

Abstract
Behavioral treatment gains established in one setting do not always maintain in other settings. The present review examines the relevance of basic and translational research to understanding failures to maintain treatment gains across settings. Specifically, studies of the renewal effect examine how transitioning away from a treatment setting could evoke a return of undesirable behavior, rather than newly trained appropriate behavior. Studies of renewal typically arrange three phases, with a response trained and reinforced under a particular set of contextual stimuli in the first phase. Next, that response is extinguished, often under a different set of contextual stimuli. Finally, that response returns despite extinction remaining in effect upon returning to the original training context or transitioning to a novel context. Thus, removing the extinction context is sufficient to produce a recurrence of the response. The findings suggest treatment effects can become specific to the context in which the treatment was delivered. This literature offers promising methods for systematically assessing the factors contributing to treatment maintenance and improving generalization of treatment gains across contexts. Therefore, the present review suggests basic and translational research on renewal provides an empirical literature to bring greater conceptual systematization to understanding generalization and maintenance of behavioral treatment.
Keywords: extinction, generalization, maintenance, relapse, renewal, translational research
Behavioral treatments arrange contingences designed to promote appropriate and desirable behavior and reduce or eliminate problematic behaviors. Initially successful treatment, however, does not ensure sustained success. Basic and translational research provides methods to understand the factors influencing and approaches for improving long-term maintenance of treatment outcomes. The present review examines basic and translational research concerned with how changes in environmental context influence long-term maintenance of behavioral treatments.
BEHAVIORAL TREATMENTS AND CONTEXT
Differential reinforcement of alternative behavior (DRA) is a class of behavioral treatments promoting the engagement in appropriate behavior and the elimination of problem behavior (see Petscher, Rey, & Bailey, 2009, for a review). Functional communication training (FCT) is one form of DRA that trains appropriate communication responses (i.e., mands) to replace problem behavior. Functional analyses identify the environmental events reinforcing and maintaining problem behavior (Iwata, Dorsey, Slifer, Bauman, & Richman, 1982/1994). With DRA/FCT treatments, behavior analysts identify appropriate responses to serve the same function as problem behavior. Specifically, they deliver the reinforcer maintaining the problem behavior for engaging in appropriate behavior. In one example of FCT for problem behavior maintained by attention, attention is provided contingent upon engaging in verbal requests and withheld contingent upon problem behavior. FCT is one of the most common methods used to treat problem behavior and is effective in eliminating problem behavior while FCT remains in effect (see Falcomata & Wacker, 2013; Greer, Fisher, Saini, Owen, & Jones, 2016; Rooker, Jessel, Kurtz, & Hagopian, 2013; Tiger, Hanley, & Bruzek, 2008 for reviews).
Appropriate behavior trained using FCT in one setting (e.g., clinic), however, is not guaranteed to generalize to other settings (e.g., home, school, store). As suggested by Fisher, Greer, Fuhrman, and Querim (2015), a longstanding common problem in applied behavior analysis is that treatment effects established during initial assessments and implementation often do not transfer to other contexts or therapists (e.g., Kazdin & Bootzin, 1972; Lang, Sigafoos, Lancioni, Didden, & Rispoli, 2010; Luczynski, Hanley, & Rodriguez, 2014; Rincover & Koegel, 1975; see Falcomata & Wacker, 2013; Osnes & Lieblein, 2003; Stokes & Baer, 1977; Stokes & Osnes, 1989, for reviews). In one example in which FCT did not successfully transfer across contexts, Schindler and Horner (2005) used FCT to decrease aggression and tantrums exhibited by three preschool children diagnosed with autism spectrum disorder (ASD). In combination with extinction of problem behavior, FCT successfully decreased problem behavior in the preschool room in which FCT occurred. However, transitioning to different settings (i.e., a different classroom and home) resulted in low levels of the functional communication response and increased levels of problem behavior (see also Durand & Carr, 1991; Wacker et al., 2005, for related findings).
Hagopian, Gonzáles, Rivet, Triggs, and Clark (2011) treated pica exhibited by two teenage individuals diagnosed with ASD and other developmental disabilities. Pica remained low when therapists blocked pica attempts and reinforced appropriate disposal of items (i.e., DRA) in a 3 × 3 m clinical treatment room. Despite treatment remaining in place, pica increased transiently for both participants when assessing pica in different settings (i.e., bathroom or classroom). Thus, behavioral treatments for problem behavior established in one setting, or context, did not generalize to other contexts. Given the importance of transferring treatment effects across contexts in applied behavior analysis and other helping professions, understanding the learning processes involved in the generalization of treatment would be an important step for approaching the development of strategies to promote generalization. This article reviews basic research on the role of contextual control of learning and its implications for generalization of behavioral treatments across contexts.
In this paper, contextual control refers to changes in behavior that occur when the same contingencies that come to control behavior in one context are implemented in a different context. The term context has been used in a variety of ways but generally refers to any environmental stimulus conditions that might come to influence behavior. Basic research on contextual control usually refers to global aspects of the environment (see Bouton, Winterbauer, & Todd, 2012; Podlesnik & Kelley, 2015, for discussions). With rats in a conditioning chamber, olfactory, visual, tactile, and location cues of the conditioning chambers have defined the context. With humans, room location, temperature, décor, odor, color of training and testing materials, illumination, etc. have defined the context (e.g., Collins & Brandon, 2002; Kelley, Liddon, Ribeiro, Greif, & Podlesnik, 2015; Vansteenwegen et al., 2006). Therefore, the environmental features tend to be more global than the specific discriminative stimuli arranged in studies of stimulus control with humans and nonhumans (e.g., Fisher, Kuhn, & Thompson, 1998; Fox, Smethells, & Reilly, 2013; Guttman & Kalish, 1956). In some circumstances, researchers may observe clear changes in responding following changes in context without isolating the specific stimulus characteristic(s) responsible for the change in responding (see Podlesnik & Kelley, 2015, for a related discussion). In these circumstances, contextual control is commensurate with a change in behavior linked to changes in settings often involved in the treatment of problem behavior and skill acquisition (e.g., from clinic to home or school).
Changing other more well defined features of the environment also come to control behavior in ways similar to global environmental contexts (Bouton et al., 2012; Podlesnik & Kelley, 2015). Therefore, we use the term contextual control in this paper to describe the full complement of antecedent and consequent events that may control the occurrence or nonoccurrence of behavior. For example, a target response may decrease due to changing (a) the global environmental setting (e.g., Bouton, Todd, Vurbic, & Winterbauer, 2011); (b) the discriminative stimulus, (e.g., Podlesnik & Miranda-Dukoski, 2015); (c) the establishing operation (e.g., food deprivation vs. satiety, Davidson, 1987, 1993); (d) the presence of punishment (e.g., Azrin & Holz, 1966; Estes, 1944), drug effects (e.g., Bouton, Kenney, & Rosengard, 1990); or (f) the presentation of alternative reinforcers (e.g., Trask & Bouton, 2016). Thus, the term contextual control, as used here, is broader than the term stimulus control—contextual control includes operant control by a broader set of antecedent and consequent events than stimulus control. Contextual control includes any environmental change co-occurring with a contingency change that comes to influence behavior.
EXTINCTION AND RENEWAL
Operant extinction eliminates reinforcers previously delivered contingent upon a target response. Behavior generally declines during extinction at a rate and pattern influenced by a range of variables, including those arranged during prior training and while extinction is in effect (see Bouton & Todd, 2014; Nevin & Wacker, 2013; Vurbic & Bouton, 2014 for reviews). Eliminating behavior with extinction, however, does not guarantee that the behavior will not reoccur. Extinguished behavior can wholly or partially return under a range of circumstances while extinction remains in effect (see Bouton et al., 2012; Pritchard, Hoerger, & Mace, 2014, for reviews).
Understanding extinction is important to applied behavior analysts. Despite extinction rarely being used as the sole component of behavioral treatments, extinction often is used in combination with other procedures to form the bedrock of many treatments designed to teach new skills and to eliminate problem behavior (Lerman & Iwata, 1996). As mentioned previously, clinicians often eliminate inappropriate behavior by determining the functional reinforcer(s) maintaining the problem behavior (Iwata et al., 1982/1994) and then withholding those reinforcers when problem behavior occurs. With DRA/FCT treatments, clinicians train appropriate responses to increase independence and/or communication skills (Petscher et al., 2009). Clinicians place less functional and socially acceptable responses on extinction when teaching progressively more functional and socially acceptable responses (e.g., shaping eye contact: Hall, Maynes, & Reiss, 2009; shaping vocalizations: Shillingsburg, Hollander, Yosick, Bowen, & Muskat, 2015). Despite strong support for such treatments in the relative short term, long-term maintenance of behavioral treatments remains a challenge (see Luczynski et al., 2014; Nevin & Wacker, 2013; Osnes & Lieblein, 2003). Instances in which previously eliminated undesirable behavior returns has been referred to as treatment relapse (e.g., Mace & Critchfield, 2010; Pritchard et al., 2014). Understanding the circumstances under which extinguished behavior returns, or relapses, can provide insight into strategies for maintaining gains achieved during behavioral treatments across a range of situations and in the face of challenges.
Despite the extensive use of extinction in behavioral treatments, little applied research is devoted to understanding the processes underlying extinction’s effects in the context of treatment (Lerman & Iwata, 1996). Fortunately, there is an extensive literature on extinction outside of applied behavior analysis upon which to draw (see Bouton et al., 2012; Lattal, St. Peter, & Escobar, 2013; Lerman & Iwata, 1996; Rescorla, 2001; Vurbic & Bouton, 2014, for reviews). Laboratory models can be used to assess fundamental learning processes during extinction and, more importantly for the present discussion, factors contributing to treatment relapse with the goal of improving long-term effectiveness (see Podlesnik & Kelley, 2015, for a discussion).
One effect of extinction particularly relevant to addressing long-term treatment efficacy is the influence of the current context on relapse of previously eliminated behavior—an effect often termed renewal. As noted previously, context refers to any environmental stimulus conditions that might come to influence behavior. Smith (2007) defined context generally as “that which surrounds” (p. 112) the learning situation confronting the organism. In basic laboratory studies, different contexts most often are defined by changes in aspects of the conditioning chamber, although a wide range of events has been shown to function as contexts (e.g., drug effects, time) and a wide range of relapse effects has been attributed to contextual changes (e.g., resurgence, reinstatement, spontaneous recovery; see Bouton et al., 2012; Podlesnik & Kelley, 2015, for discussions).
Renewal of extinguished behavior has been demonstrated most extensively in studies of respondent conditioning (see Bouton, 2002, 2004; McConnell & Miller, 2014, for reviews). For example, in a study by Bouton and Bolles (1979), rats trained to associate a tone with shock in one type of chamber (Context A) produced a conditional freezing response to the tone in that context, as indicated by decreases in rates of lever pressing maintained by food. However, rats no longer engaged in a conditional freezing response to the tone after extinguishing the tone in a different type of chamber (Context B). Upon returning to the original chamber (Context A) or transitioning to an entirely novel chamber (Context C), the tone conditional stimulus once again evoked conditional freezing in the absence of the shock. Training and extinguishing the tone in the same type of chamber (Context A) but transitioning to a novel chamber (Context C) also resulted in the tone conditional stimulus again evoking conditional freezing (Bouton & Ricker, 1994). With ABA, ABC, and AAC context arrangements, the return of responding in the third phase is a renewal effect (Bouton & King, 1983; Bouton & Peck, 1989).
