

Abstract
Autoimmune diseases (AIDs) are the result of specific immune responses directed against structures of the self. In normal conditions, the molecules recognized as “self” are tolerated by immune system, but when the self-tolerance is lost, the immune system could react against molecules from the body, causing the loss of self-tolerance, and subsequently the onset of AID that differs for organ target and etiology. Autoimmune thyroid disease (ATD) is caused by the development of autoimmunity against thyroid antigens and comprises Hashimoto’s thyroiditis and Graves disease. They are frequently associated with other organ or non-organ specific AIDs, such as myasthenia gravis (MG). In fact, ATD seems to be the most associated pathology to MG. The etiology of both diseases is multifactorial and it is due to genetic and environmental factors, and each of them has specific characteristics. The two pathologies show many commonalities, such as the organ-specificity with a clear pathogenic effect of antibodies, the pathological mechanisms, such as deregulation of the immune system and the implication of the genetic predisposition. They also show some differences, such as the mode of action of the antibodies and therapies. In this review that focuses on ATD and MG, the common features and the differences between the two diseases are discussed.
Keywords: acetylcholine receptor antibodies, interferon type I, germinal centers, genetics, etiology
Introduction
Autoimmune diseases (AIDs) are the result of specific immune responses directed against structures of the self. Under normal conditions, the immune system is tolerant to molecules recognized as “self” and does not react to antigens expressed in endogenous tissues. If self-tolerance is missing, the immune system could develop an immune response against self-molecules, causing the development of AIDs that include autoimmune thyroid diseases (ATDs) and myasthenia gravis (MG).
The etiology of AIDs is multifactorial and involves genetic and environmental factors. ATDs are endocrine diseases due to an autoimmune reaction against thyroid antigens, in a specific genetic background triggered by exposure to environmental factors (1).
The two main ATDs are Graves disease (GD) and Hashimoto’s thyroiditis (HT) that are characterized by hypothyroidism and thyrotoxicosis, respectively, by the production of thyroid autoantibodies such as thyroid peroxidase (TPO), thyroglobulin (TG), and thyroid-stimulating hormone receptor (TSHR), as well as by lymphocytic infiltration of the thyroid (1). MG is a neuromuscular disorder due to a defective transmission of the nerve impulse to muscles, causing muscle weakness and abnormal fatigability. In most cases, MG is mediated by antibodies targeting the acetylcholine receptor (AChR) while in a minority of patients, the autoantibodies are specific for muscle-specific kinase (MuSK) or agrin receptor LRP4 (low-density lipoprotein receptor-related protein-4). Other targets, such as titin, and ryanodine, have been investigated (2).
Concomitant Thyroiditis and MG: Epidemiological Features
The prevalence of ATDs is high and estimated to be 5% in the general population (3) while MG is a rare disease with an incidence of 8–10 cases per one million persons/year and a prevalence of 150–250 cases per one million (4) (Table 1). Although ATD is one of the most representative organ-specific autoimmune disorders, it is associated with other autoimmune endocrine failures or non-endocrine diseases (5). Among the non-endocrine diseases, we can mention vitiligo, pernicious anemia, MG, autoimmune gastritis, celiac disease, and hepatitis (6, 7). Interestingly, in these associated diseases, the presence of anti-TPO antibodies is more frequent than the prevalence of ATDs (6).
Table 1.
Epidemiological and clinical features of patients with autoimmune thyroid diseases and myasthenia gravis (MG).
| Hashimoto’s thyroiditis | Graves’ disease | AChR-MG | MuSK-MG | |
|---|---|---|---|---|
| Hypothyroidism | Hyperthyroidism | |||
| Incidence | About 2% | About 2% | About 0.1% | About 0.01% |
| Female/male ratio | Around 10 | Around 10 | Early onset: F > M (ratio around 3) | Around 6 |
| Late onset: F = M | ||||
| Tissue pathology | Damage of the thyroid gland | Enlarged thyroid (diffuse goiter) | Thymic pathologies, hyperplasia among the young patients, and thymoma among the oldest patients | No thymic pathology |
| Therapy | Levothyroxine (LT4) | Anti-thyroid drugs, radioactive iodine, and surgery | Anticholinesterase drugs, thymectomy, immunosuppressive drugs (azathioprine, corticosteroids) | Corticosteroids |
| Rituximab |
The aim of this review is to focus on the association between MG and ATDs. The most common diseases coexisting with MG are GD and HT, with a frequency of 7 and 3%, respectively (13). ATDs were diagnosed in 26.8% of MG Polish patients including 4.4% with GD, 9% with HT, and 13.4% with anti-thyroid antibodies (14). In British and German populations, 16% of early-onset MG (EOMG), 9% of late-onset MG, and 17% of thymoma-MG patients had antibodies against TPO or TG (15). About 0.2% of patients affected by ATDs show MG that is much higher than the general incidence of MG (0.01%). MG could be ocular or generalized, even if ATDs are more frequent in the ocular group (16–18). When associated to ATDs, MG shows specific features, such as the young age of onset, mild clinical symptoms, low levels of AChR antibodies, and low frequency of thymic alterations (18–20). These data highlight that the association between MG and ATDs is much more frequent than expected.
Etiology
The etiology of AIDs is still unknown. Drugs, virus infections, radiation, stress are some of the environmental factors that may be involved in the development of ATDs and MG in susceptible individuals, contributing to the activation of an innate immune response (8, 10, 21, 22).
Factors of Predisposition
Autoimmune thyroid diseases are more common among women than men with a female:male ratio of 5–10:1. There is a difference in prevalence and incidence in the base of geographic area, race, and age. The frequency of anti-thyroid antibodies increases with age, showing a peak ranging from 45 to 55 years. In females, one of the two X chromosomes is inactivated during early embryonic stage (23). The inactivation of the same X chromosome, that occurs in more than 80% of cells, could result in defect in immunological tolerance to X-linked antigens that could lead to autoimmunity. Moreover, fetal microchimerism was observed in blood and thyroid tissues from women with either HT or GD. During pregnancy, the production of maternal regulatory T cells (Treg) early in pregnancy could lead to a decrease in the circulating anti-thyroid antibodies, maintaining a state of tolerance to fetal alloantigens in order to avoid fetus rejection. After birth, anti-thyroid antibodies rebound with a transient increase. The persistence of fetal cells in maternal tissues leads to fetal microchimerism (24).
