

Abstract
Glyphosate has been used for more than 15 years for weed management in citrus groves in the Gulf of Mexico, at up to 3–4 applications per year. Goosegrass (Eleusine indica (L.) Gaertn.) control has sometimes failed. In this research, the mechanisms governing three goosegrass biotypes (Ein-Or from an orange grove, and Ein-Pl1 and Ein-Pl2 from Persian lime groves) with suspected resistance to glyphosate were characterized and compared to a susceptible biotype (Ein-S). Dose-response and shikimate accumulation assays confirmed resistance of the resistant (R) biotypes. There were no differences in glyphosate absorption, but the R biotypes retained up to 62–78% of the herbicide in the treated leaf at 96 h after treatment (HAT), in comparison to the Ein-S biotype (36%). The 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS) activity in the Ein-Or and Ein-S biotypes was over 100-fold lower than the Ein-Pl1 and Ein-Pl2 ones. The latter showed a high EPSPS-basal activity, a mutation at Pro-106-Ser position in the EPSPS gene, and EPSPS overexpression. The EPSPS basal and EPSPS overexpression were positively correlated. The R goosegrass biotypes displayed poor glyphosate translocation. Furthermore, this grassweed showed, for the first time, two mechanisms at the target-site level (Pro-106-Ser mutation + EPSPS overexpression) acting together simultaneously against glyphosate.
Introduction
Goosegrass (Eleusine indica (L.) Gaertn.), an annual-C4 summer grass plant, is one of the world’s worst weeds. It comes from Africa and is widely spread over tropical, subtropical regions and temperate regions of the world1, 2. Goosegrass is a problematic weed mainly on lawns, in annual crops and in fruit orchards3, 4. In Mexico, goosegrass is widespread in 25 states5. Depending on the crop, different herbicides have been recommended for goosegrass control6. However, herbicide use as the primary weed management approach has resulted in goosegrass resistance to different herbicide groups, such as acetohydroxyacid synthase (ALS) inhibitors, acetyl-CoA carboxylase (ACCase) inhibitors, photosystem I and II (PSI and PSII) inhibitors, microtubule inhibitor, glutamine synthetase and 5-enolpyruvylshikimate-3-phosphate synthase (EPSPS; EC 2.5.1.19) inhibitors7. Even one biotype from Malaysia showed multiple resistance to three non-selective herbicides, glufosinate, glyphosate and paraquat8.
Glyphosate (N-phosphonomethyl-glycine) is a post-emergence, non-selective and systemic herbicide that is used extensively to control a wide spectrum of weeds in orchards, soil conservation systems and non-cropped (fallow) areas9. Glyphosate inhibits EPSPS, disrupting the biosynthesis of phenylalanine, tyrosine, tryptophan and others secondary aromatic products10. This is due to the complex effects caused by glyphosate on the shikimate pathway, the photosynthetic processes and the oxidative events of plants11, triggering the expression of other chloroplast enzymes12. Recently, the expression of the phosphofructokinase (PFK: EC 2.7.1.11) enzyme has been strongly associated with EPSPS expression13.
The widespread reliance on glyphosate for weed control in recent decades, and the introduction of glyphosate-resistant crops has resulted in a large number of weeds becoming resistant to this herbicide7, 14. Glyphosate resistance mechanisms can be categorized into two classes: target site resistance (TSR) and non-target site resistance (NTSR)15. TSR mechanisms are a result of mutations during encoding of the EPSPS gene1, 4, 13, 15, 16, overexpression of the EPSPS enzyme4, 13, 17, and/or enhanced basal EPSPS activity17, 18. NTSR mechanisms are due to reduced absorption and translocation18–21, vacuolar sequestration22, and metabolism of the herbicide into non-toxic compounds23.
The state of Veracruz, on the Gulf of Mexico, is the main citrus fruit producing region in Mexico, with Persian lime being the most important crop because of its large production and its economic value24. In addition to mowing, few other approaches or herbicides are employed to manage weeds in citrus production systems (Dominguez-Valenzuela, personal communication) other than glyphosate25. To date, Bidens pilosa 16 and Leptochloa virgata 17 have been identified as resistant to this herbicide in citrus groves in the states of Veracruz and Puebla. However, weed control with glyphosate has also failed for other weeds in the citrus production systems of this region.
The objectives of this study were as follows: 1) to confirm resistance of goosegrass to glyphosate; and 2) to study the resistance mechanisms involved in the resistant biotypes collected from citrus fruit groves in Mexico.
Results
Dose-response
The survival and fresh weight of the four goosegrass biotypes decreased with increasing glyphosate doses (Fig. 1). The three suspected resistant (R) biotypes collected from the citrus-growing region of Veracruz, Mexico [Ein-Pl1 and Ein-Pl2 biotypes from Persian lime (Citrus latifolia Tan.) groves and Ein-Or biotype from an orange (Citrus sinensis Osbeck) grove]; presented resistance to glyphosate. Therefore, the glyphosate doses used to reduce the fresh weight (GR50) and to kill a plant population (LD50) by 50% of the R biotypes ranged from 136 to 556, and from 974 to 1938 g per ha, respectively. According to these parameters, the resistance indexes (RI) ranged between 2.6 and 15.9. LD90, and GR90 corroborated the resistance of these biotypes (Table 1).
Figure 1.
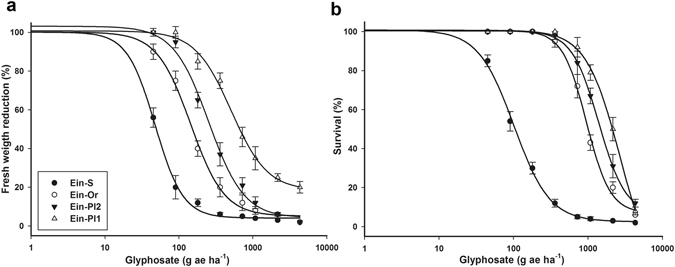
Log–logistic curves of glyphosate -susceptible and -resistant goosegrass (Eleusine indica) biotypes collected in citrus groves in Veracruz, Mexico, evaluated at 21 DAT. (a) Dose-response curve with respect to percentage of fresh mass reduction. (b) Dose-response curve with respect to percentage of survival. Vertical bars represent the standard error of the mean (n = 10).
Table 1.
