

Abstract
The majority of Osteosarcoma (OS) patients are treated with a combination of chemotherapy, resection, and limb salvage protocols. These protocols include distraction osteogenesis (DO), which is characterized by direct new bone formation. Cisplatin (CDP) is extensively used for OS chemotherapy and recent studies, using a mouse DO model, have demonstrated that CDP has profound negative effects on bone repair. Recent oncological therapeutic strategies are based on the use of standard cytotoxic drugs plus an assortment of biologic agents. Here we demonstrate that the previously reported CDP-associated inhibition of bone repair can be modulated by the administration of a small molecule p53 inducer (nutlin-3). The effects of nutlin-3 on CDP osteotoxicity were studied using both pre- and post-operative treatment models. In both cases the addition of nutlin-3, bracketing CDP exposure, demonstrated robust and significant bone sparing activity (p < 0.01–0.001). In addition the combination of nutlin-3 and CDP induced equivalent OS tumor killing in a xenograft model. Collectively, these results demonstrate that the induction of p53 peri-operatively protects bone healing from the toxic effects of CDP, while maintaining OS toxicity.
Keywords: cisplatin, nutlin-3, distraction osteogenesis, chemotherapy, limb salvage
There is an increasing interest in new therapeutic options that include a combination of standard cytotoxic drugs and biologic agents in the oncology setting.1 The improvement in quality-of-life and overall survival achieved within the last 10 years highlight the impact of combining standard chemotherapy with novel biologic agents.1 The results of the studies presented here provide a preclinical basis supporting the addition of p53 inducers (e.g., nutlin-3) to the standard cisplatin (CDP) –based therapy used for osteosarcoma (OS) patients.
OS is the most common primary bone cancer that affects children and adolescents.2 The majority of OS patients are treated by a closely timed combination of chemotherapy (often CDP) and tumor resection, in many cases followed by a type of limb salvage procedure (Cure Search: Children’s Oncology Group/COG). Distraction osteogenesis (DO), surgical reconstruction, and endoprostheses are all routinely used for limb salvage, yet the patients continue to be at risk for postoperative complications relating to inherent poor bone repair associated with the potent chemotherapies employed.2–8
In practice, exposure to chemotherapy can occur preoperatively, postoperatively, and even simultaneously with bone repair/regeneration.9 Therefore, to develop and standardize mouse models for CDP toxicity we previously studied the effects of CDP dosing both during and before surgery on bone formation during DO.10 These studies demonstrated that CDP has significant negative peri-operative effects on bone repair in DO.10 This provided the platform to identify a treatment, detailed here, that spares normal cells from the inhibitory effects of CDP, while maintaining or enhancing the anti-tumor (therapeutic) effects of CDP treatment.
Nutlin-3 was originally developed as an anti-cancer agent and was shown to be effective against OS xenografts.11,12 The small molecule inhibitor targets the MDM2-p53 interaction, and displaces p53 from the MDM2 binding pocket thereby releasing it from inhibition and proteasomal degradation.11 Nutlin-3 activity leads to the induction of downstream p53 targets, cell cycle arrest, and apoptosis; although p53-independent nutlin-3 effects have also been noted.12–15 Nutlin-3 and several second generation analogs have been shown to regress human tumors (including osteosarcoma) in preclinical studies.15–20
The idea that agents that target particular cell cycle checkpoints could protect normal cells from the effects of chemotherapy predated the discovery of nutlin-3; however, it has been demonstrated that indeed nutlin-3 could protect normal cells from various chemotherapeutic drugs by inducing reversible quiescence in vitro.1,21–29 Further, the addition of nutlin-3 to standard chemotherapeutic drugs (including CDP), in vitro, potentiated their tumor toxicities against human osteosarcoma cell lines.29,30 Based on these observations we hypothesized that nutlin-3 would protect bone healing from the toxic effects of CDP while maintaining or enhancing OS toxicity. These studies demonstrate that the peri-operative administration of nutlin-3 (bracketing CDP exposure) results in enhanced bone formation during DO with no effect on the CDP-induced killing of OS xenografts.
MATERIALS AND METHODS
Animals
Adult male C57BL/6 (B6, Jax #000664) and adult female athymic nude (Jax nu/nu, Jax #002019) mice were purchased from Jackson Labs (Bar Harbor, ME). Animals were housed in individual cages in temperature (22°C) and humidity (50%) controlled rooms having a 12 h light/12 h dark cycle. All mice were acclimated to the facility by animal care personnel for 5–7 days prior to surgery/treatment. In all studies, the mice were assigned to respective experimental groups with mean body weights equal to that of the control group (±3 g) for the study. All research protocols were approved by the Institutional Animal Care and Use Committee (IACUC) of the University of Arkansas for Medical Sciences. CDP was purchased from Sigma (#P4394) and nutlin-3 was purchased from Enzo Life Sciences (#ALX-430-128-M).
Study 1: The Effects of Pre-Operative CDP and Nutlin-3 Treatment on DO (Fig. 1A)
Figure 1.

CDP and Nutlin-3 Treatment Regimens. For the CDP studies the two dose regimen (red arrows) was used. For the nutlin-3 studies the three dose regimen (blue arrows) was used. For the combined studies, nutlin-3 will bracket CDP (red and blue arrows). The harvest times are shown by purple arrows. (A) Pre-operative drug administration. (B) Post-operative drug administration.
To provide proof-of-concept that an intervention involving induction of p53, by antagonizing the MDM2/p53 interaction, could protect bone healing from the detrimental effects of CDP the following pilot study was designed. Twenty (20) 9-week-old C57BL/6 male mice were weight matched and randomized to one of two groups: CDP + vehicle (n = 10) or CDP + nutlin-3 (n = 10). All mice received an IP injection (under isoflurane anesthesia) of either vehicle (50% DMSO) or nutlin-3 (20 mg/kg in 50% DMSO; made fresh daily) on days 1, 3, and 4 prior to surgery at day 7 (total of three injections The day after the first nutlin-3 or vehicle injection, all mice received an IP injection of CDP (2 mg/kg/day in saline; made fresh daily) for 2 days (days 2 and 3 prior to day 7 surgery). On the fourth day after the second injection of CDP, the mice underwent placement of an external fixator and osteotomy to the left tibia as described (Fig. 1A).10 Distraction began 3 days after surgery (3-day latency) at a rate of 0.075 mm b.i.d and continued for until day 21 (14 days post-op, 11 days of distraction). At sacrifice the distracted and contralateral (un-operated) tibiae were harvested.
