

Abstract
BACKGROUND
Insomnia symptoms are associated with vulnerability to age-related morbidity and mortality. Cross-sectional data suggest accelerated biological aging may be a mechanism through which sleep influences risk. A novel method for determining age acceleration using epigenetic methylation to DNA has demonstrated predictive utility as an epigenetic clock and prognostic of age-related morbidity and mortality.
METHODS
We examined the association of epigenetic age and immune cell aging with sleep in the Women’s Health Initiative (WHI) study (N=2,078; Age M(SD)=64.5(7.1) with assessment of insomnia symptoms (restlessness, difficulty falling asleep, waking at night, trouble getting back to sleep, and early awakenings), sleep duration (short-sleep 5 or less; long-sleep >8hrs), epigenetic age, naïve T cell (CD8+CD45RA+CCR7+), and late differentiated T cells (CD8+CD28−CD45RA−).
RESULTS
Insomnia symptoms were related to advanced epigenetic age, B(SE)=1.02(.37), P=0.005, after adjustments for covariates. Insomnia symptoms were also associated with more late differentiated T cells (B(SE)=.59(.21), P=.006), but not with naïve T cells. Self-reported short and long sleep duration were unrelated to epigenetic age. Short sleep, but not long sleep, was associated with fewer naïve T cells (P<.005) and neither were related to late differentiated T cells.
CONCLUSIONS
Symptoms of insomnia were associated with increased epigenetic age of blood tissue, and were associated with higher counts of late differentiated CD8+ T cells. Short sleep was unrelated to epigenetic age and late differentiated cell counts, but was related to a decline in naïve T cells. In this large population based study of women in the United States, insomnia symptoms are implicated in accelerated aging.
Keywords: epigenetic, aging, methylation, insomnia, sleep, immunosenescence
INTRODUCTION
Insomnia symptoms are associated with increased vulnerability to physical and mental declines, increased frailty in the elderly, elevated inflammation, and age related morbidity and mortality (1–9) including risk for coronary heart disease(10). Sleep duration has also been linked to increased risk for disease and death in a U-shaped fashion, such that both short sleepers and long sleepers are at elevated risk(11;12). Thus both sleep duration and insomnia symptoms may have lasting health implications.
Cross-sectional epidemiological data have linked short sleep duration, poor sleep quality, and insomnia to shorter leukocyte telomere length(LTL), a proposed marker of biological aging(13–18), suggesting that inadequate quantity and quality of sleep may accelerate biological aging and be a potential mechanism through which sleep influences disease risk. Even though shortened LTL predicts age-related disease risk, including cancer incidence(19), cardiovascular disease(20;21), and mortality(22–24), LTL is thought to be an incomplete measure of biological aging(25).
An alternative biomarker of aging has recently been developed and is based on DNA methylation (DNAm), referred to as the “epigenetic clock”(26–28). This epigenetic clock method for estimating age is highly correlated with chronological age across cell types and complex tissues(26;29;30). The epigenetic clock is thought to capture aspects of biological age, supported by data demonstrating that the older epigenetic age of blood is predictive of all-cause mortality(31;32), younger epigenetic age relates to cognitive and physical fitness in the elderly(33), and epigenetic age is younger in the offspring of Italian semi-supercentenarians (i.e. subjects aged 105 or older) compared to age-matched controls(34). The epigenetic clock method has been used in applications surrounding obesity(29), Down syndrome(35), HIV infection(36), Parkinson’s disease(37), Alzheimer’s disease related neuropathologies(38), and lung cancer(39). Estimates are that 40% of epigenetic age acceleration is inheritable(26;33), with the remaining 60% thought to be accounted for by unidentified environmental and behavioral contributions. Along this line, initial work has begun to examine the role of environmental factors that may contribute to accelerated aging, with evidence that accelerated epigenetic aging is associate with lifetime stress(40), low socioeconomic status, and psychological trauma(41; 42). However, it is not yet known whether epigenetic age acceleration, using the DNAm based biomarker of aging, relates to measures of sleep disturbances or sleep duration.
We hypothesized that insomnia symptoms and sleep duration would be associated with epigenetic age acceleration among women from the Women’s Health Initiative (WHI) study. Consistent with findings that symptoms of insomnia increase inflammation(1) and morbidity and mortality risk(7–9; 11; 43), we predicted that greater insomnia symptoms would be associated with an older epigenetic age. Similar to findings of sleep duration with mortality(44), we also predicted that both short and long sleep duration would be associated with greater epigenetic aging.
METHODS
Participants
Participants included women in the Women’s Health Initiative (WHI) study, with detailed methods previously published(45–47). Exclusion criterion for the observational study was minimal to assure generalizability. Women were eligible to participate if they were ages 50–79, postmenopausal, willing to provide written informed consent, and resided in a nearby area within proximity of 40 WHI clinical centers across the United States for 3+ years after enrollment. Recruitment for the baseline assessment occurred from 1993–1999. The current analyses include a subset of 2,078 participants who were selected for an integrative genomics study with the aim to identify genomic determinants of coronary heart disease, as reported previously(39). Included in the current study are individuals with both epigenetic and sleep data available at baseline.
DNA methylation profiling
Methylation analysis were performed at HudsonAlpha Institute of Biotechnology using the Illumina Infinium HumanMethylation450 BeadChip, which includes 485,577 different CpG sites, and was described previously(39).
Estimating blood cell counts
We estimate blood cell proportions using the advanced analysis option of the epigenetic clock software(26) available online at (https://labs.genetics.ucla.edu/horvath/dnamage/), which estimates the percentage of late differentiated CD8+ T cells (CD8+CD28−CD45RA−) and the number (count) of naïve CD8+ T cells (CD8+CD45RA+CCR7+)(36). Final counts statistically adjusted for chronological age. Additional cell subsets reported in the supplement.
DNA Methylation Age and the Epigenetic Clock
We estimated the epigenetic age (also known as DNAm age) of each blood sample using two well defined methods. First, we measure extrinsic epigenetic age acceleration (EEAA) that is highly correlated with immune senescence. EEAA is based on the DNAm Age measure proposed in Hannum et al (2013)(28) that relies on 71 CpGs, and is enhanced by forming a weighted average of this with the estimated blood cell counts from three blood cell types that are known to change with age: naive (CD45RA+CCR7+) cytotoxic T cells and late differentiated (CD28−CD45RA−) cytotoxic T cells and plasma B cells using the approach of (48). The (static) weights that are used in the weighted average are determined by the correlation between the respective variables and chronological age in the WHI data (48). By definition, EEAA is positively correlated with the estimated abundance of exhausted CD8+ T cells, plasma B-cells, and is negatively correlated with naive CD8+ T cells. Therefore, the measures of EEAA tracks both age related changes in blood cell composition and intrinsic epigenetic changes.
The second approach uses the Horvath (2013) method, using 353 CpGs and coefficient values, as reported previously(26), to define DNAm age. This measure, which we call intrinsic epigenetic age acceleration (IEAA), rather than using blood cell types to form a weighted average (as EEAA does), adjusts for imputed measures of blood cell counts: naive cytotoxic T cells and late differentiated cytotoxic T cells and plasma B cells. Further detail of these methods and comparisons between EEAA and IEAA can be found in the supplement. Both the EEAA and IEAA measure are expressed as the deviation between DNAm age and chronological age, and are used to define measures of epigenetic age acceleration, which is computed from the residual when regressing DNAm age on chronological age. A positive value indicates that epigenetic age is higher than chronological age.
