

Abstract
Hypoxia is an important feature of the microenvironment of a wide range of solid tumours. Its critical role in radio‐ and chemoresistance and its significance as an adverse prognostic factor have been well established over the last decades. On a cellular level, hypoxia evokes a complex molecular response with a central role for the HIF‐1 pathway. The cellular processes under control of HIF‐1 contain important prognostic information and comprise potential candidates for directing hypoxia‐modifying therapies. This review will provide an overview of the current knowledge on the molecular aspects of tumour hypoxia and the link to clinical practice.
Keywords: Tumour hypoxia; HIF-1; Hypoxic markers; Molecular response; Unfolded protein response, UPR
1. Introduction
The tumour microenvironment is of great importance, influencing malignant cells in various ways (Hu and Polyak, 2008). Within this microenvironment, hypoxia is an extensively studied parameter, with relevance in almost all types of solid tumours. As early as 1936 Mottram mentions the relative insensitivity of tumours under anaerobic conditions (Mottram, 1936). Gray described in 1953 the presence of hypoxia in murine tumour cells and the associated reduced sensitivity to radiotherapy (Gray et al., 1953). Hypoxia has negative implications for clinical outcome. This is probably based on two distinct principles: hypoxic cells are more resistant to radiotherapy and chemotherapy, and they give rise to genetic instability and more aggressive phenotypes.
Especially the increased resistance to radiotherapy is a well‐known phenomenon associated with tumour hypoxia, most studied in head and neck cancer and cervical carcinoma (Hockel et al., 1996; Nordsmark et al., 2005). Besides this increased resistance to radiotherapy, there is evidence that hypoxic cells are responsible for decreased sensitivity to certain chemotherapeutic agents as well, such as doxorubicin, 5‐fluorouracil and methotrexate. These data are mainly derived from animal and in vitro studies (Grau and Overgaard, 1992; Matthews et al., 2001; Wilson et al., 1989). There is scarce recent literature available on this subject.
Besides this resistance to chemo‐ and radiotherapy, surgically treated soft‐tissue sarcomas and cervical carcinomas also exhibit hypoxia as a prognostic factor for poor survival (Hockel et al., 1996; Nordsmark et al., 2001). A correlation was found between the rate of distant metastases and tumour oxygenation in soft‐tissue sarcoma (Brizel et al., 1996). Furthermore, in cervical cancer it has been proven that hypoxic tumours exhibit more frequent parametrial spread and lymph‐vascular space involvement (Hockel et al., 1996). A subgroup of hypoxic tumours with diminished apoptotic potential showed increased lymphatic spread and higher probability of recurrence as well (Hockel et al., 1999). These findings indicate that hypoxia stimulates tumour cells to develop towards a more invasive phenotype.
The measurement of hypoxia has been the subject of investigation for years, and oxygen electrode measurement has been considered the gold standard for sometime. 2‐Nitroimidazoles, such as pimonidazole and EF5, are exogenous hypoxic cell markers, which bind to viable hypoxic cells in vivo. Analysis of these exogenous hypoxic cell markers has largely replaced the oxygen electrode as standard assay for quantifying hypoxia. They have gained interest as a prognostic factor, although with varying results (Evans et al., 2007; Kaanders et al., 2002c; Nordsmark et al., 2006). Now the main research focus is on exploring the potential of endogenous hypoxia‐related markers, like hypoxia‐inducible factor‐1 (HIF‐1), carbonic anhydrase IX (CAIX), glucose transporters (GLUT‐1 and GLUT‐3), plasminogen activator inhibitor‐1 (PAI‐1), vascular endothelial growth factor (VEGF) and osteopontin (OPN). Most of these proteins are under control of the HIF‐1 pathway, which plays a central role in the cellular adaptation to hypoxic conditions. Another important adaptive mechanism is the unfolded protein response (UPR). Impairment of the tumour oxygenation status can lead to the accumulation of misfolded proteins in the endoplasmatic reticulum (ER). The accumulation of unfolded proteins activates the UPR, which may inhibit apoptosis. These cellular responses to hypoxia are complex, with many different genes and proteins involved. In this review the background and prognostic and predictive value of the potential markers will be discussed. Elucidating more about the response to hypoxia and establishing the value of the different markers may lead to the development of a predictive profile that enables the selection of patients for hypoxia‐modifying treatments.
2. Pathophysiology
In most malignant tumours there is an imbalance between the supply and consumption of oxygen, leading to hypoxic and even anoxic regions. During the rapid growth of a tumour an aberrant, chaotic microvasculature develops. Tumours exhibit a vascular network with a wide range of vessel diameters, intervascular distances and interbranching distances unlike those seen in normal tissue (Konerding et al., 1995). The morphological and functional deformed blood vessels diminish the oxygen delivery to the tumour cells, resulting in a low oxygen microenvironment. Another factor contributing to a decrease in oxygen supply is tumour‐associated and therapy‐induced anaemia, leading to a diminished oxygen transport capacity of the blood (anaemic hypoxia) (Vaupel et al., 2001).
From a pathophysiological point of view hypoxia can be divided into acute, or perfusion‐limited hypoxia and chronic, or diffusion‐limited hypoxia, although this division is somewhat arbitrary (Dewhirst, 1998). Acute hypoxia is often transient and is caused by a temporary disruption in blood flow as a result of an occlusion or rise in interstitial fluid pressure. In xenografts only a small part of the tumour hypoxia is of the intermittent type (Bennewith and Durand, 2004).
Chronic hypoxia arises with an increased diffusion distance of tumour cells from the microvessels (>70μm) in the distorted architecture of the vascular network. Acute and chronic hypoxia combine in a heterogeneous pattern throughout the tumour so that severe, intermediate and low levels of hypoxia can be identified. From a clinical standpoint it has been proposed that the intermediate hypoxic cells are the most important for prognosis (Wouters and Brown, 1997). These cells are resistant to therapy and still have the ability to proliferate (Hoogsteen et al., 2005).
Because the degree of hypoxia is extremely variable on a continuous scale, a strict cut‐off value to discriminate normoxic from hypoxic cells does not exist. Critical metabolic processes fail at different pO2‐levels (Figure 1). A median O2 partial pressure of less than 10mm Hg already results in ATP depletion. The oxidative phosphorylation gradually declines below this value till it ceases at O2 partial pressures of less than 0.5mmHg. Radioresistance may already occur below 25–30mmHg (Hockel and Vaupel, 2001). In most experiments, values from 0.5mmHg to 10mmHg have been used as a cut‐off value to discriminate normoxic from hypoxic tumour cells.
Figure 1.

Cellular adaptation to hypoxia. The bars show the approximate hypoxic values below which cellular responses gradually change. Figure adapted from Koumenis and Wouters (2006) and Hockel and Vaupel (2001).
3. Measuring hypoxia
Over the years different methods have been developed for assessing the level of hypoxia in tumours in vivo, with the polarographic oxygen electrode mostly used as the gold standard. However, the disadvantage of this method led to the development of less invasive techniques. Nowadays, the focus has shifted to exogenous markers, like EF5 and pimonidazole and a wide range of hypoxia‐related endogenous markers (Figure 2). Measuring hypoxia with imaging modalities such as magnetic resonance imaging (MRI) and positron emission tomography (PET) is another promising field of research.
Figure 2.
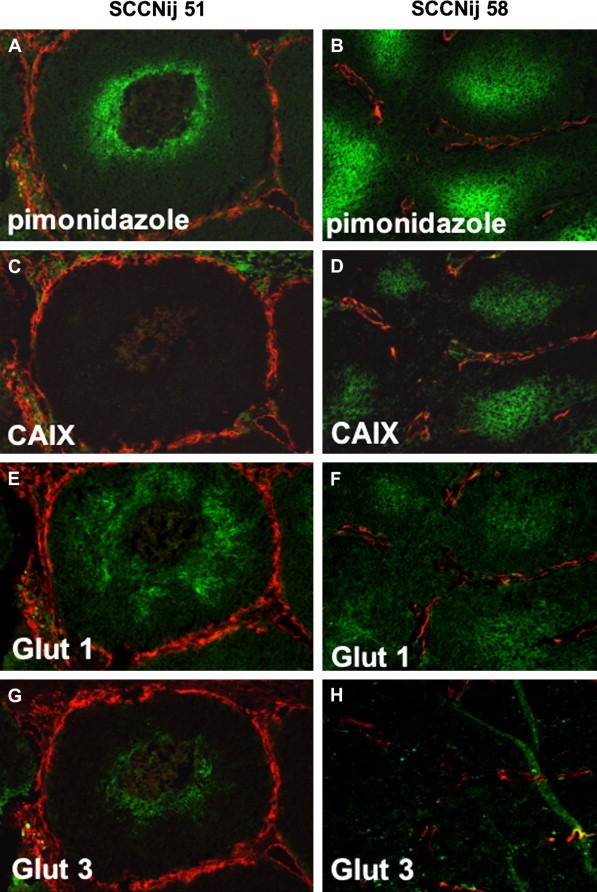
Photomicrographs of two human squamous‐cell carcinoma xenograft cell lines (SCCNij51 and SCCNij58) after immunofluorescent staining. The images show the differences in colocalisation of the exogenous marker pimonidazole (A and B) and three endogenous hypoxi a‐related markers: CAIX (C and D), GLUT‐1 (E and F) and GLUT‐3 (G and H) (all in green), relative to the vasculature (in red).
