

Abstract
A xylanase gene xynAMG1 with a 1,116-bp open reading frame, encoding an endo-β-1,4-xylanase, was cloned from a chicken cecum metagenome. The translated XynAMG1 protein consisted of 372 amino acids including a putative signal peptide of 23 amino acids. The calculated molecular mass of the mature XynAMG1 was 40,013 Da, with a theoretical pI value of 5.76. The amino acid sequence of XynAMG1 showed 59% identity to endo-β-1,4-xylanase from Prevotella bryantii and Prevotella ruminicola and 58% identity to that from Prevotella copri. XynAMG1 has two conserved motifs, DVVNE and TEXD, containing two active site glutamates and an invariant asparagine, characteristic of GH10 family xylanase. The xynAMG1 gene without signal peptide sequence was cloned and fused with thioredoxin protein (Trx.Tag) in pET-32a plasmid and overexpressed in Escherichia coli Tuner™(DE3)pLysS. The purified mature XynAMG1 was highly salt-tolerant and stable and displayed higher than 96% of its catalytic activity in the reaction containing 1 to 4 M NaCl. It was only slightly affected by common organic solvents added in aqueous solution to up to 5 M. This chicken cecum metagenome-derived xylanase has potential applications in animal feed additives and industrial enzymatic processes requiring exposure to high concentrations of salt and organic solvents.
1. Introduction
Microbial enzymes have been recognized as a major source of various types of biocatalysts which can be successfully applied in different industrial processes [1]. To be practically useful and economically competitive, industrial enzymes must display high activity and stability under harsh conditions to help reduce the production costs [2]. Carbohydrases, enzymes that degrade polymeric carbohydrates, are currently employed in various industries including food and beverages, detergent, biofuel production, textile, paper and pulp, leather industries, and animal feed. However, there are only two enzymes, xylanase and glucanase that dominate the global enzyme market by more than 80% [1, 3].
Xylanases (endo-1,4-beta-xylanase; EC 3.2.1.8) are glycosidases which randomly cleave internalβ-1,4-D-xylosidic linkages of xylan [4], a bioheteropolymer consisting of D-xylose homopolymer backbone that can be substituted to diverse degrees with glucuronosyl, 4-O-methyl-D-glucuronopyranosyl, α-L-arabinofuranosyl, acetyl, feruloyl, and/or p-coumaroyl residues and is one of the most abundant polysaccharides in nature [5–7]. Complete depolymerization of xylan needs a synergistic action of several xylanolytic enzymes. Majority of the xylanases are confined either to glycosyl hydrolase family 10 (GH10) or to family 11 (GH11) based on similarities in their hydrophobic clusters and amino acid sequences of the catalytic domains. However, some are found in other glycosyl hydrolase families including 5, 7, 8, 16, 26, 43, 52, and 62 [4, 8, 9].
Generally, intestinal microorganisms of plant-eating animals are known to be excellent sources of various hydrolytic enzymes. Considering that poultry feeds consist mainly of cereal grains which are rich in nonstarch polysaccharides including xylans and arabinoxylans, microorganisms producing nonstarch polysaccharide hydrolases including xylanase should be abundant in chicken intestine. Chicken cecal microbiota has been shown, in a culture-based study, predominated by the phyla Bacteroidetes and Firmicutes [10]. However, majority of gut microorganisms are unculturable. Therefore, metagenomic strategy is used to recover the genes of desired enzymes [11]. Recently, a metagenomic study of chicken cecal microbiome showed that up to 1.5% of the sequences represented glycosyl hydrolase (GH) domains with over two hundred different sequences of nonstarch polysaccharide-degrading enzymes found [12].
In this paper, we report the gene cloning, sequence and phylogenetic analyses, structural prediction, heterologous expression, and molecular and catalytic characterizations of a new GH10 family endo-xylanase derived from a chicken cecum metagenome.
2. Materials and Methods
2.1. Strains, Plasmids, and Chemicals
The Escherichia coli EPI300™-T1R clone harboring fosmid pCC1FOS carrying a chicken cecal metagenomic DNA fragment containing a xylanase gene was a gift from Dr Kenneth van Driel. All enzymes and dNTPs in this study were purchased from New England BioLabs Inc., USA, and Promega, USA. Plasmid DNA extraction and purification kit was purchased from GE Healthcare, UK. TALON Superflow Metal Affinity Resin (Clonetech) was purchased from TaKaRa (Otsu, Japan). The expression vector pET-32a (Novagen) was used for cloning and expressing the xylanase. E. coli Tuner (DE3)pLysS was used as expression host and was cultivated on Luria–Bertani medium (Difco). The enzyme substrates used were xylan from oat-spelt (Fluka), xylan from beechwood (Megazyme), α-cellulose (Sigma), carboxymethyl cellulose (Sigma), starch (Sigma), β-glucan from barley (Sigma), 4-nitrophenyl-β-D-xylopyranoside (Megazyme), 4-nitrophenyl-β-D-cellobioside (Sigma), and 4-nitrophenyl-α-D-galactopyranoside (Fluka). Molecular weight standard mix containing xylose, xylobiose, xylotriose, xylotetraose, xylopentaose, and xylohexaose (Megazyme) was the gift from Professor Khanok Ratanakhanokchai, KMUTT, Thailand. All other chemicals were obtained from Sigma-Aldrich (St. Louis, MO, USA).
2.2. Bioinformatic Analysis of DNA and Amino Acid Sequences
Nucleotide sequence recognized as xylanase gene was translated into amino acids. Similar sequences were retrieved from the GenBank database using the BLAST search. Sequence alignment and phylogenetic analysis were done using the CLC Main Workbench 7.7 sequence analysis software package (CLC bio). To determine the family of the xylanase, the position of glutamate residues of the active site, and the highly conserved motifs of GH10, the ExPASy–PROSITE (http:/www.expasy.org/prosite) was used. The signal peptide was predicted by using SignalP 4.1 server (http://www.cbs.dtu.dk/services/SignalP). The theoretical pI and molecular weight were predicted using an online prediction tool (http://www.expasy.org/tools/pi_tool.html). The structure of XynAMG1 protein was predicted using the SWISSMODEL (https://swissmodel.expasy.org/) and IntFOLD (http://www.reading.ac.uk/bioinf/IntFOLD/) servers. Images were generated using PyMOL software (http://www.pymol.org/).