Measurement of Sleep
Sleep Duration
Subjective reports of sleep duration were obtained at the baseline visit, in which subjects reported how many hours of sleep they got on a typical night during the past 4 weeks. Response options included the following: 5 or less, 6, 7, 8, 9, or 10 or more hours. Consistent with research linking sleep duration with mortality risk(44), and consistent with the newly released joint consensus statement of the American Academy of Sleep Medicine and the Sleep Research Society on the recommended amount of sleep for healthy adults, we created dummy variables to categorize short sleepers as 5 or less (sensitivity analyses categorized as 6 or less given disagreement as to whether the risk for morbidity/mortality is also elevated in this group), normal sleepers as 7–8 hours (the recommended amount of optimal sleep), and long sleepers as >8 hours (identified in the optimal dose of sleep model as elevated risk for disease)(49).
Global sleep disturbances
Subjective reports of sleep disturbances were derived from the Women’s Health Initiative Insomnia Rating Scale (WHIIRS)(50). Five items (0 to 4) from the scale are summed to produce an overall global sleep disturbances score ranging from 0 to 20, with higher scores reflecting greater sleep disturbance and predictive of CVD(10). Previous literature has used an elevated WHIIRS scores of >10 to indicate a significant sleep disturbance(51), which has reasonable specificity for identifying insomnia cases (50; 52).
Insomnia Symptoms
Whereas the WHIIRS reflects a global sleep disturbances assessment, the global score does not provide specificity and sensitivity for detecting discrete insomnia symptoms. Thus, we examined the five individual insomnia symptoms reported in the WHIIRS to create an insomnia symptoms specific measure, allowing us a more sensitive assessment of the relationship between insomnia symptoms and epigenetic age. Individuals were classified as having insomnia symptoms if they reported any one of the following occurring 1 to 2 or more times per week (and in sensitivity analyses occurring 3 or more times a week): restlessness, difficulty falling asleep, difficulty waking during the night, early awakenings, and inability to fall back to sleep. We computed a sum score reflecting the number of insomnia symptoms reported (0 to 5) to estimate severity, and a yes/no score if reporting any insomnia symptom.
Snoring
Self-reported snoring was also included as a covariate in models to adjust for possible sleep apnea, given the predictive utility of this measure in WHI for CVD(53).
Depressive Symptoms
The Center for Epidemiological Studies Depression Scale (CES-D)(54) 6-item version was administered to participants, as was reported previously(45; 55), and is used to control for depressive symptoms.
Statistical Analysis
The measures of epigenetic age and cell counts include adjustment for chronological age (i.e., an estimate of accelerated aging), therefore further adjustment for chronological age in the models was not necessary. Cross-sectional associations of sleep measures with epigenetic age acceleration, naïve CD8+ cell counts, and late differentiated CD8+ cell counts were performed using bi-weight mid-correlation analyses. To test for differences in epigenetic age acceleration between sleep duration categories, sleep disturbances categories, and insomnia symptoms, we used a linear multivariate models. Model 1 adjusts for race/ethnicity (Black/non-Black, Hispanic/non-Hispanic), education (less than high school, high school diploma, some college, Bachelor’s degree, graduate school), and BMI categories (<18.5, 18.5–24.9, 25–29.9, 30–34.9, 35–39.9, 40+) and tested differences in EEAA and IEAA between: 1) any WHIIRS insomnia symptoms (Yes/No), 2) sleep disturbances groups with normal quality sleepers (WHIRS 10 or less) as the reference, and 3) short duration sleepers, long duration sleepers, and normal duration sleepers (reference). In addition to adjusting for Model 1 covariates, Model 2 adjusts for comorbid chronic conditions including diabetes, hypertension, and CVD. Secondary analyses employed pairwise comparisons using the least significant difference (LSD) method to test differences in EEAA between number of WHIIRS insomnia symptoms categories (0, 1–2, 3–4, and 5 symptoms).
Results
Descriptive statistics on demographic data and distribution of sleep measures can be seen in Table 1. Among the sample of 2,078 women, mean age was 64.5 years (SD=7.1), with variations in epigenetic age relative to chronological age (−27.6 to +27.9 years for EEAA; −21.5 to +42.7 years for IEAA. The WHI sample is racially and ethnically diverse with approximately half non-Hispanic white, a third Black, and the remainder of Hispanic ethnicity.
Table 1.
Demographic Characteristics of the WHI Participants
| Variable | N (%) or Mean (SD) |
|---|---|
| Race/ethnicity | |
| White | 989 (47.6%) |
| Black | 668 (32.1%) |
| Hispanic | 421 (20.3%) |
| Chronological Age at Draw | |
| 50–59 | 526 (25.3%) |
| 60–69 | 972 (46.9%) |
| 70–80 | 580 (27.9%) |
| Educational Attainment | |
| Less than High School | 264 (12.7%) |
| High School Diploma | 697 (33.5%) |
| Some College | 523 (25.2%) |
| Bachelor’s Degree | 342 (16.5%) |
| Graduate School | 252 (12.1%) |
| Health Conditions | |
| Cardiovascular Disease | 293 (14.1%) |
| Diabetes | 271 (13%) |
| Hypertension | 907 (43.6%) |
| BMI Category | |
| <18.5 | 13 (.6%) |
| 18.5–24.9 | 443 (21.3%) |
| 25–29.9 | 716 (34.5%) |
| 30–34.9 | 530 (25.5%) |
| 35–39.9 | 246 (11.8%) |
| 40+ | 130 (6.3%) |
| Sleep Duration | |
| < 6 hours | 271 (13.1%) |
| 6 hours | 626 (30.1%) |
| 7–8 hours | 1096 (52.7%) |
| > 8 hours | 85 (4.1%) |
| WHI Insomnia Rating Scale | |
| No Sleep Disturbances (0–10) | 1667 (80.2%) |
| Sleep Disturbance (>10) | 411 (19.8%) |
| Wake up several times at night | |
| No, not in past 4 weeks | 430 (20.7%) |
