

Abstract
A key function of the proximal tubule is retrieval of most of the vast quantities of NaCl and water filtered by the kidney. Physiological studies using brush border vesicles and perfused tubules have indicated that a major fraction of Cl− reabsorption across the apical membrane of proximal tubule cells occurs via Cl−-formate exchange. The molecular identity of the transporter responsible for renal brush border Cl−-formate exchange has yet to be elucidated. As a strategy to identify one or more anion exchangers responsible for mediating Cl− reabsorption in the proximal tubule, we screened the expressed sequence tag database for homologs of pendrin, a transporter previously shown to mediate Cl−-formate exchange. We now report the cDNA cloning of CFEX, a mouse pendrin homolog with expression in the kidney by Northern analysis. Sequence analysis indicated that CFEX very likely represents the mouse ortholog of human SLC26A6. Immunolocalization studies detected expression of CFEX, but not pendrin, on the brush border membrane of proximal tubule cells. Functional expression studies in Xenopus oocytes demonstrated that CFEX mediates Cl−-formate exchange. Taken together, these observations identify CFEX as a prime candidate to mediate Cl−-formate exchange in the proximal tubule and thereby to contribute importantly to renal NaCl reabsorption. Given its wide tissue distribution, CFEX also may contribute to transcellular Cl− transport in additional epithelia such as the pancreas and contribute to transmembrane Cl− transport in nonepithelial tissues such as the heart.
The majority of NaCl and water filtered by the kidney is reabsorbed in the proximal tubule. Although passive paracellular transport contributes to NaCl reabsorption in this nephron segment, an important component of NaCl reabsorption in the proximal tubule is active and transcellular (1). Studies using isolated brush border vesicles and perfused tubules are consistent with the concept that a major fraction of NaCl entry across the apical membrane of proximal tubule cells occurs via Cl−-formate exchange in parallel with Na+-H+ exchange and H+-coupled recycling of formate (2–7). Whereas several lines of evidence indicate that the transporter primarily responsible for apical Na+-H+ exchange in the proximal tubule is NHE3 (8–13), the transporter responsible for apical Cl−-formate exchange has yet to be identified.
Pendrin is a monovalent anion exchanger that has been found capable of mediating Cl−-formate and Cl−-OH−/HCO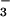 exchange (14–16). Although mRNA encoding pendrin was detected in rat proximal tubules by reverse transcriptase–PCR (15), it has been immunolocalized to the apical membrane of cells in the cortical collecting tubule rather than the proximal tubule (17). Studies in knockout mice indicate pendrin plays an essential role in base secretion by the collecting tubule (17).
exchange (14–16). Although mRNA encoding pendrin was detected in rat proximal tubules by reverse transcriptase–PCR (15), it has been immunolocalized to the apical membrane of cells in the cortical collecting tubule rather than the proximal tubule (17). Studies in knockout mice indicate pendrin plays an essential role in base secretion by the collecting tubule (17).
As a strategy to identify one or more anion exchangers responsible for mediating Cl− reabsorption in the proximal tubule, we screened the expressed sequence tag (EST) database for homologs of pendrin that are expressed in the mouse kidney. We now report the cDNA cloning of CFEX, a pendrin homolog that mediates Cl−-formate exchange when expressed in Xenopus oocytes. Immunocytochemical analysis using anti-CFEX antibodies indicates that this transporter is expressed on the brush border membrane of proximal tubule cells. Hence, CFEX is a candidate to mediate Cl−-formate exchange in the proximal tubule and thereby to contribute importantly to renal NaCl reabsorption.
Materials and Methods
cDNA Cloning of CFEX.
The reported amino acid sequence for human pendrin was used to perform a tblastn search of the GenBank EST database. Four unique mouse EST clones were identified that had neither overlap with each other nor close identity to cDNAs encoding known proteins. Primers were synthesized based on the reported sequences and used to screen a mouse renal cDNA library (CLONTECH) by PCR to identify those that were expressed in kidney. Products of the expected size were obtained by using primers based on two of the EST sequences (GenBank accession nos. AA871419 and AI747461). These two EST clones then were obtained (Research Genetics, Huntsville, AL) and sequenced. The 0.7 kb of AA747461 sequence was found to be entirely contained within the 1.2-kb sequence of AA871419. Analysis of the sequence of AA871419 indicated the presence of an ORF encoding the C terminus of a predicted protein with ≈40% identity to pendrin. 5′ Rapid amplification of cDNA ends then was performed by using Marathon-Ready mouse kidney cDNA (CLONTECH) as template for nested PCR. A 1.8-kb PCR product was successfully amplified and subcloned into pCR2.1-TOPO (Invitrogen). Sequencing indicated that this PCR product contained 144 bp of 5′ untranslated region and the first ATG of a single ORF that was continuous with the reading frame in AA871419. The presence of upstream stop codons in the same reading frame confirmed that the start codon had been identified.