The presence of ABC and AAC renewal in the above studies reveals that the effects of extinction do not fully generalize to other contexts. Despite the continued absence of shock, conditional freezing returned when transitioning out of the extinction context but not when remaining in the extinction context. Merely removing the context in which extinction occurred was sufficient to produce a return of conditioned responding. Thus, the current environmental stimulus context determines whether or not the conditional stimulus evokes the conditional response. The role of environmental context after conditioning and extinction can be understood using an analogy with verbal behavior (Bouton, 2002). The function of many words can be ambiguous without context. For example, shouting “fire” has no obvious function in the absence of context. Shouting “fire” has different effects, or meanings, depending on whether it is shouted in a crowed movie theater versus at a firing range. As with ambiguous words like fire and duck, the function of a conditional stimulus learned in one context and extinguished in another comes to depend on the current context. In the former context, the conditional stimulus is associated with unconditional stimuli but in the latter context it is not. Whether the response is performed depends on the prevailing context.
Although originally documented most extensively with respondent conditioning and in studies using nonhuman animals, renewal has recently been studied more intensively with operant behavior (Bouton, Todd, Vurbic, & Winterbauer, 2011), and its relevance has been linked to a range of human behavioral problems, including anxiety (Vervliet, Craske, & Hermans, 2013), drug abuse (Crombag, Bossert, Koya, & Shaham, 2008), and obesity and overeating (Boutelle & Bouton, 2015; Bouton, 2011). In addition, classic studies of memory (e.g., Godden & Baddeley, 1975; Goodwin, Powell, Bremer, Hoine, & Stern, 1969) reveal that lists of words learned in one context (e.g., underwater, intoxicated) are often better recalled when tested in the same context than in different contexts (e.g., on dry land, sober). These types of findings in which the environmental context mediates performance of learned behaviors are general, having been observed across a range of fields in experimental psychology (see Bouton, 1993, 2002, 2004, for reviews).
Recently, renewal has been extended to operant behavior in humans in a translational study with two children with developmental disabilities (Kelley et al., 2015). Color of the task materials, tee shirt worn by the experimenter, and poster board on the wall defined two different contexts. In the training context (A) during Phase 1, the experimenter delivered small edibles to participants on a fixed-ratio (FR) 1 of number/letter tracing for one participant (John) and matching pictures for the other participant (Drew). The investigators then arranged extinction in Phase 2 in a different context (B). Finally, Kelley et al. (2015) also arranged extinction in Phase 3 but in the original training context (A). Figure 1 shows that, despite extinction remaining in effect in Phase 3, response rates temporarily increased upon returning to the original training context—a renewal effect. Moreover, this study reported similar findings in a pigeon model arranging analogous contingency and contextual changes (see Podlesnik & Kelley, 2015, for a discussion). Thus, context influences the performance of operant behavior.
Figure 1.
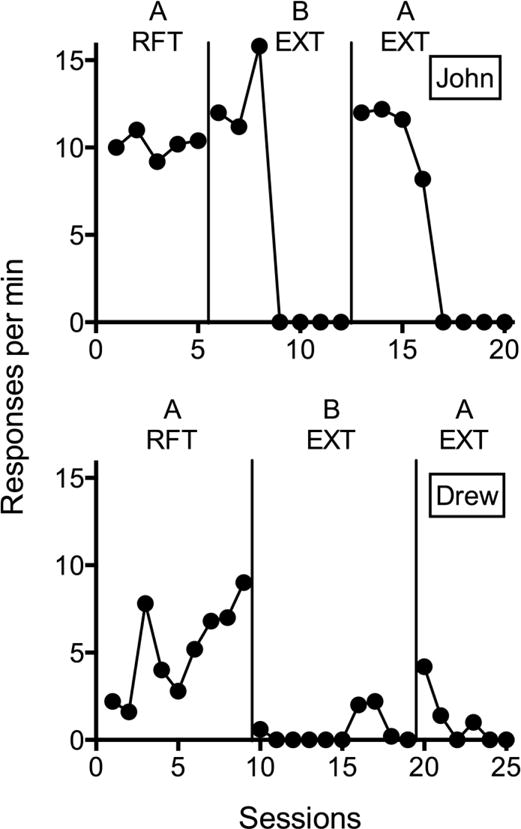
Response rates across sessions of Phases 1, 2, and 3 from Kelley et al. (2015). Phase 1 arranged reinforcement (RFT) during Context A. Phase 2 arranged extinction (EXT) during Context B. Phase 3 arranged extinction (EXT) during Context A. The top and bottom panels show responding across two children, John and Drew, respectively. (Copyright ©2015 by the Society for the Experimental Analysis of Behavior. Reproduced with permission.)
The implication for behavioral treatments from the translational research of Kelley et al. (2015) is that problem behavior learned in one context (e.g., home) and eliminated through treatment in a different context (e.g., clinic) could return upon returning to the original context or entering a novel context, even when treatment integrity remains intact. If the effects of extinction depend on the prevailing context, as observed with renewal, the effectiveness of a range of behavioral treatments that involve extinction might also depend on context. Therefore, we now will review basic and translational research on the contextual control of operant behavior and discuss the relevance of those findings to understanding and improving behavioral treatments. Readers interested in examining related theory and conceptual issues relevant to this research should explore the following references (Bouton & Todd, 2014; McConnell & Miller, 2014; Podlesnik & Kelley, 2015).
BASIC RESEARCH ON RENEWAL
Basic Findings
In an example of operant renewal from basic research, Bouton et al. (2011) trained rats to press levers according to a variable-interval (VI) 30-s of food reinforcement in Context A during Phase 1. Olfactory (lemon vs. pine), tactile (flat vs. staggered grid flooring), and visual (presence vs. absence of striped walls) stimuli comprised the different contexts across conditions, counterbalanced across rats. Figure 2 shows mean responses per min for three groups of rats. In Phase 2, lever pressing for the group labeled ABA was extinguished to near-zero rates in Context B. Testing for a single session in the extinction context (B) then had no effect. However, testing in Context A for a single session while extinction remained in effect produced a recovery to approximately 40% of the original rate of lever pressing. Therefore, similar to the findings of Kelley et al. (2015), simply returning to the original training context produced a substantial increase in extinguished responding—termed ABA renewal. These findings have profound implications for behavioral treatments conducted in clinical settings—merely returning to the original training context (A) was sufficient to instigate a return of the response while the extinction contingency remained in full effect. As previously stated, the implication is problem behavior learned in one context (A; e.g., home) but treated in a different context (B; e.g., clinic), might return when the client returns to the original context, even when perfectly maintaining treatment fidelity. Moreover, tripling the exposure to extinction did not reduce renewal in a related experiment by Bouton et al. (2011), suggesting problem behavior could be susceptible to renewal effects even following extensive treatment (but see Denniston, Chang, & Miller, 2003; Laborda, Polack, Miguez, & Miller, 2014; Thomas, Vurbic, & Novak, 2009).
Figure 2.
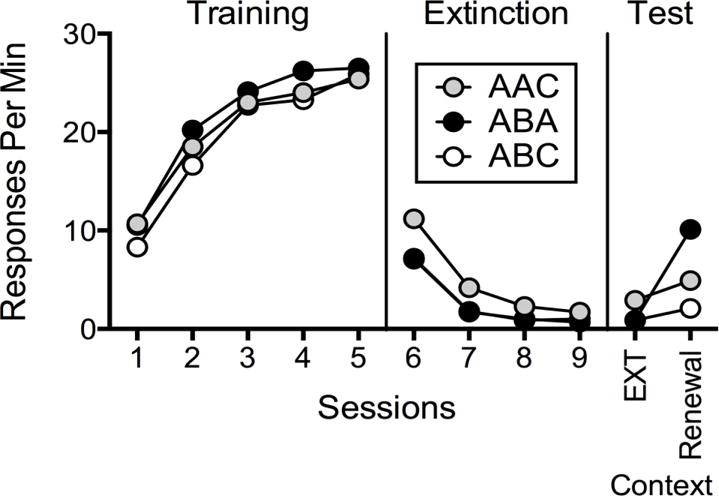
Mean response rates across Phases 1, 2, and 3 in tests of renewal of extinguished operant responding in rats from Bouton et al. (2011). Sixteen rats served in each group. After operant training in Context A, the response was extinguished in either Context A (Group AAC) or Context B (Groups ABA and ABC). Testing then occurred both in the extinction context and in the original conditioning context (Context A; Group ABA) or in a new context (Context C; Groups ABC and AAC). The data from Groups ABA and AAC were collected in the same experiment and are directly comparable. Data from Group ABC were collected in a separate experiment, so comparisons of data from Groups ABA and AAC with Group ABC should be made with caution. This figure was replotted from data appearing in Figures 1 and 3 in the original paper.
Further, Bouton et al. (2011) showed that renewal does not require returning to the original training context after extinguishing the response in another one (i.e., ABA renewal). Transitioning from the training context (A) to a novel context (B) in Phase 2 produced a decrement in operant responding (see Figure 2, ABA and ABC groups) compared with remaining in the training context (A) during extinction (see AAC group; see also Bouton, Todd, & León, 2014; see Bouton, 2014, for a review). During testing, the ABC group continued to show little responding in the extinction context (B), but transitioning to a third context (C) produced renewal—termed ABC renewal. Finally, in the AAC group, lever pressing was trained in Context A but also extinguished in Context A. Transitioning to a novel context also produced a recovery in responding—termed AAC renewal (also known as AAB renewal; Bouton et al., 2011); remaining in the extinction context (A) did not produce renewal.
The results depicted in Figure 2 suggest that ABA renewal is larger than ABC or AAC. The pattern is consistent with other findings for both respondent (e.g., Bouton & Bolles, 1979; Rescorla, 2008; Thomas, Larson, & Ayres, 2003, but see Polack, Laborda, & Miller, 2013) and operant conditioning (e.g., Berry, Sweeney, & Odum, 2014; Nakajima, Urushihara, & Masaki, 2002; Todd, Winterbauer, & Bouton, 2012; but see Todd, 2013, for evidence that AAC renewal can be equivalent to ABA under some conditions). Yet, 16 out of 16 rats showed AAC renewal (more responding in C than A during testing), 15 out of 16 rats showed ABC renewal (more responding in C than B), and 16 out of 16 rats showed ABA renewal. Moreover, ABC and AAC renewal are important because they indicate that removal from the extinction context can be sufficient to cause a return of the extinguished response. These findings with ABC and AAC renewal suggest that merely transitioning out of the extinction context is sufficient to produce renewal (see Bouton et al., 2012, for a discussion). This tendency for renewal to occur when transitioning from the extinction context to any other context may be one of the reasons why treatment relapse occurs.
The presence of renewal when transitioning to novel contexts in which extinction was never experienced (i.e., ABC and AAC renewal) further suggests problem behavior could be susceptible to renewal effects following behavioral treatments in clinical settings. With ABC renewal, problem behavior learned in one context (A; e.g., home) but treated in a different context (B; e.g., clinic), will return when transitioned to a novel context (C; e.g., school, new clinic). With AAC renewal, even problem behaviors learned and treated in the same context (A; e.g., home) will return when transitioned to a novel context (C; e.g., school, novel clinic). It should be emphasized that these tests of renewal effects reported here occur with extinction in effect, thereby simulating perfect treatment integrity. Perfect treatment integrity is ideal but often difficult to maintain (Betz, Fisher, Roane, Mintz, & Owen, 2013; Fisher et al., 2015; Hanley, Iwata, & Thompson, 2001). The unfortunate implication for treatment is that caregivers might inadvertently or unintentionally reinforce instances of problem behavior, termed commission errors (St. Peter Pipkin, Vollmer, & Sloman, 2010). Thus, any increase in problem behavior with a context change increases the possibility of commission errors and reacquisition of problem behavior. Therefore, understanding factors contributing to and reducing renewal could improve the chances of maintaining long-term treatment efficacy.