In MG, instead, the early-onset forms, characterized by the age of onset before 50 years, are more frequent in female than male with a ratio female:male of 3:1. Different studies suggest an important role of estrogens in MG (25), since estrogen receptors are expressed on thymic epithelial cells and on thymocytes (21). The female bias in AIDs could be due to reduced expression by estrogens of AIRE, a transcription factor involved in negative selection, resulting in a decreased quality of autoreactive cells elimination (26).
By case–control studies, and more recent genome-wide association studies, different genes have been associated with the ATDs and MG and the presence of specific autoantibodies. Genes involved in T-cell activation and regulation, such as protein tyrosine phosphatase non-receptor 22 (PTPN22), cytotoxic T-lymphocyte antigen-4 (CTLA4), and human leukocyte antigens (HLA), are associated with both ATDs and MG. PTPN22 is an intracellular protein tyrosine phosphatase bound to c-src tyrosine kinase, involved in T-cell activation (27); CTLA4 plays a role in inhibiting T-cell signaling, and the HLA is essential for presenting exogenous antigens for recognition by CD4+ T-helper cells (28). Other genes have been associated to a single disease, as indicated in Figure 1. Therefore, in both ATDs and MG, factors of predisposition include not only genetic background, but also the potential role of sexual hormones.
Figure 1.
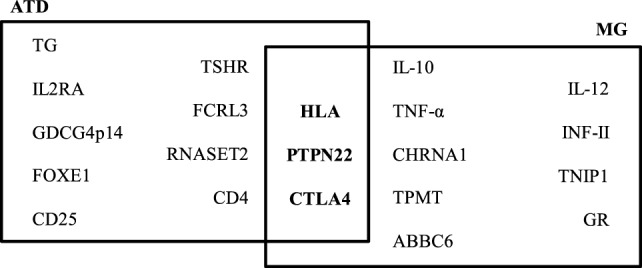
Predisposing genes in autoimmune thyroid diseases (ATDs) and myasthenia gravis (MG). The data on this figure were collected from the following Ref. (28–30).
Triggering Factors
Hepatitis C virus (HCV) infection is the most associated to ATDs both in adults and children, in fact, infected HCV patients show dysfunctions in the thyroid (10, 31). In patients with chronic hepatitis C (CHC), the thyroid disorders are characterized by an increased risk of ATD and hypothyroidism in females, elevated levels of anti-TPO antibodies, and by papillary thyroid cancer risk (32, 33). The hypothesized mechanism is that HCV envelope protein E2 induces strong inflammatory responses in human thyrocytes, resulting in the production of interleukin (IL)-8, IL-6, and tumor necrosis factor-α (TNF-α). The E2 protein also induces the upregulation of molecules involved in innate immune pathways (34). Also human herpes virus-6 (HHV-6) infection is associated with ATD onset, in fact, a high level of HHV-6 activation marker was found in thyroid tissue of patients with ATD (35). In myasthenic patients, the existence of a chronic inflammatory state in the thymus could alter innate immune responses leading to self-tolerance failure (36–39). The inflammatory state could be due to persistent viral replications, in fact, Epstein Barr virus (EBV), cytomegalovirus, human foamy virus, and Nile virus were found to be associated to MG (40, 41). Pathogen infections could play a role in AIDs through dysregulation of toll-like receptor-mediated innate immune responses, which can result in altered innate immune responses and long-term inflammation, rendering the thymus vulnerable to auto-sensitization (40, 41). EBV is one of the main candidates suspected to play a role in MG initiation, since it is able to promote B-cell abnormal activation and survival, and to disrupt critical B-cell tolerance checkpoints (40, 42–44).
Recent data have confirmed a strong association between ATD development and interferon (IFN)-α therapy in patients with CHC. About 40% of CHC patients acquire thyroid disorders while receiving IFN-α. IFN-induced thyroiditis is visible as clinical thyroiditis in about 15% of HCV patients receiving IFN-α and subclinical thyroiditis in up to 40% of patients (45). Moreover, it was observed that the generation of anti-thyroid antibodies tends to continue also after IFN therapy (30). IFN-α could induce thyroiditis by both direct toxic effects on the thyroid and by immune recruitment mechanisms (46).
Interestingly, IFN-I therapies can also prime the development of MG (47). IFN-I, especially IFN-β, could play a central role in the thymic follicular hyperplasia of MG patients by inducing high expression of α-AChR and of CXCL13 chemokine in thymic epithelial cells, and of the chemokine CCL21 in endothelial lymphatic cells, two chemokines involved in the abnormal recruitment of B cells in EOMG thymuses. IFN-β also increases B-cell activating factor expression, which promotes the development of autoreactive B cells (48). Also, IFN-β overexpression in MG thymus can mediate the effects of dsRNA activation and causes α-AChR subunit overexpression, suggesting that IFN-β can play a central role in MG development (36).
Other drugs can induce AIDs, including immunomodulatory agents used to treat melanoma, such as monoclonal antibodies inhibiting the immune checkpoint pathways, as CTLA4 and programmed cell death protein 1 (PD-1), two-cell surface receptors on T cells which down-regulate immune response (49). Ipilimumab is a human immunostimulatory antibody targeting CTLA4 that can cause thyroiditis and/or hypothyroidism in 6% of cases after several cycles. Pembrolizumab and nivolumab act against PD-1 and, if combined with ipilimumab to inhibit both CLT4 and PD-1, show a stronger effect with thyroiditis in 22% of cases (50).
Differential and Common Features in ATDs and MG
Antibodies
Both MG and ATD diseases are organ specific and antibody-mediated, and both kinds of disorders combine many different pathologies. Patients with ATDs have antibodies against proteins of the thyroid, but the characteristics of the disease differ according to the autoantigen. Patients with HT have serum antibodies reacting with TG, TPO, while patients with GD have antibodies against the receptor of TSH (51) (Table 2).
Table 2.
