

ABSTRACT
It is undeniably very logical to first formulate an unambiguous definition of “Life” before engaging in defining the parameters instrumental to Life's evolution. Because nearly everybody assumes, erroneously in my opinion, that catching Life's essence in a single sentence is impossible, this way of thinking remained largely unexplored in evolutionary theory. Upon analyzing what exactly happens at the transition from “still alive” to “just dead,” the following definition emerged. What we call “Life” (L) is an activity. It is nothing other than the total sum (∑) of all communication acts (C) executed, at moment t, by entities organized as sender-receiver compartments: L = ∑C Such “living” entities are self-electrifying and talking ( = communicating) aggregates of fossil stardust operating in an environment heavily polluted by toxic calcium. Communication is a multifaceted, complex process that is seldom well explained in introductory textbooks of biology. Communication is instrumental to adaptation because, at the cellular level, any act of communication is in fact a problem-solving act. It can be logically deduced that not Natural Selection itself but communication/problem-solving activity preceding selection is the universal driving force of evolution. This is against what textbooks usually claim, although doubt on the status of Natural Selection as driving force has been around for long. Finally, adopting the sender-receiver with its 2 memory systems (genetic and cognitive, both with their own rules) and 2 types of progeny (”physical children” and “pupils”) as the universal unit of architecture and function of all living entities, also enables the seamless integration of cultural and organic evolution, another long-standing tough problem in evolutionary theory. Paraphrasing Theodosius Dobzhansky, the very essence of biology is: “Nothing in biology and evolutionary theory makes sense except in the light of the ability of living matter to communicate, and by doing so, to solve problems.”
KEYWORDS: adaptation, Calcigender, cultural evolution, definition of Life, EES, electrome, evo-devo, Metadarwinism, Neo-Darwinism, selection
Introduction
Thanks to Charles Darwin1 and Alfred Russel Wallace2 evolutionary theory with focus on species formation with Natural Selection as its driving force was founded. It became the very heart of Biology. Over the years its foundations became ever better documented. Micro-evolution is the change in allele frequencies that occur over time within a population. It is relevant to the emergence of new species. Macro-evolution acts on a scale of separated gene pools. It occurs at or above the level of species. Mega-evolution is a more recent approach.3 It does not specifically focus on genetic changes but it attempts to describe the evolution of Life in its totality, irrespective of the way Life manifests itself in the wealth of prokaryotic and eukaryotic species and their communities. Although viruses are not living entities themselves, their role in evolution should not be neglected. Indeed, viruses outnumber prokaryotic and cellular life forms 10 times, and all cellular life is constantly in interaction with genetic parasites. Hence, the virosphere should be integrated into the realm of “life.”4 The Mega-evolution approach requires that one first unambiguously defines what Life exactly is before engaging in analyzing the various mechanisms/parameters instrumental to its change in the long run (evolution).
From the standpoint of philosophy of science this is the logical way to proceed. Yet, such approach did not really get ground in the mainstream of current evolutionary theory, as exemplified in the formulation of the neo-Darwinian New Synthesis (NS),5,6,7,8,9,10,11 Bowler11,12 and Kutschera and Niklas13 summarized the historical development of the term New Synthesis, from George Romanes who introduced the term to refer to the version of evolution advocated by Alfred Russel Wallace2 and August Weismann (1834–1914) with its heavy dependence on Natural Selection, to Stephen J. Gould.10 In the context of this paper, it deserves mentioning that Natural Selection has not always been accepted as the main driving force of evolution. In his book “Evolution: the modern synthesis” Julian Huxley14 used the phrase “the eclipse of Darwinism” to describe the state of affairs before the modern evolutionary synthesis, when relatively few biologists thought that natural selection was the primary driving force of evolution.11,12 Thus, the question: “What is cause and what is result in evolution?” has been asked repeatedly over a long time. To date the question could be formulated this way: Should, perhaps, Natural Selection be replaced by Problem-solving activity as the universal driving force of evolution?
The main reason for the duality in which theory and practice do not match resides in the widely accepted idea that it is impossible to catch the nature of Life in a single sentence, a statement that features in textbooks of general biology, edition after edition for at least half a century up to the present.15 Such (unwarranted) statement discourages younger newcomers with an interest in such a fundamental question to engage in searching for a plausible answer.
This paper summarizes (at the end of my scientific career) the key elements of my ideas on an alternative approach of evolutionary theory that were published during the past 20 y (more information in References list). The goal is to unite the different aspects into an integrative concept that is accessible to both the exact sciences and the humanities. The essence of my approach has been that, if one first defines “Life”16 and the parameters instrumental to its incessant change, in particular the undervalued electrical properties of cells and the cognitive memory system, a novel paradigm (originally launched as “Mega-Evolution”3) emerges. It can upgrade the often undervalued input of the humanities in improving the wording of evolutionary theory, resulting in much more unity in Biology. An unavoidable consequence of my approach is that there are many more parameters instrumental to evolutionary change than the genetic- and epigenetic changes that dominate the thinking in the New Synthesis. Indeed, the 4 pillars of Life, in digital era wording its hardware-, software-, energy- and motivation aspects,3 all can change in many different ways. Yet, a simple concept, namely the sender-receiver, allows to integrate them all.
An unambiguous definition of Life has already been deduced 20 y ago
In the late 1980s, I was challenged by my undergraduate students to come up with a plausible definition to “What is Life?,” this to make me more credible as a professor of biology, the science of Life. Their reasoning was: “Why should we engage in the study of Life ( = Biology) if one cannot define what Life is?” I accepted the challenge, but I soon experienced that many had tried before me, without much success. No wonder as one gets confronted with all the criteria a good definition of Life should meet according to the philosophers of science Schejter and Agassi.17 Their wording was: “Apart from its not being trite and uninformative (circular, to use a traditional term), it should be neither too wide nor too narrow; it should not exclude living things and it should not include dead ones. Furthermore, it should not make biology part-and-parcel of chemistry and physics (meaning that there should be room for an immaterial dimension).” I add: “and it should organize all known dimensions and properties of living matter in a logical order and context, and it should pave the way for defining what exactly happens at the moment of death.”
But why did (and still does) nearly everybody assume that Life cannot be defined? To my own surprise, the very reason turned out to be the result of the combination of asking the wrong question, with an understandable but nevertheless fatal thinking error. Indeed, the common procedure at that time (the 1980s-90s) was to try to deduce the properties of Life by comparing the properties of living matter with those of non-living or inanimate matter, assuming that these 2 conditions are true opposites like warm-cold, high-low etc. True opposites can only have one counterpart. But a given living entity, e.g. a dog can be opposed to a myriad of non-living entities: a bottle, a ring, a brick, a ship etc. Thus they are false opposites. The true opposites with respect to Life are: “still alive” versus “just dead”. This urges for answering: “What exactly changes at the very moment of Death?” Answering that question in a non-circular way e.g., “death ensues when life ends” was less simple than I had expected. It yielded the insights that: 1. Death ends an activity of a given system. That activity turned out to be communication activity of systems organized as sender-receiver communicating compartments; 2. There are more levels of (communicational) complexity in most living systems than the 4–5 that textbooks usually list, at least 16 in my classification system (see later).
