

Abstract
Background
Rice plays an extremely important role in food safety because it feeds more than half of the world’s population. Rice grain yield depends on biomass and the harvest index. An important strategy to break through the rice grain yield ceiling is to increase the biological yield. Therefore, genes associated with organ size are important targets for rice breeding.
Results
We characterized a rice mutant gns4 (grain number and size on chromosome 4) with reduced organ size, fewer grains per panicle, and smaller grains compared with those of WT. Map-based cloning indicated that the GNS4 gene, encoding a cytochrome P450 protein, is a novel allele of DWARF11 (D11). A single nucleotide polymorphism (deletion) in the promoter region of GNS4 reduced its expression level in the mutant, leading to reduced grain number and smaller grains. Morphological and cellular analyses suggested that GNS4 positively regulates grain size by promoting cell elongation. Overexpression of GNS4 significantly increased organ size, 1000-grain weight, and panicle size, and subsequently enhanced grain yields in both the Nipponbare and Wuyunjing7 (a high-yielding cultivar) backgrounds. These results suggest that GNS4 is key target gene with possible applications in rice yield breeding.
Conclusion
GNS4 was identified as a positive regulator of grain number and grain size in rice. Increasing the expression level of this gene in a high-yielding rice variety enhanced grain yield. GNS4 can be targeted in breeding programs to increase yields.
Electronic supplementary material
The online version of this article (doi:10.1186/s12284-017-0171-4) contains supplementary material, which is available to authorized users.
Keywords: Rice, GNS4/D11, Grain number, Grain size, Cell elongation
Background
The world’s population is estimated to grow to around 8.5 billion by 2030 (http://esa.un.org/unpd/wpp/Publications/). To feed the growing population, it is estimated that agricultural production needs to increase by 60% (Yamaguchi and Hwang 2015). Rice (Oryza sativa L.) is a staple food of more than 3.5 billion people, mainly in Asia (Seck et al. 2012). A significant improvement in rice yield per unit ground area would significantly reduce the global food shortage.
Rice grain yield is defined as the product of yield sink capacity and filling efficiency (Kato and Takeda 1996). To achieve new breakthroughs in yield, breeding efforts have focused on expanding the yield sink capacity, mainly by increasing the number of grains per panicle and grain size. Strategies including high fertilizer inputs and optimized cultivation methods have been used to increase grain number and enhance grain filling to maximize rice production. New varieties, especially the so-called ‘super rice’ cultivars that produce large numbers of grains per panicle with a large yield potential have been bred and cultivated. There have also been breakthroughs in elucidating the molecular mechanisms underlying rice yield traits. Using molecular genetic approaches, researchers have identified several genes that control the size of rice panicle and grain. For example, mutants of LAX1, FZP, LOG, APO1, SP1, FON, DEP2, DEP3, and PAY1 genes were found to produce abnormal inflorescences and smaller panicles (Chu et al. 2006; Ikeda et al. 2007; Komatsu et al. 2003; Komatsu et al. 2001; Kurakawa et al. 2007; Li et al. 2010; Li et al. 2009; Qiao et al. 2011; Suzaki et al. 2004; Zhao et al. 2015). Several quantitative trait loci (QTL) controlling grain number have been identified. Among them, Gn1a, IPA1, PROG1, An-1, and An-2 negatively regulate grain number per panicle (Ashikari et al. 2005; Gu et al. 2015; Jiao et al. 2010; Jin et al. 2008; Luo et al. 2013), while SPIKE and qGP5–1 are related to increased grain number (Dong et al. 2013; Fujita et al. 2013).
Rice grain size is defined by grain length, width, length-width ratio, and grain weight, and is another important factor in determining rice yield. Generally, dwarf mutants of the genes involved in gibberellin (GA) and brassinosteroid (BR) biosynthesis and signaling, such as D1, D2, D11, D18, D61, BRD1, BRD2, and DSG1, produce smaller grains (Ashikari et al. 1999; Hong et al. 2003; Itoh et al. 2001; Mori et al. 2002; Tanabe et al. 2005; Yamamuro et al. 2000). Several QTL related to grain size have been isolated. For example, GS3, GL3.1, GW2, GW5 and GS5 control grain size (Fan et al. 2006; Li et al. 2011; Qi et al. 2012; Song et al. 2007; Weng et al. 2008), GW8 and GL7/GW7/SLG7 regulate grain shape (Wang et al. 2015a; Wang et al. 2012; Wang et al. 2015b; Zhou et al. 2015), and GIF1 and TGW6 control grain filling (Ishimaru et al. 2013; Wang et al. 2008).
In this study, we characterized a rice mutant, gns4 (grain number and size on chromosome 4), which showed reduced grain number per panicle and smaller grains compared with those of wild type (WT). The GNS4 gene, isolated via a map-based cloning approach, was found to be a novel allele of DWARF11 (D11), which encodes a cytochrome P450 protein. GNS4 regulates the expression levels of genes involved in BR synthesis and BR response. Overexpression of GNS4 in a high-yielding cultivar background significantly enhanced grain weight and increased grain yield, suggesting that GNS4 is key target gene with possible applications in yield breeding.
Results
Characters of gns4 mutant
To investigate the mechanism underlying panicle and grain development in rice, we conducted a genetic screen for mutants with altered panicle and grain size. The gns4 mutant was isolated from EMS-treated japonica variety Zhonghua 11C (ZH11C). At maturity, gns4 plants were shorter than WT plants (Fig. 1a, d), and produced smaller panicles and grains than those of WT (Fig. 1b, c). The average grain number per panicle of gns4 was 86.4% of that in ZH11C (Fig. 1e). As well as the reduced grain number, the main axes of gns4 were vestigial. The degree of spikelet clustering mainly depended on the length of the secondary branches. Some secondary branches of the gns4 mutant were significantly shortened, which caused spikelet clustering. The grain length, grain width, and grain thickness were significantly smaller in the gns4 mutant than in WT (Fig. 1f–h), resulting in reduced 1000-grain weight (9.4% lower than that of WT) (Fig. 1i). Together, these results indicated that GNS4 influences panicle and grain size in rice.
Fig. 1.