Parameters of the log-logistic equationa used to calculate the glyphosate dose (g ae per ha) required to reduce the fresh weight (GR) and survival percentage (LD) by 50 or 90% of the glyphosate-resistant and -susceptible goosegrass (Eleusine indica) biotypes collected in citrus groves from Veracruz, Mexico.
| Fresh weight reduction (GR) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Biotypes | c | d | b | GR 50 | CI95% b | RI c | P | GR 90 | CI95% b | RI c | P |
| Ein-Pl1 | 2.34 | 101.90 | 1.54 | 555.6 | 46.6 | 10.4 | 0.0001 | 2297.4 | 267.8 | 20.5 | 0.0038 |
| Ein-Pl2 | 7.17 | 102.07 | 1.97 | 267.1 | 24.9 | 5.0 | 0.0001 | 811.0 | 147.3 | 7.2 | 0.0001 |
| Ein-Or | 2.92 | 102.49 | 2.04 | 137.5 | 13.4 | 2.6 | 0.0001 | 418.9 | 72.0 | 3.7 | 0.0006 |
| Ein-S | 0.93 | 100.65 | 2.91 | 53.4 | 7.6 | — | — | 111.9 | 15.8 | — | — |
| Survival percentage (LD) | |||||||||||
| Biotypes | c | d | b | LD 50 | CI95% b | RI c | P | LD 90 | CI95% b | RI c | P |
| Ein-Pl1 | 0.12 | 99.45 | 3.69 | 1938.3 | 366.1 | 15.9 | 0.0001 | 3956.0 | 521.4 | 18.9 | 0.0001 |
| Ein-Pl2 | 0.62 | 100.08 | 4.96 | 1442.6 | 244.3 | 11.9 | 0.0001 | 2244.5 | 396.0 | 10.7 | 0.0001 |
| Ein-Or | 2.72 | 100.15 | 2.99 | 973.8 | 184.3 | 8.0 | 0.0001 | 2028.7 | 337.6 | 9.7 | 0.0001 |
| Ein-S | 0.57 | 100.40 | 2.89 | 121.7 | 23.7 | — | — | 209.2 | 33.8 | — | — |
a Y = c + {(d − c)/[1 + (x/g)b]} where; Y is the percentage of fresh weight and/or survival with respect to the control, c and d are the the lower and upper asymptotes, b is the slope of the curve at the inflection point, g the herbicide dose at the inflection point (i.e. GR50 or LD50), and x (independent variable) is the glyphosate dose. bCI values are the upper and lower limits (±) of the 95% confidence intervals (n = 10). cRI = Resistance index (R/S) calculated using the corresponding ED50, or LD50 values of the resistant biotype with respect to the susceptible one.
Shikimic acid accumulation
Both the R and S biotypes of goosegrass accumulated shikimic acid as the glyphosate concentrations increased. The greater accumulation of shikimic acid (approximately 160 µg g−1 fresh weight) was exhibited by the Ein-S biotype. The R biotypes accumulated between 3.3 and 8.1 times less shikimic acid at 1000 µM glyphosate, with the lowest amounts accumulated by the Ein-Pl1 and Ein-Pl2 biotypes (Fig. 2).
Figure 2.
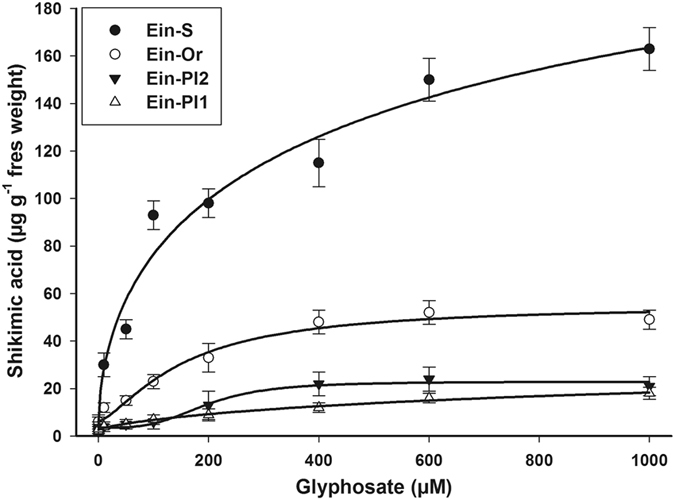
Shikimic acid accumulation of the glyphosate -susceptible and -resistant goosegrass (Eleusine indica) biotypes collected in citrus groves in Veracruz, Mexico, at different glyphosate concentrations. Vertical bars represent the standard error of the mean (n = 3).
Absorption and translocation
Foliar 14C-glyphosate absorption showed no differences between goosegrass biotypes. At 12 h after treatment (HAT), the 14C-glyphosate absorption rate ranged from 14.3 to 20.1%, and from 58.4 to 66.2% at 96 HAT (Fig. 3).
Figure 3.
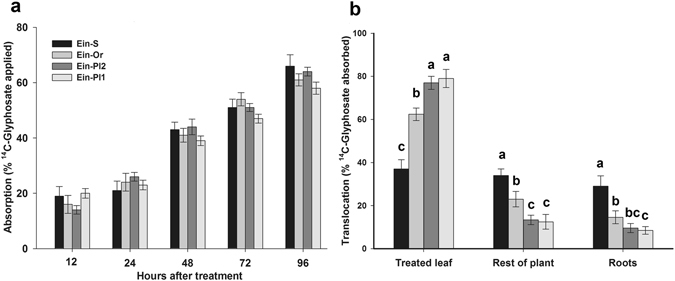
14C-Glyphosate absorption and translocation in glyphosate-susceptible and -resistant plants of goosegrass (Eleusine indica) biotypes collected in citrus groves in Veracruz, Mexico. (a) 14C-glyphosate absorption in goosegrass plants from 12 to 96 HAT. (b) 14C-glyphosate translocation in goosegrass plants at 96 HAT. Different letters are significantly different at 95% probability determined by Tukey’s test. The vertical bars represent the standard error of the mean (n = 5).
However, the 14C-glyphosate translocation exhibited differences between 12 to 96 HAT. The Ein-S biotype presented the highest translocation rate at 96 HAT, moving most of the absorbed glyphosate from the treated leaf to other plant parts (33.8 and 29.3% to the rest of the plant and roots, respectively). Conversely, the R biotypes retained a larger amount of herbicide in the treated leaf, with 62.4, 76.3 and 79.6% of the glyphosate absorbed by the Ein-Or, Ein-Pl2 and Ein-Pl1 biotypes, respectively, evaluated simultaneously. The R biotype Ein-Or translocated slightly more glyphosate to the rest of the plant and roots than the others two R biotypes (Fig. 3).
Glyphosate metabolism
More than 90% of the applied glyphosate was not metabolized by the goosegrass plants at 4 DAT. At this time, small amounts of AMPA (aminomethyl phosphonate) (36.−6.6%) and glyoxylate (1.6–2.4%) were detected in all goosegrass biotypes. A similar amount of metabolized glyphosate in the R and S biotypes, as well as the non-detection of sarcosine, suggested that metabolism is not a resistance mechanism (Table 2).
Table 2.
Glyphosate metabolism expressed as percentage of total glyphosate and their metabolites in the glyphosate-resistant and -susceptible goosegrass (Eleusine indica) biotypes collected in citrus groves from Veracruz, Mexico.
| Biotypes | glyphosate | AMPA | glyoxylate | sarcosine |
|---|---|---|---|---|
| Ein-Pl1 | 93.2 ± 2.6 | 4.6 ± 1.4 | 2.1 ± 0.6 | ND |
| Ein-Pl2 | 91.7 ± 2.3 | 6.6 ± 2.2 | 1.7 ± 1.1 | ND |
| Ein-Or | 92.8 ± 5.9 | 4.8 ± 1.8 | 2.4 ± 1.2 | ND |
| Ein-S | 95.3 ± 4.1 | 3.6 ± 0.9 | 1.6 ± 0.4 | ND |
| P | 0.3765 | 0.1395 | 0.0672 | ND |
Values represent mean (n = 6). ±Standard error. AMPA aminomethyl phosphonate. ND not detected.
