

Abstract
The purpose of this study was to investigate whether a tethered-swimming incremental test comprising small increases in resistive force applied every 60 seconds could delineate the isocapnic region during rapidly-incremented exercise. Sixteen competitive swimmers (male, n = 11; female, n = 5) performed: (a) a test to determine highest force during 30 seconds of all-out tethered swimming (Favg) and the ΔF, which represented the difference between Favg and the force required to maintain body alignment (Fbase), and (b) an incremental test beginning with 60 seconds of tethered swimming against a load that exceeded Fbase by 30% of ΔF followed by increments of 5% of ΔF every 60 seconds. This incremental test was continued until the limit of tolerance with pulmonary gas exchange (rates of oxygen uptake and carbon dioxide production) and ventilatory (rate of minute ventilation) data collected breath by breath. These data were subsequently analyzed to determine whether two breakpoints defining the isocapnic region (i.e., gas exchange threshold and respiratory compensation point) were present. We also determined the peak rate of O2 uptake and exercise economy during the incremental test. The gas exchange threshold and respiratory compensation point were observed for each test such that the associated metabolic rates, which bound the heavy-intensity domain during constant-work-rate exercise, could be determined. Significant correlations (Spearman’s) were observed for exercise economy along with (a) peak rate of oxygen uptake (ρ = .562; p < 0.025), and (b) metabolic rate at gas exchange threshold (ρ = −.759; p < 0.005). A rapidly-incremented tethered-swimming test allows for determination of the metabolic rates that define zones for domain-specific constant-work-rate training.
Key words: isocapnic region, gas exchange threshold, respiratory compensation point, exercise economy, constant-work-rate exercise, heavy intensity
Introduction
In both theory and practice, it is accepted that endurance training for athletes should comprise time spent in different exercise-intensity zones that are based on definable physiological responses (Midgley et al., 2007). For example, a typical week for an endurance athlete might include both easy training in the moderate-intensity domain where a metabolic steady state is achieved rapidly and steady training in the heavy domain where a steady state is attainable, although delayed (Jones and DiMenna, 2011). Furthermore, training at intensities where a steady state is unattainable (e.g., tempo training in the severe domain and/or interval training with work intervals in the extreme domain) might also comprise part of the athlete’s weekly load (Jones and DiMenna, 2011). It is, therefore, important to quantify the metabolic rates that separate athletes’ exercise-intensity domains so that domain-specific training can be prescribed.
In 1981, Whipp et al. developed an incremental cycle-ergometer test where the work rate was applied as a smooth function of time (i.e., as a ramp; e.g., one watt every two seconds) (Whipp et al., 1981). Unlike slowly-incremented tests (e.g., traditional step tests with prolonged stages), this test allowed for clear delineation of the isocapnic region that separated two thresholds that could be identified from gas exchange and ventilatory data during incremental exercise (referred to in this article as the gas exchange threshold and respiratory compensation point; GET and RCP, respectively). Furthermore, small increments in the work rate that were continuously applied allowed for precise determination of the metabolic rates (i.e., rates of oxygen uptake) at each of these breakpoints (i.e., O2GET and O2RCP). These increased aspects of sensitivity are important because O2GET and O2RCP are aligned with the metabolic rates that serve as lower and upper boundaries for the heavy-intensity domain during constant-work-rate exercise (Jones and Poole, 2005; Keir et al., 2015; Stanula et al., 2014; Whipp and Wasserman, 1972). Consequently, a rapidly-incremented test with small work-rate increases is useful for prescribing domain-specific training and it is, therefore, not surprising that variations of this cycling protocol are used in the athletic, clinical and research setting. However, athletes must be tested in their specific mode of exercise and this is particularly the case for swimmers (Pinna et al., 2013). Hence, it is important to develop rapidly-incremented protocols that allow for precise determination of O2GET and O2RCP to prescribe endurance swim training.
Incremental tests that are used to assess cardiorespiratory capacity during free swimming typically involve intervals of set distance (e.g., 200 m) performed at progressively-increasing velocities (Fernandes et al., 2003, 2011; Ribeiro et al., 2015). This means that unlike the smooth-ramp cycling test described above, these tests consist of lengthy stages comprising large unequal work-rate increments. It is, therefore, not surprising that researchers typically use these tests to identify only one of the aforementioned thresholds (e.g., often referred to as the anaerobic threshold) (Fernandes et al., 2011; Ribeiro et al., 2015). Indeed, due to its very nature, it is unlikely that this type of testing can identify the isocapnic region to derive the three-phase model that is best suited for exercise prescription (Binder et al., 2008; Skinner and McLellan, 1980). However, it is difficult to envision how rapidly-incremented small symmetrical changes in the work rate can be applied during free swimming because control of pace is imprecise. One alternative is to perform stationary swimming against a resistive load that can be increased with greater precision (i.e., tethered swimming). Research has confirmed that the maximal rate of oxygen uptake (O2max) derived from an incremental tethered-swimming test is highly correlated with and not significantly different from that which is measured during free swimming (Bonen et al., 1980). However, in that study, no attempt was made to discern the threshold(s) that was/were encountered as the resistive load was increased and a discontinuous protocol was used with work bouts of 2-4 minutes separated by recovery intervals of ≥ 5 minutes of rest (Bonen et al., 1980). Consequently, it is unlikely that this test would have been sensitive to threshold differentiation. Nevertheless, the ability of the tethered methodology to serve as a swim ergometer raises the intriguing possibility that a rapidly-incremented protocol similar to that which is used for stationary cycling could be developed using this approach.