Physiopathological features of patients with autoimmune thyroid diseases and myasthenia gravis (MG).
| Hashimoto’s thyroiditis | Graves’ disease | AChR-MG | MuSK-MG | ||
|---|---|---|---|---|---|
| Humoral immunity | Target of the autoantibodies | TG (20–50%), TPO (90–95%) | TSHR | AChR | MuSK |
| Mechanism of the Abies | Thyroid destruction by cytotoxic cells, death receptors, and impairment of thyroid hormone production | Overactivation of the gland: thyroid stimulatory, blocking, and neutral Abies | AChR blocking, internalization, and degradation | Disruption of neuromuscular junction and inhibition of the retrograde signaling | |
| Role of complement | Yes | Yes | Yes | No | |
| Cellular mechanisms | Infiltration of the target organ | +++ Thyroid | + Thyroid, but not destruction | Neg in the muscle | Neg in the muscle |
| +++ in the thymus | Neg in the thymus | ||||
| Ectopic GC | Yes (thyroid) | Yes (thyroid) | Yes (thymus) | No | |
| T-cell involvement | Th1, Th17 | Th2, Th17 | Th1, Th2, Th17 | Th1, Th17 | |
| Role of epithelial cells | Overproduction of pro-inflammatory cytokines and chemokines by thyroid epithelial cells | Overproduction of pro-inflammatory cytokines and chemokines by thymic epithelial cells | Unknown | ||
| Regulatory B cells | Normal B10 number | Normal B10 number | Decreased B10 cell number | Decreased B10 cell number | |
Myasthenia gravis is due to antibodies against the neuromuscular junction (59). Similarly to thyroiditis, in MG, several antigens are the targets of the autoantibodies, and the disease features depend upon the nature of the antibodies. Patients with anti-AChR, but not with anti-MuSK antibodies, have thymic pathologies, hyperplasia among the young patients, and thymoma among the oldest patients (60).
Interestingly, in both MG and ATDs, some forms of the disease are IgG4 dependent, an Ig subclass that does not bind to the complement. In MG, anti-MuSK antibodies are IgG4 (61). In ATDs, several subcategories of IgG4-mediated diseases have been identified including a fibrosing variant of HT, IgG4-related HT, and GD with elevated IgG4 levels (62). These IgG4 diseases share common mechanisms that involve the mechanical interference of extracellular ligand–receptor interactions by the IgG4 antibodies (63).
The mechanisms of action of the antibodies are quite different in MG and ATD, likely due to the nature of the target antigen and its localization. In HT, together with cytotoxic cells, the antibodies contribute to the destruction of the thyroid, leading to hypothyroidism (Table 2). In the case of GD, the antibodies against TSHR could be stimulatory, blocking or neutral; when the stimulating antibodies predominate, clinical features become obvious (56). Thus, the antibodies are functional, able to stimulate or to inhibit the secretion of thyroid hormones. Fluctuating antibody levels can lead to syndromes alternating between hyperthyroidism and hypothyroidism (64). In the case of MG, anti-AChR antibodies induce its degradation dependent upon the complement, and its internalization (2), while anti-MuSK antibodies disrupt the neuromuscular junction and inhibit the retrograde signaling (65, 66). Recent findings suggest that the anti-AChR antibodies could also have a functional effect, by inducing the overproduction of IL-6, a cytokine that plays a role in muscle biology (67). It is not clear yet if this mechanism participates to the pathogenic mechanisms or is a compensatory mechanism.
Most of the autoantibodies have a clinical usefulness. Anti-TPO and anti-TSHR antibodies are relevant for the diagnosis of HT and GD, respectively (30, 68). Anti-TSH receptor antibodies are of interest in GD as they correlate with the disease severity and their levels decrease with therapies (69). However, anti-TPO and anti-TG Abs are not unique to HT patients since these antibodies are detectable in the majority of GD patients (70). In the case of MG, the anti-AChR antibodies are very useful for the diagnosis but not for the follow-up. On the other hand, for the group of patients with anti-MuSK antibodies, monitoring its level is relevant, since it correlates with the clinical course (2).
Infiltration of the Target Organ and Germinal Centers
Patients with GD can have an infiltration of the thyroid gland, while in the case of HD, the infiltration is severe and accompanied by the destruction of the thyroid (71). Ectopic B-cell follicles are observed in the thyroid gland in HT (72). Autoreactive B cells within these lymphoid follicles were recognized by their ability to bind thyroid antigens (72). In MG, the neuromuscular junctions displays minimal lymphocytic infiltration, while the thymus at least in the young patients includes many infiltrating cells, signs of inflammation, and germinal centers (53). The degree of hyperplasia is related to the level of anti-AChR antibodies (73).
Immune Dysregulation
In both MG and ATDs, T-cell immune-mediated mechanisms are involved. In ATDs, cellular immunity targeting thyroid antigens is very common (74, 75). This mechanism is also a feature of experimental thyroiditis obtained in animals by injection of thyroid antigen with adjuvants (76). In MG, similar data are observed; in the patients, and in the experimental models of MG, T-cell proliferation using the autoantigen or peptides from the AChR has been shown (77–79).
In addition, inflammatory cells such as Th1 and Th17 were shown to be involved in the different forms of thyroid or myasthenic diseases (Table 2). Th1 cytokines are increased in MG patients and its experimental model (EAMG) and normalized with therapies (80, 81). Th1 cells and their cytokines are required for EAMG development (82), through the production of complement-dependent anti-AChR antibodies that are pathogenic (82, 83). In addition, TNF-α has been shown to contribute to the chronic inflammation observed in the MG thymus (84). In ATDs, Th1 cells recruited in the thyroid may be responsible for increased production of IFN-γ and TNF-α, which in turn stimulates the secretion of the pro-inflammatory chemokine CXCL10 from the thyroid cells, resulting in an amplification feedback loop, which could perpetuate the autoimmune process (1). Th17 cells and IL-17 have an inflammatory and pathogenic role in MG and ATD (85). Interestingly, IL-17 also contributes to B-cell responses. Indeed, mice mutated for IL-17 receptor have reduced humoral responses and germinal center development (86). In MG patients, the seric level of IL-17 is increased (84, 87). In the mouse model, IL-17 deficient mice are resistant to develop MG, and the pathogenic anti-murine AChR antibodies are lower compared with wild-type mice (88). In ATDs, an increased differentiation of Th17 lymphocytes and an enhanced synthesis of Th17 cytokines were shown, mainly in HT (89).
Finally, the defects of immune regulation are a hallmark of AIDs. In both MG and thyroiditis, functional defects of Treg cells have been shown while the cell number is normal (90–92). In addition, there is a shift from Treg cells to Th17 cells, suggesting that Treg/Th17 balance is altered (84, 93). However, in MG, it was also demonstrated that the Teff cells are resistant to suppression (84). To our knowledge, there is no equivalent study in ATDs disorders. The number of B reg cells has been shown to be decreased in MG but not ATDs (55, 58, 94).
Microbiota is essential for immunologic and digestive homeostasis (95) and is involved in many AIDs (52). In animals, the lack of microbiota is associated with reduced intestinal surface areas with shorter villi, changes in mucus layer, and permeability (5, 96), together with reduced B- and T-cell production (97). Interestingly, in humans, a morphological and functional damage of the intestinal barrier was similar in patients bearing type-1 diabetes and with ATD (98). In addition, in hyperthyroid patients, the microbiota composition was shown to be altered (99). This aspect has not yet been investigated in MG.
