

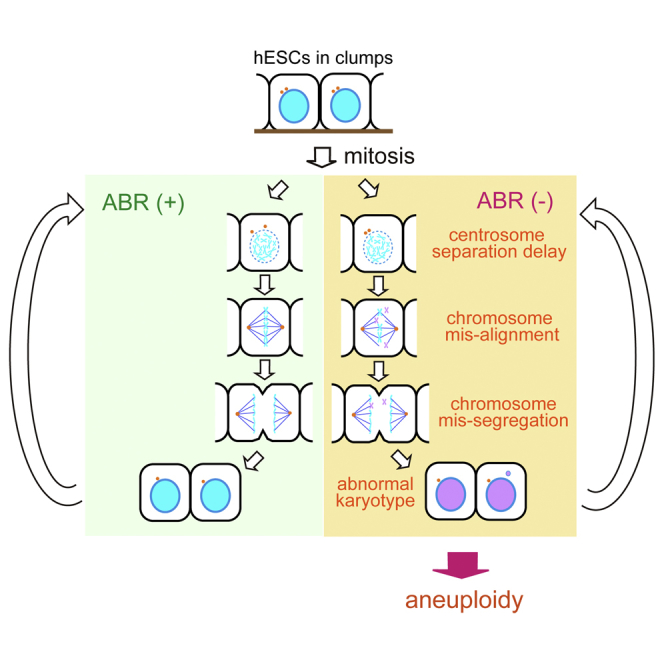
Summary
Pluripotent stem cells can undergo repeated self-renewal while retaining genetic integrity, but they occasionally acquire aneuploidy during long-term culture, which is a practical obstacle for medical applications of human pluripotent stem cells. In this study, we explored the biological roles of ABR, a regulator of RHO family small GTPases, and found that it has pivotal roles during mitotic processes in human embryonic stem cells (hESCs). Although ABR has been shown to be involved in dissociation-induced hESC apoptosis, it does not appear to have direct effects on cell survival unless cell-cell contact is impaired. Instead, we found that it is important for faithful hESC division. Mechanistically, ABR depletion compromised centrosome dynamics and predisposed the cell to chromosome misalignment and missegregation, which raised the frequency of aneuploidy. These results provide insights into the mechanisms that support the genetic integrity of self-renewing hESCs.
Keywords: human embryonic stem cells, mitotic fidelity, aneuploidy, RHO family small GTPases, ABR, cell-cell communication
Graphical Abstract
Highlights
-
•
ABR depletion leads to G2/M accumulation in hESCs
-
•
Centrosome dynamics and mitotic fidelity are compromised upon ABR depletion
-
•
When mitosis progresses without ABR, hESCs show a high incidence of aneuploidy
-
•
ABR safeguards faithful chromosome inheritance during hESC division
In this article, Ohgushi and colleagues report that ABR depletion in hESCs causes severe defects in several mitotic processes, including centrosome separation and chromosome alignment/segregation. Importantly, ABR deficiency increases the frequency of aneuploidy, highlighting its key role in conferring robust way to faithful inheritance of genetic information on self-renewing hESCs.
Introduction
The faithful inheritance of genetic material during repetitive cell division is fundamental for animal development and tissue regeneration in multicellular organisms. Several quality control mechanisms survey the organism for genetic normality and then activate programs for error correction or elimination of abnormal cells. These mechanisms could suppress aneuploidy, a genetic aberration that arises from missegregation of whole chromosomes during mitosis. If aneuploid cells override these barriers and continue proliferating, they can acquire cancerous properties. It is well recognized that chromosomal instability, the condition in which aneuploidy occurs at a high rate, underlies genetic abnormalities found in many types of tumor cells. Actually, aneuploidy is commonly observed in a wide range of tumor tissues and cancer-derived cell lines (reviewed in Santaguida and Amon, 2015).
Pluripotent stem cells, such as embryonic stem cells (ESCs) and induced pluripotent stem cells (iPSCs), have special abilities to differentiate into cells of all three germ layers (pluripotency) and to undergo unlimited proliferation while retaining their identities (self-renewal) (Nichols and Smith, 2012). In addition, they are known to be able to maintain genetic integrity, which is an essential requirement for their utilization in genetic studies or medical applications. Maintaining chromosome number is particularly important in pluripotent stem cells because aneuploidy can lead not only to oncogenic transformation but also to differentiation dysregulation (Peterson and Loring, 2014, Ben-David et al., 2014, Lamm et al., 2016, Zhang et al., 2016). Nevertheless, aneuploidy is often observed in some human ESC (hESC) and iPSC lines (Spits et al., 2008, Mayshar et al., 2010, Taapken et al., 2011). A screening study of a large number of hESC/iPSC lines documented a progressive tendency to acquire karyotypic abnormality during long-term culture, indicating a culture-associated susceptibility to aneuploidy (International Stem Cell Initiative et al., 2011). Although previous reports describe several putative risks contributing to chromosome instability, including excessive replication stresses and DNA damage responses (Zhao et al., 2015, Lamm et al., 2016, Jacobs et al., 2016), safeguarding mechanisms to counteract these threats remain to be elucidated.
