

Abstract
Background
Most gastrointestinal stromal tumors (GISTs) harbor mutually exclusive gain of function mutations in the receptor tyrosine kinase (RTK) KIT (70–80%) or in the related receptor PDGFRA (~10%). These GISTs generally respond well to therapy with the RTK inhibitor imatinib mesylate (IM), although initial response is genotype-dependent. An alternate mechanism leading to GIST oncogenesis is deficiency in the succinate dehydrogenase (SDH) enzyme complex resulting from genetic or epigenetic inactivation of one of the four SDH subunit genes (SDHA, SDHB, SDHC, SDHD, collectively referred to as SDHX). SDH loss of function is generally seen only in GIST lacking RTK mutations, and SDH-deficient GIST respond poorly to imatinib therapy.
Methods
Tumor and normal DNA from a GIST case carrying the IM-resistant PDGFRA D842V mutation was analyzed by whole exome sequencing (WES) to identify additional potential targets for therapy. The tumors analyzed were separate recurrences following progression on imatinib, sunitinib, and the experimental PDGFRA inhibitor crenolanib. Tumor sections from the GIST case and a panel of ~75 additional GISTs were subjected to immunohistochemistry (IHC) for the SDHB subunit.
Results
Surprisingly, a somatic, loss of function mutation in exon 4 of the SDHB subunit gene (c.291_292delCT, p.I97Mfs*21) was identified in both tumors. Sanger sequencing confirmed the presence of this inactivating mutation, and IHC for the SDHB subunit demonstrated that these tumors were SDH-deficient. IHC for the SDHB subunit across a panel of ~75 GIST cases failed to detect SDH deficiency in other GISTs with RTK mutations.
Conclusions
This is the first reported case of a PDGFRA mutant GIST exhibiting SDH-deficiency. A brief discussion of the relevant GIST literature is included.
Electronic supplementary material
The online version of this article (doi:10.1186/s12885-017-3499-7) contains supplementary material, which is available to authorized users.
Keywords: Gastrointestinal stromal tumor, Kit, Platelet derived growth factor receptor alpha, Succinate dehydrogenase, Imatinib mesylate, Crenolanib
Background
Gastrointestinal stromal tumor, or GIST, is the most common mesenchymal tumor of the GI tract, with an estimated annual incidence of 14–20 cases per million [1]. GIST generally presents as a sporadic disease in older adults (median age 60–65 years), and affects men and women equally. GIST may originate throughout the GI tract but occurs most commonly in the stomach (~60%) or small intestine (~25%) [2]. These tumors are related to spindle-shaped pacemaker cells of the gut known as the interstitial cells of Cajal (ICC) with which they share phenotypic characteristics including the expression of the type III receptor tyrosine kinase (RTK) KIT (CD117) [3, 4]. Mutually exclusive gain of function mutations in KIT, or in the related RTK PDGFRA, are found in ~85% of GIST, and lead to increased kinase signaling including MAPK and PI3K-AKT [5, 6]. The discovery of activating mutations in these receptors in GIST led to the approval of the RTK inhibitor imatinib mesylate (IM) as front-line therapy for GIST, and subsequent approval of sunitinib malate and regorafenib for IM-resistant disease. While most RTK-mutated GISTs are IM-sensitive, therapeutic response is influenced by genotype. Among the most common mutation types, GISTs with mutations in the juxtamembrane domain encoded by KIT exon 11 are generally IM-sensitive while GISTs harboring KIT exon 9 mutations require IM dose-escalation. The most common PDGFRA mutation, the D842V mutation in exon 18 encoding the kinase activation loop, confers primary clinical resistance to IM [7, 8].
The 10–15% of GIST that lack mutations in KIT and PDGFRA, known as wild type or, more accurately, RTK-wild type GIST, also respond poorly to IM therapy [9, 10]. RTK-wild type GIST may harbor mutations in genes that activate kinase signaling downstream of the receptors, particularly in the RAS/RAF/MAPK pathway. Germline loss of function mutations in neurofibromin, a RAS-GAP that acts as a negative regulator of RAS signaling, are linked to neurofibromatosis type 1 (NF1), and predispose NF1 individuals to development of a variety of tumor types including GIST. While NF1-related GIST has been reported to comprise ~1.5% of the patient population of GIST [11], reports of sporadic or constitutional NF1 gene mutations in undiagnosed NF1 individuals suggests this percentage may be somewhat higher [12, 13]. The gain of function V600E mutation in the BRAF gene has been found in ~7% of GISTs lacking KIT/PDGFRA mutations [14, 15], while a small number of cases with activating mutations in the KRAS gene have also been reported [16]. There is limited clinical data on the efficacy of IM or other RTK inhibitors in RAS pathway mutated GIST [2].
A distinct mechanism of oncogenesis seen in approximately 40% of RTK-wild type GIST is deficiency in the mitochondrially located tumor suppressor complex succinate dehydrogenase (SDH). SDH is a heterotetrameric enzyme complex that connects the oxidation of succinate to fumarate in the Krebs cycle to the reduction of coenzyme Q in the mitochondrial electron transport chain. Mutation or silencing of any of the four SDH genes (SDHA-D, or collectively SDHX) destabilizes the complex and results in accumulation of succinate and activation of cellular pathways that lead to increased angiogenesis and cellular proliferation [17]. SDH-deficient GIST includes GIST that occur in association with rare multitumor syndromes, the majority of GIST that occur in children, and a subset of sporadic adult gastric GISTs. The Carney-Stratakis syndrome (CSS), an inherited multi-tumor syndrome characterized by the occurrence of multifocal gastric GIST and multicentric paraganglioma (PGL) [18], is caused by germline mutations in the SDHB, SDHC, or SDHD subunit genes [19, 20]. The Carney triad (CT) is a non-familial association of gastric GIST, PGL and pulmonary chondromas that occurs primarily in young females [21]. CT and most pediatric GIST lack SDHX mutations but exhibit SDH deficiency due to epigenetic silencing of the SDHC gene through promoter hyper-methylation [22–25]. More recently, mutations in the SDHA gene subunit have been identified in a subset of sporadic adult, RTK-wild type gastric GISTs [26–31]. Analysis of large GIST sample sets has established that SDH deficiency is largely mutually exclusive to KIT/PDGFRA/BRAF/NF1 mutation [23, 32, 33].
Here we report whole exome sequencing (WES) analysis of two GISTs harboring the IM-insensitive PDGFRA D842V mutation. These tumors, resected at different times from the same patient, were found to harbor an inactivating mutation in the SDHB gene in addition to the PDGFRA mutation. Immunohistochemical (IHC) analysis demonstrated complete lack of SDHB expression in the GISTs, confirming SDH-deficiency. This report describes for the first time the co-existence of PDGFRA and SDH gene mutations in a GIST case. A brief review of the literature describing the co-occurrence of mutations in PDGFRA or KIT and SDH subunit genes follows.
Methods
Sequence analysis of patient tumor samples
The Fox Chase Biosample Repository (BSR) obtains, deposits and maintains patient samples following informed written consent. De-identified patient pathological and molecular reports available from the BSR were queried for GIST samples with mutations in PDGFRA. Tumor samples and normal blood from four patients with the IM-resistant D842V mutation were obtained from the BSR under a protocol approved by the Fox Chase Cancer Center Institutional Review Board (#03–848). Additional associated de-identified clinical data from the case in question were obtained from the BSR data warehouse that contains demographic data, clinical information and treatment outcomes. The isolation and characterization of genomic DNA for whole-exome sequencing (WES) from frozen tumor specimens has been described [12]. Exome-enriched genomic libraries (Sureselect human all exon V4, Agilent Technologies, Santa Clara, CA) from normal and tumor DNA were subjected to paired-end 100 bp sequencing on the Illumina HiSeq 2000 instrument (Illumina, San Diego, CA). Reads were mapped to the reference human genome (Hg19 corresponding v37) using the BWA aligner [34] and mapped reads were sorted, merged, and de-duplicated (Picard). Local realignments were done using GATK in areas surrounding insertions and deletions (indels) [35, 36]. Variant calling was performed using GATK UnifiedGenotyper [35, 36] and single nucleotide variants (SNVs) annotation and variant effect predictions were done with ANNOVAR [37] by querying various databases. Non-synonymous, potentially deleterious coding region variants, splice-site mutations, and insertions or deletions (indels) that were predicted to be present in the tumor only, were visually confirmed on the Integrative Genomics Viewer (IGV) [38], and confirmed by exon-based Sanger sequencing. Primer sequences for confirmation of mutations listed in Table 1 are shown in Additional file 1. Relevant exons were PCR-amplified from genomic DNA and subjected to Sanger sequencing by the Fox Chase Cancer Center DNA Sequencing Facility.