The following definitions emerged:
-
Communication. Numerous definitions of communication have already been formulated one more complex than the other, but seldom all-encompassing.3 My preferred definition reads: “Communication is transfer/handling of information in a system organized as a sender-receiver communicating compartment” (Fig. 1A). Any act of communication is generated as follows. A sender or the environment produces and releases a message(s) which is always written in coded form into what is called ‘a communication channel’ (blood, water, air etc.). The message (usually transported with the help of some carrier) will eventually arrive at a competent receiver ( = with matching receptors). Here it will be captured, decoded, amplified and responded to by causing the mobilization of part of the stockpiled energy to do some sort of ‘work’ sooner or later, e.g., by engaging in feedback. Depending upon the complexity of the system, numerous acts of communication can be simultaneously executed. All parts of any communicating compartment are subject to change; the sender, the message/messenger system, the transmission channel, the receiver, the feedback loops etc.
Feedback is not a circular, but a spiral-like unidirectional activity (Fig. 1B). When the complexity of a signaling system increases, the possibility for generating more than one answer may arise; this happens at bifurcation points (Fig. 1B). In my opinion this necessity to make a choice between 2 or more possibilities is the very basis of free will. The more bifurcation points, the more possibilities for making use of free will.
- Compartment. A biologic communicating compartment, or simply “compartment,” is a unit based on carbon chemistry and on electricity carried by inorganic ions. This unit
-
–is limited by a moderately leaky boundary with appropriate holes;
-
–can stockpile the right form(s) and amounts of energy;
-
–can generate gradients that can be used for communication for the purpose of enabling the compartment to function from its lowest to its highest level of compartmental organization.3
-
–
Information. Many definitions exist. My definition of information, intended for use in a physiologic context, reads: A message contains information when, upon being decoded by a competent receiver ( = a receiver with the proper receptor(s)), part of the stored energy in that receiver is mobilized for doing some sort of work. This is the meaning of AT WORK in Fig. 1A. Information is itself immaterial, but it usually needs a carrier for being transported.3,16,20
-
Life. A logically deduced unambiguous definition of Life (as an activity) reads: Life sounds like a noun (in English), but it denotes an activity, making it more into the meaning of a verb. What we call ‘Life’ (L) is nothing other than the total sum (∑) of all acts of communication (C) exerted by a given sender-receiver compartment at moment t, at all levels of its compartmental organization (cell organelle, cell, tissue,…, whole organism,…, population, community, Gaia level). The simplest symbolic notation reads: L = ∑C.3,16,21 Because Life is an activity of a given sender-receiver compartment of which there exist many different forms, one can specify the definition further as:
L = Life; S = type of compartment; t = moment at which the communication acts are executed; 1 = lowest level of compartmental organization (1 = prokaryotic cell or cell organelle in a eukaryotic cell); j = highest level of compartmental organization (cell, tissue, organ, organism, aggregate, …, population, community, the Gaia-level). For a symbolic notation that highlights how to compare biological and mechanical life (e.g., computer-life), see refs. 3, 21. Thus Life has both a qualitative (nature of the communication acts) and a quantitative (number of communication acts) aspect.
As to the origin of Life: it came into being at the very moment that the first act of communication was executed.22 Fig. 2. illustrates this event in cartoon form.23 How in pre-biotic conditions a living entity could have come into existence chemically has been discussed by Guth.24,25 The reasons why I think that the synthesis of actin-like molecules was as important as that of RNA or DNA as information carriers, namely for introducing polarity and for serving as an anchor for other macromolecules have been outlined before.3,20,22
Death ensues when a given communicating compartment irreversibly (to exclude regeneration) loses its ability to communicate at its highest level of compartmental organization, the total number of such levels . It follows that the essence of being alive, or of Life as an activity, is communication activity. At the level of the cell, death ensues when its self-generated ‘electrical dimension’ (see later in section 5) irreversibly collapses.
Time. If one thinks that the definition of Life also requires that Time has to be defined as well, I tried to formulate a definition that appeals to biologists, more than the more abstract ones worded by theoretical physicists. In my opinion Time is invariably a property of a given energy-converting system. It is a measure for the inertia of the conversion of a given form of energy (heat, light, chemical etc.) into another form(s), plus increase in entropy of the system (second law of thermodynamics). There are as many different times as there are energy converting systems.3 This definition does not at all unveil why there is inertia in energy conversion, thus why such conversions do not proceed with an infinitely high speed. This continues to be a big mystery in physics.
Figure 1.
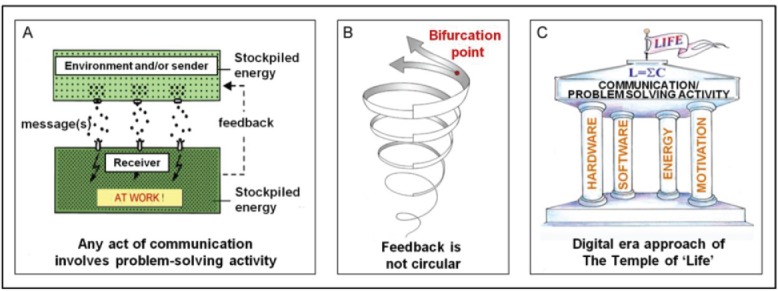
The classical sender-receiver or communicating compartment (A) is a better alternative than the cell for functioning as the universal unit of structure and function of all living matter. Feedback is a spiral-like, unidirectional process (B). At bifurcation points, a choice has to be made as to how to proceed with communication. In digital-era wording, the Temple of Life has only 4 pillars (C), in contrast to the classical PICERAS Temple of Life that has 7.18,19
Figure 2.

Cartoon illustrating my view that Life came into being at the very moment that the first act of communication was executed. Which act and under which environmental conditions this happened is unknown. Adapted from ref. 19.
Evolution of Life
If L = ∑C is an acceptable symbolic notation for Life, the simplest symbolic notation for its evolution becomes:
Some of the novelties in Mega-Evolution as compared with classical evolutionary theory (neo-Darwinian new synthesis)
The common descent principle was never better documented
This principle represents the very heart of Darwinian/Lamarckian evolutionary theory in both the New Synthesis and in the Mega-Evolution approach. Today it is very well experimentally documented.26 In the past, a truly major novel insight has been formulated by the late Lynn Margulis. According to her symbiogenesis theory,27 the eukaryotic cell came into being when at least 3 different ancient prokaryotic species established a functional symbiotic novel level of compartmental organization. Later in evolution, ever more complex multicellular eukaryotic entities came into being, requiring ever more complex coordinating signaling systems. The consequence of Margulis' theory is that, in fact, all life forms on the planet earth, thus both the contemporary genuine prokaryotes as well as all eukaryotes are manifestations of the only existing planetary form of life, which is prokaryotic (Archaea + bacteria) in origin and nature. No other forms of life are known. According to28 acquiring genomes was an important issue in the origins of species.
Not “the cell” but “the sender-receiver” as the universal unit of structure and function of all living matter
It has been outlined before3,23 that the sender-receiver (Fig. 1A) better serves the role of universal unit of both structure and function of all living matter than ‘the cell’. In origin the term cellulae was used by Robert Hooke (1635–1703) to denote the small chambers in cork. Later Matthias Jakob Schleiden and Theodor Schwann described that all living matter is made up of cellulae. The prokaryotic cell is the smallest sender-receiver. The Gaia-level is the highest one. In my opinion, any level can act as unit of selection.
Levels of complexity in communicating compartments: More numerous than in classical biology
In introductory textbooks of biology, the usual levels of complexity are; cell organelle, cell, organism, population and community. In the Mega-evolution approach which uses communication as criterion for grouping ‘entities’, there are at least 16 levels of compartmental organization in living matter. This has been described at length elsewhere.3,21,23 The ≥ 16 levels can be grouped into 3 categories. Witzany29 handles a similar communication-based classification system.