For expression studies, the coding sequence was amplified by PCR using primers designed to incorporate upstream and downstream EcoRI and XbaI sites, respectively (5′ primer: CCAAGAATTCACCATGGGGCTGCAGAGGAGAGACTA; 3′ primer:CCGGTCTAGATCAGAGTTTGGTGGCCAAAACA). This PCR product was subcloned into pCR2.1-TOPO and fully sequenced. The EcoRI and XbaI sites then were used to excise and ligate the CFEX coding region into either the Xenopus expression plasmid pGH19 between the 5′ and 3′ untranslated regions of the Xenopus β-globin gene (18) or the mammalian expression vector pcDNA3.1(+) (Invitrogen).
Northern Analysis.
A mouse multiple tissue Northern blot (CLONTECH) was probed with a 32P-labeled PCR-generated cDNA clone spanning nucleotides 1556–1992 of the CFEX coding sequence. This region has low sequence similarity to pendrin and was specifically chosen to minimize the potential for cross-hybridization to the pendrin transcript. The Northern blot was prehybridized in Church-Gilbert solution containing 50% formamide and 100 μg of denatured salmon sperm DNA/ml for 24 h at 42°C. The blot was hybridized for 24 h at 42°C in the same solution containing 106 cpm/ml of either the 32P random prime-labeled CFEX probe or a similarly labeled β-actin probe as a RNA loading control. After hybridization, the blot was washed in 2× SSC containing 0.5% SDS for 20 min at room temperature and then 1× SSC containing 0.5% SDS for 20 min at 55°C.
Preparation of Antibodies.
Peptides corresponding to the C-terminal 29 aa of rat pendrin (CKDPLDLMEAEMNAEELDVQDEAMRRLAS) and mouse CFEX (CVFASVHDAVTFALSHRKSVPKSPVLATKL), the latter including an additional N-terminal cysteine, were synthesized by the Howard Hughes Medical Institute Biopolymer Laboratory and W. M. Keck Foundation Biotechnology Resource Laboratory at Yale University. Each peptide was coupled to keyhole limpet hemocyanin with the Imject Maleimide Activated Immunogen Conjugation Kit (Pierce) and used to immunize rabbits to generate polyclonal antisera (Pocono Rabbit Farm, Canadensis, PA). Antisera were purified with affinity columns prepared by immobilizing each peptide with SulfoLink Coupling Gel (Pierce).
Transient Expression of CFEX in COS-7 Cells.
To verify antibody specificity, immunoblotting was performed by using COS-7 cells transfected with CFEX cDNA that had been subcloned in pcDNA3.1. COS-7 cells were grown in DMEM with 10% FCS, 50 units/ml penicillin, and 50 mg/ml streptomycin at 37°C in 5% CO2-95% air. Cells were plated on 6-well tissue culture dishes for 24 h and used at 80–90% confluence. Cells were transfected with 2 μg plasmid and 10 μl Superfect Reagent (Qiagen, Valencia, CA) according to the manufacturer's protocol. After transfection, cells were incubated for 72 h and then solubilized in SDS sample buffer for immunoblotting experiments.
SDS/PAGE and Immunoblotting.
COS-7 cells or renal microsomes isolated from mouse kidney cortex (19) were solubilized in sample buffer, and proteins were separated by SDS/PAGE using 7.5% polyacrylamide gels according to Laemmli (20). For immunoblotting, proteins were transferred to poly(vinylidene difluoride) (PVDF, Immobilon-P, Millipore) with a Transphor transfer electrophoresis unit (Hoefer) and stained with Ponceau S in 0.5% trichloroacetic acid. For immunoblotting studies, PVDF strips were incubated first in Blotto (5% nonfat dry milk and 0.1% Tween 20 in PBS, pH 7.4) for 1 h to block nonspecific binding, followed by overnight incubation in primary antibody diluted 1:100 in Blotto. The strips then were washed in Blotto and incubated for 1 h with 1:2,000 dilution horseradish peroxidase-conjugated secondary antibody (goat anti-rabbit IgG, Zymed). Bound antibody was detected with the enhanced chemiluminescence system (Amersham Pharmacia) according to the manufacturer's protocol.
Tissue Preparation and Immunofluorescence Staining.
Adult female C57BL/6J mice were anesthetized by i.p. injection of sodium pentobarbital. Animals were perfusion-fixed with PLP (2% paraformaldehyde, 750 mM lysine, and 10 mM sodium periodate in phosphate buffer, pH 7.4, 22°C) as described by Larsson (21). The kidneys were removed, cut into 2–4 mm blocks, and postfixed in the same fixative for an additional 4 h at room temperature. The tissue was cryoprotected by incubation in a phosphate buffer (pH 7.2) containing 2.3 M sucrose and 50% polyvinylpyrrolidone (22), mounted on aluminum nails, frozen, and stored in liquid nitrogen.
