

Abstract
Oxidized phospholipids are products of lipid oxidation that are found on oxidized low-density lipoproteins and apoptotic cell membranes. These biologically active lipids were shown to affect a variety of cell types and attributed pro-as well as anti-inflammatory effects. In particular, macrophages exposed to oxidized phospholipids drastically change their gene expression pattern and function. These ‘Mox,’macrophages were identified in atherosclerotic lesions, however, it remains unclear how lipid oxidation products are sensed by macrophages and how they influence their biological function. Here, we review recent developments in the field that provide insight into the structure, recognition, and downstream signaling of oxidized phospholipids in macrophages.
Keywords: Oxidized phospholipids, macrophages, inflammation
Introduction
Since the discovery of lipid oxidation products as biologically active compounds by Hermann Esterbauer[1–3], the presence of oxidized lipids has been confirmed in a variety of tissues that are either chronically inflamed or oxidatively damaged. Lipid oxidation is associated with the pathogenesis of various diseases including atherosclerosis, diabetes, cancer, Alzheimer’s disease, rheumatoid arthritis, and the pathophysiology of aging. In all of these settings, macrophages play a key role in controlling the initiation and progression of the disease. Phospholipids that contain unsaturated fatty acids are major constituents of cell membranes and lipoproteins and are particularly prone to oxidative modification, which results in the formation of a large variety of biologically active compounds. Oxidized phospholipids (OxPL) derived from oxidation of lipoproteins or from apoptotic cell membranes were shown to accumulate at sites of chronic inflammation and oxidative tissue damage, where they not only affect biophysical properties of membranes, but also can be considered endogenous danger associated molecular patterns (DAMPs) that modulate immune responses (reviewed in [4–7]). After the identification of OxPL as biologically active components of minimally oxidized LDL[8, 9], OxPL have been ascribed pro- as well as anti-inflammatory properties, depending on their structural features and functional groups, which also determine their recognition by extracellular or intracellular receptors. The effects of OxPL on modulation of inflammatory responses have been extensively reviewed[10–14], however, little is known about the mechanisms that control the formation and degradation of OxPL in different cell types. Moreover, the effects oxidized lipids have on cells of the immune system are poorly understood. OxPL are recognized by macrophages via various mechanisms involving intra- and extracellular receptors, eliciting a series of responses that result in context-specific phenotypic adaptation. Here, we will summarize recent findings that illustrate how macrophages and other cells recognize and respond to OxPL and we will discuss the functional consequences of these responses for inflammatory control and tissue homeostasis.
Structures and biological functions of oxidized phospholipids
Oxidative modification of lipids is mediated by free radical-induced mechanisms involving enzymes such as NADPH oxidase and myeloperoxidase. Thus, at sites of inflammation, where neutrophils and other cells generate an environment of high oxidative stress, lipid oxidation products accumulate and exert a variety of biological activities. Polyunsaturated fatty acids and especially arachidonic acid are highly susceptible to lipid peroxidation, which leads to the generation of lipid hydroperoxides, which can undergo carbon-carbon bond cleavage giving rise to the formation of short chain, unesterified aldehydes and aldehydes still esterified to the parent lipid, termed core-aldehydes. It has been shown that activation of the NADPH oxidase during apoptosis leads to oxidation of the membrane phosphatidylserine (PS), but also phosphatidylcholine (PC) and phosphatidylethanolamine (PE)[15]. The presence of oxidized PC (OxPC) on the surface of apoptotic cells has been demonstrated using the monoclonal antibody EO6, which exclusively binds to OxPC[16–19]. Furthermore, enzymatic oxidation of phospholipids involving 12/15 lipoxygenase produces biologically active mediators[20, 21]. The cholesteryl esters found within LDL are also subject to oxidative modification, which may contribute to endothelial activation[22]. Miller et al. found that oxidized cholesteryl esters (OxCE) use toll-like receptor 4 (TLR4) and spleen tyrosine kinase (Syk) to induce a pro-inflammatory response in macrophages[23].
Initially, three biologically active phospholipids present in minimally modified low-density lipoprotein (mm-LDL), which were derived from oxidation of 1-palmitoyl-2-arachidonoyl-sn-glycero-3-phosphorylcholine (PAPC), were structurally identified: 1-palmitoyl-2-oxovaleroyl-sn-glycero-3-phosphorylcholine (POVPC) and 1-palmitoyl-2-glutaroyl-sn-glycero-3-phosphorylcholine (PGPC), and 1-palmitoyl-2-(5,6-epoxyisoprostane)-PC (PEIPC)[8, 24]. These lipids were shown to activate endothelial cells in a structure-specific manner[25, 26]. Furthermore, a group of CD36-activating truncated oxidized phospholipids was described by Podrez et al. These species include an sn-2 acyl group that requires a γ-hydroxy-α,β-unsaturated carbonyl or a γ-oxo-α,β-unsaturated carbonyl and were found to be generated during LDL oxidation[27]. These lipids, collectively referred to as oxPCCD36, were found to directly contribute to the development of foam cell formation in macrophages[28]. In this study the authors describe that even trace amounts of oxPCCD36 are enough to induce CD36-dependent binding and uptake of LDL[28]. Accumulating evidence suggests that biologically active OxPL that are present in atherogenic lipoproteins, in atherosclerotic lesions, and in membrane vesicles released from activated and apoptotic cells play an essential role in the development of atherosclerosis [14, 29, 30]. Indeed, OxPL affect a variety of vascular cell types, including endothelial [31–36] and smooth muscle [37–39] cells. Furthermore, OxPL have potent effects on immune cells. Studies performed by our lab have shown that phospholipid oxidation products of PAPC (OxPAPC) inhibit basic steps of the classical dendritic cell (DC) maturation process[7, 40]. Inhibitory effects on DC, anergic effects on T-cells as well as on the formation on Treg cells have been reported[41, 42], and one report demonstrates an important role for OxPL on epigenetic modulation of DC activation[41].
Levuglandins/ketoaldehydes are derived from oxidation of arachidonic acid and contain highly reactive aldehydes, which readily form lysyl adducts with proteins[43]. Macrophages recognize LDL that has been modified with levuglandins[44], and free levuglandins as well as protein-adducts have been demonstrated at sites of inflammation and oxidative stress, including atherosclerotic lesions and kidneys of patients with end-stage renal disease[45, 46]. However, the exact role of levuglandins/ketoaldehydes in contribution to disease progression remains to be elucidated. Given the ability of levuglandins/ketoaldehydes to react with primary amines, it was postulated that levuglandins might also react with phosphatidylethanolamines. Indeed, it was shown that γ-ketoaldehydes are able to react with ethanolamine at a much faster rate than with lysine[47, 48] to produce compounds that induce inflammatory reactions in macrophages[49]. The γ-ketoaldehyde/levuglandin modification of phosphatidylethanolamines produces a highly cytotoxic phosphatidylethanolamine[50], and using mouse models of hypertension, it was shown that scavenging γ-ketoaldehydes prevented immune-mediated hypertension and associated comorbidities[51]. Another modification of phosphatidylethanolamines has been described to result in the formation N-acyl-phosphatidylethanolamines (NAPEs), which are precursors to N-acyl-ethanolamines (NAEs), thought to be highly potent satiety signals generated in the intestines[52]. Recently, a derivative of carboxyalkylpyrrole(CAP)-phosphatidylethanolamine (CAP-PE) has been identified in the plasma of ApoE−/− mice[53]. These CAP-PE derivatives directly bind and activate TLR2/1 to induce platelet activation[53]. Similarly to the phosphatidylethanolamine adducts, the CAP-protein derivatives, such as those formed by ω-(2-carboxyethyl)-pyrrole (CEP), have also been shown to induce platelet activation, but even more startlingly, they induce VEGF receptor-independent angiogenesis through TLR2 activation[54]. Moreover, the CEPs have been extensively described for their role as inducers of inflammation in age-related macular degeneration[55].
OxPL and macrophages: Identification of Mox
Macrophages sense changes in their tissue microenvironment and respond by changing their phenotype and function accordingly. In chronically inflamed tissues, the formation of OxPL may contribute to the initiation of the macrophage-dependent inflammatory process, but also to the perpetuation of inflammation, possibly by interfering with inflammatory resolution. Several studies have reported a variety of cellular responses to an oxidatively damaged microenvironment by analyzing the reaction of macrophages to OxPL[56–59]. Of special interest, it has been noted that the profiles of inflammatory gene expression and recruitment of monocytes and macrophages induced by oxidized lipids is remarkably similar to the “low-grade inflammation” seen in settings of diet-induced obesity or atherosclerosis, and generally in metabolic tissue damage[60, 61]. Furthermore, we have shown that OxPL induce specific macrophage recruitment in the air-pouch model of inflammation, by a mechanism involving CCR2[62].
A critical event leading to the resolution of inflammation is the removal of apoptotic cells[63, 64]. Delayed apoptosis results in increased oxidative stress and diminished phagocytosis, and may be associated with the prolongation and persistence of inflammatory disorders[65, 66]. These observations strongly indicate that improved tissue remodelling could be a direct consequence of increased engulfment capacity of macrophages. Taking into consideration that apoptosis is essential for successful resolution of acute inflammation, OxPL generated during apoptosis could influence this process, by upsetting the balance of macrophage phenotypes. In this context, OxPL were shown to be important recognition signals on apoptotic cells[18], which could facilitate phagocytosis by macrophages[15, 67]. The Tabas group showed that OxPL induce apoptosis in macrophages undergoing ER-stress via CD36/TLR2 activation[68] and Gerhard Kronke’s group reported that 12/15 lipoxygenase plays a major and unexpected role in the non-inflammatory clearance of apoptotic cells, via a mechanism involving the formation of oxidized phosphatidylethanolamines (OxPE)[20]. On the other hand, phagocytosis was shown to be inhibited in macrophages by mm-LDL[69] and OxPAPC[58], and Sylvia Knapp’s group discovered that this inhibition involves WAVE1[70, 71], which is a cytoskeleton bound A-kinase anchoring protein. The group showed that WAVE1 knockout macrophages are resistant to OxPAPC-inhibited phagocytosis, and that the E. coli infected chimeric WAVE1-knockout mice have a better survival rate compared to controls[71]. Altogether, this has important implications in the regulation and outcome of inflammatory responses in atherosclerosis and sepsis.
In chronically inflamed tissues, where macrophages are constantly exposed to high levels of oxidized lipids, macrophages survive for surprisingly long periods of time, despite the toxic environment[72]. This implies the upregulation of survival mechanisms, such as antioxidant pathways and phase II detoxification genes. We have previously shown that UV-light-induced oxidation of phospholipids have protective effects on skin cells, via induction of redox-sensitive transcriptional programs involving activation of nuclear factor (erythroid-derived 2)-like 2 (Nrf2)[73, 74]. In atherosclerosis, the intra-plaque milieu is complex, leading to a remarkably diverse macrophage population[75]. Within an atherosclerotic plaque, lipid mediators ranging from cholesterol crystals to OxPL activate the inflammasome and pattern recognition receptors, as well as other receptors to induce both pro- and anti-inflammatory phenotypic polarizations (M1 and M2)[76]. Our group characterized a novel OxPL-induced macrophage phenotype, “Mox”, in murine atherosclerotic lesions[58]. The Mox phenotype significantly differs from pro-inflammatory “M1” and anti-inflammatory “M2” macrophages and is characterized by an Nrf2-dependent gene expression pattern[58]. Nrf2-dependent gene expression is induced by OxPL in the vasculature and especially in endothelial cells[77, 78] where it is believed to exert protective effects. However, it was also shown that Nrf2 deficiency in macrophages had differing effects on atherosclerotic lesion formation[79–81] and we have shown that Nrf2 deficiency in macrophages did not protect against HFD-induced insulin resistance[82]. Nrf2 in macrophages was shown to exert direct anti-inflammatory effects[83] and an anti-inflammatory, reparative function in the brain was ascribed to Mox macrophages detected in microglia after stimulation of nicotinic receptors[84]. Of note, a recent report demonstrated regulation of chronic inflammatory pain by OxPAPC in neurons via activation of TRPA1 channels[85].
