

Abstract
The lymph gland (LG) is a major source of hematopoiesis during Drosophila development. In this tissue, prohemocytes differentiate into multiple lineages, including macrophage-like plasmatocytes, which comprise the vast majority of mature hemocytes. Previous studies have uncovered genetic pathways that regulate prohemocyte maintenance and some cell fate choices between hemocyte lineages. However, less is known about how the plasmatocyte pool of the LG is established and matures. Here, we report that Tiggrin, a matrix protein expressed in the LG, is a specific regulator of plasmatocyte maturation. Tiggrin mutants exhibit precocious maturation of plasmatocytes, whereas Tiggrin overexpression blocks this process, resulting in a buildup of intermediate progenitors (IPs) expressing prohemocyte and hemocyte markers. These IPs likely represent a transitory state in prohemocyte to plasmatocyte differentiation. We also found that overexpression of Wee1 kinase, which slows G2/M progression, results in a phenotype similar to Tiggrin overexpression, whereas String/Cdc25 expression phenocopies Tiggrin mutants. Further analysis revealed that Wee1 inhibits plasmatocyte maturation through upregulation of Tiggrin transcription. Our results elucidate connections between the extracellular matrix and cell cycle regulators in the regulation of hematopoiesis.
KEY WORDS: Drosophila lymph gland, Hematopoiesis, Plasmatocyte, Tiggrin, Cell cycle
Summary: The matrix protein Tiggrin, which is modulated by cell cycle regulators, inhibits the maturation of plasmatocytes in the Drosophila larval lymph gland through an integrin-independent mechanism.
INTRODUCTION
In Drosophila, hemocytes are initially specified from the procephalic mesoderm and undergo further amplification during larval development (Lebestky et al., 2000; Makhijani et al., 2011; Markus et al., 2009). In parallel, precursor cells of the lymph gland (LG) assemble in the dorsal thoracic mesoderm during embryogenesis and develop during larval stages into several pairs of lobes aligned on the dorsal vessel (Holz et al., 2003; Jung et al., 2005). The cells of this tissue undergo hematopoiesis, and the LG dissembles at the start of pupation, releasing mature hemocytes into circulation, where they assist with tissue remodeling during metamorphosis (Grigorian et al., 2011; Lanot et al., 2001).
During the 3rd instar stage, the LG contains two disk-shaped primary lobes (PLs) that typically contain a few thousand cells divided into three domains. A centrally located medullary zone (MZ) contains prohemocytes with stem cell-like properties, whereas differentiating hemocytes are located in the peripheral cortical zone (CZ) (Evans et al., 2009; Jung et al., 2005). In addition, a small group of cells termed the posterior signaling center (PSC) controls differentiation and specification of hemocytes in the CZ (Benmimoun et al., 2015; Oyallon et al., 2016). Although much remains to be understood, the fly LG has developed into a powerful model for hematopoiesis and stem cell/progenitor regulation (Crozatier and Meister, 2007; Crozatier and Vincent, 2011; Letourneau et al., 2016; Martinez-Agosto et al., 2007; Morin-Poulard et al., 2013; Shim et al., 2013).
There are three major lineages of mature hemocytes in Drosophila: plasmatocytes, crystal cells and lamellocytes, all of which can be produced by the LG. Plasmatocytes contribute about 95% of all mature hemocytes in healthy animals (Crozatier and Meister, 2007; Letourneau et al., 2016). These cells are the equivalent of mammalian macrophages, which are able to clean both apoptotic debris and foreign materials (Gold and Brückner, 2015). They also play important roles in innate immunity (Charroux and Royet, 2009; Gold and Brückner, 2015) and participate in tissue regeneration by activating stem cells near sites of injury (Ayyaz et al., 2015). Crystal cells are specialized non-phagocytic cells that facilitate immune responses and wound-healing by causing melanization (Lanot et al., 2001). Lamellocytes are rarely found in healthy animals, but their number is significantly increased when larvae are immunologically challenged with infection by a parasitic wasp (Crozatier et al., 2004; Honti et al., 2014; Rizki and Rizki, 1992).
The genetic control of cell fate in the PL has been extensively studied, and several signaling pathways are known to be important for its proper development. For example, the Wnt protein Wingless (Wg) is expressed in the MZ, where it promotes prohemocyte proliferation and maintenance (Sinenko et al., 2009). Notch signaling controls the crystal cell-lamellocyte decision, as inhibition of this pathway resulted in a reduction in crystal cells and a large increase in lamellocytes in healthy larvae (Duvic et al., 2002; Small et al., 2014). Crystal cell number in the PL is also controlled by Hippo signaling, which restricts specification of this cell type (Ferguson and Martinez-Agosto, 2014; Milton et al., 2014). By comparison with crystal cells and lamellocytes, the maturation process of the largest hemocyte population in the LG, the plasmatocytes, remains relatively obscure.
The working model of the larval LG states that plasmatocytes are derived from prohemocytes. Consistent with this, a population of cells that expresses both MZ and CZ markers has been observed in the PL (Dragojlovic-Munther and Martinez-Agosto, 2012; Sinenko et al., 2009), as well as cells that possess CZ markers but lack mature plasmatocyte markers (Minakhina et al., 2011) or lack both MZ and mature plasmatocyte markers (Krzemien et al., 2010). These intermediate progenitors (IPs) are typically found near the MZ and have a higher mitotic capacity than differentiated plasmatocytes (Krzemien et al., 2010). There are some reports of factors controlling this IP pool, e.g. the transcription factor Pannier (Minakhina et al., 2011), but it has been difficult to pin down their roles, owing to the transitory nature of this population. The ability to ‘lock’ cells in this intermediate stage would be an important tool for better understanding their role in LG hematopoiesis.
We have previously reported that Wg signaling represses the expression of Tiggrin (Tig) in embryonic hemocytes and in the PL of the LG (Blauwkamp et al., 2008; Zhang et al., 2014). Tig encodes a large extracellular matrix (ECM) protein that binds to integrins and is important for muscle attachment and cell-cell adhesion (Bunch et al., 1998; Fogerty et al., 1994; Graner et al., 1998; Zhang et al., 2010). The repression of Tig transcription by Wg signaling is noteworthy, as it occurs through a direct mechanism involving novel binding sites for the transcription factor TCF/Pangolin (TCF/Pan), which mediates Wg target gene regulation in flies (Zhang et al., 2014). However, the physiological role of this regulation is not clear.
Here, we report on the biological role of Tig in the larval LG, using a combination of loss- and gain-of-function approaches. We found that Tig mutants displayed a premature appearance of mature plasmatocytes. Conversely, overexpression of Tig blocked plasmatocyte differentiation, and caused a large buildup of IPs that express both MZ and CZ markers. These manipulations of Tig levels had little or no effect on the number of crystal cells and lamellocytes. Expression of a Tig mutant transgene lacking an integrin-binding domain had the same effect as wild-type Tig, suggesting that the function of Tig in the CZ is independent of integrin signaling. In addition, we found that regulators of G2/M transition dramatically affect plasmatocyte differentiation and likely do so through regulation of Tig expression. These results highlight the connection between cell cycle regulators and the ECM protein Tig in the regulation of hematopoiesis in the fly LG.
RESULTS
Tig is required for maintaining the hemocyte population in the PL of the LG
Tig is an essential gene, with mutants dying as pupae owing to defects in muscle attachment, morphology and function (Bunch et al., 1998). Tig is secreted at muscle attachment sites by circulating hemocytes (Bunch et al., 1998; Fogerty et al., 1994). In addition to its expression in circulating hemocytes, we previously reported that Tig protein and two reporters containing Tig cis-regulatory sequences are primarily expressed in the CZ of the PL (Zhang et al., 2014). To examine the role of Tig in the larval LG, we examined PLs in a Tig mutant transheterozygous background (TigX/TigA1). The TigX allele is a small deletion removing the entire Tig locus and parts of two adjacent genes, whereas the TigA1 allele is an EMS-induced point mutation that fails to complement the muscle phenotype of TigX (Bunch et al., 1998). Tig mutants displayed a dramatic reduction in PL size in late 3rd instars (Fig. 1A,B). Both the CZ and MZ are reduced in Tig mutants compared with wild type (Fig. 1C), but the PSC cell number is unaffected (Fig. 1D). These results revealed a previously unexpected role for Tig in the larval LG development.
Fig. 1.
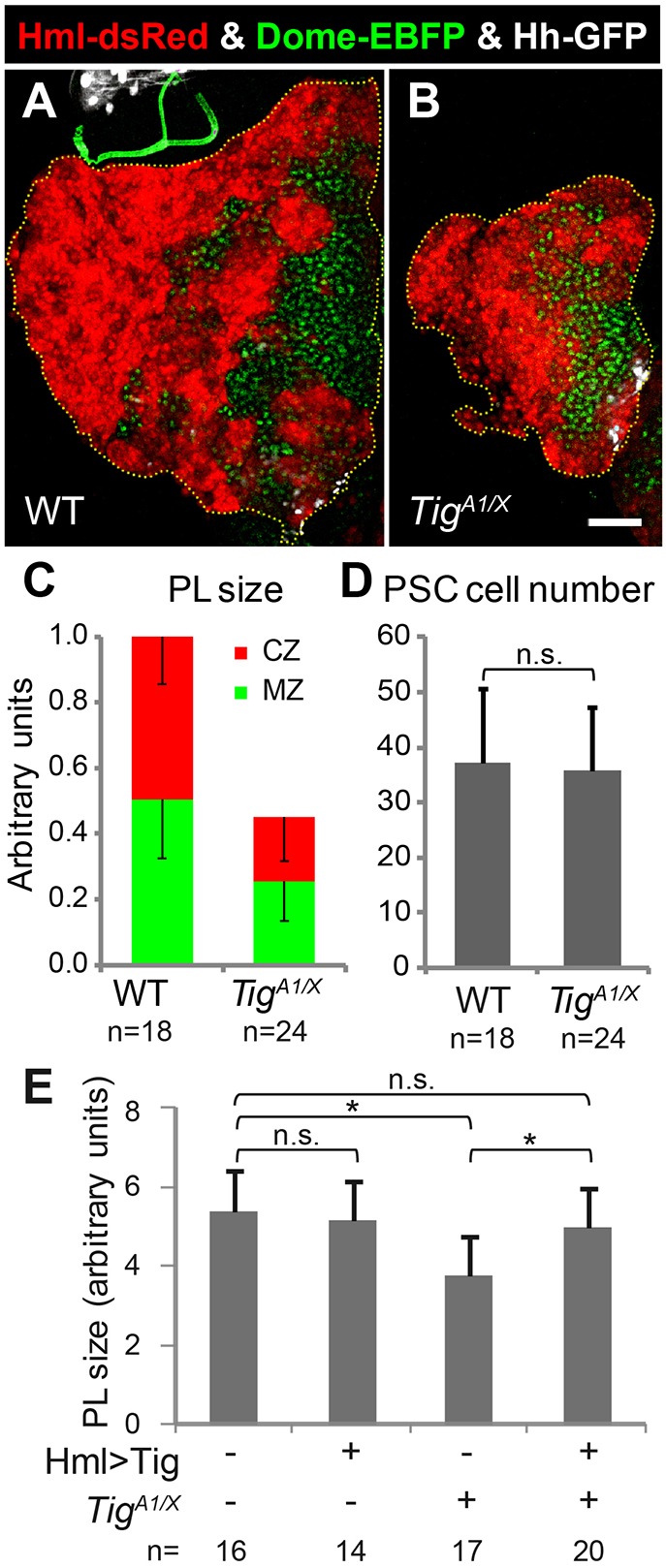
Tig is important for development of the PL of the LG. (A,B) Confocal images of PLs from mid/late 3rd instar larvae from w1118 or TigA1/X mutant transheterozygotes. The CZ, MZ and PSC are marked by Hml-dsRed (red), Dome-EBFP (green) and Hh-GFP (white), respectively. Tig mutants had smaller PLs with less CZ and MZ but unchanged PSC. (C) Quantification shows that the sizes of CZ, MZ and the total PL are significantly different between wild type and Tig mutants (P<0.01 for all comparisons). (D) No detectable change in PSC cell number is observed in Tig mutants. (E) Size of PLs from mid/late 3rd instar larvae containing P[Hml-Gal4] with or without P[UAS-Tig] and Tig mutant alleles. Hml>Tig has no effect on PL size by itself but rescued the PL size reduction of Tig mutants. The reduction of PL size in Tig mutants was less dramatic in the rescue experiment than in C (see also Tables S1 and S2). This is likely due to differences in the genetic backgrounds (i.e. the inclusion of P[Hml-Gal4], P[UAS-GFP] in the rescue data). *P<0.05, n.s., not significant. Data are mean±s.e.m. Scale bar: 50 µm.