Compartments restricted to a single individual (levels 1–8): prokaryote, eukaryote, cell aggregate, syncytium, mono-epithelium, polyepithelium, segmented organism, tool-utilizing compartment.
Compartments with individuals of the same species (levels 9–14): colony, heterosexual and social compartments, baby inside mother (internal budding) compartment, population/species, electrosphere compartment (e.g., humans linked by telephone (in all its variants), radio, internet etc.).
Compartments with individuals belonging to different species (levels 15–16): the community (with nutritional and/or protective aspects), and the planetary or Gaia compartment.
In classical evolutionary theory the main focus is on the population and species (genetics), which is level 13 (out of 16) in my classification system that takes into account the signaling pathways at all levels.
Instead of body and mind/soul, rather hardware and software
One of the many reasons why it took so long before a plausible definition of Life was formulated resided in the absence of an adequate vocabulary. The dichotomy Soma or Body and - for humans - Soul or Mind reigned in Western culture for millennia. In Asian culture, that distinction was less clear-cut. Yet, defining Soul was not evident. The term disappeared from the core of Psychology as a discipline (psyche = soul in Greek), but it continues to be an essential element in (some) religious and spiritual activities. The question whether only humans have a soul or whether other organisms, in particular animals, also have a soul and are conscious20,30,31 continues to be asked again and again.32,33 It is better replaced by the questions how the cognitive memory works, how widespread such memory system is, and what its relation is with the cells “electrome” (a term recently introduced,20 consciousness and problem-solving. Since the start of the digital era, the terms hardware and software (Fig. 1C) became widely accepted for computers. In biology and in particular in evolutionary theory, they are useful, be it that this is not yet common practice. Hardware replaces Soma. Chemically the hardware of organisms is literally made up of fossil stardust.3 Software helps to describe some aspects of the still enigmatic cognitive memory. It is not a substitute for Soul, a term which is widely used in daily life and in some disciplines of the humanities, but not in Biology.20
Largely overlooked: Cells have a multifunctional, truly vital, self-generated electrical dimension–the electrome
A brief introduction to the basic principles and the multifunctionality of the electrical dimension of cells was formulated by.34 See also textbooks of physiology.
Transmembrane voltage gradient
Hitherto, the main focus of nearly all approaches to organic evolution, less to cultural evolution, was on genetics, from the molecular to the population level. Mutations of all sorts represent the very heart of genetic change and variability. In recent years, they got company from epigenetic changes (see later in Fig. 4). The fact that some simple inorganic ions like H+, K+, Na+, Ca2+, Cl−, HCO− (to name the important ones) also play a role in control of gene expression and development34,35 is seldom mentioned, and almost never taken into account in any evolutionary theory. Yet such ions are absolutely essential for being alive.
Figure 4.

Major genetic and non-genetic causes of (Communicational) variability. Not only Charles Darwin, but his contemporary Alfred Russel Wallace as well, independently conceived the theory of evolution through natural selection. Jean-Baptiste Pierre Antoine de Monet, Chevalier de Lamarck (1744–1829) but commonly referred to as simply Lamarck is best known for his theory of inheritance of acquired characteristics that was proven wrong in the context of classical genetics (pictures from Wikipedia, with thanks). Epigenetics is a form of temporary transfer of genetic information (through DNA- and/or histone modification) to the next (few) generation(s). According to some researchers such transfer is Lamarckian in nature. Cultural evolution is also mainly Lamarckian in nature.
The 2 major aspects of self-generated cellular electricity are: 1. The voltage gradient over the plasma membrane; and 2. Transcellular electric currents (Fig. 3). In animal cells, the transmembrane voltage gradient amounts to approximately 20–80 millivolt, depending upon the cell type and physiologic conditions. Expressed in Volts per cm, such seemingly small voltage difference over the very thin plasma membrane corresponds to about 50,000 Volts per cm, which is truly huge. In bacteria this figure is even higher.
Figure 3.
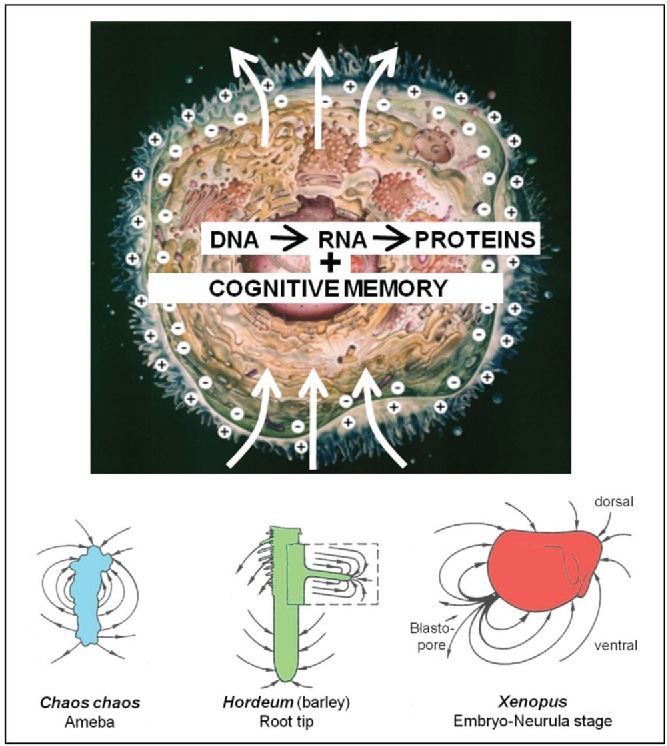
Self-generated electrical activity by cells. All cells generate an inorganic ion-based voltage difference over their plasma membrane. In resting conditions, the cytoplasmic side of the plasma membrane is at negative potential. The inorganic ion environment in the cytoplasm and (if present) in the nucleoplasm influences both gene expression and the still enigmatic cognitive memory system. Mechanisms exist that make that ion pumps and channels, the key actors instrumental to the ‘electrical dimension of cells’, do not float freely in the plane of the plasma membrane. Non-spherical symmetry can result. Under proper conditions, self-generated electrical currents traverse the cell. The current loops have to be closed outside the cell. Three examples of the electric field which surrounds a living entity are shown: an ameba, a plant and an animal embryo. They were measured with the vibrating probe technique of Jaffe and Nuccitelli.36 For more examples see ref. 19.
How cells build up a voltage gradient is very well documented by electrophysiological experiments in many cell types, but in particular in neurons and muscle cells. Here, the signaling function of action potentials has been intensively studied. One should keep in mind that the electrical dimension of cells and tissues is lost upon homogenization. Furthermore, although transport of inorganic ions requires proteins, in particular ion pumps and channels which are coded for in the genomes of all organisms, the roles of the inorganic ions themselves are not coded as such in the genome. Hence their role in control of gene expression, e.g., by changing the conformation of some proteins and of chromatin is not controlled by the genome. In addition, the electrical dimension of cells does not arise de novo at each cell division. Upon undergoing division the parental cell shares part of its plasma membrane with its inherent electrical activity to the daughter cells (for figures, see ref. 20). Thus any extended theory of evolution (EES) that only focuses upon genetic changes and transfer of only genetic information to the next generation, but that disregards the sharing of the electrical activity and, where relevant, the transfer of cognitive information (by teaching-learning) is due to be incomplete.
