

Abstract
Lung cancer is a devastating disease and a major therapeutic burden with poor survival rates. The discovery of rare cells with stem cell‐like properties in solid tumours is emerging as an important area of cancer research and may help explain the resistance of these tumours to current therapeutics. Despite rapid developments in cancer stem cell research in other solid tumours, progress in the lung has been hampered by an incomplete understanding of the epithelial stem cell hierarchy, the heterogeneity of disease and the lack of a suitable in vivo transplantation model to assess stem cell behaviour. In this review we critically discuss what is currently known about the role of normal stem cells and cancer‐initiating cells in lung tumour development, and briefly discuss strategies aimed at advancing the field of lung stem cell biology, with an emphasis on the design and manipulation of state‐of‐art mouse models.
Keywords: Lung cancer, Stem cells, Progenitor cells, Cancer-initiating cells
1. Introduction
Lung cancer is the leading cause of cancer deaths worldwide (Landis et al., 1999). Human lung cancer can be divided into two main histopathological groups: non‐small cell lung cancer (NSCLC) (van Zandwijk et al., 1995) and small‐cell lung cancer (SCLC) (Schiller, 2001). About 80% of lung cancers are NSCLC, and they can be further subdivided into adenocarcinomas, squamous cell, bronchioalveolar, and large cell carcinomas (Travis, 2002). The remaining 20% of lung cancers show properties of neuroendocrine differentiation. Lessons learnt from murine models of human NSCLC as well as SCLC cancer (Jackson et al., 2001; Meuwissen et al., 2003), indicate that the genetic alterations and the cell type in which the mutations occur dictate the tumour phenotype. To date, however, the cellular origin of lung cancer is largely unknown. The tumour subtypes generally follow a proximal‐to‐distal distribution pattern moving distally from the trachea: squamous cell carcinoma (SCC), SCLC and adenocarcinoma/bronchioalveolar carcinoma (reviewed in (Giangreco et al., 2007)). Thus, it is speculated that these different tumour subclasses arise from distinct cells of origin localised within a defined regional compartment/microenvironment. The identification of these cells of origin has significant implications, not only for our understanding of lung cancer biology, but presumably also for lung cancer therapies.
As discussed by others (Reya et al., 2001; Visvader and Lindeman, 2008), putative cancer‐initiating cells are likely to exhibit properties inherent to normal tissue stem cells. Indeed, the existence of subpopulations of cancer cells with stem cell‐like properties has been demonstrated in multiple solid tumours (Al‐Hajj et al., 2003). The identification and characterization of lung tissue stem cells has however progressed slowly. This can be largely attributed to the complex anatomical structure of the tissue, its cellular heterogeneity and the extremely slow turn‐over rate of lung epithelium. This raises the question of whether cancer‐initiating cells also originate from early progenitors (for example, normal tissue stem cells) in tissues with a slow turn‐over rate. It is becoming evident that the acquisition of deleterious mutations may not only occur in slowly proliferating stem cells, but also in more differentiated cell types with sufficient amplifying capacity (for example, transit amplifying cells) as occurs in highly proliferating tissues, like skin and gut.
2. Non‐small cell lung cancer (NSCLC)
Increasing data over recent years have indicated the existence of cancer‐initiating cells, with stem cell characteristics in lung tumours. Initial observations (Jackson et al., 2001) suggested that lung tumours initiated by oncogenic K‐Ras activation appeared to be derived from targeted cells located in the bronchioalveolar duct junction (BADJ), a region where the airways terminate and form alveoli (Fig. 1). The cellular composition of this microenvironment has been subjected to extensive examination over the years and shown to harbour label‐retaining cells (Giangreco et al., 2002). Close examination of these early lung tumour lesions revealed the presence of a rare cell population shown to co‐express the Clara cell marker CC10 (also known as CCA, CCSP, uteroglobin and Scgb1a1) and the alveolar type 2 marker, Surfactant Protein C (SPC) (Jackson et al., 2001). As an extension of this initial observation, in normal lung homeostasis, double‐positive (CC10 and SPC) cells were shown to be quiescent, but became active to proliferate upon lung injury induced by naphthalene (Kim et al., 2005). Moreover, double‐positive cells (CC10 and SPC) were shown to express stem cell markers Sca‐1 and CD34, established surface markers of hematopoietic and skin stem cells, respectively (Blanpain et al., 2004; Morrison and Weissman, 1994), and could be further purified from freshly prepared lung tissue following the exclusion of CD45‐positive hematopoietic and CD31‐positive endothelial cells (Kim et al., 2005). FACS sorted Sca‐1posCD34pos cell populations displayed enhanced self‐renewal and differentiation properties in vitro. Moreover, when plated in Matrigel, these double‐positive cells gave rise to Clara, alveolar type 2 and alveolar type I cells, suggesting that they exhibit progenitor cell‐like features. Thus, Kim et al. (2005) termed these cells Bronchioalveolar Stem Cells (BASCs).