To confirm the specificity of the Tig PL phenotype, a rescue was performed with a P[UAS-Tig] transgene via Hml-Gal4, which is active in the CZ of the PL, as well as resident and circulating hemocytes (Goto et al., 2003; Makhijani et al., 2011). In an otherwise wild-type background, Hml>Tig animals displayed no detectable difference in PL size, but Hml>Tig rescued the reduced PL phenotype of TigX/TigA1 larvae (Fig. 1E), indicating that the Tig mutant PL phenotype was due to loss of Tig activity. These data suggest that Tig acts in the CZ to maintain the size of the PL, with the MZ size reduction likely a secondary effect.
TigX/TigA1 mutant larvae are more slender and elongated than controls (Bunch et al., 1998), raising the possibility that the reduced PL size is a non-specific effect. To address this, the size of wing and eye-antennal imaginal discs in Tig mutants was examined. These tissues were 18-22% smaller in Tig mutants than in controls (Fig. S1). Although these reductions are statistically significant, they are less severe than the approximate twofold reduction observed in the PL (Fig. 1C).
To address whether premature release of hemocytes from Tig mutants could account for the small PL size, we examined control and Tig mutant PLs containing a Vkg>GFP gene trap, which indicates the location of collagen IV in the basal lamina (Morin et al., 2001). Although one-quarter of wild-type PLs have regions that are Vkg+ but devoid of cells (suggesting release of hemocytes from that area), the frequency of occurrence was similar in TigX/TigA1 mutants (Fig. S2). Early release of LG hemocytes should increase the number of circulating hemocytes in mid/late 3rd instar larvae, but we observed a reduction in Tig mutants (Fig. S2). This could be due to a reduction in embryonic hematopoiesis, given the expression of Tig in hemocytes at that stage (Fogerty et al., 1994; Alfonso and Jones, 2002; Blauwkamp et al., 2008). These data revealed no evidence for early release of hemocytes from Tig mutant LGs.
Examination of earlier stages revealed that Tig mutant PL size was similar to controls at late 2nd larval instar, but were significantly reduced by the early 3rd instar stage (Fig. 2A-G,I; Table S1). A similar trend is observed in CZ size (Fig. 2G,I; Table S1). The difference in PL size between controls and Tig mutants was similar whether ascertained using optical slices or volumetrically measured with stacked projections (Table S2). These data indicate a defect in PL and CZ growth starting at the early 3rd larval instar.
Fig. 2.

Tig mutants display precocious differentiation of plasmatocytes in the 1° and 2° lobes of the LG. (A-F′) Confocal images of PLs from late 2nd (A,D), early 3rd (B,E) and mid/late 3rd (C,F) instar larvae from w1118 (A-C) or TigA1/X mutants (D-F). The CZ and MZ are marked by Hml-dsRed (red) and Dome-EBFP (green), respectively (A-F), and plasmatocytes via P1 immunostaining (A′-F′). (G) The decrease in PL and CZ size in the Tig mutants manifested during the 3rd instar larval stage. (H) The precocious appearance of P1+ plasmatocytes in Tig mutants began in early 3rd instars. Data are mean values. (I) Summary of the statistical analysis in G,H. *P<0.05, **P<0.01, ***P<0.001, n.s., not significant. More information on these data is available in Table S2. (J,K) Confocal images of mid 3rd instar larval LGs of wild type and Tig mutants in which the 2° lobes are highlighted (yellow dashed lines), marked by Hml-dsRed (red), Dome-EBFP (green) and immunostained for P1 protein (white). The 2° lobes of Tig mutants have higher P1 staining and Hml-dsRed expression. (L,M) DIC images of control and Tig mutant 2° lobes (yellow dashed lines). Most 2° lobes of wild-type animals displayed smooth tightly packed cells, similar to the MZ of the PL. In contrast, cells in the 2° lobes of most Tig mutants appeared larger and more loosely packed, as is typically found in the CZ of the PL. Further description of the phenotypic range of the 2° lobes is provided in Table S3. Scale bars: 50 µm.
Tig mutants display precocious plasmatocyte differentiation
Plasmatocytes are specified throughout the 3rd larval instar stage and constitute the major type of hemocyte in the CZ (Crozatier and Meister, 2007). Plasmatocyte differentiation was monitored with the P1 antibody, which recognizes Nimrod C1 (NimC1), a phagocytosis receptor expressed in mature plasmatocytes (Kurucz et al., 2007). At late 2nd instar, both control and TigX/TigA1 PLs had similar low numbers of P1+ cells. By early 3rd instar, Tig mutants had a significant increase in plasmatocytes compared with wild type, which increased at mid/late 3rd instar (Fig. 2A′-F′,H,I; Table S1). This increase was observed when quantification was performed on optical slices or volumetrically (Table S2). In addition, the 2° lobes of TigX/TigA1 LGs had a dramatic increase in plasmatocytes in mid 3rd instars (Fig. 2J,K). Cells in control 2° lobes were small and compact, resembling the MZ of the PL. In contrast, most 2° lobes in Tig mutants had larger more loosely packed cells, reminiscent of CZ (Fig. 2L,M; Table S3). Development to pupation occurred slightly later in Tig mutants than in controls (Fig. S3), indicating that the precocious appearance of plasmatocytes in Tig mutants cannot be explained by a faster developmental clock. These data indicate that Tig suppresses plasmatocyte maturation in the LG.
To determine whether the precocious maturation of plasmatocytes was specific for loss of Tig, rescue via Hml>Tig was performed. Indeed, the phenotype was rescued by heterologous expression of Tig (Fig. 3I-L). Tig mutants had higher levels of mature plasmatocytes than wild-type or rescued PLs whether quantified by % P1+ cells/CZ (Fig. 3K) or % P1+ cells/PL (Fig. 3L). In contrast to control and Tig rescued PLs, where P1+ cells were also Hml+, in Tig mutants there were P1+ cells that were Hml− (arrows in Fig. 3E). These P1+Hml− cells may represent ‘hyper-mature’ plasmatocytes that have passed through the P1+Hml+ stage.
Fig. 3.
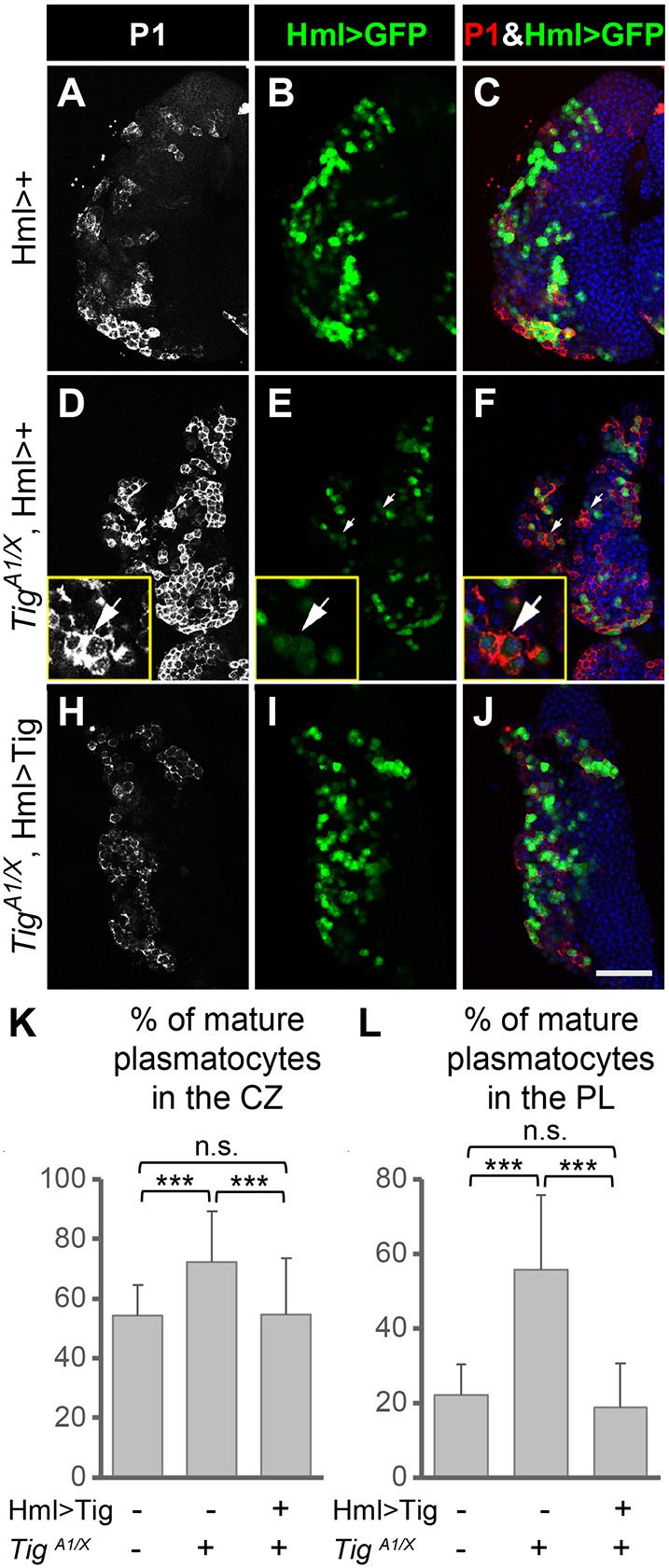
Rescue of the precocious plasmatocyte differentiation in Tig mutants by Hml>Tig. (A-J) Confocal images of PLs from mid 3rd instar larvae containing P[Hml-Gal4] and P[UAS-GFP] in a control (A-C) or a TigA1/X background without (D-F) or with (H-J) P[UAS-Tig]. Plasmatocytes were marked by P1 staining (white in A,D,H; red in C,F,J), DAPI (blue) and GFP (green). Loss of Tig resulted in an increase in P1+ mature plasmatocytes (D) that was rescued in a Hml>Tig background (H). P1+, Hml>GFP− cells are present in the Tig mutants (arrows in D-F; see also insets). (K) Quantification of the data, using the ratio of P1+, Hml>GFP+/Hml>GFP+ cells to determine the percentage of mature plasmatocytes/CZ. (L) Quantification of the data, normalizing the P1+ area to the entire PL, which includes the P1+, Hml>GFP− cells. Animals were reared at 25°C to restrict Hml>Tig expression to a moderate level. n.s., not significant; ***P<0.001. Nine PLs were examined for each condition. Data are mean±s.e.m. Scale bar: 50 µm.
The Tig plasmatocyte phenotype does not appear to be related to increased apoptosis, as expression of the caspase inhibitor P35 did not decrease the frequency of P1+ cells nor rescue PL size defect of Tig mutants (Fig. S4). Tig specifically affected plasmatocytes, as no significant difference in the number of lamellocytes or crystal cells were detected in TigX/TigA1 PLs (Fig. S5). However, partial knockdown of Tig with RNAi did not recapitulate the plasmatocyte phenotype (Fig. S6) and mutant clones of TigX did not display a detectable cell autonomous increase in P1+ cells (Fig. 4). The negative data with RNAi could be due to residual Tig in the PL, whereas the lack of increased plasmatocytes within the mutant clones could be due to non-autonomous rescue from Tig secreted from surrounding cells. Alternatively, these results could mean that loss of Tig in non-PL cells, e.g. circulating or sessile hemocytes are responsible for the PL phenotype (see Discussion for further comment).