Therapies
Although both MG and ATDs are associated with immune system defects, the treatments are different. In the case of MG, treatments include anticholinesterase molecules and immunosuppressive therapies. Among these therapies are conventional immunosuppressant, such as azathioprine, as well as corticosteroids. Recently, monoclonal antibodies against B lymphocytes have proved interesting (100). In the case of thyroid disease, therapies are aimed at regulating thyroid hormone levels (101). The treatment of choice for HT or hypothyroidism is thyroid hormone replacement. The drug is orally administered usually for life.
Surgery is applied in both pathologies. In MG, thymectomy may be proposed when the thymus is hyperplastic or when a thymoma is associated. Recently, a Thymectomy Trial in Non-Thymomatous MG Patients Receiving Prednisone Therapy was conducted in order understand if transsternal thymectomy with prednisone therapy could be more efficient than prednisone alone after 3 years. An improvement of clinical outcomes over a 3-year period in patients with non-thymomatous MG underwent thymectomy was observed (102). In ATDs, thyroid ablation is recommended in GD when the goiter is large, and in HT when a defined thyroid nodule is present (68). It is interesting to note that in both cases the organ that is operated is inflammatory and contains germinal centers with B lymphocytes participating in the pathogenic response.
Conclusion
In conclusion, MG and ATD share many commonalities. They are both organ-specific AIDs with a clear pathogenic effect of antibodies, although ATDs are 50–100 times more frequent than MG disease.
Interestingly, ocular involvement is observed in both pathologies. The pathological mechanisms high up many commonalities, such as deregulation of the immune system and the implication of predisposing genes, such as HLA and PTNP22 genes. However, the mode of action of the antibodies is different: if the antibodies in ATDs deregulate the level of the hormones, in the case of MG they reduce the expression of receptors at the motor plate leading to a functional defect of the synapse.
It is interesting to note that certain drugs are capable of inducing MG and ATDs, for example, IFN-I or monoclonal antibodies against immune checkpoints have proved to be inducers of these pathologies.
Finally, therapies in MG and ATDs are different. In ATDs, the use of molecules to regulate the level of the hormones is satisfactory. In MG, anticholinesterases are generally insufficient, and immunosuppressive therapy is very frequently associated.
Author Contributions
AL and SB-A discussed the content of the review and wrote the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The reviewer, CR, and handling editor declared their shared affiliation, and the handling editor states that the process nevertheless met the standards of a fair and objective review.
References
- 1.Antonelli A, Ferrari SM, Corrado A, Di Domenicantonio A, Fallahi P. Autoimmune thyroid disorders. Autoimmun Rev (2015) 14:174–80. 10.1016/j.autrev.2014.10.016 [DOI] [PubMed] [Google Scholar]
- 2.Berrih-Aknin S, Frenkian-Cuvelier M, Eymard B. Diagnostic and clinical classification of autoimmune myasthenia gravis. J Autoimmun (2014) 4(8–49):143–8. 10.1016/j.jaut.2014.01.003 [DOI] [PubMed] [Google Scholar]
- 3.Hollowell JG, Staehling NW, Flanders WD, Hannon WH, Gunter EW, Spencer CA, et al. Serum TSH, T(4), and thyroid antibodies in the United States population (1988 to 1994): National Health and Nutrition Examination Survey (NHANES III). J Clin Endocrinol Metab (2002) 87:489–99. 10.1210/jcem.87.2.8182 [DOI] [PubMed] [Google Scholar]
- 4.Gilhus NE. Myasthenia gravis. N Engl J Med (2016) 375:2570–81. 10.1056/NEJMra1602678 [DOI] [PubMed] [Google Scholar]
- 5.Betterle C, Dal Pra C, Mantero F, Zanchetta R. Autoimmune adrenal insufficiency and autoimmune polyendocrine syndromes: autoantibodies, autoantigens, and their applicability in diagnosis and disease prediction. Endocr Rev (2002) 23:327–64. 10.1210/edrv.23.3.0466 [DOI] [PubMed] [Google Scholar]
- 6.Weetman AP. Diseases associated with thyroid autoimmunity: explanations for the expanding spectrum. Clin Endocrinol (Oxf) (2011) 74:411–8. 10.1111/j.1365-2265.2010.03855.x [DOI] [PubMed] [Google Scholar]
- 7.Cellini M, Santaguida MG, Virili C, Capriello S, Brusca N, Gargano L, et al. Hashimoto’s thyroiditis and autoimmune gastritis. Front Endocrinol (2017) 8:92. 10.3389/fendo.2017.00092 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Topliss DJ. Clinical update in aspects of the management of autoimmune thyroid diseases. Endocrinol Metab (Seoul) (2016) 31:493–9. 10.3803/EnM.2016.31.4.493 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Drachman DB. Myasthenia gravis. Semin Neurol (2016) 36:419–24. 10.1055/s-0036-1586265 [DOI] [PubMed] [Google Scholar]
- 10.Dong YH, Fu DG. Autoimmune thyroid disease: mechanism, genetics and current knowledge. Eur Rev Med Pharmacol Sci (2014) 18:3611–8. [PubMed] [Google Scholar]
- 11.Gilhus NE, Skeie GO, Romi F, Lazaridis K, Zisimopoulou P, Tzartos S. Myasthenia gravis—autoantibody characteristics and their implications for therapy. Nat Rev Neurol (2016) 12:259–68. 10.1038/nrneurol.2016.44 [DOI] [PubMed] [Google Scholar]
- 12.Jacobson DL, Gange SJ, Rose NR, Graham NM. Epidemiology and estimated population burden of selected autoimmune diseases in the United States. Clin Immunol Immunopathol (1997) 84:223–43. 10.1006/clin.1997.4412 [DOI] [PubMed] [Google Scholar]