Semithin cryosections (0.5 μm) were cut with a Reichert Ultracut E ultramicrotome fitted with an FC-4E cryoattachment and then mounted on gelatin-coated slides. The sections were washed in TBS and then incubated with 1% BSA and 0.5 M ammonium chloride in TBS for 15 min at 20°C in a humidified chamber. The ammonium chloride quenching wash was removed and the sections were incubated with 1% SDS in TBS for 5 min. The sections were washed twice with TBS and then blocked with 10% goat serum and 0.1% BSA in TBS for 15 min at 20°C. The sections then were incubated with anti-CFEX or antipendrin primary antibody diluted in TBS containing 10% goat serum and 0.1% BSA for 16 h at 4°C. The primary antibodies were removed and the sections were washed three times (10 min each wash) with high-salt TBS (3.4% NaCl) containing 0.1% BSA and then incubated for 1 h with a fluorescein-conjugated secondary antibody (Alexa Fluor 488 goat anti-rabbit IgG, Molecular Probes) diluted 1:200 with the TBS/BSA/goat serum blocking buffer. The slides were washed for 10 min with high-salt TBS, twice with standard TBS (0.9% NaCl; 10 min each wash), and then mounted in VectaShield (Vector Laboratories) to inhibit fading of the immunofluorescent signal. The slides then were visualized on a Zeiss Axiophot phase-contrast microscope.
Before use for immunofluorescence, 10-μl aliquots of the affinity-purified primary antisera were incubated with 30 μg of either the CFEX or pendrin peptide described above in 500 μl of TBS for 16 h at 4°C. The samples were centrifuged at 18,000 × g for 15 min at 4°C. The supernatants then were removed and stored at 4°C until needed. The anti-CFEX/peptide and antipendrin/peptide mixtures were used at dilutions of 1:4 and 1:6, respectively.
Functional Expression in Xenopus Oocytes.
Oocytes were isolated from Xenopus laevis frogs by partial ovarectomy under tricane (3-aminobenzoic acid ethyl ester) anesthesia (0.2% for 5–10 min). A small incision was made in the abdomen and a lobe of ovary was removed. The removed tissue was manually dissected into small clumps of 5–10 oocytes by using small forceps and washed for 20 min in Ca-free hypotonic medium (85 mM NaCl, 2 mM KCl, 1 mM MgCl2, 5 mM Hepes titrated with Tris base to pH 7.5) to remove blood and damaged tissue. Oocytes then were defolliculated by treatment with 2 mg/ml collagenase (Worthington type I or Sigma type I) in Ca-free hypotonic solution for 45–90 min with gentle agitation at room temperature. The oocytes were periodically checked and the digestion was terminated when the oocyte follicle had been removed with minimal damage to the oocytes. After this incubation oocytes were washed three times in Ca-free hypotonic media, then washed three times in isotonic solution (96 mM NaCl, 2.0 mM KCl, 1.8 mM CaCl2, 1.0 mM MgCl2, 5 mM Hepes, pH 7.5). Stage V-VI oocytes then were selected and maintained at 18°C in the same isotonic solution supplemented with 50 mg/ml gentamicin and 2.5 mM sodium pyruvate.
pGH19 plasmid DNA was linearized by XhoI digestion, and cRNA was transcribed by using T7 RNA polymerase (mMESSAGE mMACHINE, Ambion, Austin, TX). Precipitated cRNA was dissolved in sterile H2O, and yield and quality were assessed by spectroscopy and agarose gel electrophoresis. On the day of their isolation, oocytes were loaded with 50 nl of sterile H2O or 50 nl of a cRNA solution containing 25 ng of CFEX cRNA by use of a pneumatic injector. The injected oocytes then were incubated at 18°C for ≈48 h to allow for expression of the protein.
For uptake experiments, oocytes were washed twice at room temperature in 1 ml chloride-free buffer (98 mM K-gluconate/1.8 mM hemi-calcium-gluconate/1 mM hemi-magnesium-gluconate/5 mM Hepes, pH 7.5) before incubation in 500 μl of the same solution containing 3.4 mM 36Cl or 38 μM [14C]formate. After 30-min incubation, external isotope was removed by washing the oocytes three times with 1 ml of ice-cold chloride-free buffer. For chloride efflux measurements, oocytes were incubated for 60 min in a chloride-free buffer containing 36Cl. After three washes in ice-cold chloride-free buffer, the 36Cl content of oocytes was measured both initially and 30 min after suspension in solutions containing test anions. Net efflux was calculated as the difference between the initial oocyte 36Cl content and that remaining after 30-min reincubation. For experiments in which 10 mM anions (Cl−, formate, HCO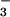 , acetate) were tested for effects on [14C]formate uptake or 36Cl efflux, gluconate was isosmotically replaced by each test anion, and media were buffered with 5 mM Tris titrated with Hepes to pH 8.5. An external pH of 8.5 was used because it allowed the pH of the HCO3 solution to remain stable due to the ampholyte effect (23). Radioisotope content of each individual oocyte was measured by scintillation spectroscopy after solubilization in 0.2 ml of 10% SDS and addition of 3 ml scintillation fluid (Opti-Fluor, Packard). Results shown in the bar graphs represent means ± SE for the number of oocytes indicated. In some cases the SE values are too small to be visible in the figures.
