

Abstract
Hepatitis C virus (HCV) infection is associated with B cell abnormality; however the phenotypic profiles of immunoglobulin (Ig)M+B cell subsets in patients with HCV infection remain unclear. In the current study, the effect of HCV infection on IgM+B cell subsets was evaluated. The percentages, as well as the differentiation and activation features of peripheral IgM+B naive subsets [cluster of differentiation (CD)27-IgM+B cells] and IgM+B memory subsets (CD27+IgM+B cells) were assessed using flow cytometry in 27 patients with chronic hepatitis C (CHC) and 20 healthy controls (HCs). The frequency of CD27+IgM+B memory subsets detected in patients with CHC was significantly higher than that in HCs (P<0.05). Although the frequency of CD27-IgM+B naive subsets was similar in both groups, there was a significantly higher proportion of CD5+B cells detected in the CD27-IgM+B subsets of patients with CHC compared with HCs (P<0.05). Among CD27-IgM+B subsets, abnormal differentiation was associated with HCV infection, with significantly increased percentages of IgD+B cells and CD38+B cells in patients with CHC compared with HCs (P<0.05). In CD27+IgM+B memory subsets, the abnormality of cell differentiation was associated with a significantly increased percentage of CD38+B cells in patients with CHC compared with HCs (P<0.05). In addition, the percentage of activated CD27+IgM+B subsets in patients with CHC were significantly higher than those observed in HCs (P<0.05). The number of CD27-IgD+IgM+B, CD27-CD38+IgM+B and CD27+CD38+IgM+B cells were negatively correlated with HCV RNA in patients with CHC. These results suggest that HCV infection contributes to abnormalities in the percentage, differentiation and activation of IgM+B cell subsets and may disrupt the immune response mediated by IgM+B cells.
Keywords: hepatitis C, immunoglobulin M+B cells, activation, differentiation
Introduction
Chronic hepatitis C virus (HCV) infection may cause chronic hepatitis and is associated with liver cirrhosis and hepatocellular carcinoma (1). Although HCV infection primarily affects the liver, the virus is capable of evading the immune response of the host by suppressing specific T and B cell responses (2). Furthermore, a number of autoimmune diseases have been identified in patients with chronic hepatitis C (CHC) (3), indicating that HCV infection disrupts the normal activity of the immune system. Therefore, improving understanding regarding immune-associated abnormalities in patients with CHC may help to control HCV progression and lead to the development of more effective treatment strategies.
Normal B cell development contributes to an effective immune response to remove pathogens, whereas disturbance of B cell differentiation and activation leads to the disruption of B cell homeostasis in chronic infection and autoimmunity disease (4–6). Previous studies have demonstrated that B cell disorders mediated by HCV infection are not only associated with B cell subset skewing, with an increased proportion of immature B cells and a decreased proportion of memory B cells (7–10), but are also associated with the increased activation of naive and memory B cells (11–13). Immunoglobulin (Ig) M+B cells are responsible for the production of IgM antibodies and are involved in the pathogenesis of viral infections and autoimmunity diseases (14–16). Roughan et al (17) determined that there is a higher frequency of circulating IgM+B memory subsets in the peripheral blood of patients with CHC; however the phenotypic features of different IgM+B cell subsets and associated risk factors in patients with CHC remain unknown.
The present study aimed to identify the effects of HCV infection on IgM+B cell subsets. The percentage, differentiation and activation status of peripheral IgM+B cell subsets was evaluated in patients with CHC using flow cytometry. In addition, the association between IgM+B cell subsets and different clinical parameters was investigated in patients with CHC.
Patients and methods
Patients and controls
The study population consisted of 27 patients with CHC with genotype 1b, including 11 male and 16 female patients (mean age 54.8 years), and 20 age- and sex-matched healthy controls (HCs; 9 male and 11 female patients; mean age 50 years). Patients with CHC were recruited in November 2013 from Guan county (China), in which the majority of patients are infected with HCV genotype 1b (18). Inclusion criteria of patients with chronic HCV infection were: Positive HCV antibodies and HCV RNA levels (>2,000 IU/ml) in the past 6 months. Exclusion criteria were: Co-infection with hepatitis B virus or human immunodeficiency virus. The HCs were tested negative for HCV antibodies. In addition, liver damage induced by HCV infection was monitored via measuring serum alanine aminotransferase (ALT). The baseline characteristics of all study subjects are presented in Table I. The present study was approved by the Ethics Committee of Peking University People's Hospital (Beijing, China) and all participants provided written informed consent.
Table I.
Baseline characteristics of subjects.
| Parameters | CHC | HC | P-values |
|---|---|---|---|
| Subjects (n) | 27 | 20 | N/A |
| Age (years) | 54.8±10.4 | 50.0±5.7 | 0.06a |
| Sex, male/female | 11/16 | 9/11 | 0.77 |
| ALT | 45.9±36.6 | 26.7±15.4 | 0.03a |
| Genotype (1b), n | 27 | N/A | N/A |
| HCV-RNA (log10 IU/ml) | 6.9±0.8 | N/A | N/A |
All data are presented as the mean ± standard deviation, unless otherwise specified.