EPSPS enzyme activity
Significant differences were found in the basal EPSPS activity. The Ein-Pl1 biotype presented the highest basal activity (0.35 μmol of phosphate released per μg total soluble protein (TSP)−1 min−1), whereas the Ein-Pl2, Ein-Or and Ein-S biotype averages were 0.20, 0.03 and 0.04 of μmol μg TPS−1 min−1, respectively (Fig. 4).
Figure 4.
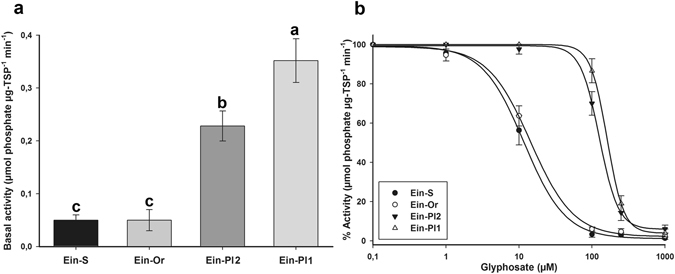
EPSPS activity in glyphosate-susceptible and -resistant plants of goosegrass (Eleusine indica) biotypes collected in citrus groves in Veracruz, Mexico. (a) Basal EPSPS activity in glyphosate -susceptible and -resistant goosegrass plants. Histograms represent the treatment means and vertical bars ± standard error (n = 3). (b) EPSPS enzyme activity expressed as a percentage of the untreated control in leaf extracts of plants from glyphosate-susceptible and resistant goosegrass biotypes. Vertical bars represent ± standard error (n = 3).
The EPSPS enzyme was inhibited by glyphosate in the S and R biotypes. The glyphosate concentration that inhibited EPSPS activity by 50% (I50) in the Ein-S biotype was 2.8 µM. With respect to this parameter, the RIs of Ein-Or, Ein-Pl2 and Ein-Pl1 biotypes were 1.2, 10.8 and 13.8, respectively, compared to the S biotype. There were no differences in the EPSPS activity between the Ein-Or and Ein-S biotypes (Table 3).
Table 3.
Parameters of the log-logistic equationa used to calculate the glyphosate concentration dose (µM) required to reduce the EPSPS activity reduction (I50) by 50% of the glyphosate-resistant and -susceptible goosegrass (Eleusine indica) biotypes collected in citrus groves from Veracruz, Mexico.
| Biotypes | c | d | b | I50 | CI95%b | RIc | P |
|---|---|---|---|---|---|---|---|
| Ein-Pl1 | 3.91 | 100.00 | 3.82 | 161.1 | 18.8 | 13.8 | <0.0001 |
| Ein-Pl2 | 5.90 | 99.42 | 3.37 | 125.9 | 14.6 | 10.8 | <0.0001 |
| Ein-Or | 2.14 | 98.93 | 1.48 | 14.4 | 2.8 | 1.2 | <0.0001 |
| Ein-S | 1.07 | 99.37 | 1.53 | 11.7 | 3.3 | — | <0.0001 |
a Y = c + {(d − c)/[1 + (x/g)b]} where; Y is the percentage of EPSPS activity with respect to the control, c and d are the lower and upper asymptotes, b is the slope of the curve at the inflection point, I50 the herbicide dose at the inflection point, and x (independent variable) is the glyphosate concentration. bCI values are the upper and lower limits (±) of the 95% confidence intervals (n = 3). cRI = Resistance index = I50R/I50S.
EPSPS gene sequence
The amplified fragment of the EPSPS gene included Thr-102 and Pro-106 positions, the point mutations conferring glyphosate resistance, presenting a homology of above 98% between goosegrass biotypes. The obtained EPSPS sequences were aligned with the GenBank accessions (R-J417033 and S-J417034) of goosegrass that were reported by Baerson et al.1. In the Thr-102 position, no mutation was found. At the Pro-106 position, the Ein-Pl1 and Ein-Pl2 biotypes exhibited a codon change from CCA to TCA, resulting in an amino acid substitution from Proline to Serine (Table 4).
Table 4.
Partial alignment of nucleotides of EPSPS gene of the glyphosate-resistant and -susceptible goosegrass (Eleusine indica) biotypes collected in citrus groves from Veracruz, Mexico.
| Positiona | 99 | 100 | 101 | 102 | 103 | 104 | 105 | 106 | 107 |
|---|---|---|---|---|---|---|---|---|---|
| Accession b | |||||||||
| R-J417033 | AAT | GCT | GCA | ACT | GCA | ATG | CGA | TCA c | TTG |
| S-J417034 | AAT | GCT | GCA | ACT | GCA | ATG | CGA | CCA | TTG |
| Biotype | |||||||||
| Ein-Pl1 | AAT | GCT | GCA | ACT | GCA | ATG | CGA | TCA | TTG |
| Ein-Pl2 | AAT | GCT | GCA | ACT | GCA | ATG | CGA | TCA | TTG |
| Ein-Or | AAT | GCT | GCA | ACT | GCA | ATG | CGA | CCA | TTG |
| Ein-S | AAT | GCT | GCA | ACT | GCA | ATG | CGA | CCA | TTG |
aAmino acid position based on the start codon (ATG) of Arabidopsis thaliana (GenBank: CAA29828.1) EPSPS sequence. bGenbank accession reported by Baerson et al.1. cCodon change from CCA to TCA resulting in an amino acid substitution from Proline to Serine that confers resistance to glyphosate.
EPSPS gene expression
The EPSPS gene expression quantified from cDNA in the goosegrass biotypes was determined before and after glyphosate application. EPSPS gene expression relative to the β-Actin gene before treatment (control) ranged from 0.11 to 0.28, 0.10 to 0.25, 1.09 to 1.36 and 1.90 to 2.58 for the Ein-S, Ein-Or, Ein-Pl2 and Ein-Pl1 biotypes, respectively. After glyphosate treatment, the EPSPS expression level ranged between 0.23 to 0.54 for the Ein-S and Ein-Or biotypes, and 5.18–6.73 and 3.22–4.48 for the Ein-P1 and Ein-Pl2 biotypes, respectively. Furthermore, the EPSPS expression level of the control was positively correlated with the EPSPS basal activity (Fig. 5).
Figure 5.
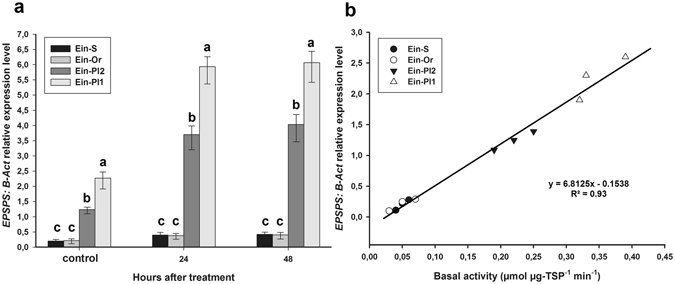
EPSPS gene expression relative to β-Actin for glyphosate -susceptible and -resistant plants of goosegrass (Eleusine indica) biotypes. (a) EPSPS expression level for glyphosate-susceptible and -resistant goosegrass plants before and after glyphosate treatment. Histograms represent the treatment means and vertical bars ± standard error (n = 6). (b) correlation between the EPSPS expression (control) and EPSPS basal activity.
