

Abstract
A prominent feature of comparative life histories in fish (and other indeterminate growers) is the approximate invariance across species of certain dimensionless numbers made up from reproductive and timing variables. The two best known are the age at maturity (α) divided by the average adult lifespan (E), and the proportion of a body mass given to reproduction per year (c) multiplied by E. This article uses evolutionary life-history theory for nongrowing populations to predict the numeric value of these numbers for fish and lizards, with several new implications for the dynamics of ontogenetic development.
Keywords: dimensional analysis, ESS, fish, ontogeny, reptiles
Comparisons of life histories across species usually look for body-size scalings (allometries) or the matching of life history to features of the environment (1–5). More recently (history discussed in ref. 1), it has been shown that interesting general rules exist for life histories if they are viewed in a dimensionless way. For example, measure the average adult lifespan (E) as a ratio of the age at first reproduction (α); then E/α is (approximately) invariant within a closely related taxon like mammals. After removing absolute scale (e.g., age), the life histories of, say, elephants and squirrels look much alike and they look very different from fish, reptiles, or birds. Although there are disagreements in the literature (e.g., refs. 1–3 vs. refs. 4 and 5) as to how well a correlation between (say) E and α is sufficient to claim that E/α is invariant, r values are often above 0.8 and data are well fit by lines through the origin (or double logarithm relations with a slope of 1). Evolutionary life-history theory can be used to predict when such dimensionless invariance is to be expected (1–5).
This article develops a new evolutionary model for life histories (1–5) by combining a new production-growth model (6) with some general, yet specific, constraints on reproductive allocation, and by allowing natural selection to adjust the age of first breeding and the metabolic/productive capacity of cells. The optimal life histories correctly predict the numeric value of dimensionless numbers that combine maturation age (size), mortality, and reproductive allocation. The life-history prediction also yields a new lifetime growth model for body mass under indeterminate growth and predicts that the fastest growth rate will be near the size at first reproduction (7). Table 1 is a glossary of terms, whereas Appendix A is a brief overview of the optimization problem.
Table 1.
Glossary of terms and units
| Term | Definition | Units | Where defined |
|---|---|---|---|
| m | Body mass | Mass | Fig. 1 |
| α | Age at first reproduction | Time | Fig. 1 |
| mα | Size at first reproduction | Mass | Fig. 1 |
| m2 | Asymptotic size without reproduction | Mass | Fig. 1 |
| m∞ | Asymptotic size with reproduction | Mass | Fig. 1 |
| a⋅m0.75 | Assimulation of energy | Mass/time | Fig. 1, Appendix B |
| b | Maintenance metabolism per cell divided by the cost of a new cell | 1/time | Fig. 1, Appendix B |
| dm/dt | a⋅m0.75 − b⋅m, growth equation prior to mα | Mass/time | Fig. 1, Appendix B |
| dm/dt | a⋅m0.75 − b⋅m − c⋅m, growth equation after mα | Mass/time | Fig. 1, Appendix B |
| c⋅m | Reproductive allocation | Mass/time | Fig. 1 |
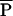 |
Average of c⋅m for an adult within a species | Mass/time | Text, Fig. 5 |
| q = c/b | Reproductive allocation constraint | Dimensionless | Appendix B |
| GSI | Gonadosomatic index (proportion of somatic body mass given to reproduction per year; estimates c) | 1/time | Text, Fig. 4 |
| Z(x) | Instantaneous mortality rate at age x | 1/time | Fig. 2, Appendix C |
| Z | Adult (m > mα) instantaneous mortality rate | 1/time | Fig. 2, Appendix C |
| E = 1/Z | Average adult lifespan | Time | Text, Appendix C |
Body-Size Growth
Many equations have been statistically fit to body-size growth data (8, 9) and most merely describe rather than explain. A new growth model from West et al. (6) is an exception because it derives net production from the first principles of energy intake minus maintenance and activity cost. The argument (Appendix B) leads to a differential equation for change in mass (m) of the form: (dm/dT) = am0.75 − bm (Eq. 1), where a is expected to be similar for species with similar metabolic scaling (e.g., within mammals, within fish). b is equal to the maintenance metabolic rate per existing cell divided by the cost of building a new cell. Eq. 1 excludes reproduction and would result in sigmoid growth to an asymptotic size [m2 = (a/b)4], as shown in Fig. 1. To add reproduction, we note that gonad mass in fish is commonly proportional to body mass (10–12); thus, after the onset of reproduction (age α) at size mα, growth follows dm/dT = am0.75 − (b + c)m, where c⋅m is the reproductive allocation. The asymptotic size has been shifted downward from m2 to m∞ (= [a/(b + c)]4). Thus, lifetime growth reflects production (Eq. 1) and the timing (α) and magnitude (c⋅m) of reproduction (refs. 1, 4, 13, and 14; see hatched area in Fig. 1).