P<0.05; CHC, chronic hepatitis C; HC, healthy control; HCV, hepatitis C virus; ALT, alanine transaminase; N/A, not applicable.
Clinical tests and peripheral blood mononuclear cell (PBMC) preparation
Levels of HCV antibodies, HCV RNA and HCV genotypes were determined as previously described (18). ALT was detected using a Hitachi 7600 automated biochemical analyzer (Hitachi, Ltd., Tokyo, Japan). PBMCs were isolated using density gradient centrifugation with Ficoll-Paque Plus (GE Healthcare Life Sciences, Uppsala, Sweden), as described previously (19). PBMCs were kept in liquid nitrogen until analysis.
Flow cytometric analysis
To measure the phenotypic features of peripheral IgM+B cell subsets, 1×106 PBMCs were incubated in 200 µl blocking solution consisting of 2% bovine serum albumin (Genview Corp., Houston, TX, USA) in PBS for 30 min at room temperature to block non-specific binding. Cells were stained with different fluorescent conjugated anti-human antibodies: Peridinin chlorophyll protein (PerCP)-conjugated anti-cluster of differentiation (CD)19 antibody (catalogue no. 340421; 1:100), allophycocyanin (APC)-conjugated anti-CD5 antibody (catalogue no. 340583; 1:100), APC-conjugated anti-IgD antibody (catalogue no. 561303; 1:100), APC-conjugated anti-CD10 antibody (catalogue no. 340923; 1:100) and phycoerythrin (PE)-conjugated anti-CD21 antibody (catalogue no. 555422; 1:100) were purchased from BD Pharmingen (BD Biosciences, San Jose, CA, USA). Fluorescein isothiocyanate (FITC)-conjugated anti-CD27 antibody (catalogue no. 11-0279; 1:100), APC-conjugated anti-CD38 antibody (catalogue no. 17-0389; 1:100), APC-conjugated anti-IgM antibody (catalogue no. 17-9998; 1:100), PE-conjugated anti-IgM antibody (catalogue no. 12-9998; 1:100), PE-conjugated anti-CD86 antibody (catalogue no. 12-0869; 1:100) and PE-conjugated anti-CD95 (catalogue no. 12-0959; 1:100) antibody were purchased from eBioscience (Thermo Fisher Scientific, Inc., Waltham MA, USA). Cells were incubated with these antibodies for 20 min at room temperature in the dark.
In addition, PBMCs were incubated with isotype-matched control antibodies listed below for 20 min at room temperature in the dark to determine background levels of staining. PerCP Mouse IgG1 κ Isotype Control (catalogue no. 559425; 1:100), APC Mouse IgG1, κ Isotype Control (catalogue no. 550854; 1:100) and APC Mouse IgG2b κ Isotype Control (catalogue no. 555745; 1:100) were purchased from BD Biosciences. Mouse IgG1 K isotype control FITC (catalogue no. 11-4714; 1:100), Mouse IgG2b K isotype control PE (catalogue no. 12-4732; 1:100), and Mouse IgG1 K isotype control PE (catalogue no. 12-4714; 1:100) were purchased from eBioscience (Thermo Fisher Scientific, Inc.).
Following washing twice with PBS, the cells were fixed with 1% paraformaldehyde in the dark at 4°C for 12 h. Then the expression of surface molecules on cells were assayed using the BD FACSCalibur (BD Biosciences) and data were analyzed using FlowJo 7.6 software (Tree Star, Inc., Ashland, OR, USA). Based on expression of CD5, B cells were divided into CD5+B cells (B1 cells) and CD5-B cells (B2 cells) (20). Depending on the expression of CD27, a distinct memory B cell marker, IgM+B cells were divided into naive subsets (CD27-IgM+B cells) and memory subsets (CD27+IgM+B cells) (17). In addition, CD21 was used to identify activated subsets (CD21-cells) and resting subsets (CD21+cells) in naïve B cells and memory B cells (7).
Statistical analysis
Data are presented as the mean ± standard deviation. Comparisons in all experiments were performed using Student's t-test or two-sample Wilcoxon rank-sum (Mann-Whitney) tests, according to data distribution measured using a Kolmogorow-Smirnov test with SPSS 16.0 software (SPSS, Inc., Chicago, IL, USA). The linear correlation between variables was analyzed using GraphPad Prism 5 (GraphPad Software, Inc., La Jolla, CA, USA). P<0.05 was considered to indicate a statistically significant difference.
Results
Frequency of CD27+IgM+B memory subsets in total B cells of patients with CHC
To characterize IgM+B cells (CD19+IgM+B cells) in patients infected with HCV, the frequency of IgM+B cells in patients with CHC and HCs was detected using flow cytometry (Fig. 1A). There were no significant differences between the frequency of total IgM+B cells in patients with CHC and HCs (Fig. 1B). The percentage of IgM+B naive subsets (CD27-IgM+B cells) and IgM+B memory subsets (CD27+IgM+B cells) were further assessed in both groups (Fig. 1A). As presented in Fig. 1B, there was no significant difference between the frequency of CD27-IgM+B cells in patients with CHC and HCs. However, the frequency of CD27+IgM+B cells was significantly higher in patients with CHC than in HCs (P<0.05; Fig. 1B). This suggests that infection with HCV may disrupt the homeostasis of IgM+B cell subsets.