Fig. 4.
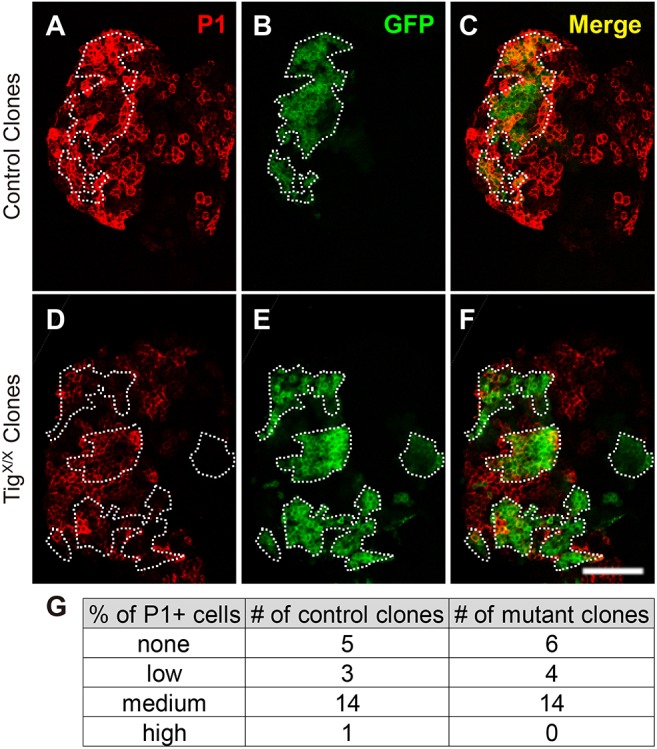
Tig mutant clones in the PL do not display a higher number of plasmatocytes. (A-F) Confocal images of PLs stained for P1 (red) and containing MARCM clones of control (A-C) and TigX mutant clones (D-F). Clones are marked by GFP (green). (G) Table summarizing P1 staining levels in control and Tig mutant clones. No obvious difference in P1 expression was observed inside the Tig mutant clones. Animals were reared at 25°C. Scale bar: 50 µm.
Tig overexpression inhibits plasmatocyte maturation
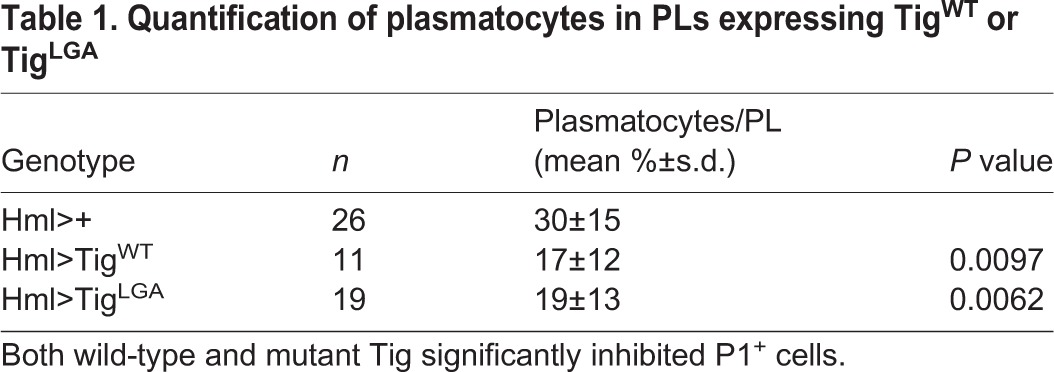
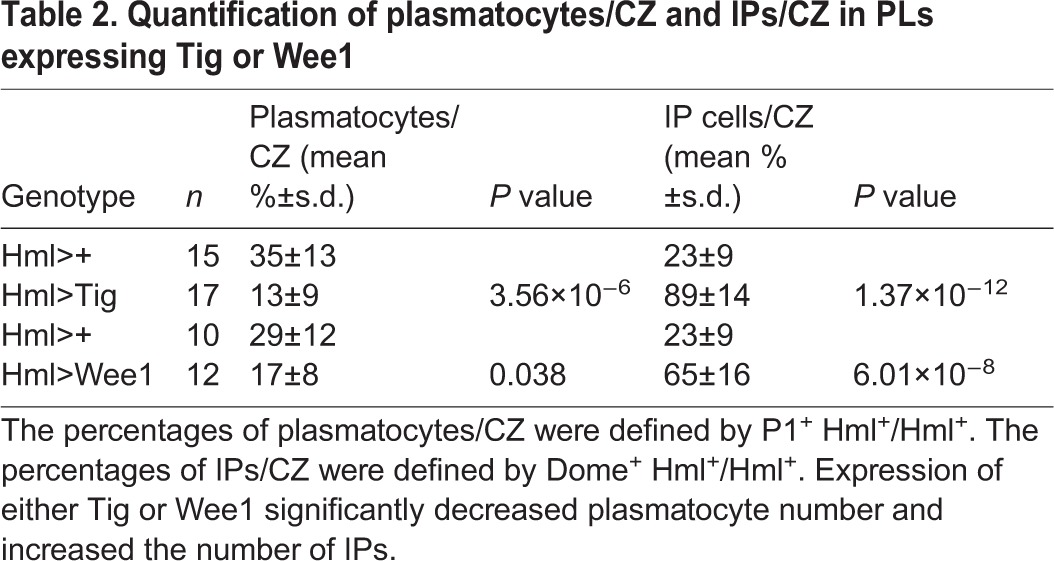
Hml>Tig animals reared at 25°C had no obvious differences in plasmatocyte differentiation (Fig. S7), but increasing the level of Tig expression (via culturing at 29°C) caused a dramatic reduction in the number of P1+ cells (Fig. 5A,F), while having no effect on the CZ marker Hml>GFP (Fig. 5B,G). The level of Tig expression in Hml>Tig PLs was much higher than controls (Fig. 5C,H; Fig. S8), but the residual P1+ cells had no detectable Tig signal (Fig. 5J). Quantification of either optical slices or z-stacks demonstrated that the reduction of plasmatocytes was significant (Table 1, Table S4). Hml>Tig animals displayed no developmental delay (Fig. S3), were viable as adults and expression of the caspase inhibitor P35 had no effect on the ability of Tig to inhibit plasmatocyte maturation (Fig. S9). To confirm the results obtained with P1 immunostaining, we examined a second marker: Eater-dsRed. This reporter is driven by an enhancer from the Eater locus, which encodes a phagocytosis receptor expressed specifically in plasmatoctyes (Kocks et al., 2005; Tokusumi et al., 2009). Hml>Tig PLs displayed a strong repression of Eater-dsRed expression (Fig. 5P-Q′). Taken together, these results suggest that overexpression of Tig blocks plasmatocyte differentiation.
Fig. 5.

Tig overexpression represses plasmatocyte differentiation independently of an integrin-binding domain. All confocal images are of PLs from mid/late 3rd instar larvae containing P[Hml-Gal4] and P[UAS-GFP], without or with transgenes expressing wild-type Tig (P[UAS-TigWT]) or a Tig transgene with a mutated integrin-binding motif (P[UAS-TigLGA]). (A-O) PLs stained for Tig and P1. When overexpressed at similar levels (C,H,M), both TigWT and TigLGA strongly repressed P1 expression (A,F,K). (J,O) Magnification of boxed areas in I and N showing minimal overlap between residual P1 signal and Tig. Quantification of the data is shown in Table 1 and Table S4. (P-R′) PLs containing an Eater-dsRed transgene expressing Tig proteins. Eater-dsRed was strongly repressed by either TigWT or TigLGA. (S-U′) PLs stained for the crystal cell marker Lz. TigWT and TigLGA did not cause a detectable change in the number of crystal cells (see Fig. S7 for quantification). Animals were reared at 29°C. Scale bars: 25 µm in K for A-D, F-I and K-N; 25 µm in O for E,J,O; 50 µm in R′,U′.
Table 1.
Quantification of plasmatocytes in PLs expressing TigWT or TigLGA
During the course of this study, we became aware that several of our stocks contained a mutant allele of nimC1 (Honti et al., 2013). Importantly, this deletion allele expresses a truncated protein that is not recognized by the P1 antibody. All stocks used in this report contain the wild-type nimC1 allele, except for the P[Hml-Gal4] P[UAS-GFP] chromosome, the transgenic inserts of which were too close to nimC1 to recombine away. To examine whether the presence of a mutant nimC1 allele affected plasmatocyte number, we examined Eater-dsRed expression in the nimC1 mutants. Interestingly, we found that the nimC1 mutants had increased Eater-dsRed expression (Fig. S10). In addition, another CZ driver, Pxn-Gal4 (which was wild-type for nimC1), also inhibited P1+ positive cells when combined with UAS-Tig (Fig. S11). We conclude that the presence of the nimC1 mutant allele in the Hml-Gal4 experiments does not alter the conclusion that Tig overexpression blocks plasmatocyte maturation.
Based on results from a combination of genetic and biochemical experiments, Tig is a ligand for αPSβPS2 integrin (Brabant et al., 1996; Bunch et al., 1998; Stevens and Jacobs, 2002). Tig contains a RGD motif commonly found in integrin ligands (Fogerty et al., 1994). Substitution of these residues (to LGA) greatly reduced integrin-mediated cell spreading and dramatically lowered the ability of transgenic Tig to rescue the muscle attachment defects and lethality of Tig mutants (Bunch et al., 1998). However, TigLGA expression via Hml-Gal4 resulted in the same phenotype, i.e. inhibition of P1+ cells while retaining Hml+ cells, as TigWT (Fig. 5K-O; Table 1). TigLGA and TigWT also inhibited Eater-dsRed expression similarly (Fig. 5P-R′). Expression levels of both transgenes were similar, as judged by Tig immunostaining (Fig. 5H,M). In addition, overexpression of either Tig had no effect on the frequency of crystal cell formation (Fig. 5S-U′; Fig. S12) or lamellocytes (Fig. S5). These data suggest that Tig specifically inhibits plasmatocyte differentiation in an integrin-independent manner.
Tig prolongs a pre-plasmatocyte, IP cell fate in the PL
Overexpression of Tig in the CZ caused the accumulation of Hml+ cells that lack the plasmatocyte markers P1 and Eater-dsRed (Fig. 5). They are reminiscent of the IPs that have been previously noted in wild-type PLs (Dragojlovic-Munther and Martinez-Agosto, 2012; Krzemien et al., 2010; Makhijani et al., 2011). Indeed, our examination of Hml>GFP PLs revealed a significant population of cells that were Hml+ but were P1− (Fig. 6A-C′). Hml+ cells with very low levels of P1 staining are also evident (arrow in Fig. 6B). The Hml>GFP signal does not overlap with Lz staining, indicating that the Hml+ cells were not crystal cells (Fig. 6D-E′). Another line of evidence for the presence of IPs is the existence of cells at the MZ/CZ border that are positive for both MZ and CZ markers (Sinenko et al., 2009; Dragojlovic-Munther and Martinez-Agosto, 2012). Cells with these characteristics (i.e. Dome+, Hml+) were relatively rare in control PLs, but Hml+ cells with intermediate levels of Dome signal were readily apparent in the CZ of Hml>Tig PLs (Fig. 6F-I; Table 2). The data indicate that Tig expression causes a buildup of IPs that cannot proceed with plasmatocyte differentiation.
Fig. 6.