Figure 1.

Schematic diagram of the growth model (dm/dT, m = mass) from Appendix B. Growth follows the domed curve until size at first reproduction (mα), then it follows the curve minus c⋅m, the reproductive allocation. Largest size (dm/dT = 0) is m2 without reproduction, m∞ with reproduction.
Life History Optimization
We assume nongrowing populations; thus, the appropriate fitness measure is the lifetime reproductive allocation (1)—the time integral of c⋅m, including, of course, mortality (see Eq. 1 of Appendix A). It seems reasonable to assume that α and c are the control variables, the two parameters most easily adjusted by natural selection across species (ref. 1; but see below). Appendix C details the evolutionary optimization argument; briefly, α and c are chosen to maximize a quantity proportional to lifetime production of offspring in the face of a mortality rate (Z) externally imposed on a nongrowing population. The optimization makes some quite specific assumptions about mortality (Z) and the limitations on c, the reproductive allocation as a fraction of body mass (m). Because these are important assumptions, we discuss them here.
Fig. 2 shows the mortality rate (Z) assumption. For eggs, larval and immature fish, mortality is high (and often density-dependent). Z is assumed to drop to some constant value before feasible ages of first breeding (α) and to remain constant until late in life when senescence may increase it (15–18). Contrary to suggestions by Kozlowski and coworker (4, 5), Z is probably not body-size (or age)-dependent over an ontogeny within a species, at least after the early life history and before senescence; the vast majority of fish species studied show no within-species body-size dependence of Z (15–18). Notice that the average adult lifespan (E) is Z−1.
Figure 2.

The instantaneous mortality rate Z(x) is externally imposed and is assumed age and size independent over most of the lifespan. See the text for a detailed discussion.
Theory for life-history evolution presents a major puzzle with reference to Fig. 1; many formal evolutionary models (19) predict that growth should cease with the onset of reproduction (i.e., mα → m∞). Because such determinate growth is uncommon outside of birds, mammals, and insects, the models must be ignoring something very basic and widespread. One logical candidate is that all production (Eq. 1) is simply not available to be funneled into reproduction (G. Williams, personal communication).
West et al. (ref. 6; Fig. 1 and Eq. 1) present us with a second puzzle: b is the metabolic maintenance cost per existing cell, which raises the question of why any species would have a high b—Why build a body of cells expensive to maintain? We hypothesize that expensive cells (high b) allow greater reproductive effort, c. As developed in Appendix C, c is determined by two factors: (i) the proportion of the body cells given over to reproduction, and (ii) the net production from the reproductive cells. Morphology (e.g., body shape and space for the gonad) may limit constraint i, but we hypothesize that to get greater production from the cells devoted to reproduction requires increased maintenance costs (higher b), and the individual is constrained to build all of its cells this way. Thus, we make c = b⋅q, where q includes constraints i and ii. We further suggest that q itself is more or less fixed by a combination of morphology and how cells produce material, so that evolutionary adjustment of c, the reproductive allocation, requires adjustment of b, the background maintenance cost per cell. Constraining c to be a multiplier of b effectively limits the amount of personal production (Eq. 1) that can be funneled to reproduction, and it leads to indeterminate growth where the onset of reproduction (mα) is at a much smaller size than the asymptotic size (m∞). Thus, the Darwinian fitness optimization in Appendix C really adjusts α and b in the face of the externally imposed mortality (Z). The optimal life histories are shown in Fig. 3, where we plot the predicted values of the dimensionless numbers q⋅b/Z (= c⋅E) and α⋅Z (= α/E) as a function of q.
Figure 3.

Appendix C derives the optimal life histories by adjusting the age of first reproduction (α) and the reproductive allocation (c = qb; see Fig. 1 for c) in the face of externally imposed mortality (Z) to maximize lifetime production of offspring (R0). q constrains reproductive allocation (c) to be a multiplier of the maintenance metabolic rate per cell (b). The optimal life history has the value of the dimensionless numbers b/Z (= b⋅E), α⋅Z (= α/E), and qb/Z (= c⋅E) determined solely by q. Z−1 is E, the average adult lifespan.