Figure 1.
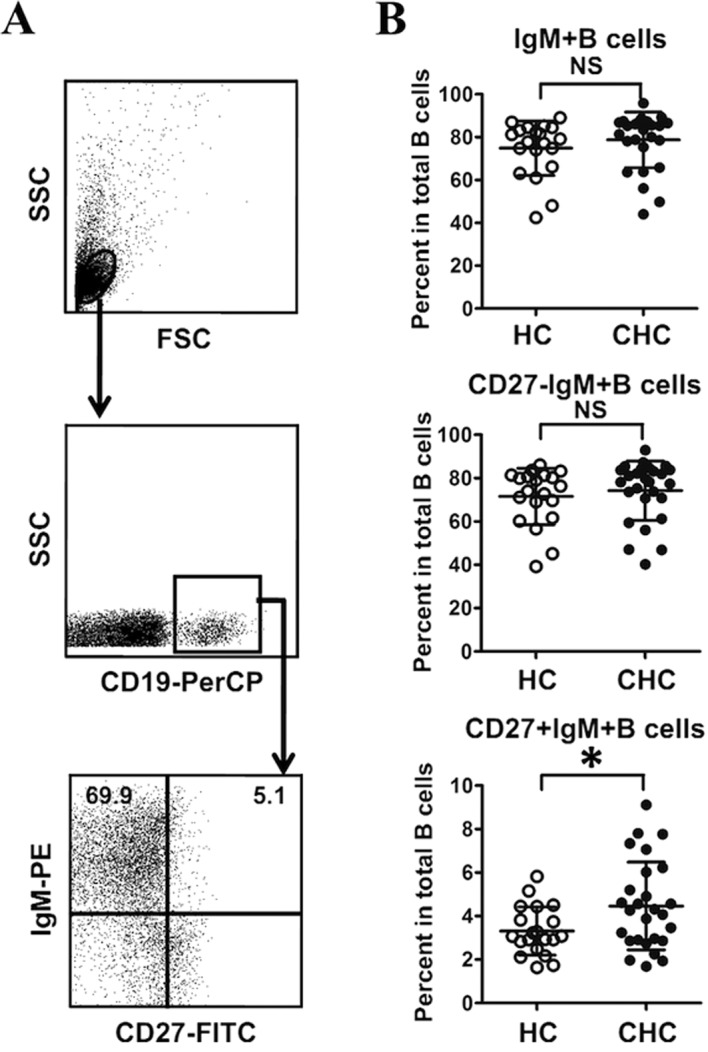
Frequency of CD27+IgM+B cells in patients with CHC and HCs. (A) Gating strategy for IgM+B cell subsets by flow cytometry. (B) Frequencies of IgM+B, CD27-IgM+B and CD27+IgM+B cells in total B cells. *P<0.05. PerCP, peridinin chlorophyll; PE, phycoerythrin; CHC, chronic hepatitis C; HC, healthy control; NS, no significant difference; FSC, forward scatter; SSC, side scatter; Ig, immunoglobulin; FITC, Fluorescein isothiocyanate; CD, cluster of differentiation.
Percentage of CD5+B cells in CD27-IgM+B cells of patients with CHC
Previous research has demonstrated that expansion of CD5+B cells is associated with HCV infection (21), whereas the association between CD5+B cells and IgM+B cell subsets is unclear in patients with HCV infection. In the current study, the percentage of CD5+B cells in total B cells was significantly higher in patients with CHC compared with HCs (P<0.05; Fig. 2), which was consistent with a previous report (21). The percentage of CD5+B cells in IgM+B cell subsets was also measured in the current study. The results indicated that the frequency of CD5+B cells was significantly higher in CD27-IgM+B cells from patients with CHC compared with HCs (P<0.05; Fig. 2), while there was no significant difference in the frequency of CD5+B in CD27+IgM+B cells between the two groups (Fig. 2). This suggests that only CD27-IgM+B cells are associated with expansion of CD5+B cells in HCV infection.
Figure 2.

Frequency of CD5+B cells in patients with CHC. (A) Gating strategy for measuring the frequency of CD5+B cells in different B cell subsets by flow cytometry. (B) Frequency of CD5+B cells in total B cells. (C) Frequency of CD5+B cells in CD27-IgM+B cells. (D) Frequency of CD5+B cells in CD27+IgM+B cells. *P<0.05. CHC, chronic hepatitis C; HC, healthy control; NS, no significant difference; SSC, side scatter; PE, phycoerythrin; FITC, fluorescein isothiocyanate; PerCP, peridinin chlorophyll; APC, allophycocyanin; Ig, immunoglobulin; CD, cluster of differentiation.
