

Abstract
Objective
Epstein-Barr virus (EBV) encodes at least 25 pri-microRNAs (miRNAs) in two regions of its DNA genome, BART and BHRF. B95-8, an EBV reference strain, has a deletion in the BART region. However, no information is available on the deletions or mutations in the BART and BHRF regions in clinical samples of EBV-associated lymphoma.
Results
Nine DNA fragments encoding miR-BARTs and two coding miR-BHRF1s were amplified by PCR from DNA samples extracted from 16 cases of EBV-associated lymphoma. All the PCR products were sequenced directly. DNA fragments encoding miR-BARTs and miR-BHRF1-1 were successfully amplified from all samples. An adenine-to-guanine mutation in the DNA fragment encoding miR-BART2-3p was detected in four of the 16 cases, and a cytosine-to-thymidine mutation in the DNA fragment encoding miR-BART11-3p was detected in one of the 16 samples. These mutations were not associated with any histological categories of lymphoma. In conclusion, mutations were rarely observed in the DNA encoding viral miRNAs in cases of lymphoma. This suggests that the DNA sequences of EBV-encoded miR-BARTs and miR-BHRF1-1 are conserved in EBV-associated lymphoma.
Keywords: Epstein-Barr virus, miRNA, Lymphoma, PCR, Sequence
Introduction
Epstein-Barr virus (EBV) is a common herpesvirus that infects more than 90% of all adults throughout the world [1]. Primary EBV infections in adults cause infectious mononucleosis [2]. EBV is also associated with several types of lymphoma and cancer. According to a recent classification of lymphoma, EBV is associated with certain cases of Burkitt lymphoma, diffuse large-B-cell lymphoma (DLBCL) in immunodeficient patients, EBV-positive DLBCL of the elderly (ELD), DLBCL consistent with methotrexate (MTX)-associated lymphoproliferative disorder, NK/T-cell lymphoma (NKT), plasmablastic lymphoma, Hodgkin lymphoma, etc. [3]. Recently, it has been demonstrated that EBV encodes at least 25 pre-microRNAs (miRNAs) in its genome and that 44 mature miRNAs are registered in miRBase (http://www.mirbase.org/) from these pre-miRNAs [4, 5]. miRNAs are small RNAs (20–22 nucleotides) with various demonstrated biological functions in cells [6]. Representative functions of miRNAs include RNA silencing and the posttranscriptional regulation of gene expression by binding their target mRNAs [7]. The miRNAs of EBV are encoded in two primary transcripts, BHRF1 and BART. BHRF1 encodes miR-BHRF1-1, 1-2, and 1-3, which are expressed in EBV latency III infection [8]. BART encodes 22 miR-BARTs in three clusters [9]. Various functions of the EBV-encoded miRNAs have been reported. However, B95-8, an EBV reference strain, has a deletion in the BART DNA [10], although it displays full transformational activity in human B cells. In contrast, a study of insertion mutants of miR-BARTs in a recombinant EBV (B95-8) genome demonstrated that miR-BARTs promote cell-cycle progression and prevent the apoptosis of primary human B cells [11]. Another study showed that miR-BHRF1s accelerate B-cell expansion with low latent gene expression levels and reduce the viral antigenic load, which potentially facilitate the persistence of the virus in the infected host [12]. Therefore, the functions of the miR-BARTs and miR-BHRF1s in B-cell transformation differ.
A recent study of the full-genome sequences of EBV using next-generation sequencing demonstrated that BART and other regions were deleted in the full EBV genomes of some EBV-positive cell lines [13]. However, because the BART region contains a repetitive sequence, the next-generation sequencer did not correctly read the full sequence of BART. Therefore, there is no information about the deletions or mutations in the BART and BHRF regions in clinical samples of EBV-associated lymphoma.
In this study, DNA fragments encoding miR-BARTs and miR-BHRF1 were amplified by PCR from DNA samples extracted from EBV-positive lymphoma patients and were directly sequenced to determine the mutation, deletion, and/or insertion status of the regions encoding miRNAs.
Main text
Clinical specimens
Sixteen frozen samples of EBV-associated lymphoma were examined. All samples were obtained in biopsy or autopsy for pathological diagnosis. The samples included seven ELD, three AIDS-related DLBCLs (ARL), two NKTs, two low-grade B-cell lymphomas, one MTX-related lymphoma (MTX), and one plasmablastic lymphoma (PBL). All the patients were residents of Tokyo or prefectures neighboring Tokyo. The histological diagnosis of lymphoma was based on the 4th edition of the World Health Organization classification of lymphoma [3]. All cases were confirmed as positive for EBV infection with the in situ hybridization of EBV-encoded small RNAs (EBER). In situ hybridization was performed using EBER PNA Probe/Fluorescein and PNA-ISH Detection Kit (Dako, Glostrup, Denmark) according to the manufacturer’s instructions.