Data Results: Fish and Reptiles
α⋅Z has been estimated for various fish groups (1) by a variety of means, and α⋅Z ≃ 2. α⋅Z has been estimated for various parasitic nematodes (20) and Pandalid shrimp (1), with averages again near 2. This α⋅Z value predicts (Fig. 3) that b/Z ≃ 0.17–0.13, or that q⋅b/Z (= c⋅E) should equal 0.50–0.65. Fisheries biologists (17, 18) often estimate the mass of mature gonads divided by the somatic body mass (multiplied by the spawning per year) as a measure of the reproductive allocation rate. This is termed the gonadosomatic index (GSI) and is an estimate of c. Gunderson and coworker (17, 18) showed that the GSI was highly correlated with the adult instantaneous mortality rate across fish species. Fig. 4 shows Gunderson's (17) plot of GSI vs. adult mortality for 28 fish stocks; the relation is proportional [ref. 17; fitted lines (y on x or x on y) go through origin; 95% confidence intervals include zero] with a high correlation (r = 0.87). The average of the 28 GSI/mortality ratios is 0.60 (SE = 0.05), near the center of the predicted 0.50–0.65 range.
Figure 4.

The adult instantaneous mortality rate is proportional to the gonadosomatic index for 28 spp. of fish (data and analysis from ref. 17).
α⋅Z is not near 2 for all indeterminate growers; indeed Charnov and coworker (1, 21) showed that Z and α were inversely proportional across 45 snake and lizard populations (36 spp.), resulting in α⋅Z ≃ 1.32. But this reptile α takes age zero to be hatching when the offspring are a small but non-zero size. The optimal life histories (Appendix C) remain the same whether the offspring begin their independent growth at non-zero size, but the α of Fig. 3 now refers to age zero as being when the posthatching growth curve is extrapolated back to zero. Charnov (ref. 1, p. 75) reestimated the αs for the snakes and lizards, extending the body-size growth curves back to zero. The Zs and the new αs were still inversely proportional, and the new α⋅Z was ≃2.1, near the fish, nematode, and shrimp values. This α⋅Z again predicts c⋅E to equal ≃0.6. Shine and Schwarzkopf (22) compiled data on yearly clutch mass divided by adult (somatic) mass [termed relative clutch mass (RCM)], and yearly adult mortality (Z) for 18 spp. (35 populations, all sexual species) of lizards and 10 spp. of snakes. The lizards look similar to the fish with the average of RCM/Z equal to 0.64 and a moderate, positive correlation between RCM and Z (r = 0.56, n = 35, P < 0.01). However, the snakes are quite different with an average RCM/Z equal to 1.17, almost double the lizard/fish value. The snakes show little variation in Z, and no significant correlation between Z and RCM. Thus, lizards weakly support the model, whereas snakes do not at all.
The life-history theory developed here aims to solve the two problems of why any organism would build cells with high-maintenance cost (high b) and just what limitations on reproductive allocation (c of Fig. 1) act to yield the indeterminate growth so common outside birds, mammals, and insects. We suggest that c/b is constrained to be a fixed value (= q). The resulting optimal life history (Fig. 3) uses the commonly observed α/E value of 2 to successfully predict (for fish and, less so, for lizards) that c⋅E should be about 0.6. Fig. 3 also predicts that q ≃ 3–5 and b⋅E ≃ 0.15; both of these predictions may be testable.