We previously reported that the aberrant activation of the RHO-ROCK pathway was responsible for dissociation-induced hESC apoptosis (Watanabe et al., 2007, Ohgushi et al., 2010). We also identified ABR, a modulator of RHO family small GTPase activities, as an upstream factor controlling the survival-or-death decision of dissociated hESCs. The ROCK activation is thought to affect cellular motility (Li et al., 2010), but whether this phenomenon represents any biological implications has remained a mystery. To tackle this question, we sought to explore ABR function. We found that ABR did not have direct effects on cell survival unless cell-cell contact was impaired. Instead, we obtained unexpected data indicating that ABR depletion increased the frequency of chromosome missegregation. These findings shed light on the safeguarding mechanism that prevents chromosomal instability in hESCs.
Results
ABR Depletion Caused Cellular Accumulation at the G2-M Phase of the Cell Cycle
To examine ABR functions in hESCs, we applied a doxycycline (dox)-inducible short hairpin RNA expression strategy (Figure S1A, and refer to Ohgushi et al., 2015). This method permitted the selective depletion of target molecules with controlled timing and under the same genotypic background. We succeeded in reducing ABR protein to an undetectable level after dox addition (Figure 1A), and we refer to these genetically engineered cells as tet-shABR hESCs. To address the putative primary responses caused by ABR depletion, we first examined cellular behaviors on day 3 of dox treatment when the ABR protein level seemed to reach a minimum (Figure S1B). The expression levels of pluripotent markers were nearly equal between control and dox-treated cells (Figures S1C and S1D). The number of dead cells significantly increased after dox treatment, but the extent was not substantial (Figure S1E). At this time point, it was the cell cycle profile that we found remarkably different between dox-treated and untreated tet-shABR cells (Figures 1B and 1C).
Figure 1.

ABR Depletion Leads to G2-M Accumulation
(A) Western blotting analyses. The tet-shABR cells were cultured with or without dox for 3 days. HSC70 was examined as a loading control.
(B and C) Cell cycle profile of dox-treated (right) or untreated (left) tet-shABR cells. The histograms show representative results from three independent experiments (B). The occupancy of each phase in the analyzed cells is indicated in these histograms and also shown as a bar graph (C).
(D) Population dominance of S versus G2-M phase is represented as the ratio of S to G2-M phase cells.
(E) FUCCI-expressing tet-shABR cells were cultured with or without dox for 72 hr. Cells were classified into the indicated five categories according to the time length of one round of cell cycle (n = 20 from three independent imaging experiments).
(F) Growth curve of tet-shABR cells that were cultured with or without dox for 8 days.
(G) Rescue experiments. The expression of RNAi-resistant ABR mutants (Abr∗) lacking the GEF domain (ΔDH) but not GAP domain (ΔDH) in ABR-depleted hESC restores S phase dominance. ABR∗ was used as a positive control.
All experiments were repeated three times and data are shown as representative (A and B), bar graphs (C, D, and G), or a scatterplot (F). Error bars in graphs represent SD (C, D, F, and G). Statistics: Dunnett's test (G, n = 3) versus lane 2; n.s., not significant and ∗∗p < 0.05. See also Figure S1.
ESCs are known to exhibit a characteristic cell cycle pattern that includes an abbreviated G1 phase and dominant occupancy of replicating S phase cells (Boward et al., 2016). Indeed, our control cells exhibited this typical pattern (Figure 1B, left). Interestingly, in dox-treated cells, the S phase population was decreased while the G2 and M populations were greatly increased (Figure 1B, right), resulting in an inversion in population dominancy (Figures 1C and 1D). These observations demonstrate that ABR-depleted hESCs accumulate at the G2-to-M stage. To further confirm this, S phase cells were labeled with a transient bromodeoxyuridine supplementation, and then traced during the subsequent 12 hr (Figures S1F–S1G). In control cells, the labeled population passed through G2-M into the next G1 phase. In dox-treated cells, however, labeled cells seemed to be trapped at the 4N state and struggled to proceed into the next cycle, suggesting that ABR-depleted cells had trouble entering or exiting mitosis. In addition to these population analyses, we performed single-cell tracing using tet-shABR cells expressing a FUCCI reporter (Figure S1H, Sakaue-Sawano et al., 2008). This revealed the tendency of ABR-depleted cells to take longer times to complete one round of a cell cycle than did control cells (Figure 1E). Consequently, ABR-depleted cells showed significant growth retention when cultured for a further extended period (Figure 1F).