PLs contain a pool of Hml+/P1− IPs that is expanded by Tig expression. Confocal images of PLs from mid/late 3rd instar larvae. (A,A′) Stack projections of the surface layer of a P[Hml-Gal4]/P[UAS-GFP] PL with GFP (green) and P1 immunostaining (red). (B-C′) Magnified views illustrating areas with mostly IPs, i.e. GFP+ with no detectable P1 (B,B′; arrows indicate cells with low P1 levels) or largely mature plasmatocytes, i.e. GFP+ P1+ (C,C′). (D-E′) Stack projection of the surface layer of a wild-type PL expressing Hml>GFP (green) and immunostained for Lz (red). Crystal cells (Lz+) typically have little or no GFP, suggesting that the IPs are not crystal cells. Twelve PLs were examined for each condition. (F-I) PLs expressing P[Hml-Gal4] with or without P[UAS-Tig], also containing zone markers Hml-dsRed (green) and Dome-EBFP (red), and stained for P1 (white). Hml>Tig expanded the population of Hml+ P1− cells, which also contained intermediate levels of Dome-EBFP. See Table 2 for further quantification. There was also a reproducible increase in Dome-EBFP expression in the MZ (compare F with H), the reason for which is not clear. All animals were reared at 29°C. Scale bars: 25 µm.
Table 2.
Quantification of plasmatocytes/CZ and IPs/CZ in PLs expressing Tig or Wee1
Tig protein is found throughout the CZ (Zhang et al., 2014), which is seemingly in conflict with a model where Tig promotes an IP fate while inhibiting plasmatocyte maturation. To examine the expression of Tig in more detail, in situ hybridization was performed to determine the pattern of Tig mRNAs. This analysis revealed the presence of significant levels of Tig transcripts in both the MZ and CZ, although expression is higher in the CZ (Fig. S13). Examination of the Tig transcriptional reporters minR-lacZ and Tig-lacZ also revealed a complex expression pattern (Fig. 7). Tig-lacZ, containing a 1.8 kb stretch of genomic DNA that included sequences upstream of the Tig endogenous promoter and the first intron (Zhang et al., 2014), was expressed in the IP and adjacent MZ cells, but was most prominently found in the CZ (Fig. 7A-C,M; Table S5). minR-lacZ, containing two repeats of a 40 bp minimal Wg-responsive element from the Tig first intron (Zhang et al., 2014), had a more patchy expression where ∼40% of the pattern overlapped with the MZ and IP (Fig. 7G-I,M; Table S5). The Tig-lacZ reporter displayed greater overlap with P1 than did minR (Fig. 7D-F,J-L,M,O). Strikingly, although nearly 60% of minR-lacZ expression was found in the CZ (Table S5) fewer than 20% of P1+ cells expressed this reporter (Fig. 7O). These data indicate that although the Tig expression pattern overlaps with mature plasmatocytes, Tig transcription also occurs in the MZ and IPs.
Fig. 7.

Expression patterns of Tig transcriptional reporters in the PL. (A-L) Confocal images of wild-type PLs expressing Tig-lacZ (A-F) or minR-lacZ (G-L). PLs were labeled with Hml-dsRed and Dome-EBFP (A-C,G-I, mid/late 3rd instar) or immunostained for P1 (D-F,J-L, mid 3rd instar). Tig-lacZ was predominantly found in the CZ (A-C) but many P1+ cells did not express Tig-lacZ (D-F). minR-lacZ had a patchier expression pattern and was clearly found in both MZ and CZ (G-I), and most of the P1+ cells were not minR-lacZ+ (J-L). (M) Quantification of the data showing that both reporters were more likely found in IPs and the CZ rather than the MZ. More information on the quantification can be found in Table S5. (N,O) Quantification from D-F and J-L showing the degree of overlap between the lacZ reporters and P1. n.s., not significant; ***P<0.001. Seven PLs were examined for Tig-lacZ and 22 for minR-lacZ. Data are mean±s.e.m. Scale bars: 50 µm.
To examine the effect of Tig overexpression in the MZ, Dome-Gal4 was crossed to a strong UAS-Tig line. Dome>Tig PLs displayed a similar reduction in plasmatocytes as Hml>Tig (Fig. S14). However, Tig immunostaining revealed that Tig protein was detectable only in the CZ (Fig. S14). These results suggest zone-specific regulation of Tig protein, i.e. CZ-specific translation of Tig mRNA or MZ-specific degradation of Tig protein. Although the mechanism for these observations requires further investigation, overall these data combined with the Tig transcript and transcriptional reporter analysis suggest multiple levels of regulation of Tig expression in the PL, which is not inconsistent with a role in acting as a brake on plasmatocyte maturation.
G2/M regulators control plasmatocyte differentiation and Tig expression
Hml>Tig PLs had a higher S-phase index than controls (Fig. S15), suggesting that Tig is promoting cell proliferation. However, Hml>Tig PLs also had a dramatic reduction in M-phase index (Fig. S16), suggesting many cells are arrested in G2 phase. Given that Tig affects both plasmatocyte maturation and cell cycle progression in the PL, we wondered whether these two processes were related, e.g. whether manipulation of cell cycle regulators affected PL cell fate. By screening a collection of known cell cycle regulators, we discovered that expression of Wee1 kinase had a profound effect on plasmatocyte differentiation (Fig. 8). Wee1 regulates G2/M transition by inhibiting Cdk1, the kinase subunit of maturation-promoting factor (MPF), which promotes the onset of M phase (Campbell et al., 1995; Price et al., 2002; Russell and Nurse, 1987). Hml>Wee1 PLs had a reduction in the number of plasmatocytes (Fig. 8A-H; Table 2 and Table S4), with no significant change in crystal cells and lamellocytes (Fig. 8I-L; Figs S5 and S12). As observed with Tig overexpression, Wee1 caused an accumulation of IPs in the PL (Table 2).
Fig. 8.
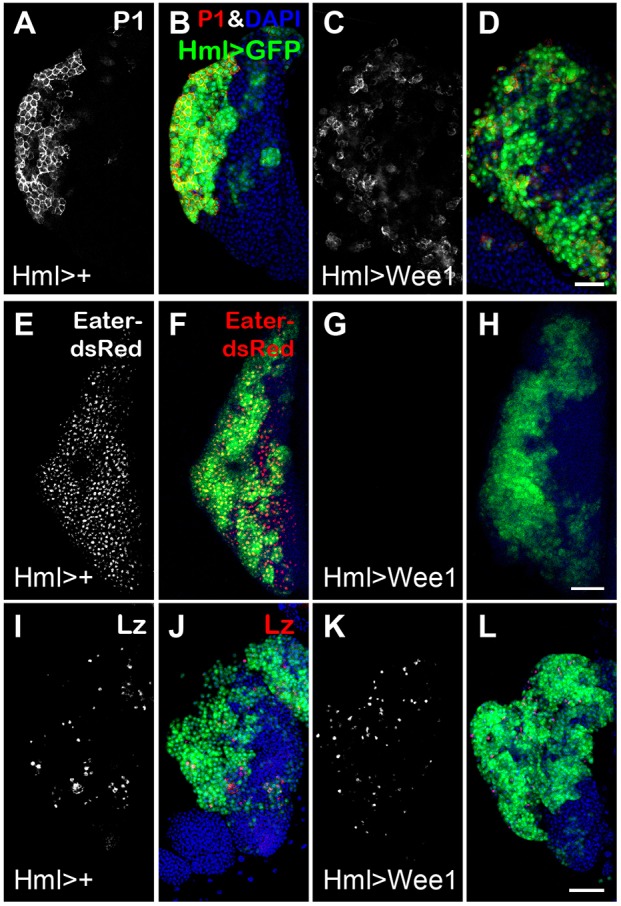
Wee1 kinase blocks plasmatocyte differentiation. (A-L) Confocal images of PLs from mid/late 3rd instar larvae containing P[Hml-Gal4] and P[UAS-GFP] with or without P[UAS-Wee1] labeled with different cell fate markers. In Hml>Wee1 PLs, P1 (A-D) and Eater-dsRed (E-H) signals were strongly repressed, whereas Lz (I-L) displayed no detectable difference. See Table 2 and Fig. S7 for quantification. Animals were reared at 29°C. Scale bars: 50 µm.
To confirm that Wee1 expression caused a slowdown of the G2/M transition, we used the RGB cell cycle tracker (Handke et al., 2014). Wee1 caused a marked increase in cells that were positive for EBFP, Tomato and EGFP (arrowheads in Fig. S17), indicative of late G2 (Handke et al., 2014). Interestingly, the slowing of the G2/M transition did not reduce the number of Hml+ cells, perhaps owing to increased proliferation of MZ cells (Fig. S17).
To extend the Wee1 results, we examined PLs expressing String/CDC25 (Stg), a phosphatase that antagonizes Wee1 function to activate Cdk1 (Edgar and O'Farrell, 1990; Russell and Nurse, 1986). Hml>Stg phenocopied Tig mutants, i.e. PLs were smaller than controls with an increase in mature plasmatocytes (Fig. 9A-F). Interestingly, Hml>Stg PLs had a significant reduction in minR-lacZ expression (Fig. 9G-K). Taken together, the Wee1 and Stg data link the G2/M transition to Tig expression and plasmatocyte differentiation.
Fig. 9.
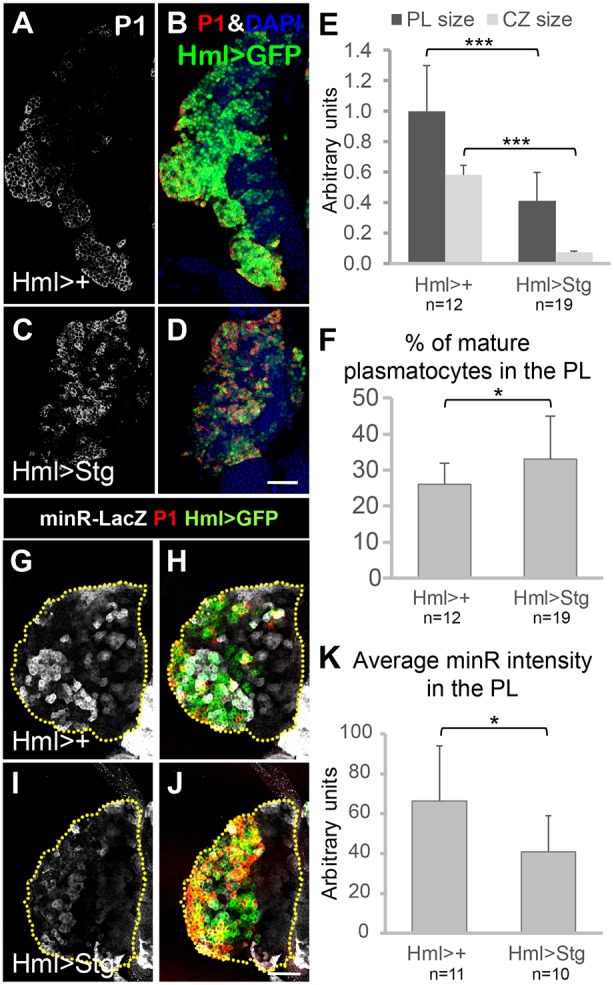
The phosphatase Stg promotes plasmatocyte differentiation and inhibits Tig reporter expression. (A-D) Confocal images of mid/late 3rd instar PLs containing P[Hml-Gal4] and P[UAS-GFP] with or without P[UAS-Stg], and immunostained for P1. In Hml>Stg PLs, the vast majority of Hml>GFP+ cells were also P1+, and the IP population (GFP+, low/no P1) was greatly reduced. (E) Quantification confirmed that PL and CZ sizes were greatly reduced in Hml>Stg PLs. (F) Quantification of P1 staining demonstrated an increase in mature plasmatocytes in Hml>Stg PLs. (G-J) Confocal images of mid 3rd instar PLs containing the minR-lacZ reporter, along with P[Hml-Gal4] and P[UAS-GFP] with or without P[UAS-Stg]. Stg expression inhibited minR-lacZ expression. (K) Quantification of the data in G-J. All animals were reared at 29°C. Data are mean±s.e.m. *P<0.05; ***P<0.001. Scale bars: 50 µm.