Self-generated transcellular electric currents: The cell as a miniature electrophoresis chamber-concept, the cell's electrome
It took the development of a very sensitive piece of equipment called “the vibrating probe” by Jaffe and Nuccitelli36 before it became technically possible to detect weak self-generated electric fields around very small entities, e.g., a single ameba, a root tip, a slime mold, a very young fruit fly embryo etc. (see Fig. 3 in ref. 19). Before, one was not even aware that such a hidden extracellular electric field even existed. If e.g., an ameba is surrounded by a self-generated electric field, the ionic current leaves the cell at a particular location(s) of the cell and enters at one or more other sites, this to close the current loops (Fig. 3). This polarity is due to the fact that the ion transporting proteins (ion pumps and ion channels) which reside in the plasma membrane are not spherically symmetrically distributed over the whole surface of the cell. They do not float freely in the plane of the liquid (lipid) membrane because they are held in place by some mechanisms, e.g., by being anchored to specific proteins of the cytoskeleton. This way, some pumps and channels can be kept segregated from each other and realize the transcellular transport of inorganic ions and an extracellular electric field.
The transcellular ionic current system means that charged macromolecules which are in solution in the cytoplasmic fluid can be forced to undergo electrophoretic transport. This system of self-electrophoresis was originally discovered in ovarioles (egg tubes) of a silk moth.37 Several years later, it yielded the concept of the cell as a miniature electrophoresis chamber.34 More recently, it also yielded the concept: The cell's self-generated electrome.20
Adaptation
The possible role of genetic and epigenetic changes in adaptation is well documented. Yet, other mechanisms have to be taken into account as well.
Also overlooked: Organisms have two memory systems, two possible types of progeny—namely physical children and pupils
Like any sender-receiver all prokaryotic and eukaryotic cells on earth probably have 2 memory systems, a genetic- and a cognitive one, each with its own set of rules. The central dogma of molecular biology (DNA → RNA → Proteins)38 represents the very heart of the functioning of the genetic memory. Today its functioning is well understood. In contrast, despite all progress in the neurosciences, the biochemical functioning of the cognitive memory remains largely a black box.39 One of the results is that “inclusive inheritance”40 uses heredity ( = through genes) for all transfer of information to the next generation instead of transferability of information to the next generation (and laterally as well), which allows taking into account teaching-learning involving the non-genetic aspects of the cognitive memory.
Physical children are the progeny generated through the principles of the genetic memory that underlies the formation of the hardware of organisms. Pupils are the progeny generated through the cognitive memory system, where relevant.
Any act of communication is a problem-solving act by definition and can hence be instrumental to adaptation: Semiosis
Why is an act of communication, at the cellular level, invariably a problem-solving act (Fig. 1A)? This follows from the fact that any message, whatever its nature, is coded. Hence, when the message (often, if not always transported with the help of some carrier) arrives at the receiver and is captured there, it next needs to be decoded before it can trigger the receiver to ‘do something with it’, either instantly, or later after storage for some time, or it can be deleted. We understand our mother tongue but no other (foreign) languages because in our childhood our parents, family members, our broad environment etc. installed - by teaching - in our brain the decoding programs for our mother tongue. Mimicry was also important. That gives us the impression that understanding our mother tongue is not a problem-solving activity. This interpretation is wrong; it is an automated decoding activity. The causal link between signaling and problem-solving is not commonly emphasized in the “exact biologic sciences” contrary to its status in the humanities, in particular in linguistics. However, one should always keep in mind that since Kurt Gödel (1931) we know that, contrary to what many intuitively assume, it is not possible to generate exact scientific sentences in principle.
In linguistics the term ‘semiosis’ or ‘sign process’ is routinely used.41,42,43,44 It was introduced by Charles Sanders Peirce (1830–1914) to denote any form of activity, conduct, or process that involves signs, including the production of meaning. I agree with Kull and Emmeche43 that because it incessantly interprets signs and signals, “Life is semiosis.” My wording L = ∑C3,21 said the same, but in the wording of the exact biologic sciences.
Adaptation to an environment poisoned by high Ca2+-concentrations
Organisms have to adapt to changing external conditions. The environment can become dryer, wetter, colder, warmer, less rich in food supply, populated by more parasites etc. When chemical pollution as an adverse condition is at stake, one usually thinks at man-caused pollution by pesticides, heavy metals, CO2 etc. Yet the most toxic pollutant on earth (O2 not taken into account) is the omnipresent Ca2+-ion. This may look strange because we encounter the beneficial aspects of Ca2+ in our daily life: our calcareous bony skeleton, Ca2+-rich milk, the egg shell of birds, and Ca2+ as a secondary messenger.45,46 Yet, because above a very low threshold a rise in cytoplasmic Ca2+ concentration is very toxic as it causes changes in the conformation of some essential macromolecules, in particular proteins. In fact it is because of this toxic effect on proteins that Ca2+ can act as secondary messenger. The intracellular Ca2+ concentration in the cytoplasm of unstimulated cells amounts to the very low value of about 100 nM. The extracellular concentration is many orders of magnitude higher, namely about 1–60 mM (about 2 mM in blood). Thus there is a gradient of about 100,000 times in Ca2+-concentration cytoplasm-outside cells. If the intracellular Ca2+-concentration rises too much, for too long, cells can get damaged and may even enter the apoptosis cell death cycle (Calcium-induced apoptosis.47 The duration of the heart contraction cycle which is based upon periodic Ca2+-release from the SER followed by fast re-uptake is an indication of what “too long” means, namely in the order of seconds rather than of minutes in most cell types. The toxicity of Ca2+ means that cells have to continuously fight against the influx of excess Ca2+ from the outside world (environment). Their major weapon is the different types of ATP-driven Ca2+-ATPases in both the plasma membrane and in the internal membrane systems. The cellular system for maintaining Ca2+-homeostasis, and other types of homeostasis as well, a most important issue in cellular physiology and evolution, is complex. It is in this context that the self-generated inorganic ion-based cellular electricity and the lipid nature of cell membranes has to be understood. This is well worded by Torday48,49 as “The history of physiologic cellular-molecular interrelationships can be traced all the way back to the unicellular state by following the pathway formed by lipids ubiquitously accommodating calcium homeostasis, and its consequent adaptive effects on oxygen uptake by cells, tissues and organs.”
Lipid membranes are not permeable to inorganic ions unless they harbor proteinaceous ion channels and pumps. They are permeable to electrons, which means that self-generated cellular electricity could not function if it were electron-based. Self-generated inorganic ion-based electricity is vital to life. A cell is dead when its electrical dimension collapses.20,50 An overlooked key feature of cells is that all cells are able to drive an electrogenic electrical current through themselves, at least during part of their developmental cycle, and that they are polarized, meaning that cells are almost never spherically symmetric (for figures, see ref. 19). This is contained in “The cell as a miniature electrophoresis chamber concept.”34
The universal “driving force” of evolution?
Not natural selection, but problem-solving activity preceding selection is the universal driving force of evolution
Neo-Darwinists hearing somebody contesting the generally accepted view that Darwin's Natural Selection is the universal driving force of evolution probably experience this as cursing in a cathedral. Yet, Metadarwinists may consent (see website: The third way of evolution51). The problem for both is that at present it remains difficult to clearly define the mode of action of Natural Selection and to present examples where it has been at work. If selection would nevertheless not be the driving force, what is the alternative? I argue that if one changes paradigm away from the New Synthesis by beginning with first defining “Life” and next looks at what mechanisms may be instrumental to its variability, problem-solving activity preceding selection emerges as the long-sought for alternative.