3.1. Imaging hypoxia
The non‐invasive assessment of tumour hypoxia is an active area of research, with an emphasis on PET and blood oxygenation level dependent magnetic resonance imaging (BOLD‐MRI) as promising modalities (review by Padhani et al., 2007).
With PET it is possible to quantify hypoxia in a well‐tolerated non‐invasive procedure with the use of short‐lived positron emitting radionuclides. Several 18F‐ or 64Cu‐containing compounds have been developed to estimate the hypoxic fraction, like 18F‐EF1, 18F‐MISO and 60/64Cu‐ATSM, with 18F‐MISO being the most widely used and investigated. These tracers have been tested in several clinical studies for their prognostic and predictive value, showing encouraging results (Dehdashti et al., 2003; Rajendran et al., 2006). Eventually, hypoxia guided radiotherapy with radiation dose distributions shaped to the hypoxic image, in such a way that the most hypoxic regions of a tumour receive the highest radiation dose, could be a strategy to obtain better tumour control.
BOLD‐MRI is an experimental technique for detecting hypoxia with good spatial and temporal resolution (Rijpkema et al., 2002). It is based on the paramagnetic properties of deoxyhaemoglobin, offering a way to indirectly visualise the pO2 in blood vessels and surrounding tissue. An added advantage is that the administration of contrast material can be omitted. Unfortunately, it does not provide quantitative information about the oxygen concentration, a major drawback of all these techniques. Further investigation is necessary to validate this method and establish its clinical importance.
3.2. Polarographic needle electrode
Being regarded as the gold standard for a long time, the polarographic oxygen electrode has been extensively used for quantifying hypoxia in both animal studies and human tumours (Brizel et al., 1997; Gatenby et al., 1988). With this invasive technique oxygen concentrations can be measured directly at many different positions in the tumour and within a short time frame. Although the Eppendorf electrode, introduced in the late 1980s, is a vast improvement compared to the older systems, the method still has some major drawbacks. It is limited to accessible tumour sites like head and neck and cervical cancer, disrupts the tissue and it has a large inter‐observer variability (Nozue et al., 1997). Another restriction of this technique is the failure to distinguish necrotic areas from viable tumour tissue and to discern the patterns of hypoxia.
3.3. Exogenous markers
The limitations of the needle electrode led to the development of exogenous markers, the 2‐nitroimidazoles, to measure hypoxia. Two markers are approved for clinical use: pimonidazole, (1‐[(2‐hydroxy‐3‐piperidinyl)propyl]‐2‐nitroimidazole hydrochloride, and EF5, [2‐(2‐nitro‐1H‐imidazole‐1‐yl)‐N‐(2,2,3,3,3‐pentafluoropropyl)acetamide]. These markers can be administered intravenously and are reduced and bound to thiol‐containing proteins in viable hypoxic cells. Pimonidazole and EF5 have been proven to be reliable hypoxic markers, with a good correlation with the radiobiologically hypoxic fraction (Lee et al., 1996; Raleigh et al., 1999) and pO2 (Raleigh et al., 1999).
Several studies have shown the prognostic value of the 2‐nitroimidazoles in a clinical setting. In head and neck cancer a correlation was found between the degree of hypoxia estimated by 2‐nitroimidazole binding and the locoregional control and event‐free‐survival (Evans et al., 2007; Kaanders et al., 2002c). One study performed in patients with cervical carcinoma did not show this association (Nordsmark et al., 2006) In patients with head and neck cancer treated with ARCON (accelerated radiotherapy with carbogen and nicotinamide), a hypoxia‐modifying therapy, pimonidazole demonstrated a predictive value as well (Kaanders et al., 2002c).
An oral prescription of pimonidazole has become available, which remains to be validated (Bennewith et al., 2002). Recently, it has been FDA approved. For clinical use it would be a progress, making the administration of pimonidazole even more convenient.
3.4. Endogenous hypoxia‐related markers
An alternative method not relying on injected markers to quantify hypoxia is the use of endogenous markers, an extensive range of potential markers having been studied over the last years (Bussink et al., 2003). We prefer the term hypoxia‐related marker to hypoxic marker, as the association between these markers and the oxygen status of the tissue is weak and has only been reported in a few cases. Endogenous markers are proteins upregulated in association with hypoxia and can be measured in blood plasma or immunohistochemically on tumour biopsies. No single marker has consistently demonstrated strong prognostic power in clinical practice as of yet, although a correlation with patient outcome has been found with CAIX, osteopontin and PAI‐1. Attempts have been made to combine various markers to create a hypoxia‐prognostic profile, with moderate success (Koukourakis et al., 2006; Le et al., 2007). The most important markers will be highlighted, with emphasis on their role in the hypoxic molecular response and their clinical significance.
3.4.1. HIF‐1
Hypoxia initiates a complicated response involving a plethora of different molecular pathways. These pathways modulate several cellular functions, like proliferation, apoptosis, angiogenesis, pH balance and anaerobic glycolysis. The HIF‐1 pathway with its numerous downstream targets is the key controller in this reaction (Figure 3). The transcription factor HIF‐1 is a heterodimer consisting of two subunits: HIF‐1α and HIF‐1β. HIF‐1β is constitutively active, while HIF‐1α is rapidly degraded under normoxic conditions and stabilised by hypoxia (Wang et al., 1995). Upon activation HIF‐1 binds to the hypoxia responsive element (HRE), thereby promoting the transcription of numerous genes including VEGF and the genes encoding for the glucose transporters. Several cofactors are involved in the transcriptional regulation of the various target genes.
Figure 3.
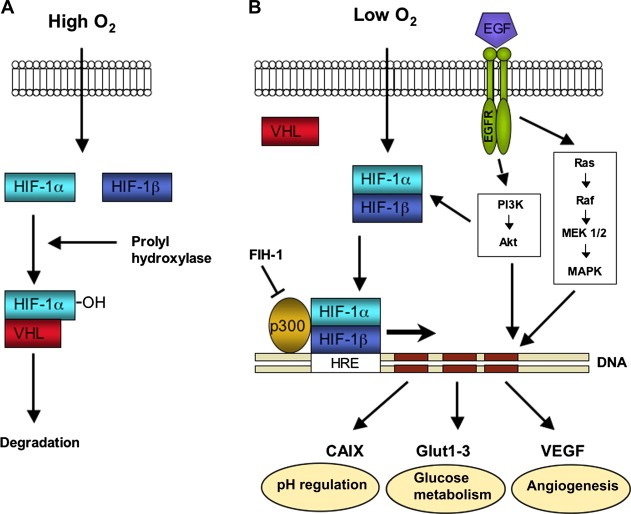
Schematic representation of the HIF‐1 pathway. Under normoxic conditions HIF‐1α is hydroxylated and rapidly degraded (A). Under hypoxic conditions the HIF‐1 complex is stabilised and initiates the transcription of its target genes. EGFR can activate HIF‐1α in an oxygen‐independent way (B).
Under normoxic conditions two inhibitory pathways of HIF‐1α are essential. Prolyl hydroxylases (PHDs) hydroxylate proline sites in the oxygen‐dependent degradation domain (ODD) of the HIF‐1α protein. This enables binding of the von Hippel Lindau protein (VHL), which leads to the proteasomal degradation of HIF‐1α. Under hypoxic conditions hydroxylation does not occur, leading to accumulation of HIF‐1. The second main inhibitory pathway of HIF‐1α is factor inhibiting HIF‐1 (FIH‐1). FIH‐1 hydroxylates the C‐terminal transactivation domain (CAD) of HIF‐1α, which inhibits binding of p300/CBP to the HIF‐1 complex, which is a cofactor necessary for transcription. Besides the oxygen‐dependent activation of HIF‐1α, certain receptors of the tyrosine kinase family, like insulin‐like growth factor receptor (IGFR), epidermal growth factor receptor (EGFR) and HER2/neu, can activate HIF‐1α in an oxygen‐independent way. They regulate the transcriptional activity through the P13K/Akt/mTOR pathway (Semenza, 2000).
HIF‐1α has gained interest as an endogenous hypoxia‐related marker, after the discovery of its overexpression in a wide variety of malignant tumours (Zhong et al., 1999). Its significance as a prognostic factor for aggressive tumour behaviour has been proven in various types of cancer; clear cell carcinoma, ovarian carcinoma, gastric carcinoma, breast cancer, soft‐tissue sarcoma, bladder cancer, head and neck cancer, rectal, lung cancer and cervical carcinoma (Moon et al., 2007), although some studies show an opposite effect (Beasley et al., 2002; Fillies et al., 2005). Moreover, the correlation between HIF‐1α expression and oxygen electrode or pimonidazole measurements is weak (Mayer et al., 2004). Therefore, the value of HIF‐1α quantification as a hypoxia‐related assay remains questionable.
Given the widespread overexpression of HIF‐1 and its influence on multiple cellular functions, it is a promising therapeutic target, although the heterogeneity of the gene response also makes it a complex target. Drugs aimed at the HIF‐1 pathway can intervene in multiple ways; decreasing HIF‐1α mRNA or protein levels, inhibiting DNA binding of HIF‐1 or decreasing HIF‐1‐mediated transactivation (Semenza, 2007). So far, clinical studies show limited success of these approaches.