ABR protein has a unique domain structure: a guanine nucleotide exchanging factor (GEF) domain at the N terminus and a GTPase-activating protein (GAP) domain at the C terminus (Figure S1I). When isolated and tested by in vitro assay, these domains were shown to possess GEF and GAP activities for the selected members of RHO family small GTPases (Heisterkamp et al., 1993, Chuang et al., 1995). We sought to determine which domain is responsible for ABR's ability to drive cell cycle progression by restoring ABR expression using RNAi-resistant or domain-deleted mutants (Figure 1G). The introduction of codon-swapped RNAi-immune mutant (ABR∗) into tet-shABR hESCs restored the S phase dominance. A partial restoration was observed when an ABR mutant lacking a GEF domain was introduced. On the other hand, a GAP-dead mutant showed little rescuing effects, indicating the importance of GAP activity for ABR.
In sum, these results show that ABR plays a key role in cell cycle progression from G2-M to the next G1 phase through its GAP activity.
Compromised Centrosome Dynamics upon ABR Depletion
To obtain mechanistic insights into the accumulation of ABR-depleted cells in the G2-M phase, we focused on the centrosome, a central organelle that operates multiple mitotic events (Tanenbaum and Medema, 2010). Centrosomes were replicated during S phase, matured at G2 phase and separated bilaterally in parallel with M phase entry (Figure 2A), all of which are important prerequisites for proper cell division. In both control and dox-treated cells, duplicated centrosomes were evident at prophase (data not shown). The phosphorylation level of a centrosomal kinase AURORA-A (AURKA) did not demonstrate a substantial difference in centrosome maturation (Figures S2A–S2C). Otherwise, by monitoring centrosome dynamics using tet-shABR hESCs expressing mVenus-fused centrin-2 (CETN2), a component of the centrosome, we found that it took longer in dox-treated cells for each centrosome to move to the opposite side (Figure 2B and Movie S1). Notably, whereas in control cells the nuclear envelope breakdown (NEB) occurred immediately after centrosomes started to move bilaterally, a much longer time was needed for NEB to take place in dox-treated cells (Figures 2B and 2C). As a consequence, inter-centrosomal distances at the time of NEB were significantly increased in ABR-depleted cells (Figure 2D). These observations raise the possibility that anomalous centrosome behaviors could be a mechanistic link between ABR dysfunction and G2-M accumulation. In support of this idea, it has been reported that RAC, a downstream small GTPase of ABR, modulates centrosome movement during G2-to-M progression in cultured epithelial cells (Woodcock et al., 2010, Whalley et al., 2015).
Figure 2.

ABR Controls Centrosome Dynamics
(A) Centrosomes are visualized by γTUBULIN staining (green). Mitosis or interphase is determined by chromosomal staining pattern and morphology (gray).
(B–E) Live imaging analyses of control or dox-treated tet-shABR cells expressing mVenus-CENT2. (B) Snapshots from Movie S1. The distance between centrosomes was measured with 2-min intervals. t = 0 corresponds to separation starting time, defined as a no-return point of bilateral movement. Arrows indicate NEB onsets. (D) The durations from separation initiation to NEB. The y axis corresponds to the red line-gated periods indicated in (C) control (n = 50) and dox-treated cells (n = 36) were analyzed. (E) The distance between centrosomes at the time of NEB. The y axis corresponds to blue line-gated lengths indicated in (C) control (n = 41) and dox-treated cells (n = 32) were analyzed.
The imaging experiments were performed three times. Scale bars represent 10 μm. Error bars in the graphs represent SD (D and E). Statistics: Student's t test (D and E); ∗∗∗p < 0.001 and ∗∗p < 0.01. See also Figure S2 and Movie S1.