Table 1.
Confirmed deleteriousa or truncating somatic variants in GIST1 and GIST2
| Gene symbol | UniProt accessionb | Genomic coordinatec | Exon | Mutation (cDNA) | Mutation (protein) | MAFd (%), (GIST 1;GIST2) |
|---|---|---|---|---|---|---|
| PDGFRA | P16234 | chr4:55,152,093 | 18 | c.2525A > T | p.D842V | 33; 48 |
| SDHB | P21912 | chr1:17,355,226 | 4 | c.291_292delCT | p.I97Mfs*21 | 42; 61 |
| CERS2 | Q96G23 | chr1:150,939,337 | 9 | c.743C > G | p.S248* | 36; 45 |
| DNAH3 | Q8TD57 | chr16:21,049,229 | 34 | c.4804G > A | p.G1602S | 76; 86 |
| CAPN9 | O14815 | chr1:230,928,629 | 16 | c.1825G > A | p.G609S | 32; 32 |
| DIS3 | Q9Y2L1 | chr13:73,347,835 | 8 | c.1226C > T | p.S409F | 17; 39 |
| GJD2 | Q9UKL4 | chr15:35,044,921 | 2 | c.724 T > C | p.C242R | 32; 19 |
| EDN3 | P14138 | chr20:57,876,697 | 2 | c.285G > T | p.R95S | 41; 42 |
| PI4KA | P42356 | chr22:21,104,246 | 28 | c.3190A > G | p.I1064V | 43; 29 |
| TENM2 | Q9NT68 | chr5:167,645,590 | 23 | c.4667 T > A | p.I1556N | 60; 64 |
| JPH1 | Q9HDC5 | chr8:75,227,712 | 3 | c.523G > A | p.V175 M | 25; 40 |
| DIS3L2 | Q8IYB7 | chr2:232,880,352 | 2 | c.181G > T | p.E61* | 0; 89 |
| SENP6 | Q9GZR1 | chr6:76,369,037 | 7 | c.610A > G | p.K204E | 0; 58 |
| PLCG2 | P16885 | chr16:81,944,244 | 18 | c.1853G > A | p.R618H | 0; 24 |
ahttp://www.mypeg.info; bhttp://www.uniprot.org; cHg19; dmutant allele frequency
The symbol "*" in a description of a protein variant is standard nomenclature that denotes a stop codon in the protein sequence
SNP array analysis
SNP array analysis was performed using Affymetrix CytoScan HD arrays (Santa Clara, California, USA). Genomic DNA was digested with NspI restriction enzyme, adaptor-ligated, and amplified using a primer recognizing the adapter sequence. Amplification products were purified using magnetic beads, fragmented, biotin-labeled, and hybridized to arrays according to the manufacturer’s recommendations. The hybridized array was washed, scanned with a GeneChip Scanner 3000 7G, and intensities of probe hybridization were analyzed using Affymetrix GeneChip Command Console. Copy number and genotyping analyses were performed using Affymetrix Chromosome Analysis Suite software with default settings. Whole genome SNP copy number data were visualized using the Affymetrix Chromosome Analysis Suite.
Immunohistochemical analysis
IHC for CD117 (KIT) was performed as described [39]. DOG1 IHC was performed by the Fox Chase Cancer Center Clinical Pathology Laboratory using an antibody from Cell Marque (Sigma Aldrich, St. Louis, MO, USA). IHC for SDHB, and criteria used for assessing SDHB protein expression have been previously described [26, 40]. The construction of tissue microarrays (TMAs) containing ~75 clinically annotated GIST specimens has been previously described [12]. TMAs and whole tissue sections were evaluated for punctate cytoplasmic staining for SDHB. Lack of SDHB staining in GIST cells was considered informative only if typical granular cytoplasmic staining (a mitochondrial pattern) was seen in internal controls such as endothelial cells. IHC for CD31 was performed using the anti-CD31 antibody M083, clone JC70A (Dako N.A., Carpenteria, CA, USA).
Literature search strategy
Combinations of the search terms “GIST or gastrointestinal stromal tumor” and “KIT or PDGFRA” and “SDH or succinate dehydrogenase” were used to generate a reference list of ~100 reports at the time of manuscript preparation. Abstracts from the search were individually examined for relevance and relevant studies were reviewed for the Discussion section.
Results
The patient’s primary tumor, an intermediate to high risk, ~10 cm gastric GIST, was resected at an outside institution. Per report, the tumor exhibited weak and focal positivity for CD117 (KIT), vimentin and calretinen, and was negative for SMA, S100, CD34, and desmin. A recurrent GIST was detected approximately 1 year later on surveillance CT, and IM therapy (400 mg daily) was initiated for a period of 3 months. Additional masses were detected in the upper abdomen and left lower quadrant, and progressive disease was confirmed by biopsy. Treatment with sunitinib was initiated (50 mg daily, 4 weeks on 2 weeks off) for 3 cycles. Restaging CT scan again showed disease progression, and sunitinib was discontinued. Subsequently the patient was referred to Fox Chase Cancer Center (FCCC), where a left lower quadrant abdominal wall metastasis was resected and banked by the FCCC Biosample repository (described herein as GIST 1). Direct Sanger sequence analysis of DNA from this tumor by the FCCC Clinical Molecular Genetics Laboratory detected an exon 18 D842V mutation in the PDGFRA gene. The patient was enrolled in a Phase II trial (NCT01243346) testing the RTK inhibitor crenolanib in GIST with IM-resistant PDGFRA mutations. The patient remained on crenolanib for 3 months until progression. The patient was off trial for a period of ~1.5 months, and again underwent surgery at FCCC for removal of metastatic small bowel and abdominal wall GISTs (GIST 2). Both GIST 1 and GIST 2 retained a high degree of cellularity despite the patient’s history of treatment with targeted therapeutics (Fig. 1a, d). The tumors displayed the epithelioid cell morphology seen in many PDGFRA-mutated GISTs, with focal areas displaying a partially developed organoid pattern of cells. Both recurrent tumors were essentially negative for KIT and DOG1 staining (Fig. 1b, e, c, and f). While DOG1 has been shown to be a somewhat more sensitive marker for GIST than KIT and is useful in evaluating KIT-negative tumors, one report has demonstrated that just over 1/3 of KIT-negative GIST stain positive for DOG1 [41], and evaluation of large GIST series for both markers have identified cases, including those harboring mutations in PDGFRA, that are both KIT- and DOG1-negative [42, 43].
Fig. 1.