One Other Prediction: Reproductive Allometry
This article focuses on dimensionless predictions for the optimal life histories. This approach is taken partly because invariance of the dimensionless numbers is one of the strongest rules for comparative (across species) life histories, often much stronger than body-size rules (1, 17, 18). Notice that none of the dimensionless results (Fig. 3) depend on the numeric value of a, the “height” of the production in Eq. 1. To incorporate a into this dimensionless analysis requires that we construct a quantity with the same (or opposite) dimensions; a has units of (mass)0.25/time, thus it is quite natural to multiply it by (mass)0.75 to yield a term with units mass/time, or yearly production. It is straightforward to show that if 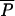 is the adult average mass devoted to reproduction per year (average of c⋅m within a species) and mα is the mass at first reproduction,
is the adult average mass devoted to reproduction per year (average of c⋅m within a species) and mα is the mass at first reproduction, 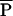 /a⋅m
/a⋅m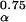 is a unique function of the three other dimensionless numbers α/E, b⋅E, and c/b. Because these three numbers are predicted (or for c/b, assumed) to take on the same values across species,
is a unique function of the three other dimensionless numbers α/E, b⋅E, and c/b. Because these three numbers are predicted (or for c/b, assumed) to take on the same values across species, 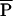 /a⋅m
/a⋅m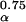 equals “a constant” and we have
equals “a constant” and we have 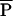 ∝ a⋅m0.75 for species with similar a values. Winemiller and Rose (23) estimated reproductive parameters for 139 marine and freshwater fish species. Fig. 5 plots the average mass given to reproduction per year (
∝ a⋅m0.75 for species with similar a values. Winemiller and Rose (23) estimated reproductive parameters for 139 marine and freshwater fish species. Fig. 5 plots the average mass given to reproduction per year (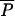 ) vs. length at maturity cubed (to give mα) for the species; as predicted, the double logarithm plot is linear with a slope near ¾. We use ordinary least squares regression because we believe the y variate is estimated with much greater error than the x variate. The filled square is the mean (±SE) for a sample of 48 spp. of North American freshwater darters (small, perch-like fish) compiled in ref. 24. These species were not included in the regression, but clearly are not different from the other species.
) vs. length at maturity cubed (to give mα) for the species; as predicted, the double logarithm plot is linear with a slope near ¾. We use ordinary least squares regression because we believe the y variate is estimated with much greater error than the x variate. The filled square is the mean (±SE) for a sample of 48 spp. of North American freshwater darters (small, perch-like fish) compiled in ref. 24. These species were not included in the regression, but clearly are not different from the other species.
Figure 5.

Reproductive allometry for fish. The average mass devoted to reproduction scales with an exponent near ¾ when plotted against the size at first reproduction for 139 spp. of North American fishes (methods and species list in ref. 23). The y variate is estimated as numbers of eggs laid per year by an adult multiplied by the egg diameter cubed. The x variate is the length at first reproduction cubed. Statistics: SE, slope = 0.04; intercept = 0.64. (All 95% confidence intervals are ± 2 SE.) The solid square is the mean (± SE) for 48 spp. of freshwater darters (from ref. 24) not included in the regression. They are on the same fitted line. Eleven species from ref. 23 excluded from the analysis: semelparous salmon, cave fish, and deep-sea eels; the latter two groups have very low production/growth rates and the length cubed is not a good estimate of body mass for eels. Regression is ordinary least squares, because measurement error is much greater for y variate.
Discussion
Probably the most interesting assumption of the model is that c/b is constrained to a fixed number, so that adjustment of reproduction c requires adjustment of b. This assumption gives an evolutionary reason for high b and it also outputs indeterminate growth. Indeed, the c/b constraint suggests that it ought to be difficult to evolve determinate growth; why birds and mammals may have determinate growth is discussed elsewhere (25, 26).
The model has three more general implications for the structure of life histories and body-size growth. First, Fig. 1 is schematic but slightly misleading in that the c⋅m line should be shifted far to the left. The shift in asymptotic size caused by reproduction (m2 → m∞) is large for q estimated to be 3–5, which means that prereproductive growth is on the ascending part of the am0.75 − bm curve. The fastest dm/dT should be at mα, because the slowing of growth is caused only by reproductive allocation; this result seems to be true for fish (7). Second, reproductive allocation (c⋅m) is optimized with respect to the mortality rate (Z), but there is no tradeoff between the two in the sense that higher c⋅m does not cause higher mortality. Rather, the causality is just the reverse; shorter lifespan (high Z) selects for greater per-unit time allocation to reproduction (higher c). Finally, the c⋅E is the average proportion of a body mass expended in reproduction over an average adult lifetime. Thus, a female fish expends a mass equal to about 60% of her average adult body mass per lifetime of reproduction (Fig. 4). This number (0.6) is very low when compared with, say, altricial birds, where a typical female rears about five daughters over a lifetime, each fed to adult size (25, 27, 28), so that c⋅E ≃ 5. One wonders why birds are almost 10 times more productive by this measure (flight? endothermy? habitat productivity?).