, acetate) were tested for effects on [14C]formate uptake or 36Cl efflux, gluconate was isosmotically replaced by each test anion, and media were buffered with 5 mM Tris titrated with Hepes to pH 8.5. An external pH of 8.5 was used because it allowed the pH of the HCO3 solution to remain stable due to the ampholyte effect (23). Radioisotope content of each individual oocyte was measured by scintillation spectroscopy after solubilization in 0.2 ml of 10% SDS and addition of 3 ml scintillation fluid (Opti-Fluor, Packard). Results shown in the bar graphs represent means ± SE for the number of oocytes indicated. In some cases the SE values are too small to be visible in the figures.
Results
In an attempt to identify an anion exchanger capable of mediating Cl− reabsorption in the proximal tubule, we screened the EST database for homologs of pendrin. A novel mouse EST was identified, and 5′ rapid amplification of cDNA ends was used to obtain the full coding sequence of the corresponding transcript. The sequence (GenBank accession no. AY032863) encodes a predicted protein of 735 aa. A database search indicated very high similarity to human SLC26A6 (79% amino acid identity) (24, 25) and lower, but significant, similarity (30–40% identity) to pendrin (26) and other members of the SLC26 family of anion transporters such as down-regulated in adenoma (27), diastrophic dysplasia sulfate transporter (28), the sulfate transporter SAT1 (29), and prestin (30). SLC26A6 was previously identified as encoding a putative anion exchanger expressed in renal tubules and pancreatic ducts (24, 25), although functional expression studies failed to detect anion transport activity (25). A tblastn search of the National Center for Biotechnology Information working draft sequence of the human genome indicated no higher match than to exons of SLC26A6. Accordingly, it is very likely that the cDNA we have cloned is the mouse ortholog of human SLC26A6. For the purpose of the present discussion, we will refer to the novel mouse protein as CFEX, in view of its ability to mediate Cl−-formate exchange (see below). The predicted CFEX protein has a large hydrophobic domain predicted to contain 10–12 membrane-spanning segments and a hydrophilic domain containing a consensus site for tyrosine phosphorylation and a possible PDZ interacting domain at the C terminus, as described for SLC26A6 (24, 25).
The expression of CFEX in mouse tissues was evaluated by Northern analysis, as indicted in Fig. 1. The principal transcript of ≈2.7 kb was most abundant in heart, liver, and kidney, with lesser degrees of expression noted in testis, brain, lung, and skeletal muscle. The ubiquitous pattern of expression of a 2.7-kb transcript with greatest abundance in heart and kidney is similar to that reported for SLC26A6 in human tissues (25). However, whereas CFEX was well expressed in liver, SLC26A6 was barely, if at all, detected in this organ. Thus, if CFEX and SLC26A6 represent orthologs of the same gene, there are significant species differences in its level of expression in different organs.
Figure 1.

(a) Northern analysis of CFEX expression in mouse tissues. (b) Actin control.
To evaluate whether CFEX is expressed on the brush border membrane of proximal tubule cells, we generated a CFEX-specific antipeptide antibody for immunocytochemical studies. To verify that the antibody is capable of reacting with the intact CFEX protein, immunoblotting was performed by using COS-7 cells transiently transfected with CFEX cDNA. As demonstrated in Fig. 2, the antibody labeled a polypeptide of ≈78 kDa in CFEX-transfected cells that was absent in control cells. Antibody staining was blocked by preincubation of the antibody with the CFEX peptide against which it had been generated (not shown). These findings confirmed the CFEX specificity of the antibody. In mouse kidney membranes, the antibody labeled a single band of ≈90 kDa, indicating expression of CFEX in native renal tissue. The small disparity in molecular mass may reflect incomplete glycosylation in the transfected overexpressing COS-7 cells.
Figure 2.

Immunoblot to verify specificity of anti-CFEX antibody. (Left) Untransfected COS-7 cells. (Center) CFEX-transfected COS-7 cells. (Right) Mouse kidney membranes.
To determine the cellular and subcellular localization of CFEX in the kidney, indirect immunofluorescence microscopy was carried out on semithin (0.5 μm) cryosections of PLP-fixed mouse kidney, as illustrated in Fig. 3. The anti-CFEX antibody stained the apical surface of proximal tubules (Fig. 3A). Confirming its specificity, this staining was absent when the antibody was preincubated with the CFEX peptide against which it was generated (Fig. 3B). A higher-power view confirmed the staining of the brush border membrane of proximal tubule cells (Fig. 3 C and D). It may be noted that not all proximal tubule segments were stained, indicating heterogeneity with respect to CFEX expression.