| Yes, less than once a week | 354 (17.1%) |
| Yes, 1 or 2 times a week | 481 (23.1%) |
| Yes, 3 or 4 times a week | 345 (16.6%) |
| Yes, 5 or more times a week | 468 (22.5%) |
| Wake earlier than planned | |
| No, not in past 4 weeks | 904 (43.5%) |
| Yes, less than once a week | 452 (21.7%) |
| Yes, 1 or 2 times a week | 364 (17.5%) |
| Yes, 3 or 4 times a week | 211 (10.2%) |
| Yes, 5 or more times a week | 147 (7.1%) |
| Trouble getting back to sleep | |
| No, not in past 4 weeks | 1069 (51.4%) |
| Yes, less than once a week | 390 (18.8%) |
| Yes, 1 or 2 times a week | 299 (14.4%) |
| Yes, 3 or 4 times a week | 174 (8.4%) |
| Yes, 5 or more times a week | 146 (7%) |
| Snore | |
| No, not in past 4 weeks | 315 (15.1%) |
| Yes, less than once a week | 97 (4.7%) |
| Yes, 1 or 2 times a week | 102 (4.9%) |
| Yes, 3 or 4 times a week | 111 (5.3%) |
| Yes, 5 or more times a week | 313 (15.1%) |
| Don’t know | 1140 (54.8%) |
| Trouble falling asleep | |
| No, not in past 4 weeks | 1176 (56.6%) |
| Yes, less than once a week | 378 (18.2%) |
| Yes, 1 or 2 times a week | 286 (13.8%) |
| Yes, 3 or 4 times a week | 144 (6.9%) |
| Yes, 5 or more times a week | 94 (4.5%) |
| Restless Sleep | |
| Very restless | 52 (2.5%) |
| Restless | 286 (13.8%) |
| Average quality | 890 (42.8%) |
| Sound or restful | 590 (28.4%) |
| Very sound or restful | 260 (12.5%) |
| Any Insomnia Symptom | |
| Yes | 1706 (82.1%) |
| No | 372 (17.9%) |
| Number of Insomnia Symptom | |
| 0 | 372 (17.9%) |
| 1–2 | 1091 (52.5%) |
| 3–4 | 467 (22.5%) |
| 5 | 148 (7.1%) |
| Epigenetic age | |
| EEAA | .00 (6.4) a |
| IEAA | .00 (5.0) b |
range −27.6 to 27.9 years
range −24.5 to 42.7 years
Insomnia Symptoms, Sleep Duration, and Epigenetic Age Acceleration
Initial unadjusted analyses of differences in EEAA between women with and without WHIIRS insomnia symptoms revealed a significant effect (See Table 2). Further adjustment in Model 1 for race/ethnicity, BMI, educational attainment, and snoring found that the linear coefficient for insomnia symptoms was significant (P=0.005; Table 4). Further adjustment for health conditions in Model 2 did not modify this effect (P=.02). A closer examination of whether the number of insomnia symptoms was related to EEAA suggested that greater difference in epigenetic age correlated with increasing number of WHIIRS insomnia symptoms (P=0.007; See Figure 1) such that the epigenetic age (EEAA) of women with no insomnia symptoms were “younger” than those with 1–2 symptoms (mean difference = 1.17, P=0.002), 3–4 symptoms (mean difference = 1.10, P=0.01), and 5 symptoms (mean difference = 1.77, P=0.005). Further analyses adjusting for depressive symptoms did not modify these results. In addition, we performed sensitivity analyses to examine whether the effect was predominantly from those reporting insomnia symptoms 3 or more times per week, as opposed to 1 to 2 times per week. We found that individuals reporting any insomnia symptoms, either selected if occurring 3 or more times per week or 1–2 times per week or more, exhibited significantly older epigenetic age when compared to those with no insomnia symptoms (P’s<.05).
Table 2.
Point-Biserial Correlations with extrinsic (EEAA) and intrinsic (IEAA) epigenetic age acceleration.
| EEAA | IEAA | |||
|---|---|---|---|---|
|
| ||||
| r | P value | r | P value | |
| Short Sleep Duration (<6 hrs) | −.01 | .73 | −.06 | .06 |
| Long Sleep Duration (9–10 hrs) | −.02 | .46 | −.02 | .50 |
| WHIIRS Sleep disturbance (>10) | .05* | .02 | .01 | .53 |
| Trouble Falling Asleep | .04 | .08 | −.01 | .83 |
| Waking at Night | .07* | .001 | .04 | .09 |
| Waking too early | .03 | .23 | .01 | .64 |
| Trouble going back to sleep | .04 | .10 | −.01 | .69 |
| Snore | .03 | .13 | −.001 | .97 |
| Restless Sleep | .02 | .28 | .02 | .50 |
| Any Insomnia Symptom | .08* | .001 | .03 | .20 |
p<.05
Table 4.
Linear effect model coefficient (B) and standard error (SE) of each sleep characteristic predicting Extrinsic Epigenetic Age (EEAA).
| Independent Predictor | Model 1 | Model 2 | ||
|---|---|---|---|---|
| B(SE) | P value | B(SE) | P value | |
| Sleep Duration | ||||
| Short Sleep (< 6 hours) | −.005(.44) | .99 | −.08(.44) | .86 |
| Normal Sleep (7–8 hours; Reference) | REF | REF | REF | REF |
| Long Sleep (> 8 hours) | −.75(.70) | .29 | −.73(.71) | .31 |
| Sleep Disturbance (WHIIRS >10 vs 10 or less) | .61(.35) | .09 | .64(.36) | .08 |
| Wake at night | 1.00(.35) | .004 | .92(.35) | .008 |
| Restless | .12(.38) | .74 | .11(.38) | .78 |
| Trouble Falling Asleep | .11(.32) | .74 | .07(.32) | .82 |
| Waking too early | .20(.29) | .51 | .19(.29) | .52 |
| Trouble going back to sleep | .20(.31) | .50 | .18(.31) | .57 |
| Any Insomnia Symptom (Yes vs. No) | 1.02(.37) | .005 | .85(.36) | .02 |
Each independent predictor is entered in a separate model with EEAA as the dependent variable. Model 1 adjusts for race (Black vs. non-Black; Hispanic vs. non-Hispanic), education (category), BMI (category), and snore (yes=1). Model 2 adjusts for comorbid chronic conditions: diabetes, hypertension, and CVD.
Figure 1.
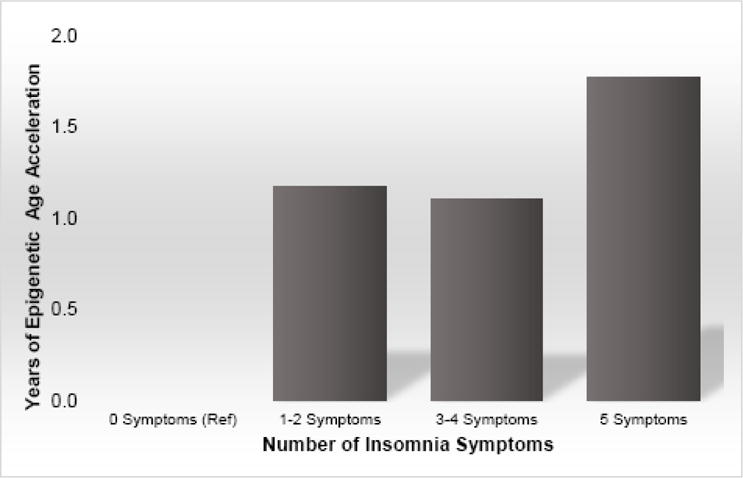
Mean difference in extrinsic epigenetic age acceleration by number of insomnia symptoms.
To characterize whether unique dimensions of insomnia were more strongly related to epigenetic age, we further analyzed the individual insomnia symptoms: trouble going to sleep, frequent waking at night, trouble getting back to sleep, waking earlier than planned, and restless sleep. Older epigenetic age (higher EEAA) was present in women who reported waking in the night compared to women with no night time awakenings, P=0.004. Model 2 adjustments did not alter this effect, P=0.008. Women reporting waking at night were on average one year older epigenetically than those without night time awakenings. The other insomnia symptoms alone did not predict epigenetic age acceleration (See Table 4).
