


Abstract
In Arabidopsis a SWI2/SNF2 chromatin remodeling factor-related protein DDM1 and a cytosine methyltransferase MET1 are required for maintenance of genomic cytosine methylation. Mutations in either gene cause global demethylation. In this work we have assessed the effects of these mutations on the PAI tryptophan biosynthetic gene family, which consists of four densely methylated genes arranged as a tail-to-tail inverted repeat plus two unlinked singlet genes. The methylation mutations caused only partial demethylation of the PAI loci: ddm1 had a strong effect on the singlet genes but a weaker effect on the inverted repeat, whereas met1 had a stronger effect on the inverted repeat than on the singlet genes. The double ddm1 met1 mutant also displayed partial demethylation of the PAI genes, with a pattern similar to the ddm1 single mutant. To determine the relationship between partial methylation and expression for the singlet PAI2 gene we constructed a novel reporter strain of Arabidopsis in which PAI2 silencing could be monitored by a blue fluorescent plant phenotype diagnostic of tryptophan pathway defects. This reporter strain revealed that intermediate levels of methylation correlate with intermediate suppression of the fluorescent phenotype.
INTRODUCTION
Many eukaryotes, including mammals and higher plants, modify their genomic DNA by methylation of cytosine residues at the 5 position. The bulk of methylation is found on repetitive sequences, including centromere-associated repeats, rDNA arrays and transposon- or retrotransposon-derived repeats (1,2). However, some methylation is also found in single or low copy regions of the genome. Methylation is associated with suppressed gene expression and suppressed homologous recombination and is thought to have evolved as a defense mechanism against invasive repetitive sequences and/or as a means of regulating gene expression.
Mutational analysis of methylation genes in eukaryotes indicates that multiple methylation systems with different specificities contribute to the overall methylation pattern of the genome. For example, mouse ES cells deleted for the cytosine methyltransferase gene Dnmt1, Dnmt3a or Dnmt3b display distinct methylation changes (3,4). In the higher plant model system Arabidopsis thaliana, two complementation groups of recessive mutants strongly deficient for methylation of repetitive sequences, ddm1 and ddm2, were isolated by a Southern blot methylation assay (5; E.Richards, Washington University, St Louis, personal communication). The DDM1 locus encodes a SWI2/SNF2-related protein and might therefore act by influencing the chromatin structure in methylated regions of the genome (6). The DDM2 locus encodes a cytosine methyltransferase gene MET1 (E.Richards, personal communication), which has sequence identity to the mouse and human Dnmt1 methyltransferases (2,7). The MET1 gene has also been implicated in methylation of repetitive sequences by reverse genetic approaches with antisense transgenes (8,9). Both DDM1- and MET1-deficient strains are viable and fertile, but accumulate a number of morphological defects upon inbreeding, including floral homeotic changes that lead to partial or complete sterility, delayed time to flowering, alteration of leaf shape and reduced size (8–11). These progressive phenotypic changes are likely to be caused, either directly or indirectly, by progressive changes in methylation patterns. In addition, certain floral homeotic changes in ddm1, met1 and antisense MET1 strains have been shown to be due to local hypermethylation of the SUPERMAN gene (12). Hypermethylation of selected genomic regions suggests that DDM1- and MET1-deficient strains retain a de novo methylation system.
In this work we have assessed the effects of the ddm1 and met1 mutations on the methylated endogenous PAI gene family, which encodes the tryptophan biosynthetic enzyme phosphoribosylanthranilate isomerase. In most strains, such as the commonly used laboratory strain Columbia (Col), the PAI gene family consists of three unlinked singlet genes (PAI1, PAI2 and PAI3) that are completely unmethylated. In contrast, in a few strains, such as Wassilewskija (WS), the PAI gene family consists of a tandem inverted repeat duplication of two genes (PAI1-PAI4) and two unlinked singlet genes (PAI2 and PAI3) and all four genes are densely cytosine methylated at both 5′-CG-3′ (CG) and non-5′-CG-3′ (non-CG) cytosines (13–15; Figs 1 and 2 and Table 1). No instability or variability in this dense methylation has been detected in WS tissues grown under standard laboratory conditions. The WS strain is phenotypically normal due to expression of the PAI1 gene in the inverted repeat (14). This expression might be due to the presence of duplicated promoter sequences from another gene fused in an upstream unmethylated region to the PAI1-proximal methylated promoter sequences. The PAI3 and PAI4 genes do not encode functional enzyme and are not expressed and the PAI2 gene, although functional, is silenced by cytosine methylation.