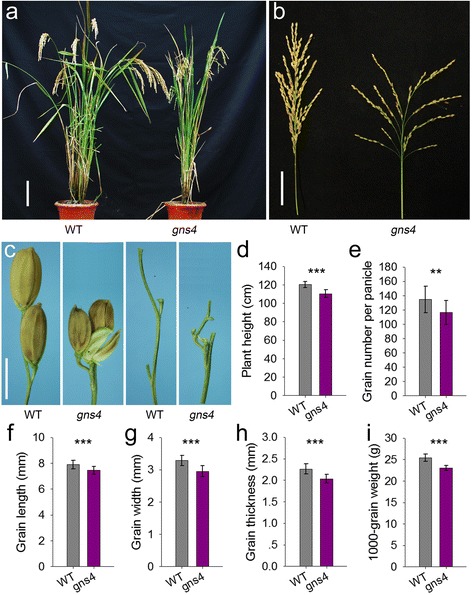
Morphological characteristics of the gns4 mutant. a Gross morphologies of WT and gns4 mutant at mature stage. Scale bar, 15 cm. b Panicle phenotypes of WT and gns4 mutant. Scale bar, 5 cm. c Grains and pedicels of WT and gns4 mutant. Scale bar, 5 mm. d-i Comparison of agronomic traits between WT and gns4 mutant. Data are given as means ± s.e. (n ≥ 20). Significant at ***0.1% and **1%
Map-based cloning of GNS4
To investigate the genetic basis of the mutation, we crossed gns4 with an indica variety 9311 to develop F1 and F2 populations. All the F1 plants showed a wild type phenotype. In the F2 population, plants with the WT and mutant phenotypes conformed to a 3:1 segregation ratio (X 2 = 0.98 < X 2 0.05 = 3.84), suggesting that this mutation was controlled by a single recessive gene.
We then isolated the GNS4 gene using map-based cloning. Firstly, the gns4 gene was limited between two molecular markers, LYH-71 and LYH-54, on chromosome 4 (Fig. 2a). The gns4 mutation was further fine-mapped to a 167-kb interval between the markers LYH-91 and LYH-52. Within this chromosome segment, there were 24 predicted open reading frames (ORFs). We failed to develop more polymorphic markers to further narrow down the candidate region, so we sequenced and analyzed all 24 ORFs. Only one SNP in the promoter region of LOC_Os04g39430 differed between WT and the gns4 mutant (Fig. 2b). The transcript level of LOC_Os04g39430 was significantly lower in gns4 leaves than in WT leaves (Fig. 2c), suggesting that LOC_Os04g39430 was a good candidate for GNS4.
Fig. 2.
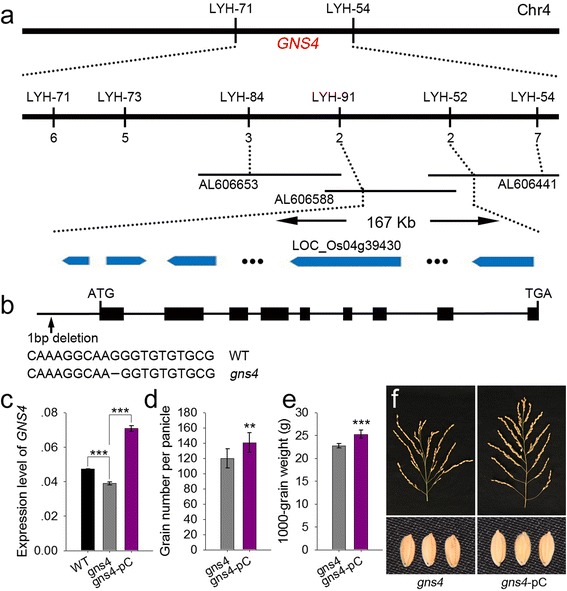
Map-based cloning of GNS4. a Fine mapping of GNS4. GNS4 was primarily mapped between markers LYH-54 and LYH-71 on chromosome 4. Then, GNS4 was fine mapped into a 167-kb segment between markers LYH-52 and LYH-91, using 856 recessive plants from the segregating population. The numbers underneath each marker indicate the numbers of recombinants between GNS4 and the molecular markers. The candidate genes within this region were showed in blue. b Gene structure and the sequence variance of GNS4 between WT and the gns4 mutant. c The expression levels of GNS4 among WT, gns4 and the complementary transgenic plants. The expression level of the rice Actin gene was amplified as a control. Values are means ± s.e. of three independent experiments. d Comparison of grain number per panicle between gns4 mutant and complementary transgenic plants. Data are given as means ± s.e. (n = 11). e Comparison of grain weight between gns4 mutant and complementary transgenic plants. Data are given as means ± s.e. (n = 11). f Panicle and grain phenotypes of gns4 and complementary transgenic plants. gns4-pC presents the transgenic gns4 plants with pGNS4::GNS4ZH11C construct. Significant at ***0.1% and **1%
To test this prediction, we generated a plasmid expressing the LOC_Os04g39430-coding region under the control of its native promoter. We introduced this construct into the gns4 mutant by Agrobacterium-mediated transformation. Twelve transgenic plants were generated, and all the positive lines showed complementation of the mutant phenotypes (Fig. 2d–f). This result confirmed that LOC_Os04g39430 was GNS4.
Next, a BLASTP analysis revealed that GNS4 was allelic to DWARF 11 (D11), which encodes a cytochrome P450 superfamily protein CYP724B1 (Tanabe et al. 2005). A previous study showed that D11 plays a role in BR synthesis and may be involved in the supply of typhasterol (TY) and 6-deoxoTY to the BR synthesis network in rice. In the gns4 mutant, a nucleotide deletion in the promoter region of GNS4 caused reduced grain number and smaller grains. We compared the transcript levels of GNS4 between WT and the gns4 mutant, and observed a slightly lower level of GNS4 transcripts in the mutant. Because the leaf phenotype of gns4 was not as stiff as those of the d11–1 and d11–2 mutants, we concluded that gns4 was likely to be a weak mutation of D11.
Manipulation of GNS4 has large effects on grain number and grain size
To further determine the roles of GNS4 in panicle and grain development, we created transgenic plants in Nipponbare (NIP) background by expressing a pUbi:RNAi–GNS4 construct driven by a constitutively expressed maize ubiquitin promoter. The positive lines (RNAi1 and RNAi2) with down-regulated GNS4 expression displayed a semi-dwarf stature (Fig. 3a, c). Similar to the gns4 mutant, the RNAi1 and RNAi2 plants had shortened plant height and panicle length (Table 1), and fewer grains per panicle (10.3% and 11.1% lower than that in WT, respectively) (Fig. 3b). Compared with Nipponbare plants, the transgenic lines had significantly decreased grain size (Table 1), resulting in approximately 7.8% and 11.1% decreases in 1000-grain weight, respectively (Fig. 3d, e). Down-regulation of GNS4 also led to reduced panicle number in RNAi1 and RNAi2 lines (Table 1).