- 13.Mao ZF, Yang LX, Mo XA, Qin C, Lai YR, He NY, et al. Frequency of autoimmune diseases in myasthenia gravis: a systematic review. Int J Neurosci (2011) 121:121–9. 10.3109/00207454.2010.539307 [DOI] [PubMed] [Google Scholar]
- 14.Kubiszewska J, Szyluk B, Szczudlik P, Bartoszewicz Z, Dutkiewicz M, Bielecki M, et al. Prevalence and impact of autoimmune thyroid disease on myasthenia gravis course. Brain Behav (2016) 6:e00537. 10.1002/brb3.537 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Klein R, Marx A, Ströbel P, Schalke B, Nix W, Willcox N. Autoimmune associations and autoantibody screening show focused recognition in patient subgroups with generalized myasthenia gravis. Hum Immunol (2013) 74:1184–93. 10.1016/j.humimm.2013.06.020 [DOI] [PubMed] [Google Scholar]
- 16.Tellez-Zenteno JF, Cardenas G, Estanol B, Garcia-Ramos G, Weder-Cisneros N. Associated conditions in myasthenia gravis: response to thymectomy. Eur J Neurol (2004) 11:767–73. 10.1111/j.1468-1331.2004.00968.x [DOI] [PubMed] [Google Scholar]
- 17.Nakamura H, Usa T, Motomura M, Ichikawa T, Nakao K, Kawasaki E, et al. Prevalence of interrelated autoantibodies in thyroid diseases and autoimmune disorders. J Endocrinol Invest (2008) 31:861–5. 10.1007/BF03346432 [DOI] [PubMed] [Google Scholar]
- 18.Chen YL, Yeh JH, Chiu HC. Clinical features of myasthenia gravis patients with autoimmune thyroid disease in Taiwan. Acta Neurol Scand (2013) 127:170–4. 10.1111/j.1600-0404.2012.01693.x [DOI] [PubMed] [Google Scholar]
- 19.Mistry N, Wass J, Turner MR. When to consider thyroid dysfunction in the neurology clinic. Pract Neurol (2009) 9:145–56. 10.1136/jnnp.2008.167163 [DOI] [PubMed] [Google Scholar]
- 20.Nacu A, Andersen JB, Lisnic V, Owe JF, Gilhus NE. Complicating autoimmune diseases in myasthenia gravis: a review. Autoimmunity (2015) 48:362–8. 10.3109/08916934.2015.1030614 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Kawashima A, Tanigawa K, Akama T, Yoshihara A, Ishii N, Suzuki K. Innate immune activation and thyroid autoimmunity. J Clin Endocrinol Metab (2011) 96:3661–71. 10.1210/jc.2011-1568 [DOI] [PubMed] [Google Scholar]
- 22.Drutel A, Archambeaud F, Caron P. Selenium and the thyroid gland: more good news for clinicians. Clin Endocrinol (Oxf) (2013) 78:155–64. 10.1111/cen.12066 [DOI] [PubMed] [Google Scholar]
- 23.Brix TH, Knudsen GP, Kristiansen M, Kyvik KO, Orstavik KH, Hegedüs L. High frequency of skewed X-chromosome inactivation in females with autoimmune thyroid disease: a possible explanation for the female predisposition to thyroid autoimmunity. J Clin Endocrinol Metab (2005) 90:5949–53. 10.1210/jc.2005-1366 [DOI] [PubMed] [Google Scholar]
- 24.Lepez T, Vandewoestyne M, Hussain S, Van Nieuwerburgh F, Poppe K, Velkeniers B, et al. Fetal microchimeric cells in blood of women with an autoimmune thyroid disease. PLoS One (2011) 6:e29646. 10.1371/journal.pone.0029646 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Nancy P, Berrih-Aknin S. Differential estrogen receptor expression in autoimmune myasthenia gravis. Endocrinology (2005) 146:2345–53. 10.1210/en.2004-1003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Dragin N, Bismuth J, Cizeron-Clairac G, Biferi MG, Berthault C, Serraf A, et al. Estrogen-mediated downregulation of AIRE influences sexual dimorphism in autoimmune diseases. J Clin Invest (2016) 126:1525–37. 10.1172/JCI81894 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Lee YH, Rho YH, Choi SJ, Ji JD, Song GG, Nath SK, et al. The PTPN22 C1858T functional polymorphism and autoimmune diseases—a meta-analysis. Rheumatology (Oxford) (2007) 46:49–56. 10.1093/rheumatology/kel170 [DOI] [PubMed] [Google Scholar]
- 28.Simmonds MJ. GWAS in autoimmune thyroid disease: redefining our understanding of pathogenesis. Nat Rev Endocrinol (2013) 9:277–87. 10.1038/nrendo.2013.56 [DOI] [PubMed] [Google Scholar]
- 29.Avidan N, Le Panse R, Berrih-Aknin S, Miller A. Genetic basis of myasthenia gravis—a comprehensive review. J Autoimmun (2014) 52:146–53. 10.1016/j.jaut.2013.12.001 [DOI] [PubMed] [Google Scholar]
- 30.Tomer Y, Huber A. The etiology of autoimmune thyroid disease: a story of genes and environment. J Autoimmun (2009) 32:231–9. 10.1016/j.jaut.2009.02.007 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Fallahi P, Ferrari SM, Giuggioli D, Manfredi A, Mancusi C, Fabiani S, et al. Thyroid involvement in hepatitis C—associated mixed cryoglobulinemia. Hormones (Athens) (2014) 13:16–23. [DOI] [PubMed] [Google Scholar]
- 32.Hammerstad SS, Jahnsen FL, Tauriainen S, Hyöty H, Paulsen T, Norheim I, et al. Inflammation and increased myxovirus resistance protein A expression in thyroid tissue in the early stages of Hashimoto’s thyroiditis. Thyroid (2013) 23:334–41. 10.1089/thy.2012.0264 [DOI] [PubMed] [Google Scholar]
- 33.Fallahi P, Ferrari SM, Vita R, Benvenga S, Antonelli A. The role of human parvovirus B19 and hepatitis C virus in the development of thyroid disorders. Rev Endocr Metab Disord (2016) 17(4):529–35. 10.1007/s11154-016-9361-4 [DOI] [PubMed] [Google Scholar]
- 34.Hammerstad SS, Stefan M, Blackard J, Owen RP, Lee HJ, Concepcion E, et al. Hepatitis C virus E2 protein induces upregulation of IL-8 pathways and production of heat shock proteins in human thyroid cells. J Clin Endocrinol Metab (2017) 102:689–97. 10.1210/jc.2016-3403 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Sultanova A, Cistjakovs M, Gravelsina S, Chapenko S, Roga S, Cunskis E, et al. Association of active human herpesvirus-6 (HHV-6) infection with autoimmune thyroid gland diseases. Clin Microbiol Infect (2017) 23:50.e51–e55. 10.1016/j.cmi.2016.09.023 [DOI] [PubMed] [Google Scholar]