Figure 1.
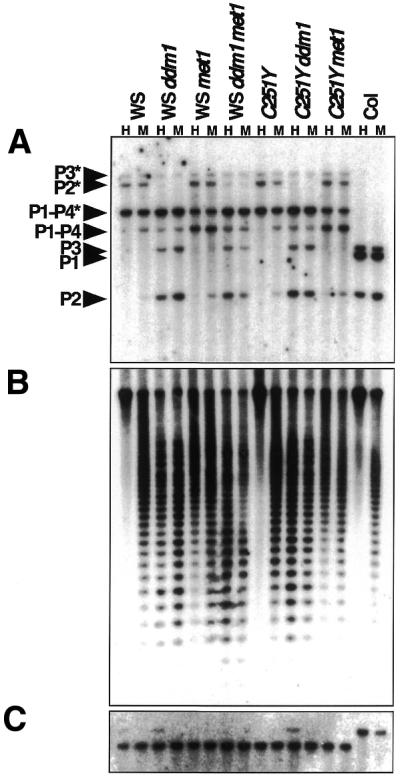
(A) PAI internal methylation patterns in methylation mutant strains. A genomic Southern blot of DNA prepared from whole 4-week-old plants of the indicated strains cleaved with either HpaII (H) or MspI (M) and probed with an internal PAI probe is shown. The WS ddm1 and WS met1 DNAs were extracted from a representative line of each strain inbred for six generations in the presence of the homozygous mutation. The WS ddm1 met1 DNA was extracted from a representative line that was inbred for one generation in the presence of both homozygous mutations. The C251Y (WS pai1C251Y) ddm1 and C251Y met1 DNAs were extracted from a representative line of each strain inbred for two generations in the presence of the homozygous mutation. The MspI restriction maps of the WS and Col PAI genes were previously described (13–15). The band sizes predicted for the PAI loci (P1–P4 is WS PAI1-PAI4, P1 is Col PAI1, P2 is WS or Col PAI2, P3 is WS or Col PAI3) are indicated in the left margin, with the fully cleaved species unmarked and the internally methylated species marked with an asterisk. (B) Centromere repeat methylation patterns. The same blot shown in (A) was stripped and reprobed with a fragment that detects the 180 bp centromere repeats. (C) Digestion pattern of the ASA1 gene. The same blot shown in (A) was stripped and reprobed with an ASA1 (anthranilate synthase α subunit 1) cDNA probe. Because the ASA1 region carries very little methylation, HpaII and MspI give similar restriction patterns. The probe detects a 4.9 kb band in WS and a 5.9 kb band in Col due to an MspI polymorphism. The ASA1 blot thus controls for equal loading and complete digestion of DNA samples
Figure 2.

PAI promoter methylation patterns in methylation mutant strains. Bisulfite genomic sequencing of methylation patterns was performed for the top and bottom strands of the indicated PAI gene promoters in a representative line of four generation inbred WS ddm1 and a representative line of four generation inbred WS met1, with eight independent molecules sequenced for each strand. Vertical lines indicate the positions of cytosines, with the height of each line representing how many sequenced molecules had 5-methylcytosine (5-MeC) at that position. Black indicates cytosines in the context CG, blue indicates cytosines in the context CNG and red indicates cytosines in other contexts. Asterisks indicate sites where none of the sequenced molecules had a 5-MeC. The black horizontal line indicates the region of PAI identity and the gray horizontal line indicates flanking upstream heterologous sequence unique to each gene. For the purposes of comparison our previously published sequencing data for the wild-type WS PAI1 and PAI2 promoter regions are also shown; these data are reprinted from Luff et al. (15) with permission from Elsevier Science.
