keijzers G, Hansen AM, et al. Associations of subjective vitality with DNA damage, cardiovascular risk factors and physical performance. Acta Physiol (Oxf) 2015;213:156–170. doi: 10.1111/apha.12296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Garm C, Moreno-Villanueva M, Bürkle A, Petersen I, Bohr VA, Christensen K, Stevnsner T. Age and gender effects on DNA strand break repair in peripheral blood mononuclear cells. Aging Cell. 2013;12:58–66. doi: 10.1111/acel.12019. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Garm C, Moreno-Villanueva M, Bürkle A, Larsen LA, Bohr VA, Christensen K, Stevnsner T. Genetic and environmental influence on DNA strand break repair: a twin study. Environ Mol Mutagen. 2013;54:414–420. doi: 10.1002/em.21791. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Morath J, Moreno-Villanueva M, Hamuni G, Kolassa S, Ruf-Leuschner M, Schauer M, Elbert T, Bürkle A, Kolassa IT. Effects of psychotherapy on DNA strand break accumulation originating from traumatic stress. Psychother Psychosom. 2014;83:289–297. doi: 10.1159/000362739. [DOI] [PubMed] [Google Scholar]
- 136.Singh NP. Microgels for estimation of DNA strand breaks, DNA protein crosslinks and apoptosis. Mutat Res. 2000;455:111–127. doi: 10.1016/s0027-5107(00)00075-0. [DOI] [PubMed] [Google Scholar]
- 137.Gedik CM, Collins A ESCODD (European Standards Committee on Oxidative DNA Damage) Establishing the background level of base oxidation in human lymphocyte DNA: results of an interlaboratory validation study. FASEB J. 2005;19:82–84. doi: 10.1096/fj.04-1767fje. [DOI] [PubMed] [Google Scholar]
- 138.Hastwell PW, Chai LL, Roberts KJ, Webster TW, Harvey JS, Rees RW, Walmsley RM. High-specificity and high-sensitivity genotoxicity assessment in a human cell line: validation of the Green-Screen HC GADD45a-GFP genotoxicity assay. Mutat Res. 2006;607:160–175. doi: 10.1016/j.mrgentox.2006.04.011. [DOI] [PubMed] [Google Scholar]
- 139.Birrell L, Cahill P, Hughes C, Tate M, Walmsley RM. GADD45a-GFP GreenScreen HC assay results for the ECVAM recommended lists of genotoxic and non-genotoxic chemicals for assessment of new genotoxicity tests. Mutat Res. 2010;695:87–95. doi: 10.1016/j.mrgentox.2009.12.008. [DOI] [PubMed] [Google Scholar]
- 140.Hughes C, Rabinowitz A, Tate M, Birrell L, Allsup J, Billinton N, Walmsley RM. Development of a high-throughput Gaussia luciferase reporter assay for the activation of the GADD45a gene by mutagens, promutagens, clastogens, and aneugens. J Biomol Screen. 2012;17:1302–1315. doi: 10.1177/1087057112453312. [DOI] [PubMed] [Google Scholar]
- 141.Simpson K, Bevan N, Hastwell P, Eidam P, Shah P, Gogo E, Rees S, Brown A. The BlueScreen-384 assay as an indicator of genotoxic hazard potential in early-stage drug discovery. J Biomol Screen. 2013;18:441–452. doi: 10.1177/1087057112470858. [DOI] [PubMed] [Google Scholar]
- 142.Rosemary Siafakas A, Richardson DR. Growth arrest and DNA damage-45 alpha (GADD45alpha) Int J Biochem Cell Biol. 2009;41:986–989. doi: 10.1016/j.biocel.2008.06.018. [DOI] [PubMed] [Google Scholar]
- 143.Hendriks G, Derr RS, Misovic B, Morolli B, Calléja FM, Vrieling H. The Extended ToxTracker Assay Discriminates Between Induction of DNA Damage, Oxidative Stress, and Protein Misfolding. Toxicol Sci. 2016;150:190–203. doi: 10.1093/toxsci/kfv323. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 144.Gliga AR, Skoglund S, Wallinder IO, Fadeel B, Karlsson HL. Size-dependent cytotoxicity of silver nanoparticles in human lung cells: the role of cellular uptake, agglomeration and Ag release. Part Fibre Toxicol. 2014;11:11. doi: 10.1186/1743-8977-11-11. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Behl M, Stout MD, Herbert RA, Dill JA, Baker GL, Hayden BK, Roycroft JH, Bucher JR, Hooth MJ. Comparative toxicity and carcinogenicity of soluble and insoluble cobalt compounds. Toxicology. 2015;333:195–205. doi: 10.1016/j.tox.2015.04.008. [DOI] [PubMed] [Google Scholar]