Abnormal differentiation of CD27-IgM+B and CD27+IgM+B subsets in patients with CHC
B cell homeostasis is primarily dependent on B cell differentiation and activation (22,23). Thus, the differentiation of IgM+B cells in both IgM+B cell subsets was evaluated in the current study. IgM+B cells at the differentiation stage transition from immature B cells to plasmablasts (22,23). In the current study, immature B cells were CD27-CD10+B cells (Fig. 3A). The percentage of CD27-CD10+B cells in IgM+B cells was determined in both groups, and the results demonstrated that the frequency of CD27-CD10+IgM+B cells in IgM+B cells was significantly higher in patients with CHC compared with HCs (P<0.05; Fig. 3B). At naïve B cells differentiate into plasmablasts, B cell differentiation is associated with alterations in IgD and CD38 expression (22,23) and the current study measured the expression of IgD and CD38 in IgM+B cell subsets. As presented in Fig. 3C, a significantly higher percentage of IgD+B cells in CD27-IgM+B cells was identified in patients with CHC compared with HCs (P<0.05), whereas there were no significant differences in the frequency of IgD+B cells in CD27+IgM+B cells between the two groups. Furthermore, a significantly higher percentage of CD38+B cells was observed in CD27-IgM+B and CD27+IgM+B cells in patients with CHC compared with HCs (P<0.05; Fig. 3D). Together, these results indicate that HCV infection is associated with the abnormal differentiation of CD27-IgM+B and CD27+IgM+B cells.
Figure 3.

Abnormal differentiation of IgM+B cell subsets in patients with CHC. (A) Gating strategy for measuring the differentiation of IgM+B cell subsets by flow cytometry. (B) Frequency of CD27-CD10+B cells in IgM+B cells. (C) Frequency of IgD+B cells in IgM+B cell subsets. (D) Frequency of CD38+B cells in IgM+B cell subsets. *P<0.05. CHC, chronic hepatitis C; HC, healthy control; NS, no significant difference; PE, phycoerythrin; PerCP, peridinin chlorophyll; Ig, immunoglobulin; CD, cluster of differentiation; APC, allophycocyanin.
Activation of CD27+IgM+B subsets in patients with CHC
The activation of IgM+B subsets in both groups was investigated (Fig. 4A). As shown in Fig. 4B, compared with HCs, the number of activated CD27+IgM+B cells (CD27+IgM+CD21-B cells) increased significantly in patients with CHC (P<0.05); however, there were no significant differences between the number of activated CD27-IgM+B cells (CD27-IgM+CD21-B cells) in patients with CHC compared with HCs (Fig. 4B).
Figure 4.
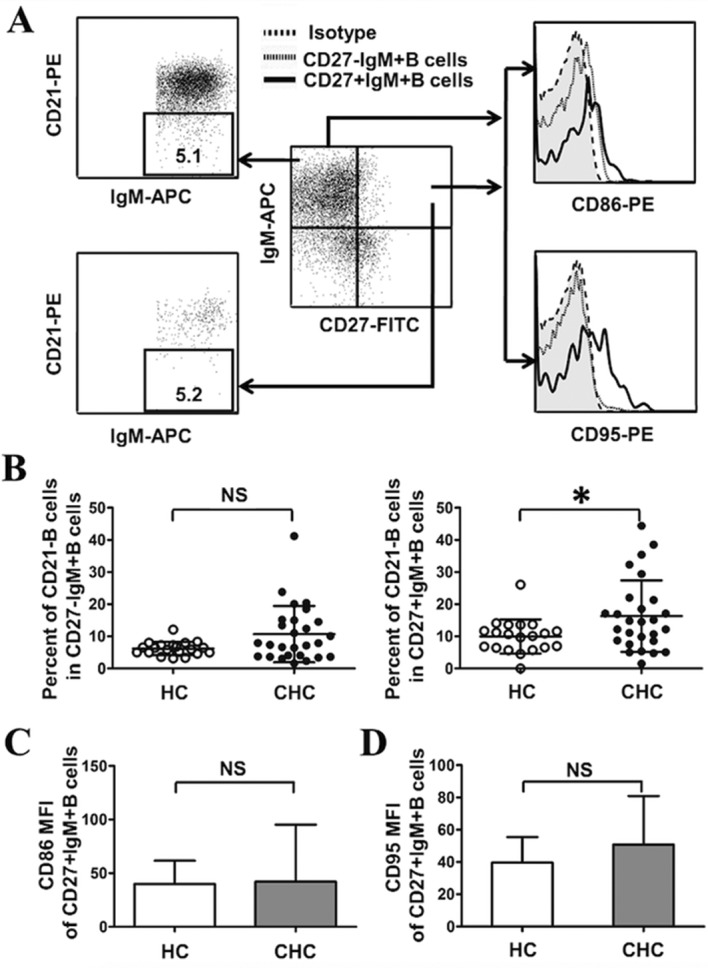
Activation of CD27+IgM+B cell subsets in patients with CHC. (A) Gating strategy to detect IgM+B cell subsets and activation markers. (B) Frequencies of CD21-B cells in IgM+B subsets. (C) Expression of CD86 in CD27+IgM+B cells. (D) Expression of CD95 in CD27+IgM+B subsets. *P<0.05. CHC, chronic hepatitis C; HC, healthy control; NS, no significant difference; PE, phycoerythrin; APC, allophycocyanin; FITC, fluorescein isothiocyanate; MFI, mean fluorescence intensity; Ig, immunoglobulin; CD, cluster of differentiation.