In addition to its ability to discriminate the phases in which bicarbonate buffering is/is not sufficient to maintain homeostatic blood pH during incremental exercise, the smooth-ramp rapidly-incremented cycling test allows for determination of O2max if the test is continued until the limit of tolerance (Bogaard et al., 2008; Whipp et al., 1981; Whipp and Wasserman, 1972). The rapidly-incremented nature of this test is also important in this regard because longer incremental protocols (e.g., ≥ 12 minutes) can result in underestimation of O2max (Astorino et al., 2004; Yoon et al., 2007). This has resonance for swimming because the incremental protocol that is typically employed (Fernandes et al., 2003, 2011; Ribeiro et al., 2015) comprises 7-8 steps and, therefore, overall test duration can exceed 12 minutes. Despite its non-steady-state nature, the smooth-ramp test also reveals a O2/work-rate slope that serves as a measure of exercise economy (Whipp et al., 1981). It is generally believed that improving O2max, O2GET/ O2RCP and exercise economy might each require different training strategies (Jones and Carter, 2000); hence, the information provided by this singular test can be used to tailor athletic training to the characteristics of a given athlete and to monitor specific training-induced changes.
The purpose of the present study was to investigate whether a rapidly-incremented tethered-swimming protocol with small increases in resistive force could be used to delineate the isocapnic region that separates O2GET and O2RCP. For this reason, we had competitive swimmers of both sexes perform a novel incremental tethered-swimming test comprising work-rate increments equivalent to 5% of the difference between the maximal resistive force against which they could swim and the force required to maintain body alignment applied every 60 seconds. We hypothesized that two distinct breakpoints in gas-exchange/ventilatory kinetics would be identifiable such that O2GET and O2RCP could be determined. We also measured O2peak and the O2/load slope to explore relationships that might be present between exercise economy and the other variables of aerobic fitness.
Material and Methods
Participants
Eleven male (mean ± SD: age, 18 ± 4 yr; stature, 1.80 ± 0.07 m; body mass, 72 ± 10 kg) and five female (age, 17 ± 4 yr; stature, 1.66 ± 0.06 m; body mass, 61 ± 10 kg) swimmers volunteered to participate in this study. The swimmers were competitive at the regional/national level and had each accumulated at least three years of competition training. The subjects were required to give their written informed consent prior to initiation of testing after the experimental procedures, associated risks and potential benefits of participation had been explained. For subjects under the age of 18 (n = 13; range, 14-17 years), signed consent from a parent or guardian was also obtained. This study was approved by the São Paulo State University ethics committee. The subjects were instructed to: 1.) avoid strenuous exercise in the 24 hours preceding each testing session; and 2.) arrive at the pool in a rested and fully-hydrated state at least three hours postprandial. Subjects were also asked to refrain from stimulant beverages and alcohol for 24 hours prior to each test.
Procedures
The competitive swimmers who agreed to participate reported to the pool for testing on two different occasions separated by 48 hours. During the first visit, subjects were familiarized with tethered swimming after which they performed an all-out tethered-swim test. Results from this test were used to calculate the resistive forces that would be applied during the incremental test, which was performed during the second visit. Both tests were undertaken at the same time of day for a given subject in a semi-Olympic swimming pool with water temperature of ~28º C.
The all-out tethered-swim test was performed with a 4905-N load cell attached to the swimmer’s hip by an inelastic rope. For this test, subjects swam all-out for 30 seconds using a full front-crawl stroke with the averaged peaks of the wave frequency from the force-time signal defined as the trial’s average force. Subjects performed this test twice separated by 20 minutes of rest and the higher value for average force (Favg) was recorded for further analysis. The load cell was calibrated for 100 Hz signal acquisition prior to each test and the acquired signal was smoothed by the manufacturer’s software package (N2000PRO, Cefise). We then determined the difference between Favg and the force that was required to maintain the swimmer’s body alignment prior to initiation of the all-out swim (i.e., baseline force production; Fbase) to derive ΔF. The Fbase and ΔF for each subject were used to calculate the starting resistive force and the increments in resistive force that would be applied during the incremental test.
During the second visit to the pool, subjects completed a maximal incremental tethered-swim test to determine whether the isocapnic region during rapidly-incremented exercise could be delineated during swimming. For this test, a custom-built weight-bearing pulley-rope system similar to a power rack, but modified for instantaneous weight-plate loading (≥ 0.4-kg increments) was used (Figure 1). Test administrators loaded the weight plates onto the system’s carriage manually after receiving time cues from an associate. As was the case for the all-out tethered-swim test, the rope of the load-application system was attached to the subject’s hip and the subject swam using the front-crawl style. Importantly, attachment of the rope in this manner allowed for the leg kick to be unimpeded while providing a near-horizontal opposing force which resulted in minimal alteration of the standard swimming posture. Subjects were instructed to swim at a sufficient rate to avoid rearward/forward displacement of their body position as load increments were applied during each stage. Stage length was 60 seconds. The initial stage was performed against a load that exceeded Fbase by 30% of ΔF and from that point, each stage comprised a load increment of 5% of ΔF. Two markers on the bottom of the pool provided visual reference points that allowed the swimmers to maintain a relatively-fixed position (e.g., ± 1 m from the desired position) and the test was terminated at the point at which this was no longer possible despite strong verbal encouragement from the testers. Breath-by-breath pulmonary gas-exchange data were collected using a portable metabolic unit designed for cardiopulmonary exercise testing (CPET K4b2; Cosmed, Rome, Italy). For this assessment, subjects breathed through a snorkel apparatus (new AquaTrainer®) that had been validated for pulmonary O2 measurement during swimming (Baldari et al., 2013). Before each test, the unit was calibrated according to the manufacturer’s recommendations. After this procedure prior to attachment of the rope, subjects rested quietly on the pool border for 10 minutes with gas-exchange data collected in order to establish baseline parameters. Breath-by-breath O2 data collected during the baseline and exercise periods were averaged over consecutive nine-second periods after being smoothed by the collection unit’s software. O2peak was defined as the highest three-point rolling average of consecutive nine-second O2 values recorded prior to the limit of tolerance. The final three-point rolling average for each completed 60 s stage was used to determine the O2/load slope via linear regression. When O2 failed to increase by an appreciable amount (determined by visual inspection) for ≥ 2 stages immediately prior to the limit of tolerance, a O2 plateau was considered to be present and datum from that stage was removed from the fit. Attempts to identify both GET and RCP were made by consensus from a panel of independent reviewers experienced at making these determinations from a cluster of measurements. For GET, these included: 1.) the first disproportionate increase in the rate of carbon dioxide production (CO2) from visual inspection of individual plots of CO2 vs. O2; 2.) an increase in the expired rate of ventilation (E)/O2 with no increase in E/CO2; and 3) an increase in end-tidal O2 tension with no fall in end-tidal CO2 tension. For RCP, criteria included: 1.) the first disproportionate increase in E in relation to CO2; and 2.) a fall in end-tidal CO2 tension.