Multiple Mitotic Failures in ABR-Depleted hESCs
A number of previous studies indicate that compromised centrosome separation often leads to severe failures in mitotic processes (Nam et al., 2015). To observe mitosis processes in cells with reduced ABR expression, we monitored cell cycle progression using tet-shABR hESCs expressing fluorescence protein-fused H2B (a marker for chromosomes), α-tubulin (TUBA, a marker for mitotic spindle) and Lifeact (a marker for actin filament) (Figures 3A–3C, Movie S2, part 1 and Figure S2D for a control experiment). Through these live imaging studies, we first found that a substantial number of dox-treated cells faced unrecoverable mitotic errors, including cell death or cytokinesis failures (Figures 3A, 3B and 3D; Movie S2, parts 2 and 3). Most of these cells had encountered problems in chromosomal alignment before these serious errors. Looking into these data more carefully, we also found that the majority of dox-treated cells struggled to align chromosomes at the central plane and spent significantly extended times before exiting from mitosis, even if they were finally able to divide (Figures 3C and 3E; Movie S2, part 4). In addition to these live imaging data, our immunostaining analyses using metaphase-arrested cells showed a high frequency of spindle malformation, which might arise from defects in centrosome separation, as well as misaligned chromosomes in dox-treated cells (Figures 3G and 3H).
Figure 3.

Multiple Mitotic Failures upon ABR Depletion
(A–E) Snap shots from Movie S2. Live imagings were performed using dox-treated tet-shABR cells expressing fluorescent protein-tagged H2B (chromosome, blue or gray), α-tubulin (TUBA, mitotic spindle, green) and LifeAct (F-actin, red). Examples of cytokinesis error (A), cell death (B), and extended mitosis (C) are shown. Yellow arrowheads indicate misaligned chromosomes. (D) The incidence of mitotic errors. Cellular behaviors were categorized into the indicated three groups. Control (n = 146) and dox-treated cells (n = 87) were analyzed. (E) The mitosis duration. In the cells that progressed into the next stage (classified as “division completion” in D), the time length from NEB to abscission was categorized into the indicated five groups. Control (n = 144) and dox-treated cells (n = 72) were analyzed.
(F) Immunostaining analyses on metaphase-arrested tet-shABR cells that were treated or untreated with dox for 3 days. Nuclei (gray or red) and TUBA (green) are shown. According to spindle morphology or chromosome positions, cellular phenotypes were classified into the indicated five categories.
(G and H) The incidence of spindle malformation (G) and chromosome misalignment (H). Control (n = 94) and dox-treated cells (n = 52) were analyzed.
The imaging experiments were repeated three times, and representative examples were shown (A, B, and C). The immunostaining was performed two times with three replicates in each experiment (F). Scale bars represent 10 μm. See also Figure S2 and Movie S2.
Thus, ABR-depleted cells encountered serious difficulties in chromosome alignment at metaphase, which delayed their transition into anaphase. This might be another cause for the accumulation of ABR-depleted cells in the G2-M stage (Figure 1B).
Chromosomal Missegregation and Aneuploidy in ABR-Depleted hESCs
Despite these troubles during prophase or metaphase, a large fraction of ABR-depleted cells did proceed to anaphase. This indicates that ABR is not absolutely required for hESCs to complete cell division. The extended period for metaphase-to-anaphase transition implies the activation of salvage mechanisms that serve as a backup when normal processes are disrupted (Musacchio, 2015). In these cases, however, we repeatedly observed lagging chromosomes during anaphase-to-telophase progression and a resultant micronucleus formation in the daughter cells (Figure 4A; Movie S3). Immunostaining analyses revealed that ABR depletion increased the incidence of these signs for chromosome missegregation (Figures 4B, 4C, and S3B; S3A shows typical staining patterns). From these data, we speculated that hESCs are able to bypass a mitotic necessity of ABR with the help of salvage mechanisms, but this process renders the cell susceptible to erroneous chromosome segregation.
Figure 4.

Chromosome Missegregation and Aneuploidy in ABR-Depleted Cells
(A) Snapshots from Movie S3. Yellow, magenta, and green arrowheads indicate misaligned chromosomes at metaphase, lagging chromosomes at anaphase, and micronuclei in daughter cells, respectively.
(B and C) Immunostaining analyses for lagging chromosomes. Two representatives from dox-treated samples are shown in (B) (nuclei, red; centromeres, green). Magenta arrowheads indicate centromere-positive lagging chromosomes. Mitotic cells with lagging chromosomes were counted and the incidence was shown in (C). Control (n = 315) and dox-treated cells (n = 318) were analyzed.