Figure 1.
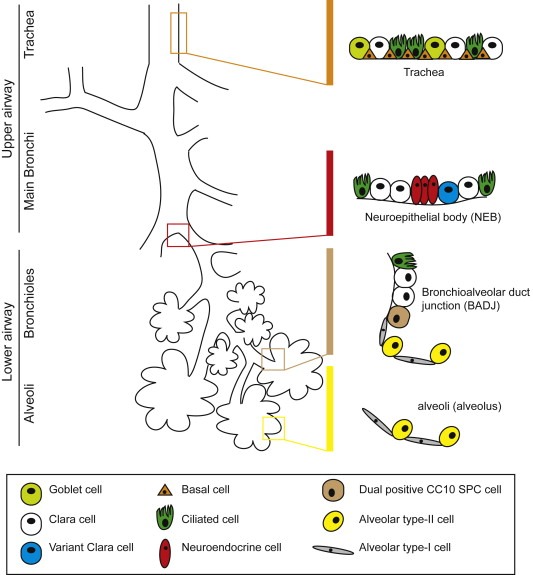
A schematic diagram of the mouse lung highlighting the spatially distinct cellular environments shown to harbour airway stem/progenitor cells. The mouse lung consists of three main levels of conducting airways: the trachea, bronchi and bronchioles. Candidate epithelial niches (depicted on the right hand side) have been identified and found to exist in spatially defined regions: the tracheal submucosal gland ducts, neuroendocrine bodies (NEB) of the bronchi/bronchioles, and the broncholalveolar duct junction (BADJ).
The progenitor‐like properties of BASCs have come under critical scrutiny over the past few years. This can be in part attributed to the lack of bona fide in vivo evidence to verify that BASCs are indeed stem cells for the bronchioalveolar and alveoli epithelium. Recent findings obtained from elegant lineage tracing studies performed by Rawlins et al. (2009) do not support a role for BASCs in maintenance, growth and repair of the bronchiolar and alveolar regions. Furthermore the utility of Sca‐1 as a lung stem cell marker has also been recently questioned. In contrast to observations made by Kim et al. (2005), freshly isolated CD45negCD31negSca‐1pos cells were shown to exhibit a mesenchymal gene expression profile, while CD45negCD31negSca‐1neg cells exhibited a profile of epithelial cells (McQualter et al., 2009). This epithelial cell population was further divided on the basis of the expression of a panel of cell surface markers. In a series of in vitro assays, EpCAMhiCD49fposCD104posCD24low epithelia exhibited colony‐forming potential and gave rise to committed clonogenic airway and alveolar progenitor cells (McQualter et al., 2010). It is feasible that differences observed when using Sca‐1 as a prospective lung stem cell marker could arise from variations in mouse strain, cell isolation, culture conditions and FACS sorting parameters. These factors vary between studies and thus could influence the interpretation of results (reviewed in (Raiser and Kim, 2009)). Taken together, these studies highlight the importance and necessity of in vivo approaches and meticulous cell surface marker examination in order to clarify stem cell identity and biology.