Interestingly, based on their specific gene expression profile, Mox macrophages have been described to play a role in iron metabolism, favouring iron storage due to compromised iron release[86]. These and other observations also indicate that Mox contribute to inflammation. We have shown that OxPL induce a low level of inflammatory gene expression through a TLR2-dependent mechanism[87]. Moreover, we were able to show that Mox macrophages secrete IL-1β[58]. In macrophages, the events leading up to IL-1β secretion involves the transcription of pro-IL-1β, activation of the NLRP3 inflammasome, which constitutes an activated caspase-1, which then cleaves pro-IL-1β. However, the endogenously formed activators of IL-1β production in chronically inflamed tissue are not known. In settings of ‘sterile inflammation,’ such as in obesity and atherosclerosis, the underlying question whether products of lipid oxidation could evoke an inflammatory response via inflammasome-dependent cytokine production remains unanswered. In this regard, OxPL were shown to induce NLRP3 inflammasome activation in macrophages[88] and Jonathan Kagan’s group recently showed that endogenously formed OxPAPC is able to directly activate caspase 11, thereby activating the NLRP3 inflammasome in a non-canonical fashion in dendritic cells[89]. These findings could have important implications for a variety of diseases where caspase 11 activation and inflammasome-dependent activation of the immune response has been reported[90].
The cellular metabolic programming, or bioenergetics, of macrophages has been linked to a macrophage’s inflammatory phenotype: pro-inflammatory “M1” macrophages rely upon glycolysis for their bioenergetic needs while anti-inflammatory “M2” macrophages rely on oxidative phosphorylation[91–95]. Recently, multiple studies identified the importance of cellular metabolism in the determination of macrophage pro-[96] or anti-inflammatory[97] function. Further investigation revealed that the mitochondrial dysfunction caused by pro-inflammatory stimuli prevented macrophages from assuming an anti-inflammatory polarization state[98].
However, the metabolic profile of Mox has not been examined. Furthermore, the mechanisms by which OxPL induce inflammatory signaling and metabolic changes in Mox remain poorly defined. OxPL can induce ceramide accumulation in macrophages[99], and since mitochondrial damage can be mediated by ceramides, Mox macrophages may have a compromised bioenergetic profile. Activation of TLR2 signaling by OxPL may contribute to the low-grade chronic inflammation, and ceramide synthesis was shown to be induced by TLR2 stimulation[100]. TLR2 is known to hetero-dimerize with either TLR1 or TLR6, which collaborate with OxPL receptors CD14 or CD36 respectively[68, 101, 102]. Together, these studies indicate that OxPL may affect macrophage bioenergetics via mechanisms involving TLR2-dependent ceramide production.
How are OxPL recognized?
Among a number of recently characterized danger signals, phospholipid oxidation products accumulate under conditions of increased oxidative stress and cell death. They serve as indicators of inflammation-induced tissue damage and were shown to act as endogenous regulators of the innate immune response[4]. For instance, diet-induced oxidative stress is a major cause for vascular, liver and adipose tissue damage, hallmarks of atherosclerosis, fatty liver disease and insulin resistance and diabetes[103]. Disturbed redox balance due to chronic inflammation in neurological disorders may contribute to formation and accumulation of OxPL in the brain[104]. Nevertheless, how the recognition of an altered tissue microenvironment by immune cells and the mechanism by which oxidative damage ultimately translates into an inflammatory reaction is not clear.
Based on the current literature, OxPL may bind to and initiate a response through soluble acceptors in the blood plasma, by membrane-bound receptors, or by intracellular sensor proteins. Depending on their functional reactive groups, they are able to either covalently modify these receptor proteins, or possibly bind to and interact in a reversible manner. In inflamed tissue, OxPL can either act pro-inflammatory or anti-inflammatory, and for most OxPL whose biological functions have been investigated, dual pro and anti-inflammatory effects were described. So far, OxPL have been documented to directly bind to LPS binding protein (LBP), CD14, MD-2, and CD36, which may lead to activation or inhibition of TLR4 or TLR2 signaling. Intracellularly, OxPL interact with caspase 11 or Keap1/Nrf2, exerting pro- or anti-inflammatory effects, respectively. Here we summarize the evidence for the various systems that recognize and respond to OxPL. Table 1 catalogs the evidence for OxPL effects as distinguished by phospholipid class (i.e. PC, PE, PS, PG, PA, and PI). It is important to keep in mind that many studies were performed with mixtures, which contain a large number of diverse oxidized moieties, making it difficult to conclude which oxidized moieties are tied to which downstream consequences. Nonetheless, the studies that include enriched, purified, or synthetic OxPL species are included in Table 2.
Table 1.
Evidence for oxidized phospholipid recognition sorted by head group
| Oxidized Phospholipid Class | Head Group | Structure (X) | Activation | Inhibition |
|---|---|---|---|---|
| OxPC | Choline |
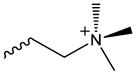
|
CD36[120], Nrf2[77, 78], Casp11/NLRP3[89], TLR2[87] | TLR2[40, 111], TLR4[111, 112], CD14/MD2[111], LBP[101] |
| OxPE | Ethanolamine |
|
CD36[67, 78, 118], Nrf2[78] | TLR2[111], TLR4[110, 111], CD14[101], LBP[101] |
| OxPS | Serine |
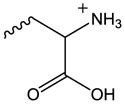
|
CD36[67, 78], Nrf2[78] | CD14[101], LBP[101] |
| OxPA | Phosphatidic Acid |
|
Nrf2[78] | LBP[101] |
| OxPG | Glycerol |
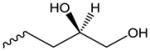
|
Nrf2[78] | N/A |
| OxPI | Inositol |
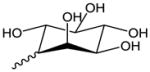
|
N/A | N/A |
Table 2.
Catalog of oxidized phospholipids by epitope
| General Phospholipid Structure |
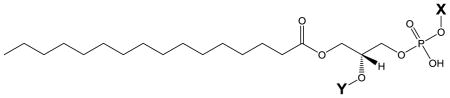
|
||
|---|---|---|---|
| sn-2 Moiety | Structure (Y) | Abbreviation (C16:0 in sn-1) | Associated Head Group (X) |
| Arachidonoyl |
|
PA- | PC, PE, PS, PA, PG, PI |
| Oxygenation | |||
| Hydroxy- arachidonoyl |
|
PA(X)-OH or HETE-(X) | PC[78, 156–158], PE[156–158], PS |
| Hydroperoxy- arachidonoyl |
|
PA(X)-OOH or HPETE-(X) | PC[78], PE, PS |
| 5,6-Epoxyisoprostane E2 |
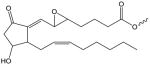
|
PEI-(X) | PC[24, 78, 89] |
| 5,6- Epoxycyclopentenone |
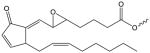
|
PEC-(X) | PC[119, 159] |
| 15-deoxy-Δ12,14- Prostaglandin J2 |
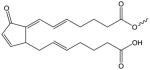
|
15d-PGJ2-(X) | PC[119, 160] |
| F2-Isoprostane |
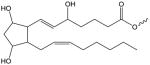
|
F2-IP-(X) | PC[161] |
| H2-Endoperoxide Isoprostane |
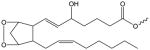
|
H2-IP-(X) | N/A |
| E2-Isoprostane |
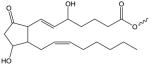
|
E2-IP-(X) | N/A |
| D2-Isoprostane |
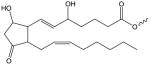
|
D2-IP-(X) | N/A |
| E2-Isolevuglandin |
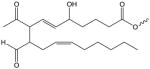
|
E2-IL-(X) | PC[162, 163] |
| Chain Fragmentation | |||
| Glutaroyl |
|
PG-(X) | PC[8, 78] [25, 26, 76, 136, 164], PE, PS |
| Azelaoyl |
|
PAze-(X) or PAz- (X) | PC[78, 165, 166], PE, PS |
| 5-Oxovaleroyl |
|
POV-(X) | PC[8, 26, 78, 164], PE, PS |
| 9-Oxononanoyl |
|
PON-(X) | PC[16, 164], PE, PS |
| 5-Hydroxy-8-oxo-6- octenoyl |
|
HOOA-(X) | PC[27, 28], PS[67] |
| 9-Hydroxy-12-oxo- 10-dodecenoyl |
|
HODA-(X) | PC[27, 28], PS[67] |
| 5-Hydroxy-8-oxo-6- octendioyl |
|
HOdiA-(X) | PC[27, 28], PS[67] |
| 9-Hydroxy-10- dodecenedioyl |
|
HDdiA-(X) | PC[27, 28], PS[67] |
| 5-Keto-8-oxo-6- octenoyl |
|
KOOA-(X) | PC[27, 28], PS[67] |
| 9-Keto-12-oxo-10- dodecenyl |
|
KODA-(X) | PC[27, 28], PS[67] |
| 5-Keto-6-octendioyl |
|
KOdiA-(X) | PC[27, 28], PS[67] |
| 9-Keto-10- dodecenedioyl |
|
KDdiA-(X) | PC[27, 28], PS[67] |
| Ester Hydrolysis | |||
| Hydroxy |
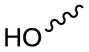
|
Lyso-(X) | PC[126], PE[167], PS[127] |
OxPL modify proteins at the cell membrane and in the cytosol
Using cultured RAW 264.7 macrophages and fluorescently labeled POVPC or PGPC Hermetter’s group identified primary protein targets of these phospholipids by mass spectrometry[105]. They showed that the aldehyde-containing POVPC binds to proteins in the cell membrane by covalently reacting with amino groups of proteins. Interestingly, POVPC was exchangeable from lipoproteins to cells, where only a selective group of proteins was modified by labeled POVPC. PGPC on the other hand, freely travelled through the cell membrane into the cytosol[105]. Berliner’s group took a different approach to identify the proteome that is modified by OxPL in endothelial cells, using biotin as affinity tag at the polar head group of the phospholipids[106]. They then showed that the interaction of OxPL with proteins involves modification of cysteines[107].
OxPL are DAMPS that are recognized by pattern recognition receptors
Upon oxidative modification, phospholipids structurally resemble DAMPs that are recognized by pattern recognition receptors (PRRs)[5]. Sensing of these endogenously formed danger or “altered-self” molecules by the innate immune system is mediated by immune-modulating and scavenger receptors such as CD36, TLRs, CD14, LBP, and C-reactive protein[108, 109]. The fact that the oxidation process renders phospholipids “visible” to the innate immune system indicates a crucial role for phospholipid oxidation products in the pathogenesis of both chronic inflammatory and autoimmune diseases[4].
First indications that OxPL bear similarities with compounds that can be recognized by TLRs came from our studies showing that these lipids potently inhibit bacterial ligand-induced TLR4 activation[102]. Our group was first to show that OxPAPC inhibited LPS-induced NF-κB-mediated inflammatory gene expression in human umbilical-vein endothelial cells (HUVEC) and in LPS-injected mice protecting mice from lethal endotoxin shock[102]. We further demonstrated that OxPAPC bound to LPS-binding protein (LBP) and CD14, which blocked the interaction of LPS with both proteins in vitro[102]. These results were supported by others[101, 110–112], showing that OxPAPC can bind to CD14 and MD2 to interfere with LPS-TLR4/MD2 activation. Moreover, OxPAPC inhibited N-palmitoyl-S-dipalmitoylglyceryl-Cys-Scr-(lys)4 (Pam3CSK4)-induced TLR2 activation, which was reversed by serum and sCD14, suggesting that CD14 is one of the targets of OxPAPC.
Oxidative modification of phospholipids provides epitopes for the adaptive immune system
The patterns that are generated during oxidation of phospholipids are also recognized by the humoral part of the adaptive immune system. Both IgG and IgM antibodies directed against oxidized low-density lipoprotein (oxLDL) are present in the plasma of humans and animals and their titers have been shown to correlate with atherosclerosis progression and measures of lipid peroxidation[113, 114], as well as in several autoimmune disorders such as systemic lupus erythematosus and rheumatoid arthritis[115]. Detailed studies in ApoE-deficient mice, which show increased levels of oxLDL, led to the cloning of a set of abundant monoclonal IgM antibodies directed against oxLDL, which includes the prototypic EO6 antibody that specifically binds to OxPL on the surface of oxLDL and apoptotic cells[17, 116, 117]. This antibody recognizes the PC headgroup, which is exposed after conformational changes following the oxidation of the phospholipid[118].