Examination of EEAA with the global WHIIRS sleep disturbance scores, cutoff >10, revealed differences in women with elevated WHIIRS scores (M=.573,n=399) compared to women with scores 10 or less (M=−.183, n=1,615), P<0.05. Linear effect Model 1 (See Table 4) adjusted for race/ethnicity, BMI, educational attainment, and snoring, and Model 2 further adjusted for health conditions, with subsequent reduction in the strength of the effect of sleep disturbances, P=0.09 and P=0.08 respectively. Additional adjustment for depressive symptoms did not modify these results, P=0.10.
Analyses examining differences in EEAA by short sleep and long sleep compared to normal sleep duration found no significant effects, P’s>0.10 (See Tables 2 and 4). Further sensitivity analyses tested whether categorization of short sleep as 6 hours per night or less would generate different findings. Sleeping 6 hrs or less compared to 7–8 hrs was unrelated to EEAA, P=.95. Given findings by Vgontzas and colleagues that demonstrate a significant increased risk for mortality among short sleepers with insomnia symptoms (8), further analyses examined the interaction of short sleep (6 or less; 5 or less) with insomnia symptoms. There were no significant interaction effects (P’s>.4). Analyses examining associations of sleep duration and disturbances with IEAA found no significant differences in relation to insomnia symptoms, nor extremes of sleep duration. See Supplemental Table S1 for model coefficients and P values.
Insomnia Symptoms, Sleep Duration, and Cell Subtypes: Naïve and Late Differentiated T Cells
Analyses of CD8+ naïve cell with sleep dimensions were performed. While WHIIRS insomnia symptoms were not associated with naïve CD8+ cells, short sleep and trouble falling asleep were significantly associated with fewer naïve CD8+ cells (P’s<0.05; See Table 3).
Table 3.
Point-Biserial Correlations with age adjusted naïve and late differentiated subsets.
| Naïve: CD8+CD45RA+CCR7+ | Late Differentiated: CD8+CD28−CD45RA− | |||
|---|---|---|---|---|
|
| ||||
| r | P value | r | P value | |
| Short Sleep Duration (<6 hrs) | −.07 | .008 | .0 | .29 |
| Long Sleep Duration (9–10 hrs) | .03 | .39 | .04 | .22 |
| WHIIRS Sleep disturbance (>10) | −.03 | .14 | .06 | .005 |
| Trouble Falling Asleep | −.05 | .03 | .07 | .003 |
| Waking at Night | −.01 | .58 | .05 | .02 |
| Waking too early | −.03 | .25 | .03 | .17 |
| Trouble going back to sleep | −.03 | .12 | .05 | .03 |
| Snore | −.001 | .97 | .02 | .41 |
| Restless Sleep | −.03 | .19 | .01 | .65 |
| Any Insomnia Symptom | −.03 | .18 | .08 | <.001 |
CD8+CD28−CD45− late differentiated T cells were significantly positively correlated with WHIIRS global sleep disturbance scores >10, trouble falling asleep, waking at night, trouble going back to sleep, and having any insomnia symptom (All P’s<0.05; See Table 3). The findings were such that having a high global sleep disturbance score, discrete insomnia symptoms, or any insomnia symptom was related to a greater amount of these late differentiated, exhausted T cells in circulating blood. Sleep duration was unrelated to the late differentiated T cell subtype (See Table 3).
In subsequent adjusted models, reporting any insomnia symptoms continued to be significantly associated with having more CD8+CD28−CD45RA− T cells, after adjustments for race/ethnicity, BMI, educational attainment, snoring, and health conditions, B(SE)=.59(.21), P=0.006. Further adjustment for depressive symptoms had no effect on these results, P=0.008. Examination of each insomnia symptom, adjusting for model 1 and 2 covariates, revealed modest effects for trouble falling asleep, B(SE)=.32(.19), P=0.09, waking at night B(SE)=.40(.20), P=0.05, and going back to sleep, B(SE)=.34(.18), P=0.06. Linear effect models, adjusting for race/ethnicity, BMI, educational attainment, snoring, and health conditions demonstrated the significant associations of the WHIIRS measure of global sleep disturbances with CD8+CD28−CD45RA− was retained, B(SE)=.41(.21), P=0.05.
Discussion
Overall, we find modest correlations between self-reported insomnia symptoms and measures of epigenetic age acceleration in peripheral blood cells. Insomnia symptoms were significantly associated with the extrinsic measure of age acceleration, which measures the age of the immune system. Moreover, and consistent with the findings with extrinsic age acceleration, we report that insomnia symptoms were significantly associated with a greater proportion of late differentiated cytotoxic T cells (CD8+CD28−CD45RA−), indicative of an aged immune system(56). Insomnia symptoms did not seem to relate to intrinsic age acceleration, which is independent of age related changes in blood cell counts. More careful examination of insomnia symptoms revealed that the association with the extrinsic measure of epigenetic age was graded; increasing number of insomnia symptoms was associated with an older epigenetic age, in which those reporting 5 symptoms had the oldest epigenetic age compared to women with no insomnia symptoms (See Figure 1). This effect was present after adjustment for potential confounds including race/ethnicity, BMI, educational attainment, snoring, depressive symptoms, and health conditions. When we analyzed whether specific dimensions of insomnia symptoms uniquely related to epigenetic age acceleration, we found that self-reported waking at night was significantly related to an older epigenetic age, more so than other insomnia symptoms reported. In parallel to the findings with the number of insomnia symptoms, the global measure of sleep disturbances showed a more modest relationship with epigenetic age, suggesting use of the WHIIRS cutoff >10, which has high specificity for identifying insomnia cases, is a less sensitive measure than examination of the cumulative effect of individual insomnia symptoms in the context of age acceleration.
In addition, we found that late differentiated T cells, which are in a senescent or near senescent state, were higher in those reporting insomnia symptoms. This finding suggests poor sleep may accelerate CD8+ cell replication and progression to cellular senescence. This is in line with findings of Prather and colleagues(17), reporting marked telomere shortening in CD8+ T cells with sleep disturbances. Importantly, CD8+CD28− T cells have noticeably shorter telomere length and reduced telomerase activity, significantly impaired replicative capacity, and are at an increased proportion in circulation among those with increasing chronological age, individuals who are HIV+ (an accelerated aging disease)(36) and those with diseases and conditions of later life(56–58). Likewise, telomeric shortening within CD8+CD28− cells was predictive of susceptibility to a novel virus, further implicating this cell subset in cellular aging and immune compromise(59). Taken together, our results advocate that experiencing sleep disturbances is associated with accelerated aging, particularly within the immune system.