Discussion
Determining resistance
Glyphosate resistance was confirmed in three goosegrass biotypes, even in the Ein-Or biotype from the orange grove, which had the lowest GR50 values, because the glyphosate field dose (720 g per ha) used in citrus groves from Veracruz was not enough to achieve full control, because this biotype presented a LD50 of 973.8 g per ha. This is consistent with the fact that producers care more about weed management in Persian lime groves than in orange groves due to the greater economic profitability of limes, which induces more frequent glyphosate use in these groves. Outside the flowering and fruit set seasons, eventually 2,4-D is applied in the mixture with lower glyphosate doses to improve the control of broadleaf weeds (Domínguez-Valenzuela, personal communication). However, this practice has not reduced the selection pressure exerted by glyphosate and, unfortunately, these results confirm a third case of glyphosate resistance in the citrus region of the Gulf of Mexico, after identification of L. virgata in 201025 and B. pilosa in 201416. The RIs estimated based on GR50 and LD50 parameters, are in agreement with the RIs estimated for other biotypes/populations of goosegrass2–4, 26–28.
The amount of shikimic acid accumulated by goosegrass R biotypes indicated that the selection pressure exerted by glyphosate in these biotypes causes scant sensitivity to this herbicide. Differential accumulation of shikimic acid occurs when glyphosate does not reach the target-site in sufficient amounts, inhibiting EPSPS in different proportions17. For glyphosate to be lethal in plants with a low shikimic acid accumulation, larger amounts of herbicide are required16. The different shikimic acid accumulation patterns between R and S goosegrass biotypes were similar to those observed in the dose-response assays, indicated high glyphosate susceptibility of the S biotype and a different resistance level of R biotypes. This suggests that the mechanisms of resistance involved in the R biotypes are different from each other. The shikimic acid accumulation has been widely reported as discriminating resistance to glyphosate4, 23, 29–31.
Exploring the NTSR mechanism involved
The absorption of glyphosate is influenced by leaf morphological and physiological traits32, which has been suspected to occur through active transporters and by passive diffusion down a concentration gradient in cells12. Goosegrass biotypes presented similar 14C-glyphosate absorption patterns over time after application. This suggests that the R and S goosegrass biotype leaf traits have not been altered by glyphosate selection pressure, thereby allowing absorption in the same proportion. Therefore, absorption is not implicated in the resistance to glyphosate of R goosegrass biotypes. This mechanism is not usually involved in glyphosate resistance, and only some grassweed species, such as Digitaria insularis 23, L. virgata 18, Lolium multiflorum 20 and Sorghum halepense 21, have registered differences in their absorption patterns.
Glyphosate is a systemic herbicide and must reach the enzymatic sites of root and shoot meristems in order to act10. Any reduction in its translocation to these active and sensitive sites has a negative effect on its efficiency33. R goosegrass biotypes (Ein-Or, Ein-Pl1 and Ein-Pl2) exhibited less glyphosate translocation to the rest of the plant and roots than the S biotype (Ein-S), retaining it mainly within the treated leaves. The exact mechanism that reduces glyphosate translocation is not fully known, but a possible change in the plasma membrane transporters that restricts the glyphosate movement inside of resistant plants19 has been considered. It has also been suggested that there is an unknown barrier altering the subcellular distribution of glyphosate and keeping it away from the target site34. This barrier may exist either in the phloem system or in the mesophyll cells, where glyphosate has to enter to be translocated35. The Ein-Or biotype was selected by glyphosate less often than the Ein-Pl1 and Ein-Pl2 biotypes. This explains why its translocation rate is higher with respect to the latter biotypes but was significantly lower than the Ein-S biotype. Therefore, differential translocation of the herbicide can be considered one of the glyphosate resistant mechanisms in these R goosegrass biotypes. Similar restrictions in glyphosate translocation were reported for different grassweed species, such as the following: D. insularis 23 , Lolium multiflorum 36 , L. perenne 37, L. rigidum 30 , Leptochloa virgata 18, S. halepense 21, among others.
On the other hand, glyphosate metabolism has been verified as not involved in the resistance to glyphosate in R goosegrass biotypes. Degradation of glyphosate has been documented in a few weed species, such as the following: D. insularis 23 and Parthenium hyterophorus 38. Although metabolism could influence resistance to glyphosate, there is no evidence that it plays an important role in it39, and it has been rarely studied or reported as endowing resistance to glyphosate15. Our results were consistent with other studies in B. pilosa 16, C. canadensis 40 , Echinoclhoa colona 17 and L. virgata 18, which demonstrated that this mechanism does not contribute to glyphosate resistance.
Exploring the TSR mechanism involved
The goosegrass biotypes of Ein-S and Ein-Or presented similar basal EPSPS levels and glyphosate response patterns. This indicates that the R biotype Ein-Or is susceptible to glyphosate at a target-site level. The EPSPS enzyme of glyphosate R plants could be sensitive to glyphosate when the EPSPS gene had no mutations18, 41. This explains why the Ein-Or biotype had similar susceptibility to glyphosate as that of the Ein-S biotype, suggesting that this R biotype does not employ target-site mechanisms against this herbicide. On the other hand, the Ein-Pl1 and Ein-Pl2 biotypes presented high basal EPSPS activity, and larger amounts of glyphosate were required to inhibit EPSPS activity by 50%. Basal activity differences are associated with a greater EPSPS gene amplification, and consequently, the overexpression of its enzyme42. This implies that, in addition to reduced translocation, Ein-Pl1 and Ein-Pl2 biotypes presented at least one target-site mechanism against glyphosate. Different R goosegrass populations from China4, 13, and other glyphosate resistant grassweed species have displayed high levels of basal EPSPS activity compared to their respective susceptible ones17, 42, and higher glyphosate concentrations were necessary to inhibit the EPSPS activity by 50%.
EPSPS gene sequencing demonstrated that the Ein-Pl1 and Ein-Pl2 biotypes possessed a mutation at Pro-106 position. Amino acid substitution in this genomic EPSPS position from Proline to Serine, Alanine, Threonine and/or Leucine, has been widely reported in mono- and dicotyledonous weeds, endowing partial resistance to glyphosate, in some cases accompanied by a NTSR mechanism15. Some grassweed species, which have shown a mutation in combination with other mechanism (TSR + NTSR) in the last two years, are C. virgata 31 , E. colona 17 , L. virgata 18, L. rigidum 30 and Poa annua 29. Recently, the double mutation commonly known as TIPS (Thr-102-Ile + Pro-106-Ser), has been documented in glyphosate resistant populations of goosegrass4, 26, and B. pilosa 16, endowing them with a high degree of resistance.