3.4.2. Carbonic anhydrase IX
One of the downstream targets of HIF‐1 is carbonic anhydrase 9 (CA9). The upregulation of CA9 under hypoxic conditions is controlled by the HIF‐1 binding at the hypoxia responsive element (HRE) in its promoter region (Figure 3). This is supported by the fact that in VHL‐defective renal cell carcinoma, in which the inhibition of HIF‐1 is lost, extensive overexpression of CAIX can be found (Grabmaier et al., 2004). CAIX is one of the 14 members of the CA‐family, existing of cytosolic, membrane‐associated, mitochondrial and secreted carbonic anhydrases (Potter and Harris, 2003). CAIX (or MN, or G250) is a membrane‐associated enzyme with a zinc‐containing extracellular catalytic domain. This highly active domain catalyses the reversible hydration of carbon dioxide to carbonic acid: H2O+CO2↔H++HCO3 −. It is involved in the respiratory gas exchange and acid–base balance, maintaining the intracellular and lowering the extracellular pH. CAIX can form a close interaction with certain ion‐transport systems, the so‐called metabolons. The association with the bicarbonate transporter is recently established, affirming the role of CAIX in ion‐transport and electrolyte‐secretion (Morgan et al., 2007).
CAIX is only limitedly present in normal tissue; it is found in gastric mucosa, small intestine and muscle. The overexpression of CAIX is demonstrated in different types of cancer (Ivanov et al., 2001). The CAIX‐positive cells are mainly present in the perinecrotic areas of a tumour, in contrast with tumours with inactivated VHL, where a more general staining pattern is observed (Wykoff et al., 2000). The expression shows overlap with the staining pattern of pimonidazole, but a strong correlation is not present (Kaanders et al., 2002c; Troost et al., 2005). There is no correlation between the amount of CAIX and direct oxygen measurement with the needle electrode in cervical carcinoma (Mayer et al., 2005a). Notwithstanding, CAIX has proven to be a prognostic marker in various tumours, as high CAIX expression is associated with worse locoregional control and overall survival (Potter and Harris, 2003). Especially, the combination of CAIX and a proliferation marker such as IdUrd or Ki67, showed encouraging results. These markers identify cells that are proliferating under hypoxic conditions, perhaps the most crucial subpopulation of tumour cells (Hoogsteen et al., 2005; Kim et al., 2007). In breast cancer, a quantifying assay of CAIX could identify patients who would respond least to adjuvant treatment, demonstrating its predictive value as well (Span et al., 2003).
Blocking the function of CAIX, resulting in an extracellular rise in pH, can be an interesting approach in cancer treatment. An acid tumour microenvironment can add to increased chemoresistance, decreasing the uptake and consequently the cytotoxicity of chemotherapeutic agents and enhances metastatic potential (Tredan et al., 2007). Specific inhibitors of CAIX have been developed, of which the sulphonamides are most promising. Indisulam, a potent inhibitor of CAIX is under current clinical investigation (Talbot et al., 2007). Furthermore, a few monoclonal antibodies to block CAIX are under investigation in renal cell carcinoma (Davis et al., 2007; Stillebroer et al., 2007), a perfect candidate for these studies, considering the high CAIX‐overexpression found in this tumour type.
3.4.3. Glucose transporters
Another group of genes that is upregulated in hypoxic conditions is that of genes encoding for the glucose transporters (GLUTs). These transmembrane glycoproteins are omnipresent in normal tissue, facilitating glucose transport across the cell membrane. Malignant tumours generally have a higher rate of metabolism, are more dependent on glycolysis as an energy source (Warburg effect) and therefore have a higher glucose need. Under hypoxic conditions the cell's demand for glucose increases as the anaerobic glycolysis becomes even more important. This also involves recruitment and overexpression of the glucose transporters in many malignant (hypoxic) tumours. The two glucose transporters most associated with invasive cancer are GLUT‐1 and GLUT‐3, being overexpressed in cervical carcinoma, head and neck cancer, colorectal and bladder cancer (Macheda et al., 2005). A third participant has been discovered more recently in breast and prostate cancer: GLUT‐12 (Rogers et al., 2002).
The comparison of immunohistochemical staining on biopsies of cervical carcinoma of GLUT‐1 and pimonidazole showed similar staining patterns (Airley et al., 2003), but a strong correlation with pO2 measurements by Eppendorf histography has never been found. Several studies have assessed the applicability of the glucose transporters as prognostic markers. In cervical carcinoma high GLUT‐1 expression has been associated with lower metastasis‐free‐survival (Airley et al., 2001), although in another study this correlation was not overtly present (Mayer et al., 2005b). Additionally, in invasive bladder cancer, high GLUT‐1 expression has been shown to be associated with poor survival (Palit et al., 2005). GLUT‐1 and GLUT‐3 have been proposed as prognostic factors in head and neck cancer, but with equivocal conclusions (Baer et al., 2002; Jonathan et al., 2006; Oliver et al., 2004). Summarizing, GLUT expression has some prognostic potential in solid tumours, but it is not a very robust hypoxia‐related marker.
3.4.4. VEGF
VEGF is one of the genes upregulated by HIF‐1 in a hypoxic microenvironment. It can also be activated in an oxygen‐independent way through the PI3K‐AKT pathway, for example by EGFR or loss of PTEN (Bussink et al., 2008; Mizukami et al., 2007; Pore et al., 2004). VEGF plays a crucial role in angiogenesis, physiologically and also pathologically in malignant tumour growth (review by Ferrara et al., 2003). VEGF activates signalling pathways like the Ras‐Raf‐MAPK and the PI3K‐AKT pathway and increases urokinase‐type plasminogen activator (uPA), tissue‐type plasminogen activator (tPA) and plasminogen activator inhibitor‐1 (PAI‐1) expression (see Section 3.4.5), thereby influencing tumour progression and metastatic spread in multiple ways. Of the two tyrosine kinase receptors for VEGF, VEGFR‐1 (Flt‐1) and VEGFR‐2 (Flk‐1), the latter seems the most important in enhancing mitosis of endothelial cells and angiogenesis (Ferrara et al., 2003).
The expression of VEGF in different tumour types and its significance for prognosis has been investigated in numerous studies (as reviewed by Moon et al., 2007). Almost all of them showed a correlation of VEGF with poor prognosis. Several VEGF‐ and VEGFR‐inhibiting drugs have been developed and tested in vitro and in vivo (Petersen, 2007). For instance SU5416, a selective Flk‐1 inhibitor (Fong et al., 1999) exhibited a substantial anti‐tumour activity when combined with irradiation (Schuuring et al., 2005). Bevacizumab is a monoclonal antibody against VEGF, approved for use in stage IIIB/IV non‐small cell lung cancer, which in a phase III ECOG trial showed improved progression‐free survival when combined with chemotherapy (Sandler et al., 2006). As the absolute benefit of these monoclonal antibodies is small, the main focus is now on developing multitargeted receptor tyrosine kinase inhibitors (RTKI), like vandetanib, which inhibits EGFR and RET besides VEGFR (Hanrahan and Heymach, 2007). These compounds are currently under investigation in clinical phase III trials.
3.4.5. uPA/uPAR system
Another important signal route that can add to the aggressiveness of the tumour is the urokinase plasminogen activator (uPA) system (Dass et al., 2007). It includes uPA with its membrane‐bound receptor (uPAR), tPA and the inhibitors (PAIs). UPA and tPA are serine proteases that catalyse the activation of plasminogen in plasmin. Plasmin in turn can activate many other enzymes, among which the matrix metalloproteases. Eventually, this leads to the degradation of the basement membrane and extracellular matrix, enhancing invasion and metastatic spread. Even though uPA can exhibit some functions without binding to uPAR, it is most active when associated with its receptor.
PAI‐1 and PAI‐2 are two inhibitors of uPA and tPA. PAI‐I binds to the uPA/uPAR complex, leading to internalization of the whole complex and degradation of uPA and PAI‐1 in the lysosomes, while uPAR is transported back to the cell membrane (Figure 4).
Figure 4.
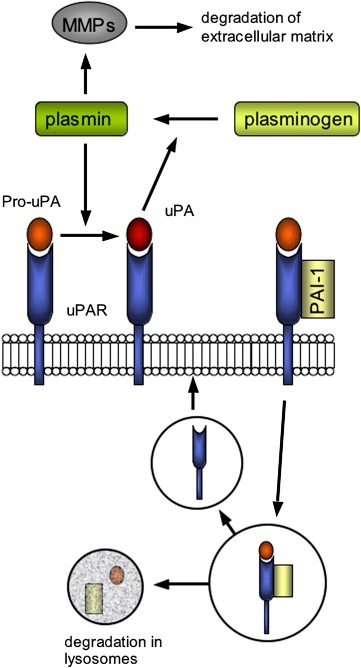
The uPA/uPAR system; its activation, the effects and the inhibitory action of PAI‐1.
uPA, uPAR and PAI‐1 are all reported to be overexpressed in malignant tumours (Dass et al., 2007). In addition to the inhibiting effect on uPA, PAI‐1 manifests biological functions with an inverse effect, like inhibiting apoptosis and stimulating angiogenesis (Bajou et al., 2001; Kwaan et al., 2000). Another interesting fact is the presence of a hypoxia responsive element in the promoter region of the PAI‐1 gene, suggesting the possibility of upregulation of PAI‐1 under hypoxic conditions. This has been confirmed to occur in hepatoma and head and neck cell lines (Fink et al., 2002; Schilling et al., 2007). Both uPA and PAI‐1 are associated with a higher relapse rate and a poor survival in various types of cancer. In breast cancer, a large meta‐analysis showed a poor prognosis for uPA and PAI‐1 positive tumours (Look et al., 2002). The prognostic value of uPA and PAI‐1 in breast cancer is now being assessed in a multicentre prospective trial, the node‐negative breast cancer (NNBC)‐3 Europe trial, where participating centres can perform risk estimation for breast cancer patients by the uPA and PAI‐1 concentration in tumour tissue biopsies. uPA/PAI‐1 is the first novel protein biomarker combination recommended for clinical routine use in breast cancer by the current 2007 ASCO guidelines (Harris et al., 2007).