In a former paper,19 I used the example of students doing an exam to illustrate this principle. The general perception is that the examiner, not the students taking the exam does the selection. Yet, if one analyses the system, the opposite conclusion emerges. The teacher-examiner formulates the questions. In evolutionary wording, he/she constructs some gradient, like nature would build temperature-, light- etc. gradients. It is up to the students to show their ability to overcome the exam-gradient. Thus, they engage in self-selection, a principle advanced as Gradient-Provoked Swelling/Shrinking Self-Selection or GP-Triple S Principle.3 The examiner only lists their success or failure. The principle of self-selection is further strengthened when the student succeeds in solving the problem by feedback, i.e. by answering in such a way that the sender/teacher will (deliberately or not) lower the gradient (e.g., by changing the subject of examination).
But problem-solving activity is inherent to communication activity which itself is a synonym for Life (as an activity). This leads to the unexpected and counterintuitive conclusion that Life itself is the driving force of its own evolution. In other words, the principle of Life being an activity of compartments that are invariably organized in sender-receiver entities contains the endogenous mechanism for driving its own evolution. In my opinion, this is a splendid principle.
Cultural evolution is evolution “the software way”
Neo-Darwinism did not yet succeed in plausibly incorporating cultural evolution into the mainstream of evolutionary theory.23 The main reason is that the New Synthesis reduces all causes of variability under the common denominator of genetic changes (Fig. 4).
As long as one assumes that the principles of the cognitive memory are inherent to those of DNA → RNA → Proteins, this assumption is the only possible one. But the assumption is wrong. The cognitive memory system has its own rules and mechanisms which include self-generated electrical activity.20 This activity is based on the transport of inorganic ions, and thus only partially dependent upon the central dogma of molecular biology.38 Cultural evolution is mainly achieved through the possibilities of the cognitive memory. In digital era wording, it is evolution ‘the software way’ while organic evolution is evolution ‘the hardware way’, using the principles of the genetic memory.23 In fact, organic- and cultural evolution are the 2 sides of the very same coin, which is evolution of Life with its 2 memory systems. As stated before, cultural evolution is achieved through pupils who function as the software progeny counterpart of physical children.
Differentiation, Evo-devo, Haeckel's “ontogeny recapitulates phylogeny”
The key issue in development is that all cells of a differentiated organism have the same genome, a few exceptions like e.g., the random inactivation of one of the X chromosomes in female mammals not taken into account.52,53 This follows from the fact that the type of cell division during differentiation is mitosis, not meiosis.
If a universal mechanism for guiding development in multicellular organisms exists, it may read as follows: “Keep, during the successive mitotic divisions, the genome constant, but change over and over again its inorganic ion and/or macromolecular environment.”54,55 Changes in inorganic ionic environment result from the fact that all cells of a differentiated organism differ in their plasma membrane-cytoskeletal properties. The inorganic ionic environment around the genes is causal to coarse regulation of control of gene expression, e.g., by its effect on chromatin structure. Transcription factors are better suited for the fine tuning of gene expression.
Although the cited principle looks logical and simple, the generation of differences in membrane properties requires a sequence of events in which polarity, asymmetric cell division and other mechanisms play key roles. This sequence has been summarized in the double asymmetry principle as outlined (with illustrations) in refs. 3, 19, 54 (Fig. 5).
Figure 5.
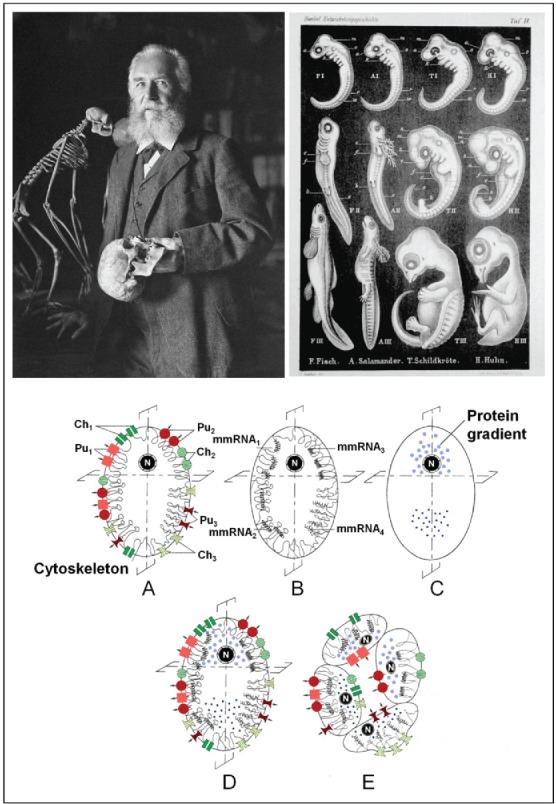
Development and evolution. Ernst Haeckel's ‘biogenetic law’56 “Ontogeny recapitulates phylogeny,” although not perfect, strongly influenced the thinking about the relationship between development and evolution (pictures from Wikipedia, with thanks). It took until the second half of the 20th century before the molecular mechanisms underlying differentiation and development were gradually unraveled. The lower part of the figure illustrates the essence of the ‘double asymmetry principle’3,54,55 as instrumental to governing differentiation during development. It reads: “Keep during the successive mitotic divisions the genome constant (thus no mutations), but change again and again the inorganic ionic as well as the macromolecular environment around the genes. Figures A-E illustrate the major mechanisms instrumental to generating functional asymmetry in the stem cell (zygote) of a hypothetical 4-celled, epithelially organized organism (animal) (A-E).19 The black color of all nuclei is meant to indicate that the genome does not change during differentiation. It is differentially used. First, an asymmetric distribution is realized (i) of the plasma membrane-cytoskeletal complex (A: Pu1-Pu3 = 3 types of ion pumps; Ch1-Ch3 = 3 types of ion channels); (ii) and/or of maternal mRNAs (B: mmRNA1-mmRNA4, some of which can be anchored to the cytoskeleton; (iii) and/or of gradients of certain proteins in the cytoplasm/yolk (C) (e.g., of the bicoid and nanos gradients in eggs of Drosophila). Next the egg is cleaved in an asymmetric way. Sometimes this happens already during the first cleavage, but never later than during the third. In D, which represents the superposition of A, B, and C, it is the second cleavage which will give rise to 4 different cell types, all sharing the same genome (E). This part of the figure legend was copied from De Loof.19
Ernst Haeckel's law,56 Ontogeny Recapitulates Phylogeny (Fig. 5) has continued to receive criticism over the nearly 150 y since its proposal by Haeckel. In my opinion, it remains a valid and appealing key concept in evolutionary theory. It strongly influenced my way of thinking on development and evolution.
Discussion
The Nature paper of Laland et al.57 shows that among evolutionary biologists the conviction is gaining ground that the neo-Darwinian New Synthesis needs an upgrade, but unanimity on this opinion has not yet been reached. Whether one is pro or contra, an upgrade may be influenced by one's major study object(s). If one focuses on sessile organisms like e.g., plants, one may be inclined to assume that the NS explains well enough the mechanisms of evolution, some details not taken into account. If one focuses more on free living organisms like e.g., animals, one may favor the view that some systems partially direct their own evolution,58 and that therefore an upgrade is urgently needed. Free living organisms benefit more from adaptations in mobility and from the possibilities offered by the cognitive memory system for elaborating strategies for improving their survival and reproductive success. Another cause of pro-contra thinking may concern the type of evolution one is interested in. The humanities are primarily interested in cultural evolution of the Homo sapiens. The exact biologic sciences do not consider the Homo sapiens as a special case for which another type of evolution needs to be invoked, but as one of the numerous terrestrial species. They are more interested in the organic-chemical evolution of this and other species, no matter whether they live in an aquatic or terrestrial habitat.