EPSPS expression in cDNA was differed in the R goosegrass plants of the Ein-Pl1 and Ein-Pl2 biotypes with respect to plants of the Ein-S and Ein-Or biotypes. The EPSPS expression of the R and S biotypes before glyphosate treatment (control) was strongly and positively correlated with their EPSPS basal activity, indicating that the EPSPS expression was proportionate to the amount of transcribed EPSPS enzyme. There is a positive correlation between the EPSPS gene copy number and the EPSPS transcription43. It is possible that the goosegrass biotypes have different EPSPS genomic copy numbers from each other. However, the EPSPS gene copy numbers were not studied in the goosegrass biotypes, because the manifestation of more EPSPS gene copies does not always result in higher protein transcription44. Furthermore, an increase in EPSPS expression was observed after treatment in response to the stress exerted by glyphosate. This level of expression remained constant at 24 and 48 HAT. Similar results were observed in R goosegrass populations from China4, where the EPSPS expression levels from 6 to 72 HAT treated at 922.5 g per ha were evaluated and remained constant after application. The EPSPS expression level depends more on the dose of glyphosate applied than the time period evaluated. Goosegrass (R and S) plants exhibited a gradual increase in the EPSPS expression with increased concentrations of glyphosate2. Some resistant grassweed species presenting EPSPS copy numbers and/or overexpression as glyphosate resistance mechanisms are as follows: B. diandrus 45 , Kochia scoparia 44 and L. perenne 42, 46. These results indicate that the Ein-Pl1 and Ein-Pl2 goosegrass biotypes from Persian lime groves have evolved a mutation and overexpression of the target site, which gives them greater resistance to glyphosate due to the different selection pressure exerted by this herbicide in these groves compared to the orange groves.
It is important to note that, so far, known cases of glyphosate resistance involving the target-site were only due to point mutations (the single Pro-106 mutation1, 4, 16–18, 26, 29–31, or the double Thr-102 + Pro-106 one4, 16, 26) or to EPSPS amplification2, 4, 42–46, but never both TSR mechanisms acting together simultaneously against this herbicide, except in Chloris truncata 47. Ngo et al.47 suggested that the Glu-91-Ala mutation (position 86 in Escherichia coli) in a conserved region of the EPSPS gene contributed to the glyphosate resistance in C. truncata. However, the highly conserved region of the EPSPS gene was comprised of amino acid 90 to 102 in E. coli (from 95 to 107 in plants)48. Furthermore, Ngo et al.47 pointed to widely known mutations, induced in E. coli and plants, which may contribute to the glyphosate resistance, and they indicated that the Glu-91-Ala mutation had not been previously described as conferring resistance to this herbicide. Consequently, we suggest that the only TSR mechanism in C. truncata was the amplification of EPSPS. Therefore, in the case of the glyphosate resistance reported here, this is the first case of a weed involving both a mutation responsible for conferring resistance (Pro-106-Ser) as well as the overexpression of the EPSPS gene.
Goosegrass presents a great adaptive capacity to develop herbicide resistance, making it one of the most harmful weeds in agriculture; this weed was the first to achieve the following: evolved single Pro-1061 and double Thr-102-Ile + Pro-106-Ser4, 16 mutations that confer resistance to glyphosate; exhibited multiple resistance to non-selective herbicides8, and the current methodologies do not allow determination of the NTRS and TSR mechanisms involved in their resistance to glufosinate49; and now we demonstrated that goosegrass can be expressed by two mechanisms simultaneously at the target site level against glyphosate.
Conclusion
Reduced translocation is one mechanism of resistance to glyphosate that has evolved in resistant goosegrass biotypes from Mexico. Greater selection pressure exerted by glyphosate triggered a mutation at the Pro-106 position + overexpression of the EPSPS gene in the resistant goosegrass biotypes in Persian lime groves. These results confirm the first case of glyphosate resistance of this species in the country, as well as the third case of resistance to this herbicide in Mexico’s citrus groves.
Materials and Methods
Plant Material
Within a citrus fruit region in the municipality of Martinez de la Torre, in the state of Veracruz, Mexico, seeds of fully mature goosegrass plants of two suspect R biotypes (Ein-Pl1: N 20°10′34, W 97°07′36; and Ein-Pl2; N 20°10′32, W 97°07′37) were collected in Persian lime groves and one (Ein-Or: N 20°10′28, W 97°07′12) from an orange grove. A susceptible biotype (Ein-S: N 20°10′17, W 97°06′47) without history of glyphosate application also was also collected in a land urban as control. The citrus fruit groves had a history of glyphosate usage of over 15 years with 3–4 and 1–2 applications per year of 720 g per ha in Persian lime and orange, respectively.
The seeds were germinated in peat, and the seedlings were transplanted individually into 250 mL pots with a substrate mixture of sand and peat (1: 1, v/v). The plants were placed in a greenhouse at a temperature regime of 26/18 °C day/night, and were treated with three-four true leaves counted from the bottom.
Dose response assay
Different doses of glyphosate (Roundup EnergyPro 45% p/v [potassium salt]; Monsanto, Spain) were made with a Generation III Research Track Sprayer (DeVries Manufacturing Inc., Minnesota, USA) equipped with a flat-fan nozzle (Tee Jet 8002EVS) delivering 200 L ha−1 at 200 kPa. The glyphosate doses applied were: 0, 45, 90, 180, 360, 720, 1080, 2160, and 4320 g per ha. The experiments were arranged in a completely random design with 10 replications per dose and repeated twice. Survival and fresh weights of plants were measured at 21 DAT. Data were expressed as percent with respect to untreated control plants.
Shikimic assay
Young leaf discs 4 mm in diameter were taken until completing 50 mg of plant tissue from plants of the goosegrass biotypes. Shikimic acid accumulation was determined according to Shaner et al.50. The glyphosate concentrations used were: 0, 0.1, 1, 10, 50, 100, 200, 400, 600 and 1000 µM. Sample absorbance was measured in a Beckman DU-640 spectrophotometer at 380 nm. The test was performed in triplicate on five treated and non-treated plants of each biotype in a completely random design. Results were expressed in mg of shikimic acid g−1 fresh tissue.
14C-glyphosate absorption and translocation
14C-glyphosate (American Radiolabeled Chemicals, Inc., USA) was added to the commercial herbicide to prepare a solution with a specific activity of 0.834 kBq μL−1. The final glyphosate concentration corresponded to 360 g ae per ha in 200 L ha−1. Goosegrass plants were treated (0.834 kBq plant−1) at 12, 24, 48, 72 and 96 HAT. Five plants per biotype at each time evaluated in a completely random design were handled according to Bracamonte et al.38. Radioactivity was analyzed by liquid scintillation spectrometry (LSS) in a Beckman LS 6500 scintillation counter (Beckman Coulter Inc. Fullerton, USA) during 10 min per sample. The average of 14C recovered (SE) was 93.6 (4.3) %. Percentage of 14C-glyphosate absorbed was expressed as [kBq in combusted tissue/(kBq in combusted tissue + kBq in leaf washes)] × 100.