Even though it would appear that BASCs do not behave as stem cells for the bronchioalveolar and alveoli epithelium, they could still be the target cells of origin for lung cancer. Immunofluorescence studies performed by Kim et al showed elevated numbers of CC10 SPC double‐positive BASCs in early tumorigenic lesions in Lox‐K‐ras mice (Kim et al., 2005). Moreover, a continual expansion in BASC number correlated with tumour progression in Lox‐K‐ras mice, implicating BASCs as the cell of origin of lung adenocarcinoma. Furthermore, combined K‐Ras G12D activation with naphthalene treatment resulted in an increase in the number and the size of tumours (Fig. 3) (Kim et al., 2005). Taken together, findings obtained from this study were interpreted to implicate a role for BASCs in the initiation of tumorigenesis. A tumour xenograft experiment is however required to confirm the tumorigenic capacity of these double‐positive cells with respect to the remaining tumour population. The subcutaneous injection of putative “cancer‐initiating cells” in immunocompromised mice is a widely used method to test the tumour propagating capacity of a defined cell population. One caveat of this transplantation assay is that it fails to mimic the cellular niche from which the tumour arises. This could however be overcome by orthotopic transplantation of candidate cancer‐initiating cells back into their normal microenvironment. With regards to putative lung cancer‐initiating cells, this could be achieved by intratracheal delivery of FACS sorted cell subpopulations into the lung cavity using methodology employed for the delivery of an adeno‐Cre virus in sporadic murine models of human lung cancer (Meuwissen et al., 2003, 2001). Further investigation into the molecular mechanisms that regulate the maintenance and proliferation of BASCs have predominately arisen from in vivo studies whereby genes with established roles in lung development or carcinogensis have been manipulated. Targeted loss of either GATA‐6, PTEN, PI3‐K or p38α MAP kinase all resulted in increases in the abundance of double‐positive (CC10 and SPC) cells (Ventura et al., 2007; Yanagi et al., 2007; Yang et al., 2008; Zhang et al., 2008).
Figure 3.
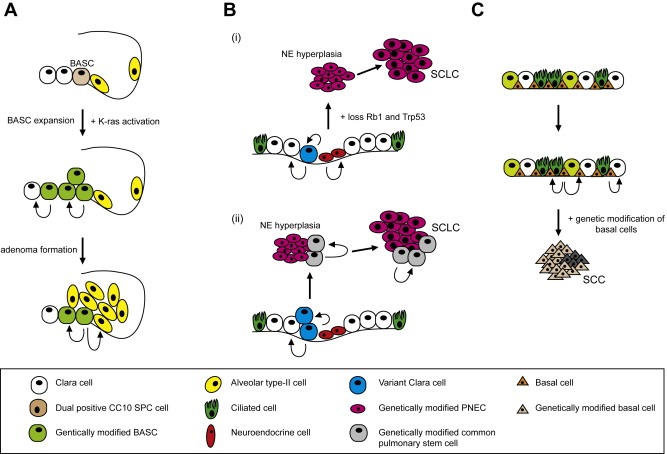
Schematic overview of the putative role of normal tissue stem/progenitor cells in lung cancer. (A) The putative role of BASCs in the ontogeny of murine NSCLC. Depicted is a bronchioalveolar duct junction (BADJ) that contains a rare cell population that expresses both Clara‐specific and alveolar‐specific markers. These cells are bronchioalveolar stem cells (BASCs). Activation of K‐Ras results in enhanced proliferation of BASCs, with no effect on the proliferation of alveolar type 2 cells. (B) Two hypotheses explaining the origin of lung NE tumours. In the epithelial lining of the bronchi there are neuroendocrine bodies (NEBs), which harbour pulmonary neuroendocrine cells (PNECs) that are associated with variant Clara expressing cells (vCEs). One current line of thinking (i) is that NE tumours arise from PNECs. The other hypothesis (ii) is that NE hyperplasia and SCLC arise from a less‐differentiated progenitor‐like cell (for example vCE). This would predict a mixture of undifferentiated and differentiated cancer cells. Whether NE hyperplasia and SCLC harbour cancer stem cells arising from a common pulmonary stem cell remains unknown. (C) A hypothesis explaining the origin of lung SCC. In the trachea a KRT5+KRT14+p63+ basal cell population exists, which exhibit properties of stem cells. Given the basal‐like phenotype of SCC, one could hypothesize that squamous cell tumours arise from these basal stem cells. Genetic modification of these cells, for example by SOX2 over expression, may result in increased proliferation of tracheal epithelium.
3. Small‐cell lung cancer (SCLC)
The specific cell population(s) that gives rise, upon genetic alteration, to SCLC is not yet known. Human and mouse SCLC predominately localise to the midlevel bronchioles and typically express a range of neuroendocrine markers, such as calcitonin‐gene related peptide (CGRP) and neural cell adhesion molecule (Ncam1), and transcription factors which play important roles in neuroendocrine differentiation, including achaete‐scute complex homolog‐like 1 (ASCL1) (Meuwissen et al., 2003). Based on these observations, it has been hypothesized that a rare population of neuroendocrine (NE) cells are the progenitors of SCLC (Fig. 3B) (Wistuba et al., 2001). In the mouse lung, microenvironments found in close proximity to neuroepithelial bodies (NEB) (Fig. 1) have been shown to maintain putative stem cell populations (Reynolds et al., 2000a). However, NEB‐associated pulmonary NE cells (PNECs) do not behave as stem cells, instead it would appear that they exhibit properties of unipotent progenitor cells (Reynolds et al., 2000, 2000). It is also not clear whether the same cell of origin is responsible for initiating both SCLC and NSCLC. The observation that a proportion of SCLCs display a mixture of SCLC‐ and NSCLC‐specific features may argue for the existence of a “common” cell of origin for these lung cancers (Yesner, 2001).