Acknowledgments
We thank G. West, J. Brown, R. Shine, R. Sibly, J. Downhower, D. Gunderson, and B. Enquist for discussion. P. Andrews, P. Taylor, and A. Irwin helped greatly with the math for Appendix C and Fig. 3. E.L.C. was supported by a MacArthur Fellowship.
Abbreviation
- RCM
relative clutch mass
Overview of the Optimization Argument
Body mass (m) follows dm/dT = a⋅m0.75 − b⋅m (Fig. 1, Appendix B), where a⋅m0.75 is proportional to metabolic scaling and b is the maintenance metabolic rate per cell divided by the cost of a new cell. At the onset of reproduction (age α, size mα), an allocation equal to a proportion c of body mass (c⋅m) is diverted to reproduction, and growth slows to dm/dT = a⋅m0.75 − (b + c)m. The optimization problem is to maximize lifetime reproduction, the time integral of c⋅m, which also includes mortality; thus, MAX ∫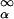 L(x)⋅c⋅m(x,α)dx (Eq. 1), where L(x) is the chance of being alive at age x [=
L(x)⋅c⋅m(x,α)dx (Eq. 1), where L(x) is the chance of being alive at age x [= 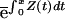 , where Z(y) is the instantaneous mortality rate at age y; Fig. 2]. The control variables are α and c (Fig. 1).
, where Z(y) is the instantaneous mortality rate at age y; Fig. 2]. The control variables are α and c (Fig. 1).
If c can take on any value up to 100% of growth, it is (almost) always optimal to give all growth to reproduction at age α (i.e., c = a⋅m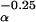 − b, called determinate growth). But we propose that “expensive cells” (high b) are needed to produce (to allow) greater reproductive effort (high c), thus c/b is a fixed value, a constraint. This constraint assumption leads to a lower value for c, with additional growth after age α (called indeterminate growth; Fig. 1).
− b, called determinate growth). But we propose that “expensive cells” (high b) are needed to produce (to allow) greater reproductive effort (high c), thus c/b is a fixed value, a constraint. This constraint assumption leads to a lower value for c, with additional growth after age α (called indeterminate growth; Fig. 1).
If Z is the adult instantaneous mortality rate, Z−1 is the average adult lifespan. Plausible values of the (assumed, constraining) ratio of c to b give (Fig. 3) αZ ≈ 2, which is a well established regularity for fish and a few other taxa with indeterminate growth, and c/Z ≈ 0.6 (a measure of lifetime reproductive allocation), which seems to be consistent with data on relative gonad sizes of fish species in relation to average adult lifetimes (Fig. 4).
The Production/Growth Model
West et al. (6) expand their previous work (29) on whole-organism metabolic rate to encompass growth. Their basic starting point is the balance of energy flow within an organism. Incoming energy and materials from the environment are transported by means of hierarchical branching network systems to supply all cells. In general, these resources are transformed into metabolic energy, which is utilized for life-sustaining activities such as maintenance of existing biomass. During ontogeny, however, some fraction of the total metabolic energy is allocated to the production of new tissue. In general, then, the rate of energy transformation is the sum of two terms, one representing the maintenance of existing tissue and the other the creation of new tissue. This scheme can be expressed by the dynamic conservation of energy equation:
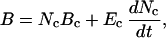 |
2 |
where the incoming rate of energy flow, B, is the average metabolic rate of the whole organism at some time t. The whole organism is composed of fundamental units, cells, subscripted c. The metabolic rate of a single cell is denoted by Bc, and the metabolic energy required to create such a cell by Ec. Nc is the total number of cells; here we consider some average typical cell as the fundamental unit. The first term of Eq. 2, Nc⋅Bc, is simply the power needed to sustain the organism in all of its activities, whereas the second term is the power allocated to the production of new cells and therefore to growth. Ec, Bc, and the mass of the cell, mc, are assumed to remain constant throughout growth and development.
At any time, t, the total body mass, m = mcNc, thus Eq. 2 can be written as:
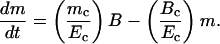 |
3 |
Now, the whole-organism metabolic rate scales (29) as B = B0m3/4, where B0 is constant for a given taxon. This equation for B immediately leads to the general growth/production equation:
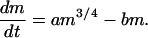 |
4 |
To include reproduction, assume that after maturity (age α, size mα), c⋅m of production is devoted to reproduction; this rule then subtracts an additional c⋅m from Eq. 4. Fig. 1 displays the results. Eq. 4 has the largest size at m2 = (a/b)4 and the fastest growth at 0.32m2; reproductive allocation moves the largest size down to m∞ = [a/(b + c)]4.