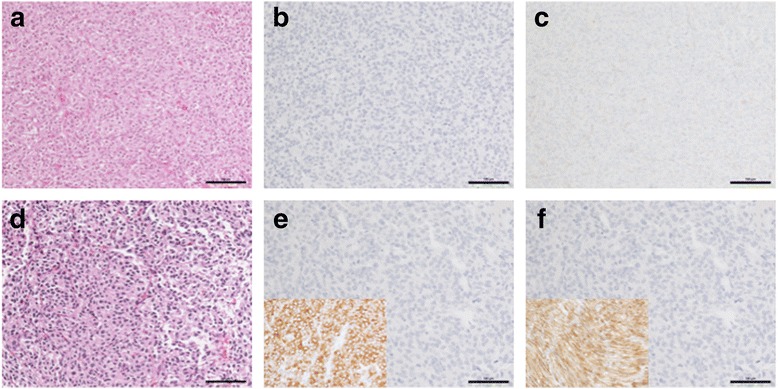
Immunohistochemical analysis of GIST specimens. Panels A, D: H&E for GIST 1 and GIST 2. Panels B, E: CD117 staining for GIST 1 and 2; insert: positive CD117 staining in a control GIST specimen. Panels C, F: DOG1 staining for GIST 1 and 2; insert: positive DOG1 staining in a control GIST specimen
Whole exome sequencing (WES) was used to investigate mechanisms of drug resistance in these GISTs. Ten deleterious missense or stop-gain somatic mutations (including the heterozygous PDGFRA D842V mutation) were found in both tumors, along with 3 mutations found only in GIST 2 (Fig. 2a, Table 1). A two-base frameshift deletion (c.291_292delCT, p.I97Mfs*21) in exon 4 of the SDHB gene was also identified in both GIST 1 and 2 (Fig. 2a). This mutation is predicted to result in a 21 amino acid residue frameshift beginning at residue 97 and ending with a stop codon. The SDHB mutant allele frequency from the WES analysis was 42 and 61% in GIST 1 and GIST 2, respectively. Sanger sequencing (Fig. 2b) confirmed the two-base deletion: the chromatograms indicate the wild type allele peaks are also present as a minority population. SNP array analysis (Cytoscan HD, Affymetrix) identified copy number loss across chromosome arm 1p, which encompasses the SDHB gene locus, in both tumors (Fig. 2c). This data suggests loss of the wild type SDHB allele and loss of SDH function in the tumors. To explore this further, tissue sections for the two tumors were evaluated for expression of SDHB. Although the WES analysis detected wild type reads in the area with the two base SDHB deletion, IHC for this subunit indicated that the tumor cells in both specimens were completely negative for SDHB staining (Fig. 3a, d), while endothelial cells surrounding tumor cells displayed strong punctate staining indicative of an intact SDH complex. IHC for the endothelial cell marker CD31 (Fig. 3b, e) indicated the tumors were highly vascularized, suggesting the wild type SDHB allele component in the WES and Sanger sequencing may derive from the substantial endothelial cell compartment in the tumors. Our interpretation of the molecular data and IHC analysis for this case is that both GISTs exhibit SDH deficiency in addition to harboring the gain of function PDGFRA mutation.
Fig. 2.
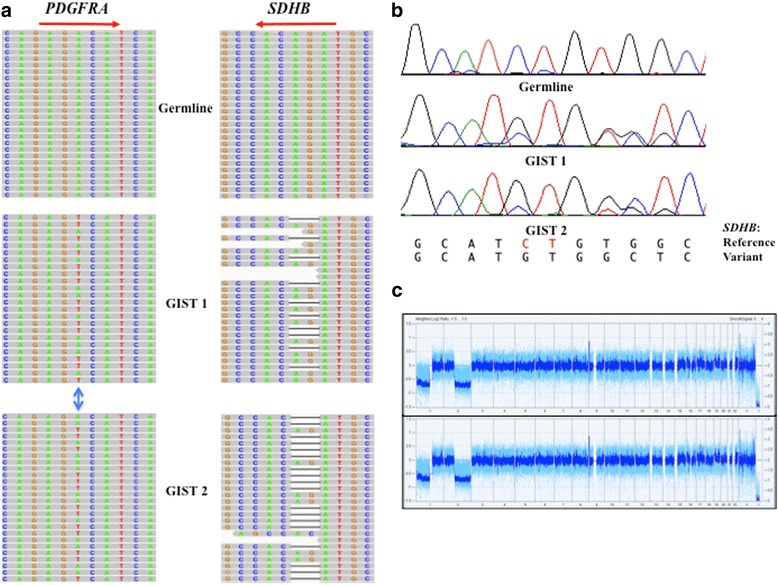
a A subset of reads from WES analysis visualized on the Integrative Genomics Viewer (IGV). Left panels show the heterozygous PDGFRA c.2525A > T mutation in the two tumors (indicated by double-headed blue arrow) that results in the p.D842V amino acid change. Right panels show the two base-pair deletion in exon 4 of the SDHB gene (represented as a bar) in reads from both tumors but not in the patient’s germline DNA. Red arrow indicates direction of transcription of PDGRA and SDHB. b Sanger sequencing confirming the frameshift SDHB deletion (c.291_292delCT, p.Iso97Metfs*21) in the patient’s GISTs. In the reference sequence the deleted bases are shown in red, and the variant sequence can be seen overlaying the reference sequence in the chromatograms from the tumors. c Whole genome view from the Chromosome Analysis Suite (Affymetrix) shows weighted SNP log-2 ratio (light blue) and smoothened signal (dark blue) for GIST 1 (top panel) and GIST 2 (bottom panel). Large-scale chromosome losses across chromosome arms 1p and 2q can be seen in both tumors
Fig. 3.
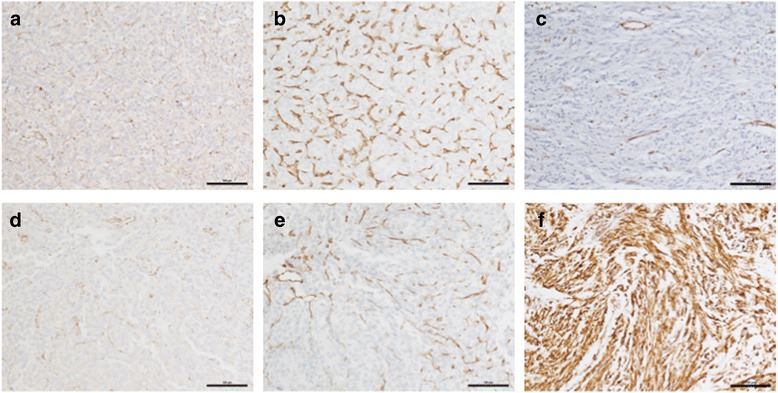
Panels a, d SDHB IHC for GIST 1 and GIST 2 shows distinct punctate staining in endothelial cells while tumor cells are negative for SDHB. Panels b, e IHC for endothelial marker CD31 for GIST 1 and GIST 2. Panel c Negative SDHB IHC for a control SDH-deficient RTK-wild type GIST. Panel f Positive SDHB staining for an SDH-competent KIT mutant GIST
As SDHX and KIT/PDGFRA mutations are generally considered to be mutually exclusive oncogenic events in GIST, we explored this phenomenon further by screening a panel of ~75 GIST cases with known genotype status from our institution. In all we analysed 19 PDGFRA mutated tumors (including the two from the index case and 10 additional D842V mutants), 50 KIT cases, and 5 previously described RTK-wild type SDH-deficient cases [26]. Several representative sections are shown in Fig. 3. While SDHB deficiency was clearly seen in RTK-wild type GIST cases (Fig. 3c shows one example), all PDGFRA- or KIT-mutated tumors were SDH-competant as indicated by punctate SDHB staining (Fig. 3f).
The WES analysis of GIST 1 and GIST 2 identified somatic mutations in 12 other genes in addition to the PDGFRA and SDHB mutations (Table 1). Although to our knowledge none of these genes have been implicated in GIST tumorigenesis, ceramide synthase 2 (CERS2) has recently been implicated in breast cancer (reviewed in [44]), while endothelin 3 (EDN3) has been shown to play a role in melanocyte differentiation, and altered endothelin signaling may be involved in melanomagenesis or progression [45].