In contrast, our hypothesis that extremes of sleep duration would be related to an accelerated epigenetic age was not supported by the data. We found no significant effects of short or long sleep on extrinsic or intrinsic epigenetic age. There was a trend for a difference in intrinsic epigenetic age to be associated with short- as compared to normal sleep duration, although this did not reach statistical significance (Table 2a). We did see a significant inverse association of naïve T cell counts with short sleep duration (Table 2b), indicating a reduction in the number of naïve T cells in the T-cell compartment among those with short sleep. These findings have implications for early immune responses to novel antigens, such as susceptibility to a new cold or flu virus or in response to vaccinations(60), which in part contributes to reduced immune competence seen in late life. Indeed, recent evidence has linked short sleep with an increased risk for developing the common cold after exposure(61).
A lack of association of epigenetic age acceleration with extremes of sleep duration is contrasted to the considerable epidemiological data linking short and long sleep with morbidity and mortality outcomes(11;12). Our null results may be due to measurement error of sleep duration. Considerable differences exist between sleep duration obtained when assessed by objective measures such as actigraphy as compared to self-reports of sleep duration, as used in the current report. Indeed, the correlation between objective and subjective measures are r=.45, with average self-report being. 8 hrs more than objectively recorded sleep duration(62). Future research should address this question using an objective assessment of sleep duration over repeated samplings and not rely solely on retrospective self-report of sleep duration.
In contrast to self-reported sleep duration, self-reported insomnia symptoms using the WHIIRS has been demonstrated to have test-retest reliability and construct validity compared to objective indicators of sleep(52). Although objectively derived assessments of waking after sleep onset are likely to capture more nuanced information about frequency and length of waking, self-reports of waking during the night, waking early, or difficulty falling asleep are all subjective self-reports of insomnia symptoms and are considered valid methods in epidemiological studies.
These cross-sectional results need to be replicated in a prospective design to determine causation, but suggest that sleep disturbances may increase risk for morbidity and mortality by accelerating the rate at which the immune, and possibly other bodily systems, age. Should replication and prospective designs support these initial findings, then this raises the possibility that maintenance of sleep health (i.e., absence of sleep disturbance) slow the aging process and is vital for the prevention of early onset of disease and premature death(63). Alternatively, persistent insomnia symptoms may accelerate aging(64) by disrupting healthy cellular function, increasing energy demands, promoting DNA damage, inflammation, and cellular senescence(65–71), which may lead to early mortality(64). Together these biological alterations result in epigenetic modifications that appear with more advanced cellular age, which supports the hypothesis that DNAm age captures the cumulative work of the epigenomic maintenance system(29).
There are several limitations to the current findings. First, the results are limited by the cross-sectional design, and an observation of associations between variables does not provide evidence of causality. Future studies should explore the alternative hypothesis that older epigenetic age is a driver of disruptions in sleep. Disruptions in circadian patterns with age may be the consequence of biological aging of the suprachiasmatic nucleus (SCN), the circadian clock of the body(72). Along this line, aging is associated with alterations in many aspects of the SCN that influence sleep timing, duration, rhythm, depth, and quality. Therefore the direction of this association cannot be determined in the present results. However, we did not find associations of sleep disturbances with IEAA (an indicator of more global aging), but rather primarily with EEAA, an indicator of immune system aging. Likewise, there are several potential unidentified confounds that might influence the data, including unmeasured subclinical disease. As mentioned previously, the current results are limited by self-report measures of sleep taken from one sampling time point, and may not be representative of long term sleep patterns, adding error to the measure. Future research should establish repeated assessments of objectively defined sleep characteristics as they relate to changes in biological aging over time. Likewise, although our findings point to a link between insomnia symptoms and epigenetic age, the effect sizes are small; further, caution should be taken when interpreting these cross-sectional results where directionality and causation is not tested. The current sample was comprised of women only, and is not representative of males in the population. Finally, we focused on blood tissue. Here we did see associations of insomnia symptoms with alterations in cell subsets representative of aging immunity including reductions in naïve cytotoxic T cells and increases in late differentiated/exhausted T cells. Future studies should explore whether sleep is associated with the epigenetic ages of other tissues outside of the immune system. Physiological factors appear to have tissue specific aging effects, e.g. obesity is strongly correlated with epigenetic age acceleration of liver tissue but much less so in blood(29).
Several strengths of the study design should also be noted, particularly the high quality of the WHI study design, measurements, and sample collection, and the diverse sampling of women across various strata of the United States of America. The study is well powered to test the hypotheses. The study also included a validated measure of sleep, the WHIIRS. Another notable strength is the highly accurate biomarkers of epigenetic age as mentioned in the introduction.
Conclusions
In summary, insomnia symptoms were significantly associated with a marker of epigenetic age acceleration in a sample of over two-thousand women from the WHI study, representing the first findings to date to document insomnia symptoms to be associated with the “epigenetic clock”. Given that sleep disturbances are also associated with increase vulnerability to physical and mental declines, and age related morbidity and mortality risk(2–10), these findings raise the possibility that accelerated epigenetic aging might serve as one of several mechanisms through which sleep disturbances influence risk for age related disease and early mortality. These findings are in concordance with work examining sleep disturbances, including insomnia(18), and blood cell telomere length(13;15–17), and extend the findings to link insomnia symptoms with accelerated epigenetic aging and evidence of an aged immune system. Given the recognized importance of aging biology for many chronic diseases seen in later life(73;74) and the growing demand to address the needs of an aging population, this work provides initial evidence that addressing behavioral factors such as sleep disturbances, which are prevalent in older adults(2), may promote an extension of healthspan, more years of healthy living without chronic disease in later life(74;75).
Supplementary Material
Acknowledgments
This study was supported by the UCLA Cousins Center for Psychoneuroimmunology; NIH/NIA K01 AG044462 (Carroll); NIH/NHLBI 60442456 BAA23 (Assimes, Absher, Horvath); National Institutes of Health NIH/NIA 5R01AG042511–02 (Horvath and Levine); NIH/NIA R01 AG034588 and R01 AG026364, NIH/NCI R01 CA160245, NIH/NIDA R01 DA032922, NIH/NHLBI R01 HL095799-01 (Irwin), P30 AG017265, R24AG037898 (Seeman). The WHI program is funded by the National Heart, Lung, and Blood Institute, National Institutes of Health, U.S. Department of Health and Human Services through contracts HHSN268201100046C, HHSN268201600003C, HHSN268201600002C, HHSN268201600004C, HHSN268201600001C, and HHSN271201100004C.
Footnotes
Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain.
Recognition of the team of WHI Investigators include the following:
Program Office: (National Heart, Lung, and Blood Institute, Bethesda, Maryland) Jacques Rossouw, Shari Ludlam, Dale Burwen, Joan McGowan, Leslie Ford, and Nancy Geller
Clinical Coordinating Center: Clinical Coordinating Center: (Fred Hutchinson Cancer Research Center, Seattle, WA) Garnet Anderson, Ross Prentice, Andrea LaCroix, and Charles Kooperberg
Investigators and Academic Centers: (Brigham and Women’s Hospital, Harvard Medical School, Boston, MA) JoAnn E. Manson; (MedStar Health Research Institute/Howard University, Washington, DC) Barbara V. Howard; (Stanford Prevention Research Center, Stanford, CA) Marcia L. Stefanick; (The Ohio State University, Columbus, OH) Rebecca Jackson; (University of Arizona, Tucson/Phoenix, AZ) Cynthia A. Thomson; (University at Buffalo, Buffalo, NY) Jean Wactawski-Wende; (University of Florida, Gainesville/Jacksonville, FL) Marian Limacher; (University of Iowa, Iowa City/Davenport, IA) Robert Wallace; (University of Pittsburgh, Pittsburgh, PA) Lewis Kuller; (Wake Forest University School of Medicine, Winston-Salem, NC) Sally Shumaker
Women’s Health Initiative Memory Study: (Wake Forest University School of Medicine, Winston-Salem, NC) Sally Shumaker
FINANCIAL DISCLOSURES
All authors report no biomedical financial interests or potential conflicts of interest.