PCR amplification and DNA sequencing
DNA was extracted from frozen samples from lymphoma patients, with the DNeasy Blood & Tissue Kit (Qiagen, Hilden, Germany), according to the manufacturer’s protocol. Fragments containing the all EBV-miRNA genes were amplified with PCR from the DNA samples using the primers listed in Table 1. PCR amplification was performed at 94 °C for 5 min (one cycle); 94 °C for 30 s, 55 °C for 30 s, and 72 °C for 30 s (35 cycles); and 72 °C for 10 min (one cycle) using the GeneAmp PCR System 9700 (Applied Biosystems, Foster City, CA, USA). The PCR products were purified with the QIAquick PCR Purification Kit (Qiagen), and then directly sequenced with an ABI 3130 Genetic Analyzer (Applied Biosystems) using the BigDye Terminator Ready Reaction Kit (Applied Biosystems), according to the manufacturer’s instructions.
Table 1.
List of PCR primers
| Set | Targets | Strand | Name of primer | Start | End | Sequence (5′-3′) | Size of PCR product |
|---|---|---|---|---|---|---|---|
| E1 | miR-BARTs3,4,1 | Forward | EBV139081F | 139,081 | 139,100 | tccctgtaaacacacaccac | 440 |
| Reverse | EBV139520R | 139,520 | 139,501 | ttctacatcatgcctggttc | |||
| E2 | miR-BARTs15,5,16,17,6 | Forward | EBV139501F | 139,501 | 139,520 | gaaccaggcatgatgtagaa | 690 |
| Reverse | EBV140190R | 140,190 | 140,171 | tttagatctgtggttacatg | |||
| E3 | miR-BARTs21,18 | Forward | EBV145451F | 145,451 | 145,470 | ttagatgttagctttgtgtt | 590 |
| Reverse | EBV146040R | 146,040 | 146,021 | ggcccaaaccttcgcagcag | |||
| E4 | miR-BART7 | Forward | EBV145911F | 145,911 | 145,930 | ttgttgccgttgaaagacgg | 610 |
| Reverse | EBV146520R | 146,520 | 146,501 | tggccacactaaacacacaa | |||
| E5 | miR-BARTs8,9 | Forward | EBV146701F | 146,701 | 146,720 | ttatttgggttacaagacct | 350 |
| Reverse | EBV147050R | 147,050 | 147,031 | cacaatgaaacccaaagccc | |||
| E6 | miR-BARTs22, 10,11 | Forward | EBV147131F | 147,131 | 147,150 | cggttgtcacaggtgctaga | 500 |
| Reverse | EBV147630R | 147,630 | 147,611 | cgtgaaaggcactccagaat | |||
| E7 | miR-BARTs12,19,20 | Forward | EBV147871F | 147,871 | 147,890 | acctaagacccgcccatcac | 550 |
| Reverse | EBV148420R | 148,420 | 148,401 | ccaaaggacccgggatcacg | |||
| E8 | miR-BARTs13, 14 | Forward | EBV148461F | 148,461 | 148,480 | catcttgacgttggaatgtc | 360 |
| Reverse | EBV148820R | 148,820 | 148,801 | ctcctgggttggcgtttccg | |||
| E9 | miR-BART2 | Forward | EBV152651F | 152,651 | 152,670 | gcagcaaaagaggaacttgc | 350 |
| Reverse | EBV153000R | 153,000 | 152,981 | ggcaaagatccccagcggag | |||
| E10 | miR-BHRF1-1 | Forward | EBV41581F | 41,581 | 41,600 | cctcaccatgacacactaag | 260 |
| Reverse | EBV41840R | 41,840 | 41,821 | ccagatgcacccaacagccc | |||
| E11 | miR-BHRF1-2,3 | Forward | EBV42991F | 42,991 | 43,010 | gggtgacacagtgcccatgc | 330 |
| Reverse | EBV43320R | 43,320 | 43,301 | acactcacctcagttatttc |
Nucleotide numbering is based on GenBank KF373730
Alignment analysis
Sequences were aligned with the Genetyx software (ver. 13, Genetyx, Tokyo, Japan). The reference sequences used were EBV strains AG876 (GenBank accession no. NC_009334), Akata (KC207813), B95-8 (V01555), HL01 (LN824226), HL02 (LN827546), Jijoye (LN827800), L591 (LN827523), Makaku (LN827551), Mutu (KC207814), Raji (KF717093), sLCL-IS1.20 (LN827576), and Wewaki1 (LN827544).