From the concept that Life is based upon 4 pillars, namely hardware, software, energy and motivation, in combination with the fact that all 4 are subject to change in many ways, it follows that genetic- and epigenetic changes are only one element of possible change, be it a very important one.
If one agrees that the numerous novel insights generated by the novel disciplines in biology19,34,48-49,59-65 need to be incorporated in an Extended Evolutionary Synthesis (EES), one faces the question how such integration and unification can be achieved. The importance of communication for understanding Life and its evolution has been approached in various ways.3,23,43 In my opinion, the most straightforward approach is to start from a plausible definition of Life that is acceptable to both the humanities and the exact biologic sciences. This represents a truly novel change in paradigm. Communication activity executed by sender-receiver compartments is the key issue in such definition.16 It leads to the question how the architecture and functioning evolved from the probably simple Progenote as the primordial sender-receiver into the multitude of organismal and supra-organismal entities that function as sender-receivers.
Neo-Darwinists and Metadarwinists both agree on the common descent principle, the very heart of Darwinism. They differ in opinion(s) on several topics (see website: Thethirdwayofevolution51), e.g., on the relative importance of epigenetics, on the weight one should give to the overall importance of genetic changes as instrumental to bringing about (all) evolutionary change as well as on the significance of Natural Selection as the universal driving force of evolution. NS primarily focuses on the effects of all kinds of mutations (Fig. 4) and on species formation through the possibilities of only one memory system, namely the genetic memory and the central dogma of molecular biology DNA → RNA → Proteins. This is apparent from the formulation of ‘the inclusive inheritance principle’40 that acknowledges that in addition to all sorts of mutations, there are indeed other causes of variability instrumental to evolution. But in the end their effects can all be explained by one memory system, the genetic one. But cells/organisms have in addition to their DNA memory, a cognitive memory system. Although it continues to be a (biochemical) black box, there is no reason to neglect its existence and importance. Darwin did not know the principles of the genetic memory but he took them into account. As a result, NS fails to adequately incorporate cultural evolution into the mainstream of evolutionary theory. This type of evolution relies more on the cognitive memory system.
As long as NS does not accept a software upgrade, it will remain a theory of the evolution of the hardware of living matter as governed by the principles of genetics. Such type of evolution is very slow. It usually (but not always) operates at the geological time scale. Mega-evolution takes 2 memory systems into account. Through teaching and learning which are mainly enabled by the cognitive memory, evolution by non-genetic mechanisms (which is ‘evolution the software way’ in my approach) can be very fast as illustrated by the recent evolution of the species Homo sapiens. Another example is the coming into existence of a new Darwin finch species on the Galápagos island Daphne Major that took only 4–5 generations, starting in 1981.66 These data illustrate the power of the introduction of a dialect in a language as instrument for reproductive isolation, an important issue in species formation.
Some people may not like the idea that our body is in fact a clump of some 100,000 billion ( = 1011) eukaryotic cells of which each is the symbiotic result of a few (3?) ancient Archaea/bacteria. Mitochondria are modified bacteria. Each eukaryotic cell contains several mitochondria. In addition, numerous bacteria live on the outer surface (skin) and in the alimentary canal of animals. All these subcellular and cellular entities have to cooperate which means that the communication networks (signaling pathways) inside any multicellular organism are numerous. The complexity can be orders of magnitude higher in populations, communities etc. In a recent (2015) internet discussion forum that followed publication of a paper by Shanta,32 Kalevi Kull posted the quote that “Life is semiosis. Life is a network of sign processes and that this is obviously the most exact and brief definition of life.”
I advocate replacing the widely accepted concept that “Natural Selection is the driving force of evolution” by: “Problem-solving activity preceding selection (like when doing an exam) is that universal force.” One could argue that in the end it does not make much difference: the best adapted (which are not necessarily the strongest ones) will do better. Yet, the formulation does make a substantial difference because it necessitates answering the question which biologic principle enables problem-solving. The answer is that problem-solving does neither follow in full from the central dogma of molecular biology38 nor from the fact that all living matter is cellularly organized. It is inherent to the organization of all living matter in senders-receivers that continuously handle information, thereby solving problems, most of them in an automated way. This approach necessitates that one rethinks several aspects of evolutionary theory. For example, should one continue to attribute so much weight to species formation? Or, what is the unit of selection: the cell, the organism, the species etc. or the signaling pathway as instrumental to problem-solving or the sender-receiver compartment? How to better incorporate the principles of physiology in evolutionary theory?19,61,67
Because of the multitude of signaling pathways and their endless interactions the scope of Metadarwinism (in particular the (Metadarwinian) Extended Evolutionary Synthesis or (M)EES-approach as explained by19 and in this paper is much broader than that of the New Synthesis. MEES is better rooted in physiology, a weak point of NS. Instead of heredity, MEES prefers “transferability of memory system, the cognitive memory. Sharing electrical activities at cell division, and sharing part of the cytoskeleton with its inherent polarity, is also a means of transfer of information to the next generation(s) (and where relevant, laterally as well) by all means”, thus also by the possibilities of a second memory system different from the DNA-system. This way the seamless integration of both organic- and cultural evolution can be realized.19
The Mega-evolution approach urges for changes in teaching biology. For the moment textbooks of biology seldom explain the principles of communication, probably because the authors assume that these principles are self-evident (because we communicate all the time without any problem), and that therefore they do not need extensive explanation. In reality, the opposite is true. The fact that most communication happens in an automated way indicates that it is far from simple. How could it become automated? Upon analysis, it becomes clear that the mechanisms of communication are at least as sophisticated as those of genetics. In particular, as cited before, the role of the cognitive memory in signaling is still a black box, despite all progress in neurobiology.39
Conclusions
In the recent past I repeatedly stated that, paraphrasing Theodosius Dobzhansky,5 “Nothing in biology and evolutionary theory makes sense except in the light of the ability of living matter to communicate, and by doing so, to solve problems.” Given its continuing observational and descriptive nature the discipline of Biology keeps missing a unifying principle comparable to E = mC2 for physics or the atomic model for chemistry. John Torday48-49 summarized this with the characterization by Earnest Rutherford as ‘stamp collecting’. In my opinion, if properly incorporated in teaching L = ∑C harbors the potential for shedding the (not fully mistaken) perception that many biologists insufficiently grasp in full the very nature and importance of the principle that can integrate all subdisciplines of Biology, namely communication. Maybe, the following one-liner may incite students to realize that evolution is much more than the story of changing genes/proteins:
“Living entities: self-electrifying and talking aggregates of fossil stardust operating in an environment heavily polluted by toxic calcium”.19
Disclosure of potential conflicts of interest
No potential conflicts of interest were disclosed.
Acknowledgments
I thank all students and colleagues who helped me to streamline my communication-based view of evolution. I also thank the editors and referees for their comments and suggestions that served as the foundations for the present paper. My thanks too to Julie Puttemans, Marijke Christiaens and son Seppe Hoste, to Katrien Becuwe for help with the figures, and to Michael Gaffney for text correction.