Training Reinforcement Conditions
Reinforcement conditions in the training context (A) impact renewal. A longer reinforcement history and greater reinforcer rates in Context A during Phase 1 enhance both ABA and ABC renewal (Berry et al., 2014; Podlesnik & Shahan, 2009, 2010; Todd et al., 2012). Todd et al. (2012) arranged either 4 or 12 training sessions in Context A during Phase 1. Following extinction in Context B, both ABA and ABC renewal consistently were greater in groups of rats receiving 12 training sessions in Phase 1. Thus, problem behavior having a more extensive learning history is more susceptible to renewal, as might be the case with older individuals having extensive histories of engaging in problem behavior.
Relatedly, Podlesnik and Shahan (2009, 2010) and Berry et al. (2014) assessed whether greater reinforcement rates also produce greater ABA and ABC renewal in pigeons. Podlesnik and Shahan (2009) arranged equal variable-interval (VI) 120-s schedules between two components of a multiple schedule. Additional food reinforcers were presented according to a variable-time (VT) 20-s schedule in one component to increase the overall reinforcement rate in that component. Two distinct colors alternated on the keys every minute to signal the different reinforcement rates. A stable house light served to signal the training context (A) in both components. During Phase 2, all reinforcers were discontinued and the house light flashed every 0.1 s to comprise Context B. Finally, the house light returned to being stable to create a return to Context A while extinction remained in effect. Responding showed greater resistance to extinction in Phase 2 and larger ABA renewal effects upon returning to Context A in Phase 3 in the component in which experimenters arranged a higher rate of reinforcement in Phase 1. Therefore, these findings revealed greater training reinforcement rates increase ABA renewal (Berry et al., 2014; Thrailkill & Shahan, 2012).
Berry et al. (2014) arranged a similar test for ABC renewal with food reinforcers delivered according to a VI 30-s schedule in the richer component and a VI 120-s schedule in the leaner component in Phase 1. A white house light and particular operant chamber comprised Context A. Responding was extinguished in Phase 2 while changing the house light color and operant chamber as Context B. Finally, they again changed the house light color and operant chamber while extinction remained in effect in Context C. As with ABA renewal in Podlesnik and Shahan (2009), Berry et al. also observed greater resistance to extinction and renewal with greater training reinforcement rates arranged in Phase 1. Therefore, more frequent reinforcement of problem behavior could generally produce more persistent problem behavior, in terms of making behavior both more difficult to eliminate and more likely to return upon changing context (see Nevin et al., in press; Nevin & Wacker, 2013; Podlesnik & DeLeon, 2015, for discussions).
Training and Extinction Response Rates
Previous literature examining other operant relapse phenomena, including resurgence (da Silva, Maxwell, & Lattal, 2008; Sweeney & Shahan, 2013; Winterbauer, Lucke, & Bouton, 2013) and reinstatement (Doughty, Reed, & Lattal, 2004), suggests that preextinction response rate influences the degree to which responding will relapse. We know of no published literature examining these effects directly with renewal of extinguished operant responding. Nevertheless, we examined whether training response rates influenced ABA renewal from two previously published studies (i.e., Bouton et al., 2011; Podlesnik & Shahan, 2009). We selected these studies because of the differences in species (rats vs. pigeons) and differences in procedures (simple schedule vs. multiple schedule). The specific methods have been described in greater detail in the previous section.
For Bouton et al. (2011), we compared average response rates during the last session of Phase 1 with response rates during the single session of the renewal test for the 16 rats in the ABA group. A Pearson correlation yielded a significant positive relation between training response rates and renewal (r [14] =.63, p = .009).
Podlesnik and Shahan (2009) examined ABA renewal in 10 pigeons across two components of a multiple schedule. We examined response rates in training as the mean of the last six sessions in Phase 1 and the mean of all four sessions from Phase 3. Nine of 10 pigeons showed lower training response rates in the component with added response-independent food, and the 10th pigeon showed equivalent response rates. Training response rates correlated positively with renewal in both components. Correlations reached statistical significance in the component with the added food (r [8] = .93, p < .0001) but not in the component without added food (r [8] = .57, p = .883). The findings from Bouton et al. (2011) and Podlesnik and Shahan (2009) show that greater rates of behavior prior to treatment tend to result in greater renewal during posttreatment shifts in context. These findings suggest relapse could be more likely with problem behavior occurring more frequently prior to treatment.
Despite an apparently general relation between training response rates and renewal in these studies, the findings of Podlesnik and Shahan (2009) suggest an interaction between training response rates and training reinforcement rates. As previously mentioned, 9 of 10 pigeons showed lower training response rates in the component with added food. However, we observed the opposite relation during the renewal test—higher response rates in the component associated with added food (see also Saini & Fisher, 2016). Therefore, additional sources of reinforcement that might initially decrease stable training response rates (e.g., NCR, DRA) could produce a response that is more susceptible to relapse with changes in context (see Nevin & Wacker, 2013; Podlesnik & DeLeon, 2015, for discussions).
Finally, we also compared response rates during tests for renewal in the presence of the extinction context (B) and training context (A) from Bouton et al. (2011). A Pearson correlation yielded a significant relation between extinction response rates and renewal (r [14] =.54, p = .032). The positive relation between level of responding in the extinction context and renewal suggests responding that is more persistent in treatment also will be more likely to relapse. This finding is consistent with those described in the previous section on training reinforcement rates by also revealing that responses that are more resistant to initial treatment will be more likely to relapse (see Nevin & Wacker, 2013; Podlesnik & DeLeon, 2015, for discussions).
Context Similarity
Treatment relapse might be more likely as the treatment setting becomes more different from contexts contacted outside of the treatment setting. Consistent with this idea, the similarity of the extinction context in Phase 2 and the renewal-test context in Phase 3 influences the size of renewal effects. Todd et al. (2012) trained all groups of rats in Context A and extinguished responding in Context B. Finally, they tested for ABA renewal in Context A, as well as ABC renewal in Context C. Importantly, they varied the degree to which the extinction context (B) differed from the renewal-test contexts (A or C) across groups of rats. Results showed greater renewal when Context B was more distinct from Contexts A and C than when Context B was less distinct than Contexts A and C (see Podlesnik & Miranda-Dukoski, 2015, for related findings). In other words, renewal was greater when extinction occurred in the presence of contexts more different than the original learning context (A) and renewal-test context (i.e., A or C).
In terms of relevance to treatment relapse, renewal of problem behavior could be more likely when treatment occurs in settings that are dramatically different (e.g., clinic) from those settings to which individuals are transitioned following treatment (e.g., home, school, store). Generally consistent with literature on stimulus control (Honig & Urcuioli, 1981; Lazareva, 2012), treatment effects generalize to new contexts to the extent to which those new contexts resemble prior training and treatment contexts. Thus, at least initially and when possible, efforts might be made to make clinical settings similar to posttreatment settings, posttreatment settings similar to clinical settings, and posttreatment settings different from the initial contexts in which problem behavior was learned. In fact, treatment ideally would occur in the same context in which problem behavior was established to avoid renewal effects upon changing contexts.
Some readers might find this suggestion at odds with the finding shown in Figure 2 of more rapid decreases in target responding in Phase 2 of the ABA and ABC groups than for the AAC group. Changing contexts in Phase 2 might produce faster decreases initially but at the expense of potentially greater renewal, depending on the posttreatment setting. Recall Todd et al. (2012) observed greater ABA and ABC renewal when treatment contexts (B) differed from the posttreatment context (A or C). Therefore, avoiding the use of new and distinct treatment settings entirely might eliminate the possibility of renewal in the natural setting by establishing treatment effects directly in the context where the problem behavior occurs.
Of course, there will be cases in which implementing treatment in the context in which severe problem behavior was established could pose practical and safety concerns (e.g., availability of potentially dangerous objects). In general, insights into treatment based on the literature on renewal should be weighed against practical issues and clinical goals of each case. In addition, there will be many clinical cases in which remaining in the natural setting will be impractical or impossible and, therefore, other strategies will need to be developed to combat renewal effects. Operant renewal can nonetheless be modified by a number of other environmental variables, including similarity of contextual conditions across phases (e.g., Podlesnik & Miranda-Dukoski, 2015; Todd et al., 2012) and training response and reinforcement history (Bouton et al., 2011; Podlesnik & Shahan, 2009, 2010; Todd et al., 2012). If environmental factors modify renewal effects, procedures could be developed to model potential interventions to mitigate renewal under clinical situations. To this end, we discuss below other approaches and techniques to mitigating renewal.
Before we do, we recognize that much of the research on renewal primarily reflects respondent or operant conditioning with nonhuman animals. We thus recognize that the extent to which one can generalize effects from studies of renewal to clinical applications requires additional research. The nascent status of the translational and clinical research on renewal, however, offers a ripe and important area for translational and clinical researchers. It is worth noting, though, that despite the absence of direct demonstrations of renewal in clinical cases published using applied behavior analysis to date, the findings from the basic literature show renewal to be robust and general across species and experimental preparations. And a number of findings established with the renewal of respondent conditioning from animal laboratories generalize to studies using humans (e.g., Vervliet et al., 2013). We now turn to techniques demonstrated to reduce renewal in basic and translational research with human and nonhuman animals.
TECHNIQUES TO MITIGATE RENEWAL
Training a novel response with reinforcement produces new learning. Similarly, the eventual absence of responding following extinction also reflects learning (e.g., Bouton, 2002, 2004, 2014). We argued above that whether one pattern of behavior or the other occurs depends in part on the prevailing context. This process is analogous to how the meaning of a word changes between different contexts (e.g., fire in theater vs. shooting range). Relatedly, clinicians can understand failures to perform newly trained skills or the return of problem behavior when testing in novel contexts as reflecting the relation between context and a particular learning history with that response (see Todd, Vurbic, & Bouton, 2014). Further, renewal also occurs following techniques to strengthen treatments in Phase 2. For example, renewal occurs following omission training in Phase 2, or differential reinforcement of other behavior schedules (DRO; Nakajima et al., 2002). Renewal also occurs if punishment is used instead of extinction (Bouton & Schepers, 2015). From this perspective, generalization failures do not reflect a failure to have learned the more recently trained responses. Instead, generalization failures reflect an influence of context on the learning that occurred during treatment. Additional learning experiences might be necessary for the appropriate response to be performed in the new context.
Stokes and Baer (1977) recommended strategies for increasing generalization of treatment gains beyond the conditions in which interventions were established. Maintaining the extinction contingency when context changes across Phases 2 and 3 in most basic studies of renewal resembles what Stokes and Baer called the train-and-hope approach to having treatment gains extend to other settings, or contexts. Specifically, generalization of treatment gains (i.e., extinction effects) is not explicitly programmed across contexts. With ABA and ABC renewal, Phase 1 within Context A models reinforcement of ongoing behavior under natural conditions, whereas Phase 2 within Context B models an extinction intervention at a clinic. Upon returning in Phase 3 to the original context (A) or transitioning to a novel context (C), no further contingency or common stimuli are provided to enhance the effects of extinction following the context change. In fact, ABA- and ABC-renewal procedures implemented in basic research may reflect best-case examples of the train-and-hope approach. This is because renewal procedures maintain the extinction contingency during context changes in Phase 3, modeling perfect maintenance of treatment integrity. However, when the train and hope is used in typical human environments (e.g., train in the clinic, transfer treatment to the home) the level of treatment integrity may often be imperfect (e.g., problem behavior may occasionally produce reinforcement when extinction is recommended). Thus, failure to generalize treatment gains under actual implementation of treatments using train and hope could be significant.