2.3. Construction of pET32a-XynAMG1 Plasmid, Gene Expression, and Purification of XynAMG1 Xylanase
The xylanase gene xynAMG1 was amplified by PCR from the pCC1FOS fosmid clones, using the Xyl524GH10CH-Fw: 5′-ATGAGCTCGCTGACACCC-3′ forward and Xyl524GH10CH-Rv: 5′-CTAAGCTTGTCACTGCTTGAAC-3′ reverse primers. The primer pair targeted the truncated gene, excluding the leader peptide encoding sequences, and introduced SacI and HindIII restriction sites at 5′- and 3′-end of the gene. The amplified PCR product was digested with SacI and HindIII and ligated, using T4 DNA polymerase, into pET-32a(+) vector previously linearized with the same restriction enzymes and dephosphorylated with Antarctic Phosphatase (New England BioLabs). The expression plasmid (pET32a–xynAMG1), having the xynAMG1 gene under the control of the T7 promoter, was used to transform chemically competent [13] E. coli Tuner (DH3)pLysS cells to give E. coli Tuner (pET32a–xynAMG1) expression strain. Positive clones were proven by colony PCR and the presence of xylanase activity in the cell lysate. Selected E. coli Tuner (pET32a–xynAMG1) clone was grown in LB broth containing 34 μg/mL and 100 μg/mL of chloramphenicol and ampicillin, respectively, incubated at 37°C with 200 rpm shaking until the culture reached an OD600 nm of 0.6. Then the xynAMG1 gene expression was induced by adding IPTG to a final concentration of 0.4 mM and the culture was further incubated for 5 h at 37°C with 200 rpm shaking. The cells were harvested by centrifugation, resuspended in nine volumes of ice-cold phosphate buffer (50 mM sodium phosphate, 1.2 M NaCl, 10 mM imidazole) pH 7.2, and subjected to ultrasonic cell disruption while keeping in an ice-bath. Unbroken cells and insoluble cell debris were removed by centrifugation at 12,000g for 25 min yielding the clear soluble cell lysate. The cell lysates were subjected to sodium dodecyl sulphate polyacrylamide gel electrophoresis (SDS-PAGE, 12% separating gel) to determine the expression profile of the XynAMG1 xylanase. The recombinant XynAMG1 was purified from cell lysate by Immobilized Metal Affinity Chromatography (IMAC) using the TALON Metal Affinity Resin (Clontech Lab, Inc.) charged with cobalt. The IMAC column (Tricon 10 × 100 mm) was operated using an ÄKTA purifier FPLC system (GE Healthcare Bio-Sciences) at the 0.2 mL/min flow rate. The bound proteins were eluted with 10 bed volumes of linear gradient of 1–150 mM imidazole in phosphate buffer (50 mM sodium phosphate, 1.2 M NaCl) pH 7.2, at 0.5 mL/min flow rate. Fractions containing xylanase activity were pooled and concentrated by the Amicon® 30 kDa cut off centrifugal filter device (EMD Millipore) at 4,000 rpm, 4°C. The concentrated XynAMG1 was repurified using the IMAC and then desalted as described above. The pET-32a derived Trx-His∙tag which fused to the N-terminus of the expressed XynAMG1 was removed by digestion with enterokinase (New England BioLabs) for 16 h at 23°C. The digestion mixture was then loaded onto the IMAC column as above, and the recombinant XynAMG1 was retrieved from the flow-through, while the Trx-His∙tag and the uncleaved Trx-His-XynAMG1 remained bound to the TALON resin. Protein concentration was determined by the Bradford method using bovine serum albumin (BSA) as the standard. The purity of XynAMG1 was analyzed on SDS-PAGE. The zymogram analysis was done on 12% gel native PAGE containing 0.2% oat-spelt xylan as the substrate [14].
2.4. Biochemical Characterization of Purified XynAMG1 Xylanase
Substrate specificity of XynAMG1 was determined by assaying activity towards different substrates at 1% (w/v) concentration in 50 mM citrate buffer pH 5.5. The tested substrates were of the following polymers: beechwood xylan, oat-spelt xylan, α-cellulose, carboxymethyl cellulose, starch, and barley β-glucan [14, 15]. The xylanase activity was estimated by measuring reducing sugar released from the reaction using the DNS method [16]. XynAMG1 was also tested against 4 mg/mL of 4-nitrophenyl-β-D-xylopyranoside, 4-nitrophenyl-β-D-cellobioside, and 4-nitrophenyl-α-D-galactopyranoside synthetic chromogenic substrates as previously described [17]. End products from XynAMG1 catalyzed hydrolysis of beechwood xylan and 4-nitrophenyl-β-D-xylopyranoside were analyzed by thin-layer chromatography (TLC) on a Silica gel 60 F254 plate (10 × 10 cm) (Merck, Darmstadt, Germany) using a developing solvent (chloroform/acetic acid/water 6 : 7 : 1, v/v). The products were visualized by spraying with ethanol/sulfuric acid (95 : 5, v/v) and heating for 10 min at 100°C as previously described [18].
2.5. Effect of pH and Temperature on XynAMG1 Activity and Stability
To determine the optimum pH for XynAMG1 activity, the enzyme was allowed to function in buffers ranging from pH 3.0 to 11.0 with 1 pH unit interval. The buffers used were 50 mM citrate buffer for pH range of 3–6, 50 mM sodium phosphate buffer for pH range of 6–8, and 50 mM glycine NaOH buffer for pH range of 8–11. XynAMG1 activities were determined using the standard enzyme assay, and a pH versus enzyme activity profile was plotted [19]. The enzyme stability at different pH was determined by preincubating XynAMG1 in different buffers from pH 3.0 to 11.0 (with 1 pH unit interval) for 30 min and 60 min at 25°C. Then, the remaining activities of the enzyme were measured under standard condition [20].
The optimum temperature for xylanase activity was determined by assaying the enzyme activity at different temperatures from 20 to 100°C with step increment of 5°C. Thermal stability was determined by incubating the enzyme in pH 5.5 buffer in the absence of substrate at the temperature range of 45 to 70°C. Then, the residual activities were measured under standard conditions [21].
2.6. Effect of Metal Ions, Salt, Chemical Agents, and Solvents on XynAMG1 Activity
XynAMG1 was incubated with 2 mM and 10 mM solution of Mn2+, Ca2+, Zn2+, Co2+, Cu2+, Mg2+, or nonmetal reagents including ethylenediaminetetraacetic acid (EDTA), dithiothreitol (DTT), β-mercaptoethanol (βME), and detergents Triton X-100, Tween 80, and SDS, for 1 h at room temperature. Residual activity was measured under standard condition [22].
To investigate the impact of sodium chloride on XynAMG1 activity, the purified XynAMG1 was incubated with 0–4 M NaCl for 0 and 2 h and then assayed under standard conditions in the reaction containing NaCl at the same concentration as previously incubated [23].
XynAMG1 was incubated in aqueous solution of different solvents (acetone, methanol, ethanol, and propanol) at different concentrations (0–5 M) for 30 min. Residual activity was determined by using assay system under optimal conditions [22].
The nucleotide sequence of the xynAMG1 gene reported in this paper has been deposited in the GenBank database with an accession number of KX347434.
3. Results and Discussion
3.1. In Silico Sequence and Structural Analyses of XynAMG1 Xylanase
An E. coli clone harboring DNA from chicken cecum metagenome was found to contain an open reading frame of 1,116 bp encoding a protein of 372 amino acids, sequence of which was related to xylanase family GH10 and it was named the XynAMG1 xylanase. The first 23 N-terminus amino acids were predicted to be the signal peptide which guides secretion of the 349 amino acids' mature enzyme having a molecular mass and a pI value of 40,013 Da and 5.76, respectively.