Table 1. Effects of methylation mutations on patterns of PAI promoter cytosine methylationa.
| Strain |
PAI gene |
CMeG |
CMeNG |
Other CMe |
Total CMe |
| WS | PAI1 | 236 (100%) | 127 (100%) | 298 (100%) | 661 (100%) |
| WSddm1 | PAI1 | 201 (85%) | 104 (81%) | 176 (60%) | 481 (73%) |
| WSmet1 | PAI1 | 61 (26%) | 52 (41%) | 142 (48%) | 255 (39%) |
| WS | PAI2 | 245 (100%) | 112 (100%) | 341 (100%) | 698 (100%) |
| WSddm1 | PAI2 | 123 (50%) | 27 (24%) | 76 (22%) | 226 (32%) |
| WS met1 | PAI2 | 131 (53%) | 44 (39%) | 136 (40%) | 311 (45%) |
aNumbers represent the indicated type of methylated cytosines for eight independent top strand plus eight independent bottom strand bisulfite sequencing clones for the indicated PAI promoter region of the indicated strain. The same data are shown in diagram form in Figure 2.
The WS PAI1-PAI4 inverted repeat locus can trigger de novo methylation of an unmethylated PAI2 gene crossed in from a different strain background (15). Moreover, when the PAI1-PAI4 inverted repeat is removed from the genome, either by deletion mutation or by segregation in crosses with a strain that carries a singlet PAI1 gene at this locus, methylation on the PAI2 gene is reduced in density and persists mainly at CG cytosines (13,15,16). Thus, the PAI1-PAI4 locus provides a signal for the dense methylation of PAI sequences.
In previous work we found that the ddm1 mutation can strongly hypomethylate the CG-methylated PAI2 gene in a Δpai1-pai4 strain background after a few generations of inbreeding (16). In this work we show that the ddm1 and met1 mutations, either singly or combined, reduce but do not eliminate PAI methylation when crossed into the WS strain background. These results suggest that the presence of the PAI1-PAI4 inverted repeat opposes the effects of the methylation mutations. We also describe a novel reporter strain derived from WS where the dense methylation and silencing of the PAI2 gene can be visualized by tryptophan deficiency phenotypes, including blue fluorescence under UV light due to accumulation of early tryptophan biosynthetic intermediates. Using this reporter strain we show that ddm1 and met1 mutations can partially relieve PAI2 silencing, consistent with their partial effects on PAI2 methylation.
MATERIALS AND METHODS
Plant strain construction
The WS ddm1 strain was made by crossing the Columbia ddm1-2 allele with wild-type WS and genotyping F2 progeny plants with polymorphic markers at PAI1, PAI2, PAI3 and m555, which lies ∼1 cM away from the DDM1 locus (http://www.arabidopsis.org/aboutcaps.html). All ddm1 single mutant genotypes were confirmed by PCR amplification of the gene with a primer pair that creates a restriction site for RsaI just beyond the end of one primer on a wild-type DDM1 template but not on a ddm1-2 mutant template due to the presence of the ddm1-2 mutation (DDM1-2F, 5′-GTTGGACAGTGTGGTAAATTCCGCT-3′; DDM1-2R, 5′-GAGCTACGAGCCATGGGTTTGTGAAACGTA-3′; where the underlined base represents a mismatch with the DDM1 gene that creates an RsaI site). The WS met1 strain was made by crossing the ddm2-1 allele, which was originally isolated in the Columbia background and then introgressed by crossing three times into the Landsberg erecta strain background, with wild-type WS. F2 progeny plants were genotyped with the PAI gene polymorphic markers and with a polymorphism created by the ddm2-1 mutation. Specifically, primers that flank the ddm2-1 mutation, MTGF (5′-CCGGTGCATATGGAGTATCCC-3′) and MTGR (5′-GCATCGAATGAGGTTAAGCTC-3′), were used to PCR amplify this region from plant genomic DNA and the PCR products were cleaved with HaeIII, which cuts once in the wild-type product but fails to cleave in the mutant product due to the ddm2-1 base change. For both WS ddm1 and WS met1 four independent lines were followed for methylation changes by Southern blot assay for four generations of inbreeding and no significant differences among lines were detected. One representative line of each construct was selected for additional inbreeding, bisulfite genomic sequencing and crossing to generate the WS ddm1 met1 double mutant lines.