References
- 1.Irwin MR, Olmstead R, Carroll JE. Sleep Disturbance, Sleep Duration, and Inflammation: A Systematic Review and Meta-Analysis of Cohort Studies and Experimental Sleep Deprivation. Biol Psychiatry. 2015 doi: 10.1016/j.biopsych.2015.05.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Ohayon MM. Epidemiology of insomnia: what we know and what we still need to learn. Sleep Med Rev. 2002;6:97–111. doi: 10.1053/smrv.2002.0186. [DOI] [PubMed] [Google Scholar]
- 3.Center for Disease Control. CDC Data & Statistics | Feature: Insufficient Sleep Is a Public Health Epidemic. 2011 Retrieved August 4, 2011, from http://www.cdc.gov/Features/dsSleep/
- 4.Colten HR, Altevogt BM. Sleep disorders and sleep deprivation: an unmet public health problem. National Academies Press; 2006. [PubMed] [Google Scholar]
- 5.Motivala SJ. Sleep and Inflammation: Psychoneuroimmunology in the Context of Cardiovascular Disease. Ann Behav Med. 2011;42:141–152. doi: 10.1007/s12160-011-9280-2. [DOI] [PubMed] [Google Scholar]
- 6.Ensrud KE, Blackwell TL, Ancoli-Israel S, Redline S, Cawthon PM, Paudel ML, et al. Sleep disturbances and risk of frailty and mortality in older men. Sleep Med. 2012;13:1217–25. doi: 10.1016/j.sleep.2012.04.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Li Y, Zhang X, Winkelman JW, Redline S, Hu FB, Stampfer M, et al. Association between insomnia symptoms and mortality: a prospective study of U.S. men. Circulation. 2014;129:737–46. doi: 10.1161/CIRCULATIONAHA.113.004500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Vgontzas AN, Liao D, Pejovic S, Calhoun S, Karataraki M, Basta M, et al. Insomnia with short sleep duration and mortality: the Penn State cohort. Sleep. 2010;33:1159–64. doi: 10.1093/sleep/33.9.1159. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Althuis MD, Fredman L, Langenberg PW, Magaziner J. The relationship between insomnia and mortality among community-dwelling older women. J Am Geriatr Soc. 1998;46:1270–3. doi: 10.1111/j.1532-5415.1998.tb04544.x. [DOI] [PubMed] [Google Scholar]
- 10.Sands-Lincoln M, Loucks EB, Lu B, Carskadon MA, Sharkey K, Stefanick ML, et al. Sleep duration, insomnia, and coronary heart disease among postmenopausal women in the Women’s Health Initiative. J Womens Health (Larchmt) 2013;22:477–86. doi: 10.1089/jwh.2012.3918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Carroll JE, Irwin MR, Merkin SS, Seeman TE. Sleep and multisystem biological risk: a population-based study. PLoS One. 2015;10:e0118467. doi: 10.1371/journal.pone.0118467. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Cappuccio FP, Cooper D, D’Elia L, Strazzullo P, Miller MA. Sleep duration predicts cardiovascular outcomes: a systematic review and meta-analysis of prospective studies. Eur Heart J. 2011;32:1484–92. doi: 10.1093/eurheartj/ehr007. [DOI] [PubMed] [Google Scholar]
- 13.Cribbet MR, Carlisle M, Cawthon RM, Uchino BN, Williams PG, Smith TW, et al. Cellular aging and restorative processes: subjective sleep quality and duration moderate the association between age and telomere length in a sample of middle-aged and older adults. Sleep. 2014;37:65–70. doi: 10.5665/sleep.3308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Jackowska M, Hamer M, Carvalho LA, Erusalimsky JD, Butcher L, Steptoe A. Short Sleep Duration Is Associated with Shorter Telomere Length in Healthy Men: Findings from the Whitehall II Cohort Study. In: Kiechl S, editor. PLoS One. Vol. 7. 2012. p. e47292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Prather AA, Puterman E, Lin J, O’Donovan A, Krauss J, Tomiyama AJ, et al. Shorter leukocyte telomere length in midlife women with poor sleep quality. J Aging Res. 2011;2011:721390. doi: 10.4061/2011/721390. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Liang G, Schernhammer E, Qi L, Gao X, De Vivo I, Han J. Associations between Rotating Night Shifts, Sleep Duration, and Telomere Length in Women. PLoS One. 2011;6:e23462. doi: 10.1371/journal.pone.0023462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Prather AA, Gurfein B, Moran P, Daubenmier J, Acree M, Bacchetti P, et al. Tired telomeres: Poor global sleep quality, perceived stress, and telomere length in immune cell subsets in obese men and women. Brain Behav Immun. 2015;47:155–162. doi: 10.1016/j.bbi.2014.12.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Carroll JE, Esquivel S, Goldberg A, Seeman TE, Effros RB, Dock J, et al. Insomnia and Telomere Length in Older Adults. Sleep. 2016;39:559–564. doi: 10.5665/sleep.5526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Willeit P, Willeit J, Mayr A, Weger S, Oberhollenzer F, Brandstätter A, et al. Telomere length and risk of incident cancer and cancer mortality. JAMA. 2010;304:69–75. doi: 10.1001/jama.2010.897. [DOI] [PubMed] [Google Scholar]
- 20.Farzaneh-Far R, Cawthon RM, Na B, Browner WS, Schiller NB, Whooley MA. Prognostic value of leukocyte telomere length in patients with stable coronary artery disease: data from the Heart and Soul Study. Arterioscler Thromb Vasc Biol. 2008;28:1379–84. doi: 10.1161/ATVBAHA.108.167049. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Fitzpatrick AL, Kronmal RA, Gardner JP, Psaty BM, Jenny NS, Tracy RP, et al. Leukocyte telomere length and cardiovascular disease in the cardiovascular health study. Am J Epidemiol. 2007;165:14–21. doi: 10.1093/aje/kwj346. [DOI] [PubMed] [Google Scholar]
- 22.Bakaysa SL, Mucci LA, Slagboom PE, Boomsma DI, McClearn GE, Johansson B, Pedersen NL. Telomere length predicts survival independent of genetic influences. Aging Cell. 2007;6:769. doi: 10.1111/j.1474-9726.2007.00340.x. [DOI] [PubMed] [Google Scholar]
- 23.Cawthon RM, Smith KR, O’Brien E, Sivatchenko A, Kerber RA. Association between telomere length in blood and mortality in people aged 60 years or older. Lancet. 2003;361:393–5. doi: 10.1016/S0140-6736(03)12384-7. [DOI] [PubMed] [Google Scholar]