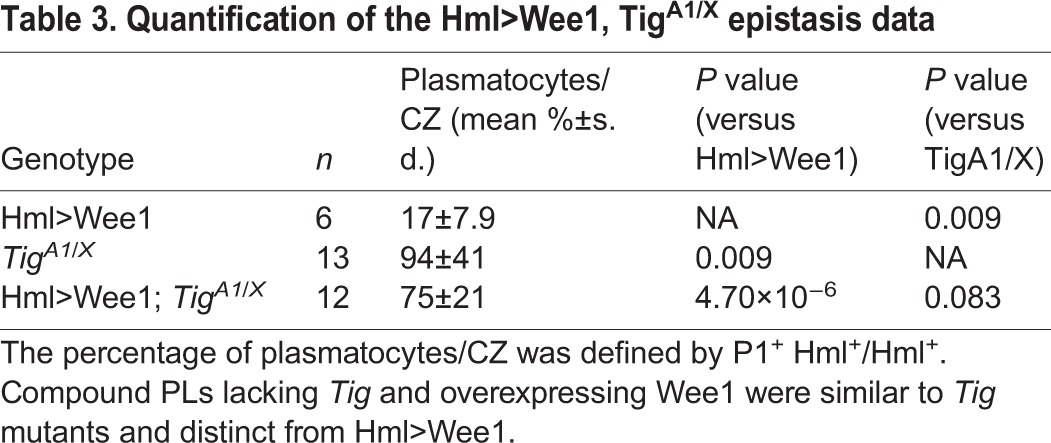
The similarity between the Tig and G2/M phenotypes raised the possibility that they act in a linear pathway. To test this, epistasis analysis was performed by overexpressing Wee1 in a Tig mutant background. As described earlier, Hml>Wee1 PLs had a reduced number of mature plasmatocytes (Fig. 10A-D). Conversely, TigA1/TigX mutants had a high level of P1+ plasmatoctyes (Fig. 10E-H). The composite (TigA1/X, Hml>Wee1) phenotype was very similar to the Tig mutant alone, i.e. numerous mature plasmatocytes and very few IPs (Hml+, no/low P1) (Fig. 10I-L; Table 3). These data suggest that Tig acts downstream of Wee1 in regulating the plasmatocyte cell fate.
Fig. 10.
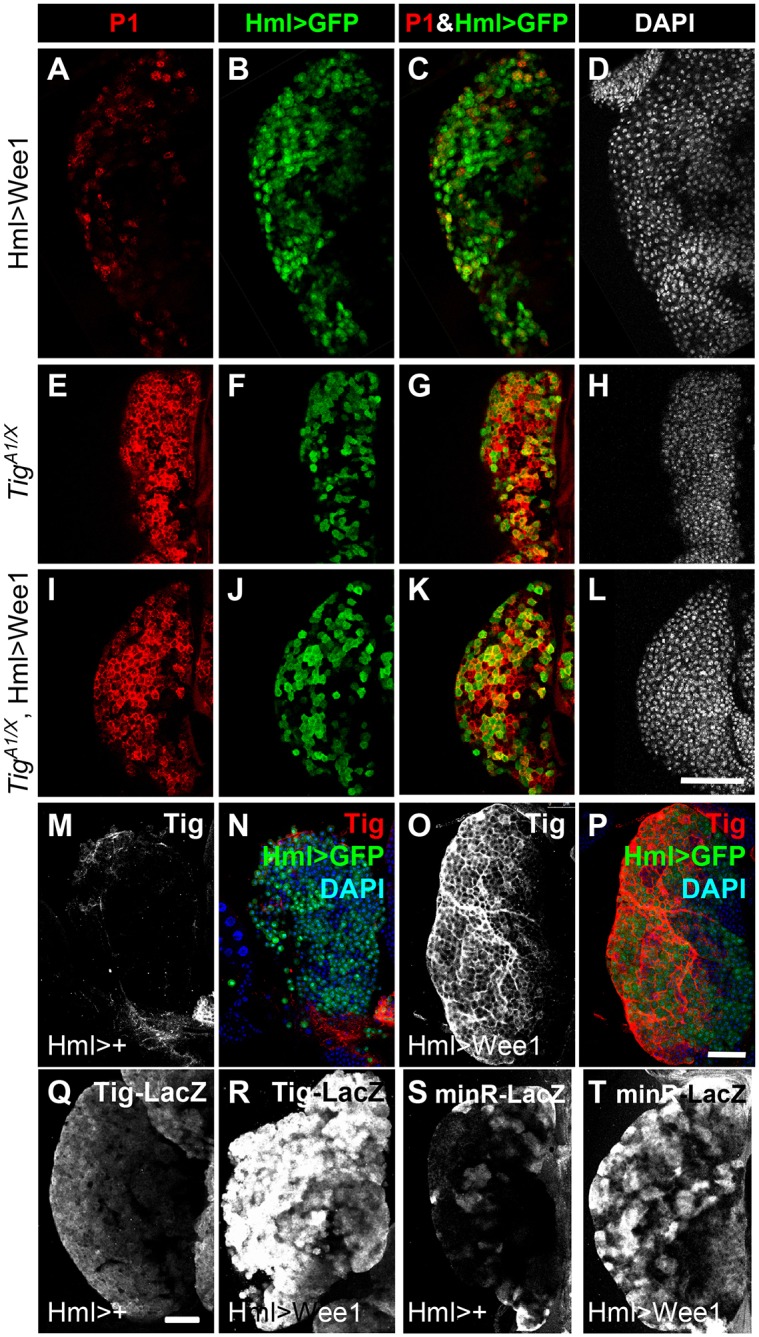
Wee1 regulates plasmatocyte differentiation through Tig expression. Confocal images of PLs from mid/late 3rd instar larva. All PLs contained P[Hml-Gal4] and P[UAS-GFP] with or without P[UAS-Wee] and Tig mutant alleles. (A-D) The CZ (Hml>GFP+) of Hml>Wee1 PLs had few plasmatocytes (GFP+ P1+) and many IPs (GFP+ low/no P1). (E-L) The CZ of Tig mutant PLs, with or without Wee1 overexpression, had high levels of mature plasmatocytes and few IPs. See Table 3 for quantification. (M-T) PLs containing P[Hml-Gal4] and P[UAS-GFP] with or without P[UAS-Wee1]. Wee1 induced strong activation of Tig protein (compare M with O) in 50% of PLs examined (n=12). For Tig-lacZ, strong Wee1-dependent activation of the reporter (compare Q with R) was observed in 75% of PLs (n=12). For minR-lacZ, strong induction (compare S with T) was observed in all PLs (n=10). All animals were reared at 29°C. Scale bars: 50 µm.
Table 3.
Quantification of the Hml>Wee1, TigA1/X epistasis data
One possibility to explain our epistasis data is that Wee1 activates Tig expression. Indeed, Tig protein and the Tig transcriptional reporters were dramatically upregulated in a Hml>Wee1 background (Fig. 10M-T), demonstrating that Wee1 expression transcriptionally activated Tig expression. Taken together, the data strongly support a model where the G2/M transition regulator Wee1 represses plasmatocyte differentiation through inducing Tig expression.
DISCUSSION
In this report, we demonstrated that the ECM protein Tig is an important negative regulator of plasmatocyte maturation in the LG of Drosophila (Fig. 11A). Overexpression of Tig inhibited the differentiation of plasmatocytes and Tig mutant primary and secondary lobes had precocious maturation of these macrophage-like cells. These manipulations in Tig gene activity had little or no effect on non-plasmatocyte lineages, i.e. crystal cell and lamellocytes. Tig mutant phenotypes were rescued by expression of transgenic Hml>Tig. Hml-Gal4 is active in the CZ of the PL, but also in circulating and residual hemocytes outside the LG (Goto et al., 2003; Makhijani et al., 2011). Given that Tig is expressed in the CZ of the PL (Zhang et al., 2014; Fig. 10M, Figs S6, S8 and S14), the simplest explanation is that Tig expressed in the LG is responsible for regulating plasmatocytes. But our data do not exclude the possibility that Tig expressed in non-LG cells could also contribute to the phenotypes we observed. In any case, our work demonstrates that, in addition to its function in muscle attachment (Bunch et al., 1998), Tig plays an important role in regulating cell fate specification during hematopoiesis in the LG.
Fig. 11.
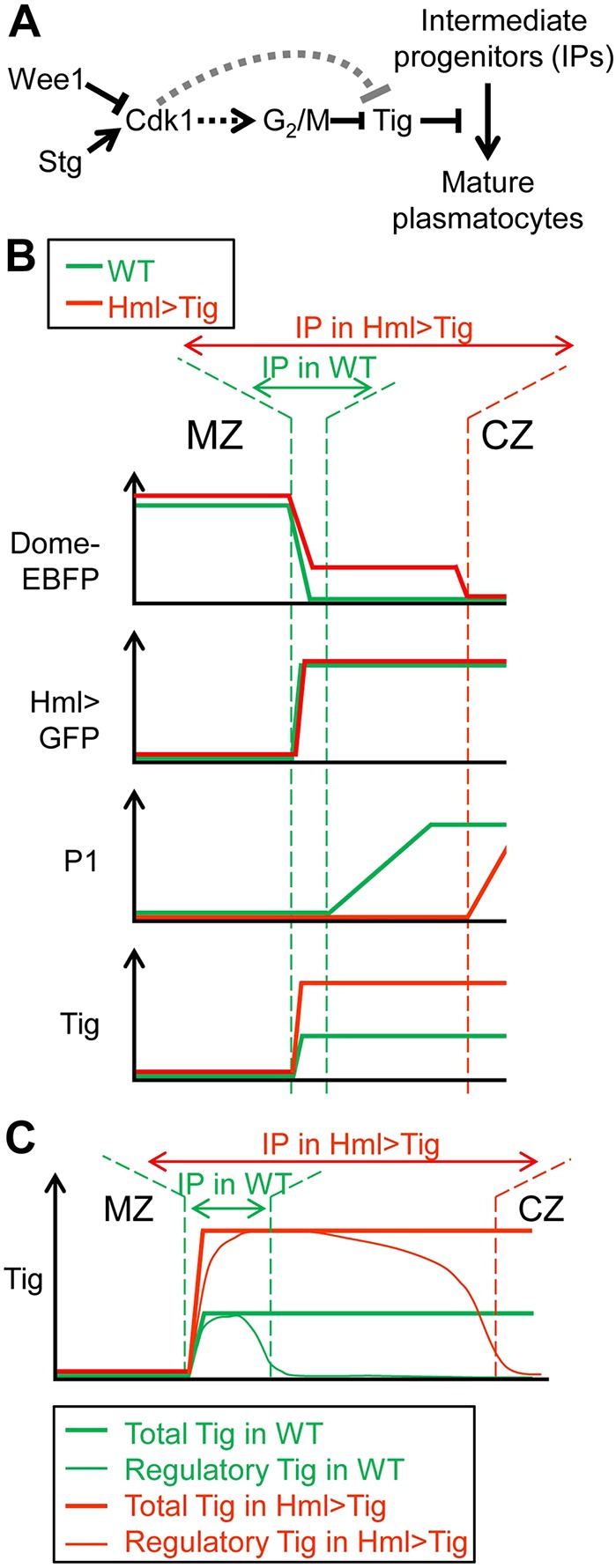
Working model of the regulation of plasmatocyte maturation by Tig and cell cycle regulators. (A) Genetic pathway controlling plasmatocyte differentiation. Tig and Wee1 inhibit the transition from IPs to plasmatocytes, while Stg accelerates it. Tig is epistatic to Wee1 and Wee1 activates Tig expression. Wee1 and Stg could affect Tig though their common target Cdk1 and may affect Tig expression by altering the G2/M transition. Alternatively, they could act on Tig transcription independently of the cell cycle. (B) Summary of the expression levels of different proteins across the MZ and CZ in wild-type and Hml>Tig PLs at the mid/late-3rd instar stage. There is a dramatic expansion of the domain containing IPs in Hml>Tig, i.e. cells that are Dome+/Hml+/P1−. (C) A speculative model for a ‘regulatory’ pool of Tig promoting the IP cell fate. Tig protein is detected at uniform levels throughout the CZ (Zhang et al., 2014), but we propose that newly synthesized Tig protein forms a regulatory pool that is enriched in the IPs, where it acts as a brake on plasmatocyte maturation.