4. Squamous cell carcinoma (SCC)
The histopathology and gene expression patterns of mouse lung SCC‐like lesions frequently resemble tracheal basal cell progenitors, thus it is attractive to speculate that these are the target cells of origin in this subclass of lung cancer (Fig. 3C). These progenitors are keratin (KRT) 5/14 positive, express the transcription factor p63 and are located at the submucosal gland duct junctions or intracartilaginous boundaries (Fig. 1) (Borthwick et al., 2001, 2004, 2004). Recently in order to directly examine the properties of basal cells, Rock et al. (2009) generated a transgenic mouse whereby a tamoxifen inducible Cre‐recombinase (CreER T2) is under the control of the human keratin 5 (KRT5) gene promoter (KRT5‐CreER T2). When used in combination with reporter animals, the author's had the ability to label and subsequently trace basal cells of the trachea. In steady‐state conditions and in response to epithelial injury, lacZ+ basal, Clara and Ciliated cells were detected in the tracheas of KRT5‐CreER T2; Rosa26R‐lacZ adult mice at defined time‐points following tamoxifen administration (Fig. 2) (Rock et al., 2009). Thus, KRT5+ basal cells have the ability to self‐renew and generate luminal cells in the trachea. Amplification of chromosome segment 3q26.33 is found in human lung and esophageal SCCs (Bass et al., 2009). Interestingly, the transcription factor Sox2, which plays a key role in the proliferation and differentiation of basal progenitor cells, resides within 3q26.33, and was shown to be the relevant gene for the amplification (Bass et al., 2009; Que et al., 2009). In order to establish a clear relationship between basal progenitors and lung SCC new mouse models whereby genetic modifications associated with SCC are restricted to basal progenitor cells need to be generated.
Figure 2.
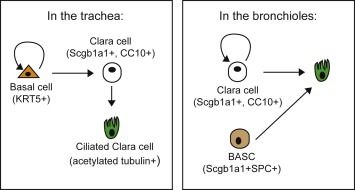
Stem/progenitor properties of lung epithelial cells present in the spatially distinct regions of the mouse lung. A summary of what is known to date about the stem/progenitor cell properties present in the trachea and bronchi/bronchioles. The solid lines depict properties based upon evidence gained from “pulse‐chase” lineage‐labelling experiments in the adult mouse. A curly arrow indicates the ability of a cell to self‐renew.
5. Cancer stem cells (CSCs)
Recently, using a similar strategy to that of Bonnet and Dick (Bonnet and Dick, 1997), Eramo et al. (2008) demonstrated that human SCLC and NSCLC tumours contain a rare population of undifferentiated cells that express the cell surface marker CD133. CD133 is a well‐characterized marker of cancer stem cells (CSCs) in different types of brain tumours (Singh et al., 2004), and appears to be a marker of normal neural stem cells in both human and mouse (Lee et al., 2005; Uchida et al., 2000). Isolated CD133pos lung cancer cells grew as spheres in serum‐free culture conditions and when subcutaneously injected into immunocompromised animals, formed xenografts that were phenotypically identical to the original tumour whereas CD133neg cells could not (Eramo et al., 2008). In normal lung tissue, the cell type which CD133 labels is not yet known. Using a well‐characterized mouse model of lung epithelial injury, an increase in CD133 cells (from 0.6 to 3.8% of total lung epithelium) was observed 7 days following naphthalene administration (Eramo et al., 2008). Thus, it is tempting to speculate that CD133 marks progenitor cells (variant Clara cells) involved in regenerating the epithelial lining following injury however, immunohistochemical analysis is required to confirm this hypothesis.