- 36.Cufi P, Dragin N, Weiss JM, Martinez-Martinez P, De Baets MH, Roussin R, et al. Implication of double-stranded RNA signaling in the etiology of autoimmune myasthenia gravis. Ann Neurol (2013) 73:281–93. 10.1002/ana.23791 [DOI] [PubMed] [Google Scholar]
- 37.Cavalcante P, Le Panse R, Berrih-Aknin S, Maggi L, Antozzi C, Baggi F, et al. The thymus in myasthenia gravis: site of “innate autoimmunity”? Muscle Nerve (2011) 44:467–84. 10.1002/mus.22103 [DOI] [PubMed] [Google Scholar]
- 38.Cavalcante P, Cufi P, Mantegazza R, Berrih-Aknin S, Bernasconi P, Le Panse R. Etiology of myasthenia gravis: innate immunity signature in pathological thymus. Autoimmun Rev (2013) 12:863–74. 10.1016/j.autrev.2013.03.010 [DOI] [PubMed] [Google Scholar]
- 39.Cordiglieri C, Marolda R, Franzi S, Cappelletti C, Giardina C, Motta T, et al. Innate immunity in myasthenia gravis thymus: pathogenic effects of toll-like receptor 4 signaling on autoimmunity. J Autoimmun (2014) 52:74–89. 10.1016/j.jaut.2013.12.013 [DOI] [PubMed] [Google Scholar]
- 40.Cavalcante P, Galbardi B, Franzi S, Marcuzzo S, Barzago C, Bonanno S, et al. Increased expression of toll-like receptors 7 and 9 in myasthenia gravis thymus characterized by active Epstein-Barr virus infection. Immunobiology (2016) 221:516–27. 10.1016/j.imbio.2015.12.007 [DOI] [PubMed] [Google Scholar]
- 41.Barzago C, Lum J, Cavalcante P, Srinivasan KG, Faggiani E, Camera G, et al. A novel infection- and inflammation-associated molecular signature in peripheral blood of myasthenia gravis patients. Immunobiology (2016) 221:1227–36. 10.1016/j.imbio.2016.06.012 [DOI] [PubMed] [Google Scholar]
- 42.Bernasconi P, Barberis M, Baggi F, Passerini L, Cannone M, Arnoldi E, et al. Increased toll-like receptor 4 expression in thymus of myasthenic patients with thymitis and thymic involution. Am J Pathol (2005) 167:129–39. 10.1016/S0002-9440(10)62960-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Le Panse R, Cizeron-Clairac G, Bismuth J, Berrih-Aknin S. Microarrays reveal distinct gene signatures in the thymus of seropositive and seronegative myasthenia gravis patients and the role of CC chemokine ligand 21 in thymic hyperplasia. J Immunol (2006) 177:7868–79. 10.4049/jimmunol.177.11.7868 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Cizeron-Clairac G, Le Panse R, Frenkian-Cuvelier M, Meraouna A, Truffault F, Bismuth J, et al. Thymus and Myasthenia gravis: what can we learn from DNA microarrays? J Neuroimmunol (2008) 20(1–202):57–63. 10.1016/j.jneuroim.2008.06.028 [DOI] [PubMed] [Google Scholar]
- 45.Hasham A, Zhang W, Lotay V, Haggerty S, Stefan M, Concepcion E, et al. Genetic analysis of interferon induced thyroiditis (IIT): evidence for a key role for MHC and apoptosis related genes and pathways. J Autoimmun (2013) 44:61–70. 10.1016/j.jaut.2013.04.002 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Akeno N, Smith EP, Stefan M, Huber AK, Zhang W, Keddache M, et al. IFN-alpha mediates the development of autoimmunity both by direct tissue toxicity and through immune cell recruitment mechanisms. J Immunol (2011) 186:4693–706. 10.4049/jimmunol.1002631 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Dionisiotis J, Zoukos Y, Thomaides T. Development of myasthenia gravis in two patients with multiple sclerosis following interferon beta treatment. J Neurol Neurosurg Psychiatry (2004) 75:1079. 10.1136/jnnp.2003.028233 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Cufi P, Dragin N, Ruhlmann N, Weiss JM, Fadel E, Serraf A, et al. Central role of interferon-beta in thymic events leading to myasthenia gravis. J Autoimmun (2014) 52:44–52. 10.1016/j.jaut.2013.12.016 [DOI] [PubMed] [Google Scholar]
- 49.Ribas A. Tumor immunotherapy directed at PD-1. N Engl J Med (2012) 366:2517–9. 10.1056/NEJMe1205943 [DOI] [PubMed] [Google Scholar]
- 50.Ryder M, Callahan M, Postow MA, Wolchok J, Fagin JA. Endocrine-related adverse events following ipilimumab in patients with advanced melanoma: a comprehensive retrospective review from a single institution. Endocr Relat Cancer (2014) 21:371–81. 10.1530/ERC-13-0499 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Stassi G, De Maria R. Autoimmune thyroid disease: new models of cell death in autoimmunity. Nat Rev Immunol (2002) 2:195–204. 10.1038/nri750 [DOI] [PubMed] [Google Scholar]
- 52.Berrih-Aknin S. Myasthenia gravis: paradox versus paradigm in autoimmunity. J Autoimmun (2014) 52:1–28. 10.1016/j.jaut.2014.05.001 [DOI] [PubMed] [Google Scholar]
- 53.Berrih-Aknin S, Le Panse R. Myasthenia gravis: a comprehensive review of immune dysregulation and etiological mechanisms. J Autoimmun (2014) 52:90–100. 10.1016/j.jaut.2013.12.011 [DOI] [PubMed] [Google Scholar]
- 54.Berrih-Aknin S, Ragheb S, Le Panse R, Lisak RP. Ectopic germinal centers, BAFF and anti-B-cell therapy in myasthenia gravis. Autoimmun Rev (2013) 12:885–93. 10.1016/j.autrev.2013.03.011 [DOI] [PubMed] [Google Scholar]
- 55.Kristensen B. Regulatory B and T cell responses in patients with autoimmune thyroid disease and healthy controls. Dan Med J (2016) 63:B5177. [PubMed] [Google Scholar]