Activated B cells are associated with high levels of the co-stimulatory molecules CD86 and CD95, and the abnormal expression of these molecules in B cells is associated with chronic diseases such as autoimmune hepatitis and rheumatoid arthritis (24,25). Based on this, levels of CD86 and CD95 were evaluated in CD27+IgM+B cells, using mean fluorescence intensity to represent molecule expression. The results demonstrated that the expression of CD86 and CD95 in CD27+IgM+B cells was similar in patients with CHC and HCs (Fig. 4C and D). Taken together, these results suggest that HCV infection is associated with the activation of CD27+IgM+B cell subsets but is not associated with the expression of CD86 and CD95 in CD27+IgM+B cells.
Correlation of HCV RNA and ALT with IgM+B cell subsets in patients with CHC
In order to evaluate the association between IgM+B cell subsets and clinical parameters in patients with CHC, it was investigated whether IgM+B subsets were correlated with HCV RNA and the liver cell damage marker ALT. As presented in Fig. 5A, the percentages of CD27-IgD+IgM+B cells (P<0.01, r2=0.49), CD27-CD38+IgM+B cells (P<0.01, r2=0.34) and CD27+CD38+IgM+B cells (P=0.01, r2=0.23) were negatively correlated with viral load in patients with CHC. However, no significant correlations between any IgM+B cell subsets and ALT were detected in patients with CHC (Fig. 5B). Thus, these data indicate that there is a HCV but not ALT is negatively correlated with the abnormal expression of certain IgM+B cell subsets.
Figure 5.
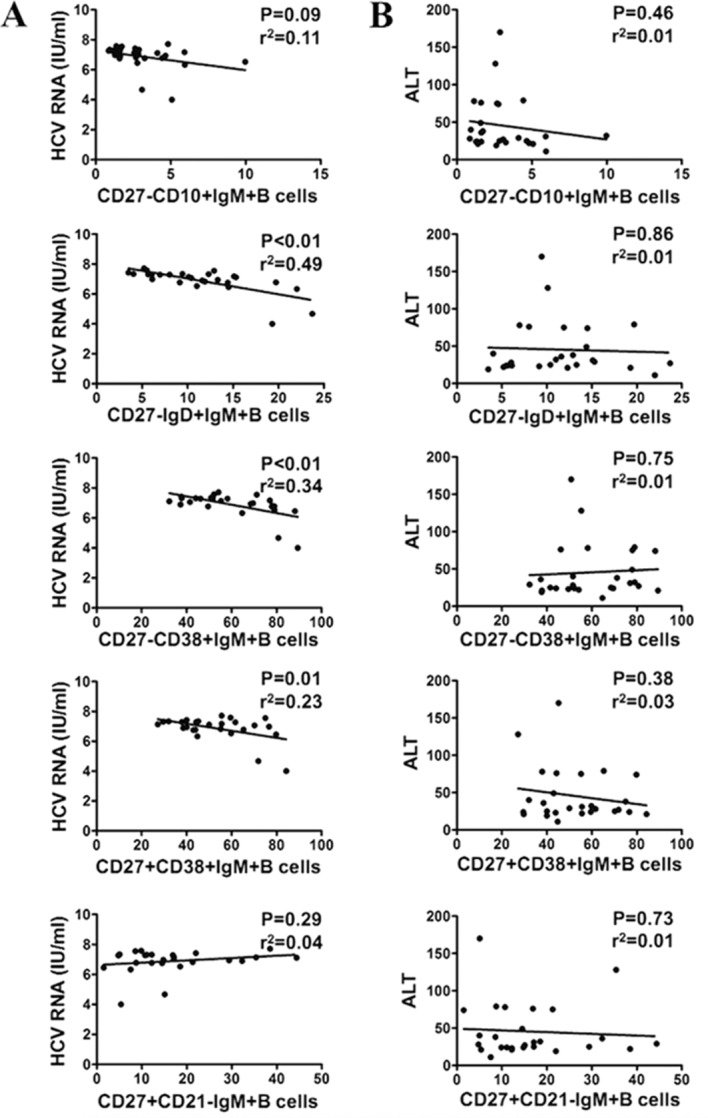
Correlation analysis of HCV RNA and ALT with different IgM+B cell subsets in patients with CHC. (A) Correlation of HCV RNA with CD27-CD10+IgM+B, CD27-IgD+IgM+B, CD27-CD38+IgM+B cells, CD27+CD38+IgM+B and CD27+CD21-IgM+B cells. (B) Correlation of ALT with CD27-CD10+IgM+B cells, CD27-IgD+IgM+B cells, CD27-CD38+IgM+B cells, CD27+CD38+IgM+B cells, and CD27+CD21-IgM+B cells. HCV, hepatitis C virus; ALT, alanine transaminase; CD, cluster of differentiation; Ig, immunoglobulin.