Chemoresistance is a claimed hallmark of CSCs and is postulated to be the mechanism behind the resistance of tumours to chemotherapy (reviewed in (Dean et al., 2005)). Given that current therapies used in the treatment of lung cancer are not efficient in eradicating the disease, it is attractive to speculate the existence of a chemoresistant population of CSCs in human lung cancer. Interestingly, CD133pos lung cancer spheres exhibited chemoresistance in vitro (Eramo et al., 2008). This was associated with increased expression of efflux transporters from the ATP‐binding cassette (ABC) gene family, which have an established role in multi‐drug resistance of tumour cells. In an independent study, cisplatin treatment of mice carrying human lung cancer xenografts resulted in a variable inhibition of tumour volume and was associated with an enrichment of the CD133pos fraction present within cisplatin‐treated tumours (Bertolini et al., 2009). In a more recent study, a sporadic mouse model of human NSCLC, based on the conditional activation of Kras in lung epithelium (Jackson et al., 2001) was used to assess the tumour response mechanisms to cisplatin (Oliver et al., 2010). Similar to their human counterparts, mouse NSCLC tumours initially responded to cisplatin, however acquired resistance following prolonged exposure. Interestingly, the DNA damage, cell cycle arrest and apoptotic response initially observed following cisplatin treatment was shown genetically to be independent of the tumour suppressor p53 and its transcriptional regulator, p21 (Oliver et al., 2010). Whether a CD133pos cell population is involved in chemoresistance in mouse lung tumours is not clear, and to our knowledge has not been explored. This would however be an interesting line of research to pursue and may clarify the importance of CD133 as a CSC marker in lung tumours.
6. The search for lung cancer‐initiating cells continues
Much work is still required before we can confidently state whether CSCs exist in lung tumours. The identity of stem and progenitor cell populations in the lung requires further scrutiny before a relationship with lung cancer can be inferred. Additional studies, discussed in greater detail below, using lineage tagging of candidate stem/progenitor cells and highly regulated mouse models of all subclasses of human lung cancer are still required.
Data arising in recent years from the Hogan Laboratory highlights the importance of lineage tracing as a method to study candidate stem/progenitor cells in vivo (Rawlins et al., 2009, 2007, 2009). This strategy will broaden our understanding of the lineage relationships between lung epithelial populations as, compared to the hematopoietic system where the cell hierarchy is well defined, little is still known about the cellular populations hypothesized to be present within the lung. However, to date the identification of these different cell types relies in a large part on the expression of secreted proteins, which is not ideal for sorting cells by FACS. Thus progress needs to be made in finding better cell surface markers, which can be used to distinguish and sort different epithelial cell types.
One of the biggest caveats hampering stem cell research in the lung field is the lack of a suitable in vivo transplantation model. The value of establishing such a model is illustrated by the advances made in the mammary gland field, where a complete mammary gland can be grown following implantation of a single adult stem cell into a cleared mammary fat pad (Shackleton et al., 2006; Stingl et al., 2006). One approach, which may help in defining stem cell populations within the tracheal niche, is the rat xenograft model (reviewed in (Liu et al., 2006)). In this model isolated epithelial cells are dissociated and seeded on the surface of a rat trachea that has been denuded of endogenous epithelium by freeze/thawing. The trachea is then grafted subcutaneously into an immunocompromised mouse. Three to 4 weeks later, a fully differentiated, normal epithelium is restored. The advantage of this approach is that it can be used for both human and mouse cells. Moreover, it can be used to assess the stem cell properties of different purified epithelial populations isolated on the basis of surface marker expression. Another approach, which may assist in identifying stem cell potential is an in vitro organ culture system. Over the past years significant advances have been reported in the ability to derive and maintain in vitro organ cultures from isolated tissue stem cells. Pioneering studies performed in the laboratory of Hans Clevers, describe in vitro culture conditions that support the establishment of small intestine crypt cultures from single Lgr5+ stem cells (Sato et al., 2009). Similar advances in this technique are currently being made with the lung, whereby culture conditions have been established to support mouse tracheal, bronchi and alveolar progenitor cells (McQualter et al., 2010; Rock et al., 2009).
The generation of mouse models of human lung cancer has been pivotal in broadening our current understanding of lung cancer biology, by unveiling critical underlying pathways that control tumour formation, maintenance and progression. Further manipulation of these models by conditional activation or inactivation of oncogenes and/or tumour suppressors in cell type specific compartments will assist in defining the cancer cell of origin in lung cancer. This strategy has been successfully employed to identify the cell of origin in pancreatic tumours initiated by activated K‐Ras (Gidekel Friedlander et al., 2009).