For double mutant construction, several hundred F2 progeny from a cross of WS ddm1 × WS met1 were grown in soil until the flowering stage and individuals displaying floral abnormalities and sterility were discarded. The remaining 86 fertile plants were scored for ddm1 and met1 genotypes with the m555 and MTG markers, which both lie on the lower arm of chromosome 5. This analysis yielded 19 plants that were homozygous for ddm1 but heterozygous for met1, eight plants that were homozygous for met1 but heterozygous for ddm1 and no plants that were double homozygous mutants. To generate double homozygous mutants one ddm1 homozygous met1 heterozygous fertile F2 individual and one ddm1 heterozygous met1 homozygous fertile F2 individual were allowed to self-pollinate and the segregating F3 progeny were scored for ddm1 and met1 genotypes. Double homozygous progeny from both the ‘ddm1 first, met1 second’ and the ‘met1 first, ddm1 second’ lineages were inbred for one generation and DNA was prepared from second generation progeny plants for Southern blot analysis. PAI and centromere methylation patterns were identical for both types of lineages. DNA from the ‘met1 first, ddm1 second’ lineage is shown in Figure 1.
The WS pai1C251Y strain was made by EMS mutagenesis of WS using standard methods (17), followed by screening ∼70 000 M2 progeny plants for blue fluorescence as described (13). Fluorescent isolates caused by spontaneous deletion of the PAI1-PAI4 inverted repeat (13) were identified by DNA analysis and discarded. The remaining fluorescent isolates were tested by complementation crosses with the WS Δpai1-pai4 fluorescent mutant. The WS pai1C251Y allele was further characterized by cloning and sequencing the PAI1 gene. The strain was backcrossed twice to wild-type WS before performing methylation and double mutant analysis. Another pai1 missense mutation isolated from the screen, pai1G226E, had more severe growth and fertility defects than pai1C251Y and therefore was not pursued as a reporter strain.
Seedlings were germinated on plant nutrient plus sucrose medium (18) under continuous illumination. Adult plants were grown in Scotts MetroMix 360 potting medium under continuous illumination.
DNA and RNA analysis
Plant genomic DNA for Southern blot and bisulfite genomic sequencing methylation analysis and plant RNA for northern blot analysis were prepared as previously described (14). The PAI probe is a 0.7 kb internal PstI fragment isolated from a Col PAI1 cDNA (13). The centromere 180 bp repeat probe was isolated as a HindIII fragment from plasmid pARR20-1 (a gift of E.Richards). Bisulfite genomic sequencing was performed as previously described (16). Briefly, 10 µg DNA was cleaved with XhoI to separate the two halves of the PAI1-PAI4 inverted repeat, mutagenized with sodium bisulfite and tested for efficiency of mutagenesis by analysis of the unmethylated ASA1 gene. Eight independent PCR products for each region (PAI1 top strand, PAI1 bottom strand, PAI2 top strand, PAI2 bottom strand) were cloned into pBlueScript KS II+ (Stratagene) for sequencing. Primer sequences for the PAI2 bottom strand were as previously described (16) and primer sequences for the other amplified regions are available upon request from J.Bender.