3.4.6. Osteopontin
OPN is an integrin‐binding protein first identified in non‐collagenous bone matrix and is involved in several physiological processes, such as cytokine production, cell adhesion and cell migration. In cancer, OPN seems to add to the aggressiveness and metastatic potential of tumour cells (Rittling and Chambers, 2004). An important factor in this malignant progression is the activation of uPA and matrix metalloproteases by OPN through the PI3K/Akt, the IKKα/β and the MAPK pathways. Two domains of OPN, the integrin‐ and CD44‐binding domain, are involved in this process. In the OPN glycoprotein these two domains are separated by a protease‐sensitive site and become functionally active upon cleavage of this site by thrombin as two distinct proteins. Another effect of the binding of OPN to the integrin receptor is the c‐Src kinase mediated activation of the EGFR, a tyrosine kinase receptor belonging to the erbB growth factor receptor family, that plays a crucial role in oncogenesis as well (Rangaswami et al., 2006).
OPN gene expression is inversely correlated with that of VHL, indicating the role of OPN as a hypoxia‐related protein. This was confirmed by the correlation between high plasma OPN levels and low tumour pO2 in patients with head and neck cancer (Le et al., 2003).
Overexpression of OPN is present in many different types of tumours (Brown et al., 1994; Coppola et al., 2004). Several studies have been performed to evaluate the prognostic significance of OPN applying different methods: measuring the plasma concentration, the mRNA level or using immunohistochemistry on tissue sections. In lung and breast cancer immunohistochemically quantified OPN has been correlated with poor prognosis (Chambers et al., 1996; Rudland et al., 2002). Plasma osteopontin has been associated with poor outcome in head and neck cancer, hepatocellular carcinoma and recently renal cell carcinoma (Overgaard et al., 2005; Ramankulov et al., 2007; Zhang et al., 2006). In the DAHANCA 5 trial, a randomised trial comparing radiotherapy alone with radiotherapy combined with the hypoxia sensitiser nimorazole, it showed predictive value as well. Only patients with a high plasma concentration of OPN had advantage of the hypoxia‐modifying therapy (Overgaard et al., 2005). As OPN is not only involved in malignant disease, but also in vascular and inflammatory disorders, its use as a hypoxia‐related marker and potential target for treatment remains questionable.
3.5. Unfolded protein response
Cellular adaptation to hypoxic stress involves upregulation of hypoxia responsive genes, along with a down regulation of oxygen‐ and energy‐consuming processes, such as cell proliferation and protein synthesis. The unfolding protein response plays an important role in converting the cell into a low energy state (recent review by Koumenis and Wouters, 2006).
In the cell mRNA translation, post‐translational modification and protein folding takes place in the endoplasmatic reticulum (ER). In times of ER stress, like hypoxia, accumulation of incorrectly folded proteins occurs. This initiates the unfolded protein response, which leads to down regulation of protein synthesis, increased degradation of unfolded proteins and, during prolonged ER stress, activation of proapoptotic genes. The reduction in protein synthesis can be attributed predominantly to a decrease in mRNA translation. The main mediator in this response is protein kinase activated by dsRNA (PKR)‐like endoplasmatic reticulum kinase (PERK), which phosphorylates and thereby inactivates eukaryotic initiating factor 2α (eIF2α), an important translation‐initiating factor (Figure 5). PERK is an ER‐resident kinase that is normally inactivated by glucose‐regulated protein 78 (GRP78), a chaperone protein that dissociates from PERK under ER stress.
Figure 5.
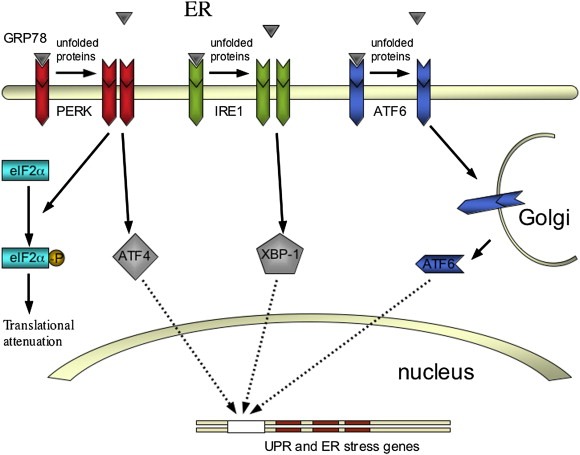
The three main mediators in the unfolded protein response (UPR). PERK, IRE1 and ATF6 effectuate a decrease in protein synthesis and an upregulation of UPR and ER stress genes.
The ER transmembrane proteins IRE1 and activating transcription factor‐6 (ATF6) regulate the expression of the UPR and ER stress genes, also the second function of PERK. IRE1 activates the transcription factor XBP‐1, which is transported subsequently to the nucleus to upregulate the UPR and ER stress genes. ATF6 exhibits the same function as a transcription factor upon activation by proteolysis in the Golgi. GRP78 is associated with IRE1 and ATF6 as well, inactivating them in the absence of ER stress.
In xenografts with reduced GRP78, an suppression of tumour growth, an increase of apoptosis and an inhibition of invasion is observed (Lee, 2007). Furthermore, knockdown of GRP78 enhances sensitivity of tumour cells to chemotherapeutic drugs by increasing apoptosis. In many different types of cancer GRP78 is a negative prognostic factor, with the exception of neuroblastoma and lung cancer, in which it is associated with a favourable outcome (Lee, 2007). GRP78 might be used in the future as a predictive marker in breast cancer to select patients for adjuvant chemotherapy.
Inactivation of PERK or eIF2α in vitro and in xenografts also results in an increased susceptibility to hypoxia and low cell survival (Blais and Bell, 2006). PERK or IRE1 inhibitors could be combined with other cytotoxic therapies as a novel approach in anti‐cancer treatment.
4. Targeting hypoxia
Given the complexity of the hypoxic response, various strategies have been devised to target hypoxic cells. The most straightforward strategy is increasing the oxygen availability. Furthermore, some therapies have been developed using hypoxic cell specific toxins, others are aimed at decreasing their resistance to radiotherapy. A new field of research involves the development of drugs that target proteins involved in the hypoxic response or angiogenesis.
4.1. Increasing oxygen availability
One of the first attempts to overcome hypoxia involved delivery of radiotherapy in hyperbaric oxygen chambers. In head and neck and cervical cancer patients treated with hyperbaric oxygen showed improved local tumour control and survival (Dische, 1978). This treatment was eventually abandoned as hypoxia‐modifying therapy due to the complex technique and poor patient compliance.
The next step was to combine anti‐cancer treatment such as radiotherapy with normobaric oxygen or carbogen (95% oxygen+5% carbon dioxide) breathing. ARCON (accelerated radiotherapy with carbogen and nicotinamide) combines radiotherapy with carbogen breathing and nicotinamide, a vasoactive agent, counteracting both diffusion‐limited and perfusion‐limited hypoxia (Kaanders et al., 2002a). In a phase II trial of head and neck cancer it demonstrated a substantial therapeutic effect in laryngeal carcinoma, notably in the more advanced tumour stages. The 3‐year local control rate was 84% for T4 tumours, compared to around 50% with radiotherapy alone (Kaanders et al., 2002b). Recently, two large phase III trials on laryngeal cancer and bladder cancer were completed. These trials will elucidate the potential benefit of ARCON as hypoxia‐modifying regimen.
4.2. Hypoxic cytotoxins
Bioreductive drugs, like tirapazamine (TPZ), exhibit a direct cytotoxic effect on hypoxic cells. In the absence of oxygen, reduction of TPZ takes place, forming a highly reactive radical, which induces DNA damage. TPZ has proven its therapeutic value in combination with cisplatin in a phase III randomised trial in stages IIIB and IV non‐small‐cell lung cancer, increasing the 1‐year survival from 23% to 34% (von Pawel et al., 2000). Conversely, the combination of TPZ with paclitaxel and carboplatin showed an increase in toxicity without any survival advantage in advanced stage lung cancer (Williamson et al., 2005). In advanced head and neck cancer, a phase II trial comparing TPZ, cisplatin and radiotherapy with 5‐fluorouracil, cisplatin and radiotherapy showed increased failure‐free survival rates with acceptable toxicity (Rischin et al., 2005). The results from phase III trials are forthcoming; hopefully these will clarify the clinical benefits of tirapazamine.