Study 2: The Effects of Post-Operative CDP and Nutlin-3 Treatment on DO (Fig. 1B)
The second study utilized the delivery of CDP at 7 and 8 days post-surgery (study days 14 and 15) with, in some cases, the addition of nutlin-3 before and after the CDP injections. Sixty-eight (68) 12–13 week old C57BL/6 male mice were weight matched and randomized into one of eight replicate groups: 2 mg/kg/day CDP (n = 7), 4 mg/kg/day CDP (n = 10), 6 mg/kg/day CDP (n = 10), 20 mg/kg/day nutlin-3 (n = 8), 2 mg/kg/day CDP + 20 mg/kg/day nutlin-3 (n = 12), 2 mg/kg/day CDP + 50% DMSO (n = 7), 50% DMSO (n = 7), or Saline (n = 7) (all made fresh daily). The CDP vehicle was saline and the nutlin-3 vehicle was 50% DMSO. All mice underwent placement of an external fixator and osteotomy to the left tibia as described.10 Distraction began 3 days after surgery (i.e., a 3-day latency) at a rate of 0.075 mm b.i.d. and continued for 14 days.10 The mice in the nutlin-3 groups received an IP injection (under isoflurane anesthesia) of nutlin-3 (20 mg/kg in 50% DMSO; made fresh daily) on days 6, 8, and 9 post surgery (total of three injections; study days 13, 15, and 16) and their specific controls received an IP injection of 50% DMSO on the same days. The mice in the CDP groups received an IP injection of CDP (2, 4, or 6 mg/kg/day in saline; made fresh daily) for 2 days (days 7 and 8 post surgery, study days 14 and 15) and their specific controls (saline group) received an IP injection of saline on the same days (Fig. 1B). At sacrifice the distracted and contralateral tibiae were harvested.
Study 3: The Effects of CDP and Nutlin-3 Treatment on Human Osteosarcoma (OS) Xenografts in Nude Mice
We next tested whether the addition of nutlin-3 to CDP treatment would compromise CDP toxicity toward OS cells in a xenographic model using a human OS cell line (SJSA-1/ ATCC: CRL 2098).11,12 Twenty (20) 14-week-old athymic female nude mice (Jax, nu/j, 002019, 6 week old) were maintained in micro-isolator cages, weight matched and randomized into groups: CDP (n = 5), CDP + nutlin-3 (n = 5), or vehicle (n = 10). Human OS cells (3 × 106) were grown in RPMI-1640 medium plus 10% fetal bovine serum and resuspended in 0.2 ml Matrigel and PBS (1:1) and injected (26 gauge needle) subcutaneously into the left flank. One week later, mice from the CDP + nutlin-3 group received an IP injection (under isoflorane anesthesia) of nutlin-3 (20 mg/kg in 50% DMSO; made fresh daily) on days 8, 10, and 11 post cell injection (total of 3 injections) and their controls (vehicle group) received an IP injection of 50% DMSO. The day after the first nutlin-3 injection, mice from the CDP groups received an IP injection of CDP (6 mg/kg/day in saline first injection and 10 mg/kg for subsequent injections, made fresh daily) for 2 days (days 9 and 10 post cell injection). The same nutlin-3 and CDP injection time line was repeated one week later. Primary tumors were measured weekly (2 largest dimensions: x = larger diameter, y = smaller diameter) using Fisher Scientific digital calipers and tumor size was calculated using the formula ½ xy2. Mice were sacrificed 4 days after the last nutlin-3 injection. At the time of sacrifice the tumors were measured, dissected, and fixed formalin.
Distraction Osteognenesis Protocol
DO was performed as previously described.10,32–35 Under nembutal anesthesia, each mouse underwent placement of an external fixator and osteotomy to the left tibia. Four 27-gauge, 1.25 in needles were manually drilled through the tibia (two proximally, two distally). The titanium external fixator was then secured to the pins and a small skin incision was made distal to the tibia crest and the soft tissue carefully retracted to visualize the bone. A single hole was manually drilled through both cortices of the mid-diaphysis, and surgical scissors were used to fracture the cortex on either side of the hole. The fibula was fractured by direct lateral pressure. The periosteum and dermal tissues were closed with a single suture. Finally, buprenex (1.0 mg/kg) was given IM post-surgery for analgesia. Distraction began 3 days after surgery (3-day latency) at a rate of 0.075 mm b.i.d. (0.15 mm/day) and continued for 11 or 14 days at which time the tibias with fixators in place were harvested.
Radiographic and Histologic Analysis
After 48 h in 10% neutral buffered formalin, left tibiae were removed from the fixators for high-resolution single beam radiography, microCT evaluation, and subsequent histological processing. Radiographs were quantified as we have described.10 Following radiography and micro CT analysis the distracted tibiae were decalcified in 5% formic acid, dehydrated, and embedded in paraffin for histologic evaluation.36 Five to seven micron longitudinal sections were cut on a microtome (Leitz 1512, Wetzlar, Germany) for hematoxylin and eosin staining (H&E). Sections were selected to represent a central or near central gap location as we have described.10 To be included in both radiographic and histologic analyses the DO samples had to 1) be well aligned, 2) have no broken pin sites, 3) have few bone chips within the DO gap, 4) have an intact ankle, and 5) have had no significant weight loss or health problems during the distraction period as described previously.10
MicroCT Analysis of Distraction Osteogenesis
Following analytical radiography, formalin fixed tibia were analyzed using a MicroCT40 (Scanco Medical AG, Bassersdorf, Switzerland) and the manufacturer’s software as described.10 Briefly, distracted tibia including the entire distraction gap were scanned in cross section with an isotropic voxel size of 12 μm at 55 kV, 114 μA, and 1,000 projections per rotation with threshold and noise filter the same as that used for the trabecular analysis of intact mouse bones (sigma 0.8, support 1, threshold 245).37 The DO gaps and the extent of new bone formation were analyzed as described.37
Statistics
Significant differences between two individual group means of new bone formation after distraction were measured by the Student’s t-test. All data are reported as mean ± standard error of the mean (SEM). When assumptions of equal variances between groups failed, differences between group means were estimated using a generalized linear model assuming the underlying distribution of the outcome.38 Differences between group means of tumor volumes after transplant were estimated by Kruskal–Wallis One Way Analysis of Variance (ANOVA) on ranks. In Study 2 all pairwise comparisons were performed using ANOVA with the post-hoc Student–Newman–Keuls method. Differences were considered significant when p < 0.05 and are reported as such.