Fig. 3.
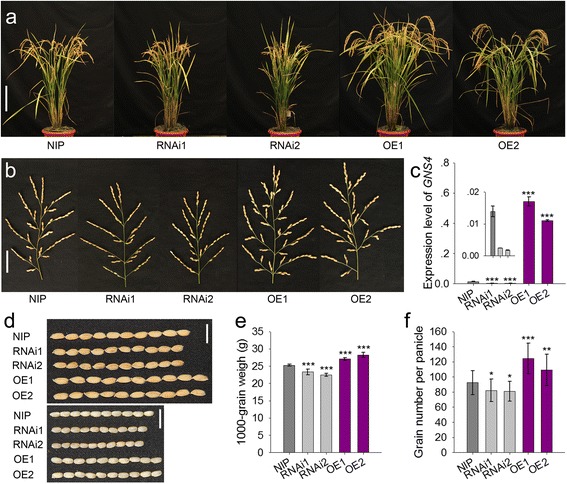
RNAi and overexpression analysis of GNS4 gene in Nipponbare background. a Phenotypic features of GNS4 RNAi and overexpression transgenic plants. Bar, 20 cm. b Panicle architecture of GNS4 RNAi and overexpression transgenic plants. Bar, 5 cm. c GNS4 mRNA expression level of the RNAi and overexpression transgenic plants. The expression level of the rice Actin gene was amplified as a control. Values are means ± s.e. of four independent experiments. d Grains of the transgenic plants. Bar, 1 cm. e-f Comparison of 1000-grain weight and grain number per plant between Nipponbare and transgenic lines. Data are given as means ± s.e. (n ≥ 13). Significant at ***0.1%, **1% and *5%. NIP, Nipponbare. RNAi1 and RNAi2 are two independent lines for RNAi analysis, and OE1 and OE2 are two independent lines for overexpression analysis
Table 1.
Comparison of agronomic traits between NIP and the transgenic lines
| NIP | RNAi1 | RNAi2 | OE1 | OE2 | |
|---|---|---|---|---|---|
| Grain length (mm) | 7.51 ± 0.12 | 6.60 ± 0.35a | 6.52 ± 0.13a | 8.01 ± 0.19a | 7.96 ± 0.17a |
| Grain width (mm) | 3.39 ± 0.18 | 3.15 ± 0.15b | 3.17 ± 0.22c | 3.27 ± 0.08NS | 3.38 ± 0.10NS |
| Grain thickness (mm) | 2.41 ± 0.07 | 2.36 ± 0.06NS | 2.28 ± 0.06a | 2.33 ± 0.07c | 2.39 ± 0.12NS |
| Plant height (cm) | 82.74 ± 4.61 | 72.69 ± 5.39a | 72.47 ± 5.99a | 86.50 ± 3.33a | 91.97 ± 4.01a |
| Panicle length (cm) | 21.47 ± 1.38 | 19.75 ± 1.13a | 20.41 ± 1.82c | 23.68 ± 1.87a | 23.98 ± 1.62a |
| Panicle number per plant | 17.00 ± 1.95 | 14.90 ± 2.08c | 12.00 ± 2.98a | 18.09 ± 3.73NS | 15.73 ± 3.32NS |
| Grain yield per plant (g) | 23.63 ± 3.43 | 10.37 ± 3.82a | 10.56 ± 3.93a | 27.60 ± 4.85c | 27.35 ± 3.86c |
| Biomass yield per plant (g) | 51.56 ± 9.57 | 33.34 ± 8.02a | 35.13 ± 5.30a | 64.08 ± 11.03b | 61.98 ± 10.04c |
Data are given as means ± s.e. (n ≥ 8). Significant at a0.1%, b1% and c5%. NS, not significant. NIP, Nipponbare. RNAi1 and RNAi2 are two independent lines for RNAi analysis, and OE1 and OE2 are two independent lines for overexpression analysis
We also produced overexpression lines of GNS4 in the Nipponbare background. Two independent positive lines with higher GNS4 transcript levels (Fig. 3c), OE1 and OE2, showed significantly higher plants, longer panicles, bigger and more grains, compared with those of WT (Fig. 3b, f; Table 1). However, no obvious increase in panicle number was observed in OE1 and OE2 lines (Table 1). At mature stage, we carried out a yield test and found that the elevated expression level of GNS4 enhanced both biomass yield and grain yield (Table 1).
Together, these results provided persuasive evidences that GNS4 has large effects on multiple traits, and improves yield production by producing more and bigger grains in rice.
Mutation of GNS4 decreased cell size in rice grains
The BRs are plant steroid hormones that regulate many aspects of plant development, such as organ size and cell elongation (Yang et al. 2011). The gns4 mutant produced smaller grains than those of WT. The size of the spikelet hull sets an upper limit for sink size and finally determines the grain size (Sakamoto and Matsuoka 2008). To investigate whether cell number or cell size contributes to the difference in grain size, we observed the outer glume epidermal cells under a scanning electron microscope. The outer epidermal cells were smaller in gns4 than in WT, for example, there was a ~ 6.6% decrease in longitudinal cell length on gns4 spikelet hulls (Fig. 4). As mentioned above, the grain length was 5.5% shorter in gns4 than in WT. The SEM analysis of the outer glume epidermal cells of RNAi and overexpression plants showed that the cell length varied with the transcript levels of GNS4 (Fig. 4). These results indicated that GNS4 regulates spikelet hull size and grain weights largely by controlling cell elongation.
Fig. 4.
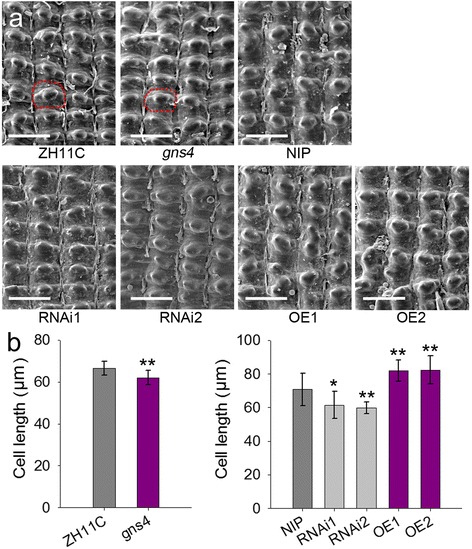
The effect of GNS4 on cell size of glume. a Scanning electron microscope photographs of the glume outer surfaces of mature seeds. The outline of one glume cell in WT and gns4 hulls, respectively, is boxed by the red dotted line. Bar, 100 μm. b Comparison of average length of outer glume surface cells. Data are shown as means ± s.e. (n ≥ 10). Significant at **1% and *5%
Previous studies reported that BR-deficient or BR-insensitive rice mutants exhibit erect leaves (Zhang ea. al., 2014). We examined the lamina joint bending angles and found that down-regulation of GNS4 led to erect leaves, while GNS4 overexpression caused an enlarged leaf angle (Fig. 5). We also compared the lamina joint angle of the flag (I), second (II), third (III), and fourth (IV) leaves (counted from the flag leaf downwards) at the heading stage between Nipponbare and transgenic plants. The angles of the bottom leaves were larger than those of the flag leaves (Fig. 5), implying that the effect of GNS4 on leaf angle increases with leaf age or stage of development.