Taken together, lessons taught by these studies will hopefully assist in the development of more effective cellular and molecular targeted therapies for the treatment of lung disease.
Sutherland Kate D., Berns Anton, (2010), Cell of origin of lung cancer, Molecular Oncology, 4, doi: 10.1016/j.molonc.2010.05.002.
Contributor Information
Kate D. Sutherland, Email: k.sutherland@nki.nl
Anton Berns, Email: a.berns@nki.nl.
References
- Al-Hajj, M. , Wicha, M.S. , Benito-Hernandez, A. , Morrison, S.J. , Clarke, M.F. , 2003. Prospective identification of tumorigenic breast cancer cells. Proc. Natl. Acad. Sci. U. S. A. 100, 3983–3988. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bass, A.J. , Watanabe, H. , Mermel, C.H. , Yu, S. , Perner, S. , Verhaak, R.G. , 2009. SOX2 is an amplified lineage-survival oncogene in lung and esophageal squamous cell carcinomas. Nat. Genet. 41, 1238–1242. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bertolini, G. , Roz, L. , Perego, P. , Tortoreto, M. , Fontanella, E. , Gatti, L. , 2009. Highly tumorigenic lung cancer CD133+ cells display stem-like features and are spared by cisplatin treatment. Proc. Natl. Acad. Sci. U. S. A. 106, 16281–16286. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blanpain, C. , Lowry, W.E. , Geoghegan, A. , Polak, L. , Fuchs, E. , 2004. Self-renewal, multipotency, and the existence of two cell populations within an epithelial stem cell niche. Cell. 118, 635–648. [DOI] [PubMed] [Google Scholar]
- Bonnet, D. , Dick, J.E. , 1997. Human acute myeloid leukemia is organized as a hierarchy that originates from a primitive hematopoietic cell. Nat. Med. 3, 730–737. [DOI] [PubMed] [Google Scholar]
- Borthwick, D.W. , Shahbazian, M. , Krantz, Q.T. , Dorin, J.R. , Randell, S.H. , 2001. Evidence for stem-cell niches in the tracheal epithelium. Am. J. Respir. Cell. Mol. Biol. 24, 662–670. [DOI] [PubMed] [Google Scholar]
- Dean, M. , Fojo, T. , Bates, S. , 2005. Tumour stem cells and drug resistance. Nat. Rev. Cancer. 5, 275–284. [DOI] [PubMed] [Google Scholar]
- Eramo, A. , Lotti, F. , Sette, G. , Pilozzi, E. , Biffoni, M. , Di Virgilio, A. , 2008. Identification and expansion of the tumorigenic lung cancer stem cell population. Cell Death Differ. 15, 504–514. [DOI] [PubMed] [Google Scholar]
- Giangreco, A. , Groot, K.R. , Janes, S.M. , 2007. Lung cancer and lung stem cells: strange bedfellows?. Am. J. Respir. Crit. Care Med. 175, 547–553. [DOI] [PubMed] [Google Scholar]
- Giangreco, A. , Reynolds, S.D. , Stripp, B.R. , 2002. Terminal bronchioles harbor a unique airway stem cell population that localizes to the bronchoalveolar duct junction. Am. J. Pathol. 161, 173–182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gidekel Friedlander, S.Y. , Chu, G.C. , Snyder, E.L. , Girnius, N. , Dibelius, G. , Crowley, D. , 2009. Context-dependent transformation of adult pancreatic cells by oncogenic K-Ras. Cancer Cell. 16, 379–389. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hong, K.U. , Reynolds, S.D. , Watkins, S. , Fuchs, E. , Stripp, B.R. , 2004. Basal cells are a multipotent progenitor capable of renewing the bronchial epithelium. Am. J. Pathol. 164, 577–588. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hong, K.U. , Reynolds, S.D. , Watkins, S. , Fuchs, E. , Stripp, B.R. , 2004. In vivo differentiation potential of tracheal basal cells: evidence for multipotent and unipotent subpopulations. Am. J. Physiol. Lung Cell. Mol. Physiol. 286, L643–L649. [DOI] [PubMed] [Google Scholar]