OxPL bind to soluble immune receptors
CD14
CD14 is a co-receptor for both TLR2 and TLR4 that has been shown to exist in both a membrane-bound and soluble form. CD14 has been shown to enhance the sensitivity of TLR2 activation to Pam3CSK4, and is essential for LPS-induced activation of TLR4. As shown by Erridge et al., OxPAPC, in particular the components PGPC and POVPC, are able to potently inhibit Pam3CSK4 activation of TLR2, in a HEK cell model with TLR2-overexpression[111]. The authors concluded that this inhibition was likely due to the binding of OxPL directly to CD14, due to the rescue of the inhibition upon supplementation with soluble CD14. Furthermore, Valery Bochkov’s group found that OxPL mixtures with the head groups PE, PS, and PA induced a mobility shift in soluble CD14, indicating a direct binding interaction of these phospholipids to CD14[101]. OxPL with the PC head group also bind to CD14, but do not cause a mobility shift due to the overall neutral charge of the complex. The structural requirements for OxPL binding to CD14 require further investigation, so far there is experimental evidence for PECPC, PEIPC, OxPAPE, OxPAPC, and OxPAPS to interact with CD14[101, 111, 119] (Table 2). Walton et al. ruled out an involvement of CD14 in transmitting pro-inflammatory effects of OxPAPC on endothelial cells, instead identifying another GPI-anchored protein recognizing OxPL[120].
LPS binding protein
OxPAPC and a number of purified components of OxPAPC are able to directly bind to LBP and inhibit LPS action[101, 102, 111, 112, 121]. Unlike for CD14, it was found that oxidized phospholipids with any head group (PC, PE, PS, and PA) were able to inhibit the binding of LPS to LBP (Table 1). Eighteen total OxPL have been documented to interact with LBP, out of the twenty-four investigated. This implies that there is little chemical specificity in the interaction, and that for the most part, oxidation of the phospholipid is the only prerequisite for inhibitory interaction with LBP. It has never been determined whether LBP is required for OxPL-induced inflammation.
MD2
Another study found that the oxidized phospholipid KOdiA-PC directly inhibits the binding of LPS to MD2, providing yet another mechanism by which OxPL are able to inhibit LPS action[122]. This was supported by other studies using both binding experiments and functional assays to indicate that OxPAPC can compete with LPS to bind MD2[111]. The structural requirements of OxPL needed to activate or inhibit MD2-dependent signaling remain unknown. Taken together, these findings indicate that OxPL can compete with serum-soluble factors to inhibit LPS, and likely other bacterial or viral components, which induce systemic inflammation. This mechanism of scavenging accessory proteins by OxPAPC, or by inhibiting TLR-signaling may represent a negative feedback during inflammation to blunt innate immune responses and provide protection from overshooting inflammatory reactions.
Membrane receptors recognizing OxPL
CD36
CD36 is membrane protein known to have properties of fatty acid binding and uptake, and is expressed on a wide variety of cells, including macrophages. Podrez et al. classified a series of short-chain OxPCs, which bind to and activate CD36 on macrophages and platelets[27, 123]. They also found that adding these OxPL to cholesterol-containing liposomes enhanced their CD36-mediated uptake into macrophages[28]. The Hazen group further identified OxPS on apoptotic cells as an essential component for CD36-mediated macrophage phagocytosis of apoptotic cells[67]. Within this same study, it was shown that loading cells with oxidized phosphatidylserines, but not non-oxidized PS, facilitated their uptake by macrophages. There is evidence for CD36 activation by the following species of OxPL: OxPAPC, LysoPC, PGPC, POVPC, HOOA-PC, KOOA-PC, HOdiA-PC, and KOdiA-PC (Table 2). As mentioned previously, it was also found that the same individual species with PS head-group also bind CD36.
TLR2
Our lab has shown that OxPAPC induces pro-inflammatory gene expression in macrophages via a mechanism that involves TLR2[87] . TLR2 dimerizes with other TLRs, namely TLR1 and TLR6, to recognize distinct sets of ligands[124]. TLR2 heterodimers interact with CD36 or CD14, so it is possible that OxPL induce TLR2-dependent signaling via binding to these accessory receptors. The crystal structure of TLR2 with TLR1 and ligand Pam3CSK4 was elucidated in 2007 and with it, a better understanding of the TLR2 binding pocket[125]. Compared to conventional TLR2 agonists, such as lipoteichoic acid (LTA) or Pam3CSK4, OxPL have been shown to function as weak agonists for TLR2, with agonistic activity mainly residing in the long-chain fraction of OxPAPC[87]. Previously, only LysoPC[126] and LysoPS[127] purified species have been shown to activate TLR2. Based on these studies, the activation of TLR2 is not head group specific, as noted by the inclusion of both PC and PS head groups in the TLR2-activating lipids. As of now, there is no published evidence to support whether PE species activate TLR2. OxPL species that have been described to inhibit the action of TLR2 include PGPC, POVPC, PECPC, PEIPC, KOdia-PC, OxPAPC and OxPAPE[40, 110, 111].
TLR2-deficiency in mice was shown to be protective in various settings of non-infectious inflammatory disease models including atherosclerosis, reperfusion injury, and diabetes[128–130]. Oxidative tissue damage is a prominent feature in these diseases, and as such, TLR2 recognition of oxidatively modified DAMPs may influence disease initiation and progression. However, further studies are necessary to fully understand the structural requirements of OxPL for either activation or inhibition of TLR2.
TLR4
While there are many reports demonstrating that oxidized phospholipids inhibit TLR4-mediated effects[40, 101, 102, 110, 111], there is controversial evidence to date to support a role for TLR4 in OxPL-induced inflammation. While some studies imply TLR4 in mediating effects of OxPL[131] or LysoPC[126], others have ruled out an involvement of TLR4 in OxPL-mediated pro-inflammatory effects[132]. OxPL species that have been shown to inhibit LPS-induced TLR4 activation include PGPC, POVPC, PECPC, PEIPC, OxPAPC and OxPAPE (Table 2).
TLR9
A recent report showed TLR9 to be activated by CEP in platelets, thereby increasing platelet reactivity[133]. These findings further imply an important role for lipid oxidation in the control of thrombogenicity[53, 123, 134, 135], and increase the spectrum of immune receptors recognizing endogenously formed oxidation products.
GPCRs
A number of GPCRs have been implicated to play a role in recognizing and transmitting signals induced by OxPL. Due to structural similarities of fragmented OxPL with platelet activating factor (PAF) and the finding that PAF-receptor antagonists could inhibit some effects of OxPL[136, 137], the PAF-receptor has been implicated as one of the major receptors[138–140]. Other GPCRs that have been shown to be involved in mediating effects of OxPL include the prostaglandin E2 (EP2) receptor[141], the S1P1 receptor[142], VEGF receptor[143], and GRP-78[144].
Intracellular ‘receptors’ that sense OxPL
Intracellular sensors for OxPL exist, however, whether sufficient amounts of OxPL are produced inside the cell or if they have to be transported across cell membranes has not been elucidated. However, it has been shown that some OxPL species, including PGPC, readily cross the cell membrane[105] and Tom McIntyre’s group identified TMEM30a as a transport channel, facilitating OxPL traffic from outside the cell to the mitochondria[145].
Keap1/Nrf2
Nrf2 is a cytosolic redox-regulated transcription factor that is bound by Keap1, a redox-sensitive chaperone protein that promotes the degradation of Nrf2. Upon oxidative or electrophilic stress, Keap1 is thought to undergo a conformational change, disrupting its ability to bind Nrf2, allowing Nrf2 to translocate to the nucleus and promote transcription of antioxidant response systems[146]. The mechanism by which OxPL activate Nrf2-dependent gene expression may be by direct binding of electrophilic functional groups (such as those containing cyclopentenones) to Keap-1. On the other hand, OxPL have been shown to activate protein kinase C (PKC)-dependent signaling[10], and Nrf2 was shown to be phosphorylated at Ser-40 by PKC[147, 148].
Caspase 11/NLRP3
Recently a study showed the ability of OxPAPC to directly activate caspase11 and subsequently the NLRP3 inflammasome in dendritic cells[89]. These authors demonstrated direct binding of certain species of OxPAPC, likely PEIPC, to caspase11 in a similar manner as intracellular LPS would bind to and activate caspase 11. Others have shown that OxPL induce NLRP3 activation in macrophages[88].
PPARγ
Among nuclear hormone receptors, peroxisome proliferator-activated receptor-γ (PPARγ) was shown to recognize OxPL. McIntyre’s group showed that OxPL activate PPARγ-dependent gene expression, leading to the expression of cyclooxygenase 2 in monocytes[149, 150].
Conclusion and Outlook
Together, oxidative modification of phospholipids represents a common underlying mechanism in many diseases where tissue damage is involved. The formation of OxPL seems to be a general feature in chronic inflammatory settings that often lead to debilitating states in many patients. Therefore, attempts to lower OxPL levels are promising approaches to combat inflammation and tissue damage[151]. Not surprisingly, OxPL can be used as predictive biomarkers for certain diseases[152]. On the other hand, some commercial enterprises have explored the use of Lecinoxoids, which are ‘oxidized phospholipid-like small molecules,’ for anti-inflammatory therapies[153–155]. For instance, Lecinoxoid VB-201 had a notable effect in a mouse model of experimental autoimmune encephalomyelitis[155], and inhibited CD14 and TLR2-dependent inflammation[154]. Devising novel strategies for lowering OxPL levels as well as exploiting anti-inflammatory properties by developing small molecules modeled after OxPL should produce novel therapies against chronic inflammatory diseases.
Highlights.
Oxidized phospholipids (OxPL) are endogenous danger-associated molecular patterns.
Receptors that interact with OxPL include CD14, TLR2, CD36, Nrf2, and Caspase 11.
OxPL induce a pro-inflammatory response in macrophages.
OxPL antagonize the effects of pathogen-associated molecular patterns.
We provide a summary of structure-function relationships of diverse OxPL species.
Acknowledgments
This work has been supported by NIH grant R01 DK096076 to N.L.; V.S. is a recipient of a SFRBM Mini-Fellowship and is supported by an NIH F31 pre-doctoral fellowship. D.D and V.S. are also supported by a Double-Hoo grant from the University of Virginia. The authors apologize to the many authors whose excellent work could not be cited due to space limitations.