Sometimes this train-and-hope approach is effective in maintaining low levels of the target behavior both in research and practice. For instance, there have been failures to observe operant renewal effects with ABC and AAC renewal in the basic literature (Crombag & Shaham, 2002; Nakajima, Tanaka, Urushihara, & Imada, 2000; Podlesnik & Kelley, 2014; Zironi, Burattini, Aicardi, & Janak, 2006). There is also evidence of transfer/generalization of treatment gains across settings when treatment occurred in a different context from an assessment of generalization of those treatment effects (e.g., Durand & Carr, 1991).
Nevertheless, there is also substantial evidence indicating that treatment effects often do not generalize across contexts or settings. For example, the role of context mitigating treatment gains is revealed both by the existence of robust renewal effects in the basic and translational literature (Bouton et al., 2012; Kelley et al., 2015) and instances of treatment relapse when changing settings in application (e.g., Durand & Carr, 1991; Lalli, Casey, & Kates, 1997; Luczynski et al., 2014; Schindler & Horner, 2005). Generalization failures across settings when using multiple-baseline designs also reveal how context can negatively impact treatment generality. Changing context from the treatment setting can result in the absence of generalization of treatment effects both in terms of problem behavior (e.g., Roane, Kelly, & Fisher, 2003) and skill acquisition (e.g., Cushing & Kennedy, 1997; Dunlap, Kern-Dunlap, Clarke, & Robbins, 1991; Griffiths & Craighead, 1972). Moreover, as Stokes and Baer (1977) suggested, evidence of generalization in the applied literature could reflect a bias toward reporting successes rather than failures (i.e., “the file-drawer problem”; Rosenthal, 1979). Given the prevalence of treatment relapse in a wide range of disciplines concerned with treating behavioral problems (e.g., anxiety, drug abuse, overeating), developing methods to mitigate renewal could be broadly beneficial (see Bouton, 2014).
Next, we turn to techniques employed in the basic literature to mitigate renewal when arranging extinction in one context and testing in another. We will link these techniques with specific recommendations made by Stokes and Baer (1977) to promote generalization across settings and contexts (see also Falcomata & Wacker, 2013; Osnes & Lieblein, 2003; Stokes & Osnes, 1989). Although we focus on how context influences effectiveness of treatments for problem behavior, the literature on generalization and maintenance (e.g., Dunlap, 1993; Stokes & Baer, 1977) suggests context likely influences maintenance of skills and treatment of problem behavior in similar ways.
Multiple-Context Training
The implication of ABA, ABC, and AAC renewal is that learning new appropriate behaviors during treatment is not general or absolute but specific to the particular context in which treatment occurred. For example, a communication response replacing aggressive behavior maintained by attention might be trained in a clinical setting (Context B). However, the communication response may not generalize to other settings (e.g., home, school). Instead, clients might emit the communication response only in the clinical setting, Context B (see Bouton, 1988, for a related discussion). When this occurs, generalization beyond Context B must be directly trained.
Following this general logic, Gunther, Denniston, and Miller (1998) hypothesized that treatment arranged across multiple contexts would promote generalization of treatment effects and reduce renewal when tested in novel contexts. In a laboratory model assessing respondent fear conditioning in rats, Gunther et al. paired white noise with shock in a distinct context in Phase 1. In Phase 2, they extinguished fear by presenting white noise in the absence of shock either in one different context or across three different contexts. Testing for renewal of the fear response in a novel context during Phase 3 revealed lower renewal in the group in which white noise was extinguished across multiple contexts (see also Bandrian Balooch, Neumann, & Boschen, 2012). In addition to transitioning to a novel context (C), similar findings have been observed when returning to the original training context (A, e.g., Bandrian Balooch & Neumann, 2011; Chaudhri, Sahuque, & Janak, 2009). These findings imply that training new skills to replace problem behavior across multiple contexts could enhance generalization of those skills outside of those training contexts.
Thus far, the effect of establishing generalization across multiple contexts during Phase 2 has only been assessed in respondent-conditioning preparations (see also Glautier, Elgueta, & Nelson, 2013; Shiban, Pauli, & Mühlberger, 2013; Thomas, Vurbic, & Novak, 2009). Nevertheless, Todd et al. (2012) revealed similar effects in rats when training an operant response in either one or across two contexts in Phase 1. When changing context during extinction in Phase 2, responding extinguished more slowly in the rats trained in two contexts, indicating greater generalization to the new context in extinction. Similarly, another contextual change in Phase 3 also produced greater renewal in the rats trained in two contexts. Thus, operant responses trained across multiple contexts are more likely to persist and return when transitioning to novel contexts. These effects should also apply to problem behavior and appropriate behavior learned across multiple contexts. Additional translational and clinical research assessing multiple-context training would be useful for assessing its efficacy in clinical situations.
These renewal studies assessing performance trained or extinguished across multiple contexts reveal that exposure to multiple contexts increases generalization. These findings align with the suggestions of Stokes and Baer (1977) in that responses trained in one setting or context that do not generalize to other contexts should be taught the “same generalization lesson, and then another, and then another, and so on until the induction is formed” (p. 355). Stokes and Baer provided examples in which training appropriate conversational speech generalized to novel therapists only after being reinforced across multiple therapists (Garcia, 1974) and to novel settings after being reinforced across multiple settings (Allen, 1973). Stokes and Baer called this form of generalization training training sufficient exemplars and suggested this is probably one of the most important strategies for promoting generalization of performance in novel situations. This approach forms a more general strategy Stokes and Osnes (1989) called training diversely.
Similarly, Luczynski et al. (2014) trained a range of functional-communication responses (e.g., asking for items from peers or help from teachers) to six students in a preschool classroom. Later, they assessed generalization to novel classrooms and teachers but found decreases in functional-communication responses and increases in problem behavior in all six children. Performance in the new classroom achieved training levels only after additional instruction in the new classroom. Thus, findings both from basic studies of renewal and clinical research reveal the importance of training appropriate responses across a range of settings if those responses are to generalize to novel settings. Generalization to novel settings is of upmost importance because the training setting (e.g., clinic) is probably not the most relevant place to perform newly learned appropriate responses in the long term (e.g., home, school, community).
Extinction Cue
The perspective that a context mediates learning implies that the prevailing context during original learning evokes responding, and the context prevailing during extinction or treatment evokes little or no responding (see Bouton & Todd, 2014; Bouton et al., 2012, for reviews). From this perspective, operations signaling the extinction/treatment context should produce little or no responding. The question is whether such an operation signaling the extinction/treatment context can promote the maintenance of treatment effects outside the extinction/treatment context.
In a respondent conditioning preparation, Brooks and Bouton (1994) arranged for a tone to signal food presentation in Context A during Phase 1. In Phase 2, the tone was extinguished through presentations in the absence of food in Context B. In addition, an extinction cue in which the house light turned off between tone presentations further signaled the extinction contingency in Context B—note the extinction cue was temporally separated from the tone. Upon returning to Context A in Phase 3, the extinction cue eliminated renewal (see also Brooks & Bouton, 1993, for related findings). On the other hand, renewal was robust in a group returning to Context A without the extinction cue. Similar effects of extinction cues presented in training contexts have been demonstrated to decrease operant alcohol self-administration in rats (Willcocks & McNally, 2014) and, in humans, both reactions to alcohol cues (Collins & Brandon, 2002) and fear conditioning (Barlow, O’Brien, & Last, 1984; Dibbets, Havermans, & Arntz, 2008; Dibbets & Maes, 2011; Nelson, Sanjuan, Vadillo-Ruiz, Pérez, & León, 2011; Vansteenwegen et al., 2006). Finally, presenting a new and qualitatively different reinforcer independent of responding in Phases 2 and 3 reduces renewal when returning to the original training context in Phase 3 (Trask & Bouton, 2016). These findings imply that stimuli present during behavioral treatments in clinical settings could be used to bridge treatment effects to nontreatment settings (e.g., home, school, community). Additional translational and clinical research examining the types of cues improving treatment efficacy would be useful. For example, instructions could be useful to reduce renewal of some types of problem behavior (see Mystkowski, Craske, Echiverri, & Labus, 2006, for relevant findings).
That an extinction cue present during the extinction context can reduce renewal is consistent with what Stokes and Baer (1977) called programming common stimuli. Later, Stokes and Osnes (1989) categorized this strategy within a more general approach of incorporating common mediators. The strategy of training an extinction cue to mitigate renewal in basic studies (e.g., Brooks & Bouton, 1994; Willcocks & McNally, 2014) could be used to enhance generalization of treatment effects. The strategy is to (1) establish cues learned as part of the treatment context mediating appropriate behavior and (2) employ those cues during contexts in which appropriate behavior is unlikely. If the cues effectively signal the extinction context, they should reduce renewal when transitioning to the original training context (A) or to novel contexts (C). Thus, in clinical situations, stimuli associated with the treatment context could increase appropriate behavior in the presence of the nontreatment context relative to the absence of such common stimuli.
In a relevant clinical example, Fisher et al. (2015) arranged FCT for three individuals with developmental disabilities to reduce problem behavior (destructive behavior, aggression, elopement) in a clinical setting. They employed a multiple schedule in which they reinforced a functional communication response in the presence of the discriminative stimulus (SD) and did not reinforce the response in the presence of a different stimulus (SΔ). Therapists wore different colored wristbands to signal SD and SΔ. Once high rates of functional communication responses occurred in the presence of SD and near-zero rates occurred in the presence of SΔ, the effectiveness of SD and SΔ maintained across different contexts (i.e., rooms) and therapists for all participants. Therefore, the wristbands generally maintained the effectiveness of FCT and reductions in problem behavior in novel situations, perhaps analogous to mitigating ABC renewal effects with extinction cues (see Brooks & Bouton, 1994). These findings suggest extinction cues can be implemented as an effective strategy for mitigating renewal outside treatment contexts. In other words, extinction cues aid in increasing generalization of treatment effects (see also Fuhrman, Fisher, & Greer, 2016).
The findings of Fisher et al. (2015) indicate the promise for arranging arbitrary stimuli to increase generalization of new skills across settings and reducing renewal of problem behavior when transitioning among contexts. Other studies have shown similar effects when the common stimulus is another person (e.g., Barlow et al., 1984; Stokes, Doud, Rowbury, & Baer, 1978), a discriminative stimulus signaling alternative behavior (Mace et al., 2010; Podlesnik & Bai, 2015; Podlesnik, Bai, & Elliffe, 2012; Podlesnik, Bai, & Skinner, 2016), or the reinforcer used during FCT (Charlop, Schreibman, & Thibodeau, 1985). Similarly, Trask and Bouton (2016) found that reinforcers can play a discriminative role and reduce renewal of extinguished operant behavior in rats. In Phase 1, one type of food reinforcer, or outcome (O1), was delivered in Context A. In Phase 2, responding no longer produced O1 and a different reinforcer (O2) was delivered on a time-based schedule in Context B. Maintaining the response-independent O2 reinforcers when returning to Context A in Phase 3 attenuated ABA renewal compared to eliminating O2 reinforcers in Phase 3. Thus, reinforcers presented during the extinction context (B) could reduce renewal when returning to the original training context (A). These findings are consistent with the findings of Fisher et al. (2015) and the suggestion that a range of stimuli arranged during treatment could be used to facilitate generalization of appropriate behavior and reduce relapse of problem behavior.