- 56.Khan FA, Al-Jameil N, Khan MF, Al-Rashid M, Tabassum H. Thyroid dysfunction: an autoimmune aspect. Int J Clin Exp Med (2015) 8:6677–81. [PMC free article] [PubMed] [Google Scholar]
- 57.Ajjan RA, Weetman AP. The pathogenesis of Hashimoto’s thyroiditis: further developments in our understanding. Horm Metab Res (2015) 47:702–10. 10.1055/s-0035-1548832 [DOI] [PubMed] [Google Scholar]
- 58.Santaguida MG, Gatto I, Mangino G, Virili C, Stramazzo I, Fallahi P, et al. BREG cells in Hashimoto’s thyroiditis isolated or associated to further organ-specific autoimmune diseases. Clin Immunol (2017). 10.1016/j.clim.2017.04.012 [DOI] [PubMed] [Google Scholar]
- 59.Le Panse R, Berrih-Aknin S. Autoimmune myasthenia gravis: autoantibody mechanisms and new developments on immune regulation. Curr Opin Neurol (2013) 26:569–76. 10.1097/WCO.0b013e328364d6cd [DOI] [PubMed] [Google Scholar]
- 60.Berrih-Aknin S. Role of the thymus in autoimmune myasthenia gravis. Clin Exp Neuroimmunol (2016) 7:226–37. 10.1111/cen3.12319 [DOI] [Google Scholar]
- 61.Huijbers MG, Zhang W, Klooster R, Niks EH, Friese MB, Straasheijm KR, et al. MuSK IgG4 autoantibodies cause myasthenia gravis by inhibiting binding between MuSK and Lrp4. Proc Natl Acad Sci USA (2013) 110:20783–8. 10.1073/pnas.1313944110 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Kottahachchi D, Topliss DJ. Immunoglobulin G4-related thyroid diseases. Eur Thyroid J (2016) 5:231–9. 10.1159/000452623 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Huijbers MG, Querol LA, Niks EH, Plomp JJ, van der Maarel SM, Graus F, et al. The expanding field of IgG4-mediated neurological autoimmune disorders. Eur J Neurol (2015) 22:1151–61. 10.1111/ene.12758 [DOI] [PubMed] [Google Scholar]
- 64.Tamai H, Kasagi K, Takaichi Y, Takamatsu J, Komaki G, Matsubayashi S, et al. Development of spontaneous hypothyroidism in patients with Graves’ disease treated with antithyroidal drugs: clinical, immunological, and histological findings in 26 patients. J Clin Endocrinol Metab (1989) 69:49–53. 10.1210/jcem-69-1-49 [DOI] [PubMed] [Google Scholar]
- 65.Cole RN, Reddel SW, Gervasio OL, Phillips WD. Anti-MuSK patient antibodies disrupt the mouse neuromuscular junction. Ann Neurol (2008) 63:782–9. 10.1002/ana.21371 [DOI] [PubMed] [Google Scholar]
- 66.Koneczny I, Cossins J, Vincent A. The role of muscle-specific tyrosine kinase (MuSK) and mystery of MuSK myasthenia gravis. J Anat (2014) 224:29–35. 10.1111/joa.12034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Maurer M, Bougoin S, Feferman T, Frenkian M, Bismuth J, Mouly V, et al. IL-6 and Akt are involved in muscular pathogenesis in myasthenia gravis. Acta Neuropathol Commun (2015) 3:1. 10.1186/s40478-014-0179-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Caturegli P, De Remigis A, Rose NR. Hashimoto thyroiditis: clinical and diagnostic criteria. Autoimmun Rev (2014) 13:391–7. 10.1016/j.autrev.2014.01.007 [DOI] [PubMed] [Google Scholar]
- 69.Barbesino G, Tomer Y. Clinical review: clinical utility of TSH receptor antibodies. J Clin Endocrinol Metab (2013) 98:2247–55. 10.1210/jc.2012-4309 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Menconi F, Marcocci C, Marino M. Diagnosis and classification of Graves’ disease. Autoimmun Rev (2014) 13:398–402. 10.1016/j.autrev.2014.01.013 [DOI] [PubMed] [Google Scholar]
- 71.Wang SH, Baker JR. The role of apoptosis in thyroid autoimmunity. Thyroid (2007) 17:975–9. 10.1089/thy.2007.0208 [DOI] [PubMed] [Google Scholar]
- 72.Armengol MP, Juan M, Lucas-Martín A, Fernández-Figueras MT, Jaraquemada D, Gallart T, et al. Thyroid autoimmune disease: demonstration of thyroid antigen-specific B cells and recombination-activating gene expression in chemokine-containing active intrathyroidal germinal centers. Am J Pathol (2001) 159:861–73. 10.1016/S0002-9440(10)61762-2 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Truffault F, de Montpreville V, Eymard B, Sharshar T, Le Panse R, Berrih-Aknin S. Thymic germinal centers and corticosteroids in myasthenia gravis: an immunopathological study in 1035 cases and a critical review. Clin Rev Allergy Immunol (2017) 52:108–24. 10.1007/s12016-016-8558-3 [DOI] [PubMed] [Google Scholar]
- 74.Weetman AP. Determinants of autoimmune thyroid disease. Nat Immunol (2001) 2:769–70. 10.1038/ni0901-769 [DOI] [PubMed] [Google Scholar]
- 75.Dayan CM, Daniels GH. Chronic autoimmune thyroiditis. N Engl J Med (1996) 335:99–107. 10.1056/NEJM199607113350206 [DOI] [PubMed] [Google Scholar]
- 76.Flynn JC, Conaway DH, Cobbold S, Waldmann H, Kong YC. Depletion of L3T4+ and Lyt-2+ cells by rat monoclonal antibodies alters the development of adoptively transferred experimental autoimmune thyroiditis. Cell Immunol (1989) 122:377–90. 10.1016/0008-8749(89)90085-3 [DOI] [PubMed] [Google Scholar]
- 77.Sun JB, Harcourt G, Wang ZY, Hawke S, Olsson T, Fredrikson S, et al. T cell responses to human recombinant acetylcholine receptor-alpha subunit in myasthenia gravis and controls. Eur J Immunol (1992) 22:1553–9. 10.1002/eji.1830220631 [DOI] [PubMed] [Google Scholar]
- 78.Berrih-Aknin S, Cohen-Kaminsky S, Lepage V, Neumann D, Bach JF, Fuchs S. T-cell antigenic sites involved in myasthenia gravis: correlations with antibody titre and disease severity. J Autoimmun (1991) 4:137–53. 10.1016/0896-8411(91)90013-3 [DOI] [PubMed] [Google Scholar]
- 79.Fujii Y, Lindstrom J. Specificity of the T cell immune response to acetylcholine receptor in experimental autoimmune myasthenia gravis. Response to subunits and synthetic peptides. J Immunol (1988) 140:1830–7. [PubMed] [Google Scholar]