Fig. 5.
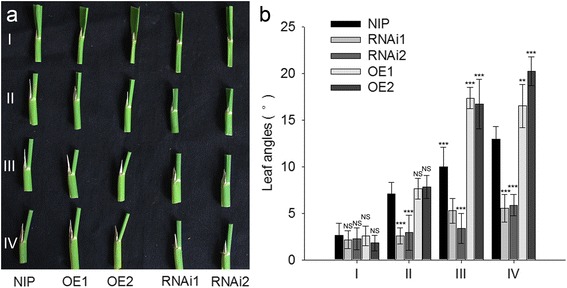
The effect of GNS4 on leaf angle. a Comparison of the lamina joint angle of the flag (I), second (II), third (III), and fourth (IV) leaves (counted from the flag leaf downwards) between Nipponbare and transgenic plants. b Quantification of the leaf lamina joint angles. Data are shown as means ± s.e. (n = 7). Significant at ***0.1%, and **1%. NS, not significant
Transcript levels of GNS4
The results of the qRT-PCR analysis indicated that in ZH11C plants, the highest transcript levels of GNS4 were in the young panicle, with much lower transcript levels in the stem, leaf, and leaf sheath (Fig. 6a). We also examined the transcript levels of four BR biosynthesis genes and nine BR-signaling genes in WT and the gns4 mutant. Interestingly, no feedback regulation of these BR-related genes was detected. The transcript levels of all 13 detected genes were significantly lower in the gns4 mutant than in WT (Fig. 6b, c).
Fig. 6.
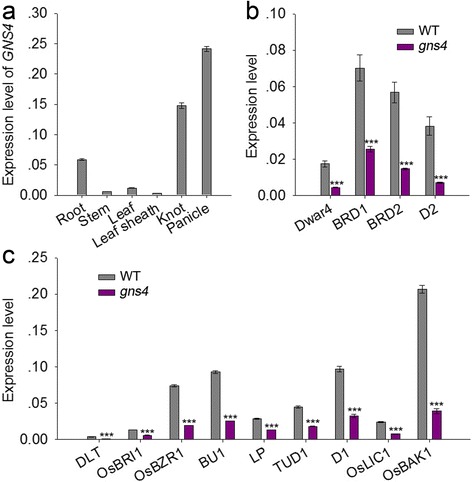
Expression analysis of GNS4 by qRT-PCR. a Relative expression levels of GNS4 in different tissues. b Comparison of transcription level of genes involved in rice BR biosynthesis between WT and gns4 mutant. c Relative transcription levels of the genes related to BR signaling in WT and gns4 mutant. The expression level of the rice Actin gene was amplified as a control. Values are means ± s.e. of four independent experiments
Elevated GNS4 expression in a high-yielding variety background increases yields
As mentioned above, overexpression of GNS4 in the Nipponbare background led to an obviously enlarged grain size. To further evaluate the potential use of the GNS4 gene, we generated transgenic lines using Wuyunjing 7 (WYJ7) as the recipient parent. WYJ7 is high-yielding variety that is widely cultivated in Jiangsu Province, China. More than 10 independent transgenic lines were obtained, and two homozygous T3 lines with elevated GNS4 transcript levels were selected for further analysis (Fig. 8a). At maturity, the GNS4-overexpressing lines exhibited dramatically improved growth, compared with WYJ7. The plant height of WYJ7-OE1 and WYJ7-OE2 was 6.1% and 10.5% greater, respectively, than that of WYJ7 (Figs. 7a and 8b). The most obvious improvement by GNS4 overexpression was the enlarged sink size, as reflected by the increased grain number per panicle and larger grains. The grain number per plant in WYJ7-OE1 and WYJ7-OE2 was increased by 10.4% and 6.8%, respectively, compared with that of WYJ7 (Figs. 7b and 8c). The 1000-grain weight of WYJ7-OE1 and WYJ7-OE2 was increased by 5.9% and 7.9%, respectively, compared with that of WYJ7 (Figs. 7c and 8e). And the enlarged grain size mainly resulted from grain length, rather than grain width and thickness (Fig. 8f-h). Compared with WYJ7, the overexpression lines had a little more panicle number per plant (significant in WYJ7-OE1 but not significant in WYJ7-OE2) (Fig. 8d). The biomass yield per plant was remarkably increased in the WYJ7-OE1 and WYJ7-OE2 (Fig. 8i). Finally, the grain yield per plant was improved by 16.5% and 14.6%, respectively, compared with that of WYJ7 (Fig. 8j).
Fig. 8.
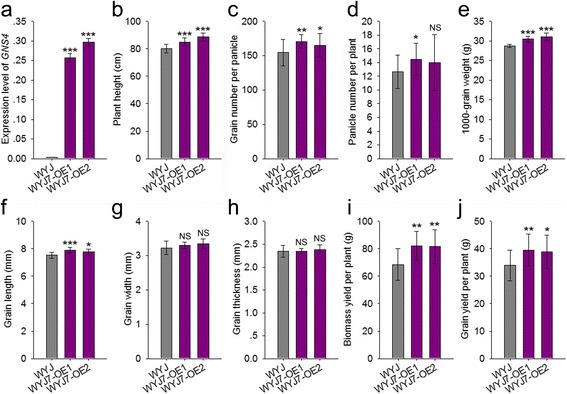
Agronomic traits of GNS4-overexpressing lines in high-yield variety Wuyunjing 7 (WYJ7) background. a GNS4 mRNA expression level of WYJ7, WYJ-OE1 and WYJ-OE2 plants. The expression level of the rice Actin gene was amplified as a control. Values are means ± s.e. of three independent experiments. b-j: Agronomic traits of WT and transgenic lines. b Plant height. c Grain number per panicle. d Panicle number per plant. e 1000-grain weight. f Grain length. g Grain width. h Grain thickness. i Biomass yield per plant. j Grain yield per plant. Traits were measured at the mature stage. Data are given as means ± s.e. (n ≥ 10). Significant at ***0.1%, **1% and *5%. NS, not significant. WYJ-OE1 and WYJ-OE2 are two independent GNS4-overexpressing lines
Fig. 7.