Metabolism
Six plants of each biotype were treated with 360 g ae per ha of glyphosate (as described in the dose-response assays) in a completely randomized design. Untreated plants were used as controls. Leaf tissues were washed with distilled water at 4 DAT, flash-frozen in liquid nitrogen, and stored at −40 °C until use. Following the methodology described by Rojano-Delgado et al.51, glyphosate and its metabolites (aminomethyl phosphonate (AMPA), glyoxylate, sarcosine and formaldehyde) were determined by reversed polarity capillary electrophoresis using a 3D Capillary Electrophoresis Agilent G1600A instrument equipped with a diode array detector (DAD, wavelength range 190–600 nm). Standard compounds used (glyphosate, AMPA, sarcosine, formaldehyde, and glyoxylate), were provided by Sigma-Aldrich, Spain. Glyoxylate naturally produced (untreated plants) was subtracted from the average of glyoxylate produced from glyphosate metabolism (treated plants) for each biotype.
EPSPS enzyme assays
Five g of the leaf tissue of each biotype plants was ground to fine powder in liquid nitrogen in a chilled mortar. The enzyme extraction was conducted according to the protocol described by Sammons et al.52. The total soluble protein (TPS) in the extract was measured using a Modified Lowry Kit for Protein Determination (Sigma-Aldrich, Madrid, Spain) in accordance with the manufacturer’s instructions. The specific EPSPS activity was determined using the EnzCheck Phosphate Assay Kit (Invitrogen, Carlsbad, CA) following the manufacturer’s instructions, to determine the inorganic phosphate release. The glyphosate concentrations used were: 0, 0.1, 1, 10, 100, 200 and 1000 µM. The EPSPS activity was measured during 10 minutes at 360 nm in a spectrophotometer (Beckman DU-640) to determine the amount of phosphate (µmol) released µg of TSP−1 min−1 and expressed as a percentage with respect to the control (without glyphosate). The experiment was repeated three times for all samples.
EPSPS gene sequence
Samples of young leaf tissue (≈100 mg) from 10 individual plants within each goosegrass biotype were ground to a fine powder in liquid nitrogen. Genomic DNA (gDNA) was extracted using Speedtools plant DNA extraction kit (Biotools, Madrid, Spain). Integrity of DNA was verified in 0.8% agarose gel and quantified in a NanoDrop ND-1000 spectrophotometer (Thermo Scientific, Walthman, MA, USA), and diluted to a final concentration of 50 ng 1 µL−1 and at −20 °C until use. The primers (forward: 5′-GGTGGATAACCTTTTAAACAGTGAG-3′; and reverse: 5′-TTAGTTCTTGACGAAAGTGCTGAGC-3′) designed by Chen et al.4, were used to amplify a fragment of 1300 pb in length, and the PCR conditions described by them were followed. An aliquot of the PCR product was loaded in a 1% agarose gel to check the correct band amplification. Then, the PCR product was purified using ExoSAP-IT® for PCR Product Clean-Up (USB, Ohio, USA) as indicated by the manufacturer. Fifteen µL of purified PCR product per sample was sequenced (STAB VIDA, Caparica, Portugal). Each individual was sequenced in triplicate. The resulting fragments were aligned with known EPSPS sequences of goosegrass from the GenBank.
EPSPS gene expression
Six untreated plants with 5–6 true leaves of each biotype were sampled (one expanded leaf) before glyphosate treatment. Then, plants were treated with 360 g ae per ha of glyphosate (as previously described in the dose-response assays), and plant were sampled again at 24 and 48 HAT. Samples were immediately stored at −80 °C. RNA was extracted using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. RNA was then treated with TURBO DNase (RNase-Free; Ambion, Warrington, UK) to eliminate any DNA contamination. Integrity of RNA was verified in 0.8% agarose gel and quantified in a NanoDrop ND-1000. First strand complementary DNA (cDNA) synthesis was carried out using one µg of RNA in all the samples. An iScript cDNA Synthesis Kit (Bio-Rad Laboratories, Inc. CA, USA) was employed following the manufacturer’s instructions. For qPCR analyses, the EPSPS (GF: 5′-CTGATGGCTGCTCCTTTAGCTC-3′; and GR: 5′-CCCAGCTATCAGAATGCTCTGC-3′), and β-Actin (F1a: 5′-ATGGTAGGGATGGGACAGAA-3′; and R1a: 5′-TCCATGTCATCCCAGTTGCT-3′) primers designed by Chen et al.4 and Alcántara-de la Cruz et al.18, respectively were used. The PCR reactions were carried out using an ABI Prism 7500 sequence detection system (Applied Biosystems, USA). Quantitative RT-PCR conditions and EPSPS gene expression analyzes were performed according to Alcántara-de la Cruz et al.18.
EPSPS expression level was calculated for each qPCR reaction. The PCR efficiency of each biotype sample and the stability of the β-Actin, as a reference gene, were determined using geNorm software according to Vandesompele et al.53. Three-four technical replications per plant were analyzed in a randomized design.
Statistical analysis
Dose-response and EPSPS enzyme activity data were subjected to non-linear regression analysis using a log-logistic equation: Y = c + {(d − c)/[1 + (x/g)b]}; where Y is the percentage of fresh weight, survival and/or EPSPS-inhibiting with respect to the control, c and d are the parameters corresponding to the lower and upper asymptotes, b is the slope of the curve at the inflection point, g the herbicide rate at the inflection point (i.e. GR50, LD50 or I50), and x (independent variable) is the glyphosate dose; to find out the amount of glyphosate needed to reduce the fresh weight (GR50), mortality (LD50), and to inhibit EPSPS activity (I50) by 50% of each goosegrass biotype. Regression analyses were conducted using the drc package with program R version 3.2.554, and the data were plotted using SigmaPlot 11.0 (Systat Software, Inc., USA).
Data of absorption and translocation, metabolism and EPSPS expression were subjected to ANOVA using Statistix version 9.0 from Analytical Software (Tallahassee, FL, USA). Model assumptions of normal distribution of errors and homogeneous variance were graphically inspected. When necessary, the means were compared using Tukey’s test’s at the 95% probability level.
Acknowledgements
This work was funded by the CONACYT-231972 and -242088 (Mexico) and by the AGL2016-78944-R (Spain) projects. The authors are grateful for the assistance and technical help the Rafael A. Roldán-Gómez for your technical help.
Author Contributions
J.D.-V.: Provided the seeds used in this work. H.C.-H., J.D.-V. and R.P.: Idea and designed the experiments. J.G., P.F.-M., R.A.-C. and E.S.-G.: Performed the research. J.G., R.A.C., P.F.-M., and E.S.-G.: Analyzed the results and written the draft of this manuscript. All authors corrected and approved the manuscript.
Competing Interests
The authors declare that they have no competing interests.
Footnotes
Publisher's note: Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Contributor Information
Ricardo Alcántara-de la Cruz, Email: ricardo.la@ufv.br.