RESULTS
PAI genes are partially hypomethylated in WS ddm1 and WS met1 strains
The Arabidopsis hypomethylation mutations ddm1 and met1 were isolated in the Col strain background, which lacks a PAI inverted repeat gene structure and PAI methylation (13,14). The mutations were therefore crossed into the WS background to assess effects on PAI methylation. Polymorphisms associated with each locus were used to identify plants that were homozygous for the relevant methylation mutation from Col and the PAI1-PAI4, PAI2 and PAI3 loci from WS (WS ddm1 and WS met1). The ddm1 allele ddm1-2 creates a splice site mutation that results in misprocessing of the DDM1 transcript and is likely to be a null allele (6). The met1 allele ddm2-1 is a missense mutation in an unconserved position of the cytosine methyltransferase catalytic domain (E.Richards, personal communication).
PAI methylation in WS ddm1 and WS met1 strains was monitored by a HpaII/MspI Southern blot assay over six generations of inbreeding. The restriction enzymes HpaII and MspI are isoschizomers that recognize the sequence 5′-CCGG-3′; HpaII is sensitive to methylation of either the inner or the outer cytosine of the sequence, whereas MspI is sensitive only to methylation of the outer cytosine of the sequence. Each PAI locus is cleaved once at a conserved HpaII/MspI site in the second intron and at flanking unmethylated sites that lie at different distances away from the central site for each locus (13–15). Thus, each PAI locus gives unique fragment sizes both when fully cleaved (diagnostic of no PAI methylation) or when uncleaved (diagnostic of PAI methylation). Previous analyses showed that the HpaII/MspI methylation correlates with methylation patterns over the rest of the PAI sequences for wild-type WS (13,15).
The HpaII/MspI assay revealed that the ddm1 mutation had only a weak effect on methylation of the PAI1-PAI4 inverted repeat locus, but had a strong hypomethylation effect on the singlet PAI2 and PAI3 genes (Fig. 1A). This basic pattern was established by the second generation, although the PAI2 and PAI3 genes became progressively less methylated over four subsequent generations of inbreeding. The met1 mutation had an intermediate hypomethylation effect on the inverted repeat PAI1-PAI4 locus, but had a weaker effect on the PAI2 and PAI3 genes. Note that because the PAI3 gene is divergent in sequence from the probe fragment (13,16), this locus is not as strongly detected as PAI1-PAI4 and PAI2 in hybridization experiments. Thus, the hypomethylated PAI3 species was only detectable on overexposure of the Figure 1A blot (data not shown). The PAI methylation pattern shown in Figure 1A remained constant over the entire WS met1 inbreeding regime monitored from the second generation onwards.
Both WS ddm1 and WS met1 inbred lines progressively accumulated a number of morphological defects and reduced fertility, as previously observed in the Col strain background (9–11). In particular, the most inbred WS ddm1 line developed flowers with unfused carpels and the most inbred WS met1 line was late flowering and displayed a number of floral abnormalities (data not shown).
We also crossed an antisense MET1 transgene (8) into the WS strain background and monitored effects on PAI methylation by Southern blot assay. The antisense MET1 transgene had a similar effect to the met1 missense mutation on WS PAI gene methylation in second generation DNA (data not shown). Because our antisense transgene-carrying lines became completely sterile within two generations of inbreeding we did not pursue analysis of these lines.
We also constructed a ddm1 met1 double mutant in the WS strain background (WS ddm1 met1) by crossing together WS ddm1 and WS met1 lines and using polymorphisms associated with the methylation mutations to identify double mutant recombinants. A majority of plants in the segregating population from this cross were late flowering and/or sterile, presumably due to accumulation of methylation changes during the long inbreeding regime of the parental strains (see above). However, we were able to recover two independent double mutant individuals that were fertile when newly segregated. The double mutants had a number of morphological defects and became completely sterile by the second generation. We prepared DNA from the second generation progeny of each double mutant lineage and analyzed PAI methylation by Southern blot. This analysis showed that the WS ddm1 met1 double mutants displayed strong hypomethylation of PAI2 and PAI3 but weak hypomethylation of the PAI1-PAI4 locus, similarly to the ddm1 single mutant (Fig. 1A). Thus, the combined methylation mutations were not sufficient to remove PAI methylation after two generations of inbreeding.