Figure 3.

Immunolocalization of CFEX in mouse kidney. (A) Staining of proximal tubules with anti-CFEX antibody in the presence of pendrin peptide. (B) Absence of staining when antibody was blocked with CFEX peptide. (C) Brush border staining. (D) Phase image for C. Magnifications: A and B × 160; C and D × 420.
Because it has been proposed that pendrin may mediate apical Cl−-base exchange in the proximal tubule (15), we performed additional immunocytochemical studies with an antipendrin antibody. As shown in Fig. 4 A and B, the antipendrin antibody failed to stain the apical membrane of proximal tubule cells but labeled the apical membranes of cells in the cortical collecting tubule. This staining was blocked when the antipendrin antibody was preincubated with the pendrin peptide against which it had been generated (Fig. 4 C and D). The observation of pendrin labeling on the apical membrane of collecting tubule cells rather than proximal tubule cells confirms the results of a recent report (17). Taken together, the findings in Figs. 3 and 4 indicate that the expression of CFEX, but not pendrin, can be detected on the brush border membrane of proximal tubule cells.
Figure 4.

Immunolocalization of pendrin in mouse kidney. (A) Staining of collecting tubule cells with antipendrin antibody in the presence of CFEX peptide. (B) Phase image for A. (C) Absence of staining when antibody was blocked with pendrin peptide. (D) Phase image for C. *, Collecting tubules. Magnifications: ×160.
Given that CFEX is expressed on the apical membrane of proximal tubule cells and is therefore a candidate for mediating brush border Cl−-formate exchange, we next examined its ability to mediate Cl−-gradient-stimulated formate uptake when heterologously expressed in Xenopus oocytes. Intracellular Cl− is ≈30 mM in Xenopus oocytes in isotonic saline media (31). We measured uptake of 38 μM [14C]formate after washing and reincubation of oocytes in the absence of external Cl− so that an outward Cl−-gradient was imposed. As shown in Fig. 5, formate uptake was greatly stimulated into oocytes injected with CFEX cRNA as compared with water-injected controls. The CFEX-mediated formate uptake was almost completely inhibited by the disulfonic stilbene DIDS (4,4′-diisothiocyanostilbene-2,2′-disulfonic acid), a known inhibitor of renal brush border Cl−-formate exchange (2–5). These results demonstrate that CFEX can function as a DIDS-sensitive formate transporter.
Figure 5.

CFEX-mediated formate uptake. Uptake of 38 μM [14C]formate was assayed in water- and CFEX cRNA-injected Xenopus oocytes. Effects of 0.1 mM DIDS also were measured.
As one method to test the anion specificity of CFEX, we compared the effects of various anions added to the external solution as inhibitors of [14C]formate. As indicated in Fig. 6, uptake of 38 μM [14C]formate was strongly inhibited by unlabeled formate, as expected for a saturable formate transport process. Formate uptake was very strongly inhibited by external Cl−, suggesting that Cl− may indeed be a substrate for the same transporter. Significant inhibition of formate uptake also resulted from addition of external HCO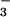 , although the effect was smaller than for Cl− and formate. In contrast, acetate, the next longer monocarboxylate, had no effect on formate uptake mediated by CFEX. The findings in Fig. 6 indicate that CFEX shares several properties with the renal brush border Cl−-formate exchanger, including the absence of affinity for acetate (2–4). However, the inhibition by HCO
, although the effect was smaller than for Cl− and formate. In contrast, acetate, the next longer monocarboxylate, had no effect on formate uptake mediated by CFEX. The findings in Fig. 6 indicate that CFEX shares several properties with the renal brush border Cl−-formate exchanger, including the absence of affinity for acetate (2–4). However, the inhibition by HCO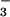 raised the possibility that CFEX might function as a more general Cl−-base exchanger as described for pendrin (15).
raised the possibility that CFEX might function as a more general Cl−-base exchanger as described for pendrin (15).
Figure 6.

Effect of external anions on [14C]formate uptake. Uptake of 38 μM [14C]formate was assayed in water- and CFEX cRNA-injected Xenopus oocytes. Effects of 10 mM of each indicated anion on CFEX-mediated [14C]formate uptake were assessed by isosmotic replacement of gluconate in the media.