References
- [1].Darwin C. On the origin of species by means of natural selection, or the preservation of favored races in the struggle for life. London: Murray; 1859 [Google Scholar]
- [2].Wallace AR. The theory of Natural Selection. A series of Essays. New York: MacMillan and Co; 1871 [Google Scholar]
- [3].De Loof A. Communication, Life, Mega-Evolution Decrypting Life's Nature. Leuven, Belgium: Leuven University Press; 2002 [Google Scholar]
- [4].Villareal LP. Viruses and the evolution of Life. Washington: ASM Press; 2005 [Google Scholar]
- [5].Dobzhansky T. Nothing in biology makes sense except in the light of evolution. Am Biol Teach 1973; 35:125-9; https://doi.org/ 10.2307/4444260 [DOI] [Google Scholar]
- [6].Stebbins G, Ayala FJ. Is a new evolutionary synthesis necessary? Science 1981; 213:967-71; PMID:17789015; https://doi.org/ 10.1126/science.213.4511.967 [DOI] [PubMed] [Google Scholar]
- [7].Mayr E. What was the evolutionary synthesis? Trends Ecol Evol 1993; 8:31-3; PMID:21236096; https://doi.org/ 10.1016/0169-5347(93)90128-C [DOI] [PubMed] [Google Scholar]
- [8].Kauffmann SA. The Origins of Order. Self-organization and Selection in Evolution. Oxford: Oxford University Press; 1993 [Google Scholar]
- [9].Mayr E, Provine WB, editors. The evolutionary synthesis: Perspectives on the unification of Biology. Cambridge: Harvard University Press; 1998 [Google Scholar]
- [10].Gould SJ. The Structure of Evolutionary Theory. Cambridge: The Belknap Press of Harvard University Press; 2002 [Google Scholar]
- [11].Bowler P. The Eclipse of Darwinism: anti-Darwinian evolutionary theories in the decades around 1900. Baltimore: John Hopkins University Press; 1983 [Google Scholar]
- [12].Bowler P. Evolution: The history of an idea. Oakland: University of California Press; 2003 [Google Scholar]
- [13].Kutschera U, Niklas KJ. The modern theory of biological evolution: an expanded synthesis. Naturwissenschaften 2004; 91(6):255-76; https://doi.org/ 10.1007/s00114-0515-y; PMID:15241603 [DOI] [PubMed] [Google Scholar]
- [14].Huxley J. Evolution: The modern synthesis. London: Allen and Urwin; 1942 [Google Scholar]
- [15].Raven P, Johnson G, Mason K, Locos J, Singer S. Biology. Columbus: McGraw Hill Education; 2013 [Google Scholar]
- [16].De Loof A. How to deduce and teach the logical and unambiguous answer, namely L = ∑C, to “What is Life?” using the principles of communication? Communicative Integrative Biol 2015; 8(5):e1059977; https://doi.org/ 10.1080/19420889.2015.1059977 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [17].Schejter A, Agassi J. On the definition of life. J. Gen. Phil Sci 1994; 25:97-106; https://doi.org/ 10.1007/BF00769279 [DOI] [Google Scholar]
- [18].Koshland DE. The seven pillars of life. Science 2002; 292:2215-6; https://doi.org/ 10.1126/science.1068489 [DOI] [PubMed] [Google Scholar]
- [19].De Loof A. From Darwin's On the Origin of Species by Means of Natural Selection. to The evolution of Life with Communication Activity as its very essence and driving force (= Mega-Evolution). Life Exc Biol 2015; 3(3):153-87; https://doi.org/ 10.9784/LEB3(3)DeLoof.01 [DOI] [Google Scholar]
- [20].De Loof A. The cell's self-generated “electrome:” the biophysical essence of the immaterial dimension of Life? Communicative Integrative Biol 2016; 9(6):e1197446; https://doi.org/ 10.1080/19420889.2016.1197446 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [21].De Loof A, Vanden Broeck J. Communication: the key to defining “life,” “death” and the force driving evolution. “Organic chemistry-based-” versus “artificial” life.” Belg J Zool 1996; 125:5-28 [Google Scholar]
- [22].De Loof A. Schrödinger 50 years ago: “What is Life?” “The ability to communicate,” a plausible reply? Int J Biochem 1993; 25:1715-21; https://doi.org/ 10.1016/0020-711X(88)90298-4 [DOI] [Google Scholar]
- [23].De Loof A. Organic and cultural evolution can be seamlessly integrated using the principles of communication and problem-solving: The foundations for an Extended Evolutionary Synthesis (EES) as outlined in the Mega-Evolution concept. Life Exc Biol 2015; 2(4):247-69; https://doi.org/ 10.9784/LEB2(4)DeLoof.01 [DOI] [Google Scholar]
- [24].Guth JH. Pre-Biotic Evolution: I. From Stellar to Molecular Evolution. Society of the Advancement of Metadarwinism 2014; 1 https://metadarwinismsociety./wordpress.com/2014/10/31/pre-biotic-evolution-from-stellar-to-molecular-evolution/ [Google Scholar]
- [25].Guth JH. Pre-Biotic Evolution: II. Pre-Biotic Chemical Oscillations and Linked Reaction Sequences. Society of the Advancement of Metadarwinism 2015; 2 https://metadarwinismsociety./wordpress.com/2015/10/31/pre-biotic-evolution-from-stellar-to-molecular-evolution/ [Google Scholar]
- [26].Ruggiero MA, Gordon DP, Orrell TM, Bailly N, Bourgoin T, Brusca RC, Cavalier-Smith T, Guiry MD, Kirk PM. 2015 A higher level classification of all living organisms. PLoS One 2015; 10(4):e0119248; https://doi.org/ 10.1371/journal.pone.0119248 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [27].Margulis L. Symbiosis in Cell Evolution; New York: W. H. Freeman and Company; 1981 [Google Scholar]
- [28].Margulis L, Sagan D. Acquiring Genomes: A Theory of the Origins of Species. New York: Basic Books; 2002 [Google Scholar]
- [29].Witzany G. Biocommunication and Natural Genome Editing. Dordrecht: Springer; 2010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [30].Trewavas AJ, Baluška F. The ubiquity of consciousness, cognition and intelligence in life. EMBO Rep 2011; 12(12):1221-5; PMID:22094270; https://doi.org/ 10.1038/embor.2011.218 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [31].Baluška F, Yokawa K, Marcuso S, Baverstock K. Understanding of anaesthesia – Why consciousness is essential for life and not based upon genes. Communicative Integrative Biol 2016; 9(6):e1238118; https://doi.org/ 10.1080/19420889.2016.1238118 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [32].Shanta BN. Life and Consciousness – The Vêdantic View. Communicative Integrative Biol 2015; 8(5):e1085138:1-11. 24; https://doi.org/ 10.1080/19420889.2015.1085138 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [33].Caetano-Anollés G. Creationism and intelligent design are incompatible with scientific progress: A response to Shanta and Vêdanta. Communicative Integrative Biol 2016; 9(1):e1123356; https://doi.org/ 10.1080/19420889.2015.1123356 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [34].De Loof A. The electrical dimension of cells: the cell as a miniature electrophoresis chamber. Int Rev Cytol 1986; 104:251-352; PMID:3531065 [DOI] [PubMed] [Google Scholar]