- Jackson, E.L. , Willis, N. , Mercer, K. , Bronson, R.T. , Crowley, D. , Montoya, R. , 2001. Analysis of lung tumor initiation and progression using conditional expression of oncogenic K-ras. Genes Dev. 15, 3243–3248. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kim, C.F. , Jackson, E.L. , Woolfenden, A.E. , Lawrence, S. , Babar, I. , Vogel, S. , 2005. Identification of bronchioalveolar stem cells in normal lung and lung cancer. Cell. 121, 823–835. [DOI] [PubMed] [Google Scholar]
- Landis, S.H. , Murray, T. , Bolden, S. , Wingo, P.A. , 1999. Cancer statistics, 1999. CA Cancer J. Clin. 49, 8–31. 31 [DOI] [PubMed] [Google Scholar]
- Lee, A. , Kessler, J.D. , Read, T.A. , Kaiser, C. , Corbeil, D. , Huttner, W.B. , 2005. Isolation of neural stem cells from the postnatal cerebellum. Nat. Neurosci. 8, 723–729. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liu, X. , Driskell, R.R. , Engelhardt, J.F. , 2006. Stem cells in the lung. Methods Enzymol. 419, 285–321. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McQualter, J.L. , Brouard, N. , Williams, B. , Baird, B.N. , Sims-Lucas, S. , Yuen, K. , 2009. Endogenous fibroblastic progenitor cells in the adult mouse lung are highly enriched in the sca-1 positive cell fraction. Stem Cells. 27, 623–633. [DOI] [PubMed] [Google Scholar]
- McQualter, J.L. , Yuen, K. , Williams, B. , Bertoncello, I. , 2010. Evidence of an epithelial stem/progenitor cell hierarchy in the adult mouse lung. Proc. Natl. Acad. Sci. U. S. A. 107, 1414–1419. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Meuwissen, R. , Linn, S.C. , Linnoila, R.I. , Zevenhoven, J. , Mooi, W.J. , Berns, A. , 2003. Induction of small cell lung cancer by somatic inactivation of both Trp53 and Rb1 in a conditional mouse model. Cancer Cell. 4, 181–189. [DOI] [PubMed] [Google Scholar]
- Meuwissen, R. , Linn, S.C. , van der Valk, M. , Mooi, W.J. , Berns, A. , 2001. Mouse model for lung tumorigenesis through Cre/lox controlled sporadic activation of the K-Ras oncogene. Oncogene. 20, 6551–6558. [DOI] [PubMed] [Google Scholar]
- Morrison, S.J. , Weissman, I.L. , 1994. The long-term repopulating subset of hematopoietic stem cells is deterministic and isolatable by phenotype. Immunity. 1, 661–673. [DOI] [PubMed] [Google Scholar]
- Oliver, T.G. , Mercer, K.L. , Sayles, L.C. , Burke, J.R. , Mendus, D. , Lovejoy, K.S. , 2010. Chronic cisplatin treatment promotes enhanced damage repair and tumor progression in a mouse model of lung cancer. Genes Dev. 24, 837–852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Que, J. , Luo, X. , Schwartz, R.J. , Hogan, B.L. , 2009. Multiple roles for Sox2 in the developing and adult mouse trachea. Development. 136, 1899–1907. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raiser, D.M. , Kim, C.F. , 2009. Commentary: Sca-1 and cells of the lung: a matter of different sorts. Stem Cells. 27, 606–611. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rawlins, E.L. , Okubo, T. , Xue, Y. , Brass, D.M. , Auten, R.L. , Hasegawa, H. , 2009. The role of Scgb1a1 + Clara cells in the long-term maintenance and repair of lung airway, but not alveolar, epithelium. Cell Stem Cell. 4, 525–534. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rawlins, E.L. , Ostrowski, L.E. , Randell, S.H. , Hogan, B.L. , 2007. Lung development and repair: contribution of the ciliated lineage. Proc. Natl. Acad. Sci. U. S. A. 104, 410–417. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reya, T. , Morrison, S.J. , Clarke, M.F. , Weissman, I.L. , 2001. Stem cells, cancer, and cancer stem cells. Nature. 414, 105–111. [DOI] [PubMed] [Google Scholar]