- 146.Nel AE, Madler L, Velegol D, Xia T, Hoek EM, Somasundaran P, Klaessig F, Castranova V, Thompson M. Understanding biophysicochemical interactions at the nano-bio interface. Nat Mater. 2009;8:543–557. doi: 10.1038/nmat2442. [DOI] [PubMed] [Google Scholar]
- 147.Godwin H, Nameth C, Avery D, et al. Nanomaterial categorization for assessing risk potential to facilitate regulatory decision-making. ACS Nano. 2015;9:3409–3417. doi: 10.1021/acsnano.5b00941. [DOI] [PubMed] [Google Scholar]
- 148.Nel AE. Implementation of alternative test strategies for the safety assessment of engineered nanomaterials. J Intern Med. 2013;274:561–577. doi: 10.1111/joim.12109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 149.Elzey S, Tsai DH, Yu LL, Winchester MR, Kelley ME, Hackley VA. Real-time size discrimination and elemental analysis of gold nanoparticles using ES-DMA coupled to ICP-MS. Anal Bioanal Chem. 2013;405:2279–2288. doi: 10.1007/s00216-012-6617-z. [DOI] [PubMed] [Google Scholar]
- 150.Mader BT, Ellefson ME, Wolf ST. Measurements of nanomaterials in environmentally relevant water matrices using liquid nebulization/differential mobility analysis. Environ Toxicol Chem. 2015;34:833–842. doi: 10.1002/etc.2865. [DOI] [PubMed] [Google Scholar]
- 151.Paget V, Sergent JA, Grall R, et al. Carboxylated nanodiamonds are neither cytotoxic nor genotoxic on liver, kidney, intestine and lung human cell lines. Nanotoxicology. 2014;8:46–56. doi: 10.3109/17435390.2013.855828. [DOI] [PubMed] [Google Scholar]
- 152.Paget V, Moche H, Kortulewski T, Grall R, Irbah L, Nesslany F, Chevillard S. Human cell line-dependent WC-Co nanoparticle cytotoxicity and genotoxicity: a key role of ROS production. Toxicol Sci. 2015;143:385–397. doi: 10.1093/toxsci/kfu238. [DOI] [PubMed] [Google Scholar]
- 153.Cho TJ, MacCuspie RI, Gigault J, Gorham JM, Elliott JT, Hackley VA. Highly stable positively charged dendron-encapsulated gold nanoparticles. Langmuir. 2014;30:3883–3893. doi: 10.1021/la5002013. [DOI] [PubMed] [Google Scholar]
- 154.Jain KK. The role of nanobiotechnology in the development of personalized medicine. Med Princ Pract. 2011;20:1–3. doi: 10.1159/000319928. [DOI] [PubMed] [Google Scholar]
- 155.Asati A, Santra S, Kaittanis C, Perez JM. Surface-charge-dependent cell localization and cytotoxicity of cerium oxide nanoparticles. ACS Nano. 2010;4:5321–5331. doi: 10.1021/nn100816s. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 156.Brown SD, Nativo P, Smith JA, et al. Gold nanoparticles for the improved anticancer drug delivery of the active component of oxaliplatin. J Am Chem Soc. 2010;132:4678–4684. doi: 10.1021/ja908117a. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Wang F, Banerjee D, Liu Y, Chen X, Liu X. Upconversion nanoparticles in biological labeling, imaging, and therapy. Analyst. 2010;135:1839–1854. doi: 10.1039/c0an00144a. [DOI] [PubMed] [Google Scholar]
- 158.Joo J, Kwon D, Yim C, Jeon S. Highly sensitive diagnostic assay for the detection of protein biomarkers using microresonators and multifunctional nanoparticles. ACS Nano. 2012;6:4375–4381. doi: 10.1021/nn301071c. [DOI] [PubMed] [Google Scholar]
- 159.Etheridge ML, Campbell SA, Erdman AG, Haynes CL, Wolf SM, McCullough J. The big picture on nanomedicine: the state of investigational and approved nanomedicine products. Nanomedicine. 2013;9:1–14. doi: 10.1016/j.nano.2012.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 160.Powell AA, Talasaz AH, Zhang H, et al. Single cell profiling of circulating tumor cells: transcriptional heterogeneity and diversity from breast cancer cell lines. PLoS One. 2012;7:e33788. doi: 10.1371/journal.pone.0033788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Shoemaker RH. The NCI60 human tumour cell line anticancer drug screen. Nat Rev Cancer. 2006;6:813–823. doi: 10.1038/nrc1951. [DOI] [PubMed] [Google Scholar]
- 162.Harris G, Palosaari T, Magdolenova Z, et al. Iron oxide nanoparticle toxicity testing using high-throughput analysis and high-content imaging. Nanotoxicology. 2015;9:87–94. doi: 10.3109/17435390.2013.816797. [DOI] [PubMed] [Google Scholar]