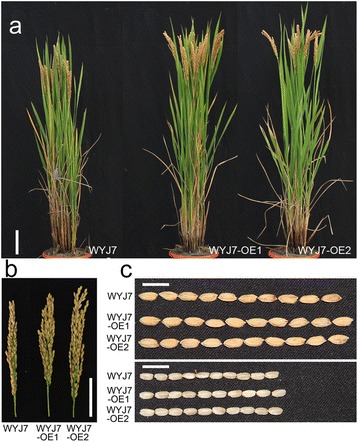
Plant morphology, panicle architecture and grain size of GNS4-overexpressing lines in high-yield variety Wuyunjing 7 (WYJ7) background. a Mature plant appearance of WYJ7 and the transgenic lines. Bar, 10 cm. b Panicle architecture of WYJ7 and the transgenic lines. Bar, 5 cm. c Comparisons of grains (top) and brown rice (bottom) between WYJ7 and the transgenic lines. Bars, 1 cm. WYJ-OE1 and WYJ-OE2 are two independent GNS4-overexpressing lines
Discussion
Rice is a very important food crop because it feeds more than half of the world’s population. Grain yield improvement is the main aim for rice breeders. Rice yield potential is determined by biomass and the harvest index. The harvest index of the cultivated rice varieties in China has almost reached its theoretical limit. Improving biomass production is an effective strategy to break through the yield ceiling. Rice biomass depends on plant organ size, which is controlled by genetic factors and environmental conditions. Although numerous genetic determinants of organ growth have been characterized, our understanding of how organ size is regulated is incomplete.
To reveal more genes related to organ size in rice, we characterized a rice mutant, gns4, with reduced panicle and grain size. Map-based cloning demonstrated that reduced transcription of GNS4 caused by an SNP in its promoter region resulted in the mutant phenotype. Down-regulation of GNS4 by RNAi also resulted in smaller panicles and grains. This finding suggested that GNS4 plays a positive role in regulating panicle and grain size in rice.
GNS4 encodes a cytochrome P450 superfamily protein CYP724B1, and is allelic to the previously reported D11, which plays roles in BR synthesis (Tanabe et al. 2005). In plants, BRs are essential steroid hormones that regulate diverse processes during plant development, such as stem elongation, vascular differentiation, male fertility, senescence, and responses to various biotic and abiotic stresses (Yang et al. 2011). As described in a previous study, both the d11–1 and d11–2 mutants exhibited shortened internodes and produced extremely small round grains. However, the gns4 mutant did not show this severe phenotype, with only a 5.5% and 9.4% reduction in grain length and grain weight, respectively, compared with those of WT. Molecular detection indicated that an SNP variation in the promoter region slightly decreased the expression level of GNS4, leading to the mutant phenotype. These data suggested that gns4 is a weak mutant of the GNS4 gene. In rice, a BR-deficient mutant displayed erect leaves, reduced plant height, and decreased tiller number and grain size (Yang et al. 2011). In addition to these common traits, the gns4 mutant has some distinct traits. For instance, gns4 has a very short main axis with most of the primary branches clustered at the base of the main axis, and few secondary branches. Together, these results suggested a novel and important role for GNS4/D11 in regulating inflorescence development.
Because GNS4 is involved in regulating panicle and grain size, we were interested in whether this gene could be used to improve rice yields. To investigate the function of GNS4, we generated several GNS4-overexpressing lines in Nipponbare background. Strong expression of GNS4 in the transgenic lines was detected by qRT-PCR (Fig. 3c). At the heading and mature stages, rice plants harboring the overexpression construct outgrew Nipponbare plants (Fig. 3), indicating that GNS4 controls vegetative growth. When grown in paddies, the biomass of GNS4-overexpressing lines was greater than that of Nipponbare (Fig. 3; Table 1). Multiple sink-related traits, including grain size and grain number per panicle, were enhanced in the GNS4-overexpressing lines. As anticipated, the grain yield per plant was increased by 16.8% and 15.7%, compared with that of Nipponbare. Next, we created GNS4-overexpressing lines using WYJ7, a high-yielding variety, as the recipient. These GNS4-overexpressing lines also showed significant biomass and grain yield improvements (Fig. 8i, j). Taken together, these results indicated that GNS4 has multiple beneficial effects on grain yield components, and is valuable for high-yield rice breeding.
Recently, Wu et al. (2016) found that the overexpression line (OE-CPB1) of CPB1/D11 under the control of the maize ubiquitin promoter showed increased grain length and 1000-grain weight. However, there was no significant increase in the grain yield per plant of OE-CPB1 plants as compared with WT, because the transgenic plants showed profound changes in plant architecture, such as larger leaf angles and narrower leaves. Transgenic plants expressing CPB1/D11 under the control of panicle-specific promoters from DEP1 and TH1 produced larger seeds and increased grain yields without changes in other agronomic traits, such as grain number per panicle. In this study, the D11-overexpressing lines in both the Nipponbare and WYJ7 backgrounds produced larger seeds, more grains, and increased yields, compared with those of WT. We noticed that the CPB1/D11 transcripts in OE-CPB1 lines accumulated more than 450-fold than in WT. In this study, however, there is only 30.0- to 93.6-fold increase in GNS4/D11 expression in the overexpression transgenic lines (Figs. 3c and 8a). As a result, the phenotypic variations in our data are not as big as Wu’ results. We speculate that the yield-increasing effect of D11 may be determined by its expression pattern, and may also be affected to some extent by genetic backgrounds.
Conclusions
The rice gns4 mutant showed reduced organ size, fewer grains per panicle, and smaller grain. Map-based cloning investigated that the GNS4 gene, encoding a cytochrome P450 protein, is allelic to DWARF11 (D11). Morphological and cellular analyses suggested that GNS4 positively regulates grain size by promoting cell elongation. Elevated expression level of GNS4 significantly increased 1000-grain weight and grain number per panicle, and subsequently enhanced grain yields in both the Nipponbare and Wuyunjing7 (a high-yielding cultivar) backgrounds. These results suggest that GNS4 is key target site to increase yields in rice breeding programs.
Methods
Plant materials
The spontaneous mutant, gns4, was identified from japonica rice Zhonghua 11C (ZH11C). The mutant was self-pollinated for several generations until the mutation was genetically proven to be truly inherited.