RESULTS
Study 1: The Effects of Pre-Operative CDP and Nutlin-3 Treatment on DO
This pilot study explored the hypothesis that a small molecule intervention, that antagonizes the MDM2/p53 interaction, could prevent the deleterious effects of CDP on bone healing. The pre-operative administration model of nutlin-3 and CDP was employed (Fig. 1A). Twenty (20) 9-week-old C57BL/6 male mice were randomized to receive either CDP + vehicle (n = 10) or CDP + nutlin-3 (n = 10). The evaluation by MicroCT identified robust rescue of the CDP-induced inhibition of bone formation in the regenerating distraction gap (Fig. 2A). Quantitation of individual specimens from both groups (Fig. 2B) demonstrated a significant increase in new bone formation in the distraction gap in Nutlin-treated animals (p < 0.001). The analysis of the distracted tibial radiographs demonstrated a significant increase in the mineralized area of distraction gaps of CDP/nutlin-3 treated (52.8% ± 3.7) versus CDP/vehicle treated mice (30.9% ± 6.5) (p < 0.010) (Fig. 2C and E). Histological analysis of the DO gaps confirmed the significant increase in bone formation in CDP/nutlin-3 treated (79.8% ± 2.1) versus CDP/vehicle treated mice (51.3% ± 4.6) (p < 0.001) (Fig. 2D and E).
Figure 2.

The effect of Nutlin on CDP treatment. (A) MicroCT renderings of the distraction gaps from cisplatin (CDP) + vehicle (VEH) treated mice and CDP + Nutlin treated mice. Increased new bone formation in the distraction gap of CDP + Nutlin-treated mice are observed. (B) Quantitation of microCT-measured new bone formation in the distraction gaps from CDP + Veh and CDP + Nutlin groups. *(p < 0.001). (C) Increased mineralized area of the distraction gaps is apparent in tibial radiographs from CDP + Veh and CDP + Nutlin groups. (D) Increased new bone formation is outlined in the histologic sections (H&E stained) distraction gaps from CDP + Veh and CDP + Nutlin groups. (E) Quantitation of both radiology and histology from C and D. The mineralized area of distraction gaps of CDP + Nutlin-treated (52.8% ± 3.7) versus CDP + VEH-treated mice (30.9% ± 6.5) (*p < 0.010) (Radiographs). A significant increase in bone formation in CDP + Nutlin-treated (79.8% ± 2.1) versus CDP + vehicle-treated mice (51.3% ± 4.6) (*p < 0.001) (Histology).
Study 2: The Effects of Post-Operative CDP and Nutlin-3 Treatment on DO
We next studied the effects of nutlin-3 and/or CDP using a treatment model that mimics the current clinical use of CDP (Cure Search/Children’s Oncology group) (Fig. 1B). Male mice were randomized into one of eight groups: 2 mg/kg/day CDP (n = 7), 4 mg/kg/day CDP (n = 10), 6 mg/kg/day CDP (n = 10), 20 mg/kg/day nutlin-3 (n = 8), 2 mg/kg/day CDP + 20 mg/kg/day nutlin-3 (n = 12), 2 mg/kg/day CDP +50% DMSO (n = 7), 50% DMSO (n = 7), or saline (n =7).
To test for any unexpected effects of the vehicles or the nutlin-3 alone, we compared new bone formation in these three groups. We found no significant effects on the mineralized area of the distraction gaps in either the saline vehicle (68.2% ± 6.0), the DMSO vehicle (66.4% ± 4.1), or the nutlin-3 (62.7% ± 5.7) groups. Histological analyses confirmed the lack of significant differences between the saline vehicle (84.0 ± 2.7), the DMSO vehicle (80.3% ± 3.5), or the nutlin-3 (71.6% ± 5.1) groups. These results suggest that the cell arresting effects of nutlin-3 were transient and the extra washout days were sufficient to protect de novo bone formation in the DO gap. This observation is consistent with the reported half-life of nutlin-3.21,23,31,39 In addition, these data suggest that neither vehicle had a significant effect on bone healing in vivo. Based on these observations and to increase the analytical power, for subsequent analyses the two vehicle groups were used as a combined vehicle control: radiography (67.4% ± 3.6) and histology (82.1% ± 2.2).
Next we compared the extent of bone formation in the DMSO vehicle and the 2 mg CDP groups. Analysis demonstrated no significant effects on the mineralized area of the distraction gaps between the 2 mg CDP (50% ± 5.8) and the 2 mg CDP/DMSO (53.4% ± 3.3) groups. Histological analyses confirmed the lack of any significant differences between the CDP (56.2% ± 8.9), and CDP/DMSO (65.4% ± 9.3) groups. For subsequent analyses, these groups were combined as the 2 mg CDP exposed groups: radiography (51.5% ± 3.4) and histology (59.7% ± 6.3).
Analysis of distracted tibial radiographs demonstrated a dose dependent inhibition of bone formation by CDP: combined CDP 2 mg/kg (51.5% ± 3.3), 4 mg/kg (36.8% ± 3.7), and 6 mg/kg (17.4% ± 4.5) versus combined vehicle controls (67.4% ± 3.6) or nutlin (65.1% ± 4.6) (p < 0.01–0.001) (Fig. 3). Histological analyses confirmed the dose dependent inhibition of bone formation by CDP: combined CDP 2 mg/kg (59.7% ± 6.3), 4 mg/kg (57.4% ± 7.7), and 6 mg/kg (28.3% ± 5.5) versus combined vehicle controls (82.1% ± 2.2) (p < 0.01–0.001).