José A. Domínguez-Valenzuela, Email: jose_dv001@yahoo.com.mx
References
- 1.Baerson SR, et al. Glyphosate-resistant. Identification of a mutation in the target enzyme 5-enolpyruvylshikimate-3-phosphate synthase. Plant Physiol. 2002;129:1265–1275. doi: 10.1104/pp.001560. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Zhang C, et al. Investigating the mechanisms of glyphosate resistance in goosegrass (Eleusine indica) population from South China. J. Integr. Agr. 2015;14:909–918. doi: 10.1016/S2095-3119(14)60890-X. [DOI] [Google Scholar]
- 3.Cha TS, Anne-Marie K, Chuah TS. Identification and characterization of RAPD–SCAR markers linked to glyphosate-susceptible and -resistant biotypes of Eleusine indica (L.) Gaertn. Mol. Biol. Rep. 2014;41:823–831. doi: 10.1007/s11033-013-2922-7. [DOI] [PubMed] [Google Scholar]
- 4.Chen J, et al. Mutations and amplification of EPSPS gene confer resistance to glyphosate in goosegrass (Eleusine indica) Planta. 2015;42:859–868. doi: 10.1007/s00425-015-2324-2. [DOI] [PubMed] [Google Scholar]
- 5.Villaseñor JL, Espinosa-García FJ. The alien flowering plants of Mexico. Diversity Distrib. 2004;10:113–123. doi: 10.1111/j.1366-9516.2004.00059.x. [DOI] [Google Scholar]
- 6.Alcantara R, Fernandez P, Smeda RJ, Alves PL, De Prado R. Response of Eleusine indica and Paspalum distichum to glyphosate following repeated use in citrus groves. Crop Prot. 2016;79:1–7. doi: 10.1016/j.cropro.2015.09.027. [DOI] [Google Scholar]
- 7.Heap I. International survey of herbicide resistant weeds. http://www.weedscience.org/ (2017).
- 8.Jalaludin A, Yu Q, Powles SB. Multiple resistance across glufosinate, glyphosate, paraquat and ACCase-inhibiting herbicides in an Eleusine indica population. Weed Res. 2015;55:82–89. doi: 10.1111/wre.12118. [DOI] [Google Scholar]
- 9.Duke, S. O. The history and current status of glyphosate. Pest Manag. Sci. doi:10.1002/ps.4652 (2017). [DOI] [PubMed]
- 10.Gomes MP, et al. Alteration of plant physiology by glyphosate and its by-product aminomethylphosphonic acid: an overview. J. Exp. Bot. 2014;65:4691–4703. doi: 10.1093/jxb/eru269. [DOI] [PubMed] [Google Scholar]
- 11.Gomes MP, et al. Glyphosate-dependent inhibition of photosynthesis in willow. Front. Plant Sci. 2017;8:207. doi: 10.3389/fpls.2017.00207. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.González-Torralva F, Brown AP, Chivasa S. Comparative proteomic analysis of horseweed (Conyza canadensis) biotypes identifies candidate proteins for glyphosate resistance. Sci. Rep. 2017;7:42565. doi: 10.1038/srep42565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chen J, et al. Investigating the mechanisms of glyphosate resistance in goosegrass (Eleusine indica (L.) Gaertn.) by RNA sequencing technology. Plant J. 2017;89:407–415. doi: 10.1111/tpj.13395. [DOI] [PubMed] [Google Scholar]
- 14.Green, J. M. The rise and future of glyphosate and glyphosate-resistant crops. Pest Manag. Sci. doi:10.1002/ps.4462 (2016). [DOI] [PubMed]
- 15.Sammons RD, Gaines TA. Glyphosate resistance: State of knowledge. Pest Manag. Sci. 2014;70:1367–1377. doi: 10.1002/ps.3743. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Alcántara-de la Cruz R, et al. Target and non-target site mechanisms developed by glyphosate-resistant hairy beggarticks (Bidens pilosa L.) populations from Mexico. Front. Plant Sci. 2016;7:1492. doi: 10.3389/fpls.2016.01492. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Alarcón-Reverte. R, et al. Concerted action of target-site mutations and high EPSPS activity in glyphosate-resistant junglerice (Echinochloa colona) from California. Pest Manag. Sci. 2015;71:996–1007. doi: 10.1002/ps.3878. [DOI] [PubMed] [Google Scholar]
- 18.Alcántara-de la Cruz R, et al. First resistance mechanisms characterization in glyphosate-resistant Leptochloa virgata. Front. Plant Sci. 2016;7:1742. doi: 10.3389/fpls.2016.01742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Lorraine-Colwill DF, et al. Investigations into the mechanism of glyphosate resistance in Lolium rigidum. Pest. Biochem. Physio. 2003;74:62–72. doi: 10.1016/S0048-3575(03)00007-5. [DOI] [Google Scholar]
- 20.Michitte P, De Prado R, Espinoza N, Ruiz-Santaella JP, Gauvrit C. Mechanisms of resistance to glyphosate in a ryegrass (Lolium multiflorum) biotype from Chile. Weed Sci. 2007;55:435–440. doi: 10.1614/WS-06-167.1. [DOI] [Google Scholar]
- 21.Vila-Aiub MM, et al. Glyphosate resistance in perennial Sorghum halepense (Johnsongrass), endowed by reduced glyphosate translocation and leaf uptake. Pest Manag. Sci. 2012;68:430–436. doi: 10.1002/ps.2286. [DOI] [PubMed] [Google Scholar]
- 22.Ge. X, et al. Vacuolar glyphosate-sequestration correlates with glyphosate resistance in ryegrass (Lolium spp.) from Australia, South America, and Europe, a31P NMR investigation. J. Agric. Food Chem. 2012;60:1243–1250. doi: 10.1021/jf203472s. [DOI] [PubMed] [Google Scholar]
- 23.Carvalho LB, et al. Pool of resistance mechanisms to glyphosate in Digitaria insularis. J. Agric. Food Chem. 2012;60:615–622. doi: 10.1021/jf204089d. [DOI] [PubMed] [Google Scholar]
- 24.U. S. Department of Agriculture (USDA) Citrus: World markets and trade. https://apps.fas.usda.gov/psdonline/circulars/citrus.pdf (2017).
- 25.Pérez-López M, et al. Characterization of glyphosate-resistant tropical sprangletop (Leptochloa virgata) and its alternative chemical control in Persian lime orchards in Mexico. Weed Sci. 2014;62:441–450. doi: 10.1614/WS-D-13-00177.1. [DOI] [Google Scholar]
- 26.Yu Q, et al. Evolution of a double amino acid substitution in the 5-enolpyruvylshikimate-3-phosphate synthase in Eleusine indica conferring high-level glyphosate resistance. Plant Physiol. 2015;167:1440–1447. doi: 10.1104/pp.15.00146. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Kaundun SS, et al. Importance of the P106S target-site mutation in conferring resistance to glyphosate in a goosegrass (Eleusine indica) population from the Philippines. Weed Sci. 2008;56:637–646. doi: 10.1614/WS-07-148.1. [DOI] [Google Scholar]
- 28.Mueller TC, Barnett KA, Brosnan JT, Steckel LE. Glyphosate-resistant goosegrass (Eleusine indica) confirmed in Tennessee. Weed Sci. 2011;59:562–566. doi: 10.1614/WS-D-11-00063.1. [DOI] [Google Scholar]
- 29.Cross R, et al. A Pro106 to Ala substitution is associated with resistance to glyphosate in annual bluegrass (Poa annua) Weed Sci. 2015;63:613–622. doi: 10.1614/WS-D-15-00033.1. [DOI] [Google Scholar]
- 30.Fernandez P, Gauvrit C, Barro F, Menendez J, De Prado R. First case of glyphosate resistance in France. Agron. Sustain. Dev. 2015;35:1469–1476. doi: 10.1007/s13593-015-0322-1. [DOI] [Google Scholar]
- 31.Ngo, T. D., Krishnan, M., Boutsalis, P., Gill, G., Preston, C. Target-site mutations conferring resistance to glyphosate in feathertop rhodes grass (Chloris virgata) populations in Australia. Pest Manag. Sci. doi:10.1002/ps.4512 (2017). [DOI] [PubMed]
- 32.De Prado, R. et al. Physiological and biochemical aspects of resistance in weeds with the use of glyphosate in A era Glyphosate: Agricultura, Meio Ambiente e Homen (eds Meschede D. K. & Pisa-Gazziero D. L.) 251–266 (Midiograf, 2016).