(D) Chromosome counting analyses. Mitotic spreads were prepared using tet-shABR cells that were treated or untreated with dox for 5 days. In each sample, 50 mitotic cells were subjected to counting.
(E) Multicolor fluorescence in situ hybridization (FISH) analyses. The dox-treated sample was stained with FISH probes for each chromosome. Two independent experiments were performed, and representative examples for normal and abnormal karyotypes are shown.
The immunostaining was repeated three times with five replicates in each experiment (B and C). The mitotic spreads for chromosome counting were prepared in three separate experiments (D). Scale bars, 10 μm. Error bars in the graphs represent SD. Statistics: Student's t test (C, n = 3); ∗∗p < 0.05. See also Figure S3 and Movie S3.
We postulated that such an error-prone situation would yield a selective pressure to facilitate the emergence of aneuploid cells. With this in mind, we carefully examined chromosome counts in the mitotic spreads that were prepared from cells treated with dox for 5 days, because at this time point most cells might undergo a few rounds of division in an ABR-independent way. Consistent with a previous report showing that the hESC line used here is stable in the karyotype during long-term culture (International Stem Cell Initiative et al., 2011), most of the control cells retained the normal number of chromosomes (Figure 4D). On the other hand, when cultured with reduced ABR expression, hESCs showed abnormal karyotypes with a higher frequency (Figures 4D and S3C). Some of them were tetraploid, which can result from mitosis skip or cytokinesis failure, and notably, others showed a gain or loss of some chromosomes (Figures 4E and S3C). Thus, ABR dysfunction actually elevated the risk of aneuploidy, highlighting a pivotal role of ABR in preventing aneuploidy in cultured hESCs.
Discussion
In this study, we explored ABR function in clump-cultured hESCs. We first noticed that ABR depletion impeded G2-to-M-to-G1 transitions. Deeper investigations at a single-cell level revealed that ABR-depleted cells struggled to complete a couple of mitotic steps, including centrosome separation at prophase and chromosome alignment at metaphase. These observations indicated that ABR has a crucial role in mitosis progression. Important information lacking now is subcellular localization of ABR. Our attempts to determine its localization in hESCs did not work well, but a large-scale proteomics analysis demonstrated ABR as a putative interactor of some centrosomal proteins (e.g., CEP25, Fogeron et al., 2013), supporting our conclusion.
ABR seems to play a safeguarding role in mitotic fidelity, in addition to being an apoptosis promoter in dissociated cells (Figure S3D), and these different outcomes upon ABR activation are dictated by the cellular adhesive state, dissociation versus clumping. A previous report demonstrated that the mitotic activation of actomyosin sometimes stimulated cell death, mirroring the dissociation-induced phenotype (Barbaric et al., 2014). Considering that cellular adhesiveness is dynamically rearranged during mitosis, spontaneous failures in the adhesion-mediated control of ABR activity could occur upon mitosis. An intriguing possibility is that mitotic cells in which ABR is inappropriately regulated might be intrinsically programmed to be eliminated, representing a mechanism restraining expansion of genetically abnormal cells. Consistently, it seems that ABR is not absolutely required for mitosis completion, but mitosis without ABR is an error-prone process leading to frequent chromosome missegregation. These results indicate that ABR sets a robust way for chromosome segregation in hESCs. This might be favorable, particularly to the self-renewing pluripotent stem cells in which the postmitotic checkpoint signaling is likely uncoupled to apoptosis-mediated elimination of genetically abnormal cells (Mantel et al., 2007).
How ABR participates in the control of mitotic fidelity remains an open question. Taking into consideration that ABR action is correlated with a cellular adhesive state, the present study suggests an unrecognized link between cell-cell contact and mitotic fidelity. In general, most types of non-transformed epithelial cells stop proliferation after forming a polarized layer in confluent culture, a phenomenon known as “contact inhibition of proliferation” (McClatchey and Yap, 2012). In this regard, hESCs are an atypical cell type: they can continue active growth within densely packed polarized colonies. We previously reported that the disconnection between cell-contact and nuclear function of transcriptional cofactors YAP/TAZ allows this type of unique proliferation (Ohgushi et al., 2015). From another viewpoint, however, this unique mode of proliferation yields complex mechanical fields for mitotic cells, because individual cells are constantly exposed to the pushing or pulling forces from contacting adjacent cells. On the basis of the observed high incidence of chromosome missegregation in ABR-depleted cells, our hypothesis is that ABR buffers the noisy mechanical cues within a multicellular society to confer robustness in the fidelity of chromosome segregation during long-term expansion of hESCs.