Figure 3.
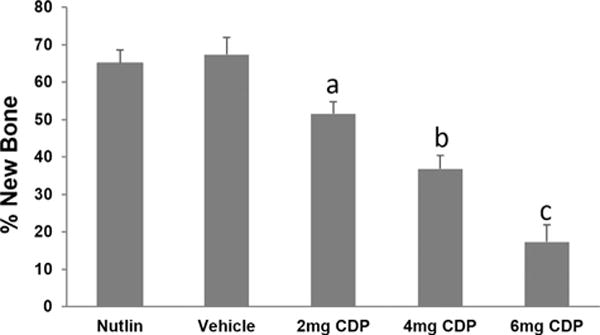
Dose-dependent inhibition of bone formation by CDP. CDP 2 mg/kg (51.5% ± 3.3), 4 mg/kg (36.8% ± 3.7), 6 mg/kg (17.4% ± 4.5) versus combined vehicle controls (67.4% ± 3.6) or nutlin (65.1% ± 4.6). Significant differences are indicated by letters all compared to vehicle. Vehicle versus 2 mg (a: p = 0.013); Vehicle versus 4 mg (b: p = 0.006); Vehicle versus 6 mg (c: p < 0.001).
To determine the magnitude of the negative effects of the low dose of CDP bone formation between the combined CDP groups versus the combined vehicle controls were compared. This comparison demonstrated an inhibition of bone formation by the combined low dose 2 mg CDP (51.5% ± 3.4) versus the combined vehicles (67.4% ± 3.6) (p = 0.011) Histological analyses confirmed the inhibition of bone formation by the combined low dose 2 mg CDP (59.7% ± 6.3) versus the combined vehicles (82.1% ± 2.2) (p = 0.013) (Fig. 4A).
Figure 4.
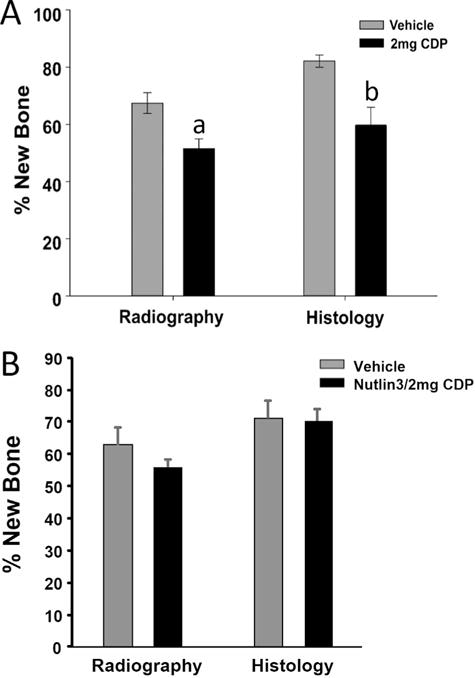
Inhibition of bone formation by CDP and protection by nutlin-3. (A) Comparison of distracted tibial radiographs demonstrated significant inhibition of bone formation by the combined low dose 2 mg CDP (51.5% ± 3.4) versus the combined vehicles (67.4% ± 3.6) (a: p = 0.011). Histological analyses confirmed the inhibition of bone formation by the combined low dose 2 mg CDP (59.7% ± 6.3) versus the combined vehicles (82.1% ± 2.2) (b: p = 0.013). (B) To directly test for the protective effects of nutlin-3 in the low dose 2 mg/kg CDP exposed mice, new bone formation in the DO gaps between the nutlin-3/vehicle versus the nutlin-3/2 mg/kg CDP groups were compared. Comparison of distracted tibial radiographs demonstrated no significant effects on the mineralized area of the DO gaps between the nutlin-3/vehicle (62.7% ± 5.7) versus nutlin-3/2 mg/kg CDP (56.6% ± 2.0). Histological analyses confirmed the lack of significant differences between the nutlin-3/vehicle (71.6% ± 5.1) versus nutlin-3 plus 2 mg/kg CDP (71.5% ± 3.6).
To directly test for the protective effects of nutlin-3 in the low dose 2 mg/kg CDP treated mice, new bone formation in the DO gaps between the nutlin-3/vehicle versus the nutlin-3/2 mg/kg CDP groups were compared (Fig. 4B). Comparison of distracted tibial radiographs demonstrated no significant effects on the mineralized area of the DO gaps between the nutlin-3/vehicle (62.7% ± 5.7) versus nutlin-3/2 mg/kg CDP (56.6% ± 2.0) (Fig. 4B). Histological analyses confirmed the lack of significant differences between the nutlin-3/vehicle (71.6% ± 5.1) versus nutlin-3 plus 2 mg/kg CDP (71.5% ± 3.6). Since the nutlin-3/vehicle groups represent normal robust bone formation during DO, these results are consistent with the protective actions of nutlin-3 on CDP exposed direct bone formation demonstrated in Study 1 (Fig. 4B).
Study 3: The Effects of CDP and Nutlin-3 Treatment on Human Osteosarcoma (OS) Xenografts in Nude Mice
This study was performed to determine the effects of nutlin-3 on CDP toxicity in a human osteosarcoma cell line (SJSA-1/ATCC: CRL 2098), a cell line isolated from an osteosarcoma11,12 that was the utilized in the initial report of nutlin-3 activity.13 Mice injected with SJSA-1 cells were assigned to three groups: vehicle, CDP, and CDP/nutlin-3. Primary tumor xenografts were dissected and measured two weeks post-treatment. Kruskal–Wallis One Way Analysis of Variance of ranked data demonstrated that CDP and CDP+ nutlin-3 resulted in, respectively, 67% (p = < 0.002) and 78% (p < 0.001) inhibition of tumor growth relative to vehicle controls (Fig. 5A and B). This result suggests enhanced OS toxicity due to the addition of nutlin-3. All pairwise multiple comparison procedures were performed using Dunn’s method.
Figure 5.