Discussion
GIST oncogenesis in the great majority of cases is due to activation of kinase signaling due primarily to gain of function mutations in the KIT and PDGFRA receptors. This class of GIST, which has been referred to as “Type 1” GIST [46], can occur throughout the GI tract, and generally presents as sporadic disease in older adults. Tumor cells are generally spindled or mixed spindled/epithelioid, and these GISTs, with notable exceptions such as the PDGFRA D842V mutation discussed here, generally respond well to front-line therapy with the RTK inhibitor IM. SDH-deficient GIST, which have have also been termed Type II GIST, are an interesting sub-class of GIST that generally lack receptor mutations. SDH impairment leads to cytosolic accumulation of the TCA intermediate succinate and inhibition of alpha-ketoglutarate dependent dioxygenases [47]. These enzymes include the prolyl hydroxylase (PHD) that normally contributes to the degradation of the hypoxia-inducible gactor-1A (HIF1A) [48] as well as TET family DNA hydroxylases that are required for demethylation of genomic DNA [49]. Stabilization of HIF1A is thought to lead to induction of transcriptional programs that foster increased angiogenesis and cell proliferation [17], while TET inhibition leads to global DNA hypermethylation that is also likely involved in the oncogenic transformation to GIST [47]. Estimates of SDH deficiency as a percentage of RTK-wild type GIST range from 40 to 85% [2, 33, 50], and more or less distinct sub-groups can be defined based on the molecular and genetic aspects of the defect. Germline mutations in the SDHB-D gene subunits are the causative factor in the familial CSS [19], a relatively rare condition which predisposes carriers to the development of gastric GIST and PGL. Mutations in the SDHA subunit gene have recently been reported by a number of investigators in apparently sporadic adult cases [26–29, 31, 50, 51]. Finally, epigenetic silencing specific to the SDHC gene has been described in syndromic CT cases, a non-familial association of gastric GIST with PGL, pulmonary chondromas and other tumors, as well as in pediatric cases [23–25]. Regardless of the oncogenic insult, SDH-deficient GIST as a class exhibit clinical, pathological, and molecular features that distinguish them from SDH-competent, predominantly RTK-mutant GIST [46]. These tumors almost always present in the stomach, often as multifocal tumors with predominantly epithelioid cell morphology, and have a greater tendency to metastasize to lymph nodes and liver, although these metastases exhibit mostly indolent growth. Patients are generally younger and predominantly female: this is especially true of the SDHC epimutant patient class. In a recent analysis of RTK-wild type GIST from the NIH Pediatric and WT GIST Clinic, Boikos et al. reported a median age of 15 y and a female predominance of 95% for patients with epimutant SDHC GIST, while the SDHX mutant GIST patient demographic was slightly older with less of a female predominance (median age of 23, 62% female) [50]. SDH-deficient GIST also exhibit distinct gene expression patterns, including over-expression of the insulin-like growth factor 1 receptor (IGF1R) [26]. Importantly, due to their distinct oncogenic mechanism these tumors exhibit primary reistance to IM.
The ability to ascertain SDH status readily using IHC for the SDHB subunit has added support to the notion that SDH deficiency in GIST is generally mutually exclusive to the other known oncogenic mechanisms. The initial studies in GIST, focused on syndromic GIST, identified SDH deficiency in CSS-associated GIST as well as GISTs from CT patients, while RTK-mutant GIST or NF1-associated GIST used as controls were SDH-competent [52, 53]. Janeway et al. used a combination of SDHX sequencing and SDHB IHC and/or immunoblotting to determine the SDH status of a series of GIST, including 34 sporadic RTK-wild type tumors (pediatric and young adult), 18 KIT-mutated tumors, and 5 NF1-associated tumors [54]. Loss of function SDHB and SDHC gene mutations were identified in 3 young adult and one pediatric RTK-wild type cases, respectively, while an additional 18/18 pediatric cases and 8/12 adult RTK-wild type tumors were deemed SDH-deficient by loss of SDHB protein expression. Only 4 of the SDH-deficient cases were analyzed for SDHA mutation, possibly accounting for the low percentage of SDHX gene mutation in this group (13%). Notably, all 5 NF1-associated GIST and 17/18 of the KIT-mutated GIST analyzed were SDH-competent. Due to the study’s focus on RTK-wild type GIST, the manuscript contains no further details on the KIT mutant, SDH-deficient case. The Miettinen group examined a series of 756 gastric GIST by SDHB IHC and identified 66 cases of SDH-deficient GIST [32]. In contrast, the 378 non-gastric GISTs they analyzed were all found to be SDH-competent. The 66 gastric SDH-deficient GIST were all found to be wild type for KIT, PDGFRA, and BRAF. The authors failed to detect mutations in SDHX genes, although a limited number of exons in the SDHB-D genes were covered, and SDHA was not analyzed. In a study focusing on NF1-associated, RTK-wild type GIST, Wang et al. failed to detect SDH deficiency in all 22 tumors analyzed, confirming previous studies that looked at smaller NF1 patient sets [55]. Doyle et al. examined SDHB expression in a large cohort of GIST with known KIT/PDGFRA mutational status (n = 264) [33]. Among 53 RTK-wild type GIST they identified 22 SDH-deficient tumors (42%), of which all 21 with known anatomical location originated in the stomach. In this study all RTK-mutant GIST examined (179 KIT, 32 PDGFR) were immunohistochemically positive for SDHB expression.
These large series confirmed that SDH deficiency is found in a subset of RTK-wild type gastric GIST cases, whereas KIT/PDGFRA-mutated and NF1-related GISTs are generally SDH-competent. Against this background we searched the GIST literature for cases such as ours that documented SDH deficiency in RTK-mutant GIST. The search identified three case studies involving CSS patients with germline SDHX mutations and somatic KIT mutations in their GIST, as well as several apparently sporadic cases. Ayala-Ramirez et al. presented a case of a 33 year old male patient with bilateral pheochromocytoma (PCC), an extra-adrenal PGL and a gastric GIST [56]. Although there was no family history of PCC/PGL, genetic analysis identified a germline truncating mutation in the SDHD gene (p.W43*), supporting the diagnosis of CSS. Genotyping of the patient’s GIST revealed a known gain of function KIT mutation (p.D579del) [57]. However, SDHB IHC was not performed, and there was no SDHD genetic analysis described for the GIST, so it is difficult to say if the tumor was SDH-deficient. Gasparotto et al. described a 38 year old female with CSS with a high risk spindle cell rectal GIST and two cervical bilateral PGLs [58]. They identified an activating KIT mutation in the GIST (p.W557_V559delinsF) along with an SDHD frameshift mutation mutation (p.C150Yfs*42). However, the SDHD mutation was heterozygous and the authors report weak focal SDH staining in the GIST, while completely absent in the PGL. In addition, the rectal location and spindle cell morphology are not characteristic of SDH-deficient GIST. These two reports leave open the possibility that the cases described were sporadic KIT driven GISTs that occurred on a genetic background of heterozygous germline SDHD mutations. Jove et al. present a case report on a CSS patient that more robustly documents the co-occurrence of two oncogenic mechanisms in the same GIST [59]. The male patient was first treated surgically for a PGL at the age of 13; a heterozygous SDHB deletion (c.166_170del5, p.P56Yfs*5) was identified from peripheral blood [60]. Subsequently at age 29, the patient underwent resection of a multinodular gastric GIST that harbored a heterozygous KIT mutation (p.L576P), previously described as an activating mutation in GIST [61]. SDHB IHC for the GIST was completely negative, with positive staining in internal control cells. Adding strength to the analysis, sequencing of the patient’s tumor DNA demonstrated LOH of the wild type SDHB allele. Two relevant cases were identified in a larger study of 95 RTK-wild type GIST reported by Boikos et al. [50]. Among their subset of SDH-deficient, SDHX mutated GIST, the case referred to as GIST 077 harbored a germline stop-gain SDHA mutation (p.R512X) and somatic loss of the wild type allele, as well as a gain of function mutation in KIT (p.L576P). The second case, GIST 117, carried a germline SDHB splice site mutation (c.423 + 1G > A) and exhibited loss of the wild type allele in the tumor. This tumor also contained an activating KRAS alteration (p.G12D), a mutation that was also described in several KIT mutant GIST cases in a single report [16]. Finally, Ondrej and colleagues described a case of 52 year old male patient with a large mass in the posterior mediastinum, originally diagnosed as pleomorphic rhabdomyosarcoma, who subsequently underwent resection for multiple recurring nodules in the greater omentum [62]. Histological diagnosis of GIST was supported by strong CD117 staining and the identification of an activating KIT mutation (p.W557_K558del) in the tumor tissue. The authors also identified a missense variation in the SDHD gene (p.G12S), and IHC for the SDHB subunit showed faint and focal staining in the tumor tissue as compared to endothelial cells serving as internal controls. Although this GIST would be an unusual example of an extra-gastric SDH-deficient GIST also harboring an oncogenic KIT mutation, the G12S variant identified may represent an SDHD polymorphism as opposed to an inactivating mutation, as it was previously identified in 8/200 (4%) of healthy individuals [63].