Combination of Multiple-Context Training and Extinction Cues
An important implication of the findings of Trask and Bouton (2016) is that changes in stimulus context can combine with aspects of behavioral interventions (see also Kearns & Weiss, 2007; Kincaid, Lattal, & Spence, 2015; King & Hayes, 2016; Nakajima et al., 2002; Podlesnik & Kelley, 2014, 2015). Specifically, Trask and Bouton also assessed the presence and absence of an alternative reinforcer (O2) during Phase 3 across groups of rats while they remained in the extinction context (B) and upon returning to the original training context (A). Renewal was greatest when simultaneously removing the alternative reinforcer (O2) and returning to Context A. Renewal was lower but similar between the group remaining in Context B but removing O2 and the group transitioning back to Context A but maintaining O2. No renewal occurred when subjects remained in Context B in the context of O2. Thus, multiple stimuli can contribute to the lack of transfer of treatment effects, or relapse. Relevant to the findings that a range of events can contribute to relapse, Falcomata and Wacker (2013) recommended assessing the effect of combinations of approaches to promote the generalization or transfer of treatment effects.
In one relevant example in which multiple treatment strategies were employed in a similar manner as described here, Piazza, Hanley, and Fisher (1996) integrated both training across multiple contexts and programming common stimuli, resulting in perfect transfer of treatment effects (see McKenzie, Smith, Simmons, & Soderlund, 2008, for related findings). Piazza et al. treated a teenage male with developmental disabilities who had engaged in severe cigarette-butt pica at home and around the community for approximately four years—the parents were chronic smokers. The patient was admitted to an inpatient clinic where assessments verified that nicotine maintained the pica. Treatment in the clinical setting occurred in a 3 m × 3 m room with a chair, table, puzzles, and workbook. Therapists planted cigarette butts within the room and taped a piece of purple construction paper to the wall to serve as a discriminative stimulus that signaled implementation of the treatment. Treatment consisted of providing noncontingent access to food and reprimands contingent on reaching for cigarette butts (“no butts”). Following a reduction in pica in the treatment setting, therapists introduced the treatment in the presence of the purple card across multiple contexts (e.g., living area, office). Then, therapists verified the effectiveness of the purple card relative to a novel yellow card in a similar 3 m × 3 m treatment room while presenting noncontingent food without the reprimand contingency. Pica remained eliminated in the presence of the purple card but not the yellow card.
After establishing the effectiveness of the purple card in the treatment room, therapists assessed its effectiveness in previously trained settings (i.e., living area, office), novel settings (i.e., walks near hospital and in mall), and the client’s home. These tests across multiple contexts are important because punishment effects can be influenced by contextual changes and, thereby, susceptible to renewal effects (Bouton & Schepers, 2015). Nevertheless, zero instances of pica occurred across all settings. Thus, the treatment package of training in multiple contexts and arranging a signal associated with treatment (i.e., purple card) effectively eliminated problem behavior in the original training situation (i.e., Context A) and prevented renewal in novel settings (i.e., Context C). These methods and findings are analogous to those described above demonstrating the effectiveness of multiple-context training (e.g., Gunther et al., 1998) and arranging extinction cues (e.g., Brooks & Bouton, 1994) in mitigating renewal. Although the findings of Piazza et al. (1996) were limited to a single participant and do not reveal the relative contribution of multiple-context training versus the extinction cue to treatment effectiveness, they reveal methods consistent with those prescribed in this review and by Falcomata and Wacker (2013).
CONCLUSION
Although relapse, or failure to generalize treatment gains, can be influenced by a range of factors, including lapses in treatment integrity (St. Peter Pipkin et al., 2010), changes in motivational variables (Murphy, McSweeney, Smith, & McComas, 2003), and changes in the function of behavior (Lerman, Iwata, Smith, Zarcone, & Vollmer, 1994), the focus of the present review addressed the role of environmental context. Context is important because all learning necessarily occurs within some context or subset of contexts. The present review focused on the research most relevant to understanding situations in which context influences operant behavior and its relevance to the generalization of behavioral treatments. We have provided evidence that context can come to mediate learning in the sense that learning can be specific to contexts. DRA/FCT treatments arranging extinction of problem behavior and reinforcement of appropriate behavior in one setting do not necessarily generalize to novel settings. These findings have been demonstrated at least since the findings motivating Stokes and Baer (1977) to write their article on treatment generalization. A primary goal of the present review is to take a step toward linking some of Stokes and Baer’s suggestions with findings from the research literature on renewal (see Bouton, 1993; Bouton et al., 2012; Bouton & Todd, 2014; Podlesnik & Kelley, 2015, for reviews).
As discussed previously (see Kelley et al., 2015; Podlesnik & Kelley, 2015), we hope contextual control of behavior comes to be anticipated by clinicians in a similar way as the transient increases in behavior at the onset of extinction, or extinction bursts (Lerman & Iwata, 1995). The possibility of observing extinction bursts with the initiation of extinction is expected. Thus, therapists can prepare for and address such increases in severity of problem behavior accordingly without the concern that the intervention is wholly ineffective or exacerbating the problem in the long term. Similarly, when transitioning among settings, behavior analysts can expect contextual effects and take appropriate measures to maximize generalization of treatment gains and minimize renewal of problem behavior. Unlike extinction bursts, for which we know relatively little about the controlling variables, the literature on renewal can serve as a guide to maximizing treatment effects across environmental contexts. Linking the literature on treatment generalization and renewal furthers the principle of applied behavior analysis that treatments be conceptually systematic (Baer, Wolf, & Risley, 1968). Treatments integrated in theoretical and conceptual frameworks derived from basic research provide principled approaches for intervention and the possibility for developing novel and unexpected approaches to implementing behavioral treatments (e.g., Mace et al., 2010).
The present review takes a general approach that one effective way of improving the generalization of treatment gains is through designing behavioral treatments with an understanding of the processes underlying those treatments (see also DeLeon, 2011; Hake, 1982; Lerman, 2003; Mace, 1994; Mace & Critchfield, 2010; Vollmer, 2011). With an understanding of how behavioral treatments impact learning and performance, researchers and clinicians can appreciate the potential range of interventions available to them. Otherwise, interventions could take the form of an “out-of-the-box” technology with a few moving parts to manipulate. Instead, it is important to understand that interventions impinge upon biological organisms whose learning processes are varied and interrelated through interactions between learning and evolutionary history (see Baum, 2012; Bouton, 2016, for relevant discussions). As a result of this view, translational research bringing together the expertise of basic researchers with applied researchers and clinicians can maximize the relevance of the basic research while providing a depth of understanding to clinically relevant issues.
Acknowledgments
The authors thank Sandhya Rajagopal for comments on a previous version of this manuscript. The contribution of MEB was supported by NIH Grant R01 DA033123.
Footnotes
Action Editor, Wayne Fisher
Contributor Information
Christopher A. Podlesnik, Florida Institute of Technology and The Scott Center for Autism Treatment, The University of Auckland
Michael E. Kelley, Florida Institute of Technology and The Scott Center for Autism Treatment
Corina Jimenez-Gomez, Florida Institute of Technology and The Scott Center for Autism Treatment.
Mark E. Bouton, University of Vermont
References
- Allen GJ. Case study: Implementation of behavior modification techniques in summer camp settings. Behavior Therapy. 1973;4:570–575. http://dx.doi.org/10.1016/S0005-7894(73)80009-7. [Google Scholar]
- Azrin NH, Holz WC. Punishment. In: Honig WK, editor. Operant behavior: Areas of research and application. New York, NY: Appleton-Century-Crofts; 1966. pp. 380–447. [Google Scholar]
- Baer DM, Wolf MM, Risley TR. Some current dimensions of applied behavior analysis. Journal of Applied Behavior Analysis. 1968;1:297–306. doi: 10.1901/jaba.1968.1-91. http://dx.doi.org/10.1901/jaba.1968.1-91. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bandrian Balooch SB, Neumann DL. Effects of multiple contexts and context similarity on the renewal of extinguished conditioned behaviour in an ABA design with humans. Learning and Motivation. 2011;42:53–63. doi: 10.1016/j.lmot.2010.08.008. [DOI] [Google Scholar]
- Bandrian Balooch SB, Neumann DL, Boschen MJ. Extinction treatment in multiple contexts attenuates ABC renewal in humans. Behaviour Research and Therapy. 2012;50:604–609. doi: 10.1016/j.brat.2012.06.003. http://dx.doi.org/10.1016/j.brat.2012.06.003. [DOI] [PubMed] [Google Scholar]
- Barlow DH, O’Brien GT, Last CG. Couples treatment of agoraphobia. Behavior Therapy. 1984;15:41–58. http://dx.doi.org/10.1016/S0005-7894(84)80040-4. [Google Scholar]
- Baum WM. Rethinking reinforcement: Allocation, induction, and contingency. Journal of the Experimental Analysis of Behavior. 2012;97:101–124. doi: 10.1901/jeab.2012.97-101. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berry MS, Sweeney MM, Odum AL. Effects of baseline reinforcement rate on operant ABA and ABC renewal. Behavioural Processes. 2014;108:87–93. doi: 10.1016/j.beproc.2014.09.009. http://dx.doi.org/10.1016/j.beproc.2014.09.009. [DOI] [PubMed] [Google Scholar]
- Betz AM, Fisher WW, Roane HS, Mintz JC, Owen TM. A component analysis of schedule thinning during functional communication training. Journal of Applied Behavior Analysis. 2013;46:219–241. doi: 10.1002/jaba.23. http://dx.doi.org/10.1002/jaba.23. [DOI] [PubMed] [Google Scholar]
- Boutelle KN, Bouton ME. Implications of learning theory for developing programs to decrease overeating. Appetite. 2015;93:62–74. doi: 10.1016/j.appet.2015.05.013. http://dx.doi.org/10.1016/j.appet.2015.05.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouton ME. Context and ambiguity in the extinction of emotional learning: Implications for exposure therapy. Behaviour Research and Therapy. 1988;26:137–149. doi: 10.1016/0005-7967(88)90113-1. http://dx.doi.org/10.1016/0005-7967(88)90113-1. [DOI] [PubMed] [Google Scholar]