Prohemocytes in the MZ (e.g. marked by Dome-EBFP) transition to hemocytes through IPs that contain residual Dome-EBFP and CZ markers such as Hml>GFP (Dragojlovic-Munther and Martinez-Agosto, 2012; Sinenko et al., 2009). Cells closer to the periphery of the CZ tend to express increasing levels of P1, a plasmatocyte marker (Krzemien et al., 2010; Makhijani et al., 2011) (Fig. 6A-C′). Thus, the prohemocytes in the MZ are Dome+, Hml− and P1−, IPs are Dome+, Hml+ and P1−, and maturing plasmatocytes are Dome−, Hml+ and P1+ (Fig. 11B). Overexpression of Tig in the CZ ‘freezes’ many cells in the IP fate, leading to an accumulation of cells expressing high levels of Hml reporters and intermediate levels of Dome-EBFP (Fig. 11B).
Our results suggest that Tig slows down plasmatocyte differentiation in the CZ, which could allow the formation of sufficient progenitors to generate the appropriate number of plasmatocytes. This model predicts that Tig expression would be highest in IPs, but this is not supported by immunostaining data (Zhang et al., 2014) and the analysis of Tig transcripts and transcriptional reporters in this report. One possibility is the existence of a ‘regulatory’ pool of Tig that is predominately active in IPs (Fig. 11C). Perhaps this regulatory pool comprises newly synthesized Tig, which can influence plasmatocyte maturation before it becomes incorporated into the ECM. An alternative explanation is that Tig is permissive for blocking plasmatocyte differentiation, and other unidentified factor(s) exist that inhibit plasmatocyte maturation and are localized to the IP compartment.
How does Tig inhibit plasmatocyte differentiation?
Tig contains an Arg-Gly-Glu (RGD) motif towards its C terminus, which was required for integrin binding in a cell-spreading assay (Bunch et al., 1998). Mutation of this tripeptide motif (RGD to LGA) greatly reduced the ability of a transgene to rescue the muscle attachment phenotype of Tig mutants (Bunch et al., 1998). However, we found that the TigLGA transgene had no detectable defect in blocking plasmatocyte differentiation. Although we cannot exclude the possibility that the LGA mutation retains the ability to bind to some integrin heterodimers, these data suggest that Tig regulates plasmatocyte differentiation independently of integrin signaling.
Are there other factors that regulate plasmatocyte development in Drosophila that could work in concert with Tig? While screening for suppressors of a LG overgrowth phenotype, three genes, visgun (vsg), SHC-adaptor protein (shc) and Adenosine deaminase-related growth factor A (Adgf-A), were identified where loss of function results in precocious plasmatocyte differentiation (Tan et al., 2012). vsg encodes an ortholog of mammalian endolyn, a endolysosomal sialomucin (Zhou et al., 2006) and shc encodes a SH2/PTB adaptor protein required for a subset of receptor tyrosine kinase receptors (Luschnig et al., 2000). Adgf-A expression is activated by JAK/STAT signaling in the CZ, which lowers extracellular adenosine levels in the MZ, maintaining the prohemocyte population (Mondal et al., 2011, 2014). The GATA transcription factor pannier (pnr) promotes plasmatocyte differentiation (Minakhina et al., 2011) so it would be possible to examine whether pnr is epistatic to Tig. Further examination is necessary to determine whether any of these factors act in conjunction with Tig to control plasmatocyte maturation.
Cell cycle regulation and cell fate determination – a case in fly hematopoiesis
Precise coordination between cell cycle progression and cell fate determination is necessary for proper development and tissue homeostasis, e.g. during neural cell lineages (Farkas and Huttner, 2008; Fichelson et al., 2005) and hematopoiesis (Nakamura-Ishizu et al., 2014). In many cases, cells exit the cell cycle upon terminal differentiation (Buttitta and Edgar, 2007) and perturbations that prolong cell cycle progression result in premature differentiation (Manansala et al., 2013; Tapias et al., 2014). Here, we report a particularly dramatic example where the specification of the plasmatocyte cell fate is tightly controlled by regulators of the G2/M transition, namely Wee1 and Stg. Interestingly, forced expression of Cdc25a (a vertebrate homolog of Stg) in zebrafish embryos blocks muscle differentiation (Bouldin et al., 2014). This is the opposite of what we observe, i.e. Stg overexpression promotes premature formation of plasmatocytes (Fig. 9). In pluripotent stem cells, cells in G1 phase are more likely to undergo differentiation (Bouldin and Kimelman, 2014; Calder et al., 2013; Coronado et al., 2013; Sela et al., 2012), possibly owing to cell cycle stage-dependent expression of key developmental regulators (Pauklin and Vallier, 2013; Singh et al., 2015). Perhaps our data showing that Wee1 and Stg regulate Tig expression is an example of G2 phase-dependent developmental regulation.
Although it is possible that Wee1 and Stg regulate plasmatocyte differentiation through their ability to regulate the cell cycle, other mechanisms are also possible. There is some evidence that Cyclin-dependent kinase 1 (Cdk1), the target of Wee and Stg (Fig. 11A), affects gene expression through phosphorylation of transcription factors (Lim and Kaldis, 2013; Hu et al., 2009, 2011). The fact that Wee1 and Stg regulate expression of minR-lacZ, a synthetic reporter that contains two TCF/Pan-binding sites (from the Tig regulatory region) placed upstream of a minimal promoter (Zhang et al., 2014), suggests that Wee1 and Stg influence Tig transcription via a mechanism that involves TCF/Pan or a factor that associates with this Wnt-regulated transcription factor. Further studies of this regulation will deepen our understanding of hematopoiesis and shed additional light on the connection between the cell cycle and cell fate determination.
MATERIALS AND METHODS
Drosophila stocks
pUAST-TigWT and pUAST-TigLGA plasmids were provided by Thomas Bunch (University of Arizona, Tucson, AZ, USA) (Bunch et al., 1998). Transgenic flies were generated by Rainbow Transgenic Flies (Camarillo, CA, USA) in a w1118 background. A pair of P[UAS-TigWT] and P[UAS-TigLGA] flies with similar relatively strong expression levels were used for all further experiments. For the rescue of Tig mutants, cultures containing P[UAS-Tig] transgenes were maintained at 25°C; in all other experiments, cultures were grown at 29°C to achieve significantly higher expression.
The other fly stocks used in this study were: TigX and TigA1 (Bunch et al., 1998); Hml-Gal4 (Goto et al., 2003); Domeless-Gal4 (Dome-Gal4) (Bourbon et al., 2002); UAS-Wee1 (Price et al., 2002); UAS-Stg (Neufeld and Edgar, 1998); UAS-RGB (Handke et al., 2014); Tig-lacZ and minR-lacZ (Zhang et al., 2014); Eater-dsRed (Kocks et al., 2005; Tokusumi et al., 2009); DHH, a line containing Dome-EBFP2, Hml-DsRed and hedgehog-GFP reporters (Evans et al., 2014); a Viking/Collagen IV gene trap (Vkg-GFP; Morin et al., 2001); Peroxidase-Gal4 (Pxn-Gal4; Stramer et al., 2005); UAS-P35 (Mergliano and Minden, 2003); and Serpent-Gal4 (Srp-Gal4; Huelsmann et al., 2006). The P[UAS-TigRNAi] is from the Vienna Drosophila Resource Center (#100036).
The TigX mutant allele was recombined onto a FRT40A chromosome as previously described (Xu and Rubin, 1993). Mutant clones were generated using the MARCM system, using a P[HS-FLP122] P[Tub-Gal4] P[UAS-GFP]; P[Tub-Gal80ts] FRT40A stock kindly provided by Chung-Yu Lee (University of Michigan, Ann Arbor, MI, USA). A FRT40A chromosome was used as a control. Clones were induced by a 1 hour heat shock at 37°C at 48-60 h AEL, and larva were dissected, fixed and analyzed 48 h later.
All crosses were initiated at 25°C. Embryos were collected within a 12 h window, transferred at 24-36 h AEL to 29°C if necessary and dissected at desired time (60-72 h AEL for late 2nd instar, 72-84 h for early 3rd instar, 90-102 h for mid 3rd instar, 96-108 h for mid/late 3rd instar and 102-114 h for late 3rd instar). See supplementary Materials and Methods for further details.
Immunohistochemistry and in situ hybridization
For PL/LG dissection, previously described protocols were used for immunostaining (Lebestky et al., 2000) or imaging of fluorescent markers (Small et al., 2012). Immunostaining was carried out as previously described (Zhang et al., 2014). Primary antibodies were used at the following dilutions: mouse α-P1 at 1:75, mouse α-L1 at 1:10 (Kurucz et al., 2007), mouse α-Lz at 1:30 (Lebestky et al., 2000; obtained from Developmental Studies Hybridoma Bank, DSHB), rabbit α-Tig at 1:50 (Deng et al., 2010), mouse α-Cut at 1:100 (Blochlinger et al., 1990; Ab 2B10, deposited by G. Rubin at DSHB) and rabbit α-lacZ (MP Biomedicals, 0855976) at 1:1000. Antisense digoxigenin-labeled probe against Tig mRNA was synthesized as previously described (Blauwkamp et al., 2008). See supplementary Materials and Methods for further details.
Imaging and data quantification
All fluorescent micrographs were taken with a Leica SP5 laser scanning confocal microscope. Bright-field imaging was conducted with a Nikon Eclipse E600 microscope. All images, except for Fig. 6A-C′, are thin optical slices, with P1 slices taken approximately one-third of the way into the PL from the dorsal side (where the Hml>GFP+, low/no P1 population of cells is the most obvious) and Lz slices taken approximately half way through the PL (where the most Lz+ cells are found). The number of PLs examined are indicated for each figure but were typically more than nine PLs/condition and were obtained in multiple experiments. There was no exclusion of samples and representative images are shown.
Images from optimal slices were quantified without blinding using Adobe Photoshop and ImageJ. For crucial data, i.e. the increase in P1+ plasmatocytes in Tig mutants and the reciprocal decrease in plasmatocytes caused by overexpression of Tig or Wee1, quantification of volumetric stacks was also performed. In all cases, data obtained from volumetric stacks and optical slices were similar (see Tables S1 and S4). The data are presented as means±s.d. Unpaired t-tests were used to determine whether means were statistically different. See supplementary Materials and Methods for further details.
Acknowledgements
We thank Utpal Banerjee, Robert Schulz, Laura Buttitta and the Bloomington Drosophila Stock Center for fly stocks. We also thank Thomas Bunch for pUAST-TigWT and pUAST-TigLGA plasmids, and Istvan Ando, Andrew Simmonds and the Developmental Studies Hybridoma Bank for primary antibodies. Thanks to Mingxue Gu for the TigRNAi data and Vinson Fan for careful reading of the manuscript. Special thanks to Laura Buttitta and members of the Buttitta lab for stimulating discussions about the link between the cell cycle and cell fate specification and for the use of their Leica DM6000 B upright microscope system.
Footnotes
Competing interests
The authors declare no competing or financial interests.