Abbreviations
- CAP
carboxyalkylpyrrole
- CAP-PE
carboxyalkylpyrrole-phosphatidylethanolamine
- CEP
carboxyethyl-pyrrole
- DAMPs
danger associated molecular patterns
- DC
dendritic cell
- HUVEC
human umbilical-vein endothelial cells
- LBP
LPS binding protein
- LTA
lipoteichoic acid
- mm-LDL
minimally modified low-density lipoprotein
- NAEs
N-acyl-ethanolamides
- NAPEs
N-acyl-phosphatidylethanolamines
- Nrf2
nuclear factor (erythroid-derived 2)-like 2
- OxCE
oxidized cholesteryl esters
- oxLDL
oxidized low-density lipoprotein
- OxPAPC
oxidation products of PAPC
- OxPC
oxidized phosphatidylcholine
- OxPL
oxidized phospholipids
- PAF
platelet activating factor
- Pam3CSK4
N-palmitoyl-S-dipalmitoylglyceryl-Cys-Scr-(lys)4
- PAPC
1-palimitoyl-2-arachidonoyl-sn-glycero-3-phosphorylcholine
- PC
phosphatidylcholine
- PE
phosphatidylethanolamine
- PEIPC
1-palmityoyl-2-(5,6-epoxyisoprostane)-sn-glycero-3-phosphorylcholine
- PGPC
1-palmityoyl-2-glutaroyl-sn-glycero-3-phosphorylcholine
- PKC
protein kinase C
- POVPC
1-palmityoyl-2-oxovaleroyl-sn-glycero-3-phosphorylcholine
- PPARγ
peroxisome proliferator-activated receptorγ
- PRR
pattern recognition receptors
- PS
phosphatidylserine
- Syk
spleen tyrosine kinase
- TLR4
toll-like receptor 4
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Bibliography
- 1.Esterbauer H, Jurgens G, Quehenberger O. Modification of human low density lipoprotein by lipid peroxidation. Basic life sciences. 1988;49:369–73. doi: 10.1007/978-1-4684-5568-7_57. [DOI] [PubMed] [Google Scholar]
- 2.Esterbauer H, Jurgens G, Quehenberger O, Koller E. Autoxidation of human low density lipoprotein: loss of polyunsaturated fatty acids and vitamin E and generation of aldehydes. Journal of lipid research. 1987;28(5):495–509. [PubMed] [Google Scholar]
- 3.Lang J, Celotto C, Esterbauer H. Quantitative determination of the lipid peroxidation product 4-hydroxynonenal by high-performance liquid chromatography. Analytical biochemistry. 1985;150(2):369–78. doi: 10.1016/0003-2697(85)90525-1. [DOI] [PubMed] [Google Scholar]
- 4.Binder CJ, Papac-Milicevic N, Witztum JL. Innate sensing of oxidation-specific epitopes in health and disease. Nature reviews Immunology. 2016;16(8):485–97. doi: 10.1038/nri.2016.63. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Miller YI, Choi SH, Wiesner P, Fang L, Harkewicz R, Hartvigsen K, Boullier A, Gonen A, Diehl CJ, Que X, Montano E, Shaw PX, Tsimikas S, Binder CJ, Witztum JL. Oxidation-specific epitopes are danger-associated molecular patterns recognized by pattern recognition receptors of innate immunity. Circulation research. 2011;108(2):235–48. doi: 10.1161/CIRCRESAHA.110.223875. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Hazen SL. Oxidized phospholipids as endogenous pattern recognition ligands in innate immunity. The Journal of biological chemistry. 2008;283(23):15527–31. doi: 10.1074/jbc.R700054200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Kronke G, Leitinger N. Oxidized phospholipids at the interface of innate and adaptive immunity. Future Lipidology. 2006;1(5):623–630. [Google Scholar]
- 8.Watson AD, Leitinger N, Navab M, Faull KF, Horkko S, Witztum JL, Palinski W, Schwenke D, Salomon RG, Sha W, Subbanagounder G, Fogelman AM, Berliner JA. Structural identification by mass spectrometry of oxidized phospholipids in minimally oxidized low density lipoprotein that induce monocyte/endothelial interactions and evidence for their presence in vivo. The Journal of biological chemistry. 1997;272(21):13597–607. doi: 10.1074/jbc.272.21.13597. [DOI] [PubMed] [Google Scholar]
- 9.Watson AD, Navab M, Hama SY, Sevanian A, Prescott SM, Stafforini DM, McIntyre TM, Du BN, Fogelman AM, Berliner JA. Effect of platelet activating factor-acetylhydrolase on the formation and action of minimally oxidized low density lipoprotein. The Journal of clinical investigation. 1995;95(2):774–82. doi: 10.1172/JCI117726. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Mauerhofer C, Philippova M, Oskolkova OV, Bochkov VN. Hormetic and anti-inflammatory properties of oxidized phospholipids. Molecular aspects of medicine. 2016;49:78–90. doi: 10.1016/j.mam.2016.02.003. [DOI] [PubMed] [Google Scholar]
- 11.Bochkov VN. Inflammatory profile of oxidized phospholipids. Thrombosis and haemostasis. 2007;97(3):348–54. [PubMed] [Google Scholar]
- 12.Freigang S. The regulation of inflammation by oxidized phospholipids. European journal of immunology. 2016;46(8):1818–25. doi: 10.1002/eji.201545676. [DOI] [PubMed] [Google Scholar]
- 13.Furnkranz A, Leitinger N. Regulation of inflammatory responses by oxidized phospholipids: structure-function relationships. Current pharmaceutical design. 2004;10(8):915–21. doi: 10.2174/1381612043452929. [DOI] [PubMed] [Google Scholar]
- 14.Berliner JA, Leitinger N, Tsimikas S. The role of oxidized phospholipids in atherosclerosis. Journal of lipid research. 2009;50(Suppl):S207–12. doi: 10.1194/jlr.R800074-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Arroyo A, Modriansky M, Serinkan FB, Bello RI, Matsura T, Jiang J, Tyurin VA, Tyurina YY, Fadeel B, Kagan VE. NADPH oxidase-dependent oxidation and externalization of phosphatidylserine during apoptosis in Me2SO-differentiated HL-60 cells. Role in phagocytic clearance. The Journal of biological chemistry. 2002;277(51):49965–75. doi: 10.1074/jbc.M204513200. [DOI] [PubMed] [Google Scholar]
- 16.Horkko S, Bird DA, Miller E, Itabe H, Leitinger N, Subbanagounder G, Berliner JA, Friedman P, Dennis EA, Curtiss LK, Palinski W, Witztum JL. Monoclonal autoantibodies specific for oxidized phospholipids or oxidized phospholipid-protein adducts inhibit macrophage uptake of oxidized low-density lipoproteins. The Journal of clinical investigation. 1999;103(1):117–28. doi: 10.1172/JCI4533. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Horkko S, Miller E, Dudl E, Reaven P, Curtiss LK, Zvaifler NJ, Terkeltaub R, Pierangeli SS, Branch DW, Palinski W, Witztum JL. Antiphospholipid antibodies are directed against epitopes of oxidized phospholipids. Recognition of cardiolipin by monoclonal antibodies to epitopes of oxidized low density lipoprotein. The Journal of clinical investigation. 1996;98(3):815–25. doi: 10.1172/JCI118854. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Chang MK, Binder CJ, Miller YI, Subbanagounder G, Silverman GJ, Berliner JA, Witztum JL. Apoptotic cells with oxidation-specific epitopes are immunogenic and proinflammatory. The Journal of experimental medicine. 2004;200(11):1359–70. doi: 10.1084/jem.20031763. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Huber J, Vales A, Mitulovic G, Blumer M, Schmid R, Witztum JL, Binder BR, Leitinger N. Oxidized membrane vesicles and blebs from apoptotic cells contain biologically active oxidized phospholipids that induce monocyte-endothelial interactions. Arteriosclerosis, thrombosis and vascular biology. 2002;22(1):101–7. doi: 10.1161/hq0102.101525. [DOI] [PubMed] [Google Scholar]
- 20.Rothe T, Gruber F, Uderhardt S, Ipseiz N, Rossner S, Oskolkova O, Bluml S, Leitinger N, Bicker W, Bochkov VN, Yamamoto M, Steinkasserer A, Schett G, Zinser E, Kronke G. 12/15-Lipoxygenase-mediated enzymatic lipid oxidation regulates DC maturation and function. The Journal of clinical investigation. 2015;125(5):1944–54. doi: 10.1172/JCI78490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Uderhardt S, Herrmann M, Oskolkova OV, Aschermann S, Bicker W, Ipseiz N, Sarter K, Frey B, Rothe T, Voll R, Nimmerjahn F, Bochkov VN, Schett G, Kronke G. 12/15-lipoxygenase orchestrates the clearance of apoptotic cells and maintains immunologic tolerance. Immunity. 2012;36(5):834–46. doi: 10.1016/j.immuni.2012.03.010. [DOI] [PubMed] [Google Scholar]
- 22.Huber J, Boechzelt H, Karten B, Surboeck M, Bochkov VN, Binder BR, Sattler W, Leitinger N. Oxidized cholesteryl linoleates stimulate endothelial cells to bind monocytes via the extracellular signal-regulated kinase 1/2 pathway. Arteriosclerosis, thrombosis and vascular biology. 2002;22(4):581–6. doi: 10.1161/01.atv.0000012782.59850.41. [DOI] [PubMed] [Google Scholar]
- 23.Choi SH, Yin H, Ravandi A, Armando A, Dumlao D, Kim J, Almazan F, Taylor AM, McNamara CA, Tsimikas S, Dennis EA, Witztum JL, Miller YI. Polyoxygenated cholesterol ester hydroperoxide activates TLR4 and SYK dependent signaling in macrophages. PloS one. 2013;8(12):e83145. doi: 10.1371/journal.pone.0083145. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Watson AD, Subbanagounder G, Welsbie DS, Faull KF, Navab M, Jung ME, Fogelman AM, Berliner JA. Structural identification of a novel pro-inflammatory epoxyisoprostane phospholipid in mildly oxidized low density lipoprotein. The Journal of biological chemistry. 1999;274(35):24787–98. doi: 10.1074/jbc.274.35.24787. [DOI] [PubMed] [Google Scholar]
- 25.Subbanagounder G, Leitinger N, Schwenke DC, Wong JW, Lee H, Rizza C, Watson AD, Faull KF, Fogelman AM, Berliner JA. Determinants of bioactivity of oxidized phospholipids. Specific oxidized fatty acyl groups at the sn-2 position. Arteriosclerosis, thrombosis and vascular biology. 2000;20(10):2248–54. doi: 10.1161/01.atv.20.10.2248. [DOI] [PubMed] [Google Scholar]
- 26.Leitinger N, Tyner TR, Oslund L, Rizza C, Subbanagounder G, Lee H, Shih PT, Mackman N, Tigyi G, Territo MC, Berliner JA, Vora DK. Structurally similar oxidized phospholipids differentially regulate endothelial binding of monocytes and neutrophils. Proceedings of the National Academy of Sciences of the United States of America. 1999;96(21):12010–5. doi: 10.1073/pnas.96.21.12010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Podrez EA, Poliakov E, Shen Z, Zhang R, Deng Y, Sun M, Finton PJ, Shan L, Gugiu B, Fox PL, Hoff HF, Salomon RG, Hazen SL. Identification of a novel family of oxidized phospholipids that serve as ligands for the macrophage scavenger receptor CD36. The Journal of biological chemistry. 2002;277(41):38503–16. doi: 10.1074/jbc.M203318200. [DOI] [PubMed] [Google Scholar]
- 28.Podrez EA, Poliakov E, Shen Z, Zhang R, Deng Y, Sun M, Finton PJ, Shan L, Febbraio M, Hajjar DP, Silverstein RL, Hoff HF, Salomon RG, Hazen SL. A novel family of atherogenic oxidized phospholipids promotes macrophage foam cell formation via the scavenger receptor CD36 and is enriched in atherosclerotic lesions. The Journal of biological chemistry. 2002;277(41):38517–23. doi: 10.1074/jbc.M205924200. [DOI] [PubMed] [Google Scholar]