Unlike somatic cells in vivo, the proliferation of which is limited to several division cycles, ESCs and iPSCs undergo numerous rounds of genome replication and cell division to fulfill the quantitative demand for their practical applications. This raises concerns about the accumulation of genetic aberrations. Among them, aneuploidy is a particular threat since some types of aneuploidy confer survival or growth advantages that outcompete normal populations (Spits et al., 2008, Avery et al., 2013, Nguyen et al., 2014). Our findings provide implications for developing hESC culture methods that are better suited for human genetic studies and cell-based therapies.
Experimental Procedures
Cell Culture
All experiments using hESC lines were approved by an institutional ethics committee and done following the hESC guidelines of the Japanese government. Undifferentiated hESCs (KhES-1, Suemori et al., 2006) were cultured on feeder layers of mouse embryonic fibroblasts in D-MEM/F12 (Sigma) supplemented with 20% KnockOut serum replacement, 2 mM glutamine, 0.1 mM non-essential amino acids (Invitrogen), 5 ng/mL recombinant human basic fibroblast growth factor (Wako), and 0.1 μM 2-mercaptoethanol. The culture medium was refreshed daily until the next passage.
Immunostaining
Immunostaining was performed as described previously (Watanabe et al., 2007). For analyses of metaphase-arrested cells, cells were treated with 1 μg/mL MG132 for 1 hr and then immediately subjected to immunostaining.
Live Imaging
For live imaging, hESC clumps were seeded onto an MEF-coated 35-mm μ-dish (Ibidi). For confocal observations, serial images were collected using a CSU-W1 unit (Yokogawa) configured with an IX81-ZDC microscope (Olympus). The maximum projection image was constructed from the obtained slices using MetaMorph software.
Statistical Analyses
All experiments were performed at least three times, and error bars in the graphs represent SDs. Statistical significance was tested by Student's t test for two-group comparison, and by one-way ANOVA for multi-group comparison with Dunnett's test using Prime4 software (GraphPad).
Author Contributions
M.O. conceived the project, performed experiments, and wrote the manuscript. M.M. performed experiments. M.E. helped M.O. in imaging experiments. Y.S. supervised the project.
Acknowledgments
We are grateful to all members of the Sasai and Eiraku laboratory for support, discussion, and encouragement. M.O. expresses special thanks for his mentor Y.S. with tremendous respect to his legacy in science. This work was supported by grants-in-aid from MEXT (to M.E. and Y.S.) and by JSPS KAKENHI 26830088 (to M.O.).
Published: June 1, 2017
Footnotes
Supplemental Information includes Supplemental Experimental Procedures, three figures and three movies and can be found with this article online at http://dx.doi.org/10.1016/j.stemcr.2017.05.003.
Supplemental Information
{kind=link}
{kind=link}
{kind=link}
References
- Avery S., Hirst A.J., Baker D., Lim C.Y., Alagaratnam S., Skotheim R.I., Lothe R.A., Pera M.F., Colman A., Robson P. BCL-XL mediates the strong selective advantage of a 20q11.21 amplification commonly found in human embryonic stem cell cultures. Stem Cell Rep. 2013;1:379–386. doi: 10.1016/j.stemcr.2013.10.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barbaric I., Biga V., Gokhale P.J., Jones M., Stavish D., Glen A., Coca D., Andrews P.W. Time-lapse analysis of human embryonic stem cells reveals multiple bottlenecks restricting colony formation and their relief upon culture adaptation. Stem Cell Rep. 2014;3:142–155. doi: 10.1016/j.stemcr.2014.05.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ben-David U., Arad G., Weissbein U., Mandefro B., Maimon A., Golan-Lev T., Narwani K., Clark A.T., Andrews P.W., Benvenisty N. Aneuploidy induces profound changes in gene expression, proliferation and tumorigenicity of human pluripotent stem cells. Nat. Commun. 2014;5:4825. doi: 10.1038/ncomms5825. [DOI] [PubMed] [Google Scholar]