Figure 1.

Depiction of the rapidly-incremented tethered-swimming protocol. Fbase is the opposing force that maintained the body position prior to loading and ΔF is the difference between Fbase and the highest force recorded for the subject.
Statistical Analysis
The O2GET, O2RCP, O2peak and O2/load slope are expressed as group mean ± SD. Spearman’s correlation coefficients were used to assess relationships between the O2/load slope and O2GET, O2RCP and O2peak. In all cases, statistical significance was accepted at p < 0.05.
Results
The Favg from the all-out tethered-swim test in absolute and relative (to body mass) terms for the entire group was 185 ± 41 N and 2.7 ± 0.6 N·kg-1BM, respectively. The Favg was 208 ± 43 N (2.9 ± 0.6 N·kg-1BM) for male subjects and 140 ± 18 N (2.3 ± 0.3 N·kg-1BM) for female subjects. The limit of tolerance during the maximal incremental tethered-swim test occurred during stage 9.1 ± 2.0 (range, 6-14 stages; only one test ≥ 11 stages), which equated to the load of 71 ± 10% of Favg. In absolute terms, the load on the final stage of the incremental test was 9.3 ± 1.5 kg (range, 7.1-13.1 kg) for the entire group and 10.0 ± 1.3 kg (8.1-13.1 kg) and 7.7 ± 0.5 kg (7.1-8.3 kg) for male and female subjects, respectively. In conjunction with the near-horizontal line of pull of the opposition that was applied, these relatively low loads in relation to the subject’s body mass (10-16%) allowed for the standard swimming posture to be maintained with the minimal alteration (e.g., swimmers’ feet were not pulled downward to an excessive degree when they reached the final stage). Four of the 16 incremental tests resulted in a O2 plateau prior to the limit of tolerance. O2peak in absolute and relative (to body mass) terms for the entire group was 3.4 ± 0.6 L·min-1 and 49.4 ± 6.3 ml·kg-1BM·min-1, respectively. O2peak was 3.7 ± 0.4 L·min-1 (51.7 ± 5.2 ml·kg1BM·min-1) for male subjects and 2.7 ± 0.1 L·min-1 (44.4 ± 5.8 ml·kg-1BM·min-1) for female subjects.
All 16 of the maximal incremental swim tests were characterized by an isocapnic region such that O2GET and O2RCP could be identified (see Figure 2). Specifically, the O2GET in absolute and relative (to body mass) terms for the entire group was 2.3 ± 0.4 L·min-1 and 33.6 ± 5.8 ml·kg-1BM·min-1 respectively. The O2GET was 2.5 ± 0.3 L·min-1 (35.3 ± 5.1 ml·kg-1BM·min-1) for male subjects and 1.8 ± 0.1 L·min-1 (29.9 ± 6.0 ml·kg-1BM·min-1) for female subjects. O2RCP in absolute and relative (to body mass) terms for the entire group was 3.0 ± 0.5 L·min-1 and 43.9 ± 5.8 ml·kg-1BM·min-1, respectively O2RCP was 3.3 ± 0.3 L·min-1 (45.8 ± 5.1 ml·kg-1BM·min-1) for male subjects and 2.4 ± 0.2 L·min-1 (39.6 ± 5.6 ml·kg-1BM·min-1) for female subjects. O2GET and O2RCP occurred at 68 ± 8% and 89 ± 4% of O2peak, respectively.
Figure 2.
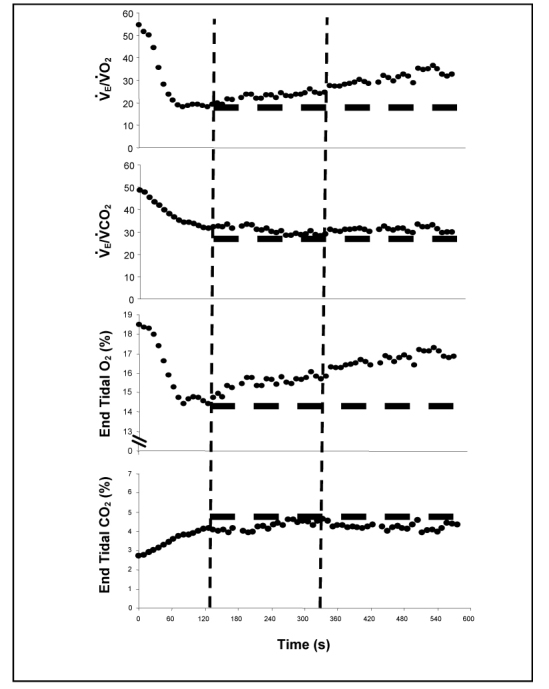
Gas exchange and ventilatory responses for a representative subject during the test. From left to right, vertical dashed lines are aligned with the GET and RCP, respectively. Horizontal dashed lines are positioned at the nadir (top three panels) or apex (bottom panel) of data points.