- 29.van der Valk FM, Bekkering S, Kroon J, Yeang C, Van den Bossche J, van Buul JD, Ravandi A, Nederveen AJ, Verberne HJ, Scipione C, Nieuwdorp M, Joosten LA, Netea MG, Koschinsky ML, Witztum JL, Tsimikas S, Riksen NP, Stroes ES. Oxidized Phospholipids on Lipoprotein(a) Elicit Arterial Wall Inflammation and an Inflammatory Monocyte Response in Humans. Circulation. 2016;134(8):611–24. doi: 10.1161/CIRCULATIONAHA.116.020838. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Tsiantoulas D, Perkmann T, Afonyushkin T, Mangold A, Prohaska TA, Papac-Milicevic N, Millischer V, Bartel C, Horkko S, Boulanger CM, Tsimikas S, Fischer MB, Witztum JL, Lang IM, Binder CJ. Circulating microparticles carry oxidation-specific epitopes and are recognized by natural IgM antibodies. Journal of lipid research. 2015;56(2):440–8. doi: 10.1194/jlr.P054569. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Oskolkova OV, Afonyushkin T, Leitner A, von Schlieffen E, Gargalovic PS, Lusis AJ, Binder BR, Bochkov VN. ATF4-dependent transcription is a key mechanism in VEGF up-regulation by oxidized phospholipids: critical role of oxidized sn-2 residues in activation of unfolded protein response. Blood. 2008;112(2):330–9. doi: 10.1182/blood-2007-09-112870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Romanoski CE, Che N, Yin F, Mai N, Pouldar D, Civelek M, Pan C, Lee S, Vakili L, Yang WP, Kayne P, Mungrue IN, Araujo JA, Berliner JA, Lusis AJ. Network for activation of human endothelial cells by oxidized phospholipids: a critical role of heme oxygenase 1. Circulation research. 2011;109(5):e27–41. doi: 10.1161/CIRCRESAHA.111.241869. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Nonas S, Miller I, Kawkitinarong K, Chatchavalvanich S, Gorshkova I, Bochkov VN, Leitinger N, Natarajan V, Garcia JG, Birukov KG. Oxidized phospholipids reduce vascular leak and inflammation in rat model of acute lung injury. American journal of respiratory and critical care medicine. 2006;173(10):1130–8. doi: 10.1164/rccm.200511-1737OC. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Isakson BE, Kronke G, Kadl A, Leitinger N, Duling BR. Oxidized phospholipids alter vascular connexin expression, phosphorylation, and heterocellular communication. Arteriosclerosis, thrombosis and vascular biology. 2006;26(10):2216–21. doi: 10.1161/01.ATV.0000237608.19055.53. [DOI] [PubMed] [Google Scholar]
- 35.Huber J, Furnkranz A, Bochkov VN, Patricia MK, Lee H, Hedrick CC, Berliner JA, Binder BR, Leitinger N. Specific monocyte adhesion to endothelial cells induced by oxidized phospholipids involves activation of cPLA2 and lipoxygenase. Journal of lipid research. 2006;47(5):1054–62. doi: 10.1194/jlr.M500555-JLR200. [DOI] [PubMed] [Google Scholar]
- 36.Bochkov VN, Philippova M, Oskolkova O, Kadl A, Furnkranz A, Karabeg E, Afonyushkin T, Gruber F, Breuss J, Minchenko A, Mechtcheriakova D, Hohensinner P, Rychli K, Wojta J, Resink T, Erne P, Binder BR, Leitinger N. Oxidized phospholipids stimulate angiogenesis via autocrine mechanisms, implicating a novel role for lipid oxidation in the evolution of atherosclerotic lesions. Circulation research. 2006;99(8):900–8. doi: 10.1161/01.RES.0000245485.04489.ee. [DOI] [PubMed] [Google Scholar]
- 37.Cherepanova OA, Pidkovka NA, Sarmento OF, Yoshida T, Gan Q, Adiguzel E, Bendeck MP, Berliner J, Leitinger N, Owens GK. Oxidized phospholipids induce type VIII collagen expression and vascular smooth muscle cell migration. Circulation research. 2009;104(5):609–18. doi: 10.1161/CIRCRESAHA.108.186064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Pidkovka NA, Cherepanova OA, Yoshida T, Alexander MR, Deaton RA, Thomas JA, Leitinger N, Owens GK. Oxidized phospholipids induce phenotypic switching of vascular smooth muscle cells in vivo and in vitro. Circulation research. 2007;101(8):792–801. doi: 10.1161/CIRCRESAHA.107.152736. [DOI] [PubMed] [Google Scholar]
- 39.Shankman LS, Gomez D, Cherepanova OA, Salmon M, Alencar GF, Haskins RM, Swiatlowska P, Newman AA, Greene ES, Straub AC, Isakson B, Randolph GJ, Owens GK. KLF4-dependent phenotypic modulation of smooth muscle cells has a key role in atherosclerotic plaque pathogenesis. Nature medicine. 2015;21(6):628–37. doi: 10.1038/nm.3866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Bluml S, Kirchberger S, Bochkov VN, Kronke G, Stuhlmeier K, Majdic O, Zlabinger GJ, Knapp W, Binder BR, Stockl J, Leitinger N. Oxidized phospholipids negatively regulate dendritic cell maturation induced by TLRs and CD40. Journal of immunology. 2005;175(1):501–8. doi: 10.4049/jimmunol.175.1.501. [DOI] [PubMed] [Google Scholar]
- 41.Bluml S, Zupkovitz G, Kirchberger S, Seyerl M, Bochkov VN, Stuhlmeier K, Majdic O, Zlabinger GJ, Seiser C, Stockl J. Epigenetic regulation of dendritic cell differentiation and function by oxidized phospholipids. Blood. 2009;114(27):5481–9. doi: 10.1182/blood-2008-11-191429. [DOI] [PubMed] [Google Scholar]
- 42.Seyerl M, Bluml S, Kirchberger S, Bochkov VN, Oskolkova O, Majdic O, Stockl J. Oxidized phospholipids induce anergy in human peripheral blood T cells. European journal of immunology. 2008;38(3):778–87. doi: 10.1002/eji.200737619. [DOI] [PubMed] [Google Scholar]
- 43.Boutaud O, Brame CJ, Salomon RG, Roberts LJ, 2nd, Oates JA. Characterization of the lysyl adducts formed from prostaglandin H2 via the levuglandin pathway. Biochemistry. 1999;38(29):9389–96. doi: 10.1021/bi990470+. [DOI] [PubMed] [Google Scholar]
- 44.Hoppe G, Subbanagounder G, O'Neil J, Salomon RG, Hoff HF. Macrophage recognition of LDL modified by levuglandin E2, an oxidation product of arachidonic acid. Biochimica et biophysica acta. 1997;1344(1):1–5. doi: 10.1016/s0005-2760(96)00160-9. [DOI] [PubMed] [Google Scholar]
- 45.Salomon RG, Bi W. Isolevuglandin adducts in disease. Antioxidants & redox signaling. 2015;22(18):1703–18. doi: 10.1089/ars.2014.6154. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Salomon RG, Subbanagounder G, O'Neil J, Kaur K, Smith MA, Hoff HF, Perry G, Monnier VM. Levuglandin E2-protein adducts in human plasma and vasculature. Chemical research in toxicology. 1997;10(5):536–45. doi: 10.1021/tx960157y. [DOI] [PubMed] [Google Scholar]
- 47.Guo L, Chen Z, Amarnath V, Davies SS. Identification of novel bioactive aldehyde-modified phosphatidylethanolamines formed by lipid peroxidation. Free radical biology & medicine. 2012;53(6):1226–38. doi: 10.1016/j.freeradbiomed.2012.07.077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Guo L, Davies SS. Bioactive aldehyde-modified phosphatidylethanolamines. Biochimie. 2013;95(1):74–8. doi: 10.1016/j.biochi.2012.07.010. [DOI] [PubMed] [Google Scholar]
- 49.Guo L, Chen Z, Amarnath V, Yancey PG, Van Lenten BJ, Savage JR, Fazio S, Linton MF, Davies SS. Isolevuglandin-type lipid aldehydes induce the inflammatory response of macrophages by modifying phosphatidylethanolamines and activating the receptor for advanced glycation endproducts. Antioxidants & redox signaling. 2015;22(18):1633–45. doi: 10.1089/ars.2014.6078. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Sullivan CB, Matafonova E, Roberts LJ, 2nd, Amarnath V, Davies SS. Isoketals form cytotoxic phosphatidylethanolamine adducts in cells. Journal of lipid research. 2010;51(5):999–1009. doi: 10.1194/jlr.M001040. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Kirabo A, Fontana V, de Faria AP, Loperena R, Galindo CL, Wu J, Bikineyeva AT, Dikalov S, Xiao L, Chen W, Saleh MA, Trott DW, Itani HA, Vinh A, Amarnath V, Amarnath K, Guzik TJ, Bernstein KE, Shen XZ, Shyr Y, Chen SC, Mernaugh RL, Laffer CL, Elijovich F, Davies SS, Moreno H, Madhur MS, Roberts J, 2nd, Harrison DG. DC isoketal-modified proteins activate T cells and promote hypertension. The Journal of clinical investigation. 2014;124(10):4642–56. doi: 10.1172/JCI74084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Chen Z, Guo L, Zhang Y, Walzem RL, Pendergast JS, Printz RL, Morris LC, Matafonova E, Stien X, Kang L, Coulon D, McGuinness OP, Niswender KD, Davies SS. Incorporation of therapeutically modified bacteria into gut microbiota inhibits obesity. The Journal of clinical investigation. 2014;124(8):3391–406. doi: 10.1172/JCI72517. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Biswas S, Xin L, Panigrahi S, Zimman A, Wang H, Yakubenko VP, Byzova TV, Salomon RG, Podrez EA. Novel phosphatidylethanolamine derivatives accumulate in circulation in hyperlipidemic ApoE−/− mice and activate platelets via TLR2. Blood. 2016;127(21):2618–29. doi: 10.1182/blood-2015-08-664300. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.West XZ, Malinin NL, Merkulova AA, Tischenko M, Kerr BA, Borden EC, Podrez EA, Salomon RG, Byzova TV. Oxidative stress induces angiogenesis by activating TLR2 with novel endogenous ligands. Nature. 2010;467(7318):972–6. doi: 10.1038/nature09421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Hollyfield JG, Bonilha VL, Rayborn ME, Yang X, Shadrach KG, Lu L, Ufret RL, Salomon RG, Perez VL. Oxidative damage-induced inflammation initiates age-related macular degeneration. Nature medicine. 2008;14(2):194–8. doi: 10.1038/nm1709. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Koller D, Hackl H, Bogner-Strauss JG, Hermetter A. Effects of oxidized phospholipids on gene expression in RAW 264.7 macrophages: a microarray study. PloS one. 2014;9(10):e110486. doi: 10.1371/journal.pone.0110486. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Groeneweg M, Vergouwe MN, Scheffer PG, Vermue HP, Sollewijn Gelpke MD, Sijbers AM, Leitinger N, Hofker MH, de Winther MP. Modification of LDL with oxidized 1-palmitoyl-2-arachidonoyl-sn-glycero-3-phosphorylcholine (oxPAPC) results in a novel form of minimally modified LDL that modulates gene expression in macrophages. Biochimica et biophysica acta. 2008;1781(6–7):336–43. doi: 10.1016/j.bbalip.2008.04.016. [DOI] [PubMed] [Google Scholar]
- 58.Kadl A, Meher AK, Sharma PR, Lee MY, Doran AC, Johnstone SR, Elliott MR, Gruber F, Han J, Chen W, Kensler T, Ravichandran KS, Isakson BE, Wamhoff BR, Leitinger N. Identification of a novel macrophage phenotype that develops in response to atherogenic phospholipids via Nrf2. Circulation research. 2010;107(6):737–46. doi: 10.1161/CIRCRESAHA.109.215715. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Orozco LD, Bennett BJ, Farber CR, Ghazalpour A, Pan C, Che N, Wen P, Qi HX, Mutukulu A, Siemers N, Neuhaus I, Yordanova R, Gargalovic P, Pellegrini M, Kirchgessner T, Lusis AJ. Unraveling inflammatory responses using systems genetics and gene-environment interactions in macrophages. Cell. 2012;151(3):658–70. doi: 10.1016/j.cell.2012.08.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Heilbronn LK, Campbell LV. Adipose tissue macrophages, low grade inflammation and insulin resistance in human obesity. Current pharmaceutical design. 2008;14(12):1225–30. doi: 10.2174/138161208784246153. [DOI] [PubMed] [Google Scholar]
- 61.Furnkranz A, Schober A, Bochkov VN, Bashtrykov P, Kronke G, Kadl A, Binder BR, Weber C, Leitinger N. Oxidized phospholipids trigger atherogenic inflammation in murine arteries. Arteriosclerosis, thrombosis and vascular biology. 2005;25(3):633–8. doi: 10.1161/01.ATV.0000153106.03644.a0. [DOI] [PubMed] [Google Scholar]