Multiple sequence alignment of the XynAMG1 protein (including the signal peptide) with similar proteins, BLASTed and retrieved from the GenBank, revealed the highest identity at 59% to those of endo-1,4-beta-xylanases from Prevotella bryantii and Prevotella ruminicola and 58% identity to that from Prevotella copri. XynAMG1 has two conserved motifs, DVVNE and TEXD, of the GH10 family xylanase which contained the two glutamate residues (E157 and E 262) predicted to be the catalytic sites, and an invariant asparagine (N156) preceding the glutamate in the first motif serving as an acid/base catalyst (Figure 1) [24–26].
Figure 1.

Amino acid sequence alignment of XynAMG1 with closely related GH10 family beta-xylanases from Prevotella bryantii B14 (CAD21011), Prevotella ruminicola B(1)4 (P48789), Prevotella copri DSM 18205 (WP_006847660), Prevotella dentalis DSM 3688 (AGB29059), Paraglaciecola mesophila KMM 241 (ACN76857), Thermobacillus xylanilyticus D3 (CAA76420), and Paenibacillus barcinonensis BP-23 (O69231). The highly conserved motifs DVVNE and TEXD are shown in boxes, each containing an active site glutamate (marked with an asterisk) and an invariant asparagine residue (marked with an arrow) preceding the active site glutamate in the first box.
Phylogenetic analysis of XynAMG1 placed it in the cluster within Prevotella xylanases clade with high bootstrap value support (Figure 2). Among these, the highest amino acid sequence identity was only 59% which was the xylanase from Prevotella bryantii and Prevotella ruminicola. This indicated that XynAMG1 is a distinctly new endo-1,4 β-xylanase member of the family GH10, probably from a bacterium in the genus Prevotella.
Figure 2.

Phylogenetic analysis based on the amino acid sequence of XynAMG1 and related GH10 family xylanases. The phylogenetic tree was constructed using the neighbor joining method (CLC Main Workbench version 7.7). The lengths of the branches indicate the relative divergence among amino acid sequences. The percentage bootstrap values based on 1,000 bootstrap replications are shown at the nodes. Xylanases of Sphingobacterium sp. TN19 and Sphingobacterium sp. HP455 are used as outgroups. Accession numbers of the xylanase amino acid sequences are shown with the names of the host organisms. The scale bar represents the number of changes per amino acid position.
Structure of XynAMG1 was modeled [27] using the protein crystal structure 2cnc.1.A of a closely related thermostable GH10 xylanase from Cellvibrio mixtus as a template. The amino acid sequence stretches in the XynAMG1 molecule predicted to form different secondary structures (alpha-helices, beta-sheet, and loops) are shown in Figure 3(a). Despite the low degrees of amino acid sequence homology of the mature forms of the two enzymes (40.2% identity; 58.6% similarity), the secondary structure profiles at corresponding positions along the amino acid sequences of the two xylanases are highly similar (Figure 3(a)). Three-dimensional structure modeling of XynAMG1 revealed the typical (β/α)8 TIM-barrel fold resembling the shape of a salad bowl (Figures 3(b) and 3(c)) which is similar to other known GH10 family xylanases [24].
Figure 3.

Structure of XynAMG1 modeled using crystal structure of a closely related GH10 endo-xylanase (2cnc.1.A) as a template. (a) Predicted secondary structure of XynAMG1; purple box: alpha-helix; green arrow: beta-sheet. (b, c) Three-dimensional representation of XynAMG1 with alpha-helices, beta-sheets, and loops folded into the typical (β/α)8 TIM-barrel structure; (b) top view; (c) side view. The XynAMG1 xylanase is shown with 2 molecules of β-D-xylopyranose (stick models) on the catalytic face.
3.2. Cloning, Expression, and Purification of XynAMG1 Xylanase
XynAMG1 without the signal peptide was expressed in E. coli with thioredoxin A peptide and hexa-histidine tags fused to its N-terminus to enhance soluble protein folding and facilitate protein purification, respectively. The purified fusion protein was digested with enterokinase to remove the peptide tags and subjected to second round of purification to yield the purified mature XynAMG1 enzyme for characterization experiments. The Trx-His-XynAMG1 fusion enzyme was catalytically active; however, its activity was around half of that of the mature XynAMG1 (data not shown). Figure 4 shows the SDS-PAGE analysis of different forms of the enzyme through successive purification steps. The molecular mass of the purified XynAMG1 after removal of the tags was determined to be around 41.0 kDa which was close to the theoretical value of 40.013 kDa calculated based on the amino acid sequence. Xylanase zymogram analysis displayed a clear band of xylan digestion corresponding with the position of the XynAMG1 protein band (Figure 4).
Figure 4.
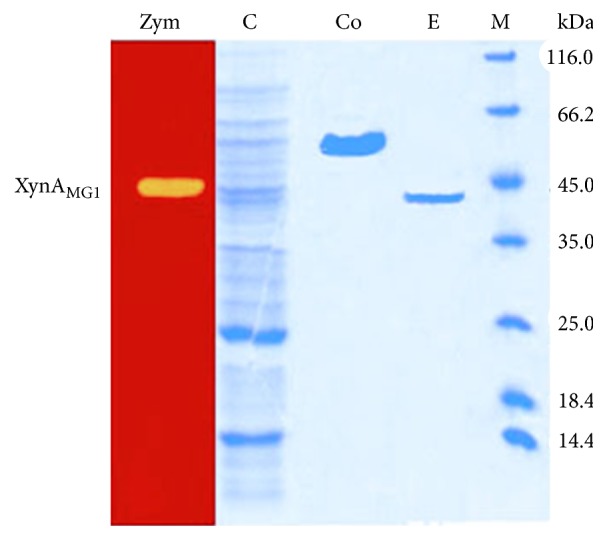
Purification steps of XynAMG1 and zymogram analysis. Proteins were separated with 12% gel SDS-PAGE. Lane (Zym) zymogram, lane (C): total soluble cell lysate from E. coli host expressing XynAMG1, lane (Co): Trx-His-XynAMG1 fusion protein purified by TALON cobalt resin, lane (E): purified XynAMG1 after digestion with enterokinase, and lane (M): protein molecular weight markers.
3.3. Substrate Specificity
The purified XynAMG1 was found to exert its highest hydrolytic activity towards the beechwood xylan while the activity towards oat-spelt xylan was 94% relative to that of the beechwood xylan. No activity could be measured with α-cellulose, carboxymethyl cellulose, starch, and β-glucan as substrates. Upon testing with synthetic chromogenic substrates, XynAMG1 could hydrolyze only the 4-nitrophenyl-β-D-xylopyranoside while 4-nitrophenyl-β-D-cellobioside or 4-nitrophenyl-α-D-galactopyranoside could not serve as substrates (Table 1). The relatively specific xylanase property of XynAMG1 is in contrast with most xylanases of family 10 which have both xylanase and cellulase activities. However, a number of xylanases of family 10 showed only xylanase activity such as xylanase Xyn10N18 derived from a bovine rumen metagenomic library [28] and Xyn10J from a compost metagenomic library which showed negligible hydrolytic activity against carboxymethyl cellulose [18]. Added to the latter group is XynAMG1 isolated in this study and the possession of the cellulase-free xylanase activity makes it potentially very useful for biobleaching of pulps [18].