The O2/load slope in absolute and relative (to body mass) terms for the entire group was 317 ± 102 ml·min-1·kg-1 and 4.7 ± 1.2 ml·kg-1BM·min-1·kg-1, respectively. The O2/load slope was 329 ± 120 ml·min-1·kg-1 (4.6 ± 1.4 ml·kg-1BM·min-1·kg-1) for male subjects and 291 ± 43 ml·min-1·kg-1 (4.8 ± 0.6 ml·kg-1BM·min-1·kg-1) for female subjects. The correlation-coefficient values for the linear fits to the O2/load data ranged from 0.828 to 0.995 (mean ± SD, 0.958 ± 0.042). The O2/load slope in absolute terms was positively correlated with the absolute O2peak (ρ = .562; p < 0.025) and negatively correlated with the percentage of O2peak at which O2GET occurred (ρ = −.759; p < 0.005) (Figure 3; top and bottom panel, respectively). There were no significant correlations between the O2/load slope and the metabolic rate at RCP. When analyzed according to sex, the negative correlation between the O2/load slope and the percent of O2peak at which VO2GET occurred was present in both sexes whereas the positive correlation between O2/load slope and O2peak remained significant only for male subjects.
Figure 3.
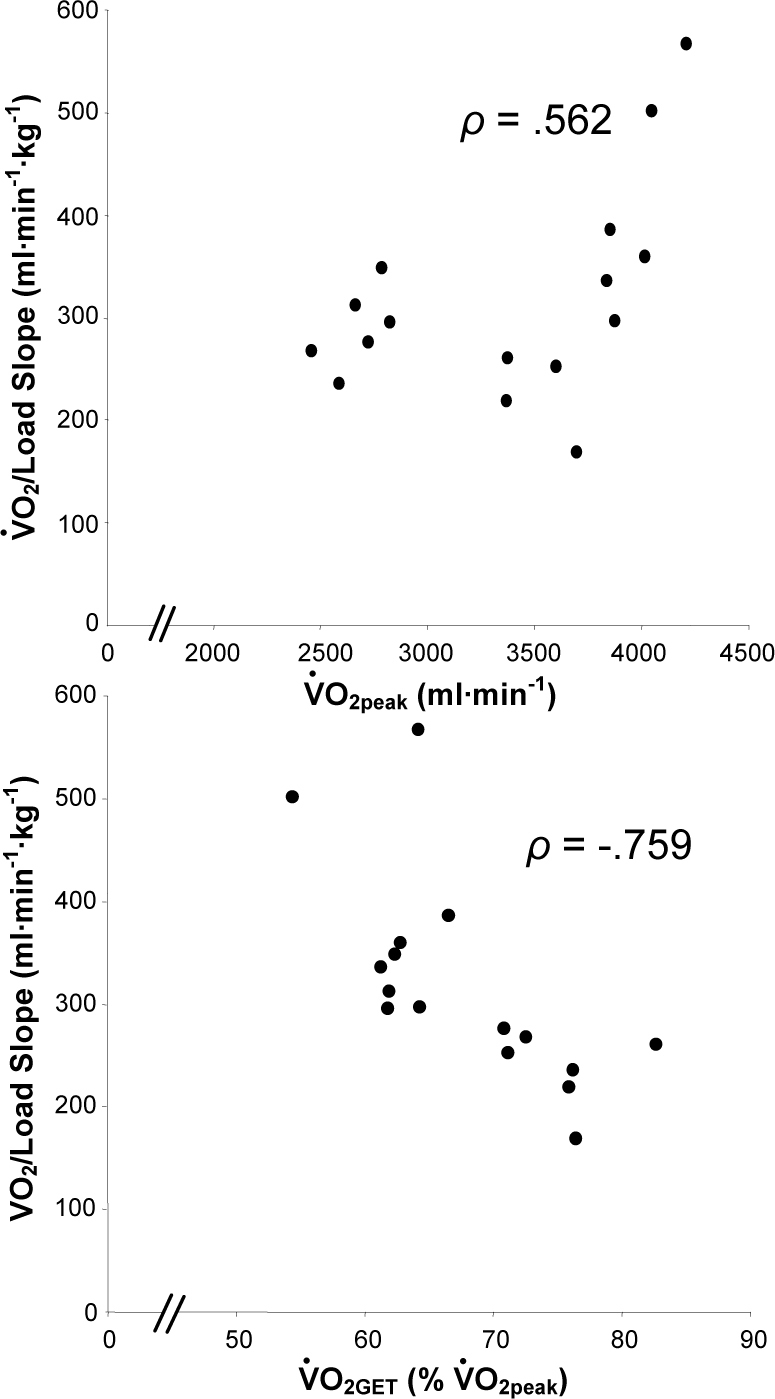
Significant correlations between O2/load slope and the peak rate of O2 uptake (Panel A) and the rate of O2 uptake at the GET relative to peak (Panel B).