- 24.Epel ES, Merkin SS, Cawthon R, Blackburn EH, Adler NE, Pletcher MJ, Seeman TE. The rate of leukocyte telomere shortening predicts mortality from cardiovascular disease in elderly men. Aging (Albany NY) 2009;1:81–8. doi: 10.18632/aging.100007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Sanders JL, Newman AB. Telomere Length in Epidemiology: A Biomarker of Aging, Age-Related Disease, Both, or Neither? Epidemiol Rev. 2013;35:112–131. doi: 10.1093/epirev/mxs008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Horvath S. DNA methylation age of human tissues and cell types. Genome Biol. 2013;14:R115. doi: 10.1186/gb-2013-14-10-r115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Bocklandt S, Lin W, Sehl ME, Sánchez FJ, Sinsheimer JS, Horvath S, Vilain E. Epigenetic predictor of age. PLoS One. 2011;6:e14821. doi: 10.1371/journal.pone.0014821. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Hannum G, Guinney J, Zhao L, Zhang L, Hughes G, Sadda S, et al. Genome-wide methylation profiles reveal quantitative views of human aging rates. Mol Cell. 2013;49:359–67. doi: 10.1016/j.molcel.2012.10.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Horvath S, Erhart W, Brosch M, Ammerpohl O, von Schönfels W, Ahrens M, et al. Obesity accelerates epigenetic aging of human liver. Proc Natl Acad Sci U S A. 2014;111:15538–43. doi: 10.1073/pnas.1412759111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Horvath S, Mah V, Lu AT, Woo JS, Choi O-W, Jasinska AJ, et al. The cerebellum ages slowly according to the epigenetic clock. Aging (Albany NY) 2015;7:294–306. doi: 10.18632/aging.100742. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Marioni RE, Shah S, McRae AF, Chen BH, Colicino E, Harris SE, et al. DNA methylation age of blood predicts all-cause mortality in later life. Genome Biol. 2015;16:25. doi: 10.1186/s13059-015-0584-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Christiansen L, Lenart A, Tan Q, Vaupel JW, Aviv A, McGue M, Christensen K. DNA methylation age is associated with mortality in a longitudinal Danish twin study. Aging Cell. 2015;15:n/a–n/a. doi: 10.1111/acel.12421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Marioni RE, Shah S, McRae AF, Ritchie SJ, Muniz-Terrera G, Harris SE, et al. The epigenetic clock is correlated with physical and cognitive fitness in the Lothian Birth Cohort 1936. Int J Epidemiol. 2015;44:1388–96. doi: 10.1093/ije/dyu277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Horvath S, Pirazzini C, Bacalini MG, Gentilini D, Di Blasio AM, Delledonne M, et al. Decreased epigenetic age of PBMCs from Italian semi-supercentenarians and their offspring. Aging (Albany NY) 2015 doi: 10.18632/aging.100861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Horvath S, Garagnani P, Bacalini MG, Pirazzini C, Salvioli S, Gentilini D, et al. Accelerated epigenetic aging in Down syndrome. Aging Cell. 2015;14:491–5. doi: 10.1111/acel.12325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Horvath S, Levine AJ. HIV-1 Infection Accelerates Age According to the Epigenetic Clock. J Infect Dis. 2015;212:1563–73. doi: 10.1093/infdis/jiv277. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Horvath S, Ritz BR. Increased epigenetic age and granulocyte counts in the blood of Parkinson’s disease patients. Aging (Albany NY) 2015 doi: 10.18632/aging.100859. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Levine ME, Lu AT, Bennett DA, Horvath S. Epigenetic age of the pre-frontal cortex is associated with neuritic plaques, amyloid load, and Alzheimer’s disease related cognitive functioning. Aging (Albany NY) 2015 doi: 10.18632/aging.100864. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Levine ME, Hosgood HD, Chen B, Absher D, Assimes T, Horvath S. DNA methylation age of blood predicts future onset of lung cancer in the women’s health initiative. Aging (Albany NY) 2015;7:690–700. doi: 10.18632/aging.100809. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Zannas AS, Arloth J, Carrillo-Roa T, Iurato S, Röh S, Ressler KJ, et al. Lifetime stress accelerates epigenetic aging in an urban, African American cohort: relevance of glucocorticoid signaling. Genome Biol. 2015;16:266. doi: 10.1186/s13059-015-0828-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Miller GE, Yu T, Chen E, Brody GH. Self-control forecasts better psychosocial outcomes but faster epigenetic aging in low-SES youth. Proc Natl Acad Sci U S A. 2015;112:10325–30. doi: 10.1073/pnas.1505063112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Boks MP, van Mierlo HC, Rutten BPF, Radstake TRDJ, De Witte L, Geuze E, et al. Longitudinal changes of telomere length and epigenetic age related to traumatic stress and post-traumatic stress disorder. Psychoneuroendocrinology. 2015;51:506–12. doi: 10.1016/j.psyneuen.2014.07.011. [DOI] [PubMed] [Google Scholar]
- 43.Carroll JE, Seeman TE, Olmstead R, Melendez G, Sadakane R, Bootzin R, et al. Improved sleep quality in older adults with insomnia reduces biomarkers of disease risk: Pilot results from a randomized controlled comparative efficacy trial. Psychoneuroendocrinology. 2015;55:184–92. doi: 10.1016/j.psyneuen.2015.02.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Cappuccio FP, D’Elia L, Strazzullo P, Miller MA. Sleep duration and all-cause mortality: a systematic review and meta-analysis of prospective studies. Sleep. 2010;33:585–92. doi: 10.1093/sleep/33.5.585. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Design of the Women’s Health Initiative clinical trial and observational study. The Women’s Health Initiative Study Group. Control Clin Trials. 1998;19:61–109. doi: 10.1016/s0197-2456(97)00078-0. [DOI] [PubMed] [Google Scholar]
- 46.Hays J, Hunt JR, Hubbell FA, Anderson GL, Limacher M, Allen C, Rossouw JE. The Women’s Health Initiative recruitment methods and results. Ann Epidemiol. 2003;13:S18–77. doi: 10.1016/s1047-2797(03)00042-5. [DOI] [PubMed] [Google Scholar]
- 47.Langer RD, White E, Lewis CE, Kotchen JM, Hendrix SL, Trevisan M. The Women’s Health Initiative Observational Study: baseline characteristics of participants and reliability of baseline measures. Ann Epidemiol. 2003;13:S107–21. doi: 10.1016/s1047-2797(03)00047-4. [DOI] [PubMed] [Google Scholar]
- 48.Klemera P, Doubal S. A new approach to the concept and computation of biological age. Mech Ageing Dev. 2006;127:240–8. doi: 10.1016/j.mad.2005.10.004. [DOI] [PubMed] [Google Scholar]