Our literature search did not identify other examples of PDGFRA mutant GIST that also exhibited SDH deficiency. The tumors we examined in our case report were separate recurrences approximately 3 years after resection of the primary gastric tumor. Both tumors had a heterozygous PDGFRA D842V mutation (Fig. 2), which has been shown to confer clinical resistance to IM [7, 9] and sunitinib [64]: indeed the patient showed rapid disease progression on these two drugs (~3 months and 5 months, respectively). The patient’s GISTs also exhibited SDH deficiency due to a somatic frameshift mutation in the SDHB subunit gene and loss of the wild type allele (Figs. 2 and 3). Patients with RTK-wild type GIST (including SDH-deficient as well as SDH-competent GIST) have lower IM response rates compared to KIT exon 11 mutant GIST [9, 10], although sunitinib response rates have been higher for RTK-wild type GIST [64]. Theoretically, either mutation could be a driver in GIST, however the patient’s primary tumor was not available for analysis. GIST oncogenesesis by PDGFRA activation requires only a single hit, and the D842V mutation accounts for oncogenesis in about 1:20 GIST cases [2]. Oncogenesis through SDHX mutations occurs less frequently and requires inactivation of both alleles (one of which is usually due to a germline mutation), so the SDH inactivation may have been a later event. Interestingly, the patient also progressed rapidly (~3 months) on the investigational PDGFRA inhibitor crenolanib, which has been shown in biochemical and cellular models to be ~100–150-fold more potent than IM against the D842V mutation [65]. It is tempting to speculate that SDH deficiency in the tumor may have contributed to the lack of response to this selective PDGFRA inhibitor.
Although the identification of an activating mutation in either KIT or PDGFRA supports the clinical diagnosis of GIST and can be used to guide therapy, it does not preclude the possibility of the presence of SDHX gene mutations in these tumors, suggesting that the inclusion of these genes may be warranted in routine testing for GISTs. As detailed above, reports that clearly document GISTs with kinase and SDHX mutations are rare but not non-existent. Most documented cases are either CSS patients with both GIST and PGL and/or PCC tumors, or carry the germline SDHX mutations that predispose to the development of these tumors. Our case is unique in that both oncogenic mutations (PDGFRA and SDHB) were somatic and there was no germline involvement. Patients with germline SDHX mutations require long-term screening for disease detection and management [56] as well as genetic counseling and/or testing of family members. For GIST patients that carry germline SDHX mutations along with a KIT or PDGFRA mutation, standard IM therapy may still be effective. Anecdotally, in the CSS case described by Gasparotto et al. [58], IM treatment targeted to the exon 11 KIT mutation led to significant response and tumor stabilization for the patient. Further investigation is required to understand the biological and clinical consequences of SDH deficiency in GISTs with activating KIT or PDGFRA gene mutations. While molecularly targeted treatment options tailored to SDH deficiency in GIST are limited, a recently completed phase 2 clinical trial of linsitinib, an inhibitor of the IGF1 receptor that is over-expressed in RTK-wild type GIST, indicated that the drug provided clinical benefit in 45% of patients, albeit without RECIST response [66]. As mentioned above, sunitinib as well as regorafenib have had some success in these tumors [64, 67], possibly due to their ability to target the vascular endothelial growth factor receptor (VEGFR) and angiogenesis [68]. Finally, the global DNA hypermethylation seen in all SDH-deficient tumors, or the specific epigenetic silencing of SDHC seen in the CT/pediatric subset, suggests the potential of demethylating agents such as decitabine for treating these GISTs [24, 47].
Conclusions
We report a unique case of an SDH-deficient GIST case with an activating PDGFRA mutation. Oncogenic mutations in GIST are generally mutually exclusive; however documented exceptions exist which may have diagnostic and therapeutic implications.
Acknowledgements
We would like to acknowledge the following facilities at FCCC for work contributing to this manuscript: the Biosample Repository, Biostatistics and Bioinformatics Facility, the DNA Sequencing Facility, the Genomics Facility, the Clinical Pathology Laboratory, the Histopathology Facility, and the Molecular Diagnostics Laboratory.
Funding
This work was supported in part by a grant from the National Cancer Institute (NIH R21 CA150831–02, M.v.M. & M.G.B.). The authors would especially like to thank the GIST Cancer Research Fund and the Young Gastrointestinal Research Fund in Tribute to Frank L. and Lous H. Young for their continued support.
Availability of data and materials
All data generated or analysed during this study are included in this published article [and its supplementary information files].
Abbreviations
- CSS
Carney-Stratakis syndrome
- CT
Carney triad
- CT
Computed tomography
- GIST
Gastrointestinal stromal tumor
- HIF
Hypoxia inducible factor
- ICC
Interstitial cells of Cajal
- IGF1R
Insulin-like growth factor 1 receptor
- IHC
Immunohistochemistry
- IM
Imatinib mesylate
- NF1
Neurofibromatosis type I
- PC
Pulmonary chondroma
- PCC
Pheochromocytoma
- PDGFRA
platelet derived growth factor receptor alpha
- PGL
Paraganglioma
- PHD
Prolyl hydroxylase
- RTK
Receptor tyrosine kinase
- SDH
Succinate dehydrogenase
- SNP
Single nucleotide polymorphism
- TMA
Tissue microarray
- WES
Whole exome sequencing
Additional file
Primers for Sanger sequencing. (DOCX 14 kb)
Authors’ contributions
MB: contributed to design and execution of the study and drafted the manuscript; KC: performed immunohistochemical analysis; YZ: carried out the WES bioinformatics analysis; BL: performed whole exome sequencing; JP: carried out SNP array analysis; LR: contributed to execution of the study and involved in drafting the manuscript; MvM: contributed to design and execution of the study; involved in drafting the manuscript, gave approval of the final manuscript. All authors read and approved the final manuscript.
Ethics approval and consent to participate
The Fox Chase Biosample Repository (BSR) obtains and archives patient samples following informed written consent. Patient samples were obtained from the BSR under a protocol approved by the Fox Chase Cancer Center Institutional Review Board (#03–848).
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Footnotes
Electronic supplementary material
The online version of this article (doi:10.1186/s12885-017-3499-7) contains supplementary material, which is available to authorized users.
Contributor Information
Martin G. Belinsky, Phone: (215) 728-2756, Email: Martin.Belinsky@fccc.edu
Kathy Q. Cai, Email: Qi.Cai@fccc.edu
Yan Zhou, Email: Yan.Zhou@fccc.edu.
Biao Luo, Email: Biao.Luo@fccc.edu.
Jianming Pei, Email: Jianming.Pei@fccc.edu.
Lori Rink, Email: Lori.Rink@fccc.edu.
Margaret von Mehren, Email: Margaret.vonMehren@fccc.edu.