No effect of Nutlin on CDP-killing of osteosarcoma cells in vivo. (A) Mice were injected with human osteosarcoma cell line (SJSA-1/ATCC: CRL 2098): Vehicle, CDP, and CDP + Nutlin. Primary tumors were dissected and measured two weeks post-treatments. Representative tumors (2 from each group) are shown. (B) Kruskal–Wallis One Way Analysis of variance of ranked data demonstrated that CDP and CDP + nutlin-3 resulted in respectively 67% and 78% inhibition of tumor growth relative to vehicle controls (cm3) (*p = < 0.002). No significant differences between the two groups receiving CDP treatment was observed.
Collectively these results demonstrate that an intervention involving induction of p53, by a small molecule designed to antagonize the MDM2/p53 interaction, could (1) protect de novo bone healing from the effects of CDP, and (2) enhance the CDP toxicity toward specific osteosarcomas as demonstrated in vitro.29,30
DISCUSSION
The majority of childhood/adolescent OS patients are treated by a closely timed combination of chemotherapy and surgery, yet these patients continue to be at risk for postoperative complications especially with regard to bone repair. The effects of CDP, a drug commonly used for treatment of OS, on bone regeneration are not well studied. We have previously demonstrated that CDP, (60 mg/M2/day patient dose ~2 mg/kg mouse dose) given both with surgery and one week before surgery, dramatically inhibited direct bone repair (DO).10 Here we report that bone repair, subsequent to CDP exposure, can be protected by administration of a small molecule inducer of p53 (nutlin-3) when given either preoperatively or postoperatively.
Study 1 was designed as a pilot study such that: (1) the nutlin-3 was given bracketing the CDP injections, and (2) the nutlin-3 was given time to decay (washout) before the start of surgery. This last point may be important since the hypothetical actions of induction of p53 would delay normal osteoprogenitor entry into the proliferative phase thus potentially compromising the bone healing for approximately 24 h.21,23,31,39 In addition, the hypothesis that the administration of nutlin-3 when given in the Pre-Operative CDP model (Fig. 1A) would protect bone repair was supported. This study demonstrated: 1) the reproducibility of the previous effects of CDP in this model, 2) that nutlin-3 (given bracketing the CDP injections) is effective in protecting new bone formation, and (3) that the time given to allow the decay of nutlin-3 (“wash out” period ~24 h) before the start of surgery was effective.21,23,31,39
In the second study the treatment paradigm was changed to the delivery of CDP 7 days post-surgery, with in some cases, addition of nutlin-3 bracketing the CDP (Fig. 1B). This protocol may better mimic the current clinical SOP (Cure Search/Children’s Oncology group). The results demonstrated the following: (1) that neither vehicle had any significant effect on the outcomes, (2) that the cell arresting effects of nutlin-3 were transient and the extra days of the “washout period” were sufficient to protect the ongoing bone formation as noted in Study 1, (3) that there was a significant dose dependent inhibition of bone formation by CDP: 2, 4, and 6 mg/kg versus vehicle controls, and (4) that the new bone formation in the nutlin-3/CDP 2 mg/kg group was protected equivalent to that seen in the nutlin-3/vehicle group. First, these results demonstrate the significant inhibitory effects of CDP in a post-operative model. This finding is consistent with the previous data that demonstrated CDP’s toxic effects on repair both at surgery and pre-surgery.6,10,40,41 Second, these results support the proof of concept that nutlin-2 has protective actions on CDP exposed direct bone formation both in the pre- and post-operative models.
In the third study, an OS xenograph model was employed to determine the effects of nutlin-3 on the demonstrated toxicity of CDP towards human osteosarcoma cells. The results demonstrated that CDP and CDP + nutlin-3 both resulted in significant inhibition of tumor growth relative to vehicle control, and that there was no tumor protection by nutlin-3, a result consistent with previous studies using a variety of osteosarcoma cell lines.29,30
Although these studies were performed in vivo, some limitations persist. We note that single studies are reported here and the studies were performed in mice and so need to be interpreted in this preclinical context. Indeed, recent literature suggests that DO is an appropriate protocol for osteosarcoma patients that may provide significant benefit to patients compared with other critical size defect repair procedures.7,8,42,43 As such, the studies described herein may provide important new information regarding the utility of DO for clinical applications.8 Also, multi-agent chemotherapy is routinely performed and we only evaluated the effects of CDP dosing. Further, the selected CDP dose of 2 mg/kg/day for 2 days (total 4 mg/kg), though hypothetically equivalent to the pediatric dose of 60 mg/m2/day by body area, may underestimate the appropriate dose due to, among other factors, the higher metabolic rate of the mouse. In fact, recent studies have utilized total CDP doses of 7, 12, 20, and 63 mg/kg to model CDP toxicity in mice.44–47 In addition, though the single individual dose of nutlin-3 used here resulted in protection of bone repair, a dose response of nutlin-3 treatments against a dose response of CDP should be informative in terms of determining a possible “threshold” of protection and thus identifying an optimal dose of nutlin-3. The timing of the post-operative drug administration should also be viewed with caution. Although the 7-day period post-op administration of CDP is similar in this model to that in the clinic, the equivalency of 7 days in mice versus 7 days in humans is uncertain. Also, in humans, a 7-day latency is normal while in mice a 3-day latency is used. Therefore in the murine model drug dosing takes place after 4 days of distraction while in humans the CDP would be given just before distraction. The effects of these changes are unknown, but are currently under investigation.
The mechanisms underlying the protective effects of nutlin-3 versus CDP are at this time speculative. The primary candidate for nutlin-3 signal transduction is p53 due (1) to the fact that the nutlin molecules were designed to enhance p53 signaling and (2) that the majority of in vitro studies with both control and cancer cells demonstrated p53 dependence of the predicted nutlin-3 effects.11,12 Alternate mechanisms are possible, as suggested by the in vitro study of kidney cells where the cytoprotective effects of nutlin-3 were not related to the regulation of p53 or consequent gene expression.13,14 This work uncovered a new pharmacological function of nutlin-3, which was suppression of two mediators of apoptosis. Further, nutlin-3 has been shown to acetylate not only p53; but also, histones and heat shock proteins.48 Whether or not these alternative pathways can be demonstrated in vivo is unknown. Finally, at the cellular level, a significant portion of the protective actions of nutlin-3, which facilitate subsequent bone healing, may target the mesenchymal osteoprogenitors, or subsets, on which bone repair is critically dependent. This hypothesis is testable and currently under interrogation in our model system. In addition, other studies are ongoing to delineate the roles of the p53/p21 signaling axis in the protection of bone repair seen after nutlin-3 administration.