Author contributions
Conceptualization: C.U.Z., K.M.C.; Methodology: C.U.Z., K.M.C.; Validation: C.U.Z., K.M.C.; Formal analysis: C.U.Z., K.M.C.; Investigation: C.U.Z., K.M.C.; Resources: C.U.Z., K.M.C.; Writing - original draft: C.U.Z.; Writing - review & editing: C.U.Z., K.M.C.; Visualization: C.U.Z., K.M.C.; Supervision: K.M.C.; Project administration: K.M.C.; Funding acquisition: K.M.C.
Funding
This work was funded by grants from the National Institutes of Health (GM108468), American Heart Association (12PRE9520018) and the University of Michigan Comprehensive Cancer Center (G012896). Deposited in PMC for release after 12 months.
Supplementary information
Supplementary information available online at http://dev.biologists.org/lookup/doi/10.1242/dev.149641.supplemental
References
- Alfonso T. B. and Jones B. W. (2002). gcm2 promotes glial cell differentiation and is required with glial cells missing for macrophage development in Drosophila. Dev. Biol. 248, 369-383. 10.1006/dbio.2002.0740 [DOI] [PubMed] [Google Scholar]
- Ayyaz A., Li H. and Jasper H. (2015). Haemocytes control stem cell activity in the Drosophila intestine. Nat. Cell Biol. 17, 736-748. 10.1038/ncb3174 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Benmimoun B., Polesello C., Haenlin M. and Waltzer L. (2015). The EBF transcription factor Collier directly promotes Drosophila blood cell progenitor maintenance independently of the niche. Proc. Natl. Acad. Sci. USA 112, 9052-9057. 10.1073/pnas.1423967112 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blauwkamp T. A., Chang M. V. and Cadigan K. M. (2008). Novel TCF-binding sites specify transcriptional repression by Wnt signalling. EMBO J. 27, 1436-1446. 10.1038/emboj.2008.80 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blochlinger K., Bodmer R., Jan L. Y. and Jan Y. N. (1990). Patterns of expression of cut, a protein required for external sensory organ development in wild-type and cut mutant Drosophila embryos. Genes Dev. 4, 1322-1331. 10.1101/gad.4.8.1322 [DOI] [PubMed] [Google Scholar]
- Bouldin C. M. and Kimelman D. (2014). Cdc25 and the importance of G2 control: insights from developmental biology. Cell Cycle 13, 2165-2171. 10.4161/cc.29537 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouldin C. M., Snelson C. D., Farr G. H. III and Kimelman D. (2014). Restricted expression of cdc25a in the tailbud is essential for formation of the zebrafish posterior body. Genes Dev. 28, 384-395. 10.1101/gad.233577.113 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bourbon H.-M., Gonzy-Treboul G., Peronnet F., Alin M.-F., Ardourel C., Benassayag C., Cribbs D., Deutsch J., Ferrer P., Haenlin M. et al. (2002). A P-insertion screen identifying novel X-linked essential genes in Drosophila. Mech. Dev. 110, 71-83. 10.1016/S0925-4773(01)00566-4 [DOI] [PubMed] [Google Scholar]
- Brabant M. C., Fristrom D., Bunch T. A. and Brower D. L. (1996). Distinct spatial and temporal functions for PS integrins during Drosophila wing morphogenesis. Development 122, 3307-3317. [DOI] [PubMed] [Google Scholar]
- Bunch T. A., Graner M. W., Fessler L. I., Fessler J. H., Schneider K. D., Kerschen A., Choy L. P., Burgess B. W. and Brower D. L. (1998). The PS2 integrin ligand tiggrin is required for proper muscle function in Drosophila. Development 125, 1679-1689. [DOI] [PubMed] [Google Scholar]
- Buttitta L. A. and Edgar B. A. (2007). Mechanisms controlling cell cycle exit upon terminal differentiation. Curr. Opin. Cell Biol. 19, 697-704. 10.1016/j.ceb.2007.10.004 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Calder A., Roth-Albin I., Bhatia S., Pilquil C., Lee J. H., Bhatia M., Levadoux-Martin M., McNicol J., Russell J., Collins T. et al. (2013). Lengthened G1 phase indicates differentiation status in human embryonic stem cells. Stem Cells Dev. 22, 279-295. 10.1089/scd.2012.0168 [DOI] [PubMed] [Google Scholar]
- Campbell S. D., Sprenger F., Edgar B. A. and O'Farrell P. H. (1995). Drosophila Wee1 kinase rescues fission yeast from mitotic catastrophe and phosphorylates Drosophila Cdc2 in vitro. Mole. Biol. Cell 6, 1333-1347. 10.1091/mbc.6.10.1333 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Charroux B. and Royet J. (2009). Elimination of plasmatocytes by targeted apoptosis reveals their role in multiple aspects of the Drosophila immune response. Proc. Natl. Acad. Sci. USA 106, 9797-9802. 10.1073/pnas.0903971106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Coronado D., Godet M., Bourillot P.-Y., Tapponnier Y., Bernat A., Petit M., Afanassieff M., Markossian S., Malashicheva A., Iacone R. et al. (2013). A short G1 phase is an intrinsic determinant of naïve embryonic stem cell pluripotency. Stem Cell Res. 10, 118-131. 10.1016/j.scr.2012.10.004 [DOI] [PubMed] [Google Scholar]
- Crozatier M. and Meister M. (2007). Drosophila haematopoiesis. Cell. Microbiol. 9, 1117-1126. 10.1111/j.1462-5822.2007.00930.x [DOI] [PubMed] [Google Scholar]
- Crozatier M. and Vincent A. (2011). Drosophila: a model for studying genetic and molecular aspects of haematopoiesis and associated leukaemias. Dis. Model. Mech. 4, 439-445. 10.1242/dmm.007351 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crozatier M., Ubeda J.-M., Vincent A. and Meister M. (2004). Cellular immune response to parasitization in Drosophila requires the EBF orthologue collier. PLoS Biol. 2, e196 10.1371/journal.pbio.0020196 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Deng H., Bell J. B. and Simmonds A. J. (2010). Vestigial is required during late-stage muscle differentiation in Drosophila melanogaster embryos. Mol. Biol. Cell 21, 3304-3316. 10.1091/mbc.E10-04-0364 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dragojlovic-Munther M. and Martinez-Agosto J. A. (2012). Multifaceted roles of PTEN and TSC orchestrate growth and differentiation of Drosophila blood progenitors. Development 139, 3752-3763. 10.1242/dev.074203 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Duvic B., Hoffmann J. A., Meister M. and Royet J. (2002). Notch signaling controls lineage specification during Drosophila larval hematopoiesis. Curr. Biol. 12, 1923-1927. 10.1016/S0960-9822(02)01297-6 [DOI] [PubMed] [Google Scholar]
- Edgar B. A. and O'Farrell P. H. (1990). The three postblastoderm cell cycles of Drosophila embryogenesis are regulated in G2 by string. Cell 62, 469-480. 10.1016/0092-8674(90)90012-4 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans C. J., Olson J. M., Ngo K. T., Kim E., Lee N. E., Kuoy E., Patananan A. N., Sitz D., Tran P. T., Do M.-T. et al. (2009). G-TRACE: rapid Gal4-based cell lineage analysis in Drosophila. Nat. Methods 6, 603-605. 10.1038/nmeth.1356 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Evans C. J., Liu T. and Banerjee U. (2014). Drosophila hematopoiesis: markers and methods for molecular genetic analysis. Methods 68, 242-251. 10.1016/j.ymeth.2014.02.038 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Farkas L. M. and Huttner W. B. (2008). The cell biology of neural stem and progenitor cells and its significance for their proliferation versus differentiation during mammalian brain development. Curr. Opin. Cell Biol. 20, 707-715. 10.1016/j.ceb.2008.09.008 [DOI] [PubMed] [Google Scholar]
- Ferguson G. B. and Martinez-Agosto J. A. (2014). Yorkie and Scalloped signaling regulates Notch-dependent lineage specification during Drosophila hematopoiesis. Curr. Biol. 24, 2665-2672. 10.1016/j.cub.2014.09.081 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fichelson P., Audibert A., Simon F. and Gho M. (2005). Cell cycle and cell-fate determination in Drosophila neural cell lineages. Trends Genet. 21, 413-420. 10.1016/j.tig.2005.05.010 [DOI] [PubMed] [Google Scholar]
- Fogerty F. J., Fessler L. I., Bunch T. A., Yaron Y., Parker C. G., Nelson R. E., Brower D. L., Gullberg D. and Fessler J. H. (1994). Tiggrin, a novel Drosophila extracellular matrix protein that functions as a ligand for Drosophila alpha PS2 beta PS integrins. Development 120, 1747-1758. [DOI] [PubMed] [Google Scholar]
- Gold K. S. and Brückner K. (2015). Macrophages and cellular immunity in Drosophila melanogaster. Semin. Immunol. 27, 357-368. 10.1016/j.smim.2016.03.010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Goto A., Kadowaki T. and Kitagawa Y. (2003). Drosophila hemolectin gene is expressed in embryonic and larval hemocytes and its knock down causes bleeding defects. Dev. Biol. 264, 582-591. 10.1016/j.ydbio.2003.06.001 [DOI] [PubMed] [Google Scholar]
- Graner M. W., Bunch T. A., Baumgartner S., Kerschen A. and Brower D. L. (1998). Splice variants of the Drosophila PS2 integrins differentially interact with RGD-containing fragments of the extracellular proteins tiggrin, ten-m, and D-laminin 2. J. Biol. Chem. 273, 18235-18241. 10.1074/jbc.273.29.18235 [DOI] [PubMed] [Google Scholar]
- Grigorian M., Mandal L. and Hartenstein V. (2011). Hematopoiesis at the onset of metamorphosis: terminal differentiation and dissociation of the Drosophila lymph gland. Dev. Genes Evol. 221, 121-131. 10.1007/s00427-011-0364-6 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Handke B., Szabad J., Lidsky P. V., Hafen E. and Lehner C. F. (2014). Towards long term cultivation of Drosophila wing imaginal discs in vitro. PLoS ONE 9, e107333 10.1371/journal.pone.0107333 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Holz A., Bossinger B., Strasser T., Janning W. and Klapper R. (2003). The two origins of hemocytes in Drosophila. Development 130, 4955-4962. 10.1242/dev.00702 [DOI] [PubMed] [Google Scholar]
- Honti V., Cinege G., Csordás G., Kurucz É., Zsámboki J., Evans C. J., Banerjee U. and Andó I. (2013). Variation of NimC1 expression in Drosophila stocks and transgenic strains. Fly 7, 263-268. 10.4161/fly.25654 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Honti V., Csordás G., Kurucz É., Márkus R. and Andó I. (2014). The cell-mediated immunity of Drosophila melanogaster: hemocyte lineages, immune compartments, microanatomy and regulation. Dev. Comp. Immunol. 42, 47-56. 10.1016/j.dci.2013.06.005 [DOI] [PubMed] [Google Scholar]
- Hu M. G., Deshpande A., Enos M., Mao D., Hinds E. A., Hu G.-F., Chang R., Guo Z., Dose M., Mao C. et al. (2009). A requirement for cyclin-dependent kinase 6 in thymocyte development and tumorigenesis. Cancer Res. 69, 810-818. 10.1158/0008-5472.CAN-08-2473 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hu M. G., Deshpande A., Schlichting N., Hinds E. A., Mao C., Dose M., Hu G.-F., Van Etten R. A., Gounari F. and Hinds P. W. (2011). CDK6 kinase activity is required for thymocyte development. Blood 117, 6120-6131. 10.1182/blood-2010-08-300517 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Huelsmann S., Hepper C., Marchese D., Knöll C. and Reuter R. (2006). The PDZ-GEF dizzy regulates cell shape of migrating macrophages via Rap1 and integrins in the Drosophila embryo. Development 133, 2915-2924. 10.1242/dev.02449 [DOI] [PubMed] [Google Scholar]