Results
Nine DNA fragments encoding miR-BARTs and two encoding miR-BHRF1 were successfully amplified with PCR from the DNA samples from all 16 patients with EBV-positive lymphoma (Fig. 1). Direct sequencing of the PCR products revealed only two mutations in the DNA fragments encoding pre-miRNAs relative to the reference sequences. An adenine-to-guanine mutation in the DNA fragment encoding miR-BART2-3p was detected in four of 16 cases (25%; Fig. 2a), which included two ELD, one T-cell lymphoma, and one low-grade B-cell lymphoma. A cytosine-to-thymidine mutation was found in a DNA fragment encoding miR-BART11-3p in one (ELD) of the 16 cases (6.25%). The ELD case with a cytosine-to-thymidine mutation in miR-BART11-3p DNA did not have an adenine-to-guanine mutation in the DNA fragment encoding miR-BART2-3p. The mutation in miR-BART11-3p was in the previously reported seed region of the miRNA [14] (Fig. 2b). There were no single-nucleotide polymorphisms, mutations, insertions, or deletions in any of the DNA fragments encoding other pre-miRNAs, including pri-miR-BHRF1s, suggesting that the miRNA sequences are strongly conserved in cases of lymphoma.
Fig. 1.
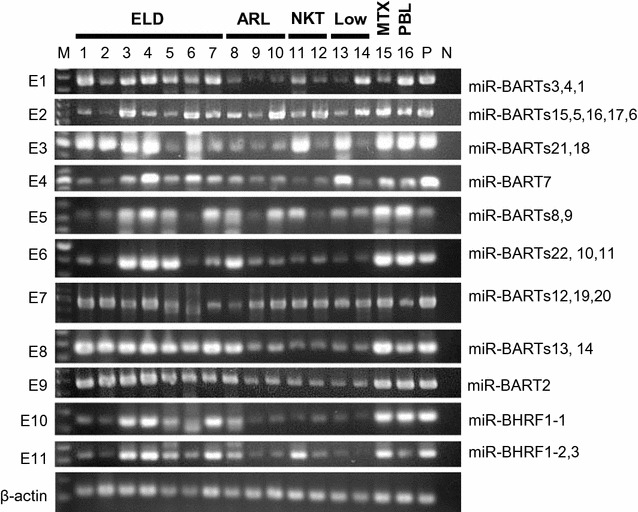
PCR products from EBV in clinical samples of lymphoma. The PCR products were separated electrophoretically in 2% agarose gel and stained with ethidium bromide. M 100-bp ladder molecular weight markers; samples 1–16, P positive control (Raji, an EBV-positive Burkitt lymphoma cell line), and N no-DNA negative control
Fig. 2.
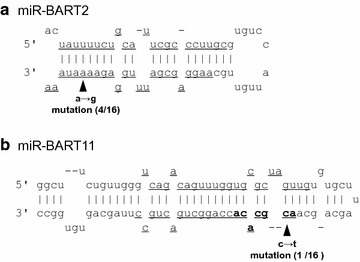
Mutations in pri-miR-BART2 and BART11. Structures of pri-miR-BART2 (a) and pri-miR-BART11 (b) are shown. Black triangles indicate single-nucleotide mutations. Mature miRNAs are indicated with underlining. Seed region of miR-BART11-3p is shown in bold letters
Discussion
The two mutations in the DNA fragment encoding miR-BARTs found in this study were not associated with any histological features of the lymphoma. In a previous study, miR-BART2-3p was not expressed in the serum samples of patients in the acute phase of infectious mononucleosis, whereas miR-BART2-5p was expressed abundantly [15]. Therefore, the mutation in miR-BART2-3p identified in the present study is unlikely to be associated with the lymphomagenesis of a specific category of lymphoma. However, miR-BART11-3p has been reported to be strongly expressed in infectious mononucleosis, gastric cancer, and nasopharyngeal carcinoma [14]. EBV-miR-BART11 plays a crucial role in the promotion of carcinogenesis in gastric cancer and nasopharyngeal carcinoma by inhibiting the tumor-suppressive effects of FOXP1 [14]. The mutation identified in the present study was in the seed region of miR-BART11-3p, suggesting that it affects the role of miR-BART11-3p.