- Boward B., Wu T., Dalton S. Control of cell fate through cell cycle and pluripotency networks. Stem Cells. 2016;34:1427–1436. doi: 10.1002/stem.2345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chuang T.H., Xu X., Kaartinen V., Heisterkamp N., Groffen J., Bokoch G.M. Abr and Bcr are multifunctional regulators of the Rho GTP-binding protein family. Proc. Natl. Acad. Sci. USA. 1995;92:10282–10286. doi: 10.1073/pnas.92.22.10282. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fogeron M.L., Müller H., Schade S., Dreher F., Lehmann V., Kühnel A., Scholz A.K., Kashofer K., Zerck A., Fauler B. LGALS3BP regulates centriole biogenesis and centrosome hypertrophy in cancer cells. Nat. Commun. 2013;4:1531. doi: 10.1038/ncomms2517. [DOI] [PubMed] [Google Scholar]
- Heisterkamp N., Kaartinen V., van Soest S., Bokoch G.M., Groffen J. Human ABR encodes a protein with GAPrac activity and homology to the DBL nucleotide exchange factor domain. J. Biol. Chem. 1993;268:16903–16906. [PubMed] [Google Scholar]
- International Stem Cell Initiative, Amps K., Andrews P.W., Anyfantis G., Armstrong L., Avery S., Baharvand H., Baker J., Baker D., Munoz M.B. Screening ethnically diverse human embryonic stem cells identifies a chromosome 20 minimal amplicon conferring growth advantage. Nat. Biotechnol. 2011;29:1132–1144. doi: 10.1038/nbt.2051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jacobs K., Zambelli F., Mertzanidou A., Smolders I., Geens M., Nguyen H.T., Barbé L., Sermon K., Spits C. Higher-density culture in human embryonic stem cells results in DNA damage and genome instability. Stem Cell Rep. 2016;6:330–341. doi: 10.1016/j.stemcr.2016.01.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lamm N., Ben-David U., Golan-Lev T., Storchová Z., Benvenisty N., Kerem B. Genomic instability in human pluripotent stem cells arises from replicative stress and chromosome condensation defects. Cell Stem Cell. 2016;18:253–261. doi: 10.1016/j.stem.2015.11.003. [DOI] [PubMed] [Google Scholar]
- Li L., Wang B.H., Wang S., Moalim-Nour L., Mohib K., Lohnes D., Wang L. Individual cell movement, asymmetric colony expansion, rho-associated kinase, and E-cadherin impact the clonogenicity of human embryonic stem cells. Biophys. J. 2010;98:2442–2451. doi: 10.1016/j.bpj.2010.02.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mantel C., Guo Y., Lee M.R., Kim M.K., Han M.K., Shibayama H., Fukuda S., Yoder M.C., Pelus L.M., Kim K.S. Checkpoint-apoptosis uncoupling in human and mouse embryonic stem cells: a source of karyotpic instability. Blood. 2007;109:4518–4527. doi: 10.1182/blood-2006-10-054247. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mayshar Y., Ben-David U., Lavon N., Biancotti J.C., Yakir B., Clark A.T., Plath K., Lowry W.E., Benvenisty N. Identification and classification of chromosomal aberrations in human induced pluripotent stem cells. Cell Stem Cell. 2010;7:521–531. doi: 10.1016/j.stem.2010.07.017. [DOI] [PubMed] [Google Scholar]
- McClatchey A.I., Yap A.S. Contact inhibition (of proliferation) redux. Curr. Opin. Cell Biol. 2012;24:685–694. doi: 10.1016/j.ceb.2012.06.009. [DOI] [PubMed] [Google Scholar]
- Musacchio A. The molecular biology of spindle assembly checkpoint signaling dynamics. Curr. Biol. 2015;25:R1002–R1018. doi: 10.1016/j.cub.2015.08.051. [DOI] [PubMed] [Google Scholar]
- Nam H.J., Naylor R.M., van Deursen J.M. Centrosome dynamics as a source of chromosomal instability. Trends Cell Biol. 2015;25:65–73. doi: 10.1016/j.tcb.2014.10.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nguyen H.T., Geens M., Mertzanidou A., Jacobs K., Heirman C., Breckpot K., Spits C. Gain of 20q11.21 in human embryonic stem cells improves cell survival by increased expression of Bcl-xL. Mol. Hum. Reprod. 2014;2:168–177. doi: 10.1093/molehr/gat077. [DOI] [PubMed] [Google Scholar]