- Jung S.-H., Evans C. J., Uemura C. and Banerjee U. (2005). The Drosophila lymph gland as a developmental model of hematopoiesis. Development 132, 2521-2533. 10.1242/dev.01837 [DOI] [PubMed] [Google Scholar]
- Kocks C., Cho J. H., Nehme N., Ulvila J., Pearson A. M., Meister M., Strom C., Conto S. L., Hetru C., Stuart L. M. et al. (2005). Eater, a transmembrane protein mediating phagocytosis of bacterial pathogens in Drosophila. Cell 123, 335-346. 10.1016/j.cell.2005.08.034 [DOI] [PubMed] [Google Scholar]
- Krzemien J., Oyallon J., Crozatier M. and Vincent A. (2010). Hematopoietic progenitors and hemocyte lineages in the Drosophila lymph gland. Dev. Biol. 346, 310-319. 10.1016/j.ydbio.2010.08.003 [DOI] [PubMed] [Google Scholar]
- Kurucz E., Márkus R., Zsámboki J., Folkl-Medzihradszky K., Darula Z., Vilmos P., Udvardy A., Krausz I., Lukacsovich T., Gateff E. et al. (2007). Nimrod, a putative phagocytosis receptor with EGF repeats in Drosophila plasmatocytes. Curr. Biol. 17, 649-654. 10.1016/j.cub.2007.02.041 [DOI] [PubMed] [Google Scholar]
- Lanot R., Zachary D., Holder F. and Meister M. (2001). Postembryonic hematopoiesis in Drosophila. Dev. Biol. 230, 243-257. 10.1006/dbio.2000.0123 [DOI] [PubMed] [Google Scholar]
- Lebestky T., Chang T., Hartenstein V. and Banerjee U. (2000). Specification of Drosophila hematopoietic lineage by conserved transcription factors. Science 288, 146-149. 10.1126/science.288.5463.146 [DOI] [PubMed] [Google Scholar]
- Letourneau M., Lapraz F., Sharma A., Vanzo N., Waltzer L. and Crozatier M. (2016). Drosophila hematopoiesis under normal conditions and in response to immune stress. FEBS Lett. 590, 4034-4051. 10.1002/1873-3468.12327 [DOI] [PubMed] [Google Scholar]
- Lim S. and Kaldis P. (2013). Cdks, cyclins and CKIs: roles beyond cell cycle regulation. Development 140, 3079-3093. 10.1242/dev.091744 [DOI] [PubMed] [Google Scholar]
- Luschnig S., Krauss J., Bohmann K., Desjeux I. and Nüsslein-Volhard C. (2000). The Drosophila SHC adaptor protein is required for signaling by a subset of receptor tyrosine kinases. Mol. Cell 5, 231-241. 10.1016/S1097-2765(00)80419-0 [DOI] [PubMed] [Google Scholar]
- Makhijani K., Alexander B., Tanaka T., Rulifson E. and Bruckner K. (2011). The peripheral nervous system supports blood cell homing and survival in the Drosophila larva. Development 138, 5379-5391. 10.1242/dev.067322 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Manansala M. C., Min S. and Cleary M. D. (2013). The Drosophila SERTAD protein Taranis determines lineage-specific neural progenitor proliferation patterns. Dev. Biol. 376, 150-162. 10.1016/j.ydbio.2013.01.025 [DOI] [PubMed] [Google Scholar]
- Markus R., Laurinyecz B., Kurucz E., Honti V., Bajusz I., Sipos B., Somogyi K., Kronhamn J., Hultmark D. and Ando I. (2009). Sessile hemocytes as a hematopoietic compartment in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 106, 4805-4809. 10.1073/pnas.0801766106 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Martinez-Agosto J. A., Mikkola H. K. A., Hartenstein V. and Banerjee U. (2007). The hematopoietic stem cell and its niche: a comparative view. Genes Dev. 21, 3044-3060. 10.1101/gad.1602607 [DOI] [PubMed] [Google Scholar]
- Mergliano J. and Minden J. S. (2003). Caspase-independent cell engulfment mirrors cell death pattern in Drosophila embryos. Development 130, 5779-5789. 10.1242/dev.00824 [DOI] [PubMed] [Google Scholar]
- Milton C. C., Grusche F. A., Degoutin J. L., Yu E., Dai Q., Lai E. C. and Harvey K. F. (2014). The Hippo pathway regulates hematopoiesis in Drosophila melanogaster. Curr. Biol. 24, 2673-2680. 10.1016/j.cub.2014.10.031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Minakhina S., Tan W. and Steward R. (2011). JAK/STAT and the GATA factor Pannier control hemocyte maturation and differentiation in Drosophila. Dev. Biol. 352, 308-316. 10.1016/j.ydbio.2011.01.035 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mondal B. C., Mukherjee T., Mandal L., Evans C. J., Sinenko S. A., Martinez-Agosto J. A. and Banerjee U. (2011). Interaction between differentiating cell- and niche-derived signals in hematopoietic progenitor maintenance. Cell 147, 1589-1600. 10.1016/j.cell.2011.11.041 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mondal B. C., Shim J., Evans C. J. and Banerjee U. (2014). Pvr expression regulators in equilibrium signal control and maintenance of Drosophila blood progenitors. Elife 3, e03626 10.7554/eLife.03626 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morin X., Daneman R., Zavortink M. and Chia W. (2001). A protein trap strategy to detect GFP-tagged proteins expressed from their endogenous loci in Drosophila. Proc. Natl. Acad. Sci. USA 98, 15050-15055. 10.1073/pnas.261408198 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Morin-Poulard I., Vincent A. and Crozatier M. (2013). The Drosophila JAK-STAT pathway in blood cell formation and immunity. Jak-Stat. 2, e25700 10.4161/jkst.25700 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nakamura-Ishizu A., Takizawa H. and Suda T. (2014). The analysis, roles and regulation of quiescence in hematopoietic stem cells. Development 141, 4656-4666. 10.1242/dev.106575 [DOI] [PubMed] [Google Scholar]
- Neufeld T. P. and Edgar B. A. (1998). Connections between growth and the cell cycle. Curr. Opin. Cell Biol. 10, 784-790. 10.1016/S0955-0674(98)80122-1 [DOI] [PubMed] [Google Scholar]
- Oyallon J., Vanzo N., Krzemień J., Morin-Poulard I., Vincent A. and Crozatier M. (2016). Two independent functions of collier/early B cell factor in the control of drosophila blood cell homeostasis. PLoS ONE 11, e0148978 10.1371/journal.pone.0148978 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pauklin S. and Vallier L. (2013). The cell-cycle state of stem cells determines cell fate propensity. Cell 155, 135-147. 10.1016/j.cell.2013.08.031 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price D. M., Jin Z., Rabinovitch S. and Campbell S. D. (2002). Ectopic expression of the Drosophila Cdk1 inhibitory kinases, Wee1 and Myt1, interferes with the second mitotic wave and disrupts pattern formation during eye development. Genetics 161, 721-731. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rizki T. M. and Rizki R. M. (1992). Lamellocyte differentiation in Drosophila larvae parasitized by Leptopilina. Dev. Comp. Immunol. 16, 103-110. 10.1016/0145-305X(92)90011-Z [DOI] [PubMed] [Google Scholar]
- Russell P. and Nurse P. (1986). cdc25+ functions as an inducer in the mitotic control of fission yeast. Cell 45, 145-153. 10.1016/0092-8674(86)90546-5 [DOI] [PubMed] [Google Scholar]
- Russell P. and Nurse P. (1987). Negative regulation of mitosis by wee1+, a gene encoding a protein kinase homolog. Cell 49, 559-567. 10.1016/0092-8674(87)90458-2 [DOI] [PubMed] [Google Scholar]
- Sela Y., Molotski N., Golan S., Itskovitz-Eldor J. and Soen Y. (2012). Human embryonic stem cells exhibit increased propensity to differentiate during the G1 phase prior to phosphorylation of retinoblastoma protein. Stem Cells 30, 1097-1108. 10.1002/stem.1078 [DOI] [PubMed] [Google Scholar]
- Shim J., Gururaja-Rao S. and Banerjee U. (2013). Nutritional regulation of stem and progenitor cells in Drosophila. Development 140, 4647-4656. 10.1242/dev.079087 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sinenko S. A., Mandal L., Martinez-Agosto J. A. and Banerjee U. (2009). Dual role of wingless signaling in stem-like hematopoietic precursor maintenance in Drosophila. Dev. Cell 16, 756-763. 10.1016/j.devcel.2009.03.003 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singh A. M., Sun Y., Li L., Zhang W., Wu T., Zhao S., Qin Z. and Dalton S. (2015). Cell-cycle control of bivalent epigenetic domains regulates the exit from pluripotency. Stem Cell Rep. 5, 323-336. 10.1016/j.stemcr.2015.07.005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Small C., Paddibhatla I., Rajwani R. and Govind S. (2012). An introduction to parasitic wasps of Drosophila and the antiparasite immune response. J. Vis. Exp. e3347 10.3791/3347 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Small C., Ramroop J., Otazo M., Huang L. H., Saleque S. and Govind S. (2014). An unexpected link between notch signaling and ROS in restricting the differentiation of hematopoietic progenitors in Drosophila. Genetics 197, 471-483. 10.1534/genetics.113.159210 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stevens A. and Jacobs J. R. (2002). Integrins regulate responsiveness to slit repellent signals. J. Neuro. 22, 4448-4455. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stramer B., Wood W., Galko M. J., Redd M. J., Jacinto A., Parkhurst S. M. and Martin P. (2005). Live imaging of wound inflammation in Drosophila embryos reveals key roles for small GTPases during in vivo cell migration. J. Cell Biol. 168, 567-573. 10.1083/jcb.200405120 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tan K. L., Goh S. C. and Minakhina S. (2012). Genetic screen for regulators of lymph gland homeostasis and hemocyte maturation in Drosophila. G3 2, 393-405. 10.1534/g3.111.001693 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tapias A., Zhou Z.-W., Shi Y., Chong Z., Wang P., Groth M., Platzer M., Huttner W., Herceg Z., Yang Y.-G. et al. (2014). Trrap-dependent histone acetylation specifically regulates cell-cycle gene transcription to control neural progenitor fate decisions. Cell Stem Cell 14, 632-643. 10.1016/j.stem.2014.04.001 [DOI] [PubMed] [Google Scholar]
- Tokusumi T., Shoue D. A., Tokusumi Y., Stoller J. R. and Schulz R. A. (2009). New hemocyte-specific enhancer-reporter transgenes for the analysis of hematopoiesis in Drosophila. Genesis 47, 771-774. 10.1002/dvg.20561 [DOI] [PubMed] [Google Scholar]
- Xu T. and Rubin G. M. (1993). Analysis of genetic mosaics in developing and adult Drosophila tissues. Development 117, 1223-1237. [DOI] [PubMed] [Google Scholar]
- Zhang L., Tran D. T. and Ten Hagen K. G. (2010). An O-glycosyltransferase promotes cell adhesion during development by influencing secretion of an extracellular matrix integrin ligand. J. Biol. Chem. 285, 19491-19501. 10.1074/jbc.M109.098145 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhang C. U., Blauwkamp T. A., Burby P. E. and Cadigan K. M. (2014). Wnt-mediated repression via bipartite DNA recognition by TCF in the Drosophila hematopoietic system. PLoS Genet. 10, e1004509 10.1371/journal.pgen.1004509 [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhou G.-Q., Zhang Y., Ferguson D. J. P., Chen S., Rasmuson-Lestander A., Campbell F. C. and Watt S. M. (2006). The Drosophila ortholog of the endolysosomal membrane protein, endolyn, regulates cell proliferation. J. Cell. Biochem. 99, 1380-1396. 10.1002/jcb.20965 [DOI] [PubMed] [Google Scholar]