The argument leading to Eq. 4 is similar to the original argument for the classic Bertalanffy growth equation (8, 9), which differs in having a 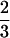 rather than a ¾ exponent. Although Bertalanffy had absorption of food by the gut (assumed to be a
rather than a ¾ exponent. Although Bertalanffy had absorption of food by the gut (assumed to be a 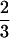 scaling) limit material (energy) delivery to cells, West et al. (6) put the delivery limit in the functioning of the hierarchical branching networks (e.g., blood) that supply cells [hence, a ¾ scaling (29)]. Lifetime growth (Fig. 1) further requires we consider the timing (mα) and magnitude of reproduction (c⋅m; refs. 1, 4, 13, 14).
scaling) limit material (energy) delivery to cells, West et al. (6) put the delivery limit in the functioning of the hierarchical branching networks (e.g., blood) that supply cells [hence, a ¾ scaling (29)]. Lifetime growth (Fig. 1) further requires we consider the timing (mα) and magnitude of reproduction (c⋅m; refs. 1, 4, 13, 14).
The Optimal Life History
Assume a nongrowing population with only survival of the very young as density-dependent (1). Let Z(x) be the instantaneous mortality rate at age x (Fig. 2); Z is high for small x but drops to a constant value some time before maturity at age α. Z is imposed externally to the organism. Growth and reproduction follow Appendix B (see Fig. 1). Lifetime production of offspring (R0) is proportional to the lifetime mass devoted to reproduction, as we have not specified offspring size; this is the time integral of the reproductive allocation, c⋅m. Thus (1),
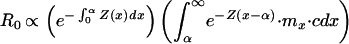 |
5 |
The use of R0 as a fitness measure can be derived from theoretical studies of evolutionary dynamics in nongrowing populations (30–32).
Natural selection is assumed to adjust α and c (the reproductive effort, from Fig. 1) to maximize Eq. 5. But it is well known (19) that if c can take on any value up to 100% of potential growth, the optimal life history is to grow to some size (mα) and then to devote all production to reproduction (i.e., mα and m∞ in Fig. 1 are the same, called determinate growth). There is a large amount of literature (19) devoted to what other factors might be added to the problem to generate an optimum where reproduction begins (mα) prior to the largest adult size (m∞), the indeterminate growth known for virtually all organisms except birds, mammals, and insects. A guess (G. Williams, personal communication) is that indeterminate growth results from strong constraints on the allowable values for c; it is simply not possible for all personal production to be funneled to reproduction (19). To reproduce, a fraction of the body cells are devoted to gonad, and those cells metabolize to yield a net gain in material. Assume that the net yield per reproductive cell is some multiplier of the maintenance metabolic rate per cell (Bc of Appendix B; the subscript c stands for cell): to get more out of a cell, one must pay a proportionally higher maintenance cost. In particular, assume (Appendix B) that c = k[(Bc/Ec)⋅δ − (Bc/Ec)], where k = proportion of the body's cells devoted to reproduction and (δ − 1)Bc/Ec is the net yield in metabolism from each reproductive cell. Because b in growth Eq. 1 is Bc/Ec, we have c = q⋅b, where q is a constant equal to k (δ − 1). Now, substitute q⋅b for c in Eq. 5, and begin growth near zero size at time zero. The math for R0 is straightforward:
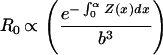 |
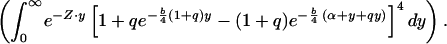 |
Thus,
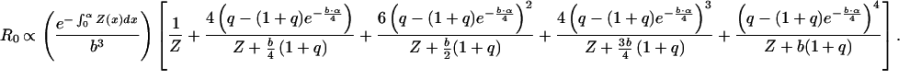 |
6 |
In the derivation of Eq. 6, we have dropped terms proportional to R0 that play no role in the optimization problem.
The optimal life history satisfies (∂R0/∂α) = 0 and (∂R0/∂b) = 0; the two derivatives (= 0) are complicated, but note that they contain only the variables α, Z, and b, and q. q is dimensionless, α is time, and Z and b are (time)−1. Thus, at the optimum, the dimensionless numbers αZ, b/Z, and bq/Z are solely functions of q. These functions are displayed in Fig. 3. Because the average adult lifespan (E) equals Z−1, these numbers may be written as α/E, b⋅E, and c⋅E.