Discussion
It has been demonstrated that abnormalities of peripheral B cell subsets are associated with HCV infection (7–13). However, the effect of HCV infection on the differentiation and activation of IgM+B cell subsets has not been fully elucidated. The current study aimed to characterize IgM+B cell subsets in patients with CHC and the results indicated that HCV infection was responsible for abnormalities in the percentage, differentiation and activation of IgM+B cell subsets. In addition, negative correlations between viral load and CD27-IgD+IgM+B cells, CD27-CD38+IgM+B cells and CD27+CD38+IgM+B cells were detected in patients with CHC.
Roughan et al (17) demonstrated that IgM+ memory B cells were expanded in patients with CHC, and identified that the unusual polyclonal expansion of the IgM+ memory B cell subset was made up of autoreactive B cells. Consistent with a previous report (17), the present study demonstrated that an increase in CD27+IgM+B cells was associated with HCV infection. The frequency of CD27-IgM+B cells in patients with CHC was similar to that in HCs. However, the percentage of CD5+B cells, which are not only characterized by the production of low-affinity IgM that recognizes a variety of self-antigens but also serve an important role in the pathogenesis of autoimmunity disease (26), was higher in the CD27-IgM+B cells of patients with CHC than in HCs. This implies that CD27-IgM+B cells may be involved in the reaction to autoantigens in patients with CHC. Taken together, these results suggest that abnormalities of CD27-IgM+B and CD27+IgM+B cells may be associated with autoreactivity and increased susceptibility to autoimmune diseases in patients with CHC.
Alterations in B cell differentiation and activation are frequently noted in pathological conditions (4–6). On this basis, the current study evaluated the differentiation status of IgM+B cell subsets in patients with CHC. Based on the immature B cell marker CD10, it was determined that the percentage of CD27-CD10+IgM+B cells was increased in patients with CHC. Previous results have demonstrated that immature B cells are CD10+CD27-IgM+B cells (27) and the current results are consistent with those from a previous study, which identified that HCV infection was associated with an increase in immature B cells (8). IgD and CD38 are associated with the development of B cells (22,23). In the present study, it was observed that the frequency of CD27-IgD+B cells in IgM+B cells was increased in patients with CHC compared with HCs. The increased frequency of this subpopulation in IgM+B cells may be associated with chronic HCV stimulation and immune dysfunction mediated by CD27-IgD-IgM+B cells. CD38 is a multifunctional ectoenzyme, which participates in lymphocyte activation and terminal differentiation (28). The increased percentage of CD27-CD38+IgM+B cells and CD27+CD38+IgM+B cells imply that chronic HCV infection may induce the activation and terminal differentiation of certain IgM+B cell subsets.
Previous studies have demonstrated that HCV infection induces the activation of naïve and memory B cell subsets (11,12,29). In the current study, the activation of IgM+B cell subsets was assessed based on the expression of the activation marker CD21. It was determined that the activation of CD27+IgM+B cells was increased in patients with CHC. Furthermore, the expression of distinct molecules associated with B cell activation was evaluated in CD27+IgM+B cells. CD86 is a critical co-stimulatory molecule to facilitate the interaction of T cells with B cells (30). CD95 is a death receptor, which initiates extrinsic pathways of apoptosis to remove activated immune cells (31). Although HCV infection was associated with an increase in the number of activated CD27+IgM+B cells, these activated CD27+IgM+B cell subsets were not correlated with CD86 and CD95 expression. This implies that the activation of CD27+IgM+B cells may not be associated with T cell stimulation or apoptosis. Further studies are required to determine the underlying mechanisms that result in increased numbers of activated CD27+IgM+B subsets in the peripheral circulation.
In the present study, the association between IgM+B cell subsets and clinical parameters was investigated in patients with CHC. Negative correlations were identified between HCV RNA and CD27-IgD+IgM+B, CD27-CD38+IgM+B and CD27+CD38+IgM+B cells. Although the relative expression of CD27-IgD+IgM+B, CD27-CD38+IgM+B and CD27+CD38+IgM+B cells was higher in patients with CHC compared with HCs, the exact roles of these IgM+B cell subsets remain unknown. It is possible that chronic HCV antigen stimulation disrupts the immune response by increasing these IgM+B cell subsets, which, in turn, helps to inhibit HCV replication to some extent in pathological conditions. Further studies are required to determine the roles of these IgM+B cell subsets in patients with CHC.
In conclusion, the current study identified the phenotypic abnormalities of CD27-IgM+B and CD27+IgM+B cells in patients with CHC and demonstrated that the frequencies of IgM+B cell subsets were negatively correlated with viral load. The alterations in IgM+B cell subset populations may be associated with the disruption of the B cell immune response and improved understanding of the association between IgM+B cells and HCV infection will be required in order to identify potential targets for HCV treatment. Despite various limitations of the current study, including the relatively small number of patients with CHC enrolled and a lack of functional analysis of IgM+B cell subsets in the pathogenic process of HCV infection, the results of the current study lay a foundation for evaluating the roles and associated mechanisms of certain IgM+B cell subsets in patients with HCV.