Genetic analysis and fine mapping of GNS4
For genetic analyses, an F2 population derived from a cross between gns4 and 9311, an indica variety, was grown in paddy fields under natural conditions. This segregating population was used for fine mapping of the GNS4 locus. Recessive individuals in the F2 segregating population were used to screen recombinants. To fine-map GNS4, several polymorphic InDel molecular markers were developed based on sequence differences between the indica variety 9311 and the japonica variety Nipponbare (Additional file 1: Table S1), according to data published at the NCBI (http://www.ncbi.nlm.nih.gov). Candidate genes were amplified and sequenced using gene-specific primers.
RNA extraction, complementary DNA synthesis, and qRT-PCR
Total RNA was isolated from various organs using an RNA extraction kit following the manufacturer’s instructions (Beijing Tiangen Biotechnology Co. Ltd., Beijing, China; www.tiangen.com/). First-strand cDNA was reverse-transcribed from approximately 1 μg gDNase-treated RNA using a FastQuant RT Kit following the manufacturer’s instructions (Beijing Tiangen Biotechnology Co. Ltd.). Gene transcript levels were determined by quantitative reverse transcription-PCR (qRT-PCR) with the rice Actin gene as the control. Each qRT-PCR was performed in a total volume of 25 μL containing 2 μL cDNA, 0.2 mM each primer, and 12.5 μL 2 × SYBR green PCR master mix (Takara, Dalian, China; http://www.takara.com.cn). The qRT-PCR was carried out using an ABI ViiA7 real-time PCR system using the following program: 95 °C for 3 min, then 40 cycles of 94 °C for 30 s, 55 °C for 30 s, and 72 °C for 40 s. Relative gene transcript levels were calculated using the 2-ΔΔCT method.
Vector construction and plant transformation
For the complementation test, the promoter (a DNA fragment ~2 kb upstream of translation start site) of GNS4 was amplified from ZH11C genomic DNA. The full coding region of GNS4 was also cloned. Both segments were then cloned into the binary pCAMBIA1301 vector to generate a construct in which GNS4 was driven by its native promoter. The full coding region of GNS4 was amplified from ZH11C cDNA and then inserted into the p1301UbiNOS vector to generate an overexpression construct in which GNS4 gene was controlled by a constitutively expressed maize ubiquitin promoter (Zhou et al. 2009). This overexpression construct was transformed into japonica varieties Nipponbare and Wuyunjing 7. For RNAi analysis, a DNA fragment of LOC_Os04g39430 was amplified and then cloned into the pMD18-T vector (Takara), before being cloned into the BamH I/Spe I and Bgl II/Xba I sites of the p1022 vector. Then, the stem-loop fragment was cloned into the p1301UbiNOS vector (Zhou et al. 2009). All the constructs were transformed into the recipient lines by Agrobacterium tumefaciens (strain EHA105) mediated transformation.
Evaluation of agronomic traits
Forty plants of each line were grown in the experimental field of Yangzhou University (E119°25′/N32°23′), from May through October in 2016. The distance between the plants in a row was 17.0 cm, and the distance between rows was 23.3 cm. Nitrogen (225 kg ha−1 as urea), together with phosphorus (50 kg ha−1 as single superphosphate) and potassium (60 kg ha−1 as KCl), were applied after transplanting. Field management and disease and pest control followed the standard procedures to prevent yield loss during the growth period.
All traits were evaluated at the mature stage. Plant height was measured from the ground surface to the tip of the tallest panicle. Panicle number per plant was the number of effective panicles with 10 or more grains. We also counted the grain number per panicle, and measured 1000-grain weight, grain yield per plant, and biomass per plant. Paddy grains were dried naturally after harvesting and stored at 37 °C for at least 1 week before testing. Fully filled grains were used for measurements. The independent sample t-test program of SPSS 10.0 for Windows was used to compare mean values among the mutants, overexpression and RNAi lines, and WT.
Morphological and cellular analyses
The glume outer surfaces of mature seeds were directly observed under a scanning electron microscope (SEM). The length of cells on the spikelet hull was measured using Image J software.
Acknowledgements
This work was supported by grants from the Key Program of Basic Research of China (2013CBA01405), the National Natural Science Foundation of China (31100863, and 31400175), the Natural Science Fund for Colleges and Universities in Jiangsu Province (15KJB210005 and 13KJA210003), the Natural Science Foundation of Jiangsu Province (BK20150010), the Prospective Agricultural Project of Yangzhou City (YZ2014165), and the Priority Academic Program Development of Jiangsu Higher Education Institutions.
Additional file
Primers used in this study. (PDF 37 kb)
Authors’ contributions
GL and ZG conceived and designed the experiments. YZ and YT conducted the experiments. JZ and ZY screened the mutant and helped to develop the mapping population. YZ and YT analyzed the data. JM, JL, YL, and CY conducted field tests. YZ and GL wrote the manuscript. All authors reviewed and approved the manuscript.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Footnotes
Yong Zhou, Yajun Tao and Jinyan Zhu contributed equally to this work.
Electronic supplementary material
The online version of this article (doi:10.1186/s12284-017-0171-4) contains supplementary material, which is available to authorized users.
Contributor Information
Zefeng Yang, Email: zfyang@yzu.edu.cn.
Zhiyun Gong, Email: zygong@yzu.edu.cn.
Guohua Liang, Email: ricegb@yzu.edu.cn.