- Nichols J., Smith A. Pluripotency in the embryo and in culture. Cold Spring Harb. Perspect. Biol. 2012;4:a008128. doi: 10.1101/cshperspect.a008128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ohgushi M., Matsumura M., Eiraku M., Murakami K., Aramaki T., Nishiyama A., Muguruma K., Nakano T., Suga H., Ueno M. Molecular pathway and cell state responsible for dissociation-induced apoptosis in human pluripotent stem cells. Cell Stem Cell. 2010;7:225–239. doi: 10.1016/j.stem.2010.06.018. [DOI] [PubMed] [Google Scholar]
- Ohgushi M., Minaguchi M., Sasai Y. Rho-signaling-directed YAP/TAZ activity underlies the log-term survival and expansion of human embryonic stem cells. Cell Stem Cell. 2015;17:448–461. doi: 10.1016/j.stem.2015.07.009. [DOI] [PubMed] [Google Scholar]
- Peterson S.E., Loring J.F. Genomic instability in pluripotent stem cells: implications for clinical applications. J. Biol. Chem. 2014;289:4578–4584. doi: 10.1074/jbc.R113.516419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sakaue-Sawano A., Kurokawa H., Morimura T., Hanyu A., Hama H., Osawa H., Kashiwagi S., Fukami K., Miyata T., Miyoshi H. Visualizing spatiotemporal dynamics of multicellular cell-cycle progression. Cell. 2008;132:487–498. doi: 10.1016/j.cell.2007.12.033. [DOI] [PubMed] [Google Scholar]
- Santaguida S., Amon A. Short- and long-term effects of chromosome mis-segregation and aneuploidy. Nat. Rev. Mol. Cell Biol. 2015;16:473–485. doi: 10.1038/nrm4025. [DOI] [PubMed] [Google Scholar]
- Spits C., Mateizel I., Geens M., Mertzanidou A., Staessen C., Vandeskelde Y., Van der Elst J., Liebaers I., Sermon K. Recurrent chromosomal abnormalities in human embryonic stem cells. Nat. Biotechnol. 2008;26:1361–1363. doi: 10.1038/nbt.1510. [DOI] [PubMed] [Google Scholar]
- Suemori H., Yasuchika K., Hasegawa K., Fujioka T., Tsuneyoshi N., Nakatsuji N. Efficient establishment of human embryonic stem cell lines and long-term maintenance with stable karyotype by enzymatic bulk passage. Biochem. Biophys. Res. Commun. 2006;345:926–932. doi: 10.1016/j.bbrc.2006.04.135. [DOI] [PubMed] [Google Scholar]
- Taapken S.M., Nisler B.S., Newton M.A., Sampsell-Barron T.L., Leonhard K.A., McIntire E.M., Montgomery K.D. Karotypic abnormalities in human induced pluripotent stem cells and embryonic stem cells. Nat. Biotechnol. 2011;29:313–314. doi: 10.1038/nbt.1835. [DOI] [PubMed] [Google Scholar]
- Tanenbaum M.E., Medema R.H. Mechanisms of centrosome separation and bipolar spindle assembly. Dev. Cell. 2010;19:797–806. doi: 10.1016/j.devcel.2010.11.011. [DOI] [PubMed] [Google Scholar]
- Watanabe K., Ueno M., Kamiya D., Nishiyama A., Matsumura M., Wataya T., Takahashi J.B., Nishikawa S., Nishikawa S., Muguruma K. A ROCK inhibitor permits survival of dissociated human embryonic stem cells. Nat. Biotechnol. 2007;25:681–686. doi: 10.1038/nbt1310. [DOI] [PubMed] [Google Scholar]
- Whalley H.J., Porter A.P., Diamantopoulou Z., White G.R., Castañeda-Saucedo E., Malliri A. Cdk1 phosphorylates the Rac activator Tiam1 to activate centrosomal Pak and promote mitotic spindle formation. Nat. Commun. 2015;6:7437. doi: 10.1038/ncomms8437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Woodcock S.A., Rushton H.J., Castañeda-Saucedo E., Myant K., White G.R., Blyth K., Sansom O.J., Malliri A. Tiam1-Rac signaling counteracts Eg5 during bipolar spindle assembly to facilitate chromosome congression. Curr. Biol. 2010;20:669–675. doi: 10.1016/j.cub.2010.02.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang M., Cheng L., Jia Y., Liu G., Li C., Song S., Bradley A., Huang Y. Aneuploid embryonic stem cells exhibit impaired differentiation and increased neoplastic potential. EMBO J. 2016;35:2285–2300. doi: 10.15252/embj.201593103. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao B., Zhang W.D., Duan Y.L., Lu Y.Q., Cun Y.X., Li C.H., Guo K., Nie W.H., Li L., Zhang R., Zheng P. Filia is an ESC-specific regulator of DNA damage response and safeguards genomic stability. Cell Stem Cell. 2015;16:684–698. doi: 10.1016/j.stem.2015.03.017. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.