Discussion
The main finding from this investigation is that a tethered swimming incremental protocol comprising relatively small work-rate increments (e.g., a resistive load increase of as little as 0.4 kg) applied every 60 seconds was sufficiently rapid to allow delineation of the isocapnic region that can be present during incremental exercise. This means that the two distinct breakpoints in the gas exchange/ventilatory response that can be identified during incremental exercise (referred to in this article as gas exchange threshold and respiratory compensation point, GET and RCP, respectively) were able to be determined from this singular test. We also calculated the O2/load slope as a measure of exercise economy and found that for the competitive swimmers we assessed, this slope was positively correlated with O2peak, but negatively correlated with the metabolic rate at GET expressed relative to O2peak. This implies that the magnitude of O2GET relative to maximal capacity can exert a considerable effect on endurance-swimming performance irrespective of O2max per se.
In the present study, we tested a novel incremental protocol that allowed for determination of O2peak, O2GET, O2RCP and exercise economy during tethered swimming. The test was novel because the increments were applied more rapidly than is typically the case during free swimming incremental protocols. For example, the protocol that is often used involves multiple even-paced 200-m swimming bouts (n × 200m) performed at increasing velocities beginning with an initial step at an easy pace and culminating with a step at maximal effort (Pyne et al., 2001). This type of testing is a modified version of the traditional step-incremental tests that are employed for cycling and running where stage length is fixed (e.g., 3-5 minutes) and the work rate is increased progressively. In addition to allowing for the determination of O2max at or near the limit of tolerance, these slowly-incremented tests were designed to provide information on metabolic responses to submaximal work; for example, the metabolic rate at which a sustained increase in blood lactate concentration above resting levels (≥ ~1 mmol·L-1) is initially observed (the lactate threshold; LT) and the change in external work produced in relation to the change in energy expenditure for completion of a step (delta efficiency). Consequently, extended stage lengths were necessary so that a metabolic steady state could be achieved upon completion of each stage. However, it is now well established that rapid attainment of a steady state (e.g., in ≤ 2 minutes) is only possible for work that is performed below LT with a steady state delayed for up to 15 minutes in what has been termed the heavy-intensity domain or even unattainable when higher work rates are encountered (Jones and Poole, 2005; Poole et al., 1988). Consequently, there is little rationale for prolonging each stage and, indeed, there are drawbacks with such an approach. For example, lengthy stage duration results in a prolonged overall test (e.g., ≥ 12 minutes) that might prevent the attainment of O2max upon exhaustion (Astorino et al., 2004; Yoon et al., 2007). Furthermore, during a slowly-incremented test (e.g., stage duration ≥ 3 minutes), non-invasive identification of LT via gas exchange/ventilatory changes corresponding to the non-metabolic production of CO2 is complicated by the fact that E will increase disproportionately compared to CO2 for all work rates above the LT (Whipp et al., 1989). Consequently, slowly-incremented tests will not allow for identification of the isocapnic region within which arterial partial pressure of CO2 is maintained above LT during incremental exercise (i.e., supra-LT work where compensatory hyperventilation is not yet required).
The range of metabolic rates within the isocapnic region during incremental exercise approximate those that comprise the heavy-intensity domain during exercise performed at a constant rate of work (Jones and Poole, 2005; Keir et al., 2015; Whipp and Wasserman, 1972). This domain is bounded on its upper end by the maximal lactate steady state, which means that heavy-intensity exercise can be sustained for extended periods despite an elevation of blood-lactate concentration above resting levels (Espada et al., 2015; Jones and Poole, 2005). Importantly, it is believed that training within this domain enhances exercise economy and shifts both the LT and lactate turnpoint (i.e., the acceleration in blood lactate accumulation during incremental exercise that typically occurs around 2.5-4.0 mmol·L-1) to higher work rates (Jones and DiMenna, 2011). Defining these metabolic rates (e.g., in the present study, a range that spanned from ~33.6 to ~49.4 ml·kg-1BM·min-1) is, therefore, important because continuous training within the heavy domain would likely make up a good portion of an endurance swimmer’s weekly training load.
Unlike the rapidly-incremented test that we employed, the n × 200m slowly-incremented test provides rest periods between stages that allow for blood to be drawn. This means that estimation of GET and RCP via gas exchange/ventilatory data is not necessary because blood lactate dynamics can be assessed directly (Fernandes et al., 2011). However, this test is typically used to identify a singular anaerobic threshold, a term that is problematic because it has been used to describe both of the thresholds that are defined according to the three-phase model (Binder at al., 2008). Furthermore, Fernandes et al. (2011) used the conventional protocol to identify an individual anaerobic threshold that occurred at a blood lactate concentration of ~2 mmol·L-1. Given this level of blood lactate accumulation, this threshold was likely greater than the LT; hence, the associated metabolic rate would be greater than the one that defines the lower boundary of the heavy-intensity domain (i.e., O2GET). Moreover, they reported that blood lactate concentration at the individual anaerobic threshold was significantly less than that which was present during constant-work-rate swimming at the maximal lactate steady state (~3 mmol·L-1). This means that the associated metabolic rate was less than the critical metabolic rate that serves as the heavy domain’s upper boundary (O2RCP) (Keir et al., 2015). Ribiero et al. (2015) also reported a singular anaerobic threshold for subjects performing the n × 200m protocol regardless of whether a direct (blood lactate concentration) or indirect (gas-exchange/ventilation) measurement was used for the determination.