Acknowledgements
The authors wish to thank all patients with CHC for providing blood samples. The current study was funded by the National Science and Technology Major Project for Infectious Diseases Control during the 12th Five-Year Plan Period (grant no. 2012ZX10002003).
References
- 1.Manns MP, Buti M, Gane E, Pawlotsky JM, Razavi H, Terrault N, Younossi Z. Hepatitis C virus infection. Nat Rev Dis Primers. 2017;3:17006. doi: 10.1038/nrdp.2017.6. [DOI] [PubMed] [Google Scholar]
- 2.Park SH, Rehermann B. Immune responses to HCV and other hepatitis viruses. Immunity. 2014;40:13–24. doi: 10.1016/j.immuni.2013.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Ferrari SM, Fallahi P, Mancusi C, Colaci M, Manfredi A, Ferri C, Antonelli A. HCV-related autoimmune disorders in HCV chronic infection. Clin Ter. 2013;164:e305–e312. doi: 10.7417/CT.2013.1594. [DOI] [PubMed] [Google Scholar]
- 4.Moir S, Fauci AS. B cells in HIV infection and disease. Nat Rev Immunol. 2009;9:235–245. doi: 10.1038/nri2524. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Sang A, Zheng YY, Morel L. Contributions of B cells to lupus pathogenesis. Mol Immunol. 2014;62:329–338. doi: 10.1016/j.molimm.2013.11.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Perry HM, Bender TP, McNamara CA. B cell subsets in atherosclerosis. Front Immunol. 2012;3:373. doi: 10.3389/fimmu.2012.00373. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sugalski JM, Rodriguez B, Moir S, Anthony DD. Peripheral blood B cell subset skewing is associated with altered cell cycling and intrinsic resistance to apoptosis and reflects a state of immune activation in chronic hepatitis C virus infection. J Immunol. 2010;185:3019–3027. doi: 10.4049/jimmunol.1000879. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Holz LE, Yoon JC, Raghuraman S, Moir S, Sneller MC, Rehermann B. B cell homeostasis in chronic hepatitis C virus-related mixed cryoglobulinemia is maintained through naïve B cell apoptosis. Hepatology. 2012;56:1602–1610. doi: 10.1002/hep.25821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Racanelli V, Frassanito MA, Leone P, Galiano M, De Re V, Silvestris F, Dammacco F. Antibody production and in vitro behavior of CD27-defined B-cell subsets: Persistent hepatitis C virus infection changes the rules. J Virol. 2006;80:3923–3934. doi: 10.1128/JVI.80.8.3923-3934.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Mizuochi T, Ito M, Saito K, Kasai M, Kunimura T, Morohoshi T, Momose H, Hamaguchi I, Takai K, Iino S, et al. Possible recruitment of peripheral blood CXCR3+ CD27+ CD19+ B cells to the liver of chronic hepatitis C patients. J Interferon Cytokine Res. 2010;30:243–252. doi: 10.1089/jir.2009.0047. [DOI] [PubMed] [Google Scholar]
- 11.Rosa D, Saletti G, De Gregorio E, Zorat F, Comar C, D'Oro U, Nuti S, Houghton M, Barnaba V, Pozzato G, Abrignani S. Activation of naïve B lymphocytes via CD81, a pathogenetic mechanism for hepatitis C virus-associated B lymphocyte disorders; Proc Natl Acad Sci USA; 2005; pp. 18544–18549. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Oliviero B, Cerino A, Varchetta S, Paudice E, Pai S, Ludovisi S, Zaramella M, Michelone G, Pugnale P, Negro F, et al. Enhanced B-cell differentiation and reduced proliferative capacity in chronic hepatitis C and chronic hepatitis B virus infections. J Hepatol. 2011;55:53–60. doi: 10.1016/j.jhep.2010.10.016. [DOI] [PubMed] [Google Scholar]
- 13.Oliviero B, Mantovani S, Ludovisi S, Varchetta S, Mele D, Paolucci S, Baldanti F, Mondelli MU. Skewed B cells in chronic hepatitis C virus infection maintain their ability to respond to virus-induced activation. J Viral Hepat. 2015;22:391–398. doi: 10.1111/jvh.12336. [DOI] [PubMed] [Google Scholar]
- 14.Valle L Della, Dohmen SE, Verhagen OJ, Berkowska MA, Vidarsson G, van der Schoot C Ellen. The majority of human memory B cells recognizing RhD and tetanus resides in IgM+ B cells. J Immunol. 2014;193:1071–1079. doi: 10.4049/jimmunol.1400706. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.D'Orsogna LJ, Krueger RG, McKinnon EJ, French MA. Circulating memory B-cell subpopulations are affected differently by HIV infection and antiretroviral therapy. AIDS. 2007;21:1747–1752. doi: 10.1097/QAD.0b013e32828642c7. [DOI] [PubMed] [Google Scholar]