4.3. Radiosensitisers
Nitroimidazoles are chemical compounds that mimic the radiosensitising effect of oxygen by inducing free‐radical mediated double strand DNA breaks. The use of the first generation drug misonidazole, the most thoroughly documented of the nitroimidazoles, is obsolete in clinical practice as a radiosensitiser, due to its severe side effects. However, it still has the attention of some researchers as a PET‐imaging compound at substantially lower doses. Of the other nitroimidazoles, nimorazole has proven its value as a hypoxic cell sensitising agent. In the Danish Head and Neck Cancer 5 study, in which 422 patients with head and neck cancer were randomised, the locoregional control rate was significantly higher (49% versus 33%) when radiotherapy was combined with nimorazole compared to radiotherapy alone (Overgaard et al., 1998). Nimorazole has not been adopted widespread as a standard treatment despite these promising results. The many negative studies with the older nitroimidazoles have contributed to this circumstance and a second randomised trial has never been performed.
4.4. Vascular targeting
The vascular network of a tumour, providing oxygen and nutrients, for long time has been an intriguing target in cancer treatment. Two main ways of vascular targeting can be distinguished: interfering in the angiogenesis, thereby preventing the formation of new blood vessels, and destroying the existent vasculature. The angiogenesis‐inhibitors have already been discussed in the paragraph above on VEGF. For the second approach, a broad range of vascular drugs is under development, the so‐called vascular disrupting agents (VDAs). These compounds are aimed at the endothelium, causing blood vessel occlusion resulting in extensive tumour necrosis. The two main groups of VDAs are the ligand‐based agents and the small molecules. The ligand‐based agents are toxins or pro‐coagulants coupled to endothelium‐directed antibodies, peptides or growth factors. Most commonly used are the small molecules, including the microtubulin destabilising drugs, disrupting the cytoskeleton of the endothelial cells, and the flavonoids, which cause massive cytokine production. Several phase I studies have proven their applicability in clinical practice (Patterson and Rustin, 2007). These VDAs are not sufficiently potent as a single agent therapy and therefore need to be combined with chemo‐ or radiotherapy. An important issue is the effect of the anti‐vascular agents on tumour oxygenation, if it is combined with chemo‐ and radiotherapy. On one side an increase in hypoxia is expected as the vascularization is diminished. On the other side VDAs typically target the tumour region with the most abnormal vasculature, usually comprising the most hypoxic and hence radioresistant cells. Furthermore, there is evidence that VDAs can cause G2/M cell cycle arrest with associated enhanced radiosensitivity. In clinical practice the VDAs have a small additive effect in combination with radiotherapy. CA4P (combretastatin‐A4‐phoshate) and DMXAA (5,6‐dimethylxanthenone‐4‐acetic acid) are two small molecules that are currently under investigation in phase II trials, combined with cytotoxic therapy. Several other compounds are tested in phase I studies (review by Horsman and Siemann, 2006).
5. Conclusion
With the abundant research performed on the molecular aspects of tumour hypoxia, several important pathways have been elucidated over the last years. The hypoxic response is complex with many genes and proteins involved that can be important prognostic factors and interesting therapeutic targets. As of yet, no single marker has shown strong predictive value to allow the selection of patients for hypoxia‐modifying treatment. Potential candidates, like pimonidazole and osteopontin, could be the subjects of further research. Furthermore, no single hypoxia‐targeted drug has exhibited substantial clinical effect as a single agent. Merging the different approaches seems to be of pivotal importance, both in creating a predictive profile and developing a combined treatment.
Acknowledgments
The Dutch Cancer Society, grant numbers 2008‐4000 and 2008‐4088, provided financial support. We thank J.P.W. Peters, J. Lok and W.J.M. Peeters for technical support. No conflict of interests.
Rademakers Saskia E., Span Paul N., Kaanders Johannes H.A.M., Sweep Fred C.G.J., van der Kogel Albert J., Bussink Johan, (2008), Molecular aspects of tumour hypoxia, Molecular Oncology, 2, doi: 10.1016/j.molonc.2008.03.006.
References
- Airley, R. , Loncaster, J. , Davidson, S. , Bromley, M. , Roberts, S. , Patterson, A. , Hunter, R. , Stratford, I. , West, C. , 2001. Glucose transporter glut-1 expression correlates with tumor hypoxia and predicts metastasis-free survival in advanced carcinoma of the cervix. Clin. Cancer Res. 7, 928–934. [PubMed] [Google Scholar]
- Airley, R.E. , Loncaster, J. , Raleigh, J.A. , Harris, A.L. , Davidson, S.E. , Hunter, R.D. , West, C.M. , Stratford, I.J. , 2003. GLUT-1 and CAIX as intrinsic markers of hypoxia in carcinoma of the cervix: relationship to pimonidazole binding. Int. J. Cancer 104, 85–91. [DOI] [PubMed] [Google Scholar]
- Baer, S. , Casaubon, L. , Schwartz, M.R. , Marcogliese, A. , Younes, M. , 2002. Glut3 expression in biopsy specimens of laryngeal carcinoma is associated with poor survival. The Laryngoscope 112, 393–396. [DOI] [PubMed] [Google Scholar]
- Bajou, K. , Masson, V. , Gerard, R.D. , Schmitt, P.M. , Albert, V. , Praus, M. , Lund, L.R. , Frandsen, T.L. , Brunner, N. , Dano, K. , 2001. The plasminogen activator inhibitor PAI-1 controls in vivo tumor vascularization by interaction with proteases, not vitronectin. Implications for antiangiogenic strategies. J. Cell Biol. 152, 777–784. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beasley, N.J. , Leek, R. , Alam, M. , Turley, H. , Cox, G.J. , Gatter, K. , Millard, P. , Fuggle, S. , Harris, A.L. , 2002. Hypoxia-inducible factors HIF-1alpha and HIF-2alpha in head and neck cancer: relationship to tumor biology and treatment outcome in surgically resected patients. Cancer Res. 62, 2493–2497. [PubMed] [Google Scholar]
- Bennewith, K.L. , Durand, R.E. , 2004. Quantifying transient hypoxia in human tumor xenografts by flow cytometry. Cancer Res. 64, 6183–6189. [DOI] [PubMed] [Google Scholar]
- Bennewith, K.L. , Raleigh, J.A. , Durand, R.E. , 2002. Orally administered pimonidazole to label hypoxic tumor cells. Cancer Res. 62, 6827–6830. [PubMed] [Google Scholar]
- Blais, J. , Bell, J.C. , 2006. Novel therapeutic target: the PERKs of inhibiting the integrated stress response. Cell Cycle 5, 2874–2877. [DOI] [PubMed] [Google Scholar]
- Brizel, D.M. , Scully, S.P. , Harrelson, J.M. , Layfield, L.J. , Bean, J.M. , Prosnitz, L.R. , Dewhirst, M.W. , 1996. Tumor oxygenation predicts for the likelihood of distant metastases in human soft tissue sarcoma. Cancer Res. 56, 941–943. [PubMed] [Google Scholar]
- Brizel, D.M. , Sibley, G.S. , Prosnitz, L.R. , Scher, R.L. , Dewhirst, M.W. , 1997. Tumor hypoxia adversely affects the prognosis of carcinoma of the head and neck. Int. J. Radiat. Oncol. Biol. Phys. 38, 285–289. [DOI] [PubMed] [Google Scholar]
- Brown, L.F. , Papadopoulos-Sergiou, A. , Berse, B. , Manseau, E.J. , Tognazzi, K. , Perruzzi, C.A. , Dvorak, H.F. , Senger, D.R. , 1994. Osteopontin expression and distribution in human carcinomas. Am. J. Pathol. 145, 610–623. [PMC free article] [PubMed] [Google Scholar]
- Bussink, J. , Kaanders, J.H. , van-der-Kogel, A.J. , 2003. Tumor hypoxia at the micro-regional level: clinical relevance and predictive value of exogenous and endogenous hypoxic cell markers. Radiother. Oncol. 67, 3–15. [DOI] [PubMed] [Google Scholar]
- Bussink, J. , van der Kogel, A.J. , Kaanders, J.H. , 2008. Acivation of the PI3-K/AKT pathway and implications for radioresistance mechanisms in head and neck cancer. Lancet Oncol. 9, 288–296. [DOI] [PubMed] [Google Scholar]