- 49.Watson NF, Badr MS, Belenky G, Bliwise DL, Buxton OM, Buysse D, et al. Joint Consensus Statement of the American Academy of Sleep Medicine and Sleep Research Society on the Recommended Amount of Sleep for a Healthy Adult: Methodology and Discussion. Sleep. 2015;38:1161–83. doi: 10.5665/sleep.4886. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Levine DW, Kripke DF, Kaplan RM, Lewis MA, Naughton MJ, Bowen DJ, Shumaker SA. Reliability and validity of the Women’s Health Initiative Insomnia Rating Scale. Psychol Assess. 2003;15:137–48. doi: 10.1037/1040-3590.15.2.137. [DOI] [PubMed] [Google Scholar]
- 51.Chen X, Wang R, Zee P, Lutsey PL, Javaheri S, Alcántara C, et al. Racial/Ethnic Differences in Sleep Disturbances: The Multi-Ethnic Study of Atherosclerosis (MESA) Sleep. 2015;38:877–88. doi: 10.5665/sleep.4732. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Levine DW, Kaplan RM, Kripke DF, Bowen DJ, Naughton MJ, Shumaker SA. Factor structure and measurement invariance of the Women’s Health Initiative Insomnia Rating Scale. Psychol Assess. 2003;15:123–36. doi: 10.1037/1040-3590.15.2.123. [DOI] [PubMed] [Google Scholar]
- 53.Sands M, Loucks EB, Lu B, Carskadon MA, Sharkey K, Stefanick M, et al. Self-reported snoring and risk of cardiovascular disease among postmenopausal women (from the Women’s Health Initiative) Am J Cardiol. 2013;111:540–6. doi: 10.1016/j.amjcard.2012.10.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Radloff LS. The CES-D Scale: A Self-Report Depression Scale for Research in the General Population. Appl Psychol Meas. 1977;1:385–401. [Google Scholar]
- 55.Wassertheil-Smoller S, Shumaker S, Ockene J, Talavera GA, Greenland P, Cochrane B, et al. Depression and cardiovascular sequelae in postmenopausal women. The Women’s Health Initiative (WHI) Arch Intern Med. 2004;164:289–98. doi: 10.1001/archinte.164.3.289. [DOI] [PubMed] [Google Scholar]
- 56.Arosa FA. CD8+CD28− T cells: certainties and uncertainties of a prevalent human T-cell subset. Immunol Cell Biol. 2002;80:1–13. doi: 10.1046/j.1440-1711.2002.01057.x. [DOI] [PubMed] [Google Scholar]
- 57.Lin J, Epel E, Cheon J, Kroenke C, Sinclair E, Bigos M, et al. Analyses and comparisons of telomerase activity and telomere length in human T and B cells: insights for epidemiology of telomere maintenance. J Immunol Methods. 2010;352:71–80. doi: 10.1016/j.jim.2009.09.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Effros RB. The role of CD8 T cell replicative senescence in human aging. Discov Med. 2005;5:293–7. [PubMed] [Google Scholar]
- 59.Cohen S. Association Between Telomere Length and Experimentally Induced Upper Respiratory Viral Infection in Healthy Adults<alt-title>Telomere Length and Respiratory Viral Infection</alt-title>. JAMA. 2013;309:699. doi: 10.1001/jama.2013.613. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Prather AA, Hall M, Fury JM, Ross DC, Muldoon MF, Cohen S, Marsland AL. Sleep and antibody response to hepatitis B vaccination. Sleep. 2012;35:1063–9. doi: 10.5665/sleep.1990. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Prather AA, Janicki-Deverts D, Hall MH, Cohen S. Behaviorally Assessed Sleep and Susceptibility to the Common Cold. Sleep. 2015;38:1353–9. doi: 10.5665/sleep.4968. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Lauderdale DS, Knutson KL, Yan LL, Liu K, Rathouz PJ. Self-reported and measured sleep duration: how similar are they? Epidemiology. 2008;19:838–45. doi: 10.1097/EDE.0b013e318187a7b0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Buysse DJ. Sleep health: can we define it? Does it matter? Sleep. 2014;37:9–17. doi: 10.5665/sleep.3298. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 64.Parthasarathy S, Vasquez MM, Halonen M, Bootzin R, Quan SF, Martinez FD, Guerra S. Persistent insomnia is associated with mortality risk. Am J Med. 2015;128:268–75.e2. doi: 10.1016/j.amjmed.2014.10.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Carroll JE, Cole SW, Seeman TE, Breen EC, Witarama T, Arevalo JMG, et al. Partial Sleep Deprivation Activates the DNA Damage Response (DDR) and the Senescence-Associated Secretory Phenotype (SASP) in Aged Adult Humans. Brain Behav Immun. 2015 doi: 10.1016/j.bbi.2015.08.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Irwin MR, Wang M, Campomayor CO, Collado-Hidalgo A, Cole S. Sleep deprivation and activation of morning levels of cellular and genomic markers of inflammation. Arch Intern Med. 2006;166:1756–62. doi: 10.1001/archinte.166.16.1756. 2006/09/20 ed. [DOI] [PubMed] [Google Scholar]
- 67.Everson CA, Henchen CJ, Szabo A, Hogg N. Cell injury and repair resulting from sleep loss and sleep recovery in laboratory rats. Sleep. 2014;37:1929–40. doi: 10.5665/sleep.4244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Naidoo N. Cellular stress/the unfolded protein response: relevance to sleep and sleep disorders. Sleep Med Rev. 2009;13:195–204. doi: 10.1016/j.smrv.2009.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Naidoo N. Roles of Endoplasmic Reticulum and Energetic Stress in Disturbed Sleep. Neuromolecular Med. 2012 doi: 10.1007/s12017-012-8179-9. 2012/04/25 ed. [DOI] [PubMed] [Google Scholar]
- 70.Naidoo N, Ferber M, Master M, Zhu Y, Pack AI. Aging impairs the unfolded protein response to sleep deprivation and leads to proapoptotic signaling. J Neurosci. 2008;28:6539–48. doi: 10.1523/JNEUROSCI.5685-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Brown MK, Naidoo N. The UPR and the anti-oxidant response: relevance to sleep and sleep loss. Mol Neurobiol. 2010;42:103–13. doi: 10.1007/s12035-010-8114-8. [DOI] [PubMed] [Google Scholar]
- 72.Hofman MA, Swaab DF. Living by the clock: The circadian pacemaker in older people. Ageing Res Rev. 2006;5:33–51. doi: 10.1016/j.arr.2005.07.001. [DOI] [PubMed] [Google Scholar]
- 73.Franceschi C, Campisi J. Chronic inflammation (inflammaging) and its potential contribution to age-associated diseases. J Gerontol A Biol Sci Med Sci. 2014;69(Suppl 1):S4–9. doi: 10.1093/gerona/glu057. [DOI] [PubMed] [Google Scholar]
- 74.Kennedy BK, Berger SL, Brunet A, Campisi J, Cuervo AM, Epel ES, et al. Geroscience: Linking Aging to Chronic Disease. Cell. 2014;159:709–713. doi: 10.1016/j.cell.2014.10.039. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Burch JB, Augustine AD, Frieden LA, Hadley E, Howcroft TK, Johnson R, et al. Advances in geroscience: impact on healthspan and chronic disease. J Gerontol A Biol Sci Med Sci. 2014;69(Suppl 1):S1–3. doi: 10.1093/gerona/glu041. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.