References
- Ashikari M, Sakakibara H, Lin S, Yamamoto T, Takashi T, Nishimura A, Angeles ER, Qian Q, Kitano H, Matsuoka M. Cytokinin oxidase regulates rice grain production. Science. 2005;309:741–745. doi: 10.1126/science.1113373. [DOI] [PubMed] [Google Scholar]
- Ashikari M, Wu J, Yano M, Sasaki T, Yoshimura A. Rice gibberellin-insensitive dwarf mutant gene Dwarf 1 encodes the alpha-subunit of GTP-binding protein. Proc Natl Acad Sci U S A. 1999;96:10284–10289. doi: 10.1073/pnas.96.18.10284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chu H, Qian Q, Liang W, Yin C, Tan H, Yao X, Yuan Z, Yang J, Huang H, Luo D, Ma H, Zhang D. The floral organ number 4 gene encoding a putative ortholog of Arabidopsis CLAVATA3 regulates apical meristem size in rice. Plant Physiol. 2006;142:1039–1052. doi: 10.1104/pp.106.086736. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dong X, Wang X, Zhang L, Yang Z, Xin X, Wu S, Sun C, Liu J, Yang J, Luo X. Identification and characterization of OsEBS, a gene involved in enhanced plant biomass and spikelet number in rice. Plant Biotech J. 2013;11:1044–1057. doi: 10.1111/pbi.12097. [DOI] [PubMed] [Google Scholar]
- Fan C, Xing Y, Mao H, Lu T, Han B, Xu C, Li X, Zhang Q. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor Appl Genet. 2006;112:1164–1171. doi: 10.1007/s00122-006-0218-1. [DOI] [PubMed] [Google Scholar]
- Fujita D, Trijatmiko KR, Tagle AG, Sapasap MV, Koide Y, Sasaki K, Tsakirpaloglou N, Gannaban RB, Nishimura T, Yanagihara S, Fukuta Y, Koshiba T, Slamet-Loedin IH, Ishimaru T, Kobayashi N. NAL1 allele from a rice landrace greatly increases yield in modern indica cultivars. Proc Natl Acad Sci U S A. 2013;110:20431–20436. doi: 10.1073/pnas.1310790110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gu B, Zhou T, Luo J, Liu H, Wang Y, Shangguan Y, Zhu J, Li Y, Sang T, Wang Z, Han B. An-2 encodes a cytokinin synthesis enzyme that regulates awn length and grain production in rice. Mol Plant. 2015;8:1635–1650. doi: 10.1016/j.molp.2015.08.001. [DOI] [PubMed] [Google Scholar]
- Hong Z, Ueguchi-Tanaka M, Umemura K, Uozu S, Fujioka S, Takatsuto S, Yoshida S, Ashikari M, Kitano H, Matsuoka M. A rice brassinosteroid-deficient mutant, ebisu dwarf (d2), is caused by a loss of function of a new member of cytochrome P 450. Plant Cell. 2003;15:2900–2910. doi: 10.1105/tpc.014712. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ikeda K, Ito M, Nagasawa N, Kyozuka J, Nagato Y. Rice ABERRANT PANICLE. ORGANIZATION 1, encoding an F-box protein, regulates meristem fate. Plant J. 2007;51:1030–1040. doi: 10.1111/j.1365-313X.2007.03200.x. [DOI] [PubMed] [Google Scholar]
- Ishimaru K, Hirotsu N, Madoka Y, Murakami N, Hara N, Onodera H, Kashiwagi T, Ujiie K, Shimizu B, Onishi A, Miyagawa H, Katoh E. Loss of function of the IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield. Nat Genet. 2013;45:707–711. doi: 10.1038/ng.2612. [DOI] [PubMed] [Google Scholar]
- Itoh H, Ueguchi-Tanaka M, Sentoku N, Kitano H, Matsuoka M, Kobayashi M. Cloning and functional analysis of two gibberellin 3 beta-hydroxylase genes that are differently expressed during the growth of rice. Proc Natl Acad Sci U S A. 2001;98:8909–8914. doi: 10.1073/pnas.141239398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jiao Y, Wang Y, Xue D, Wang J, Yan M, Liu G, Dong G, Zeng D, Lu Z, Zhu X, Qian Q, Li J. Regulation of OsSPL14 by OsmiR156 defines ideal plant architecture in rice. Nat Genet. 2010;42:541–544. doi: 10.1038/ng.591. [DOI] [PubMed] [Google Scholar]
- Jin J, Huang W, Gao JP, Yang J, Shi M, Zhu MZ, Luo D, Lin HX. Genetic control of rice plant architecture under domestication. Nat Genet. 2008;40:1365–1369. doi: 10.1038/ng.247. [DOI] [PubMed] [Google Scholar]
- Kato T, Takeda K. Associations among characters related to yield sink capacity in space-planted rice. Crop Sci. 1996;36:1135–1139. doi: 10.2135/cropsci1996.0011183X003600050011x. [DOI] [Google Scholar]
- Komatsu M, Chujo A, Nagato Y, Shimamoto K, Kyozuka J. FRIZZY PANICLE is required to prevent the formation of axillary meristems and to establish floral meristem identity in rice spikelets. Development. 2003;130:3841–3850. doi: 10.1242/dev.00564. [DOI] [PubMed] [Google Scholar]
- Komatsu M, Maekawa M, Shimamoto K, Kyozuka J. The LAX1 and FRIZZY PANICLE 2 genes determine the inflorescence architecture of rice by controlling rachis-branch and spikelet development. Dev Biol. 2001;231:364–373. doi: 10.1006/dbio.2000.9988. [DOI] [PubMed] [Google Scholar]
- Kurakawa T, Ueda N, Maekawa M, Kobayashi K, Kojima M, Nagato Y, Sakakibara H, Kyozuka J. Direct control of shoot meristem activity by a cytokinin-activating enzyme. Nature. 2007;445:652–655. doi: 10.1038/nature05504. [DOI] [PubMed] [Google Scholar]
- Li F, Liu W, Tang J, Chen J, Tong H, Hu B, Li C, Fang J, Chen M, Chu C. Rice DENSE AND ERECT PANICLE 2 is essential for determining panicle outgrowth and elongation. Cell Res. 2010;20:838–849. doi: 10.1038/cr.2010.69. [DOI] [PubMed] [Google Scholar]
- Li S, Qian Q, Fu Z, Zeng D, Meng X, Kyozuka J, Maekawa M, Zhu X, Zhang J, Li J, Wang Y. Short panicle1 encodes a putative PTR family transporter and determines rice panicle size. Plant J. 2009;58:592–605. doi: 10.1111/j.1365-313X.2009.03799.x. [DOI] [PubMed] [Google Scholar]
- Li Y, Fan C, Xing Y, Jiang Y, Luo L, Sun L, Shao D, Xu C, Li X, Xiao J, He Y, Zhang Q. Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat Genet. 2011;43:1266–1269. doi: 10.1038/ng.977. [DOI] [PubMed] [Google Scholar]