- Chambers, A.F. , Wilson, S.M. , Kerkvliet, N. , O'Malley, F.P. , Harris, J.F. , Casson, A.G. , 1996. Osteopontin expression in lung cancer. Lung Cancer (Amsterdam, Netherlands) 15, 311–323. [DOI] [PubMed] [Google Scholar]
- Coppola, D. , Szabo, M. , Boulware, D. , Muraca, P. , Alsarraj, M. , Chambers, A.F. , Yeatman, T.J. , 2004. Correlation of osteopontin protein expression and pathological stage across a wide variety of tumor histologies. Clin. Cancer Res. 10, 184–190. [DOI] [PubMed] [Google Scholar]
- Dass, K. , Ahmad, A. , Azmi, A. , Sarkar, S. , Sarkar, F. , 2007. Evolving role of uPA/uPAR system in human cancers. Cancer Treat. Rev. 10.1016/j.ctrv.2007.1010.1005 [DOI] [PubMed] [Google Scholar]
- Davis, I.D. , Wiseman, G.A. , Lee, F.T. , Gansen, D.N. , Hopkins, W. , Papenfuss, A.T. , Liu, Z. , Moynihan, T.J. , Croghan, G.A. , Adjei, A.A. , 2007. A phase I multiple dose, dose escalation study of cG250 monoclonal antibody in patients with advanced renal cell carcinoma. Cancer Immun. 7, 13 [PMC free article] [PubMed] [Google Scholar]
- Dehdashti, F. , Mintun, M.A. , Lewis, J.S. , Bradley, J. , Govindan, R. , Laforest, R. , Welch, M.J. , Siegel, B.A. , 2003. In vivo assessment of tumor hypoxia in lung cancer with 60Cu-ATSM. Eur. J. Nucl. Med. Mol. Imaging 30, 844–850. [DOI] [PubMed] [Google Scholar]
- Dewhirst, M.W. , 1998. Concepts of oxygen transport at the microcirculatory level. Semin. Radiat. Oncol. 8, 143–150. [DOI] [PubMed] [Google Scholar]
- Dische, S. , 1978. Hyperbaric oxygen: the Medical Research Council trials and their clinical significance. Br. J. Radiol. 51, 888–894. [DOI] [PubMed] [Google Scholar]
- Evans, S.M. , Du, K.L. , Chalian, A.A. , Mick, R. , Zhang, P.J. , Hahn, S.M. , Quon, H. , Lustig, R. , Weinstein, G.S. , Koch, C.J. , 2007. Patterns and levels of hypoxia in head and neck squamous cell carcinomas and their relationship to patient outcome. Int. J. Radiat. Oncol. Biol. Phys. 69, 1024–1031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ferrara, N. , Gerber, H.P. , LeCouter, J. , 2003. The biology of VEGF and its receptors. Nat. Med. 9, 669–676. [DOI] [PubMed] [Google Scholar]
- Fillies, T. , Werkmeister, R. , van-Diest, P.J. , Brandt, B. , Joos, U. , Buerger, H. , 2005. HIF1-alpha overexpression indicates a good prognosis in early stage squamous cell carcinomas of the oral floor. BMC Cancer 5, 84 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fink, T. , Kazlauskas, A. , Poellinger, L. , Ebbesen, P. , Zachar, V. , 2002. Identification of a tightly regulated hypoxia-response element in the promoter of human plasminogen activator inhibitor-1. Blood 99, 2077–2083. [DOI] [PubMed] [Google Scholar]
- Fong, T.A. , Shawver, L.K. , Sun, L. , Tang, C. , App, H. , Powell, T.J. , Kim, Y.H. , Schreck, R. , Wang, X. , Risau, W. , 1999. SU5416 is a potent and selective inhibitor of the vascular endothelial growth factor receptor (Flk-1/KDR) that inhibits tyrosine kinase catalysis, tumor vascularization, and growth of multiple tumor types. Cancer Res. 59, 99–106. [PubMed] [Google Scholar]
- Gatenby, R.A. , Kessler, H.B. , Rosenblum, J.S. , Coia, L.R. , Moldofsky, P.J. , Hartz, W.H. , Broder, G.J. , 1988. Oxygen distribution in squamous cell carcinoma metastases and its relationship to outcome of radiation therapy. Int. J. Radiat. Oncol. Biol. Phys. 14, 831–838. [DOI] [PubMed] [Google Scholar]
- Grabmaier, K. , A-de-Weijert, M.C. , Verhaegh, G.W. , Schalken, J.A. , Oosterwijk, E. , 2004. Strict regulation of CAIX(G250/MN) by HIF-1alpha in clear cell renal cell carcinoma. Oncogene 23, 5624–5631. [DOI] [PubMed] [Google Scholar]
- Grau, C. , Overgaard, J. , 1992. Effect of etoposide, carmustine, vincristine, 5-fluorouracil, or methotrexate on radiobiologically oxic and hypoxic cells in a C3H mouse mammary carcinoma in situ. Cancer Chemother. Pharm. 30, 277–280. [DOI] [PubMed] [Google Scholar]
- Gray, L. , Conger, A.D. , Ebert, M. , Hornsey, S. , Scott, O.C.A. , 1953. The concentration of oxygen dissolved in tissues at the time of irradiation as a factor in radiotherapy. Br. J. Radiol. 28, 638–648. [DOI] [PubMed] [Google Scholar]
- Hanrahan, E.O. , Heymach, J.V. , 2007. Vascular endothelial growth factor receptor tyrosine kinase inhibitors vandetanib (ZD6474) and AZD2171 in lung cancer. Clin. Cancer Res. 13, 4617–4622. [DOI] [PubMed] [Google Scholar]
- Harris, L. , Fritsche, H. , Mennel, R. , Norton, L. , Ravdin, P. , Taube, S. , Somerfield, M.R. , Hayes, D.F. , Bast, R.C. , 2007. American Society of Clinical Oncology 2007 update of recommendations for the use of tumor markers in breast cancer. J. Clin. Oncol. 25, 5287–5312. Official Journal of the American Society of Clinical Oncology [DOI] [PubMed] [Google Scholar]
- Hockel, M. , Schlenger, K. , Aral, B. , Mitze, M. , Schaffer, U. , Vaupel, P. , 1996. Association between tumor hypoxia and malignant progression in advanced cancer of the uterine cervix. Cancer Res. 56, 4509–4515. [PubMed] [Google Scholar]
- Hockel, M. , Schlenger, K. , Hockel, S. , Vaupel, P. , 1999. Hypoxic cervical cancers with low apoptotic index are highly aggressive. Cancer Res. 59, 4525–4528. [PubMed] [Google Scholar]
- Hockel, M. , Vaupel, P. , 2001. Tumor hypoxia: definitions and current clinical, biologic, and molecular aspects. J. Natl. Cancer Inst. 93, 266–276. [DOI] [PubMed] [Google Scholar]
- Hoogsteen, I.J. , Marres, H.A.M. , Wijffels, K.I.E.M. , Rijken, P.F.J.W. , Peters, J.P.W. , van den Hoogen, F.J. , Oosterwijk, E. , van der Kogel, A.J. , Kaanders, J.H.A.M. , 2005. Colocalization of carbonic anhydrase 9 expression and cell proliferation in human head and neck squamous cell carcinoma. Clin. Cancer Res. 11, 97–106. [PubMed] [Google Scholar]
- Horsman, M.R. , Siemann, D.W. , 2006. Pathophysiologic effects of vascular-targeting agents and the implications for combination with conventional therapies. Cancer Res. 66, 11,520–11,539. [DOI] [PubMed] [Google Scholar]
- Hu, M. , Polyak, K. , 2008. Microenvironmental regulation of cancer development. Curr. Opin. Genet. Dev. 10.1016/j.gde.2007.12.006 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ivanov, S. , Liao, S.Y. , Ivanova, A. , Danilkovitch-Miagkova, A. , Tarasova, N. , Weirich, G. , Merrill, M.J. , Proescholdt, M.A. , Oldfield, E.H. , Lee, J. , 2001. Expression of hypoxia-inducible cell-surface transmembrane carbonic anhydrases in human cancer. Am. J. Pathol. 158, 905–919. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Jonathan, R.A. , Wijffels, K.I. , Peeters, W. , de Wilde, P.C. , Marres, H.A. , Merkx, M.A. , Oosterwijk, E. , van der Kogel, A.J. , Kaanders, J.H. , 2006. The prognostic value of endogenous hypoxia-related markers for head and neck squamous cell carcinomas treated with ARCON. Radiother. Oncol. 79, 288–297. [DOI] [PubMed] [Google Scholar]
- Kaanders, J.H. , Bussink, J. , van der Kogel, A.J. , 2002. ARCON: a novel biology-based approach in radiotherapy. Lancet Oncol. 3, 728–737. [DOI] [PubMed] [Google Scholar]
- Kaanders, J.H. , Pop, L.A. , Marres, H.A. , Bruaset, I. , van den Hoogen, F.J. , Merkx, M.A. , van der Kogel, A.J. , 2002. ARCON: experience in 215 patients with advanced head-and-neck cancer. Int. J. Radiat. Oncol. Biol. Phys. 52, 769–778. [DOI] [PubMed] [Google Scholar]
- Kaanders, J.H. , Wijffels, K.I. , Marres, H.A. , Ljungkvist, A.S. , Pop, L.A. , van den Hoogen, F.J. , de Wilde, P.C. , Bussink, J. , Raleigh, J.A. , van der Kogel, A.J. , 2002. Pimonidazole binding and tumor vascularity predict for treatment outcome in head and neck cancer. Cancer Res. 62, 7066–7074. [PubMed] [Google Scholar]
- Kim, S.J. , Shin, H.J. , Jung, K.Y. , Baek, S.K. , Shin, B.K. , Choi, J. , Kim, B.S. , Shin, S.W. , Kim, Y.H. , Kim, J.S. , 2007. Prognostic value of carbonic anhydrase IX and Ki-67 expression in squamous cell carcinoma of the tongue. Jpn. J. Clin. Oncol. 37, 812–819. [DOI] [PubMed] [Google Scholar]