References
- 1.Charnov E L. Life History Invariants. Oxford: Oxford Univ. Press; 1993. [Google Scholar]
- 2.Charnov E L, Berrigan D. Evol Ecol. 1991;5:63–68. [Google Scholar]
- 3.Charnov E L. Proc Natl Acad Sci USA. 1991;88:1134–1137. doi: 10.1073/pnas.88.4.1134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Kozlowski J. Proc R Soc London Ser B. 1996;263:559–566. [Google Scholar]
- 5.Kozlowski J, Weiner J. Am Nat. 1997;149:352–380. [Google Scholar]
- 6.West, G. B., Brown, J. H. & Enquist, B. J. (2001) Nature (London), in press. [DOI] [PubMed]
- 7.Jensen A L. Can J Fish Aquat Sci. 1996;53:820–822. [Google Scholar]
- 8.Ricker W E. Bull Fish Res Board Can. 1975;191:1–382. [Google Scholar]
- 9.Reiss M J. The Allometry of Growth and Reproduction. Cambridge, U.K.: Cambridge Univ. Press; 1989. [Google Scholar]
- 10.Roff D A. Can J Fish Aquat Sci. 1982;39:1686–1698. [Google Scholar]
- 11.Bagenal T B. In: The Biological Basis of Freshwater Fish Production. Gerking S D, editor. Oxford: Blackwell; 1966. [Google Scholar]
- 12.Wootton R J. Symp Zool Soc London. 1979;44:133–159. [Google Scholar]
- 13.Roff D A. Can J Fish Aquat Sci. 1983;40:1395–1404. [Google Scholar]
- 14.Day T, Taylor P D. Am Nat. 1997;149:381–393. [Google Scholar]
- 15.Beverton R J H. Rapp P–V Reun–Cons Int Explor Mer. 1963;154:44–67. [Google Scholar]
- 16.Cushing D H. J Cons, Cons Int Explor Mer. 1975;36:150–157. [Google Scholar]
- 17.Gunderson D R. Can J Fish Aquat Sci. 1997;54:990–998. [Google Scholar]
- 18.Gunderson D R, Dygert P H. J Cons, Cons Int Explor Mer. 1988;44:200–209. [Google Scholar]
- 19.Perrin N, Sibly R. Annu Rev Ecol Syst. 1993;24:379–410. [Google Scholar]
- 20.Gemmill A W, Skorping A, Read A F. J Evol Biol. 1999;12:1148–1156. [Google Scholar]
- 21.Shine R, Charnov E L. Am Nat. 1992;139:1257–1269. [Google Scholar]
- 22.Shine R, Schwarzkopf L. Evolution (Lawrence, Kans) 1992;46:62–75. doi: 10.1111/j.1558-5646.1992.tb01985.x. [DOI] [PubMed] [Google Scholar]
- 23.Winemiller K O, Rose K A. Can J Fish Aquat Sci. 1992;49:2196–2218. [Google Scholar]
- 24.Bart H L, Jr, Page L M. In: Systematics, Historical Ecology, and North American Freshwater Fishes. Mayden R L, editor. Palo Alto, CA: Stanford Univ. Press; 1992. pp. 553–572. [Google Scholar]
- 25.Charnov E L. Evol Ecol Res. 2000;2:365–373. [Google Scholar]
- 26.Charnov E L. Evol Ecol Res. 2001;3:521–535. [Google Scholar]
- 27.Ricklefs R E. Condor. 2000;102:9–22. [Google Scholar]
- 28.Ricklefs R E. Am Nat. 1977;111:453–478. [Google Scholar]
- 29.West G B, Brown J H, Enquist B J. Science. 1997;276:122–126. doi: 10.1126/science.276.5309.122. [DOI] [PubMed] [Google Scholar]
- 30.Charlesworth B. Evolution in Age-Structured Populations. Cambridge, U.K.: Cambridge Univ. Press; 1994. [Google Scholar]
- 31.Bulmer M G. Theoretical Evolutionary Ecology. Sunderland, MA: Sinauer; 1994. [Google Scholar]
- 32.Stearns S C. The Evolution of Life Histories. Oxford: Oxford Univ. Press; 1992. [Google Scholar]