Table 1.
Substrate specificity of the purified recombinant XynAMG1
| Substratea | Relative activity (%)b |
|---|---|
| Beechwood xylan | 100 |
| Oat-spelt xylan | 94 |
| Carboxymethyl cellulose | 0 |
| α-Cellulose | 0 |
| Starch | 0 |
| Barley β-glucan | 0 |
| 4-nitrophenyl-β-D-xylopyranoside | 55 |
| 4-nitrophenyl-β-D-cellobioside | 0 |
| 4-nitrophenyl-α-D-galactopyranoside | 0 |
aThe test concentration for polysaccharide substrates was 1% (w/v) while that for the synthetic chromogenic substrates was 4 mg/mL. bThe activity towards beechwood xylan which was the highest activity was defined as 100%. All the values are means of three replications.
TLC analysis of the reaction mixture of XynAMG1 catalyzed hydrolysis of beechwood xylan, compared with the results reported for the novel family 10 xylanase Xyn10J from a compost metagenomic library [18], suggested that xylobiose (X2) and xylotetraose (X4) were two major end products, and xylose (X1) was produced as a minor product (Figure 5(a)). XynAMG1 showed high hydrolytic activity on 4-nitrophenyl-β-D-xylopyranoside (pNP-X), a synthetic chromogenic substrate, releasing equimolar of two products 4-nitrophenol and xylose. The extent of the reaction is determined by adding 1 mL of 1 M Na2CO3 to stop the reaction and turn the 4-nitrophenol to yellow color which was quantified by measuring the absorbance at 400 nm [17]. The present xylose as a reaction product was shown by TLC (Figure 5(b)). XynAMG1 effectively digested the synthetic substrate 4-nitrophenyl-β-D-xylopyranoside releasing large amount of xylose. However, upon digestion of beechwood xylan, a natural substrate, the major end product was not xylose (X1) but the xylotetraose (X4) and xylobiose (X2) suggested that XynAMG1 could catalyze transglycosylation reactions. Xylanases with transglycosylation activity have been previously reported in the GH10 family, such as Xyn10J from compost metagenome and XynB from Thermotoga maritima and several GH11 xylanases such as Xyn1 from Paenibacillus sp. W-61, TfxA from Thermomonospora fusca, AnxA from Aspergillus niger, and XynA from Paecilomyces thermophila [18]. With the transglycosylation activity, XynAMG1 can be useful in the application for the synthesis of alkyl oligoglycosides [18].
Figure 5.

Thin-layer chromatography analysis of end products of XynAMG1 catalyzed hydrolysis of beechwood xylan (a) and 4-nitrophenyl-β-D-xylopyranoside (b). X1, xylose; X2, xylobiose; X3, xylotriose; X4, xylotetraose; X5, xylopentaose; X6, xylohexaose; Std, molecular weight standard mix of X1 to X6; pNP-X, 4-nitrophenyl-β-D-xylopyranoside.
3.4. Effects of pH and Temperature on Enzyme Activity and Stability
The purified XynAMG1 showed the typical bell-shaped pH profile with an optimal pH of 6. It retained 32% of its initial activity at pH 9 and completely lost all of its activity at pH 10 (Figure 6(a)). This is consistent with xylanases previously reported [26, 29] and with the physiological function of xylanases in the cecum of broiler chickens since the pH values in cecum broiler chicken are generally 5.5-6 [30]. The xylanase XynAMG1 enzyme was stable between pH 5.0 and 8.0 for 60 min, retaining around 70% of its activity at pH 8.0 (Figure 6(b)). It was fairly stable at high pH similar to other bacterial xylanases [26, 31].
Figure 6.

Effects of pH and temperature on the activity and the stability of the purified XynAMG1. (a) Effect of pH on activity at pH 3.0 to 11.0. The maximum activity was detected at pH 5.5 and was taken as 100%. (b) The pH stability of XynAMG1, incubated at pH 3.0 to pH 11.0 for 30 min and 60 min, at 50°C. (c) Effect of different temperatures on the activity of XynAMG1. The maximum activity was detected at 45°C. (d) Thermal stability after incubation at 45°C to 70°C for various times. The data were presented as mean ± SD (n = 3).
The optimal temperature of XynAMG1 was 45°C (Figure 6(c)), which was close to those found in other metagenomic family 10 xylanases [26, 29]. Most xylanases of family 10 were known to have optimum temperature of 40–80°C [32]. XynAMG1 retained 72% of its activity after incubating at 60°C for 45 min (Figure 6(d)). However, when the temperature was 70°C, the enzyme completely lost the activity. Interestingly, it had around 30% activity even at 100°C in the presence of the substrate (Figure 6(c)). Many enzymes were known to be more stable in the presence of their substrates [33, 34].
3.5. Effect of Metal Ions and Chemicals on the XynMG1 Activity
As shown in Table 2, the XynAMG1 activity was slightly deactivated by metal ions Co2+, Zn2+, Mn2+, and Mg2+ at 2 mM and 10 mM in a concentration-dependent manner. Moderate inhibitions were found with Ca2+ and strong inhibitions were found with Cu2+. The inhibitions of xylanase activity by Cu2+ ion were commonly reported [35, 36].
Table 2.
Effect of metal ions, reducing agents, and ionic and nonionic surfactants on XynAMG1 xylanase activity.
| Agent | 2 mM | 10 mM |
|---|---|---|
| None | 100 ± 0.2 | 100 ± 0.2 |
| Mg2+ | 96.8 ± 0.9 | 92.4 ± 0.7 |
| Ca2+ | 91.4 ± 3.1 | 77.3 ± 2.9 |
| Mn2+ | 99.9 ± 2.5 | 97.3 ± 3.7 |
| Co2+ | 98.5 ± 2.9 | 94.2 ± 2.2 |
| Cu2+ | 86.3 ± 2.2 | 41.4 ± 1.9 |
| Zn2+ | 98.8 ± 0.9 | 95.5 ± 1.4 |
| β-Mercaptoethanol | 95.3 ± 1.0 | 93.6 ± 1.1 |
| DTT | 95.0 ± 1.6 | 94.7 ± 2.0 |
| EDTA | 91.6 ± 3.1 | 87.3 ± 3.1 |
| SDS | 22.1 ± 1.5 | 14.7 ± 1.3 |
| Triton X-100 | 96.2 ± 2.2 | 103.9 ± 1.2 |
| Tween- 80 | 97.9 ± 2.8 | 108.8 ± 2.6 |
The purified XynAMG1 was assayed in the standard assay condition in the presence of 2 mM or 10 mM test agents. The xylanase activity measured in the absence of the test agent was set as 100%. All the values are means of three replications.
Reducing agents such as β-mercaptoethanol and DTT slightly affected the XynAMG1 activity suggesting that xylanase does not seem to need disulfide bonds to achieve the hydrolysis reaction [37].