- Reynolds, S.D. , Giangreco, A. , Power, J.H. , Stripp, B.R. , 2000. Neuroepithelial bodies of pulmonary airways serve as a reservoir of progenitor cells capable of epithelial regeneration. Am. J. Pathol. 156, 269–278. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reynolds, S.D. , Hong, K.U. , Giangreco, A. , Mango, G.W. , Guron, C. , Morimoto, Y. , Stripp, B.R. , 2000. Conditional clara cell ablation reveals a self-renewing progenitor function of pulmonary neuroendocrine cells. Am. J. Physiol. Lung Cell. Mol. Physiol. 278, L1256–L1263. [DOI] [PubMed] [Google Scholar]
- Rock, J.R. , Onaitis, M.W. , Rawlins, E.L. , Lu, Y. , Clark, C.P. , Xue, Y. , 2009. Basal cells as stem cells of the mouse trachea and human airway epithelium. Proc. Natl. Acad. Sci. U. S. A. 106, 12771–12775. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sato, T. , Vries, R.G. , Snippert, H.J. , van de Wetering, M. , Barker, N. , Stange, D.E. , 2009. Single Lgr5 stem cells build crypt-villus structures in vitro without a mesenchymal niche. Nature. 459, 262–265. [DOI] [PubMed] [Google Scholar]
- Schiller, J.H. , 2001. Current standards of care in small-cell and non-small-cell lung cancer. Oncology. 61, (Suppl. 1) 3–13. [DOI] [PubMed] [Google Scholar]
- Shackleton, M. , Vaillant, F. , Simpson, K.J. , Stingl, J. , Smyth, G.K. , Asselin-Labat, M.L. , 2006. Generation of a functional mammary gland from a single stem cell. Nature. 439, 84–88. [DOI] [PubMed] [Google Scholar]
- Singh, S.K. , Hawkins, C. , Clarke, I.D. , Squire, J.A. , Bayani, J. , Hide, T. , 2004. Identification of human brain tumour initiating cells. Nature. 432, 396–401. [DOI] [PubMed] [Google Scholar]
- Stingl, J. , Eirew, P. , Ricketson, I. , Shackleton, M. , Vaillant, F. , Choi, D. , 2006. Purification and unique properties of mammary epithelial stem cells. Nature. 439, 993–997. [DOI] [PubMed] [Google Scholar]
- Travis, W.D. , 2002. Pathology of lung cancer. Clin. Chest Med. 23, 65–81. viii [DOI] [PubMed] [Google Scholar]
- Uchida, N. , Buck, D.W. , He, D. , Reitsma, M.J. , Masek, M. , Phan, T.V. , 2000. Direct isolation of human central nervous system stem cells. Proc. Natl. Acad. Sci. U. S. A. 97, 14720–14725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Ventura, J.J. , Tenbaum, S. , Perdiguero, E. , Huth, M. , Guerra, C. , Barbacid, M. , 2007. p38alpha MAP kinase is essential in lung stem and progenitor cell proliferation and differentiation. Nat. Genet. 39, 750–758. [DOI] [PubMed] [Google Scholar]
- Visvader, J.E. , Lindeman, G.J. , 2008. Cancer stem cells in solid tumours: accumulating evidence and unresolved questions. Nat. Rev. Cancer. 8, 755–768. [DOI] [PubMed] [Google Scholar]
- Wistuba, Gazdar, A.F. , Minna, J.D. , 2001. Molecular genetics of small cell lung carcinoma. Semin. Oncol. 28, 3–13. [PubMed] [Google Scholar]
- Yanagi, S. , Kishimoto, H. , Kawahara, K. , Sasaki, T. , Sasaki, M. , Nishio, M. , 2007. Pten controls lung morphogenesis, bronchioalveolar stem cells, and onset of lung adenocarcinomas in mice. J. Clin. Invest. 117, 2929–2940. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yang, Y. , Iwanaga, K. , Raso, M.G. , Wislez, M. , Hanna, A.E. , Wieder, E.D. , 2008. Phosphatidylinositol 3-kinase mediates bronchioalveolar stem cell expansion in mouse models of oncogenic K-ras-induced lung cancer. PLoS One. 3, e2220 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yesner, R. , 2001. Heterogeneity of so-called neuroendocrine lung tumors. Exp. Mol. Pathol. 70, 179–182. [DOI] [PubMed] [Google Scholar]
- Zhang, Y. , Goss, A.M. , Cohen, E.D. , Kadzik, R. , Lepore, J.J. , Muthukumaraswamy, K. , 2008. A Gata6-Wnt pathway required for epithelial stem cell development and airway regeneration. Nat. Genet. 40, 862–870. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Zandwijk, N. , Mooi, W.J. , Rodenhuis, S. , 1995. Prognostic factors in NSCLC. Recent experiences. Lung Cancer. 12, (Suppl. 1) S27–S33. [DOI] [PubMed] [Google Scholar]