- 80.Conti-Fine BM, Milani M, Kaminski HJ. Myasthenia gravis: past, present, and future. J Clin Invest (2006) 116:2843–54. 10.1172/JCI29894 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Im SH, Barchan D, Maiti PK, Raveh L, Souroujon MC, Fuchs S. Suppression of experimental myasthenia gravis, a B cell-mediated autoimmune disease, by blockade of IL-18. FASEB J (2001) 15:2140–8. 10.1096/fj.01-0072com [DOI] [PubMed] [Google Scholar]
- 82.Karachunski PI, Ostlie NS, Monfardini C, Conti-Fine BM. Absence of IFN-gamma or IL-12 has different effects on experimental myasthenia gravis in C57BL/6 mice. J Immunol (2000) 164:5236–44. 10.4049/jimmunol.164.10.5236 [DOI] [PubMed] [Google Scholar]
- 83.Souroujon MC, Brenner T, Fuchs S. Development of novel therapies for MG: studies in animal models. Autoimmunity (2010) 43:446–60. 10.3109/08916930903518081 [DOI] [PubMed] [Google Scholar]
- 84.Gradolatto A, Nazzal D, Truffault F, Bismuth J, Fadel E, Foti M, et al. Both Treg cells and Tconv cells are defective in the myasthenia gravis thymus: roles of IL-17 and TNF-alpha. J Autoimmun (2014) 52:53–63. 10.1016/j.jaut.2013.12.015 [DOI] [PubMed] [Google Scholar]
- 85.Bedoya SK, Lam B, Lau K, Larkin J, III. Th17 cells in immunity and autoimmunity. Clin Dev Immunol (2013) 2013:986789. 10.1155/2013/986789 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Hsu HC, Yang P, Wang J, Wu Q, Myers R, Chen J, et al. Interleukin 17-producing T helper cells and interleukin 17 orchestrate autoreactive germinal center development in autoimmune BXD2 mice. Nat Immunol (2008) 9:166–75. 10.1038/ni1552 [DOI] [PubMed] [Google Scholar]
- 87.Roche JC, Capablo JL, Larrad L, Gervas-Arruga J, Ara JR, Sánchez A, et al. Increased serum interleukin-17 levels in patients with myasthenia gravis. Muscle Nerve (2011) 44:278–80. 10.1002/mus.22070 [DOI] [PubMed] [Google Scholar]
- 88.Schaffert H, Pelz A, Saxena A, Losen M, Meisel A, Thiel A, et al. IL-17-producing CD4(+) T cells contribute to the loss of B-cell tolerance in experimental autoimmune myasthenia gravis. Eur J Immunol (2015) 45:1339–47. 10.1002/eji.201445064 [DOI] [PubMed] [Google Scholar]
- 89.Figueroa-Vega N, Alfonso-Pérez M, Benedicto I, Sánchez-Madrid F, González-Amaro R, Marazuela M. Increased circulating pro-inflammatory cytokines and Th17 lymphocytes in Hashimoto’s thyroiditis. J Clin Endocrinol Metab (2010) 95:953–62. 10.1210/jc.2009-1719 [DOI] [PubMed] [Google Scholar]
- 90.Mao C, Wang S, Xiao Y, Xu J, Jiang Q, Jin M, et al. Impairment of regulatory capacity of CD4+CD25+ regulatory T cells mediated by dendritic cell polarization and hyperthyroidism in Graves’ disease. J Immunol (2011) 186:4734–43. 10.4049/jimmunol.0904135 [DOI] [PubMed] [Google Scholar]
- 91.Glick AB, Wodzinski A, Fu P, Levine AD, Wald DN. Impairment of regulatory T-cell function in autoimmune thyroid disease. Thyroid (2013) 23:871–8. 10.1089/thy.2012.0514 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Balandina A, Lecart S, Dartevelle P, Saoudi A, Berrih-Aknin S. Functional defect of regulatory CD4(+)CD25+ T cells in the thymus of patients with autoimmune myasthenia gravis. Blood (2005) 105:735–41. 10.1182/blood-2003-11-3900 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Li C, Yuan J, Zhu YF, Yang XJ, Wang Q, Xu J, et al. Imbalance of Th17/Treg in different subtypes of autoimmune thyroid diseases. Cell Physiol Biochem (2016) 40:245–52. 10.1159/000452541 [DOI] [PubMed] [Google Scholar]
- 94.Sun F, Ladha SS, Yang L, Liu Q, Shi SX, Su N, et al. Interleukin-10 producing-B cells and their association with responsiveness to rituximab in myasthenia gravis. Muscle Nerve (2014) 49:487–94. 10.1002/mus.23951 [DOI] [PubMed] [Google Scholar]
- 95.Shanahan F. The gut microbiota-a clinical perspective on lessons learned. Nat Rev Gastroenterol Hepatol (2012) 9:609–14. 10.1038/nrgastro.2012.145 [DOI] [PubMed] [Google Scholar]
- 96.Natividad JM, Verdu EF. Modulation of intestinal barrier by intestinal microbiota: pathological and therapeutic implications. Pharmacol Res (2013) 69:42–51. 10.1016/j.phrs.2012.10.007 [DOI] [PubMed] [Google Scholar]
- 97.Macpherson AJ, Harris NL. Interactions between commensal intestinal bacteria and the immune system. Nat Rev Immunol (2004) 4:478–85. 10.1038/nri1373 [DOI] [PubMed] [Google Scholar]
- 98.Virili C, Centanni M. Does microbiota composition affect thyroid homeostasis? Endocrine (2015) 49:583–7. 10.1007/s12020-014-0509-2 [DOI] [PubMed] [Google Scholar]
- 99.Zhou L, Li X, Ahmed A, Wu D, Liu L, Qiu J, et al. Gut microbe analysis between hyperthyroid and healthy individuals. Curr Microbiol (2014) 69:675–80. 10.1007/s00284-014-0640-6 [DOI] [PubMed] [Google Scholar]
- 100.Tandan R, Hehir MK, II, Waheed W, Howard DB. Rituximab treatment of myasthenia gravis: a systematic review. Muscle Nerve (2017). 10.1002/mus.25597 [DOI] [PubMed] [Google Scholar]
- 101.Fallahi P, Ferrari SM, Elia G, Nasini F, Colaci M, Giuggioli D, et al. Novel therapies for thyroid autoimmune diseases. Expert Rev Clin Pharmacol (2016) 9:853–61. 10.1586/17512433.2016.1157468 [DOI] [PubMed] [Google Scholar]
- 102.Wolfe GI, Kaminski HJ, Aban IB, Minisman G, Kuo HC, Marx A, et al. Randomized trial of thymectomy in Myasthenia gravis. N Engl J Med (2016) 375:511–22. 10.1056/NEJMoa1602489 [DOI] [PMC free article] [PubMed] [Google Scholar]