References
- 1.Miettinen M, Lasota J. Gastrointestinal stromal tumors. Gastroenterol Clin N Am. 2013;42(2):399–415. doi: 10.1016/j.gtc.2013.01.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Barnett CM, Corless CL, Heinrich MC. Gastrointestinal stromal tumors: molecular markers and genetic subtypes. Hematol Oncol Clin North Am. 2013;27(5):871–888. doi: 10.1016/j.hoc.2013.07.003. [DOI] [PubMed] [Google Scholar]
- 3.Kindblom LG, Remotti HE, Aldenborg F, Meis-Kindblom JM. Gastrointestinal pacemaker cell tumor (GIPACT): gastrointestinal stromal tumors show phenotypic characteristics of the interstitial cells of Cajal. Am J Pathol. 1998;152(5):1259–1269. [PMC free article] [PubMed] [Google Scholar]
- 4.Sircar K, Hewlett BR, Huizinga JD, Chorneyko K, Berezin I, Riddell RH. Interstitial cells of Cajal as precursors of gastrointestinal stromal tumors. Am J Surg Pathol. 1999;23(4):377–389. doi: 10.1097/00000478-199904000-00002. [DOI] [PubMed] [Google Scholar]
- 5.Heinrich MC, Corless CL, Duensing A, McGreevey L, Chen CJ, Joseph N, Singer S, Griffith DJ, Haley A, Town A, et al. PDGFRA activating mutations in gastrointestinal stromal tumors. Science. 2003;299(5607):708–710. doi: 10.1126/science.1079666. [DOI] [PubMed] [Google Scholar]
- 6.Hirota S, Isozaki K, Moriyama Y, Hashimoto K, Nishida T, Ishiguro S, Kawano K, Hanada M, Kurata A, Takeda M, et al. Gain-of-function mutations of c-kit in human gastrointestinal stromal tumors. Science. 1998;279(5350):577–580. doi: 10.1126/science.279.5350.577. [DOI] [PubMed] [Google Scholar]
- 7.Heinrich MC, Corless CL, Demetri GD, Blanke CD, von Mehren M, Joensuu H, McGreevey LS, Chen CJ, Van den Abbeele AD, Druker BJ, et al. Kinase mutations and imatinib response in patients with metastatic gastrointestinal stromal tumor. J Clin Oncol. 2003;21(23):4342–4349. doi: 10.1200/JCO.2003.04.190. [DOI] [PubMed] [Google Scholar]
- 8.Cassier PA, Fumagalli E, Rutkowski P, Schoffski P, Van Glabbeke M, Debiec-Rychter M, Emile JF, Duffaud F, Martin-Broto J, Landi B, et al. Outcome of patients with platelet-derived growth factor receptor alpha-mutated gastrointestinal stromal tumors in the tyrosine kinase inhibitor era. Clin Cancer Res. 2012;18(16):4458–4464. doi: 10.1158/1078-0432.CCR-11-3025. [DOI] [PubMed] [Google Scholar]
- 9.Debiec-Rychter M, Sciot R, Le Cesne A, Schlemmer M, Hohenberger P, van Oosterom AT, Blay JY, Leyvraz S, Stul M, Casali PG, et al. KIT mutations and dose selection for imatinib in patients with advanced gastrointestinal stromal tumours. Eur J Cancer. 2006;42(8):1093–1103. doi: 10.1016/j.ejca.2006.01.030. [DOI] [PubMed] [Google Scholar]
- 10.Heinrich MC, Owzar K, Corless CL, Hollis D, Borden EC, Fletcher CD, Ryan CW, von Mehren M, Blanke CD, Rankin C, et al. Correlation of kinase genotype and clinical outcome in the north American intergroup phase III trial of imatinib mesylate for treatment of advanced gastrointestinal stromal tumor: CALGB 150105 study by cancer and leukemia group B and southwest oncology group. J Clin Oncol. 2008;26(33):5360–5367. doi: 10.1200/JCO.2008.17.4284. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Miettinen M, Fetsch JF, Sobin LH, Lasota J. Gastrointestinal stromal tumors in patients with neurofibromatosis 1: a clinicopathologic and molecular genetic study of 45 cases. Am J Surg Pathol. 2006;30(1):90–96. doi: 10.1097/01.pas.0000176433.81079.bd. [DOI] [PubMed] [Google Scholar]
- 12.Belinsky MG, Rink L, Cai KQ, Capuzzi SJ, Hoang Y, Chien J, Godwin AK, von Mehren M. Somatic loss of function mutations in neurofibromin 1 and MYC associated factor X genes identified by exome-wide sequencing in a wild-type GIST case. BMC Cancer. 2015;15:887. doi: 10.1186/s12885-015-1872-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Gasparotto D, Rossi S, Polano M, Tamborini E, Lorenzetto E, Sbaraglia M, Mondello A, Massani M, Lamon S, Bracci R, et al. Quadruple-negative GIST is a sentinel for unrecognized Neurofibromatosis type 1 syndrome. Clin Cancer Res. 2017;23(1):273–82. [DOI] [PubMed]
- 14.Agaram NP, Wong GC, Guo T, Maki RG, Singer S, Dematteo RP, Besmer P, Antonescu CR. Novel V600E BRAF mutations in imatinib-naive and imatinib-resistant gastrointestinal stromal tumors. Genes Chromosomes Cancer. 2008;47(10):853–859. doi: 10.1002/gcc.20589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Agaimy A, Terracciano LM, Dirnhofer S, Tornillo L, Foerster A, Hartmann A, Bihl MP. V600E BRAF mutations are alternative early molecular events in a subset of KIT/PDGFRA wild-type gastrointestinal stromal tumours. J Clin Pathol. 2009;62(7):613–616. doi: 10.1136/jcp.2009.064550. [DOI] [PubMed] [Google Scholar]
- 16.Miranda C, Nucifora M, Molinari F, Conca E, Anania MC, Bordoni A, Saletti P, Mazzucchelli L, Pilotti S, Pierotti MA, et al. KRAS and BRAF mutations predict primary resistance to imatinib in gastrointestinal stromal tumors. Clin Cancer Res. 2012;18(6):1769–1776. doi: 10.1158/1078-0432.CCR-11-2230. [DOI] [PubMed] [Google Scholar]
- 17.Belinsky MG, Rink L, von Mehren M. Succinate dehydrogenase deficiency in pediatric and adult gastrointestinal stromal tumors. Front Oncol. 2013;3:117. doi: 10.3389/fonc.2013.00117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Carney JA, Stratakis CA. Familial paraganglioma and gastric stromal sarcoma: a new syndrome distinct from the Carney triad. Am J Med Genet. 2002;108(2):132–139. doi: 10.1002/ajmg.10235. [DOI] [PubMed] [Google Scholar]
- 19.McWhinney SR, Pasini B, Stratakis CA. Familial gastrointestinal stromal tumors and germ-line mutations. N Engl J Med. 2007;357(10):1054–1056. doi: 10.1056/NEJMc071191. [DOI] [PubMed] [Google Scholar]
- 20.Pasini B, McWhinney SR, Bei T, Matyakhina L, Stergiopoulos S, Muchow M, Boikos SA, Ferrando B, Pacak K, Assie G, et al. Clinical and molecular genetics of patients with the Carney-Stratakis syndrome and germline mutations of the genes coding for the succinate dehydrogenase subunits SDHB, SDHC, and SDHD. Eur J Hum Genet. 2008;16(1):79–88. doi: 10.1038/sj.ejhg.5201904. [DOI] [PubMed] [Google Scholar]
- 21.Carney JA, Sheps SG, Go VL, Gordon H. The triad of gastric leiomyosarcoma, functioning extra-adrenal paraganglioma and pulmonary chondroma. N Engl J Med. 1977;296(26):1517–1518. doi: 10.1056/NEJM197706302962609. [DOI] [PubMed] [Google Scholar]
- 22.Matyakhina L, Bei TA, McWhinney SR, Pasini B, Cameron S, Gunawan B, Stergiopoulos SG, Boikos S, Muchow M, Dutra A, et al. Genetics of carney triad: recurrent losses at chromosome 1 but lack of germline mutations in genes associated with paragangliomas and gastrointestinal stromal tumors. Journal of Clinical Endocrinology & Metabolism. 2007;92(8):2938–2943. doi: 10.1210/jc.2007-0797. [DOI] [PubMed] [Google Scholar]