To test whether CFEX mediates Cl− transport, as predicted for a Cl−-formate or Cl−-base exchanger, we next measured 36Cl uptake. As demonstrated in Fig. 7, uptake of 3.4 mM 36Cl was greatly stimulated into oocytes injected with CFEX cRNA as compared with water-injected controls. As in the case of CFEX-mediated formate flux, 36Cl uptake was almost completely inhibited by DIDS. Moreover, as also shown in Fig. 7, CFEX-mediated Cl− uptake was unaffected by imposition of an outside acid pH gradient, arguing against significant Cl−-OH− exchange. In these experiments, 36Cl uptake was measured in the absence of added external Cl− so that an outward gradient of unlabeled Cl− was present. Under these conditions, CFEX-mediated 36Cl uptake most likely represented Cl−-Cl− exchange, which is a known mode of the renal brush border Cl−-formate exchanger (2, 3). Exchange of Cl− for endogenous formate also might have contributed to the measured 36Cl uptake.
Figure 7.

CFEX-mediated Cl− uptake. Uptake of 3.4 mM 36Cl was assayed in water- and CFEX cRNA-injected Xenopus oocytes. Media containing 5 mM Hepes were titrated with Tris base to pH 7.5 or 6.5. Effects of 0.1 mM DIDS also were measured.
A final series of transport studies were performed to test directly whether CFEX functions as an anion exchanger. To this end, CFEX cRNA-injected oocytes were preloaded with 36Cl by preincubation for 60 min under the same conditions used to measure 36Cl uptake in Fig. 7. The oocytes then were washed and resuspended in media containing 10 mM external Cl−, formate, HCO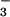 , or acetate and reincubated for an additional 30 min. As shown in Fig. 8, external Cl− and formate each markedly stimulated net loss of 36Cl compared with the control medium. In contrast, no appreciable stimulation of 36Cl efflux followed exposure to external acetate or HCO
, or acetate and reincubated for an additional 30 min. As shown in Fig. 8, external Cl− and formate each markedly stimulated net loss of 36Cl compared with the control medium. In contrast, no appreciable stimulation of 36Cl efflux followed exposure to external acetate or HCO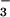 . These results are consistent with CFEX-mediated Cl−-Cl− and Cl−-formate exchange, whereas exchange of Cl− for acetate or HCO
. These results are consistent with CFEX-mediated Cl−-Cl− and Cl−-formate exchange, whereas exchange of Cl− for acetate or HCO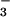 was minimal. These findings are in accord with the known modes of the renal brush border Cl−-formate exchanger (2, 3).
was minimal. These findings are in accord with the known modes of the renal brush border Cl−-formate exchanger (2, 3).
Figure 8.

Effect of external anions on 36Cl efflux. Oocyte content of 36Cl was assayed after the initial 60-min period of incubation with 3.4 mM 36Cl, and after 30-min reincubation in 36Cl-free K-gluconate media without (control) or with isosmotic replacement of gluconate by 10 mM of each indicated anion.
Discussion
We have described the cloning and preliminary characterization of CFEX, a Cl−-formate exchanger expressed on the brush border membrane of proximal tubule cells. The transport properties of CFEX, including its lack of detectable affinity for acetate and its ability to mediate Cl−-formate and Cl−-Cl− but not Cl−-OH− exchange, closely resemble those described for the renal brush border Cl−-formate exchanger in physiological studies using membrane vesicles (2, 3) and perfused tubules (4, 5). Similarly, HCO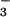 was a poor substrate for exchange with Cl− in studies of renal brush border vesicles (1, 2), and we find that CFEX mediates little, if any, Cl−-HCO
was a poor substrate for exchange with Cl− in studies of renal brush border vesicles (1, 2), and we find that CFEX mediates little, if any, Cl−-HCO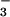 exchange. However, we did observe significant inhibition by HCO
exchange. However, we did observe significant inhibition by HCO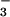 of CFEX-mediated formate transport, raising the possibility that external HCO
of CFEX-mediated formate transport, raising the possibility that external HCO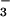 is a modifier of CFEX activity. It is interesting to note that formate stimulation of proximal tubule NaCl absorption is abolished when tubules are perfused with 25 mM compared with 5 mM HCO
is a modifier of CFEX activity. It is interesting to note that formate stimulation of proximal tubule NaCl absorption is abolished when tubules are perfused with 25 mM compared with 5 mM HCO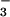 (4). This observation has been attributed to a requirement for luminal acidification to permit the pH-dependent recycling of formic acid to replenish intracellular formate and thereby sustain Cl− absorption by apical membrane Cl−-formate exchange (4). The present findings raise the additional possibility that luminal HCO
(4). This observation has been attributed to a requirement for luminal acidification to permit the pH-dependent recycling of formic acid to replenish intracellular formate and thereby sustain Cl− absorption by apical membrane Cl−-formate exchange (4). The present findings raise the additional possibility that luminal HCO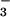 has a modifier role to regulate the rate of Cl−-formate exchange in the proximal tubule.
has a modifier role to regulate the rate of Cl−-formate exchange in the proximal tubule.
In addition to Cl−-formate exchange, a significant component of Cl− absorption in the proximal tubule can be mediated by Cl−-oxalate exchange (3, 5, 6, 32). Studies in membrane vesicles have suggested that Cl−-oxalate exchange is mediated by a separate pathway than that mediating Cl−-formate exchange (3, 33). Whether CFEX can mediate transport of oxalate remains to be evaluated.