In this study, the genetic conservation of the EBV DNA encoding miRNAs was demonstrated in patients with EBV-associated lymphoma, implying that deletions in BART are rare. The genome of Kaposi’s sarcoma-associated herpesvirus (KSHV) also encodes viral miRNAs [16], which reportedly have important functions in cell transformation and cell-cycle promotion [17, 18]. A study of clinical samples in which the DNA fragments encoding KSHV miRNAs were sequenced detected SNPs in the DNA fragments encoding miRNAs in multicentric Castleman disease and KSHV inflammatory cytokine syndrome, suggesting that sequence variations in KSHV miRNA are associated with these diseases [19]. However, no large deletion of DNA encoding an miRNA has been reported in KSHV-associated diseases. A recent study showed that a defect in the miR-BARTs region, as in B95-8, is uncommon among EBV-positive cell lines, although partial defects have been observed in two other cell lines [13]. However, the partial defects detected in that study were near the assembly gap region introduced by next-generation sequencing, suggesting that the defects were associated with the sequencing technique. The gap region was not PCR amplified in that study [13]. The lack of deletions detected in miR-BART in the present study suggests that these deletions do not occur or occur only rarely in clinical samples.
In conclusion, the sequences of the EBV genome that encode miR-BARTs and miR-BHRF1 are conserved in EBV-associated lymphoma. Further studies comparing cell lines and clinical samples are required to investigate the role of the BART deletions in EBV B95-8, and the association between the mutations in the DNA fragments encoding miR-BARTs and the pathogenesis of lymphoma.
Limitations
The limitation of this study includes small numbers of samples and PCR amplification of limited regions. Therefore, the conclusion of this study may not be generalizable to all EBV-infected cases. Any functional analysis of the mutated miRNAs was not performed in this study. Despite these limitations, these findings suggest genomic conservation of EBV at least in a subset of lymphoma because there is no similar study.
Authors’ contributions
HK conceived and designed the experiments. KS and HF performed the experiments. TH and HH contributed reagents, materials, or analysis tool. KS and HK analyzed the data and wrote the manuscript. All authors read and approved the final manuscript.
Acknowledgements
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
The dataset supporting the conclusions of this article is included within the article.
Ethics approval and consent to participate
Studies using human tissue were performed with the approval of the Institutional Review Board of the National Institute of Infectious Diseases (Approval Nos. 271 and 272) and Tokyo Metropolitan Komagome Hospital (Approval No. 628). Informed consent was not obtained due to the retrospective nature of the investigation conducted in archival biopsy specimens. Identifying information for all samples was removed before analysis for strict privacy protection; this procedure was in accordance with the “Ethical guidelines for epidemiological research” enacted by the Japanese Government.
Funding
This work was financially supported by Research Program on HIV/AIDS (Grant Numbers 16fk0410209j0102, 16fk0410108j0101, and 16fk0410107j0001) from the Agency for Medical Research and Development, and Grants-in-Aid for Scientific Research (C) from the Japan Society for the Promotion of Science (Grant Number 15K08509).
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Abbreviations
- AIDS
acquired immunodeficiency syndrome
- ARL
AIDS-related diffuse large B cell lymphoma
- DLBCL
diffuse large B-cell lymphoma
- EBV
Epstein-Barr virus
- ELD
EBV-positive DLBCL of the elderly
- KSHV
Kaposi sarcoma-associated herpesvirus
- LOW
low-grade B-cell lymphoma
- miRNA
microRNA
- MTX
methotrexate-related lymphoma
- PBL
plasmablastic lymphoma
- PCR
polymerase chain reaction
- SNP
single nucleotide polymorphism
- NKT
NK/T-cell lymphoma
Contributor Information
Keishin Sunagawa, Email: sunagawa@med.showa-u.ac.jp.
Tsunekazu Hishima, Email: hishima@cick.jp.
Hitomi Fukumoto, Email: h-fuku@nih.go.jp.
Hideki Hasegawa, Email: hasegawa@nih.go.jp.
Harutaka Katano, Phone: +81-3-4582-2704, Email: katano@nih.go.jp.