- 23.Janeway KA, Kim SY, Lodish M, Nose V, Rustin P, Gaal J, Dahia PL, Liegl B, Ball ER, Raygada M, et al. Defects in succinate dehydrogenase in gastrointestinal stromal tumors lacking KIT and PDGFRA mutations. Proc Natl Acad Sci U S A. 2011;108(1):314–318. doi: 10.1073/pnas.1009199108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Killian JK, Miettinen M, Walker RL, Wang Y, Zhu YJ, Waterfall JJ, Noyes N, Retnakumar P, Yang Z, Smith WI, Jr, et al. Recurrent epimutation of SDHC in gastrointestinal stromal tumors. Sci Transl Med. 2014;6(268):268ra177. doi: 10.1126/scitranslmed.3009961. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Haller F, Moskalev EA, Faucz FR, Barthelmess S, Wiemann S, Bieg M, Assie G, Bertherat J, Schaefer IM, Otto C, et al. Aberrant DNA hypermethylation of SDHC: a novel mechanism of tumor development in Carney triad. Endocr Relat Cancer. 2014;21(4):567–577. doi: 10.1530/ERC-14-0254. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Belinsky MG, Rink L, Flieder DB, Jahromi MS, Schiffman JD, Godwin AK, Mehren MV. Overexpression of insulin-like growth factor 1 receptor and frequent mutational inactivation of SDHA in wild-type SDHB-negative gastrointestinal stromal tumors. Genes Chromosomes Cancer. 2013;52(2):214–24. [DOI] [PMC free article] [PubMed]
- 27.Pantaleo MA, Astolfi A, Indio V, Moore R, Thiessen N, Heinrich MC, Gnocchi C, Santini D, Catena F, Formica S, et al. SDHA loss-of-function mutations in KIT-PDGFRA wild-type gastrointestinal stromal tumors identified by massively parallel sequencing. J Natl Cancer Inst. 2011;103(12):983–987. doi: 10.1093/jnci/djr130. [DOI] [PubMed] [Google Scholar]
- 28.Italiano A, Chen CL, Sung YS, Singer S, Dematteo RP, Laquaglia MP, Besmer P, Socci N, Antonescu CR. SDHA loss of function mutations in a subset of young adult wild-type gastrointestinal stromal tumors. BMC Cancer. 2012;12:408. doi: 10.1186/1471-2407-12-408. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Oudijk L, Gaal J, Korpershoek E, van Nederveen FH, Kelly L, Schiavon G, Verweij J, Mathijssen RH, den Bakker MA, Oldenburg RA, et al. SDHA mutations in adult and pediatric wild-type gastrointestinal stromal tumors. Mod Pathol. 2013;26(3):456–63. [DOI] [PubMed]
- 30.Miettinen M, Killian JK, Wang ZF, Lasota J, Lau C, Jones L, Walker R, Pineda M, Zhu YJ, Kim SY, et al. Immunohistochemical loss of Succinate Dehydrogenase subunit a (SDHA) in gastrointestinal Stromal tumors (GISTs) signals SDHA Germline mutation. Am J Surg Pathol. 2013;37(2):234–40. [DOI] [PMC free article] [PubMed]
- 31.Wagner AJ, Remillard SP, Zhang YX, Doyle LA, George S, Hornick JL. Loss of expression of SDHA predicts SDHA mutations in gastrointestinal stromal tumors. Mod Pathol. 2013;26(2):289–94. [DOI] [PubMed]
- 32.Miettinen M, Wang ZF, Sarlomo-Rikala M, Osuch C, Rutkowski P, Lasota J. Succinate dehydrogenase-deficient GISTs: a clinicopathologic, immunohistochemical, and molecular genetic study of 66 gastric GISTs with predilection to young age. Am J Surg Pathol. 2011;35(11):1712–1721. doi: 10.1097/PAS.0b013e3182260752. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Doyle LA, Nelson D, Heinrich MC, Corless CL, Hornick JL. Loss of succinate dehydrogenase subunit B (SDHB) expression is limited to a distinctive subset of gastric wild-type gastrointestinal stromal tumours: a comprehensive genotype-phenotype correlation study. Histopathology. 2012;61(5):801–809. doi: 10.1111/j.1365-2559.2012.04300.x. [DOI] [PubMed] [Google Scholar]
- 34.Li H, Durbin R. Fast and accurate short read alignment with burrows-wheeler transform. Bioinformatics. 2009;25(14):1754–1760. doi: 10.1093/bioinformatics/btp324. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.McKenna A, Hanna M, Banks E, Sivachenko A, Cibulskis K, Kernytsky A, Garimella K, Altshuler D, Gabriel S, Daly M, et al. The genome analysis toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome Res. 2010;20(9):1297–1303. doi: 10.1101/gr.107524.110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.DePristo MA, Banks E, Poplin R, Garimella KV, Maguire JR, Hartl C, Philippakis AA, del Angel G, Rivas MA, Hanna M, et al. A framework for variation discovery and genotyping using next-generation DNA sequencing data. Nat Genet. 2011;43(5):491–498. doi: 10.1038/ng.806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Wang K, Li M, Hakonarson H. ANNOVAR: functional annotation of genetic variants from high-throughput sequencing data. Nucleic Acids Res. 2010;38(16):e164. doi: 10.1093/nar/gkq603. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Robinson JT, Thorvaldsdottir H, Winckler W, Guttman M, Lander ES, Getz G, Mesirov JP. Integrative genomics viewer. Nat Biotechnol. 2011;29(1):24–26. doi: 10.1038/nbt.1754. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Tarn C, Rink L, Merkel E, Flieder D, Pathak H, Koumbi D, Testa JR, Eisenberg B, von Mehren M, Godwin AK. Insulin-like growth factor 1 receptor is a potential therapeutic target for gastrointestinal stromal tumors. Proc Natl Acad Sci U S A. 2008;105(24):8387–8392. doi: 10.1073/pnas.0803383105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Gill AJ, Benn DE, Chou A, Clarkson A, Muljono A, Meyer-Rochow GY, Richardson AL, Sidhu SB, Robinson BG, Clifton-Bligh RJ. Immunohistochemistry for SDHB triages genetic testing of SDHB, SDHC, and SDHD in paraganglioma-pheochromocytoma syndromes. Hum Pathol. 2010;41(6):805–814. doi: 10.1016/j.humpath.2009.12.005. [DOI] [PubMed] [Google Scholar]
- 41.Liegl B, Hornick JL, Corless CL, Fletcher CD. Monoclonal antibody DOG1.1 shows higher sensitivity than KIT in the diagnosis of gastrointestinal stromal tumors, including unusual subtypes. Am J Surg Pathol. 2009;33(3):437–446. doi: 10.1097/PAS.0b013e318186b158. [DOI] [PubMed] [Google Scholar]
- 42.Espinosa I, Lee CH, Kim MK, Rouse BT, Subramanian S, Montgomery K, Varma S, Corless CL, Heinrich MC, Smith KS, et al. A novel monoclonal antibody against DOG1 is a sensitive and specific marker for gastrointestinal stromal tumors. Am J Surg Pathol. 2008;32(2):210–218. doi: 10.1097/PAS.0b013e3181238cec. [DOI] [PubMed] [Google Scholar]
- 43.Miettinen M, Wang ZF, Lasota J. DOG1 antibody in the differential diagnosis of gastrointestinal stromal tumors: a study of 1840 cases. Am J Surg Pathol. 2009;33(9):1401–1408. doi: 10.1097/PAS.0b013e3181a90e1a. [DOI] [PubMed] [Google Scholar]
- 44.Levy M, Futerman AH. Mammalian ceramide synthases. IUBMB Life. 2010;62(5):347–356. doi: 10.1002/iub.319. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Saldana-Caboverde A, Kos L. Roles of endothelin signaling in melanocyte development and melanoma. Pigment Cell Melanoma Res. 2010;23(2):160–170. doi: 10.1111/j.1755-148X.2010.00678.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Gill AJ, Chou A, Vilain RE, Clifton-Bligh RJ. "pediatric-type" gastrointestinal stromal tumors are SDHB negative ("type 2") GISTs. Am J Surg Pathol. 2011;35(8):1245–1247. doi: 10.1097/PAS.0b013e3182217b93. [DOI] [PubMed] [Google Scholar]