In addition to O2GET, O2RCP and O2peak, we also calculated the O2/load slope during the rapidly-incremented test as a proxy measure of swimming efficiency. This slope, which represents the inverse of delta efficiency, provides an estimate of exercise economy that can be derived from rapidly-incremented tests that take place entirely in the non-steady state (Whipp et al., 1981). Our results showed considerable variance in this parameter with a range from 2.3 to 7.0 ml·kg-1BM·min-1 per kilogram of the load applied in the 16 swimmers that we tested. However, a consistent feature was that swimmers with a greater oxidative capacity possessed lower exercise economy. Indeed, we observed a positive correlation between O2peak and the O2/load slope (Figure 3; top panel) which, at first glance, appears counterintuitive. However, an inverse relationship between O2max and exercise economy had previously been reported for professional cyclists during cycling (Lucia et al., 2002) and premenopausal women with a wide range of O2max values during graded treadmill walking (Hunter et al., 2005). Furthermore, in the latter study, the researchers used 31P magnetic resonance spectroscopy to assess oxidative capacity and exercise economy on the muscle-tissue level during isometric plantarflexion and much like the whole-body measurement, a similar inverse relationship was found (Hunter et al., 2005). Consequently, our findings for tethered swimming are in line with what has been shown for these other modes of exercise.
In the present study, we also found a significant relationship between O2GET stated as a percentage of O2peak and the O2/load slope; however, this correlation was negative (Figure 3; bottom panel). Interestingly, it has been speculated that the inverse relationship between oxidative capacity and exercise economy might be attributable to a high proportion of type IIa fibers, which have lower exercise economy yet contribute significantly to O2max (Hunter et al., 2005). Conversely, type I fibers, which also have a profound impact on O2max, appear to possess high exercise economy (Coyle et al., 1992; Crow and Kushmerick, 1982) and it is these fibers that would likely contribute predominantly to all metabolic rates below O2GET (Henneman and Mendell, 1981). Consequently, our findings suggest that swimmers who can delay recruitment of higher-order fibers for a greater proportion of their overall capacity for work will be more economical during endurance swimming regardless of the magnitude of the O2max they possess. These results are consistent with the belief that training to improve exercise economy might require a different stimulus compared to training to improve O2max; for example, domain-specific training at a heavy intensity for prolonged duration (i.e., steady training) to increase O2GET. This type of training could, therefore, have important practical implications being that exercise economy can account for large variations in performance for endurance athletes with similar O2max values (Conley and Krahenbuhl, 1980).
Collectively, the rapid incrementation and precise control of the load afforded by the tethered-swimming test we investigated make it suitable for discerning metabolic rates that bound the heavy-intensity domain. Consequently, unlike a free-swimming incremental protocol involving distance-controlled stages, this test provides information that is useful for domain-specific exercise prescription according to the three-phase model (Binder et al., 2008; Skinner and McLellan, 1980; Whipp et al., 1989). However, it is important to note that the degree to which the swimming technique employed during tethered swimming relates to that which is present during free swimming has been questioned (Dominguez-Castells and Arellano, 2012). Thus, it is possible that the parameters derived from this test might only apply to domain-specific training utilizing the tethered methodology. However, there is evidence to suggest similarities between measurements derived during tethered and free swimming. For example, Bonen et al. (1980) had swimmers perform an incremental tethered protocol (step increases in the load applied during 2-4 minute work bouts interspersed with ≥ 5 minutes of rest) and found that the peak O2 they observed was highly correlated with (r > 0.99) and not significantly different from the peak O2 recorded when subjects performed three bouts of 200 m free swims at increasing velocities (moderate, faster and all out). Interestingly, in a separate but related experiment, these authors also found a similar peak O2 response during incremental tethered and flume swimming; however, both of these values were greater than the value observed during arm cranking (Bonen et al., 1980). This suggests that the tethered methodology achieved swim specificity that was not present with upper-body exercise per se. Moreover, Perandini et al. (2006) had subjects perform 3-4 constant-load tethered-swimming bouts to exhaustion and modelled the force/time data to reveal a critical force (i.e., force/time asymptote or y-intercept using hyperbolic and linear fits, respectively) that was significantly correlated with the critical velocity they estimated for free swimming (r ≈ 0.90). However, O2 was not measured in that study; hence, the degree to which the metabolic rates at the tethered-swimming critical force and free-swimming critical velocity were similar (e.g., presumably, a critical metabolic rate that is equivalent to O2RCP; Keir et al., 2015) could not be determined. Parameters derived from both incremental (Papoti et al., 2009) and constant-load all-out (Kalva-Filho et al., 2015; Papoti et al., 2010) tethered-swimming tests have also been shown to predict performance during free swimming. Finally, Matsumoto et al. (1999) used a discontinuous incremental-loading tethered protocol with four-minute stages to determine the LT in children with asthma (Matsumoto et al., 1999). Following this assessment, these researchers prescribed free-swim training according to the heart rate at LT and reassessed their subjects using the tethered methodology after six weeks. Importantly, the load at LT was increased by free-swim training, which suggests that a productive free-swim training regimen can be both prescribed and assessed via measurements made using the tethered methodology.
There are a number of limitations to this study that deserve mention. In addition to the inability to confirm that the parameters derived from this test are similar to those that would be identified during free swimming, we also did not verify that O2GET and O2RCP approximate the metabolic rates at the lower and upper boundaries of the heavy-intensity domain during constant-work-rate tethered swimming. While prior research suggests that this is the case for other forms of exercise (e.g., cycling and running; Jones and Poole, 2005; Keir et al., 2015; Whipp and Wasserman, 1972), future research should provide confirmation by having subjects perform the tethered incremental protocol in conjunction with a series of constant-work-rate bouts against a variety of loads to identify moderate/heavy and heavy/severe interfaces.