In general, the hypothesis that interventions using agents that impact cell cycle checkpoints (i.e., nutlin-3), could protect normal cells from the effects of chemotherapy while retaining onco-toxicity, is gaining acceptance.27 Nutlin-3 and two second generation analogs have recently been used in pre-clinical and clinical studies to regress human tumors (including osteosarcoma and liposarcoma).15–20 Furthermore several combinations of other chemotherapeutic drugs and nutlin-3 protectants have been proposed for future studies. These include TRAIL, bortezomib, VX-680, taxol, etoposide, and CDP.23,25,29,49 On a cautionary note, a recent study by Pishas et al.50 suggest a poorer response to nutlin in tumors with MDM2 amplification. However, it is important to note that of the 26 tumor-derived cell samples only one was an osteosarcoma. Regardless of these results, the use of nutlin is still indicated for sarcoma treatment by the authors and therefore the advantages of nutlin administration on bone repair remain an area of ongoing investigation.
In summary, this and previous studies have demonstrated that CDP has profound pre-, peri-, and post-operative negative effects on the process of bone repair in DO.6,10 Poor bone repair often leads to failure of the implanted endoprostheses, the engrafted bone, and/or of the lengthened bone.2–8 However, few therapies are currently available for the treatment and/or repair of chemotherapy-related bone repair deficits.4,5 Our DO model provides the platform to test a proof-of-concept that an intervention involving hypothesized induction of p53, by a small molecule designed to antagonize the MdM2/p53 interaction, could protect direct bone repair from the effects of CDP treatment while maintaining the anti-tumor potency of CDP. Finally, changes in cancer incidence and mortality have remained relatively constant over several decades; however, the number of cancer survivors has almost tripled.50 Therefore, with the increasing cohort of cancer survivors, efforts to attenuate the adverse effects of cancer therapy are receiving increased attention.51
Acknowledgments
The authors’ acknowledge the expert assistance of William Hogue and Phaedra Yount. This research was supported, in part, by (1) the Children’s University Medical Group Fund Grant Program, the Arkansas Children’s Hospital Research Institute, (2) the Arkansas Biosciences Institute, funded by the Arkansas Tobacco Settlement Plan and administered by Arkansas Children’s Hospital Research Institute, (3) UAMS Pediatric Hematology/ Oncology Division, (4) Carl L. Nelson Endowed Chair in Orthopaedic Creativity and UAMS Translational Research Institute supported by the National Institutes of Health National Center for Research Resources grant UL1 RR029884, (5) NIH National Center for Research Resources Grant #1CORR16517-01, by Arkansas Biosciences Institute and (6) National Institutes of Health R01 CA166060.
Grant sponsor: Children’s University Medical Group Fund Grant Program; Grant sponsor: Arkansas Biosciences Institute; Grant sponsor: UAMS Pediatric Hematology/Oncology Division; Grant sponsor: National Institutes of Health National Center for Research Resources; Grant number: UL1 RR029884; Grant sponsor: NIH National Center for Research Resources; Grant number: #1CORR16517-01; Grant sponsor: National Institutes of Health R01 CA166060.
Footnotes
AUTHORS’ CONTRIBUTIONS
KS: designed experiments, edited text and interpretation of data. EW: performed all experiments. LL: all animal surgery. RS: histological processing and interpretation. JS: all microCT analysis. RB: edited manuscript and provided interpretation. CM: edited text, provided clinical context. RN: edited text, provided clinical context. LS: designed experiments, edited and wrote text, interpreted results, managed efforts. JA: Provided clinical context, DO expert. DB: statistical expert. CS: statistical support, experimental design. CL: designed experiments, wrote manuscript. All authors have read and approved the final submitted manuscript.
References
- 1.Ciolli S. Effects on bone metabolism of new therapeutic strategies with standard chemotherapy and biologic drugs. Clin Cases Miner Bone Metab. 2013;10:183–186. Review. [PMC free article] [PubMed] [Google Scholar]
- 2.Yang C, Ji D, Weinstein EJ. The kinase Mirk is a potential therapeutic target in osteosarcoma. Carcinogenesis. 2010;31:552–558. doi: 10.1093/carcin/bgp330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Messerschmitt PJ, Garcia RM, Abdul-Karim FW, et al. Osteosarcoma. J Am Acad Orthop Surg. 2009;17:515–527. doi: 10.5435/00124635-200908000-00005. [DOI] [PubMed] [Google Scholar]
- 4.Bilariki K, Anagnostou E, Masse V. Low bone mineral density and high incidences of fractures and vitamin D deficiency in 52 pediatric cancer survivors. Horm Res Paediatric. 2010;74:319–327. doi: 10.1159/000313378. [DOI] [PubMed] [Google Scholar]
- 5.Bertoldo F, Pancheri S, Zenari S, et al. Emerging drugs for the management of cancer treatment induced bone loss. Expert Opin Emerg Drugs. 2010;15:323–342. doi: 10.1517/14728211003631385. Review. [DOI] [PubMed] [Google Scholar]
- 6.Ehrhart N, Eurell JA, Tommasini M. Effect of cisplatin on bone transport osteogenesis in dogs. Am J Vet Res. 2002;63:703–711. doi: 10.2460/ajvr.2002.63.703. [DOI] [PubMed] [Google Scholar]
- 7.McCoy TH, Jr, Kim HJ, Cross MB, et al. Bone tumor reconstruction with the Ilizarov method. J Surg Oncol. 2013;107:343–352. doi: 10.1002/jso.23217. [DOI] [PubMed] [Google Scholar]