- Konerding, M.A. , Miodonski, A.J. , Lametschwandtner, A. , 1995. Microvascular corrosion casting in the study of tumor vascularity: a review. Scanning Microsc. 9, 1233–1244. [PubMed] [Google Scholar]
- Koukourakis, M.I. , Bentzen, S.M. , Giatromanolaki, A. , Wilson, G.D. , Daley, F.M. , Saunders, M.I. , Dische, S. , Sivridis, E. , Harris, A.L. , 2006. Endogenous markers of two separate hypoxia response pathways (hypoxia inducible factor 2 alpha and carbonic anhydrase 9) are associated with radiotherapy failure in head and neck cancer patients recruited in the CHART randomized trial. J. Clin. Oncol. 24, 727–735. [DOI] [PubMed] [Google Scholar]
- Koumenis, C. , Wouters, B.G. , 2006. “Translating” tumor hypoxia: unfolded protein response (UPR)-dependent and UPR-independent pathways. Mol. Cancer Res. 4, 423–436. [DOI] [PubMed] [Google Scholar]
- Kwaan, H.C. , Wang, J. , Svoboda, K. , Declerck, P.J. , 2000. Plasminogen activator inhibitor 1 may promote tumour growth through inhibition of apoptosis. Br. J. Cancer 82, 1702–1708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Le, Q.T. , Kong, C. , Lavori, P.W. , O'Byrne, K. , Erler, J.T. , Huang, X. , Chen, Y. , Cao, H. , Tibshirani, R. , Denko, N. , 2007. Expression and prognostic significance of a panel of tissue hypoxia markers in head-and-neck squamous cell carcinomas. Int. J. Radiat. Oncol. Biol. Phys. 69, 167–175. [DOI] [PubMed] [Google Scholar]
- Le, Q.T. , Sutphin, P.D. , Raychaudhuri, S. , Yu, S.C. , Terris, D.J. , Lin, H.S. , Lum, B. , Pinto, H.A. , Koong, A.C. , Giaccia, A.J. , 2003. Identification of osteopontin as a prognostic plasma marker for head and neck squamous cell carcinomas. Clin. Cancer Res. 9, 59–67. [PMC free article] [PubMed] [Google Scholar]
- Lee, A.S. , 2007. GRP78 induction in cancer: therapeutic and prognostic implications. Cancer Res. 67, 3496–3499. [DOI] [PubMed] [Google Scholar]
- Lee, J. , Siemann, D.W. , Koch, C.J. , Lord, E.M. , 1996. Direct relationship between radiobiological hypoxia in tumors and monoclonal antibody detection of EF5 cellular adducts. Int. J. Cancer 67, 372–378. [DOI] [PubMed] [Google Scholar]
- Look, M.P. , van-Putten, W.L. , Duffy, M.J. , Harbeck, N. , Christensen, I.J. , Thomssen, C. , Kates, R. , Spyratos, F. , Ferno, M. , Eppenberger-Castori, S. , 2002. Pooled analysis of prognostic impact of urokinase-type plasminogen activator and its inhibitor PAI-1 in 8377 breast cancer patients. J. Natl. Cancer Inst. 94, 116–128. [DOI] [PubMed] [Google Scholar]
- Macheda, M.L. , Rogers, S. , Best, J.D. , 2005. Molecular and cellular regulation of glucose transporter (GLUT) proteins in cancer. J. Cell Physiol. 202, 654–662. [DOI] [PubMed] [Google Scholar]
- Matthews, N.E. , Adams, M.A. , Maxwell, L.R. , Gofton, T.E. , Graham, C.H. , 2001. Nitric oxide-mediated regulation of chemosensitivity in cancer cells. J. Natl. Cancer Inst. 93, 1879–1885. [DOI] [PubMed] [Google Scholar]
- Mayer, A. , Hockel, M. , Vaupel, P. , 2005. Carbonic anhydrase IX expression and tumor oxygenation status do not correlate at the microregional level in locally advanced cancers of the uterine cervix. Clin. Cancer Res. 11, 7220–7225. [DOI] [PubMed] [Google Scholar]
- Mayer, A. , Hockel, M. , Wree, A. , Vaupel, P. , 2005. Microregional expression of glucose transporter-1 and oxygenation status: lack of correlation in locally advanced cervical cancers. Clin. Cancer Res. 11, 2768–2773. [DOI] [PubMed] [Google Scholar]
- Mayer, A. , Wree, A. , Hockel, M. , Leo, C. , Pilch, H. , Vaupel, P. , 2004. Lack of correlation between expression of HIF-1alpha protein and oxygenation status in identical tissue areas of squamous cell carcinomas of the uterine cervix. Cancer Res. 64, 5876–5881. [DOI] [PubMed] [Google Scholar]
- Mizukami, Y. , Kohgo, Y. , Chung, D.C. , 2007. Hypoxia inducible factor-1 independent pathways in tumor angiogenesis. Clin. Cancer Res. 13, 5670–5674. [DOI] [PubMed] [Google Scholar]
- Moon, E.J. , Brizel, D.M. , Chi, J.T. , Dewhirst, M.W. , 2007. The potential role of intrinsic hypoxia markers as prognostic variables in cancer. Antioxid. Redox. Signal. 9, 1237–1294. [DOI] [PubMed] [Google Scholar]
- Morgan, P.E. , Pastorekova, S. , Stuart-Tilley, A.K. , Alper, S.L. , Casey, J.R. , 2007. Interactions of transmembrane carbonic anhydrase, CAIX, with bicarbonate transporters. Am. J. Physiol. Cell Physiol. 293, 738–748. [DOI] [PubMed] [Google Scholar]
- Mottram, J.C. , 1936. A factor of importance in the radiosensitivity of tumours. Br. J. Radiol. IX, 606–614. [Google Scholar]
- Nordsmark, M. , Alsner, J. , Keller, J. , Nielsen, O.S. , Jensen, O.M. , Horsman, M.R. , Overgaard, J. , 2001. Hypoxia in human soft tissue sarcomas: adverse impact on survival and no association with p53 mutations. Br. J. Cancer 84, 1070–1075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nordsmark, M. , Bentzen, S.M. , Rudat, V. , Brizel, D. , Lartigau, E. , Stadler, P. , Becker, A. , Adam, M. , Molls, M. , Dunst, J. , 2005. Prognostic value of tumor oxygenation in 397 head and neck tumors after primary radiation therapy. An international multi-center study. Radiother. Oncol. 77, 18–24. [DOI] [PubMed] [Google Scholar]
- Nordsmark, M. , Loncaster, J. , Aquino-Parsons, C. , Chou, S.C. , Gebski, V. , West, C. , Lindegaard, J.C. , Havsteen, H. , Davidson, S.E. , Hunter, R. , 2006. The prognostic value of pimonidazole and tumour pO2 in human cervix carcinomas after radiation therapy: a prospective international multi-center study. Radiother. Oncol. 80, 123–131. [DOI] [PubMed] [Google Scholar]
- Nozue, M. , Lee, I. , Yuan, F. , Teicher, B.A. , Brizel, D.M. , Dewhirst, M.W. , Milross, C.G. , Milas, L. , Song, C.W. , Thomas, C.D. , 1997. Interlaboratory variation in oxygen tension measurement by Eppendorf “Histograph” and comparison with hypoxic marker. J. Surg. Oncol. 66, 30–38. [DOI] [PubMed] [Google Scholar]
- Oliver, R.J. , Woodwards, R.T. , Sloan, P. , Thakker, N.S. , Stratford, I.J. , Airley, R.E. , 2004. Prognostic value of facilitative glucose transporter Glut-1 in oral squamous cell carcinomas treated by surgical resection; results of EORTC Translational Research Fund studies. Eur. J. Cancer 40, 503–507. [DOI] [PubMed] [Google Scholar]
- Overgaard, J. , Eriksen, J.G. , Nordsmark, M. , Alsner, J. , Horsman, M.R. , 2005. Plasma osteopontin, hypoxia, and response to the hypoxia sensitiser nimorazole in radiotherapy of head and neck cancer: results from the DAHANCA 5 randomised double-blind placebo-controlled trial. Lancet Oncol. 6, 757–764. [DOI] [PubMed] [Google Scholar]
- Overgaard, J. , Hansen, H.S. , Overgaard, M. , Bastholt, L. , Berthelsen, A. , Specht, L. , Lindelov, B. , Jorgensen, K. , 1998. A randomized double-blind phase III study of nimorazole as a hypoxic radiosensitizer of primary radiotherapy in supraglottic larynx and pharynx carcinoma. Results of the Danish Head and Neck Cancer Study (DAHANCA) Protocol 5-85. Radiother. Oncol. 46, 135–146. [DOI] [PubMed] [Google Scholar]
- von Pawel, J. , von-Roemeling, R. , Gatzemeier, U. , Boyer, M. , Elisson, L.O. , Clark, P. , Talbot, D. , Rey, A. , Butler, T.W. , Hirsh, V. , 2000. Tirapazamine plus cisplatin versus cisplatin in advanced non-small-cell lung cancer: a report of the international CATAPULT I study group. Cisplatin and tirapazamine in subjects with advanced previously untreated non-small-cell lung tumors. J. Clin. Oncol. 18, 1351–1359. [DOI] [PubMed] [Google Scholar]
- Padhani, A.R. , Krohn, K.A. , Lewis, J.S. , Alber, M. , 2007. Imaging oxygenation of human tumours. Eur. Radiol. 17, 861–872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Palit, V. , Phillips, R.M. , Puri, R. , Shah, T. , Bibby, M.C. , 2005. Expression of HIF-1alpha and Glut-1 in human bladder cancer. Oncol. Rep. 14, 909–913. [DOI] [PubMed] [Google Scholar]
- Patterson, D.M. , Rustin, G.J. , 2007. Vascular damaging agents. Clin. Oncol. 19, 443–456. [DOI] [PubMed] [Google Scholar]