In conclusion, we demonstrated that an incremental tethered-swimming test with relatively small increases in the resistive load applied every 60 seconds was sufficiently sensitive to reveal the two gas exchange/ventilatory breakpoints (GET and RCP) defining the isocapnic region during incremental exercise. The metabolic rates at these thresholds serve as lower (O2GET) and upper (O2RCP) boundaries of the heavy-intensity domain during constant-work-rate exercise; hence, this singular test can be used to define three distinct exercise-intensity zones for domain-specific training. We also found a high negative correlation between exercise economy and GET suggesting that regardless of O2max, increasing GET relative to maximal capacity can have important implications for endurance-swim performance. Training in the heavy-intensity domain provides a potent stimulus to achieve this objective. Future research should explore the degree to which these conclusions drawn from a tethered-swimming protocol are related to responses observed during free-swim training and competition.
Acknowledgements
The authors would like to thank the swimmers from the BTC team who participated in this study and Santander Totta (PROPG-UNESP) for the fellowship support. This work was supported by the Brazilian National Council for Scientific and Technological Development (CNPq; 479262/2013-6).
Authors submitted their contribution to the article to the editorial board.
References
- Astorino TA, Rietschel JC, Tam PA, Taylor K, Johnson SM, Freedman TP, Sakarya CE. Reinvestigation of optimal duration of VO2max testing. J Exerc Physiol Online. 2004;7:1–8. [Google Scholar]
- Baldari C, Fernandes RJ, Meucci M, Ribeiro J, Vilas-Boas JP, Guidetti L. Is the new AquaTrainer® snorkel valid for VO2 assessment in swimming? Int J Sports Med. 2013;34:336–344. doi: 10.1055/s-0032-1321804. [DOI] [PubMed] [Google Scholar]
- Binder RK, Wonisch M, Corra U, Cohen-Solal A, Vanhees L, Saner H, Schmid JP. Methodological approach to the first and second lactate threshold in incremental cardiopulmonary exercise testing. Eur J Cardiovasc Prev Rehabil. 2008;15:726–734. doi: 10.1097/HJR.0b013e328304fed4. [DOI] [PubMed] [Google Scholar]
- Bogaard HJ, Woltjer HH, van Keimpema AR, Serra RA, Postmus PE, de Vries PMJM. Comparison of the respiratory and hemodynamic responses of healthy subjects to exercise in three different protocols. Occup Med. 1996;46:293–98. doi: 10.1093/occmed/46.4.293. [DOI] [PubMed] [Google Scholar]
- Bonen A, Wilson BA, Yarkony M, Belcastro AN. Maximal oxygen uptake during free, tethered, and flume swimming. J Appl Physiol Respir Environ Exerc Physiol. 1980;48:232–235. doi: 10.1152/jappl.1980.48.2.232. [DOI] [PubMed] [Google Scholar]
- Conley DL, Krahenbuhl GS. Running economy and distance running performance of highly trained athletes. Med Sci Sports Exerc. 1980;12:357–360. [PubMed] [Google Scholar]
- Coyle EF, Sidossis LS, Horowitz JF, Beltz JD. Cycling efficiency is related to the percentage of type I muscle fibers. Med Sci Sports Exerc. 1992;24:782–788. [PubMed] [Google Scholar]
- Crow MT, Kushmerick MJ. Chemical energetics of slow- and fast-twitch muscles of the mouse. J Gen Physiol. 1982;79:147–66. doi: 10.1085/jgp.79.1.147. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dominguez-Castells R, Arellano R. Effect of different loads on stroke and coordination parameters during freestyle semi-tethered swimming. J Hum Kinet. 2012;32:33–41. doi: 10.2478/v10078-012-0021-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Espada MC, Reis JF, Almeida TF, Bruno PM, Vleck VE, Alves FB. Ventilatory and Physiological Responses in Swimmers Below and Above Their Maximal Lactate Steady State. J Strength Cond Res. 2015;29:2836–2843. doi: 10.1519/JSC.0000000000000504. [DOI] [PubMed] [Google Scholar]
- Fernandes RJ, Cardoso CS, Soares SM, Ascensão A, Colaço PJ, Vilas-Boas JP. Time limit and VO2 slow component at intensities corresponding to VO2max in swimmers. Int J Sports Med. 2003;24:576–581. doi: 10.1055/s-2003-43274. [DOI] [PubMed] [Google Scholar]
- Fernandes RJ, Sousa M, Machado L, Vilas-Boas JP. Step length and individual anaerobic threshold assessment in swimming. Int J Sports Med. 2011;32:940–946. doi: 10.1055/s-0031-1283189. [DOI] [PubMed] [Google Scholar]
- Henneman E, Mendell LM. Brooks VB. Handbook of Physiology I. II. Maryland: American Physiological Society; 1981. Functional organisation of motoneuron pool and its inputs; pp. 423–507. [Google Scholar]
- Hunter GR, Bamman MM, Larson-Meyer DE, Joanisse DR, McCarthy JP, Blaudeau TE, Newcomer BR. Inverse relationship between exercise economy and oxidative capacity in muscle. Eur J Appl Physiol. 2005;94:558–568. doi: 10.1007/s00421-005-1370-z. [DOI] [PubMed] [Google Scholar]
- Jones AM, Carter H. The effect of endurance training on parameters of aerobic fitness. Sports Med. 2000;29:373–86. doi: 10.2165/00007256-200029060-00001. [DOI] [PubMed] [Google Scholar]