- 16.Carsetti R, Rosado MM, Donnanno S, Guazzi V, Soresina A, Meini A, Plebani A, Aiuti F, Quinti I. The loss of IgM memory B cells correlates with clinical disease in common variable immunodeficiency. J Allergy Clin Immunol. 2005;115:412–417. doi: 10.1016/j.jaci.2004.10.048. [DOI] [PubMed] [Google Scholar]
- 17.Roughan JE, Reardon KM, Cogburn KE, Quendler H, Pockros PJ, Law M. Chronic hepatitis C virus infection breaks tolerance and drives polyclonal expansion of autoreactive B cells. Clin Vaccine Immunol. 2012;19:1027–1037. doi: 10.1128/CVI.00194-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Rao H, Wei L, Lopez-Talavera JC, Shang J, Chen H, Li J, Xie Q, Gao Z, Wang L, Wei J, et al. Distribution and clinical correlates of viral and host genotypes in Chinese patients with chronic hepatitis C virus infection. J Gastroenterol Hepatol. 2014;29:545–553. doi: 10.1111/jgh.12398. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Guo Z, Zhang H, Rao H, Jiang D, Cong X, Feng B, Wang J, Wei L, Chen H. DCs pulsed with novel HLA-A2-restricted CTL epitopes against hepatitis C virus induced a broadly reactive anti-HCV-specific T lymphocyte response. PLoS One. 2012;7:e38390. doi: 10.1371/journal.pone.0038390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Hardy RR. B-1 B cell development. J Immunol. 2006;177:2749–2754. doi: 10.4049/jimmunol.177.5.2749. [DOI] [PubMed] [Google Scholar]
- 21.Curry MP, Golden-Mason L, Nolan N, Parfrey NA, Hegarty JE, O'Farrelly C. Expansion of peripheral blood CD5+ B cells is associated with mild disease in chronic hepatitis C virus infection. J Hepatol. 2000;32:121–125. doi: 10.1016/S0168-8278(00)80198-1. [DOI] [PubMed] [Google Scholar]
- 22.Perez-Andres M, Paiva B, Nieto WG, Caraux A, Schmitz A, Almeida J, Vogt RF, Jr, Marti GE, Rawstron AC, Van Zelm MC, et al. Human peripheral blood B-cell compartments: A crossroad in B-cell traffic. Cytometry B Clin Cytom. 2010;78(Suppl 1):S47–S60. doi: 10.1002/cyto.b.20547. [DOI] [PubMed] [Google Scholar]
- 23.Bemark M, Holmqvist J, Abrahamsson J, Mellgren K. Translational Mini-Review Series on B cell subsets in disease. Reconstitution after haematopoietic stem cell transplantation-revelation of B cell developmental pathways and lineage phenotypes. Clin Exp Immunol. 2012;167:15–25. doi: 10.1111/j.1365-2249.2011.04469.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ma L, Qin J, Ji H, Zhao P, Jiang Y. Tfh and plasma cells are correlated with hypergammaglobulinaemia in patients with autoimmune hepatitis. Liver Int. 2014;34:405–415. doi: 10.1111/liv.12245. [DOI] [PubMed] [Google Scholar]
- 25.Wang J, Shan Y, Jiang Z, Feng J, Li C, Ma L, Jiang Y. High frequencies of activated B cells and T follicular helper cells are correlated with disease activity in patients with new-onset rheumatoid arthrits. Clin Exp Immunol. 2013;174:212–220. doi: 10.1111/cei.12162. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Youinou P, Renaudineau Y. CD5 expression in B cells from patients with systemic lupus erythematosus. Crit Rev Immunol. 2011;31:31–42. doi: 10.1615/CritRevImmunol.v31.i1.30. [DOI] [PubMed] [Google Scholar]
- 27.Dörner T, Giesecke C, Lipsky PE. Mechanisms of B cell autoimmunity in SLE. Arthritis Res Ther. 2011;13:243. doi: 10.1186/ar3433. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Deterre P, Berthelier V, Bauvois B, Dalloul A, Schuber F, Lund F. CD38 in T- and B-cell functions. Chem Immunol. 2000;75:146–168. doi: 10.1159/000058767. [DOI] [PubMed] [Google Scholar]
- 29.Kong FY, Feng B, Zhang HH, Rao HY, Wang JH, Cong X, Wei L. CD4+CXCR5+ T cells activate CD27+IgG+ B cells via IL-21 in patients with hepatitis C virus infection. Hepatobiliary Pancreat Dis Int. 2016;15:55–64. doi: 10.1016/S1499-3872(16)60054-1. [DOI] [PubMed] [Google Scholar]
- 30.Lim TS, Goh JK, Mortellaro A, Lim CT, Hämmerling GJ, Ricciardi-Castagnoli P. CD80 and CD86 differentially regulate mechanical interactions of T-cells with antigen-presenting dendritic cells and B-cells. PLoS One. 2012;7:e45185. doi: 10.1371/journal.pone.0045185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Strasser A, Jost PJ, Nagata S. The many roles of FAS receptor signaling in the immune system. Immunity. 2009;30:180–192. doi: 10.1016/j.immuni.2009.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]