Sequence analysis indicated that CFEX is very likely the mouse ortholog of human SLC26A6, which is very highly expressed on the apical membrane of pancreatic duct cells (24). Thus, CFEX also may play a role in transcellular Cl− transport in epithelia other than the proximal tubule. Moreover, Northern analysis indicates that CFEX expression is not limited to epithelial cells. For example, CFEX is relatively highly expressed in heart, and Cl−-formate exchange has been described in cardiac myocytes as well as vascular smooth muscle cells (34). The physiological role of Cl−-formate exchange in such nonepithelial tissues is presently unknown. Although we did not detect measurable Cl−-OH− or Cl−-HCO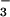 exchange activity in our assays of CFEX-mediated 36Cl flux in Xenopus oocytes, it is possible that CFEX might function as a more general Cl−-base exchanger in some native tissues under physiologic conditions.
exchange activity in our assays of CFEX-mediated 36Cl flux in Xenopus oocytes, it is possible that CFEX might function as a more general Cl−-base exchanger in some native tissues under physiologic conditions.
In conclusion, we find that CFEX mediates Cl−-formate exchange and is expressed on the brush border membrane of proximal tubule cells. Accordingly, CFEX may be responsible for mediating formate-stimulated NaCl absorption in the proximal tubule. It thereby may play an important role in renal NaCl reabsorption. Given its wide tissue distribution, CFEX also may contribute to transcellular Cl− transport in additional epithelia such as the pancreas and contribute to transmembrane Cl− transport in nonepithelial tissues such as the heart.
Acknowledgments
We thank Gordon MacGregor for his advice concerning the Xenopus oocyte expression studies. This work was supported by National Institutes of Health Grants DK-17433 and DK-33793.
Abbreviations
- EST
expressed sequence tag
- DIDS
4,4′-diisothiocyanostilbene-2,2′-disulfonic acid
Footnotes
Data deposition: The sequence reported in this paper has been deposited in the GenBank database (accession no. AY032863).
References
- 1.Aronson P S, Giebisch G. Am J Physiol. 1997;273:F179–F192. doi: 10.1152/ajprenal.1997.273.2.F179. [DOI] [PubMed] [Google Scholar]
- 2.Karniski L P, Aronson P S. Proc Natl Acad Sci USA. 1985;82:6362–6365. doi: 10.1073/pnas.82.18.6362. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Karniski L P, Aronson P S. Am J Physiol. 1987;253:F513–F521. doi: 10.1152/ajprenal.1987.253.3.F513. [DOI] [PubMed] [Google Scholar]
- 4.Schild L, Giebisch G, Karniski L P, Aronson P S. J Clin Invest. 1987;79:32–38. doi: 10.1172/JCI112803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Wang T, Giebisch G, Aronson P S. Am J Physiol. 1992;263:F37–F42. doi: 10.1152/ajprenal.1992.263.1.F37. [DOI] [PubMed] [Google Scholar]
- 6.Wareing M, Green R. J Physiol (London) 1994;477:347–354. doi: 10.1113/jphysiol.1994.sp020196. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Sheu J N, Quigley R, Baum M. Am J Physiol. 1995;268:F847–F853. doi: 10.1152/ajprenal.1995.268.5.F847. [DOI] [PubMed] [Google Scholar]
- 8.Biemesderfer D, Pizzonia J, Abu-Alfa A, Exner M, Reilly R, Igarashi P, Aronson P S. Am J Physiol. 1993;265:F736–F742. doi: 10.1152/ajprenal.1993.265.5.F736. [DOI] [PubMed] [Google Scholar]
- 9.Amemiya M, Loffing J, Lotscher M, Kaissling B, Alpern R J, Moe O W. Kidney Int. 1995;48:1206–1215. doi: 10.1038/ki.1995.404. [DOI] [PubMed] [Google Scholar]
- 10.Wang T, Yang C L, Abbiati T, Schultheis P J, Shull G E, Giebisch G, Aronson P S. Am J Physiol. 1999;277:F298–F302. doi: 10.1152/ajprenal.1999.277.2.F298. [DOI] [PubMed] [Google Scholar]