XynAMG1 enzyme retained more than 87% of its activity after incubation for 60 min with 10 mM of EDTA. The resistance of XynAMG1 to the chelating reagent suggested that it is not a metalloenzyme and no metal ion is essential for XynAMG1 activity. Its stability against the chelating agent, one of the indispensable ingredient in detergent formulations, is of great importance and worth further investigation on its application.
The anionic surfactant SDS which is well-known to cause protein denaturation severely deactivated XynAMG1. Many xylanases were known to be strongly affected by SDS. These include xylanase rMxyl from compost-soil metagenome, xylanase from Burkholderia sp. DMAX, Aspergillus awamori VTCC-F312, and Aspergillus giganteus [9, 38–40]. This indicated that hydrophobic interactions are important in maintaining the structure of XynAMG1.
Nonionic surfactants like Triton X-100 and Tween 80 at 2 mM showed mild inhibitory effects. However at 10 mM, they slightly enhanced the XynAMG1 activity which are in good agreement with other recombinant xylanases [41].
3.6. Effects of Sodium Chloride
Purified XynAMG1 showed excellent salt tolerance. While NaCl was not required for its xylanase activity, XynAMG1 could function at more than 96% of its activity in the presence of 1 to 4 M NaCl for at least 2 h (Figure 7).
Figure 7.
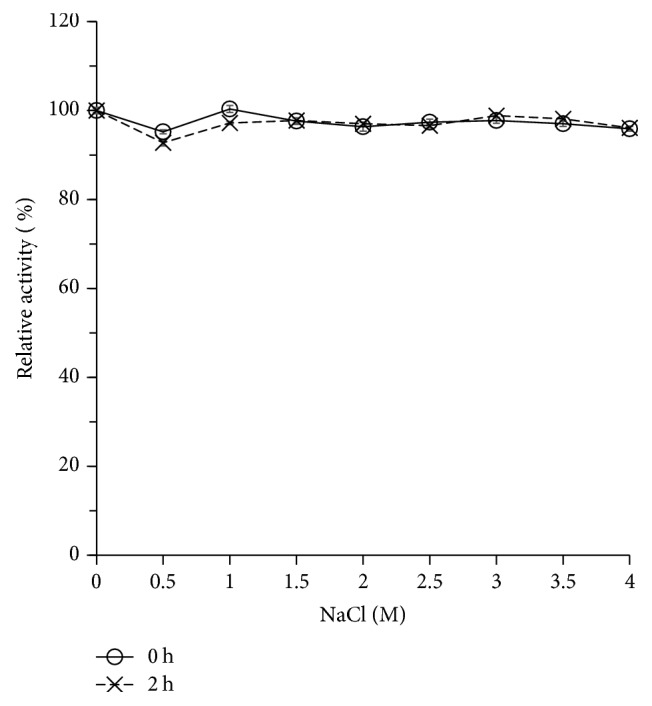
Effects of NaCl on XynAMG1 activity and stability. The xylanase activity at the starting time and without NaCl added was taken as 100%. The data were presented as mean ± SD (n = 3) (for experimental details, please see the main text).
High salt-tolerant and halophilic xylanases have been reported. Recently, a xylanase from Massilia sp. RBM26 isolated from feces of Rhinopithecus bieti was reported to maintain around 86% activity in 5 M NaCl for 1 h [26]. Xylanase from Aspergillus giganteus was shown to tolerate to up to 20% (3.4 M) of NaCl [38]. A cold-active xylanase from Glaciecola mesophila KMM 241 exhibited its highest activity at 0.5 M NaCl and retained 90% of the activity in 2.5 M NaCl at its optimal temperature of 30°C [35]. A xylanase from a marine bacterium Bacillus subtilis cho40 was reported to be greatly activated to 140% when preincubated with 0.5 M NaCl for 4 h [19]. A cold-active and halo-tolerant Xyn10A xylanase from Bacillus sp. SN5 was reported to exhibit the highest activity (134%) in 0.5 M NaCl and retain 90% activity in 2.5 M NaCl [42]. In comparison to the above-mentioned xylanases, our XynAMG1 reported here has the highest salt-tolerance property.
3.7. Effects of Organic Solvent on Xylanase XynAMG1 Activity
The XynAMG1 activity was only slightly affected by common water-miscible organic solvents like acetone, methanol, ethanol, and 1-propanol at the concentration of up to 5 M (Figure 8). At the highest concentration tested (5 M), acetone and methanol showed nearly no effect on XynAMG1 activity which is similar to the xylanase from Streptomyces rameus L2001 [43]. Comparing among alcohols, 1-propanol and ethanol which have lower polarity index values (and thus more hydrophobic) than methanol slightly inhibited XynAMG1 by 10% and 14%, respectively. These were similar to those reported for a xylanase (XynA) from Clostridium cellulovorans [44]. Hydrophobic interactions with the dissolved organic solvent molecules seem to be the major factor affecting the protein stability and the enzyme activity in this case [45].
Figure 8.
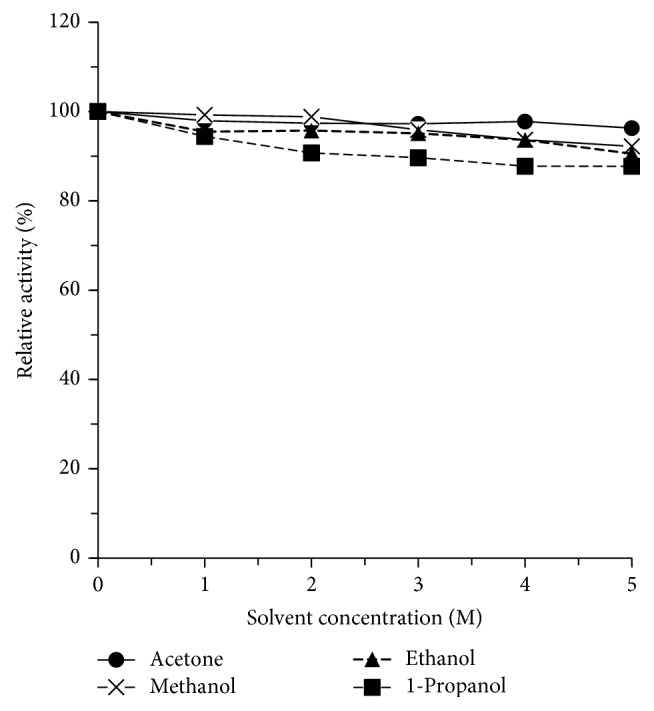
Effects of water-miscible organic solvent on XynAMG1 xylanase activity and stability. The xylanase activity at the starting time and without organic solvent added was taken as 100%. The data were presented as mean ± SD (n = 3) (for experimental details, please see the main text).
In addition to microbial xylanases mentioned in the above discussions, biochemical properties of bacterial GH10 xylanases especially those from animal intestines are listed in Table 3 for comparison with the XynAMG1 xylanase described in this study. XynAMG1 is shown to be better tolerance to organic solvent and high salt (NaCl) concentration than other xylanases compared.
Table 3.