- Luo J, Liu H, Zhou T, Gu B, Huang X, Shangguan Y, Zhu J, Li Y, Zhao Y, Wang Y, Zhao Q, Wang A, Wang Z, Sang T, Han B. An-1 encodes a basic helix-loop-helix protein that regulates awn development, grain size, and grain number in rice. Plant Cell. 2013;25:3360–3376. doi: 10.1105/tpc.113.113589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mori M, Nomura T, Ooka H, Ishizaka M, Yokota T, Sugimoto K, Okabe K, Kajiwara H, Satoh K, Yamamoto K, Hirochika H, Kikuchi S. Isolation and characterization of a rice dwarf mutant with a defect in brassinosteroid biosynthesis. Plant Physiol. 2002;130:1152–1161. doi: 10.1104/pp.007179. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qi P, Lin YS, Song XJ, Shen JB, Huang W, Shan JX, Zhu MZ, Jiang L, Gao JP, Lin HX. The novel quantitative trait locus GL3.1 controls rice grain size and yield by regulating Cyclin-T1;3. Cell Res. 2012;22:1666–1680. doi: 10.1038/cr.2012.151. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Qiao Y, Piao R, Shi J, Lee SI, Jiang W, Kim BK, Lee J, Han L, Ma W, Koh HJ. Fine mapping and candidate gene analysis of dense and erect panicle 3, DEP3, which confers high grain yield in rice (Oryza sativa L.) Theor Appl Genet. 2011;122:1439–1449. doi: 10.1007/s00122-011-1543-6. [DOI] [PubMed] [Google Scholar]
- Sakamoto T, Matsuoka M. Identifying and exploiting grain yield genes in rice. Curr Opin Plant Biol. 2008;11:209–214. doi: 10.1016/j.pbi.2008.01.009. [DOI] [PubMed] [Google Scholar]
- Seck PA, Diagne A, Mohanty S, Wopereis MCS. Crops that feed the world 7: rice. Food Sec. 2012;4:7–24. doi: 10.1007/s12571-012-0168-1. [DOI] [Google Scholar]
- Song XJ, Huang W, Shi M, Zhu MZ, Lin HX. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat Genet. 2007;39:623–630. doi: 10.1038/ng2014. [DOI] [PubMed] [Google Scholar]
- Suzaki T, Sato M, Ashikari M, Miyoshi M, Nagato Y, Hirano HY. The gene FLORAL ORGAN NUMBER1 regulates floral meristem size in rice and encodes a leucine-rich repeat receptor kinase orthologous to Arabidopsis CLAVATA1. Development. 2004;131:5649–5657. doi: 10.1242/dev.01441. [DOI] [PubMed] [Google Scholar]
- Tanabe S, Ashikari M, Fujioka S, Takatsuto S, Yoshida S, Yano M, Yoshimura A, Kitano H, Matsuoka M, Fujisawa Y, Kato H, Iwasaki Y. A novel cytochrome P 450 is implicated in brassinosteroid biosynthesis via the characterization of a rice dwarf mutant, dwarf11, with reduced seed length. Plant Cell. 2005;17:776–790. doi: 10.1105/tpc.104.024950. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wang E, Wang J, Zhu XD, Hao W, Wang LY, Li Q, Zhang LX, He W, Lu BR, Lin HX, Ma H, Zhang GQ, He ZH. Control of rice grain-filling and yield by a gene with a potential signature of domestication. Nat Genet. 2008;40:1370–1374. doi: 10.1038/ng.220. [DOI] [PubMed] [Google Scholar]
- Wang S, Li S, Liu Q, Wu K, Zhang J, Wang Y, Chen X, Zhang Y, Gao C, Wang F, Huang H, Fu X. The OsSPL16-GW7 regulatory module determines grain shape and simultaneously improves rice yield and grain quality. Nat Genet. 2015;47:949–954. doi: 10.1038/ng.3352. [DOI] [PubMed] [Google Scholar]
- Wang S, Wu K, Yuan Q, Liu X, Liu Z, Lin X, Zeng R, Zhu H, Dong G, Qian Q, Zhang G, Fu X. Control of grain size, shape and quality by OsSPL16 in rice. Nat Genet. 2012;44:950–954. doi: 10.1038/ng.2327. [DOI] [PubMed] [Google Scholar]
- Wang Y, Xiong G, Hu J, Jiang L, Yu H, Xu J, Fang Y, Zeng L, Xu E, Ye W, Meng X, Liu R, Chen H, Jing Y, Zhu X, Li J, Qian Q. Copy number variation at the GL7 locus contributes to grain size diversity in rice. Nat Genet. 2015;47:944–948. doi: 10.1038/ng.3346. [DOI] [PubMed] [Google Scholar]
- Weng J, Gu S, Wan X, Gao H, Guo T, Su N, Lei C, Zhang X, Cheng Z, Guo X, Wang J, Jiang L, Zhai H, Wan J. Isolation and initial characterization of GW5, a major QTL associated with rice grain width and weight. Cell Res. 2008;18:1199–1209. doi: 10.1038/cr.2008.307. [DOI] [PubMed] [Google Scholar]
- Wu YZ, Fu YC, Zhao SS, Gu P, Zhu ZF, Sun CQ, Tan LB. CLUSTERED PRIMARY BRANCH 1, a new allele of DWARF11, controls panicle architecture and seed size in rice. Plant Biotech J. 2016;14:377–386. doi: 10.1111/pbi.12391. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamaguchi R, Hwang LS. Food safety program in Asian countries. J Nutr Sci Vitaminol. 2015;61:S53–S54. doi: 10.3177/jnsv.61.S53. [DOI] [PubMed] [Google Scholar]
- Yamamuro C, Ihara Y, Wu X, Noguchi T, Fujioka S, Takatsuto S, Ashikari M, Kitano H, Matsuoka M. Loss of function of a rice brassinosteroid insensitive1 homolog prevents internode elongation and bending of the lamina joint. Plant Cell. 2000;12:1591–1605. doi: 10.1105/tpc.12.9.1591. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang CJ, Zhang C, Lu YN, Jin JQ, Wang XL. The mechanisms of brassinosteroids’ action: from signal transduction to plant development. Mol Plant. 2011;4:588–600. doi: 10.1093/mp/ssr020. [DOI] [PubMed] [Google Scholar]
- Zhao L, Tan L, Zhu Z, Xiao L, Xie D, Sun C. PAY1 improves plant architecture and enhances grain yield in rice. Plant J. 2015;83:528–536. doi: 10.1111/tpj.12905. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou Y, Miao J, Gu H, Peng X, Leburu M, Yuan F, Gao Y, Tao Y, Zhu J, Gong Z, Yi C, Gu M, Yang Z, Liang G. Natural variations in SLG7 regulate grain shape in rice. Genetics. 2015;201:1591–1599. doi: 10.1534/genetics.115.181115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou Y, Zhu J, Li Z, Yi C, Liu J, Zhang H, Tang S, Gu M, Liang G. Deletion in a quantitative trait gene qPE9-1 associated with panicle erectness improves plant architecture during rice domestication. Genetics. 2009;183:315–324. doi: 10.1534/genetics.109.102681. [DOI] [PMC free article] [PubMed] [Google Scholar]