- 11.Schultheis P J, Clarke L L, Meneton P, Miller M L, Soleimani M, Gawenis L R, Riddle T M, Duffy J J, Doetschman T, Wang T, et al. Nat Genet. 1998;19:282–285. doi: 10.1038/969. [DOI] [PubMed] [Google Scholar]
- 12.Wu M S, Biemesderfer D, Giebisch G, Aronson P S. J Biol Chem. 1996;271:32749–32752. doi: 10.1074/jbc.271.51.32749. [DOI] [PubMed] [Google Scholar]
- 13.Vallon V, Schwark J R, Richter K, Hropot M. Am J Physiol. 2000;278:F375–F379. doi: 10.1152/ajprenal.2000.278.3.F375. [DOI] [PubMed] [Google Scholar]
- 14.Scott D A, Wang R, Kreman T M, Sheffield V C, Karniski L P. Nat Genet. 1999;21:440–443. doi: 10.1038/7783. [DOI] [PubMed] [Google Scholar]
- 15.Soleimani M, Greeley T, Petrovic S, Wang Z, Amlal H, Kopp P, Burnham C E. Am J Physiol. 2001;280:F356–F364. doi: 10.1152/ajprenal.2001.280.2.F356. [DOI] [PubMed] [Google Scholar]
- 16.Scott D A, Karniski L P. Am J Physiol. 2000;278:C207–C211. doi: 10.1152/ajpcell.2000.278.1.C207. [DOI] [PubMed] [Google Scholar]
- 17.Royaux I E, Wall S M, Karniski L P, Everett L A, Suzuki K, Knepper M A, Green E D. Proc Natl Acad Sci USA. 2001;98:4221–4226. doi: 10.1073/pnas.071516798. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Trimmer J S, Cooperman S S, Tomiko S A, Zhou J Y, Crean S M, Boyle M B, Kallen R G, Sheng Z H, Barchi R L, Sigworth F J, et al. Neuron. 1989;3:33–49. doi: 10.1016/0896-6273(89)90113-x. [DOI] [PubMed] [Google Scholar]
- 19.Biemesderfer D, DeGray B, Aronson P S. J Biol Chem. 2001;276:10161–10167. doi: 10.1074/jbc.M008098200. [DOI] [PubMed] [Google Scholar]
- 20.Laemmli U K. Nature (London) 1970;227:680–685. doi: 10.1038/227680a0. [DOI] [PubMed] [Google Scholar]
- 21.Larsson L. J Ultrastruct Res. 1975;51:119–139. doi: 10.1016/s0022-5320(75)80013-x. [DOI] [PubMed] [Google Scholar]
- 22.Tokuyasu K T. Histochem J. 1989;21:163–171. doi: 10.1007/BF01007491. [DOI] [PubMed] [Google Scholar]
- 23.Maren T H. Kidney Int. 1978;14:395–405. doi: 10.1038/ki.1978.144. [DOI] [PubMed] [Google Scholar]
- 24.Lohi H, Kujala M, Kerkela E, Saarialho-Kere U, Kestila M, Kere J. Genomics. 2000;70:102–112. doi: 10.1006/geno.2000.6355. [DOI] [PubMed] [Google Scholar]
- 25.Waldegger S, Moschen I, Ramirez A, Smith R J, Ayadi H, Lang F, Kubisch C. Genomics. 2001;72:43–50. doi: 10.1006/geno.2000.6445. [DOI] [PubMed] [Google Scholar]
- 26.Everett L A, Glaser B, Beck J C, Idol J R, Buchs A, Heyman M, Adawi F, Hazani E, Nassir E, Baxevanis A D, et al. Nat Genet. 1997;17:411–422. doi: 10.1038/ng1297-411. [DOI] [PubMed] [Google Scholar]
- 27.Hoglund P, Haila S, Socha J, Tomaszewski L, Saarialho-Kere U, Karjalainen-Lindsberg M L, Airola K, Holmberg C, de la Chapelle A, Kere J. Nat Genet. 1996;14:316–319. doi: 10.1038/ng1196-316. [DOI] [PubMed] [Google Scholar]
- 28.Hastbacka J, de la Chapelle A, Mahtani M M, Clines G, Reeve-Daly M P, Daly M, Hamilton B A, Kusumi K, Trivedi B, Weaver A, et al. Cell. 1994;78:1073–1087. doi: 10.1016/0092-8674(94)90281-x. [DOI] [PubMed] [Google Scholar]
- 29.Bissig M, Hagenbuch B, Stieger B, Koller T, Meier P J. J Biol Chem. 1994;269:3017–3021. [PubMed] [Google Scholar]
- 30.Zheng J, Shen W, He D Z, Long K B, Madison L D, Dallos P. Nature (London) 2000;405:149–155. doi: 10.1038/35012009. [DOI] [PubMed] [Google Scholar]
- 31.Barish M E. J Physiol (London) 1983;342:309–325. doi: 10.1113/jphysiol.1983.sp014852. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Kuo S M, Aronson P S. J Biol Chem. 1996;271:15491–15497. doi: 10.1074/jbc.271.26.15491. [DOI] [PubMed] [Google Scholar]
- 33.McConnell K R, Aronson P S. J Biol Chem. 1994;269:21489–21494. [PubMed] [Google Scholar]
- 34.Soleimani M, Howard R L. Circ Res. 1994;74:48–55. doi: 10.1161/01.res.74.1.48. [DOI] [PubMed] [Google Scholar]