- 62.Kadl A, Galkina E, Leitinger N. Induction of CCR2-dependent macrophage accumulation by oxidized phospholipids in the air-pouch model of inflammation. Arthritis and rheumatism. 2009;60(5):1362–71. doi: 10.1002/art.24448. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Ravichandran KS. Beginnings of a good apoptotic meal: the find-me and eat-me signaling pathways. Immunity. 2011;35(4):445–55. doi: 10.1016/j.immuni.2011.09.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Fond AM, Ravichandran KS. Clearance of Dying Cells by Phagocytes: Mechanisms and Implications for Disease Pathogenesis. Advances in experimental medicine and biology. 2016;930:25–49. doi: 10.1007/978-3-319-39406-0_2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Tabas I, Bornfeldt KE. Macrophage Phenotype and Function in Different Stages of Atherosclerosis. Circulation research. 2016;118(4):653–67. doi: 10.1161/CIRCRESAHA.115.306256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Li S, Sun Y, Liang CP, Thorp EB, Han S, Jehle AW, Saraswathi V, Pridgen B, Kanter JE, Li R, Welch CL, Hasty AH, Bornfeldt KE, Breslow JL, Tabas I, Tall AR. Defective phagocytosis of apoptotic cells by macrophages in atherosclerotic lesions of ob/ob mice and reversal by a fish oil diet. Circulation research. 2009;105(11):1072–82. doi: 10.1161/CIRCRESAHA.109.199570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Greenberg ME, Sun M, Zhang R, Febbraio M, Silverstein R, Hazen SL. Oxidized phosphatidylserine-CD36 interactions play an essential role in macrophage-dependent phagocytosis of apoptotic cells. The Journal of experimental medicine. 2006;203(12):2613–25. doi: 10.1084/jem.20060370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Seimon TA, Nadolski MJ, Liao X, Magallon J, Nguyen M, Feric NT, Koschinsky ML, Harkewicz R, Witztum JL, Tsimikas S, Golenbock D, Moore KJ, Tabas I. Atherogenic lipids and lipoproteins trigger CD36-TLR2-dependent apoptosis in macrophages undergoing endoplasmic reticulum stress. Cell metabolism. 2010;12(5):467–82. doi: 10.1016/j.cmet.2010.09.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Miller YI, Viriyakosol S, Binder CJ, Feramisco JR, Kirkland TN, Witztum JL. Minimally modified LDL binds to CD14, induces macrophage spreading via TLR4/MD-2, and inhibits phagocytosis of apoptotic cells. The Journal of biological chemistry. 2003;278(3):1561–8. doi: 10.1074/jbc.M209634200. [DOI] [PubMed] [Google Scholar]
- 70.Knapp S, Matt U, Leitinger N, van der Poll T. Oxidized phospholipids inhibit phagocytosis and impair outcome in gram-negative sepsis in vivo. Journal of immunology. 2007;178(2):993–1001. doi: 10.4049/jimmunol.178.2.993. [DOI] [PubMed] [Google Scholar]
- 71.Matt U, Sharif O, Martins R, Furtner T, Langeberg L, Gawish R, Elbau I, Zivkovic A, Lakovits K, Oskolkova O, Doninger B, Vychytil A, Perkmann T, Schabbauer G, Binder CJ, Bochkov VN, Scott JD, Knapp S. WAVE1 mediates suppression of phagocytosis by phospholipid-derived DAMPs. The Journal of clinical investigation. 2013;123(7):3014–24. doi: 10.1172/JCI60681. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Stemmer U, Dunai ZA, Koller D, Purstinger G, Zenzmaier E, Deigner HP, Aflaki E, Kratky D, Hermetter A. Toxicity of oxidized phospholipids in cultured macrophages. Lipids in health and disease. 2012;11:110. doi: 10.1186/1476-511X-11-110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Gruber F, Mayer H, Lengauer B, Mlitz V, Sanders JM, Kadl A, Bilban M, de Martin R, Wagner O, Kensler TW, Yamamoto M, Leitinger N, Tschachler E. NF-E2-related factor 2 regulates the stress response to UVA-1-oxidized phospholipids in skin cells. FASEB journal : official publication of the Federation of American Societies for Experimental Biology. 2010;24(1):39–48. doi: 10.1096/fj.09-133520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Gruber F, Oskolkova O, Leitner A, Mildner M, Mlitz V, Lengauer B, Kadl A, Mrass P, Kronke G, Binder BR, Bochkov VN, Leitinger N, Tschachler E. Photooxidation generates biologically active phospholipids that induce heme oxygenase-1 in skin cells. The Journal of biological chemistry. 2007;282(23):16934–41. doi: 10.1074/jbc.M702523200. [DOI] [PubMed] [Google Scholar]
- 75.Leitinger N, Schulman IG. Phenotypic polarization of macrophages in atherosclerosis. Arteriosclerosis, thrombosis and vascular biology. 2013;33(6):1120–6. doi: 10.1161/ATVBAHA.112.300173. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Adamson S, Leitinger N. Phenotypic modulation of macrophages in response to plaque lipids. Current opinion in lipidology. 2011;22(5):335–42. doi: 10.1097/MOL.0b013e32834a97e4. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Jyrkkanen HK, Kansanen E, Inkala M, Kivela AM, Hurttila H, Heinonen SE, Goldsteins G, Jauhiainen S, Tiainen S, Makkonen H, Oskolkova O, Afonyushkin T, Koistinaho J, Yamamoto M, Bochkov VN, Yla-Herttuala S, Levonen AL. Nrf2 regulates antioxidant gene expression evoked by oxidized phospholipids in endothelial cells and murine arteries in vivo. Circulation research. 2008;103(1):e1–9. doi: 10.1161/CIRCRESAHA.108.176883. [DOI] [PubMed] [Google Scholar]
- 78.Afonyushkin T, Oskolkova OV, Philippova M, Resink TJ, Erne P, Binder BR, Bochkov VN. Oxidized phospholipids regulate expression of ATF4 and VEGF in endothelial cells via NRF2-dependent mechanism: novel point of convergence between electrophilic and unfolded protein stress pathways. Arteriosclerosis, thrombosis and vascular biology. 2010;30(5):1007–13. doi: 10.1161/ATVBAHA.110.204354. [DOI] [PubMed] [Google Scholar]
- 79.Collins AR, Gupte AA, Ji R, Ramirez MR, Minze LJ, Liu JZ, Arredondo M, Ren Y, Deng T, Wang J, Lyon CJ, Hsueh WA. Myeloid deletion of nuclear factor erythroid 2-related factor 2 increases atherosclerosis and liver injury. Arteriosclerosis, thrombosis and vascular biology. 2012;32(12):2839–46. doi: 10.1161/ATVBAHA.112.300345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Freigang S, Ampenberger F, Spohn G, Heer S, Shamshiev AT, Kisielow J, Hersberger M, Yamamoto M, Bachmann MF, Kopf M. Nrf2 is essential for cholesterol crystal-induced inflammasome activation and exacerbation of atherosclerosis. European journal of immunology. 2011;41(7):2040–51. doi: 10.1002/eji.201041316. [DOI] [PubMed] [Google Scholar]
- 81.Barajas B, Che N, Yin F, Rowshanrad A, Orozco LD, Gong KW, Wang X, Castellani LW, Reue K, Lusis AJ, Araujo JA. NF-E2-related factor 2 promotes atherosclerosis by effects on plasma lipoproteins and cholesterol transport that overshadow antioxidant protection. Arteriosclerosis, thrombosis and vascular biology. 2011;31(1):58–66. doi: 10.1161/ATVBAHA.110.210906. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Meher AK, Sharma PR, Lira VA, Yamamoto M, Kensler TW, Yan Z, Leitinger N. Nrf2 deficiency in myeloid cells is not sufficient to protect mice from high-fat diet-induced adipose tissue inflammation and insulin resistance. Free radical biology & medicine. 2012;52(9):1708–15. doi: 10.1016/j.freeradbiomed.2012.02.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Kobayashi EH, Suzuki T, Funayama R, Nagashima T, Hayashi M, Sekine H, Tanaka N, Moriguchi T, Motohashi H, Nakayama K, Yamamoto M. Nrf2 suppresses macrophage inflammatory response by blocking proinflammatory cytokine transcription. Nature communications. 2016;7:11624. doi: 10.1038/ncomms11624. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Hua S, Ek CJ, Mallard C, Johansson ME. Perinatal hypoxia-ischemia reduces alpha 7 nicotinic receptor expression and selective alpha 7 nicotinic receptor stimulation suppresses inflammation and promotes microglial Mox phenotype. BioMed research international. 2014;2014:718769. doi: 10.1155/2014/718769. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Liu B, Tai Y, Caceres AI, Achanta S, Balakrishna S, Shao X, Fang J, Jordt SE. Oxidized Phospholipid OxPAPC Activates TRPA1 and Contributes to Chronic Inflammatory Pain in Mice. PloS one. 2016;11(11):e0165200. doi: 10.1371/journal.pone.0165200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Marques L, Negre-Salvayre A, Costa L, Canonne-Hergaux F. Iron gene expression profile in atherogenic Mox macrophages. Biochimica et biophysica acta. 2016;1862(6):1137–46. doi: 10.1016/j.bbadis.2016.03.004. [DOI] [PubMed] [Google Scholar]
- 87.Kadl A, Sharma PR, Chen W, Agrawal R, Meher AK, Rudraiah S, Grubbs N, Sharma R, Leitinger N. Oxidized phospholipid-induced inflammation is mediated by Toll-like receptor 2. Free radical biology & medicine. 2011;51(10):1903–9. doi: 10.1016/j.freeradbiomed.2011.08.026. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 88.Yeon SH, Yang G, Lee HE, Lee JY. Oxidized phosphatidylcholine induces the activation of NLRP3 inflammasome in macrophages. Journal of leukocyte biology. 2016 doi: 10.1189/jlb.3VMA1215-579RR. [DOI] [PubMed] [Google Scholar]
- 89.Zanoni I, Tan Y, Di Gioia M, Broggi A, Ruan J, Shi J, Donado CA, Shao F, Wu H, Springstead JR, Kagan JC. An endogenous caspase-11 ligand elicits interleukin-1 release from living dendritic cells. Science. 2016;352(6290):1232–6. doi: 10.1126/science.aaf3036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 90.Kayagaki N, Warming S, Lamkanfi M, Vande Walle L, Louie S, Dong J, Newton K, Qu Y, Liu J, Heldens S, Zhang J, Lee WP, Roose-Girma M, Dixit VM. Non-canonical inflammasome activation targets caspase-11. Nature. 2011;479(7371):117–21. doi: 10.1038/nature10558. [DOI] [PubMed] [Google Scholar]
- 91.O'Neill LA, Pearce EJ. Immunometabolism governs dendritic cell and macrophage function. The Journal of experimental medicine. 2016;213(1):15–23. doi: 10.1084/jem.20151570. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 92.Bosca L, Gonzalez-Ramos S, Prieto P, Fernandez-Velasco M, Mojena M, Martin-Sanz P, Alemany S. Metabolic signatures linked to macrophage polarization: from glucose metabolism to oxidative phosphorylation. Biochemical Society transactions. 2015;43(4):740–4. doi: 10.1042/BST20150107. [DOI] [PubMed] [Google Scholar]
- 93.Tan Z, Xie N, Cui H, Moellering DR, Abraham E, Thannickal VJ, Liu G. Pyruvate dehydrogenase kinase 1 participates in macrophage polarization via regulating glucose metabolism. Journal of immunology. 2015;194(12):6082–9. doi: 10.4049/jimmunol.1402469. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Zhu L, Zhao Q, Yang T, Ding W, Zhao Y. Cellular metabolism and macrophage functional polarization. International reviews of immunology. 2015;34(1):82–100. doi: 10.3109/08830185.2014.969421. [DOI] [PubMed] [Google Scholar]
- 95.Ferrante AW., Jr Macrophages, fat, and the emergence of immunometabolism. The Journal of clinical investigation. 2013;123(12):4992–3. doi: 10.1172/JCI73658. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 96.Tannahill GM, Curtis AM, Adamik J, Palsson-McDermott EM, McGettrick AF, Goel G, Frezza C, Bernard NJ, Kelly B, Foley NH, Zheng L, Gardet A, Tong Z, Jany SS, Corr SC, Haneklaus M, Caffrey BE, Pierce K, Walmsley S, Beasley FC, Cummins E, Nizet V, Whyte M, Taylor CT, Lin H, Masters SL, Gottlieb E, Kelly VP, Clish C, Auron PE, Xavier RJ, O'Neill LA. Succinate is an inflammatory signal that induces IL-1beta through HIF-1alpha. Nature. 2013;496(7444):238–42. doi: 10.1038/nature11986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Huang SC, Everts B, Ivanova Y, O'Sullivan D, Nascimento M, Smith AM, Beatty W, Love-Gregory L, Lam WY, O'Neill CM, Yan C, Du H, Abumrad NA, Urban JF, Jr, Artyomov MN, Pearce EL, Pearce EJ. Cell-intrinsic lysosomal lipolysis is essential for alternative activation of macrophages. Nature immunology. 2014;15(9):846–55. doi: 10.1038/ni.2956. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Van den Bossche J, Baardman J, Otto NA, van der Velden S, Neele AE, van den Berg SM, Luque-Martin R, Chen HJ, Boshuizen MC, Ahmed M, Hoeksema MA, de Vos AF, de Winther MP. Mitochondrial Dysfunction Prevents Repolarization of Inflammatory Macrophages. Cell reports. 2016;17(3):684–696. doi: 10.1016/j.celrep.2016.09.008. [DOI] [PubMed] [Google Scholar]