- 47.Killian JK, Kim SY, Miettinen M, Smith C, Merino M, Tsokos M, Quezado M, Smith WI, Jr, Jahromi MS, Xekouki P, et al. Succinate dehydrogenase mutation underlies global epigenomic divergence in gastrointestinal stromal tumor. Cancer Discov. 2013;3(6):648–657. doi: 10.1158/2159-8290.CD-13-0092. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Selak MA, Armour SM, MacKenzie ED, Boulahbel H, Watson DG, Mansfield KD, Pan Y, Simon MC, Thompson CB, Gottlieb E. Succinate links TCA cycle dysfunction to oncogenesis by inhibiting HIF-alpha prolyl hydroxylase. Cancer Cell. 2005;7(1):77–85. doi: 10.1016/j.ccr.2004.11.022. [DOI] [PubMed] [Google Scholar]
- 49.Mason EF, Hornick JL. Succinate dehydrogenase deficiency is associated with decreased 5-hydroxymethylcytosine production in gastrointestinal stromal tumors: implications for mechanisms of tumorigenesis. Mod Pathol. 2013;26(11):1492–1497. doi: 10.1038/modpathol.2013.86. [DOI] [PubMed] [Google Scholar]
- 50.Boikos SA, Pappo AS, Killian JK, LaQuaglia MP, Weldon CB, George S, Trent JC, von Mehren M, Wright JA, Schiffman JD, et al. Molecular subtypes of KIT/PDGFRA wild-type gastrointestinal Stromal tumors: a report from the National Institutes of Health gastrointestinal Stromal tumor clinic. JAMA Oncol. 2016;2(7):922–928. doi: 10.1001/jamaoncol.2016.0256. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Dwight T, Benn DE, Clarkson A, Vilain R, Lipton L, Robinson BG, Clifton-Bligh RJ, Gill AJ. Loss of SDHA expression identifies SDHA mutations in Succinate Dehydrogenase-deficient gastrointestinal Stromal tumors. Am J Surg Pathol. 2013;37(2):226–33. [DOI] [PubMed]
- 52.Gill AJ, Chou A, Vilain R, Clarkson A, Lui M, Jin R, Tobias V, Samra J, Goldstein D, Smith C, et al. Immunohistochemistry for SDHB divides gastrointestinal stromal tumors (GISTs) into 2 distinct types. Am J Surg Pathol. 2010;34(5):636–644. doi: 10.1097/PAS.0b013e3181d6150d. [DOI] [PubMed] [Google Scholar]
- 53.Gaal J, Stratakis CA, Carney JA, Ball ER, Korpershoek E, Lodish MB, Levy I, Xekouki P, van Nederveen FH, den Bakker MA, et al. SDHB immunohistochemistry: a useful tool in the diagnosis of Carney-Stratakis and Carney triad gastrointestinal stromal tumors. Mod Pathol. 2011;24(1):147–151. doi: 10.1038/modpathol.2010.185. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Janeway KA, Liegl B, Harlow A, Le C, Perez-Atayde A, Kozakewich H, Corless CL, Heinrich MC, Fletcher JA. Pediatric KIT wild-type and platelet-derived growth factor receptor alpha-wild-type gastrointestinal stromal tumors share KIT activation but not mechanisms of genetic progression with adult gastrointestinal stromal tumors. Cancer Res. 2007;67(19):9084–9088. doi: 10.1158/0008-5472.CAN-07-1938. [DOI] [PubMed] [Google Scholar]
- 55.Wang JH, Lasota J, Miettinen M. Succinate Dehydrogenase subunit B (SDHB) is expressed in Neurofibromatosis 1-associated gastrointestinal Stromal tumors (Gists): implications for the SDHB expression based classification of Gists. J Cancer. 2011;2:90–93. doi: 10.7150/jca.2.90. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Ayala-Ramirez M, Callender GG, Kupferman ME, Rich TA, Chuang HH, Trent J, Perrier ND, Goodarzi M, Jimenez C. Paraganglioma syndrome type 1 in a patient with Carney-Stratakis syndrome. Nat Rev Endocrinol. 2010;6(2):110–115. doi: 10.1038/nrendo.2009.250. [DOI] [PubMed] [Google Scholar]
- 57.Nakahara M, Isozaki K, Hirota S, Miyagawa J, Hase-Sawada N, Taniguchi M, Nishida T, Kanayama S, Kitamura Y, Shinomura Y, et al. A novel gain-of-function mutation of c-kit gene in gastrointestinal stromal tumors. Gastroenterology. 1998;115(5):1090–1095. doi: 10.1016/S0016-5085(98)70079-4. [DOI] [PubMed] [Google Scholar]
- 58.Gasparotto D, Rossi S, Campagna D, Scavina P, Tiziano FD, Marzotto A, Toffolatti L, Vitelli CE, Amini M, Dei Tos AP, et al. Imatinib-sensitizing KIT mutation in a Carney-Stratakis-associated GI Stromal tumor. J Clin Oncol. 2016;34(11):e99–e103. doi: 10.1200/JCO.2012.44.7300. [DOI] [PubMed] [Google Scholar]
- 59.Jove M, Mora J, Sanjuan X, Rodriguez E, Robledo M, Farran L. Garcia del Muro X: simultaneous KIT mutation and succinate dehydrogenase (SDH) deficiency in a patient with a gastrointestinal stromal tumour and Carney-Stratakis syndrome: a case report. Histopathology. 2014;65(5):712–717. doi: 10.1111/his.12506. [DOI] [PubMed] [Google Scholar]
- 60.Mora J, Cascon A, Robledo M, Catala A. Pediatric paraganglioma: an early manifestation of an adult disease secondary to germline mutations. Pediatr Blood Cancer. 2006;47(6):785–789. doi: 10.1002/pbc.20680. [DOI] [PubMed] [Google Scholar]
- 61.Lasota J, Miettinen M. KIT and PDGFRA mutations in gastrointestinal stromal tumors (GISTs) Semin Diagn Pathol. 2006;23(2):91–102. doi: 10.1053/j.semdp.2006.08.006. [DOI] [PubMed] [Google Scholar]
- 62.Daum O, Sedivcova M, Dubova M, Michal M. KIT mutations and sequence changes in genes encoding SDH complex possibly need not be mutually exclusive in gastrointestinal stromal tumors. Appl Immunohistochem Mol Morphol. 2012;20(5):523–524. doi: 10.1097/PAI.0b013e3182494026. [DOI] [PubMed] [Google Scholar]
- 63.Cascon A, Ruiz-Llorente S, Cebrian A, Leton R, Telleria D, Benitez J, Robledo M. G12S and H50R variations are polymorphisms in the SDHD gene. Genes Chromosomes Cancer. 2003;37(2):220–221. doi: 10.1002/gcc.10212. [DOI] [PubMed] [Google Scholar]
- 64.Heinrich MC, Maki RG, Corless CL, Antonescu CR, Harlow A, Griffith D, Town A, McKinley A, Ou WB, Fletcher JA, et al. Primary and secondary kinase genotypes correlate with the biological and clinical activity of sunitinib in imatinib-resistant gastrointestinal stromal tumor. J Clin Oncol. 2008;26(33):5352–5359. doi: 10.1200/JCO.2007.15.7461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Heinrich MC, Griffith D, McKinley A, Patterson J, Presnell A, Ramachandran A, Debiec-Rychter M. Crenolanib inhibits the drug-resistant PDGFRA D842V mutation associated with imatinib-resistant gastrointestinal stromal tumors. Clin Cancer Res. 2012;18(16):4375–4384. doi: 10.1158/1078-0432.CCR-12-0625. [DOI] [PubMed] [Google Scholar]
- 66.Huss S, Elges S, Trautmann M, Sperveslage J, Hartmann W, Wardelmann E. Classification of KIT/PDGFRA wild-type gastrointestinal stromal tumors: implications for therapy. Expert Rev Anticancer Ther. 2015;15(6):623–628. doi: 10.1586/14737140.2015.1032941. [DOI] [PubMed] [Google Scholar]
- 67.Ben-Ami E, Barysauskas CM, von Mehren M, Heinrich MC, Corless CL, Butrynski JE, Morgan JA, Wagner AJ, Choy E, Yap JT, et al. Long-term follow-up results of the multicenter phase II trial of regorafenib in patients with metastatic and/or unresectable GI stromal tumor after failure of standard tyrosine kinase inhibitor therapy. Ann Oncol. 2016;27(9):1794–1799. doi: 10.1093/annonc/mdw228. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Songdej N, von Mehren M. GIST treatment options after tyrosine kinase inhibitors. Curr Treat Options in Oncol. 2014;15(3):493–506. doi: 10.1007/s11864-014-0295-3. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
All data generated or analysed during this study are included in this published article [and its supplementary information files].