- Bouton ME. Context, time, and memory retrieval in the interference paradigm of Pavlovian learning. Psychological Bulletin. 1993;114:80–99. doi: 10.1037/0033-2909.114.1.80. [DOI] [PubMed] [Google Scholar]
- Bouton ME. Context, ambiguity, and unlearning: Sources of relapse after behavioral extinction. Biological Psychiatry. 2002;52:976–986. doi: 10.1016/s0006-3223(02)01546-9. http://dx.doi.org/10.1016/S0006-3223(02)01546-9. [DOI] [PubMed] [Google Scholar]
- Bouton ME. Context and behavioral processes in extinction. Learning & Memory. 2004;11:485–494. doi: 10.1101/lm.78804. http://dx.doi.org/10.1101/lm.78804. [DOI] [PubMed] [Google Scholar]
- Bouton ME. Learning and the persistence of appetite: Extinction and the motivation to eat and overeat. Physiology and Behavior. 2011;103:51–58. doi: 10.1016/j.physbeh.2010.11.025. http://dx.doi.org/10.1016/j.physbeh.2010.11.025. [DOI] [PubMed] [Google Scholar]
- Bouton ME. Why behavior change is difficult to sustain. Preventative Medicine. 2014;68:29–36. doi: 10.1016/j.ypmed.2014.06.010. http://dx.doi.org/10.1016/j.ypmed.2014.06.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouton ME. Learning and behavior: A contemporary synthesis. 2nd. Sunderland, MA: Sinauer Associates; 2016. [Google Scholar]
- Bouton ME, Bolles RC. Contextual control of the extinction of conditioned fear. Learning and Motivation. 1979;10:445–466. doi: 10.1016/0023-9690(79)90057-2. [DOI] [Google Scholar]
- Bouton ME, Kenney FA, Rosengard C. State-dependent fear extinction with two benzodiazepine tranquilizers. Behavioral Neuroscience. 1990;104:44–55. doi: 10.1037//0735-7044.104.1.44. http://dx.doi.org/10.1037/0735-7044.104.1.44. [DOI] [PubMed] [Google Scholar]
- Bouton ME, King DA. Contextual control of the extinction of conditioned fear: Tests for the associative value of the context. Journal of Experimental Psychology: Animal Behavior Processes. 1983;9:248–265. doi: 10.1037/0097-7403.9.3.248. [DOI] [PubMed] [Google Scholar]
- Bouton ME, Peck CA. Context effects on conditioning, extinction, and reinstatement in an appetitive conditioning preparation. Animal Learning & Behavior. 1989;17:188–198. http://dx.doi.org/10.3758/BF03207634. [Google Scholar]
- Bouton ME, Ricker ST. Renewal of extinguished responding in a second context. Animal Learning & Behavior. 1994;22:317–324. http://dx.doi.org/10.3758/BF03209840. [Google Scholar]
- Bouton ME, Todd TP. A fundamental role for context in instrumental learning and extinction. Behavioural Processes. 2014;104:13–19. doi: 10.1016/j.beproc.2014.02.012. http://dx.doi.org/10.1016/j.beproc.2014.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouton ME, Todd TP, León SP. Contextual control of discriminated operant behavior. Journal of Experimental Psychology: Animal Learning and Cognition. 2014;40:92–105. doi: 10.1037/xan0000002. http://dx.doi.org/10.1037/xan0000002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouton ME, Schepers ST. Renewal after the punishment of free operant behavior. Journal of Experimental Psychology: Animal Learning and Cognition. 2015;41:81–90. doi: 10.1037/xan0000051. http://dx.doi.org/10.1037/xan0000051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouton ME, Todd TP, Vurbic D, Winterbauer NE. Renewal after the extinction of free operant behavior. Learning & Behavior. 2011;39:57–67. doi: 10.3758/s13420-011-0018-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouton ME, Vurbic D, Woods AM. D-Cycloserine facilitates context-specific fear extinction learning. Neurobiology of Learning and Memory. 2008;90:504–510. doi: 10.1016/j.nlm.2008.07.003. http://dx.doi.org/10.1016/j.nlm.2008.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bouton ME, Winterbauer NE, Todd TP. Relapse processes after the extinction of instrumental learning: Renewal, resurgence, and reacquisition. Behavioural Processes. 2012;90:130–141. doi: 10.1016/j.beproc.2012.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brooks DC, Bouton ME. A retrieval cue for extinction attenuates spontaneous recovery. Journal of Experimental Psychology: Animal Behavior Processes. 1993;19:77–89. doi: 10.1037//0097-7403.19.1.77. http://dx.doi.org/10.1037/0097-7403.19.1.77. [DOI] [PubMed] [Google Scholar]
- Brooks DC, Bouton ME. A retrieval cue for extinction attenuates response recovery (renewal) caused by a return to the conditioning context. Journal of Experimental Psychology: Animal Behavior Processes. 1994;20:366–379. doi: 10.1037//0097-7403.19.1.77. http://dx.doi.org/10.1037/0097-7403.20.4.366. [DOI] [PubMed] [Google Scholar]
- Charlop MH, Schreibman L, Thibodeau MG. Increasing spontaneous verbal responding in autistic children using a time delay procedure. Journal of Applied Behavior Analysis. 1985;18:155–166. doi: 10.1901/jaba.1985.18-155. http://dx.doi.org/10.1901/jaba.1985.18-155. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chaudri N, Sahuque LL, Janak PH. Ethanol seeking triggered by environmental context is attenuated by blocking dopamine D1 receptors in the accumbens core and shell in rats. Psychopharmacology. 2009;207:303–314. doi: 10.1007/s00213-009-1657-6. http://dx.doi.org/10.1007/s00213-009-1657-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Collins BN, Brandon TH. Effects of extinction context and retrieval cues on alcohol cue reactivity among nonalcoholic drinkers. Journal of Consulting and Clinical Psychology. 2002;70:390–397. http://dx.doi.org/10.1037/0022-006X.70.2.390. [PubMed] [Google Scholar]
- Crombag HS, Bossert JM, Koya E, Shaham Y. Context-induced relapse to drug seeking: A review. Philosophical Transactions of the Royal Society B: Biological Sciences. 2008;363:3233–3243. doi: 10.1098/rstb.2008.0090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Crombag HS, Shaham Y. Renewal of drug seeking by contextual cues after prolonged extinction in rats. Behavioral Neuroscience. 2002;116:169–173. doi: 10.1037//0735-7044.116.1.169. http://dx.doi.org/10.1037/0735-7044.116.1.169. [DOI] [PubMed] [Google Scholar]
- Cushing LS, Kennedy CH. Academic effects of providing peer support in general education classrooms on students without disabilities. Journal of Applied Behavior Analysis. 1997;30:139–151. doi: 10.1901/jaba.1997.30-139. [DOI] [PMC free article] [PubMed] [Google Scholar]
- da Silva SP, Maxwell ME, Lattal KA. Concurrent resurgence and behavioral history. Journal of the Experimental Analysis of Behavior. 2008;90:313–331. doi: 10.1901/jeab.2008.90-313. http://dx.doi.org/10.1901/jeab.2008.90-313. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Davidson TL. Learning about deprivation intensity stimuli. Behavioral Neuroscience. 1987;101:198–208. doi: 10.1037//0735-7044.101.2.198. http://dx.doi.org/10.1037/0735-7044.101.2.198. [DOI] [PubMed] [Google Scholar]
- Davidson TL. The nature and function of interoceptive signals to feed: Toward integration of physiological and learning perspectives. Psychological Review. 1993;100:640–657. doi: 10.1037/0033-295x.100.4.640. http://dx.doi.org/10.1037/0033-295X.100.4.640. [DOI] [PubMed] [Google Scholar]
- DeLeon IG. The aesthetics of intervention in the defense of the esoteric. The Behavior Analyst. 2011;34:41–45. doi: 10.1007/BF03392233. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Denniston JC, Chang RC, Miller RR. Massive extinction treatment attenuates the renewal effect. Learning and Motivation. 2003;34:68–86. http://dx.doi.org/10.1016/S0023-9690(02)00508-8. [Google Scholar]
- Dibbets P, Havermans R, Arntz A. All we need is a cue to remember: The effect of an extinction cue on renewal. Behaviour Research and Therapy. 2008;46:1070–1077. doi: 10.1016/j.brat.2008.05.007. [DOI] [PubMed] [Google Scholar]
- Dibbets P, Maes JR. The effect of an extinction cue on ABA-renewal: Does valence matter? Learning & Motivation. 2011;42:133–144. http://dx.doi.org/10.1016/j.lmot.2010.12.003. [Google Scholar]
- Doughty AH, Reed P, Lattal KA. Differential reinstatement predicted by preextinction response rate. Psychonomic Bulletin & Review. 2004;11:1118–1123. doi: 10.3758/bf03196746. http://dx.doi.org/10.3758/BF03196746. [DOI] [PubMed] [Google Scholar]
- Dunlap G. Promoting generalization. In: Van Houten R, Axelrod S, editors. Behavior analysis and treatment. New York, NY: Plenum Press; 1993. pp. 269–296. [Google Scholar]
- Dunlap G, Kern-Dunlap L, Clarke S, Robbins FR. Functional assessment, curricular revision, and severe behavior problems. Journal of Applied Behavior Analysis. 1991;24:387–397. doi: 10.1901/jaba.1991.24-387. http://dx.doi.org/10.1901/jaba.1991.24-387. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Durand VM, Carr EG. Functional communication training to reduce challenging behavior: Maintenance and application in new settings. Journal of Applied Behavior Analysis. 1991;24:251–264. doi: 10.1901/jaba.1991.24-251. http://dx.doi.org/10.1901/jaba.1991.24-251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Estes WK. An experimental study of punishment. Psychological Monographs. 1944;57:i–40. [Google Scholar]
- Falcomata TS, Wacker DP. On the use of strategies for programming generalization during functional communication training: A review of the literature. Journal of Developmental and Physical Disabilities. 2013;25:5–15. http://dx.doi.org/10.1007/s10882-012-9311-3. [Google Scholar]
- Fisher WW, Greer BD, Fuhrman AM, Querim AC. Using multiple schedules during functional communication training to promote rapid transfer of treatment effects. Journal of Applied Behavior Analysis. 2015;48:713–733. doi: 10.1002/jaba.254. http://dx.doi.org/10.1002/jaba.254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fisher WW, Kuhn DE, Thompson RH. Establishing discriminative control of responding using functional and alternative reinforcers during functional communication training. Journal of Applied Behavior Analysis. 1998;31:543–560. doi: 10.1901/jaba.1998.31-543. http://dx.doi.org/10.1901/jaba.1998.31-543. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fox AT, Smethells JR, Reilly MP. Flash rate discrimination in rats: Rate bisection and generalization peak shift. Journal of the Experimental Analysis of Behavior. 2013;100:211–221. doi: 10.1002/jeab.36. http://dx.doi.org/10.1002/jeab.36. [DOI] [PubMed] [Google Scholar]
- Fuhrman AM, Fisher WW, Greer BD. A preliminary investigation on improving functional communication training by mitigating resurgence of destructive behavior. Journal of Applied Behavior Analysis. 2016;49:884–899. doi: 10.1002/jaba.338. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Garcia E. The training and generalization of a conversational speech form in nonverbal retardates. Journal of Applied Behavior Analysis. 1974;7:137–149. doi: 10.1901/jaba.1974.7-137. http://dx.doi.org/10.1901/jaba.1974.7-137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Glautier S, Elgueta T, Nelson JB. Extinction produces context inhibition and multiple-context extinction reduces response recovery in human predictive learning. Learning & Behavior. 2013;41:341–352. doi: 10.3758/s13420-013-0109-7. http://dx.doi.org/10.3758/s13420-013-0109-7. [DOI] [PubMed] [Google Scholar]