Biochemical properties of GH10 xylanases from animal intestinal bacteria compared with XynAMG1.
| Source | Xylanase name | Optimal | Stability [residual activity (%)] |
Tolerance [residual activity (%)] |
Reference | |||
|---|---|---|---|---|---|---|---|---|
| Temp (°C) | pH | Temp | pH | Solvent | NaCl salt | |||
| Microbacterium trichothecenolyticum HY-17 from Gryllotalpa orientalis gut | rXylH | 60 | 9.0 | 55°C, 30% | pH 5.5–10, 80% | ND | ND | [46] |
|
| ||||||||
| Paenibacillus macerans IIPSP3 from termite gut | IIPSP3 | 60 | 4.5 | 90°C, 70% | pH 3.5, 40% pH 9.5, 67% |
ND | 10 mM, 119% | [47] |
|
| ||||||||
| Sphingobacterium sp. TN19 from Batocera horsfieldi larvae gut | XynA19 | 45 | 6.5 | 40°C, 90% | ND | ND | 10 mM, 95% | [48] |
|
| ||||||||
| Cellulosimicrobium sp. HY-13 from earthworm gut | XylK | 55 | 6.0 | ND | ND | ND | ND | [49] |
|
| ||||||||
| Massilia sp. RBM26 from Rhinopithecus bieti feces | XynRBM26 | 45 | 5.5 | 30–50°C, 62% | pH 5.5–10.0, 80% | ND | 5 M, 86% | [26] |
|
| ||||||||
| Caldicellulosiruptor bescii from geothermally heated freshwater pool | CbXyn10B | 70 | 7.2 | 60–75°C, 60% |
pH 8.0, 50% | ND | ND | [50] |
|
| ||||||||
| Bacteroides xylanisolvens from human gut | XB1A | 37 | 6.0 | 48°C, 80% | pH 9.0, 50% pH 5.0, 50% |
ND | ND | [51] |
|
| ||||||||
| Chicken gut metagenome | XynAMG1 | 45 | 6.0 | 60°C, 72% | pH 3, 25% pH 8, 70% |
5 M acetone, 98% 5 M ethanol, 86% |
4 M, 96% | This study |
ND = not determined.
4. Conclusion
XynAMG1 derived from a chicken cecum metagenome is a new member of the GH10 family xylanase related to the xylanases from Prevotella sp. It is resistant to metal ions, reducing agents, and certain detergents, and highly tolerant to high concentrations of salt and water-miscible organic solvents. These make XynAMG1 a potential candidate for applications in the enzymatic processes operated at high concentration of salt and organic solvent such as in food and biofuels industries.
The salt-tolerant xylanase property of XynAMG1 can be useful in the processing of sea food and food with a high salt content which contain 0.5 to 2.5 M NaCl, such as marine algae, pickles, and sauce [35]. Furthermore, food materials washing, food processing, and fermentation under high salt condition could reduce cost because sterilization is not required [26]. In bakery industry, salt-tolerant XynAMG1 xylanase has potential applications in increasing the strength of the dough and adding flavor to baked goods [52].
Modern biofuel (bioethanol) production from lignocellulose biomass employs consolidated bioprocess where both the saccharification (enzymatic hydrolysis of cellulose and hemicellulose components to simple sugar) and fermentation steps take place within the same bioreactor, which makes the process more economical [44]. Tolerance to the carried over biomass pretreatment agents (acid, alkali, and inhibitors) and ethanol tolerance have been identified as two key elements for the enzymes in this consolidated process. The biomass saccharification enzymes, including xylanase, must be able to withstand and function well in the presence of 23 to 63 g/L (0.5–1.4 M) ethanol produced from the fermentation step. XynAMG1 can be a potential candidate for this application as it could tolerate up to 5 M ethanol.
XynAMG1 functions optimally at the temperature and the pH range of the chicken intestine, its native environment where its gene was retrieved by metagenomic cloning. This xylanase has potential utility in animal feed to improve nutrient digestibility and growth performance for animals, especially broiler chickens and hens [53].
Acknowledgments
Suthep Wiyakrutta was supported by Mahidol University research grant. Hind AL-Darkazali was supported by the Scholarships for International Ph.D. Students, Faculty of Science, and the Research Assistantships for Ph.D. Students, Faculty of Graduate Studies, Mahidol University.
Conflicts of Interest
The authors declare that there are no conflicts of interest regarding the publication of this article.
References
- 1.Adrio J. L., Demain A. L. Microbial enzymes: tools for biotechnological processes. Biomolecules. 2014;4(1):117–139. doi: 10.3390/biom4010117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Bhardwaj A., Mahanta P., Ramakumar S., Ghosh A., Leelavathi S., Reddy V. S. Emerging role of N- and C-terminal interactions in stabilizing (β/a)8 fold with special emphasis on Family 10 xylanases. Computational and Structural Biotechnology Journal. 2012;2(3, article e201209014) doi: 10.5936/csbj.201209014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Adeola O., Cowieson A. J. Board-invited review: opportunities and challenges in using exogenous enzymes to improve nonruminant animal production. Journal of Animal Science. 2011;89(10):3189–3218. doi: 10.2527/jas.2010-3715. [DOI] [PubMed] [Google Scholar]
- 4.Verma D., Satyanarayana T. Molecular approaches for ameliorating microbial xylanases. Bioresource Technology. 2012;117:360–367. doi: 10.1016/j.biortech.2012.04.034. [DOI] [PubMed] [Google Scholar]
- 5.Driss D., Bhiri F., Siela M., Ghorbel R., Chaabouni S. E. Purification and properties of a thermostable xylanase GH 11 from Penicillium occitanis Pol6. Applied Biochemistry and Biotechnology. 2012;168(4):851–863. doi: 10.1007/s12010-012-9824-3. [DOI] [PubMed] [Google Scholar]
- 6.Gasparic A., Martin J., Daniel A. S., Flint H. J. A xylan hydrolase gene cluster in prevotella ruminicola B14: sequence relationships, synergistic interactions, and oxygen sensitivity of a novel enzyme with exoxylanase and β-(1,4)-xylosidase activities. Applied and Environmental Microbiology. 1995;61(8):2958–2964. doi: 10.1128/aem.61.8.2958-2964.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Gu Y., Ding Y., Ren C., et al. Reconstruction of xylose utilization pathway and regulons in firmicutes. BMC Genomics. 2010;11(1, article 255) doi: 10.1186/1471-2164-11-255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Fierens E., Rombouts S., Gebruers K., et al. TLXI, a novel type of xylanase inhibitor from wheat (Triticum aestivum) belonging to the thaumatin family. Biochemical Journal. 2007;403(3):583–591. doi: 10.1042/BJ20061291. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Shi H., Zhang Y., Li X., et al. A novel highly thermostable xylanase stimulated by Ca2+ from thermotoga thermarum: cloning, expression and characterization. Biotechnology for Biofuels. 2013;6:1–9. doi: 10.1186/1754-6834-6-26. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Mead G. C. Microbes of the avian cecum: types present and substrates utilized. Journal of Experimental Zoology. 1989;252:48–54. doi: 10.1002/jez.1402520508. [DOI] [PubMed] [Google Scholar]