- [35].De Loof A, De Haes W, Janssen T, Schoofs L. The essence of insect metamorphosis and aging: electrical rewiring of cells driven by the principles of juvenile hormone-dependent Ca(2+)-homeostasis. Gen Comp Endocr 2014; 199:70-85; PMID:24480635; https://doi.org/ 10.1016/j.ygcen.2014.01.009 [DOI] [PubMed] [Google Scholar]
- [36].Jaffe LF, Nuccitelli R. An ultrasensitive vibrating probe for measuring steady ex[tracellular electric currents. J. Cell Biol 1974; 63:614-28; PMID:4421919; https://doi.org/ 10.1083/jcb.63.2.614 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [37].Woodruff RI, Telfer WH. Electrophoresis of proteins in intercellular bridges. Nature 1980; 286:84-6; PMID:7393329; https://doi.org/ 10.1038/286084a0 [DOI] [PubMed] [Google Scholar]
- [38].Crick F. Central dogma of molecular biology. Nature 1970; 227:561-3; PMID:4913914; https://doi.org/ 10.1038/227561a0 [DOI] [PubMed] [Google Scholar]
- [39].Peixoto LL, Wimmer ME, Poplawski SG, Tudor JC, Kenworthy CA, Liu S, Mizuno K, Garcia BA, Zhang NA, Giese KP, et al.. Memory acquisition and retrieval impact different epigenetic processes that regulate gene expression. BMC Genomics 2015; 16(Suppl 5):S5; PMID:26040834; https://doi.org/ 10.1186/1471-2164-16-S5-S5 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [40].Danchin E, Charmantier A, Champagne FA, Mesoudi A, Pujol B, Blanchet S. Beyond DNA: integrating inclusive inheritance into an extended theory of evolution. Nat Rev Genet 2011; 12:475-86; PMID:21681209; https://doi.org/ 10.1038/nrg3028 [DOI] [PubMed] [Google Scholar]
- [41].Eco U. A Theory of Semiotics. Bloomington: Indiana University Press; 1978 [Google Scholar]
- [42].Wheeler W. The Whole Creature. Complexity Biosemiotics and the evolution of culture. London: Lawrence and Wishart Ltd; 2006 [Google Scholar]
- [43].Kull K, Emmeche C, editors. Towards a Semiotic Biology: Life is the action of Signs. London: Imperial College Press; 2011 [Google Scholar]
- [44].Westling L. The Logos of the Living World: Merleau-Ponty, Animals and Language. New York: Fordham University Press; 2014 [Google Scholar]
- [45].De Loof A. The essence of female-male physiological dimorphism: differential Ca2+-homeostasis enabled by the interplay between farnesol-like endogenous sesquiterpenoids and sex-steroids? The Calcigender paradigm. Gen Comp Endocr 2015; 211:131-46; PMID:25540913; https://doi.org/ 10.1016/j.ygcen.2014.12.003 [DOI] [PubMed] [Google Scholar]
- [46].De Loof A, Schoofs L, Huybrechts R. The endocrine system controlling sexual reproduction in animals: Part of the evolutionary ancient but well conserved immune system? Gen Comp Endocr 2016; 226:56-71; https://doi.org/ 10.1016/j.ygcen.2015.12.016 [DOI] [PubMed] [Google Scholar]
- [47].Orrenius S, Zhihotovsky B, Nicotera P. Regulation of cell death: the calcium-apoptosis link. Nat Rev Cell Biol 2003; 4:552-65; https://doi.org/ 10.1038/nrm1150 [DOI] [PubMed] [Google Scholar]
- [48].Torday JS. A central theory of biology. Med. Hypotheses 2015; 85:49-57 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [49].Torday JS. Homeostasis as the mechanism of evolution. Biology (Basel) 2015; 4:573-90; PMID:26389962 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [50].De Loof A, De Haes W, Boerjan B, Schoofs L. The fading electricity theory of ageing: the missing biophysical principle? Ageing Res Rev 2013; 188:35-45; https://doi.org/ 10.1016/j.arr.2012.08.001 [DOI] [PubMed] [Google Scholar]
- [51].The Third Way Evolution in the Era of Genomics and Epigenomics. www.thethirdwayof evolution.com
- [52].Gilbert SF. Developmental Biology, 10th ed Sunderland: Sinauer Associates Inc; 2013 [Google Scholar]
- [53].Wolpert L, Tickle C, Martinez Arias A. Principles of Development 5th ed Oxford: Oxford University Press; 2015 [Google Scholar]
- [54].De Loof A. Differentiation: Keep the genome constant but change over and over again its ionic and/or macromolecular environment. A conceptual synthesis. Belg J Zool 1993; 123:77-91 [Google Scholar]
- [55].De Loof A, Callaerts P, Vanden Broeck J. The pivotal role of the plasma membrane-cytoskeletal complex and epithelium formation in animals. Comp Biochem Physiol 1992; 101A:639-51; https://doi.org/ 10.1016/0300-9629(92)90338-Q [DOI] [PubMed] [Google Scholar]
- [56].Haeckel E. Natürliche Schöpfungsgeschichte. Berlin: Georg Reimer Verlag; 1868. English version entitled, The History of Creation: Or the Development of the Earth and Its Inhabitants by the Action of Natural Causes. Revised edition by Prof Lancaster E. Ray. 2 vols. London: King; 1876 [Google Scholar]
- [57].Laland K, Uller T, Feldman M, Sterelny K, Müller GB, Moczek A, Jablonka E, Odling-Smee J, Wray GA, Hoekstra HE, et al.. Does evolutionary theory need a rethink? Nature 2014; 514:161-4; PMID:25297418; https://doi.org/ 10.1038/514161a [DOI] [PubMed] [Google Scholar]
- [58].Pookottil R. B.E.E.M: Biological Emergence-based Evolutionary Mechanism: How species direct their own evolution. London: Fossil Fish Publishing; 2013. Distributed through Troubador Publishing Ltd: Wistow, Kibworth, Leicester [Google Scholar]
- [59].Pigliucci M. Do we need an extended evolutionary theory? Evolution 2007; 61:2743-9; PMID:17924956; https://doi.org/ 10.1111/j.1558-5646.2007.00246.x [DOI] [PubMed] [Google Scholar]
- [60].Noble D. The Music of Life: Biology Beyond the Genome. Oxford: Oxford University Press; 2008 [Google Scholar]
- [61].Noble D. Physiology is rocking the foundations of evolutionary biology. Exp Physiol 2013; 98:1235-43; PMID:23585325; https://doi.org/ 10.1113/expphysiol.2012.071134 [DOI] [PubMed] [Google Scholar]
- [62].Noble D. Evolution beyond Neo-Darwinism: a new conceptual framework. J Exp Biol 2015; 218(Pt1):7-13; PMID:25568446; https://doi.org/ 10.1242/jeb.106310 [DOI] [PubMed] [Google Scholar]
- [63].Shapiro JA. Evolution – A View from the 21th Century. Upper Saddle River: FT Press Science; 2011 [Google Scholar]
- [64].Shapiro JA. Rethinking the (im)possible in evolution. Prog Biophys Mol Biol 2012; 111:92-6; PMID:22986107; https://doi.org/ 10.1016/j.pbiomolbio.2012.08.016 [DOI] [PubMed] [Google Scholar]
- [65].Pigliucci M, Müller GB, editors. Evolution: The Extended Synthesis. MIT Press, Cambridge: 2011 [Google Scholar]
- [66].Grant PR, Grant BR. The second contact phase of allopatric speciation in Darwin's finches. Proc Natl Acad Sci USA 2009; 106:20141-8; PMID:19918081; https://doi.org/ 10.1073/pnas.0911761106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- [67].Danchin E, Pocheville A. Inheritance is where physiology meets evolution. J Physiol 2014; 532:2307-17; https://doi.org/ 10.1113/jphysiol.2014.272096 [DOI] [PMC free article] [PubMed] [Google Scholar]