The 180 bp centromere-associated repeats were strongly hypomethylated in the WS methylation mutant hybrid strains (Fig. 1B), consistent with previous analyses in the Col and C24 strain backgrounds (5,8,9). The centromere repeat hypomethylation was stronger in the ddm1 and ddm1 met1 mutant backgrounds than in the met1 mutant background, as estimated from the pattern of cleavage with HpaII.
The ddm1 and met1 mutations reduce both symmetrical and asymmetrical PAI methylation
To understand the effects of ddm1 and met1 mutations on PAI methylation at the sequence level we performed sodium bisulfite genomic sequencing on the promoter regions of the PAI1 inverted repeat gene and the PAI2 singlet gene in genomic DNA prepared from four generation inbred plants. In this method genomic DNA is treated with sodium bisulfite under conditions where unmethylated cytosines are deaminated to uracil but methylated cytosines remain unreacted (19). The genomic region of interest is then amplified by PCR and C→T mutations in the PCR products indicate the positions of unmethylated residues.
In WS ddm1 we found that the methylation patterns for the PAI1 promoter at the inverted repeat locus were similar to those previously determined by sequencing the same region in parental WS (15): within the region of PAI sequence identity symmetrical CG and CNG as well as asymmetrical cytosines were methylated and there was no significant spread of methylation into upstream heterologous sequences (Fig. 2). The primary difference between WS and WS ddm1 PAI1 methylation patterns was that the WS ddm1 methylation density was 73% of that found in WS (Table 1). In contrast, for the singlet PAI2 gene in WS ddm1 methylation was reduced to 32% of parental WS levels. Note that this degree of hypomethylation is likely to be an underestimate of the ultimate effect of ddm1 on PAI2, because HpaII/MspI Southern blot analysis revealed a progressive loss of PAI2 methylation between the fourth generation of inbreeding, when the sequencing was performed, and the sixth generation of inbreeding (shown in Fig. 1A). For both PAI1 and PAI2 sequences the ddm1 mutation reduced both symmetrical (CG and CNG) and asymmetrical cytosine methylation, although there was a stronger effect on non-CG methylation (Table 1).
In WS met1 both the PAI1 and PAI2 promoters had <50% residual methylation relative to parental WS (Fig. 2 and Table 1). As observed for ddm1, methylation of both symmetrical and asymmetrical cytosines was affected by the met1 mutation. We were unable to perform PAI bisulfite sequencing on the WS ddm1 met1 and WS antisense MET1 strains due to the low amounts of DNA recovered from these morphologically aberrant and sterile strains.
The bisulfite sequencing patterns were generally consistent with the WS ddm1 HpaII/MspI digestion patterns, showing weak hypomethylation at the PAI1-PAI4 inverted repeat locus and stronger hypomethylation at the singlet PAI2 locus, and with the WS met1 patterns, showing partially reduced methylation for both PAI1-PAI4 and PAI2 (Fig. 1A). However, the bisulfite sequencing revealed a stronger effect of met1 on the PAI2 gene, with ∼45% residual methylation, than was indicated by the Southern blot analysis. This discrepancy could reflect a different effect of the met1 mutation near the methylation boundary in the sequenced promoter region versus the internal HpaII/MspI site.
PAI steady-state transcript levels are not dramatically altered by ddm1 or met1 mutations
In wild-type WS the PAI1 transcript is the predominant species detected by northern blot, cDNA analysis or RT–PCR approaches and provides sufficient PAI enzyme so that WS is a phenotypically normal strain (13,14). We used the WS ddm1 and WS met1 strains to determine whether this PAI expression profile could be significantly altered by reduced methylation. From previous studies in strains that lack PAI1-PAI4 we knew that PAI3 has very low expression regardless of methylation state (14) and that PAI2 can be reactivated up to 30% of the WS PAI1 steady-state transcript levels by strong demethylation (13). However, we did not know whether PAI1 or PAI4 expression levels would be affected by methylation changes.