- 8.Sabharwal S, Fragomen A, Iobst CJ. What’s new in limb lengthening and deformity correction. Bone Joint Surg Am. 2013;95:1527–1534. doi: 10.2106/JBJS.M.00599. [DOI] [PubMed] [Google Scholar]
- 9.Jarka DE, Nicholas RW, Aronson J. Effect of methotrexate on distraction osteogenesis. Clin Orthop Relat Res. 1998;20:9–15. doi: 10.1097/00003086-199809000-00025. [DOI] [PubMed] [Google Scholar]
- 10.Stine KC, Wahl EC, Liu L, et al. Cisplatin inhibits bone healing during distraction osteogenesis. J Orthop Res. 2014;32:464–470. doi: 10.1002/jor.22527. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Vassilev LT, Vu BT, Graves B, et al. In vivo activation of the p53 pathway by small-molecule antagonists of MDM2. Science. 2004;303:844–848. doi: 10.1126/science.1092472. [DOI] [PubMed] [Google Scholar]
- 12.Vu BT, Vassilev L. Small-molecule inhibitors of the p53-MDM2 interaction. Curr Top Microbiol Immunol. 2011;348:151–172. doi: 10.1007/82_2010_110. [DOI] [PubMed] [Google Scholar]
- 13.Valentine JM, Kumar S, Moumen A. A p53-independent role for the MDM2 antagonist Nutlin-3 in DNA damage response initiation. BMC Cancer. 2011;11:79. doi: 10.1186/1471-2407-11-79. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Saha MN, Jiang H, Chang H. Molecular mechanisms of nutlin-induced apoptosis in multiple myeloma: evidence for p53-transcription-dependent and -independent pathways. Cancer Biol Ther. 2010;10:567–578. doi: 10.4161/cbt.10.6.12535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Chen L, Zhao Y, Halliday GC, et al. Structurally diverse MDM2-p53 antagonists act as modulators of MDR-1 function in neuroblastoma. Br J Cancer. 2014;111:716–725. doi: 10.1038/bjc.2014.325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Tovar C, Graves B, Packman K, et al. MDM2 small-molecule antagonist RG7112 activates p53 signaling and regresses human tumors in preclinical cancer models. Cancer Res. 2013;73:2587–2597. doi: 10.1158/0008-5472.CAN-12-2807. [DOI] [PubMed] [Google Scholar]
- 17.Ray-Coquard I, Blay JY, Italiano A, et al. Effect of the MDM2 antagonist RG7112 on the P53 pathway in patients with MDM2-amplified, well-differentiated or dedifferentiated liposarcoma: an exploratory proof-of-mechanism study. Lancet Oncol. 2012;13:1133–1140. doi: 10.1016/S1470-2045(12)70474-6. [DOI] [PubMed] [Google Scholar]
- 18.Ding Q, Zhang Z, Liu JJ, et al. Discovery of RG7388, a potent and selective p53-MDM2 inhibitor in clinical development. J Med Chem. 2013;56:5979–5983. doi: 10.1021/jm400487c. [DOI] [PubMed] [Google Scholar]
- 19.Zhang Z, Chu XJ, Liu JJ, et al. Discovery of potent and orally active p53-MDM2 inhibitors RO5353 and RO2468 for potential clinical development. ACS Med Chem Lett. 2013;5:124–127. doi: 10.1021/ml400359z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhang Z, Ding Q, Liu JJ, et al. Discovery of potent and selective spiroindolinone MDM2 inhibitor, RO8994, for cancer therapy. Bioorg Med Chem. 2014;22:4001–4009. doi: 10.1016/j.bmc.2014.05.072. [DOI] [PubMed] [Google Scholar]
- 21.Carvajal D, Tovar C, Yang H, et al. Activation of p53 by MDM2 antagonists can protect proliferating cells from mitotic inhibitors. Cancer Res. 2005;65:1918–1924. doi: 10.1158/0008-5472.CAN-04-3576. [DOI] [PubMed] [Google Scholar]
- 22.Korotchkina LG, Demidenko ZN, Gudkov AV, et al. Cellular quiescence caused by the Mdm2 inhibitor nutlin-3A. Cell Cycle. 2009;8:3777–3781. doi: 10.4161/cc.8.22.10121. [DOI] [PubMed] [Google Scholar]
- 23.Secchiero P, Bosco R, Celeghini C, et al. Recent advances in the therapeutic perspectives of Nutlin-3. Curr Pharm Des. 2011;7:569–577. doi: 10.2174/138161211795222586. [DOI] [PubMed] [Google Scholar]
- 24.Fan C, Foster BK, Wallace WH, et al. Pathobiology and prevention of cancer chemotherapy-induced bone growth arrest, bone loss, and osteonecrosis. Curr Mol Med. 2011;1:140–151. doi: 10.2174/156652411794859223. Review. [DOI] [PubMed] [Google Scholar]
- 25.Cheok CF, Kua N, Kaldis P, et al. Combination of nutlin-3 and VX-680 selectively targets p53 mutant cells with reversible effects on cells expressing wild-type p53. Cell Death Differ. 2010;17:1486–1500. doi: 10.1038/cdd.2010.18. [DOI] [PubMed] [Google Scholar]
- 26.Apontes P, Leontieva OV, Demidenko ZN, et al. Exploring long-term protection of normal human fibroblasts and epithelial cells from chemotherapy in cell culture. Oncotarget. 2011;2:222–233. doi: 10.18632/oncotarget.248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Rao B, Lain S, Thompson AM. P53-Based cyclotherapy: exploiting the “guardian of the genome” to protect normal cells from cytotoxic therapy. Br J Cancer. 2013;109:2954–2958. doi: 10.1038/bjc.2013.702. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Jiang M, Pabla N, Murphy RF, et al. Nutlin-3 protects kidney cells during cisplatin therapy by suppressing Bax/ Bak activation. J Biol Chem. 2007;282:2636–2645. doi: 10.1074/jbc.M606928200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Evdokiou A, Bouralexis S, Atkins GJ, et al. Chemotherapeutic agents sensitize osteogenic sarcoma cells, but not normal human bone cells, to Apo2L/TRAIL-induced apoptosis. Int J Cancer. 2002;99:491–504. doi: 10.1002/ijc.10376. [DOI] [PubMed] [Google Scholar]