- Godden DR, Baddeley AD. Context-dependent memory in two natural environments: On land and underwater. British Journal of Psychology. 1975;66:325–331. http://dx.doi.org/10.1111/j.2044-8295.1975.tb01468.x. [Google Scholar]
- Goodwin DW, Powell B, Bremer D, Hoine H, Stern J. Alcohol and recall: State-dependent effects in man. Science. 1969;193:1358–1360. doi: 10.1126/science.163.3873.1358. http://dx.doi.org/10.1126/science.163.3873.1358. [DOI] [PubMed] [Google Scholar]
- Greer BD, Fisher WW, Saini V, Owen TM, Jones JK. Functional communication training during reinforcement schedule thinning: An analysis of 25 applications. Journal of Applied Behavior Analysis. 2016;49:105–121. doi: 10.1002/jaba.265. http://dx.doi.org/10.1002/jaba.265. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Griffiths H, Craighead WE. Generalization in operant speech therapy for misarticulation. Journal of Speech and Hearing Disorders. 1972;37:485–494. doi: 10.1044/jshd.3704.485. [DOI] [PubMed] [Google Scholar]
- Gunther LM, Denniston JC, Miller RR. Conducting exposure treatment in multiple contexts can prevent relapse. Behaviour Research and Therapy. 1998;36:75–91. doi: 10.1016/S0005-7967(97)10019-5. [DOI] [PubMed] [Google Scholar]
- Guttman N, Kalish HI. Discriminability and stimulus generalization. Journal of Experimental Psychology. 1956;51:79–88. doi: 10.1037/h0046219. http://dx.doi.org/10.1037/h0046219. [DOI] [PubMed] [Google Scholar]
- Hagopian LP, Gonzáles ML, Rivet TT, Triggs M, Clark SB. Response interruption and differential reinforcement of alternative behavior for the treatment of pica. Behavioural Interventions. 2011;26:309–325. http://dx.doi.org/10.1002/bin.339. [Google Scholar]
- Hake DF. The basic-applied continuum and the possible evolution of human operant social and verbal research. The Behavior Analyst. 1982;5:21–28. doi: 10.1007/BF03393137. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hall SS, Maynes NP, Reiss AL. Using percentile schedules to increase eye contact in children with Fragile X syndrome. Journal of Applied Behavior Analysis. 2009;42:171–176. doi: 10.1901/jaba.2009.42-171. http://dx.doi.org/10.1901/jaba.2009.42-171. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hanley GP, Iwata BA, Thompson RH. Reinforcement schedule thinning following treatment with functional communication training. Journal of Applied Behavior Analysis. 2001;34:17–38. doi: 10.1901/jaba.2001.34-17. http://dx.doi.org/10.1901/jaba.2001.34-17. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Honig WK, Urcuioli P. The legacy of Guttman & Kalish (1956): 25 years of research on stimulus generalization. Journal of the Experimental Analysis of Behavior. 1981;36:405–445. doi: 10.1901/jeab.1981.36-405. http://dx.doi.org/10.1901/jeab.1981.36-405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Iwata BA, Dorsey MF, Slifer KJ, Bauman KE, Richman GS. Toward a functional analysis of self-injury. Journal of Applied Behavior Analysis. 1994;27:197–209. doi: 10.1901/jaba.1994.27-197. Reprinted from Analysis and Intervention in Developmental Disabilities, 2, 3–20, 1982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kazdin AE, Bootzin RR. The token economy: an evaluative review. Journal of Applied Behavior Analysis. 1972;5:343–372. doi: 10.1901/jaba.1972.5-343. http://dx.doi.org/10.1901/jaba.1972.5-343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kearns DN, Weiss SJ. Contextual renewal of cocaine seeking in rats and its attenuation by the conditioned effects of an alternative reinforcer. Drug and Alcohol Dependence. 2007;90:193–202. doi: 10.1016/j.drugalcdep.2007.03.006. [DOI] [PubMed] [Google Scholar]
- Kelley ME, Liddon CJ, Ribeiro A, Greif AE, Podlesnik CA. Basic and translational evaluation of renewal of operant responding. Journal of Applied Behavior Analysis. 2015;48:390–401. doi: 10.1002/jaba.209. http://dx.doi.org/10.1002/jaba.209. [DOI] [PubMed] [Google Scholar]
- Kincaid SL, Lattal KA, Spence J. Super-resurgence: ABA renewal increases resurgence. Behavioural Processes. 2015;115:70–73. doi: 10.1016/j.beproc.2015.02.013. http://dx.doi.org/10.1016/j.beproc.2015.02.013. [DOI] [PubMed] [Google Scholar]
- King JE, Hayes LJ. The role of discriminative stimuli on response patterns in resurgence. The Psychological Record. 2016;66:325–335. doi: 10.1007/s40732-016-0175-2. [DOI] [Google Scholar]
- Laborda MA, Polack CW, Miguez G, Miller RR. Behavioral techniques for attenuating the expression of fear associations in an animal model of anxiety. Journal of Behavior Therapy and Experimental Psychiatry. 2014;45:343–350. doi: 10.1016/j.jbtep.2014.02.005. http://dx.doi.org/10.1016/j.jbtep.2014.02.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lalli JS, Casey S, Kates K. Reducing escape behavior and increasing task completion with functional communication training, extinction, and response chaining. Journal of Applied Behavior Analysis. 1997;28:261–268. doi: 10.1901/jaba.1995.28-261. http://dx.doi.org/10.1901/jaba.1995.28-261. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lang R, Sigafoos J, Lancioni G, Didden R, Rispoli M. Influence of assessment setting on the results of functional analyses of problem behavior. Journal of Applied Behavior Analysis. 2010;43:565–567. doi: 10.1901/jaba.2010.43-565. http://dx.doi.org/10.1901/jaba.2010.43-565. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lattal KA, St Peter C, Escobar R. Operant extinction: Elimination and generation of behavior. In: Madden GJ, Dube WV, Hackenberg TD, Hanley GP, Lattal KA, editors. APA handbook of behavior analysis, Vol 2: Translating principles into practice. Washington, DC: American Psychological Association; 2013. pp. 77–107. http://dx.doi.org/10.1037/13938-004. [Google Scholar]
- Lazareva O. Relational learning in a context of transposition: A review. Journal of the Experimental Analysis of Behavior. 2012;97:231–248. doi: 10.1901/jeab.2012.97-231. http://dx.doi.org/10.1901/jeab.2012.97-231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lerman DC. From the laboratory to community application: Translational research in behavior analysis. Journal of Applied Behavior Analysis. 2003;36:415–419. doi: 10.1901/jaba.2003.36-415. http://dx.doi.org/10.1901/jaba.2003.36-415. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lerman DC, Iwata BA. Prevalence of the extinction burst and its attenuation during treatment. Journal of Applied Behavior Analysis. 1995;28:93–94. doi: 10.1901/jaba.1995.28-93. http://dx.doi.org/10.1901/jaba.1995.28-93. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lerman DC, Iwata BA. Developing a technology for the use of operant extinction in clinical settings: An examination of basic and applied research. Journal of Applied Behavior Analysis. 1996;29:345–382. doi: 10.1901/jaba.1996.29-345. http://dx.doi.org/10.1901/jaba.1996.29-345. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Lerman DC, Iwata BA, Smith RG, Zarcone JR, Vollmer TR. Transfer of behavioral function as a contributing factor in treatment relapse. Journal of Applied Behavior Analysis. 1994;27:357–370. doi: 10.1901/jaba.1994.27-357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luczynski KC, Hanley GP, Rodriguez NM. An evaluation of the generalization and maintenance of functional communication and self-control skills with preschoolers. Journal of Applied Behavior Analysis. 2014;47:246–263. doi: 10.1002/jaba.128. [DOI] [PubMed] [Google Scholar]
- Mace FC. Basic research needed for stimulating the development of behavioral technologies. Journal of the Experimental Analysis of Behavior. 1994;61:529–550. doi: 10.1901/jeab.1994.61-529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mace FC, Critchfield TS. Translational research in behavior analysis: Historical traditions and imperative for the future. Journal of the Experimental Analysis of Behavior. 2010;93:293–312. doi: 10.1901/jeab.2010.93-293. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mace FC, McComas JJ, Mauro BC, Progar PR, Ervin R, Zangrillo AN. Differential reinforcement of alternative behavior increases resistance to extinction: Clinical demonstration, animal modeling, and clinical test of one solution. Journal of the Experimental Analysis of Behavior. 2010;93:349–367. doi: 10.1901/jeab.2010.93-349. http://dx.doi.org/10.1901/jeab.2010.93-349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McConnell BL, Miller RR. Associative accounts of recovery-from-extinction effects. Learning and Motivation. 2014;46:1–15. doi: 10.1016/j.lmot.2014.01.003. http://dx.doi.org/10.1016/j.lmot.2014.01.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McKenzie SD, Smith RG, Simmons JN, Soderlund MJ. Using a stimulus correlated with reprimands to suppress automatically maintained eye poking. Journal of Applied Behavior Analysis. 2008;41:255–259. doi: 10.1901/jaba.2008.41-255. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Murphy ES, McSweeney FK, Smith RG, McComas JJ. Dynamic changes in reinforcer effectiveness: Theoretical, methodological, and practical implications for applied research. Journal of Applied Behavior Analysis. 2003;36:421–438. doi: 10.1901/jaba.2003.36-421. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mystkowski JL, Craske MG, Echiverri AM, Labus JS. Mental reinstatement of context and return of fear in spider-fearful participants. Behavior Therapy. 2006;37:49–60. doi: 10.1016/j.beth.2005.04.001. http://dx.doi.org/10.1016/j.beth.2005.04.001. [DOI] [PubMed] [Google Scholar]
- Nakajima S, Tanaka S, Urushihara K, Imada H. Renewal of extinguished lever- press responses upon return to the training context. Learning and Motivation. 2000;31:416–431. http://dx.doi.org/10.1006/lmot.2000.1064. [Google Scholar]
- Nakajima S, Urushihara K, Masaki T. Renewal of operant performance formerly eliminated by omission or non-contingency training upon return to the acquisition context. Learning and Motivation. 2002;33:510–525. http://dx.doi.org/10.1016/S0023-9690(02)00009-7. [Google Scholar]
- Nelson JB, Sanjuan MDC, Vadillo-Ruiz S, Pérez J, León SP. Experimental renewal in human participants. Journal of Experimental Psychology: Animal Behavior Processes. 2011;37:58–70. doi: 10.1037/a0020519. [DOI] [PubMed] [Google Scholar]
- Nevin JA, Craig AR, Cunningham PJ, Podlesnik CA, Shahan TA, Sweeney MM. Quantitative models of persistence and relapse from the perspective of behavioral momentum theory: Fits and misfits. Behavioural Processes. doi: 10.1016/j.beproc.2017.04.016. in press. https://doi.org/10.1016/j.beproc.2017.04.016. [DOI] [PMC free article] [PubMed]
- Nevin JA, Wacker DP. Response strength and persistence. In: Madden GJ, Dube WV, Hackenberg T, Hanley GP, Lattal KA, editors. APA handbooks in psychology: APA handbook of behavior analysis, Vol 2: Translating principles into practice. Washington, DC: American Psychological Association; 2013. pp. 109–128. http://dx.doi.org/10.1037/13938-005. [Google Scholar]
- Osnes PG, Lieblein T. An explicit technology of generalization. The Behavior Analyst Today. 2003;3:364–374. http://dx.doi.org/10.1037/h0099994. [Google Scholar]
- Petscher ES, Rey C, Bailey JA. A review of empirical support for differential reinforcement of alternative behavior. Research in Developmental Disabilities. 2009;30:409–425. doi: 10.1016/j.ridd.2008.08.008. http://dx.doi.org/10.1016/j.ridd.2008.08.008. [DOI] [PubMed] [Google Scholar]