- Jones AM, DiMenna FJ. Cardinale M, Newton R, Nosaka K. Strength and Conditioning: Biological Principles and Practical Applications. London: John Wiley & Sons, Ltd.; 2011. Cardiovascular Assessment and Aerobic Training Prescription; pp. 291–304. [Google Scholar]
- Jones AM, Poole DC. Jones AM, Poole DC. Oxygen Uptake Kinetics in Sport, Exercise and Medicine. London: Routledge; 2005. Introduction to Oxygen Uptake Kinetics; pp. 3–35. [Google Scholar]
- Kalva-Filho CA, Zagatto AM, Araújo MI, Santiago PR, da Silva AS, Gobatto CA, Papoti M. Relationship between aerobic and anaerobic parameters from 3-minute all-out tethered swimming and 400-m maximal front crawl effort. J Strength Cond Res. 2015;29:238–245. doi: 10.1519/JSC.0000000000000592. [DOI] [PubMed] [Google Scholar]
- Keir DA, Fontana FY, Robertson TC, Murias JM, Paterson DH, Kowalchuk JM, Pogliaghi S. Exercise Intensity Thresholds: Identifying the Boundaries of Sustainable Performance. Med Sci Sports Exerc. 2015;47:1932–40. doi: 10.1249/MSS.0000000000000613. [DOI] [PubMed] [Google Scholar]
- Lucía A, Hoyos J, Pérez M, Santalla A, Chicharro JL. Inverse relationship between VO2max and economy/efficiency in world-class cyclists. Med Sci Sports Exerc. 2002;34:2079–2084. doi: 10.1249/01.MSS.0000039306.92778.DF. [DOI] [PubMed] [Google Scholar]
- Matsumoto I, Araki H, Tsuda K, Odajima H, Nishima S, Higaki Y, Tanaka H, Tanaka M, Shindo M. Effects of swimming training on aerobic capacity and exercise induced bronchoconstriction in children with bronchial asthma. Thorax. 1999;54:196–201. doi: 10.1136/thx.54.3.196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Midgley AW, McNaughton LR, Jones AM. Training to enhance the physiological determinants of longdistance running performance: can valid recommendations be given to runners and coaches based on current scientific knowledge? Sports Med. 2007;37:857–880. doi: 10.2165/00007256-200737100-00003. [DOI] [PubMed] [Google Scholar]
- Papoti M, Vitório R, Araújo GG, Martins LEB, Cunha SA, Gobatto CA. Critical force during tethered swimming for the evaluation of aerobic capacity and prediction of performances in freestyle swimming. Rev. Bras. Cineantropom. 2010;12:14–20. [Google Scholar]
- Papoti M, Vitório R, Araújo GG, DaSilva ASR, Santhiago V, Martins LEB, Cunha SA, Gobatto CA. Determination of force corresponding to maximal lactate steady state in tethered swimming. Int J Exerc Sci. 2009;2:269–279. doi: 10.70252/JJCI8675. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Perandini LAB, Okuno NM, Kokubun E, Nakamura FY. Correlation between critical force and critical velocity and their respective stroke rates. Rev. Bras. Cineantropom. 2006;8:58–65. [Google Scholar]
- Pinna M, Milia R, Roberto S, Marongiu S, Olla S, Loi A, Ortu M, Migliaccio GM, Tocco F, Concu A, Crisafulli A. Assessment of the specificity of cardiopulmonary response during tethered swimming using a new snorkel device. J Physiol Sci. 2013;63:7–16. doi: 10.1007/s12576-012-0226-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Poole DC, Ward SA, Gardner GW, Whipp BJ. Metabolic and respiratory profile of the upper limit for prolonged exercise in man. Ergonomics. 1988;31:1265–79. doi: 10.1080/00140138808966766. [DOI] [PubMed] [Google Scholar]
- Pyne DB, Lee H, Swanwick KM. Monitoring the lactate threshold in world-ranked swimmers. Med Sci Sports Exerc. 2001;33:291–297. doi: 10.1097/00005768-200102000-00019. [DOI] [PubMed] [Google Scholar]
- Ribeiro J, Figueiredo P, Sousa M, De Jesus K, Keskinen K, Vilas-Boas JP, Fernandes RJ. Metabolic and ventilatory thresholds assessment in front crawl swimming. Sports Med Phys Fitness. 2015;55:701–707. [PubMed] [Google Scholar]
- Skinner JS, McLellan TM. The transition from aerobic to anaerobic metabolism. Res Q Exerc Sport. 1980;51:234–248. doi: 10.1080/02701367.1980.10609285. [DOI] [PubMed] [Google Scholar]
- Stanula A, Roczniok R, Maszczyk A, Pietraszewski P, Zając A. The role of aerobic capacity in high-intensity intermittent efforts in ice-hockey. Biol. Sport. 2014;31:193–199. doi: 10.5604/20831862.1111437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Whipp BJ, Davis JA, Torres F, Wasserman K. A test to determine parameters of aerobic function during exercise. J Appl Physiol Respir Environ Exerc Physiol. 1981;50:217–21. doi: 10.1152/jappl.1981.50.1.217. [DOI] [PubMed] [Google Scholar]
- Whipp BJ, Davis JA, Wasserman K. Ventilatory control of the ‘isocapnic buffering’ region in rapidly-incremental exercise. Respir Physiol. 1989;76:357–367. doi: 10.1016/0034-5687(89)90076-5. [DOI] [PubMed] [Google Scholar]
- Whipp BJ, Wasserman K. Oxygen uptake kinetics for various intensities of constant-load work. J Appl Physiol. 1972;33:351–356. doi: 10.1152/jappl.1972.33.3.351. [DOI] [PubMed] [Google Scholar]
- Yoon BK, Kravitz L, Robergs R. VO2max, protocol duration, and the VO2 plateau. Med Sci Sports Exerc. 2007;39:1186–1192. doi: 10.1249/mss.0b13e318054e304. [DOI] [PubMed] [Google Scholar]
- Zhang YY, Johnson MC, Chow N, Wasserman K. Effect of exercise testing protocol on parameters of aerobic function. Med Sci Sports Exerc. 1991;23:625–630. [PubMed] [Google Scholar]