- 99.Halasiddappa LM, Koefeler H, Futerman AH, Hermetter A. Oxidized phospholipids induce ceramide accumulation in RAW 264.7 macrophages: role of ceramide synthases. PloS one. 2013;8(7):e70002. doi: 10.1371/journal.pone.0070002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 100.Chen BC, Chang HM, Hsu MJ, Shih CM, Chiu YH, Chiu WT, Lin CH. Peptidoglycan induces cyclooxygenase-2 expression in macrophages by activating the neutral sphingomyelinase-ceramide pathway. The Journal of biological chemistry. 2009;284(31):20562–73. doi: 10.1074/jbc.M109.028084. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.von Schlieffen E, Oskolkova OV, Schabbauer G, Gruber F, Bluml S, Genest M, Kadl A, Marsik C, Knapp S, Chow J, Leitinger N, Binder BR, Bochkov VN. Multi-hit inhibition of circulating and cell-associated components of the toll-like receptor 4 pathway by oxidized phospholipids. Arteriosclerosis, thrombosis and vascular biology. 2009;29(3):356–62. doi: 10.1161/ATVBAHA.108.173799. [DOI] [PubMed] [Google Scholar]
- 102.Bochkov VN, Kadl A, Huber J, Gruber F, Binder BR, Leitinger N. Protective role of phospholipid oxidation products in endotoxin-induced tissue damage. Nature. 2002;419(6902):77–81. doi: 10.1038/nature01023. [DOI] [PubMed] [Google Scholar]
- 103.Henriksen EJ, Diamond-Stanic MK, Marchionne EM. Oxidative stress and the etiology of insulin resistance and type 2 diabetes. Free radical biology & medicine. 2011;51(5):993–9. doi: 10.1016/j.freeradbiomed.2010.12.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 104.Haider L, Fischer MT, Frischer JM, Bauer J, Hoftberger R, Botond G, Esterbauer H, Binder CJ, Witztum JL, Lassmann H. Oxidative damage in multiple sclerosis lesions. Brain : a journal of neurology. 2011;134(Pt 7):1914–24. doi: 10.1093/brain/awr128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 105.Stemmer U, Ramprecht C, Zenzmaier E, Stojcic B, Rechberger G, Kollroser M, Hermetter A. Uptake and protein targeting of fluorescent oxidized phospholipids in cultured RAW 264.7 macrophages. Biochimica et biophysica acta. 2012;1821(4):706–18. doi: 10.1016/j.bbalip.2012.01.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Gugiu BG, Mouillesseaux K, Duong V, Herzog T, Hekimian A, Koroniak L, Vondriska TM, Watson AD. Protein targets of oxidized phospholipids in endothelial cells. Journal of lipid research. 2008;49(3):510–20. doi: 10.1194/jlr.M700264-JLR200. [DOI] [PubMed] [Google Scholar]
- 107.Springstead JR, Gugiu BG, Lee S, Cha S, Watson AD, Berliner JA. Evidence for the importance of OxPAPC interaction with cysteines in regulating endothelial cell function. Journal of lipid research. 2012;53(7):1304–15. doi: 10.1194/jlr.M025320. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 108.Stewart CR, Stuart LM, Wilkinson K, van Gils JM, Deng J, Halle A, Rayner KJ, Boyer L, Zhong R, Frazier WA, Lacy-Hulbert A, El Khoury J, Golenbock DT, Moore KJ. CD36 ligands promote sterile inflammation through assembly of a Toll-like receptor 4 and 6 heterodimer. Nature immunology. 2010;11(2):155–61. doi: 10.1038/ni.1836. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Bochkov VN, Oskolkova OV, Birukov KG, Levonen AL, Binder CJ, Stockl J. Generation and biological activities of oxidized phospholipids. Antioxidants & redox signaling. 2010;12(8):1009–59. doi: 10.1089/ars.2009.2597. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 110.Walton KA, Cole AL, Yeh M, Subbanagounder G, Krutzik SR, Modlin RL, Lucas RM, Nakai J, Smart EJ, Vora DK, Berliner JA. Specific phospholipid oxidation products inhibit ligand activation of toll-like receptors 4 and 2. Arteriosclerosis, thrombosis and vascular biology. 2003;23(7):1197–203. doi: 10.1161/01.ATV.0000079340.80744.B8. [DOI] [PubMed] [Google Scholar]
- 111.Erridge C, Kennedy S, Spickett CM, Webb DJ. Oxidized phospholipid inhibition of toll-like receptor (TLR) signaling is restricted to TLR2 and TLR4: roles for CD14, LPS-binding protein, and MD2 as targets for specificity of inhibition. The Journal of biological chemistry. 2008;283(36):24748–59. doi: 10.1074/jbc.M800352200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.Oskolkova OV, Afonyushkin T, Preinerstorfer B, Bicker W, von Schlieffen E, Hainzl E, Demyanets S, Schabbauer G, Lindner W, Tselepis AD, Wojta J, Binder BR, Bochkov VN. Oxidized phospholipids are more potent antagonists of lipopolysaccharide than inducers of inflammation. Journal of immunology. 2010;185(12):7706–12. doi: 10.4049/jimmunol.0903594. [DOI] [PubMed] [Google Scholar]
- 113.Binder CJ, Shaw PX, Chang MK, Boullier A, Hartvigsen K, Horkko S, Miller YI, Woelkers DA, Corr M, Witztum JL. The role of natural antibodies in atherogenesis. Journal of lipid research. 2005;46(7):1353–63. doi: 10.1194/jlr.R500005-JLR200. [DOI] [PubMed] [Google Scholar]
- 114.Leibundgut G, Witztum JL, Tsimikas S. Oxidation-specific epitopes and immunological responses: Translational biotheranostic implications for atherosclerosis. Current opinion in pharmacology. 2013;13(2):168–79. doi: 10.1016/j.coph.2013.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Alves JD, Ames PR. Atherosclerosis, oxidative stress and auto-antibodies in systemic lupus erythematosus and primary antiphospholipid syndrome. Immunobiology. 2003;207(1):23–8. doi: 10.1078/0171-2985-00215. [DOI] [PubMed] [Google Scholar]
- 116.Friedman P, Horkko S, Steinberg D, Witztum JL, Dennis EA. Correlation of antiphospholipid antibody recognition with the structure of synthetic oxidized phospholipids. Importance of Schiff base formation and aldol condensation. The Journal of biological chemistry. 2002;277(9):7010–20. doi: 10.1074/jbc.M108860200. [DOI] [PubMed] [Google Scholar]
- 117.Palinski W, Horkko S, Miller E, Steinbrecher UP, Powell HC, Curtiss LK, Witztum JL. Cloning of monoclonal autoantibodies to epitopes of oxidized lipoproteins from apolipoprotein E-deficient mice. Demonstration of epitopes of oxidized low density lipoprotein in human plasma. The Journal of clinical investigation. 1996;98(3):800–14. doi: 10.1172/JCI118853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Greenberg ME, Li XM, Gugiu BG, Gu X, Qin J, Salomon RG, Hazen SL. The lipid whisker model of the structure of oxidized cell membranes. The Journal of biological chemistry. 2008;283(4):2385–96. doi: 10.1074/jbc.M707348200. [DOI] [PubMed] [Google Scholar]
- 119.Bretscher P, Egger J, Shamshiev A, Trotzmuller M, Kofeler H, Carreira EM, Kopf M, Freigang S. Phospholipid oxidation generates potent anti-inflammatory lipid mediators that mimic structurally related pro-resolving eicosanoids by activating Nrf2. EMBO molecular medicine. 2015;7(5):593–607. doi: 10.15252/emmm.201404702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Walton KA, Hsieh X, Gharavi N, Wang S, Wang G, Yeh M, Cole AL, Berliner JA. Receptors involved in the oxidized 1-palmitoyl-2-arachidonoyl-sn-glycero-3-phosphorylcholine-mediated synthesis of interleukin-8. A role for Toll-like receptor 4 and a glycosylphosphatidylinositol-anchored protein. The Journal of biological chemistry. 2003;278(32):29661–6. doi: 10.1074/jbc.M300738200. [DOI] [PubMed] [Google Scholar]
- 121.Erridge C, Spickett CM. Oxidised phospholipid regulation of Toll-like receptor signalling. Redox report : communications in free radical research. 2007;12(1):76–80. doi: 10.1179/135100007X162121. [DOI] [PubMed] [Google Scholar]
- 122.Kim MJ, Choi NY, Koo JE, Kim SY, Joung SM, Jeong E, Lee JY. Suppression of Toll-like receptor 4 activation by endogenous oxidized phosphatidylcholine, KOdiA-PC by inhibiting LPS binding to MD2. Inflammation research : official journal of the European Histamine Research Society .. [et al.] 2013;62(6):571–80. doi: 10.1007/s00011-013-0609-0. [DOI] [PubMed] [Google Scholar]
- 123.Podrez EA, Byzova TV, Febbraio M, Salomon RG, Ma Y, Valiyaveettil M, Poliakov E, Sun M, Finton PJ, Curtis BR, Chen J, Zhang R, Silverstein RL, Hazen SL. Platelet CD36 links hyperlipidemia, oxidant stress and a prothrombotic phenotype. Nature medicine. 2007;13(9):1086–95. doi: 10.1038/nm1626. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Farhat K, Riekenberg S, Heine H, Debarry J, Lang R, Mages J, Buwitt-Beckmann U, Roschmann K, Jung G, Wiesmuller KH, Ulmer AJ. Heterodimerization of TLR2 with TLR1 or TLR6 expands the ligand spectrum but does not lead to differential signaling. Journal of leukocyte biology. 2008;83(3):692–701. doi: 10.1189/jlb.0807586. [DOI] [PubMed] [Google Scholar]
- 125.Jin MS, Kim SE, Heo JY, Lee ME, Kim HM, Paik SG, Lee H, Lee JO. Crystal structure of the TLR1-TLR2 heterodimer induced by binding of a tri-acylated lipopeptide. Cell. 2007;130(6):1071–82. doi: 10.1016/j.cell.2007.09.008. [DOI] [PubMed] [Google Scholar]
- 126.Carneiro AB, Iaciura BM, Nohara LL, Lopes CD, Veas EM, Mariano VS, Bozza PT, Lopes UG, Atella GC, Almeida IC, Silva-Neto MA. Lysophosphatidylcholine triggers TLR2- and TLR4-mediated signaling pathways but counteracts LPS-induced NO synthesis in peritoneal macrophages by inhibiting NF-kappaB translocation and MAPK/ERK phosphorylation. PloS one. 2013;8(9):e76233. doi: 10.1371/journal.pone.0076233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Makide K, Uwamizu A, Shinjo Y, Ishiguro J, Okutani M, Inoue A, Aoki J. Novel lysophosphoplipid receptors: their structure and function. Journal of lipid research. 2014;55(10):1986–95. doi: 10.1194/jlr.R046920. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Mullick AE, Tobias PS, Curtiss LK. Modulation of atherosclerosis in mice by Toll-like receptor 2. The Journal of clinical investigation. 2005;115(11):3149–56. doi: 10.1172/JCI25482. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 129.Leemans JC, Stokman G, Claessen N, Rouschop KM, Teske GJ, Kirschning CJ, Akira S, van der Poll T, Weening JJ, Florquin S. Renal-associated TLR2 mediates ischemia/reperfusion injury in the kidney. The Journal of clinical investigation. 2005;115(10):2894–903. doi: 10.1172/JCI22832. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 130.Ehses JA, Meier DT, Wueest S, Rytka J, Boller S, Wielinga PY, Schraenen A, Lemaire K, Debray S, Van Lommel L, Pospisilik JA, Tschopp O, Schultze SM, Malipiero U, Esterbauer H, Ellingsgaard H, Rutti S, Schuit FC, Lutz TA, Boni-Schnetzler M, Konrad D, Donath MY. Toll-like receptor 2-deficient mice are protected from insulin resistance and beta cell dysfunction induced by a high-fat diet. Diabetologia. 2010;53(8):1795–806. doi: 10.1007/s00125-010-1747-3. [DOI] [PubMed] [Google Scholar]
- 131.Imai Y, Kuba K, Neely GG, Yaghubian-Malhami R, Perkmann T, van Loo G, Ermolaeva M, Veldhuizen R, Leung YH, Wang H, Liu H, Sun Y, Pasparakis M, Kopf M, Mech C, Bavari S, Peiris JS, Slutsky AS, Akira S, Hultqvist M, Holmdahl R, Nicholls J, Jiang C, Binder CJ, Penninger JM. Identification of oxidative stress and Toll-like receptor 4 signaling as a key pathway of acute lung injury. Cell. 2008;133(2):235–49. doi: 10.1016/j.cell.2008.02.043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Erridge C, Webb DJ, Spickett CM. Toll-like receptor 4 signalling is neither sufficient nor required for oxidised phospholipid mediated induction of interleukin-8 expression. Atherosclerosis. 2007;193(1):77–85. doi: 10.1016/j.atherosclerosis.2006.08.032. [DOI] [PubMed] [Google Scholar]