- 11.Zhao S., Wang J., Bu D., et al. Novel glycoside hydrolases identified by screening a chinese holstein dairy cow rumen-derived metagenome library. Applied and Environmental Microbiology. 2010;76(19):6701–6705. doi: 10.1128/AEM.00361-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Sergeant M. J., Constantinidou C., Cogan T. A., Bedford M. R., Penn C. W., Pallen M. J. Extensive microbial and functional diversity within the chicken cecal microbiome. PLoS ONE. 2014;9(3) doi: 10.1371/journal.pone.0091941.e91941 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Tu Z., He G., Li K. X., et al. An improved system for competent cell preparation and high efficiency plasmid transformation using different Escherichia coli strains. Electronic Journal of Biotechnology. 2005;8(1):113–120. doi: 10.2225/vol8-issue1-fulltext-8. [DOI] [Google Scholar]
- 14.Vikramathithan J., Nirmal Kumar G., Muthuraman P., Srikumar K. Purification and characterization of thermophilic xylanase isolated from the xerophytic-Cereus pterogonus SP. The Protein Journal. 2010;29(7):481–486. doi: 10.1007/s10930-010-9276-y. [DOI] [PubMed] [Google Scholar]
- 15.Zheng H.-C., Sun M.-Z., Meng L.-C., et al. Purification and characterization of a thermostable xylanase from Paenibacillus sp. NF1 and its application in xylooligosaccharides production. Journal of Microbiology and Biotechnology. 2014;24(4):489–496. doi: 10.4014/jmb.1312.12072. [DOI] [PubMed] [Google Scholar]
- 16.Miller G. L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Analytical Chemistry. 1959;31(3):426–428. doi: 10.1021/ac60147a030. [DOI] [Google Scholar]
- 17.Gallardo O., Diaz P., Pastor F. I. J. Characterization of a Paenibacillus cell-associated xylanase with high activity on aryl-xylosides: a new subclass of family 10 xylanases. Applied Microbiology and Biotechnology. 2003;61(3):226–233. doi: 10.1007/s00253-003-1239-1. [DOI] [PubMed] [Google Scholar]
- 18.Jeong Y. S., Na H. B., Kim S. K., et al. Characterization of Xyn10J, a novel family 10 xylanase from a compost metagenomic library. Applied Biochemistry and Biotechnology. 2012;166(5):1328–1339. doi: 10.1007/s12010-011-9520-8. [DOI] [PubMed] [Google Scholar]
- 19.Khandeparker R., Verma P., Deobagkar D. A novel halotolerant xylanase from marine isolate Bacillus subtilis cho40: Gene cloning and sequencing. New Biotechnology. 2011;28(6):814–821. doi: 10.1016/j.nbt.2011.08.001. [DOI] [PubMed] [Google Scholar]
- 20.Guimaraes N. C. D. A., Sorgatto M., Peixoto-Nogueira S. D. C., et al. Bioprocess and biotechnology: effect of xylanase from Aspergillus niger and Aspergillus flavus on pulp biobleaching and enzyme production using agroindustrial residues as substract. SpringerPlus. 2013;2(1, article 380):1–7. doi: 10.1186/2193-1801-2-380. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.da Silva L., Terrasan C. R. F., Carmona E. C. Purification and characterization of xylanases from Trichoderma inhamatum. Electronic Journal of Biotechnology. 2015;18(4):307–313. doi: 10.1016/j.ejbt.2015.06.001. [DOI] [Google Scholar]
- 22.Pradeep G. C., Choi Y. H., Choi Y. S., et al. A novel thermostable cellulase free xylanase stable in broad range of pH from Streptomyces sp. CS428. Process Biochemistry. 2013;48(8):1188–1196. doi: 10.1016/j.procbio.2013.06.007. [DOI] [Google Scholar]
- 23.Gao Z., Ruan L., Chen X., Zhang Y., Xu X. A novel salt-tolerant endo-β-1,4-glucanase Cel5A in Vibrio sp. G21 isolated from mangrove soil. Applied Microbiology and Biotechnology. 2010;87(4):1373–1382. doi: 10.1007/s00253-010-2554-y. [DOI] [PubMed] [Google Scholar]
- 24.Henrissat B., Callebaut I., Fabrega S., Lehn P., Mornon J.-P., Davies G. Conserved catalytic machinery and the prediction of a common fold for several families of glycosyl hydrolases. Proceedings of the National Academy of Sciences of the United States of America. 1995;92(15):7090–7094. doi: 10.1073/pnas.92.15.7090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Roberge M., Dupont C., Morosoli R., Shareck F., Kluepfel D. Asparagine-127 of xylanase A from Streptomyces lividans, a key residue in glycosyl hydrolases of superfamily 4/7: kinetic evidence for its involvement in stabilization of the catalytic intermediate. Protein Engineering. 1997;10(4):399–403. doi: 10.1093/protein/10.4.399. [DOI] [PubMed] [Google Scholar]
- 26.Xu B., Dai L., Li J., et al. Molecular and biochemical characterization of a novel xylanase from Massilia sp. RBM26 isolated from the feces of Rhinopithecus bieti. Journal of Microbiology and Biotechnology. 2015;26(1):9–19. doi: 10.4014/jmb.1504.04021. [DOI] [PubMed] [Google Scholar]
- 27.Biasini M., Bienert S., Waterhouse A., et al. SWISS-MODEL: modelling protein tertiary and quaternary structure using evolutionary information. Nucleic Acids Research. 2014;42(1):W252–W258. doi: 10.1093/nar/gku340. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Gong X., Gruniniger R. J., Forster R. J., Teather R. M., McAllister T. A. Biochemical analysis of a highly specific, pH stable xylanase gene identified from a bovine rumen-derived metagenomic library. Applied Microbiology and Biotechnology. 2013;97(6):2423–2431. doi: 10.1007/s00253-012-4088-y. [DOI] [PubMed] [Google Scholar]
- 29.Alvarez T. M., Goldbeck R., Santos C. R. D., et al. Development and biotechnological application of a novel endoxylanase family GH10 identified from sugarcane soil metagenome. PLoS ONE. 2013;8(7, article e70014) doi: 10.1371/journal.pone.0070014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Van Der Wielen P. W. J. J., Biesterveld S., Notermans S., Hofstra H., Urlings B. A. P., Van Knapen F. Role of volatile fatty acids in development of the cecal microflora in broiler chickens during growth. Applied and Environmental Microbiology. 2000;66(6):2536–2540. doi: 10.1128/AEM.66.6.2536-2540.2000. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Takahashi Y., Kawabata H., Murakami S. Analysis of functional xylanases in xylan degradation by Aspergillus niger E-1 and characterization of the GH family 10 xylanase XynVII. SpringerPlus. 2013;2, article 447 doi: 10.1186/2193-1801-2-447. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Polizeli M. L. T. M., Rizzatti A. C. S., Monti R., Terenzi H. F., Jorge J. A., Amorim D. S. Xylanases from fungi: properties and industrial applications. Applied Microbiology and Biotechnology. 2005;67(5):577–591. doi: 10.1007/s00253-005-1904-7. [DOI] [PubMed] [Google Scholar]