- Petersen, I. , 2007. Antiangiogenesis, anti-VEGF(R) and outlook. Recent Results Cancer Res. 176, 189–199. [DOI] [PubMed] [Google Scholar]
- Pore, N. , Liu, S. , Shu, H.K. , Li, B. , Haas-Kogan, D. , Stokoe, D. , Milanini-Mongiat, J. , Pages, G. , O'Rourke, D.M. , Bernhard, E. , 2004. Sp1 is involved in Akt-mediated induction of VEGF expression through an HIF-1-independent mechanism. Mol. Biol. Cell 15, 4841–4853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Potter, C.P. , Harris, A.L. , 2003. Diagnostic, prognostic and therapeutic implications of carbonic anhydrases in cancer. Br. J. Cancer 89, 2–7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rajendran, J.G. , Schwartz, D.L. , O'Sullivan, J. , Peterson, L.M. , Ng, P. , Scharnhorst, J. , Grierson, J.R. , Krohn, K.A. , 2006. Tumor hypoxia imaging with [F-18] fluoromisonidazole positron emission tomography in head and neck cancer. Clin. Cancer Res. 12, 5435–5441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raleigh, J.A. , Chou, S.C. , Arteel, G.E. , Horsman, M.R. , 1999. Comparisons among pimonidazole binding, oxygen electrode measurements, and radiation response in C3H mouse tumors. Radiat. Res. 151, 580–589. [PubMed] [Google Scholar]
- Ramankulov, A. , Lein, M. , Kristiansen, G. , Meyer, H.A. , Loening, S.A. , Jung, K. , 2007. Elevated plasma osteopontin as marker for distant metastases and poor survival in patients with renal cell carcinoma. J. Cancer Res. Clin. 133, 643–652. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rangaswami, H. , Bulbule, A. , Kundu, G.C. , 2006. Osteopontin: role in cell signaling and cancer progression. Trends Cell Biol. 16, 79–87. [DOI] [PubMed] [Google Scholar]
- Rijpkema, M. , Kaanders, J.H. , Joosten, F.B. , van der Kogel, A.J. , Heerschap, A. , 2002. Effects of breathing a hyperoxic hypercapnic gas mixture on blood oxygenation and vascularity of head-and-neck tumors as measured by magnetic resonance imaging. Int. J. Radiat. Oncol. Biol. Phys. 53, 1185–1191. [DOI] [PubMed] [Google Scholar]
- Rischin, D. , Peters, L. , Fisher, R. , Macann, A. , Denham, J. , Poulsen, M. , Jackson, M. , Kenny, L. , Penniment, M. , Corry, J. , 2005. Tirapazamine, cisplatin, and radiation versus fluorouracil, cisplatin, and radiation in patients with locally advanced head and neck cancer: a randomized phase II trial of the Trans-Tasman Radiation Oncology Group (TROG 98.02). J. Clin. Oncol. 23, 79–87. [DOI] [PubMed] [Google Scholar]
- Rittling, S.R. , Chambers, A.F. , 2004. Role of osteopontin in tumour progression. Br. J. Cancer 90, 1877–1881. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rogers, S. , Macheda, M.L. , Docherty, S.E. , Carty, M.D. , Henderson, M.A. , Soeller, W.C. , Gibbs, E.M. , James, D.E. , Best, J.D. , 2002. Identification of a novel glucose transporter-like protein-GLUT-12. Am. J. Physiol. Endocrinol. Metab. 282, E733–E738. [DOI] [PubMed] [Google Scholar]
- Rudland, P.S. , Platt-Higgins, A. , El-Tanani, M. , De-Silva-Rudland, S. , Barraclough, R. , Winstanley, J.H. , Howitt, R. , West, C.R. , 2002. Prognostic significance of the metastasis-associated protein osteopontin in human breast cancer. Cancer Res. 62, 3417–3427. [PubMed] [Google Scholar]
- Sandler, A. , Gray, R. , Perry, M.C. , Brahmer, J. , Schiller, J.H. , Dowlati, A. , Lilenbaum, R. , Johnson, D.H. , 2006. Paclitaxel-carboplatin alone or with bevacizumab for non-small-cell lung cancer. New Engl. J. Med. 355, 2542–2550. [DOI] [PubMed] [Google Scholar]
- Schilling, D. , Bayer, C. , Geurts-Moespot, A. , Sweep, F.C. , Pruschy, M. , Mengele, K. , Sprague, L.D. , Molls, M. , 2007. Induction of plasminogen activator inhibitor type-1 (PAI-1) by hypoxia and irradiation in human head and neck carcinoma cell lines. BMC Cancer 7, 143 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schuuring, J. , Bussink, J. , Bernsen, H.J. , Peeters, W. , van-Der-Kogel, A.J. , 2005. Irradiation combined with SU5416: microvascular changes and growth delay in a human xenograft glioblastoma tumor line. Int. J. Radiat. Oncol. Biol. Phys. 61, 529–534. [DOI] [PubMed] [Google Scholar]
- Semenza, G.L. , 2000. Hypoxia, clonal selection, and the role of HIF-1 in tumor progression. Crit. Rev. Biochem. Mol. Biol. 35, 71–103. [DOI] [PubMed] [Google Scholar]
- Semenza, G.L. , 2007. Evaluation of HIF-1 inhibitors as anticancer agents. Drug Discov. Today 12, 853–859. [DOI] [PubMed] [Google Scholar]
- Span, P.N. , Bussink, J. , Manders, P. , Beex, L.V. , Sweep, C.G. , 2003. Carbonic anhydrase-9 expression levels and prognosis in human breast cancer: association with treatment outcome. Br. J. Cancer 89, 271–276. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stillebroer, A.B. , Oosterwijk, E. , Oyen, W.J. , Mulders, P.F. , Boerman, O.C. , 2007. Radiolabeled antibodies in renal cell carcinoma. Cancer Imaging 7, 179–188. The official publication of the International Cancer Imaging Society [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talbot, D.C. , von Pawel, J. , Cattell, E. , Yule, S.M. , Johnston, C. , Zandvliet, A.S. , Huitema, A.D. , Norbury, C.J. , Ellis, P. , Bosquee, L. , 2007. A randomized phase II pharmacokinetic and pharmacodynamic study of indisulam as second-line therapy in patients with advanced non-small cell lung cancer. Clin. Cancer Res. 13, 1816–1822. [DOI] [PubMed] [Google Scholar]
- Tredan, O. , Galmarini, C.M. , Patel, K. , Tannock, I.F. , 2007. Drug resistance and the solid tumor microenvironment. J. Natl. Cancer Inst. 99, 1441–1454. [DOI] [PubMed] [Google Scholar]
- Troost, E.G. , Bussink, J. , Kaanders, J.H. , van-Eerd, J. , Peters, J.P. , Rijken, P.F. , Boerman, O.C. , van-der-Kogel, A.J. , 2005. Comparison of different methods of CAIX quantification in relation to hypoxia in three human head and neck tumor lines. Radiother. Oncol. 76, 194–199. [DOI] [PubMed] [Google Scholar]
- Vaupel, P. , Thews, O. , Hoeckel, M. , 2001. Treatment resistance of solid tumors: role of hypoxia and anemia. Med. Oncol 18, 243–259. [DOI] [PubMed] [Google Scholar]
- Wang, G.L. , Jiang, B.H. , Rue, E.A. , Semenza, G.L. , 1995. Hypoxia-inducible factor 1 is a basic-helix-loop–helix-PAS heterodimer regulated by cellular O2 tension. Proc. Natl. Acad. Sci. USA 92, 5510–5514. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Williamson, S.K. , Crowley, J.J. , Lara, P.N. , McCoy, J. , Lau, D.H. , Tucker, R.W. , Mills, G.M. , Gandara, D.R. , 2005. Phase III trial of paclitaxel plus carboplatin with or without tirapazamine in advanced non-small-cell lung cancer: Southwest Oncology Group Trial S0003. J. Clin. Oncol. 23, 9097–9104. [DOI] [PubMed] [Google Scholar]
- Wilson, R.E. , Keng, P.C. , Sutherland, R.M. , 1989. Drug resistance in Chinese hamster ovary cells during recovery from severe hypoxia. J. Natl. Cancer Inst 81, 1235–1240. [DOI] [PubMed] [Google Scholar]
- Wouters, B.G. , Brown, J.M. , 1997. Cells at intermediate oxygen levels can be more important than the “hypoxic fraction” in determining tumor response to fractionated radiotherapy. Radiat. Res. 147, 541–550. [PubMed] [Google Scholar]
- Wykoff, C.C. , Beasley, N.J.P. , Watson, P.H. , Turner, K.J. , Pastorek, J. , Sibtain, A. , Wilson, G.D. , Turley, H. , Talks, K.L. , Maxwell, P.H. , 2000. Hypoxia-inducible expression of tumor-associated carbonic anhydrases. Cancer Res. 60, 7075–7083. [PubMed] [Google Scholar]
- Zhang, H. , Ye, Q.H. , Ren, N. , Zhao, L. , Wang, Y.F. , Wu, X. , Sun, H.C. , Wang, L. , Zhang, B.H. , Liu, Y.K. , 2006. The prognostic significance of preoperative plasma levels of osteopontin in patients with hepatocellular carcinoma. J. Cancer Res. Clin. 132, 709–717. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhong, H. , De-Marzo, A.M. , Laughner, E. , Lim, M. , Hilton, D.A. , Zagzag, D. , Buechler, P. , Isaacs, W.B. , Semenza, G.L. , Simons, J.W. , 1999. Overexpression of hypoxia-inducible factor 1alpha in common human cancers and their metastases. Cancer Res. 59, 5830–5835. [PubMed] [Google Scholar]