- 133.Panigrahi S, Ma Y, Hong L, Gao D, West XZ, Salomon RG, Byzova TV, Podrez EA. Engagement of platelet toll-like receptor 9 by novel endogenous ligands promotes platelet hyperreactivity and thrombosis. Circulation research. 2013;112(1):103–12. doi: 10.1161/CIRCRESAHA.112.274241. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Hartwich J, Dembinska-Kiec A, Gruca A, Motyka M, Partyka L, Skrzeczynska J, Bzowska M, Pryjma J, Huber J, Leitinger N, Schmitz G. Regulation of platelet adhesion by oxidized lipoproteins and oxidized phospholipids. Platelets. 2002;13(3):141–51. doi: 10.1080/09533710022149368. [DOI] [PubMed] [Google Scholar]
- 135.Leitinger N, Blazek I, Sinzinger H. The influence of isoprostanes on ADP-induced platelet aggregation and cyclic AMP-generation in human platelets. Thrombosis research. 1997;86(4):337–42. doi: 10.1016/s0049-3848(97)00077-7. [DOI] [PubMed] [Google Scholar]
- 136.Subbanagounder G, Leitinger N, Shih PT, Faull KF, Berliner JA. Evidence that phospholipid oxidation products and/or platelet-activating factor play an important role in early atherogenesis : in vitro and In vivo inhibition by WEB 2086. Circulation research. 1999;85(4):311–8. doi: 10.1161/01.res.85.4.311. [DOI] [PubMed] [Google Scholar]
- 137.Leitinger N, Watson AD, Faull KF, Fogelman AM, Berliner JA. Monocyte binding to endothelial cells induced by oxidized phospholipids present in minimally oxidized low density lipoprotein is inhibited by a platelet activating factor receptor antagonist. Advances in experimental medicine and biology. 1997;433:379–82. doi: 10.1007/978-1-4899-1810-9_82. [DOI] [PubMed] [Google Scholar]
- 138.Chen R, Chen X, Salomon RG, McIntyre TM. Platelet activation by low concentrations of intact oxidized LDL particles involves the PAF receptor. Arteriosclerosis, thrombosis and vascular biology. 2009;29(3):363–71. doi: 10.1161/ATVBAHA.108.178731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 139.Marathe GK, Prescott SM, Zimmerman GA, McIntyre TM. Oxidized LDL contains inflammatory PAF-like phospholipids. Trends in cardiovascular medicine. 2001;11(3–4):139–42. doi: 10.1016/s1050-1738(01)00100-1. [DOI] [PubMed] [Google Scholar]
- 140.Marathe GK, Davies SS, Harrison KA, Silva AR, Murphy RC, Castro-Faria-Neto H, Prescott SM, Zimmerman GA, McIntyre TM. Inflammatory platelet-activating factor-like phospholipids in oxidized low density lipoproteins are fragmented alkyl phosphatidylcholines. The Journal of biological chemistry. 1999;274(40):28395–404. doi: 10.1074/jbc.274.40.28395. [DOI] [PubMed] [Google Scholar]
- 141.Li R, Mouillesseaux KP, Montoya D, Cruz D, Gharavi N, Dun M, Koroniak L, Berliner JA. Identification of prostaglandin E2 receptor subtype 2 as a receptor activated by OxPAPC. Circulation research. 2006;98(5):642–50. doi: 10.1161/01.RES.0000207394.39249.fc. [DOI] [PubMed] [Google Scholar]
- 142.Singleton PA, Chatchavalvanich S, Fu P, Xing J, Birukova AA, Fortune JA, Klibanov AM, Garcia JG, Birukov KG. Akt-mediated transactivation of the S1P1 receptor in caveolin-enriched microdomains regulates endothelial barrier enhancement by oxidized phospholipids. Circulation research. 2009;104(8):978–86. doi: 10.1161/CIRCRESAHA.108.193367. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Zimman A, Mouillesseaux KP, Le T, Gharavi NM, Ryvkin A, Graeber TG, Chen TT, Watson AD, Berliner JA. Vascular endothelial growth factor receptor 2 plays a role in the activation of aortic endothelial cells by oxidized phospholipids. Arteriosclerosis, thrombosis and vascular biology. 2007;27(2):332–8. doi: 10.1161/01.ATV.0000252842.57585.df. [DOI] [PubMed] [Google Scholar]
- 144.Birukova AA, Singleton PA, Gawlak G, Tian X, Mirzapoiazova T, Mambetsariev B, Dubrovskyi O, Oskolkova OV, Bochkov VN, Birukov KG. GRP78 is a novel receptor initiating a vascular barrier protective response to oxidized phospholipids. Molecular biology of the cell. 2014;25(13):2006–16. doi: 10.1091/mbc.E13-12-0743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 145.Chen R, Brady E, McIntyre TM. Human TMEM30a promotes uptake of antitumor and bioactive choline phospholipids into mammalian cells. Journal of immunology. 2011;186(5):3215–25. doi: 10.4049/jimmunol.1002710. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Suzuki T, Yamamoto M. Molecular basis of the Keap1-Nrf2 system. Free radical biology & medicine. 2015;88(Pt B):93–100. doi: 10.1016/j.freeradbiomed.2015.06.006. [DOI] [PubMed] [Google Scholar]
- 147.Huang HC, Nguyen T, Pickett CB. Phosphorylation of Nrf2 at Ser-40 by protein kinase C regulates antioxidant response element-mediated transcription. The Journal of biological chemistry. 2002;277(45):42769–74. doi: 10.1074/jbc.M206911200. [DOI] [PubMed] [Google Scholar]
- 148.Bloom DA, Jaiswal AK. Phosphorylation of Nrf2 at Ser40 by protein kinase C in response to antioxidants leads to the release of Nrf2 from INrf2, but is not required for Nrf2 stabilization/accumulation in the nucleus and transcriptional activation of antioxidant response element-mediated NAD(P)H:quinone oxidoreductase-1 gene expression. The Journal of biological chemistry. 2003;278(45):44675–82. doi: 10.1074/jbc.M307633200. [DOI] [PubMed] [Google Scholar]
- 149.Pontsler AV, St Hilaire A, Marathe GK, Zimmerman GA, McIntyre TM. Cyclooxygenase-2 is induced in monocytes by peroxisome proliferator activated receptor gamma and oxidized alkyl phospholipids from oxidized low density lipoprotein. The Journal of biological chemistry. 2002;277(15):13029–36. doi: 10.1074/jbc.M109546200. [DOI] [PubMed] [Google Scholar]
- 150.Davies SS, Pontsler AV, Marathe GK, Harrison KA, Murphy RC, Hinshaw JC, Prestwich GD, Hilaire AS, Prescott SM, Zimmerman GA, McIntyre TM. Oxidized alkyl phospholipids are specific, high affinity peroxisome proliferator-activated receptor gamma ligands and agonists. The Journal of biological chemistry. 2001;276(19):16015–23. doi: 10.1074/jbc.M100878200. [DOI] [PubMed] [Google Scholar]
- 151.Viney NJ, van Capelleveen JC, Geary RS, Xia S, Tami JA, Yu RZ, Marcovina SM, Hughes SG, Graham MJ, Crooke RM, Crooke ST, Witztum JL, Stroes ES, Tsimikas S. Antisense oligonucleotides targeting apolipoprotein(a) in people with raised lipoprotein(a): two randomised, double-blind, placebo-controlled, dose-ranging trials. Lancet. 2016;388(10057):2239–2253. doi: 10.1016/S0140-6736(16)31009-1. [DOI] [PubMed] [Google Scholar]
- 152.Bertoia ML, Pai JK, Lee JH, Taleb A, Joosten MM, Mittleman MA, Yang X, Witztum JL, Rimm EB, Tsimikas S, Mukamal KJ. Oxidation-specific biomarkers and risk of peripheral artery disease. Journal of the American College of Cardiology. 2013;61(21):2169–79. doi: 10.1016/j.jacc.2013.02.047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Mendel I, Yacov N, Shoham A, Ishai E, Breitbart E. Treatment with Oxidized Phospholipids Directly Inhibits Nonalcoholic Steatohepatitis and Liver Fibrosis Without Affecting Steatosis. Digestive diseases and sciences. 2016;61(9):2545–53. doi: 10.1007/s10620-016-4159-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 154.Mendel I, Feige E, Yacov N, Salem Y, Levi I, Propheta-Meiran O, Shoham A, Ishai E, George J, Harats D, Breitbart E. VB-201, an oxidized phospholipid small molecule, inhibits CD14- and Toll-like receptor-2-dependent innate cell activation and constrains atherosclerosis. Clinical and experimental immunology. 2014;175(1):126–37. doi: 10.1111/cei.12212. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Mendel I, Shoham A, Propheta-Meiran O, Ishai E, Halperin G, Feige E, Breitbart E. A Lecinoxoid, an oxidized phospholipid small molecule, constrains CNS autoimmune disease. Journal of neuroimmunology. 2010;226(1–2):126–35. doi: 10.1016/j.jneuroim.2010.06.011. [DOI] [PubMed] [Google Scholar]
- 156.Clark SR, Guy CJ, Scurr MJ, Taylor PR, Kift-Morgan AP, Hammond VJ, Thomas CP, Coles B, Roberts GW, Eberl M, Jones SA, Topley N, Kotecha S, O'Donnell VB. Esterified eicosanoids are acutely generated by 5-lipoxygenase in primary human neutrophils and in human and murine infection. Blood. 2011;117(6):2033–43. doi: 10.1182/blood-2010-04-278887. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 157.Arai M, Imai H, Metori A, Nakagawa Y. Preferential esterification of endogenously formed 5-hydroxyeicosatetraenoic acid to phospholipids in activated polymorphonuclear leukocytes. European journal of biochemistry / FEBS. 1997;244(2):513–9. doi: 10.1111/j.1432-1033.1997.00513.x. [DOI] [PubMed] [Google Scholar]
- 158.Stenson WF, Parker CW. Metabolism of arachidonic acid in ionophore-stimulated neutrophils. Esterification of a hydroxylated metabolite into phospholipids. The Journal of clinical investigation. 1979;64(5):1457–65. doi: 10.1172/JCI109604. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 159.Egger J, Bretscher P, Freigang S, Kopf M, Carreira EM. Synthesis of epoxyisoprostanes: effects in reducing secretion of pro-inflammatory cytokines IL-6 and IL-12. Angewandte Chemie. 2013;52(20):5382–5. doi: 10.1002/anie.201300739. [DOI] [PubMed] [Google Scholar]
- 160.Egger J, Fischer S, Bretscher P, Freigang S, Kopf M, Carreira EM. Total Synthesis of Prostaglandin 15d-PGJ(2) and Investigation of its Effect on the Secretion of IL-6 and IL-12. Organic letters. 2015;17(17):4340–3. doi: 10.1021/acs.orglett.5b02181. [DOI] [PubMed] [Google Scholar]
- 161.Morrow JD, Awad JA, Boss HJ, Blair IA, Roberts LJ., 2nd Non-cyclooxygenase-derived prostanoids (F2-isoprostanes) are formed in situ on phospholipids. Proceedings of the National Academy of Sciences of the United States of America. 1992;89(22):10721–5. doi: 10.1073/pnas.89.22.10721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Salomon RG, Sha W, Brame C, Kaur K, Subbanagounder G, O'Neil J, Hoff HF, Roberts LJ., 2nd Protein adducts of is [4]levuglandin E2, a product of the isoprostane pathway, in oxidized low density lipoprotein. The Journal of biological chemistry. 1999;274(29):20271–80. doi: 10.1074/jbc.274.29.20271. [DOI] [PubMed] [Google Scholar]
- 163.Salomon RG, Batyreva E, Kaur K, Sprecher DL, Schreiber MJ, Crabb JW, Penn MS, DiCorletoe AM, Hazen SL, Podrez EA. Isolevuglandin-protein adducts in humans: products of free radical-induced lipid oxidation through the isoprostane pathway. Biochimica et biophysica acta. 2000;1485(2–3):225–35. doi: 10.1016/s1388-1981(00)00038-x. [DOI] [PubMed] [Google Scholar]
- 164.Itabe H, Yamamoto H, Suzuki M, Kawai Y, Nakagawa Y, Suzuki A, Imanaka T, Takano T. Oxidized phosphatidylcholines that modify proteins. Analysis by monoclonal antibody against oxidized low density lipoprotein. The Journal of biological chemistry. 1996;271(52):33208–17. doi: 10.1074/jbc.271.52.33208. [DOI] [PubMed] [Google Scholar]
- 165.Mattila JP, Sabatini K, Kinnunen PK. Interaction of cytochrome c with 1-palmitoyl-2-azelaoyl-sn-glycero-3-phosphocholine: evidence for acyl chain reversal. Langmuir : the ACS journal of surfaces and colloids. 2008;24(8):4157–60. doi: 10.1021/la703940s. [DOI] [PubMed] [Google Scholar]
- 166.Sabatini K, Mattila JP, Megli FM, Kinnunen PK. Characterization of two oxidatively modified phospholipids in mixed monolayers with DPPC. Biophysical journal. 2006;90(12):4488–99. doi: 10.1529/biophysj.105.080176. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Madesh M, Balasubramanian KA. Activation of liver mitochondrial phospholipase A2 by superoxide. Archives of biochemistry and biophysics. 1997;346(2):187–92. doi: 10.1006/abbi.1997.0288. [DOI] [PubMed] [Google Scholar]