- 33.Rupert C. S. Photoenzymatic repair of ultraviolet damage in DNA. II. formation of an enzyme-substrate complex. The Journal of general physiology. 1962;45:725–741. doi: 10.1085/jgp.45.4.725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Varga A., Beáta F., Gráczer É., Osváth S., Szilágyi A. N., Vas M. Correlation between conformational stability of the ternary enzyme-substrate complex and domain closure of 3-phosphoglycerate kinase. FEBS Journal. 2005;272(8):1867–1885. doi: 10.1111/j.1742-4658.2005.04618.x. [DOI] [PubMed] [Google Scholar]
- 35.Guo B., Chen X.-L., Sun C.-Y., Zhou B.-C., Zhang Y.-Z. Gene cloning, expression and characterization of a new cold-active and salt-tolerant endo-β-1,4-xylanase from marine Glaciecola mesophila KMM 241. Applied Microbiology and Biotechnology. 2009;84(6):1107–1115. doi: 10.1007/s00253-009-2056-y. [DOI] [PubMed] [Google Scholar]
- 36.Heo S., Kwak J., Oh H.-W., et al. Characterization of an extracellular xylanase in paenibacillus sp. HY-8 isolated from an herbivorous longicorn beetle. Journal of Microbiology and Biotechnology. 2006;16(11):1753–1759. [Google Scholar]
- 37.Margolles A., De los Reyes-Gavilán C. G. Purification and functional characterization of a novel α-L-arabinofuranosidase from Bifidobacterium longum B667. Applied and Environmental Microbiology. 2003;69(9):5096–5103. doi: 10.1128/AEM.69.9.5096-5103.2003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Fialho M. B., Carmona E. C. Purification and characterization of xylanases from Aspergillus giganteus. Folia Microbiologica. 2004;49(1):13–18. doi: 10.1007/BF02931639. [DOI] [PubMed] [Google Scholar]
- 39.Mohana S., Shah A., Divecha J., Madamwar D. Xylanase production by Burkholderia sp. DMAX strain under solid state fermentation using distillery spent wash. Bioresource Technology. 2008;99(16):7553–7564. doi: 10.1016/j.biortech.2008.02.009. [DOI] [PubMed] [Google Scholar]
- 40.Verma D., Kawarabayasi Y., Miyazaki K., Satyanarayana T. Cloning, expression and characteristics of a novel alkalistable and thermostablexylanase encoding gene ( Mxyl ) retrieved from compost-soil metagenome. PLoS ONE. 2013;8(1, article e52459) doi: 10.1371/journal.pone.0052459. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Rahman M. A., Choi Y. H., Pradeep G. C., et al. An alkaline and metallo-protein type endo xylanase from Streptomyces sp. CSWu-1. Biotechnology and Bioprocess Engineering. 2014;19(2):311–319. doi: 10.1007/s12257-013-0782-6. [DOI] [Google Scholar]
- 42.Bai W., Xue Y., Zhou C., Ma Y. Cloning, expression and characterization of a novel salt-tolerant xylanase from Bacillus sp. SN5. Biotechnology Letters. 2012;34(11):2093–2099. doi: 10.1007/s10529-012-1011-7. [DOI] [PubMed] [Google Scholar]
- 43.Li X., She Y., Sun B., et al. Purification and characterization of a cellulase-free, thermostable xylanase from Streptomyces rameus L2001 and its biobleaching effect on wheat straw pulp. Biochemical Engineering Journal. 2010;52(1):71–78. doi: 10.1016/j.bej.2010.07.006. [DOI] [Google Scholar]
- 44.Morrison D., van Dyk J. S., Pletschke B. I. The effect of alcohols, lignin and phenolic compounds on the enzyme activity of Clostridium cellulovorans XynA. Bioresources. 2011;6:3132–3141. [Google Scholar]
- 45.Lemos J. L. S., Bon E. P. S., Santana M. D. F. E., Pereira N., Jr. Thermal stability of xylanases produced by Aspergillus awamori. Brazilian Journal of Microbiology. 2000;31:206–211. [Google Scholar]
- 46.Bae K. S., Sung C. K., Rhee Y. H., et al. Novel alkali-tolerant GH10 endo-ß-1, 4-xylanase with broad substrate specificity from Microbacterium trichothecenolyticum HY-17, a gut bacterium of the mole cricket Gryllotalpaorientalis. Journal of Microbiology and Biotechnology. 2014;24(7):943–953. doi: 10.4014/jmb.1405.05032. [DOI] [PubMed] [Google Scholar]
- 47.Dheeran P., Nandhagopal N., Kumar S., Jaiswal Y. K., Adhikari D. K. A novel thermostable xylanase of Paenibacillus macerans IIPSP3 isolated from the termite gut. Journal of Industrial Microbiology and Biotechnology. 2012;39(6):851–860. doi: 10.1007/s10295-012-1093-1. [DOI] [PubMed] [Google Scholar]
- 48.Zhou J., Huang H., Meng K., et al. Molecular and biochemical characterization of a novel xylanase from the symbiotic Sphingobacterium sp. TN19. Applied Microbiology and Biotechnology. 2009;85(2):323–333. doi: 10.1007/s00253-009-2081-x. [DOI] [PubMed] [Google Scholar]
- 49.Kim D. Y., Han M. K., Lee J. S., et al. Isolation and characterization of a cellulase-free endo-β-1,4-xylanase produced by an invertebrate-symbiotic bacterium, Cellulosimicrobium sp. HY-13. Process Biochemistry. 2009;44(9):1055–1059. doi: 10.1016/j.procbio.2009.05.005. [DOI] [Google Scholar]
- 50.An J., Xie Y., Zhang Y., et al. Characterization of a thermostable, specific GH10 xylanase from Caldicellulosiruptor bescii with high catalytic activity. Journal of Molecular Catalysis B: Enzymatic. 2015;117:13–20. doi: 10.1016/j.molcatb.2015.04.003. [DOI] [Google Scholar]
- 51.Mirande C., Mosoni P., Béra-Maillet C., Bernalier-Donadille A., Forano E. Characterization of Xyn10A, a highly active xylanase from the human gut bacterium Bacteroides xylanisolvens XB1A. Applied Microbiology and Biotechnology. 2010;87(6):2097–2105. doi: 10.1007/s00253-010-2694-0. [DOI] [PubMed] [Google Scholar]
- 52.Butt M. S., Tahir-Nadeem M., Ahmad Z., Sultan M. T. Xylanases and their applications in baking industry. Food Technology and Biotechnology. 2008;46(1):22–31. [Google Scholar]
- 53.Lei Z., Shao Y., Yin X., Yin D., Guo Y., Yuan J. Combination of xylanase and debranching enzymes specific to wheat arabinoxylan improve the growth performance and gut health of broilers. Journal of Agricultural and Food Chemistry. 2016;64(24):4932–4942. doi: 10.1021/acs.jafc.6b01272. [DOI] [PubMed] [Google Scholar]