- 30.Ohnstad HO, Paulsen EB, Noordhuis P, et al. MDM2 antagonist Nutlin-3a potentiates antitumour activity of cytotoxic drugs in sarcoma cell lines. BMC Cancer. 2011;11:211–222. doi: 10.1186/1471-2407-11-211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Zhang F, Tagen M, Throm S, et al. Whole-body physiologically based pharmacokinetic model for nutlin-3a in mice after intravenous and oral administration. Drug Metab Dispos. 2011;39:15–21. doi: 10.1124/dmd.110.035915. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Aronson J, Liu L, Liu Z, et al. Decreased endosteal intramembranous bone formation accompanies aging in a mouse model of distraction osteogenesis. J Regen Med. 2002;3:7–16. [Google Scholar]
- 33.Wahl EC, Aronson J, Liu L, et al. Restoration of regenerative osteoblastogenesis in aged mice: modulation of TNF. J Bone Miner Res. 2010;25:114–123. doi: 10.1359/jbmr.090708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Fowlkes JL, Bunn RC, Liu L, et al. Runt-related transcription factor 2 (RUNX2) and RUNX2-related osteogenic genes are down-regulated throughout osteogenesis in type 1 diabetes mellitus. Endocrinology. 2008;149:1697–1704. doi: 10.1210/en.2007-1408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Thrailkill KM, Liu L, Wahl EC, et al. Bone formation is impaired in a model of type 1 diabetes. Diabetes. 2005;54:2875–2881. doi: 10.2337/diabetes.54.10.2875. [DOI] [PubMed] [Google Scholar]
- 36.Skinner RA, Hickmon SG, Lumpkin CK, Jr, et al. Decalcified bone: 20 years of successful specimen management. J Histotech. 1997;20:267–277. [Google Scholar]
- 37.Perrien DS, Nicks KM, Liu L, et al. Inhibin A enhances bone formation during distraction osteogenesis. J Orthop Res. 2012;30:288–295. doi: 10.1002/jor.21501. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Swearingen CJ, Tilley BC, Adams RJ, et al. Application of beta regression to analyze ischemic stroke volume in NINDS rt-PA clinical trials. Neuroepidemiology. 2011;37:73–82. doi: 10.1159/000330375. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Chang LJ, Eastman A. Differential regulation of p21 (waf1) protein half-life by DNA damage and Nutlin-3 in p53 wild-type tumors and its therapeutic implications. Cancer Biol Ther. 2012;13:1047–1057. doi: 10.4161/cbt.21047. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Barth E, Roenningen H, Solheim LF, et al. Effect of cis-platinum on bone ingrowth into porous fiber titanium. Mechanical and biochemical correlations. Int J Oral Maxillofac Implants. 1986;1(1):123–127. [PubMed] [Google Scholar]
- 41.Monsell FP, Barnes JR, Bellemore MC, et al. Cytotoxic agents are detrimental to bone formed by distraction osteogenesis. Strategies Trauma Limb Reconstr. 2013;8:173–180. doi: 10.1007/s11751-013-0179-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Betz M, Dumont CE, Fuchs B, et al. Physeal distraction for joint preservation in malignant metaphyseal bone tumors in children. Clin Orthop Relat Res. 2012;470:1749–1754. doi: 10.1007/s11999-011-2224-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Watanabe K, Tsuchiya H, Yamamoto N, et al. Over 10-year follow-up of functional outcome in patients with bone tumors reconstructed using distraction osteogenesis. J Orthop Sci. 2013;18:101–109. doi: 10.1007/s00776-012-0327-4. [DOI] [PubMed] [Google Scholar]
- 44.Sherif IO, Abdel-Aziz A, Sarhan OM. Cisplatin-induced testicular toxicity in rats: the protective effect of arjunolic acid. J Biochem Mol Toxicol. 2014;28:515–521. doi: 10.1002/jbt.21593. [DOI] [PubMed] [Google Scholar]
- 45.Ma B, Wang Y, Zhou X, et al. Synergistic suppression effect on tumor growth of hepatocellular carcinoma by combining oncolytic adenovirus carrying XAF1 with cisplatin. Cancer Res Clin Oncol. 2015;141:419–429. doi: 10.1007/s00432-014-1835-8. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Mondal J, Bishayee K, Panigrahi AK, et al. Low doses of ethanolic extract of Boldo (Peumus boldus) can ameliorate toxicity generated by cisplatin in normal liver cells of mice in vivo and in WRL-68 cells in vitro, but not in cancer cells in vivo or in vitro. J Integr Med. 2014;12:425–438. doi: 10.1016/S2095-4964(14)60045-5. [DOI] [PubMed] [Google Scholar]
- 47.Ren Z, Wang Y, Jiang W, et al. Anti-tumor effect of a novel soluble recombinant human endostatin: administered as a single agent or in combination with chemotherapy agents in mouse tumor models. PLoS ONE. 2014;9:e107823. doi: 10.1371/journal.pone.0107823. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Haaland I, Opsahl JA, Berven FS, et al. Molecular mechanisms of nutlin-3 involve acetylation of p53, histones and heat shock proteins in acute myeloid leukemia. Mol Cancer. 2014;13:116. doi: 10.1186/1476-4598-13-116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Carrillo AM, Hicks M, Khabele D, et al. Pharmacologically increasing Mdm2 inhibits DNA repair and cooperates with genotoxic agents to kill p 53 inactivated ovarian cancer cells. Mol Cancer Res. 2015 doi: 10.1158/1541-7786.MCR-15-0089. [Epub ahead of print] [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Pishas KI, Neuhaus SJ, Clayer MT, et al. Nutlin-3a efficacy in sarcoma predicted by transcriptomic and epigenetic profiling. Cancer Res. 2014;74:921–931. doi: 10.1158/0008-5472.CAN-13-2424. [DOI] [PubMed] [Google Scholar]
- 51.Hauer-Jensen M, Denham JW, Andreyev HJ. Radiation enteropathy—pathogenesis, treatment and prevention. Nat Rev Gastroenterol Hepatol. 2014;11:470–479. doi: 10.1038/nrgastro.2014.46. [DOI] [PMC free article] [PubMed] [Google Scholar]