- 33.Preston C, Wakelin AM. Resistance to glyphosate from altered herbicide translocation patterns. Pest Manag. Sci. 2008;64:372–376. doi: 10.1002/ps.1489. [DOI] [PubMed] [Google Scholar]
- 34.Kleinman Z, Rubin B. Non-target-site glyphosate resistance in Conyza bonariensis is based on modified subcellular distribution on the herbicide. Pest Manag. Sci. 2017;73:246–253. doi: 10.1002/ps.4293. [DOI] [PubMed] [Google Scholar]
- 35.Yanniccari M, Istilart C, Giméneza DO, Castro AM. Effects of glyphosate on the movement of assimilates of two Lolium perenne L. populations with differential herbicide sensitivity. Environ. Exp. Bot. 2012;82:14–19. doi: 10.1016/j.envexpbot.2012.03.006. [DOI] [Google Scholar]
- 36.González-Torralva F, Gil-Humanes J, Barro F, Brants I, De Prado R. Target site mutation and reduced translocation are present in a glyphosate-resistant Lolium multiflorum Lam. population from Spain. Plant Physiol. Biochem. 2012;58:16–22. doi: 10.1016/j.plaphy.2012.06.001. [DOI] [PubMed] [Google Scholar]
- 37.Ghanizadeh H, Harrington KC, James TK, Woolleya DJ, Ellison NE. Mechanisms of glyphosate resistance in two perennial ryegrass (Lolium perenne) populations. Pest Manag. Sci. 2015;71:1617–1622. doi: 10.1002/ps.3968. [DOI] [PubMed] [Google Scholar]
- 38.Bracamonte E, Fernández-Moreno PT, Barro F, De Prado R. Glyphosate-resistant Parthenium hysterophorus in the Caribbean Islands: Non target site resistance and target site resistance in relation to resistance levels. Front. Plant Sci. 2016;7:1845. doi: 10.3389/fpls.2016.01845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Duke SO. Glyphosate degradation in glyphosate-resistant and -susceptible crops and weeds. J. Agric. Food Chem. 2011;59:5835–41. doi: 10.1021/jf102704x. [DOI] [PubMed] [Google Scholar]
- 40.Dinelli G, et al. Physiological and molecular bases of glyphosate resistance in Conyza bonariensis biotypes from Spain. Weed Res. 2008;48:257–265. doi: 10.1111/j.1365-3180.2008.00623.x. [DOI] [Google Scholar]
- 41.Powles SB. Gene amplification delivers glyphosate-resistant weed evolution. Proc. Natl. Acad. Sci. USA. 2010;107:955–956. doi: 10.1073/pnas.0913433107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Salas RA, et al. EPSPS gene amplification in glyphosate-resistant Italian ryegrass (Lolium perenne ssp. multiflorum) from Arkansas. Pest Manag. Sci. 2012;68:1223–1230. doi: 10.1002/ps.3342. [DOI] [PubMed] [Google Scholar]
- 43.Gaines TA, et al. Identification of genetic elements associated with EPSPS gene amplification. PLoS ONE. 2013;8:e65819. doi: 10.1371/journal.pone.0065819. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Wiersma AT, et al. Gene amplification of 5-enol-pyruvylshikimate-3-phosphate synthase in glyphosate-resistant Kochia scoparia. Planta. 2015;241:463–474. doi: 10.1007/s00425-014-2197-9. [DOI] [PubMed] [Google Scholar]
- 45.Malone JM, Morran S, Shirley N, Boutsalis P, Preston C. EPSPS gene amplification in glyphosate-resistant. Bromus diandrus. Pest Manag. Sci. 2016;72:81–88. doi: 10.1002/ps.4019. [DOI] [PubMed] [Google Scholar]
- 46.Salas RA, Scott RC, Dayan FE, Burgos NR. EPSPS gene amplification in glyphosate-resistant Italian ryegrass (Lolium perenne ssp. multiflorum) populations from Arkansas (United States) J. Agric. Food Chem. 2015;63:5885–5893. doi: 10.1021/acs.jafc.5b00018. [DOI] [PubMed] [Google Scholar]
- 47.Ngo, T. D., Malone, J. M., Boutsalis, P., Gill, G., Preston, C. EPSPS gene amplification conferring resistance to glyphosate in windmill grass (Chloris truncata) in Australia. Pest Manag Sci. doi:10.1002/ps.4573 (2017). [DOI] [PubMed]
- 48.Padgette S, et al. Site-directed mutagenesis of a conserved region of the 5-enolpyruvylshikimate-3-phosphate synthase active site. J. Biol. Chem. 1991;266:22364–22369. [PubMed] [Google Scholar]
- 49.Jalaludin A, Yu Q, Zoellner P, Beffa R, Powles SB. Characterisation of glufosinate resistance mechanisms in Eleusine indica. Pest Manag. Sci. 2017;73:1091–1100. doi: 10.1002/ps.4528. [DOI] [PubMed] [Google Scholar]
- 50.Shaner DL, Nadler-Hassar T, Henry WB, Koger CH. A rapid in vivo shikimate accumulation assay with excised leaf discs. Weed Sci. 2005;53:769–774. doi: 10.1614/WS-05-009R.1. [DOI] [Google Scholar]
- 51.Rojano-Delgado AM, Ruiz-Jiménez J, Castro MDL, De Prado R. Determination of glyphosate and its metabolites in plant material by reversed-polarity CE with indirect absorptiometric detection. Electrophoresis. 2010;31:1423–1430. doi: 10.1002/elps.200900583. [DOI] [PubMed] [Google Scholar]
- 52.Sammons, R. D., Meyer, J., Hall, E., Ostrander, E. & Schrader, S. A simple continuous assay for EPSP synthase from plant tissue. http://www.cottoninc.com/2007-Glyphosate-Resistant-Palmer-Amaranth/11a-Industry-Sammons-NCWSS07-poster.pdf (2007).
- 53.Vandesompele, J. et al. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 3, research0034; doi:10.1186/gb-2002-3-7-research0034 (2002). [DOI] [PMC free article] [PubMed]
- 54.Ritz C, Baty F, Streibig JC, Gerhard D. Dose-response analysis using R. PLoS ONE. 2015;10:e0146021. doi: 10.1371/journal.pone.0146021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Székács, A. & Darvas, B. Forty years with glyphosate in Herbicides-properties, synthesis and control of weeds (ed. Hasaneen, M. N.) 247–284 (InTech, 2012).