- Piazza CC, Hanley GP, Fisher WW. Functional analysis and treatment of cigarette pica. Journal of Applied Behavior Analysis. 1996;29:437–450. doi: 10.1901/jaba.1996.29-437. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Podlesnik CA, Bai JYH. Method of stimulus combination impacts resistance to extinction. Journal of the Experimental Analysis of Behavior. 2015;104:30–47. doi: 10.1002/jeab.155. http://dx.doi.org/10.1002/jeab.155. [DOI] [PubMed] [Google Scholar]
- Podlesnik CA, Bai JYH, Elliffe D. Resistance to extinction and relapse in combined stimulus contexts. Journal of the Experimental Analysis of Behavior. 2012;98:169–189. doi: 10.1901/jeab.2012.98-169. http://dx.doi.org/10.1901/jeab.2012.98-169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Podlesnik CA, Bai JYH, Skinner KA. Assessing the role of alternative response rate and reinforcer rates in resistance to extinction of target responding when combining stimuli. Journal of the Experimental Analysis of Behavior. 2016;105:427–444. doi: 10.1002/jeab.206. http://dx.doi.org/10.1002/jeab.206. [DOI] [PubMed] [Google Scholar]
- Podlesnik CA, DeLeon IG. Behavioral momentum theory: Understanding persistence and improving treatment. In: DiGennaro Reed FD, Reed DD, editors. Bridging the gap between science and practice in autism service delivery. New York: Springer; 2015. pp. 327–351. [Google Scholar]
- Podlesnik CA, Kelley ME. Resurgence: Response competition, stimulus control, and reinforcer control. Journal of the Experimental Analysis of Behavior. 2014;102:231–240. doi: 10.1002/jeab.102. http://dx.doi.org/10.1002/jeab.102. [DOI] [PubMed] [Google Scholar]
- Podlesnik CA, Kelley ME. Translational research on the relapse of operant behavior. Mexican Journal of Behavior Analysis. 2015;41:226–251. [Google Scholar]
- Podlesnik CA, Miranda-Dukoski L. Stimulus generalization and operant context renewal. Behavioural Processes. 2015;119:93–98. doi: 10.1016/j.beproc.2015.07.015. http://dx.doi.org/10.1016/j.beproc.2015.07.015. [DOI] [PubMed] [Google Scholar]
- Podlesnik CA, Shahan TA. Behavioral momentum and relapse of extinguished operant responding. Learning & Behavior. 2009;37:357–364. doi: 10.3758/LB.37.4.357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Podlesnik CA, Shahan TA. Extinction, relapse, and behavioral momentum. Behavioural Processes. 2010;84:400–411. doi: 10.1016/j.beproc.2010.02.001. http://dx.doi.org/10.1016/j.beproc.2010.02.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Polack CW, Laborda MA, Miller RR. On the differences in degree of renewal produced by the different renewal designs. Behavioural Processes. 2013;99:112–120. doi: 10.1016/j.beproc.2013.07.006. http://dx.doi.org/10.1016/j.beproc.2013.07.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pritchard D, Hoerger M, Mace FC. Treatment relapse and behavioral momentum theory. Journal of Applied Behavior Analysis. 2014;47:814–833. doi: 10.1002/jaba.163. http://dx.doi.org/10.1002/jaba.163. [DOI] [PubMed] [Google Scholar]
- Rescorla RA. Experimental extinction. In: Mowrer RR, Klein SB, editors. Handbook of contemporary learning theories. Mahwah, NJ: Lawrence Erlbaum Associates Publishers; 2001. pp. 119–154. [Google Scholar]
- Rescorla RA. Within-subject renewal in sign tracking. Quarterly Journal of Experimental Psychology. 2008;61:1793–1802. doi: 10.1080/17470210701790099. [DOI] [PubMed] [Google Scholar]
- Rincover A, Koegel RL. Setting generality and stimulus control in autistic children. Journal of Applied Behavior Analysis. 1975;8:235–246. doi: 10.1901/jaba.1975.8-235. http://dx.doi.org/10.1901/jaba.1975.8-235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Roane HS, Kelly ML, Fisher WW. The effects of noncontingent access to food on the rate of object mouthing across three settings. Journal of Applied Behavior Analysis. 2003;36:579–582. doi: 10.1901/jaba.2003.36-579. http://dx.doi.org/10.1901/jaba.2003.36-579. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rooker GW, Jessel J, Kurtz PF, Hagopian LP. Functional communication training with and without alternative reinforcement and punishment: an analysis of 58 applications. Journal of Applied Behavior Analysis. 2013;46:708–722. doi: 10.1002/jaba.76. http://dx.doi.org/10.1002/jaba.76. [DOI] [PubMed] [Google Scholar]
- Rosenthal R. The “file drawer problem” and tolerance for null results. Psychological Bulletin. 1979;86:838–641. http://dx.doi.org/10.1037/0033-2909.86.3.638. [Google Scholar]
- Saini V, Fisher WW. Evaluating the effects of discriminability on behavioral persistence during and following time-based reinforcement. Journal of the Experimental Analysis of Behavior. 2016;106:195–209. doi: 10.1002/jeab.225. http://dx.doi.org/10.1002/jeab.225. [DOI] [PubMed] [Google Scholar]
- Schindler HS, Horner RH. Generalized reduction of challenging behavior of young children with autism: Building transituational interventions. American Journal on Mental Retardation. 2005;110:36–47. doi: 10.1352/0895-8017(2005)110<36:GROPBO>2.0.CO;2. [DOI] [PubMed] [Google Scholar]
- Shiban Y, Pauli P, Mühlberger A. Effect of multiple context exposure on renewal in spider phobia. Behaviour Research and Therapy. 2013;51:68–74. doi: 10.1016/j.brat.2012.10.007. http://dx.doi.org/10.1016/j.brat.2012.10.007. [DOI] [PubMed] [Google Scholar]
- Shillingsburg MA, Hollander DL, Yosick RN, Bowen C, Muskat LR. Stimulus-stimulus pairing to increase vocalizations in children with language delays: A review. The Analysis of Verbal Behavior. 2015;31:215–235. doi: 10.1007/s40616-015-0042-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Smith SM. Context: a reference for focal experience. In: Roediger HL, Dudai Y, Fitzpatrick SM, editors. Science of memory concepts. New York: Oxford University Press; 2007. pp. 111–114. [Google Scholar]
- Stokes TF, Baer DM. An implicit technology of generalization. Journal of Applied Behavior Analysis. 1977;10:349–367. doi: 10.1901/jaba.1977.10-349. http://dx.doi.org/10.1901/jaba.1977.10-349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Stokes TF, Doud CL, Rowbury TG, Baer DM. Peer facilitation of generalization in a preschool classroom. Journal of Abnormal Child Psychology. 1978;6:203–209. doi: 10.1007/BF00919125. http://dx.doi.org/10.1007/BF00919125. [DOI] [PubMed] [Google Scholar]
- Stokes TF, Osnes PG. An operant pursuit of generalization. Behavior Therapist. 1989;20:337–355. doi: 10.1016/j.beth.2016.08.012. http://dx.doi.org/10.1016/S0005-7894(89)80054-1. [DOI] [PubMed] [Google Scholar]
- St Peter Pipkin C, Vollmer TR, Sloman KN. Effects of treatment integrity failures during differential reinforcement of alternative behavior: A translational model. Journal of Applied Behavior Analysis. 2010;43:47–70. doi: 10.1901/jaba.2010.43-47. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sweeney MM, Shahan TA. Effects of high, low, and thinning rates of alternative reinforcement on response elimination and resurgence. Journal of the Experimental Analysis of Behavior. 2013;100:102–116. doi: 10.1002/jeab.26. http://dx.doi.org/10.1002/jeab.26. [DOI] [PubMed] [Google Scholar]
- Thomas BL, Larsen N, Ayres JJB. Role of context similarity in ABA, ABC, and AAB renewal paradigms: Implications for theories of renewal and for treating human phobias. Learning and Motivation. 2003;34:410–436. doi: 10.1016/S0023-9690(03)00037-7. [DOI] [Google Scholar]
- Thomas BL, Vurbic D, Novak C. Extensive extinction in multiple contexts eliminates the renewal of conditioned fear in rats. Learning and Motivation. 2009;40:147–159. http://dx.doi.org/10.1016/j.lmot.2008.10.002. [Google Scholar]
- Thrailkill EA, Shahan TA. Resistance to change and relapse of observing. Journal of the Experimental Analysis of Behavior. 2012;97:281–304. doi: 10.1901/jeab.2012.97-281. http://dx.doi.org/10.1901/jeab.2012.97-281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tiger JH, Hanley GP, Bruzek J. Functional communication training: A review and practical guide. Behavior Analysis in Practice. 2008;1:16–23. doi: 10.1007/BF03391716. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todd TP. Mechanisms of renewal after the extinction of instrumental behavior. Journal of Experimental Psychology: Animal Behavior Processes. 2013;39:193–207. doi: 10.1037/a0032236. http://dx.doi.org/10.1037/a0032236. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todd TP, Vurbic D, Bouton ME. Mechanisms of renewal after the extinction of discriminated operant behavior. Journal of Experimental Psychology: Animal Learning and Cognition. 2014;40:355–368. doi: 10.1037/xan0000021. http://dx.doi.org/10.1037/xan0000021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Todd TP, Winterbauer NE, Bouton ME. Effects of the amount of acquisition and contextual generalization on the renewal of instrumental behavior after extinction. Learning & Behavior. 2012;40:145–157. doi: 10.3758/s13420-011-0051-5. http://dx.doi.org/10.3758/s13420-011-0051-5. [DOI] [PubMed] [Google Scholar]
- Trask S, Bouton ME. Discriminative properties of the reinforcer can be used to attenuate the renewal of extinguished operant behavior. Learning & Behavior. 2016;44:151–161. doi: 10.3758/s13420-015-0195-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vansteenwegen D, Vervliet B, Hermans D, Beckers T, Baeyens F, Eelen P. Stronger renewal in human fear conditioning when tested with an acquisition retrieval cue than with an extinction retrieval cue. Behaviour Research and Therapy. 2006;44:1717–1725. doi: 10.1016/j.brat.2005.10.014. [DOI] [PubMed] [Google Scholar]
- Vervliet B, Craske MG, Hermans D. Fear extinction and relapse: State of the art. Annual Review of Clinical Psychology. 2013;9:215–248. doi: 10.1146/annurev-clinpsy-050212-185542. http://dx.doi.org/10.1146/annurev-clinpsy-050212-185542. [DOI] [PubMed] [Google Scholar]
- Vollmer TR. Three variations of translational research: Comments on Critchfield (2011) The Behavior Analyst. 2011;34:31–35. doi: 10.1007/BF03392231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Vurbic D, Bouton ME. A contemporary behavioral perspective on extinction. In: McSweeney FK, Murphy ES, editors. The Wiley Blackwell handbook of operant and classical conditioning. Boston: Wiley-Blackwell; 2014. pp. 53–76. http://dx.doi.org/10.1002/9781118468135.ch3. [Google Scholar]
- Wacker DP, Berg WK, Harding JW, Barretto A, Rankin B, Ganzer J. Treatment effectiveness, stimulus generalization, and acceptability to parents of functional communication training. Educational Psychology. 2005;25:233–256. http://dx.doi.org/10.1080/0144341042000301184. [Google Scholar]
- Willcocks AL, McNally GP. An extinction retrieval cue attenuates renewal but not reacquisition of alcohol seeking. Behavioral Neuroscience. 2014;128:83–91. doi: 10.1037/a0035595. http://dx.doi.org/10.1037/a0035595. [DOI] [PubMed] [Google Scholar]
- Winterbauer NE, Lucke S, Bouton ME. Some factors modulating the strength of resurgence after extinction of an instrumental behavior. Learning & Motivation. 2013;44:60–71. doi: 10.1016/j.lmot.2012.03.003. http://dx.doi.org/10.1016/j.lmot.2012.03.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zironi I, Burattini C, Aircardi G, Janak PH. Context is a trigger for relapse to alcohol. Behavioural Brain Research. 2006;167:150–155. doi: 10.1016/j.bbr.2005.09.007. http://dx.doi.org/10.1016/j.bbr.2005.09.007. [DOI] [PubMed] [Google Scholar]