References
- 1.Cohen JI. Epstein-Barr virus infection. N Engl J Med. 2000;343:481–492. doi: 10.1056/NEJM200008173430707. [DOI] [PubMed] [Google Scholar]
- 2.Rickinson AB, Kieff ED. Epstein-Barr virus. In: Knipe DM, Howley PM, editors. Fields virology. Philadelphia: Lippincott Williams & Wilkins; 2007. pp. 2655–2700. [Google Scholar]
- 3.Swerdlow SH, Campo E, Harris NL, Jaffe ES, Pileri SA, Stein H, Thiele J, Vardiman JW. WHO classification of tumours of haematopoetic and lymphoid tissues. 4. Lyon: International Agency for Research on Cancer (IARC); 2008. [Google Scholar]
- 4.Pfeffer S, Zavolan M, Grasser FA, Chien M, Russo JJ, Ju J, John B, Enright AJ, Marks D, Sander C, et al. Identification of virus-encoded microRNAs. Science. 2004;304:734–736. doi: 10.1126/science.1096781. [DOI] [PubMed] [Google Scholar]
- 5.Kincaid RP, Sullivan CS. Virus-encoded microRNAs: an overview and a look to the future. PLoS Pathog. 2012;8:e1003018. doi: 10.1371/journal.ppat.1003018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116:281–297. doi: 10.1016/S0092-8674(04)00045-5. [DOI] [PubMed] [Google Scholar]
- 7.Ambros V. The functions of animal microRNAs. Nature. 2004;431:350–355. doi: 10.1038/nature02871. [DOI] [PubMed] [Google Scholar]
- 8.Amoroso R, Fitzsimmons L, Thomas WA, Kelly GL, Rowe M, Bell AI. Quantitative studies of Epstein-Barr virus-encoded microRNAs provide novel insights into their regulation. J Virol. 2011;85:996–1010. doi: 10.1128/JVI.01528-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Edwards RH, Marquitz AR, Raab-Traub N. Epstein-Barr virus BART microRNAs are produced from a large intron prior to splicing. J Virol. 2008;82:9094–9106. doi: 10.1128/JVI.00785-08. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Baer R, Bankier AT, Biggin MD, Deininger PL, Farrell PJ, Gibson TJ, Hatfull G, Hudson GS, Satchwell SC, Seguin C, et al. DNA sequence and expression of the B95-8 Epstein-Barr virus genome. Nature. 1984;310:207–211. doi: 10.1038/310207a0. [DOI] [PubMed] [Google Scholar]
- 11.Seto E, Moosmann A, Gromminger S, Walz N, Grundhoff A, Hammerschmidt W. Micro RNAs of Epstein-Barr virus promote cell cycle progression and prevent apoptosis of primary human B cells. PLoS Pathog. 2010;6:e1001063. doi: 10.1371/journal.ppat.1001063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Feederle R, Linnstaedt SD, Bannert H, Lips H, Bencun M, Cullen BR, Delecluse HJ. A viral microRNA cluster strongly potentiates the transforming properties of a human herpesvirus. PLoS Pathog. 2011;7:e1001294. doi: 10.1371/journal.ppat.1001294. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Palser AL, Grayson NE, White RE, Corton C, Correia S, Ba Abdullah MM, Watson SJ, Cotten M, Arrand JR, Murray PG, et al. Genome diversity of Epstein-Barr virus from multiple tumor types and normal infection. J Virol. 2015;89:5222–5237. doi: 10.1128/JVI.03614-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Song Y, Li X, Zeng Z, Li Q, Gong Z, Liao Q, Li X, Chen P, Xiang B, Zhang W, et al. Epstein-Barr virus encoded miR-BART11 promotes inflammation-induced carcinogenesis by targeting FOXP1. Oncotarget. 2016;7:36783–36799. doi: 10.18632/oncotarget.6346. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Gao L, Ai J, Xie Z, Zhou C, Liu C, Zhang H, Shen K. Dynamic expression of viral and cellular microRNAs in infectious mononucleosis caused by primary Epstein-Barr virus infection in children. Virol J. 2015;12:208. doi: 10.1186/s12985-015-0441-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Umbach JL, Cullen BR. In-depth analysis of Kaposi’s sarcoma-associated herpesvirus microRNA expression provides insights into the mammalian microRNA-processing machinery. J Virol. 2010;84:695–703. doi: 10.1128/JVI.02013-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Ziegelbauer JM. Functions of Kaposi’s sarcoma-associated herpesvirus microRNAs. Biochim Biophys Acta. 2011;1809:623–630. doi: 10.1016/j.bbagrm.2011.05.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Liang D, Lin X, Lan K. Looking at Kaposi’s sarcoma-associated herpesvirus-host interactions from a microRNA viewpoint. Front Microbiol. 2011;2:271. doi: 10.3389/fmicb.2011.00271. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Ray A, Marshall V, Uldrick T, Leighty R, Labo N, Wyvill K, Aleman K, Polizzotto MN, Little RF, Yarchoan R, et al. Sequence analysis of Kaposi sarcoma-associated herpesvirus (KSHV) microRNAs in patients with multicentric Castleman disease and KSHV-associated inflammatory cytokine syndrome. J Infect Dis. 2012;205:1665–1676. doi: 10.1093/infdis/jis249. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The dataset supporting the conclusions of this article is included within the article.