We monitored PAI expression levels in WS methylation mutant strains versus parental WS by northern blot analysis of 4-week-old whole plant RNA with a PAI1 cDNA probe which can detect all of the highly identical PAI sequences (Fig. 3). In wild-type WS the signal detected by this probe is >90% PAI1 transcripts (14). The signal level was only slightly increased in the methylation mutant strains, indicating that overall PAI expression is not dramatically altered by reduced methylation. In order to determine which PAI genes might contribute to the slight changes in expression we performed RT–PCR reactions on mutant RNA templates with primer pairs that discriminate PAI1 and PAI4 transcripts from PAI2 and PAI3 transcripts via a restriction site polymorphism or that discriminate PAI4 transcripts from other PAI transcripts via a length polymorphism (14). We were only able to detect PAI1 species by these assays (data not shown). However, based on control reactions, we estimate that the RT–PCR primers can only reliably detect PAI transcript species that are present at ≥10% of the template population. Thus, it was inconclusive as to which PAI genes might be slightly up-regulated by partial demethylation using these assays.
Figure 3.
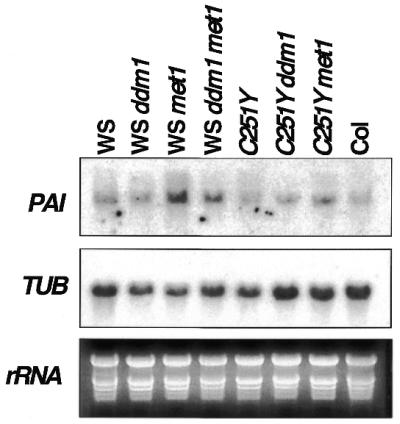
PAI steady-state transcript levels are not dramatically perturbed by methylation mutation-induced hypomethylation. Total RNA was prepared from 4-week-old plants of the same strains described in Figure 1. Duplicate northern blots prepared from these samples were probed with either the PAI cDNA 0.7 kb internal PstI fragment (3 day exposure) or with a β-tubulin probe (TUB) as a control for gel loading (16 h exposure). The ethidium bromide stained gel of the PAI northern blot (rRNA) is also shown as a control for gel loading.
Hypomethylation by the ddm1 and met1 mutations activates PAI2 expression
Previous northern blot analysis of WS PAI2 expression in a strain background where the PAI1-PAI4 genes are deleted and where the residual methylation of PAI2 is reduced and destabilized indicated that the partially methylated PAI2 gene has only ∼10% of the steady-state PAI transcript levels observed for parental WS (13). Thus, increased expression from a partially methylated PAI2 gene in methylation mutant lines would not be detected by northern blot or RT–PCR over the strong signal from PAI1. As a more sensitive means of following WS PAI2 expression we isolated a derivative of WS that displayed a number of PAI-deficient phenotypes due to a missense mutation in the PAI1 gene. In this strain background changes in PAI expression are easily monitored as changes in pai mutant phenotypes.
The pai1 missense mutant reporter strain WS pai1C251Y was strongly blue fluorescent under UV light due to the accumulation of early intermediates in the tryptophan pathway (20,21) and had yellow-green leaf pigmentation, reduced size and increased bushiness relative to parental WS (Fig. 4). However, the strain was viable and fertile even on unsupplemented agar medium, indicating that there was sufficient residual PAI enzyme produced from the mutant pai1 gene and/or the silenced PAI2 gene, so that the strain was not a tryptophan auxotroph. Neither PAI methylation nor PAI gene expression were significantly affected by the point mutation in the WS pai1C251Y strain (Figs 1 and 3). The strong blue fluorescence phenotype of the mutant was completely stable in thousands of plants analyzed over several generations of inbreeding, indicating that WS pai1C251Y does not segregate progeny with spontaneously hypomethylated and activated PAI genes at a significant frequency.
Figure 4.

The methylation mutants suppress PAI-deficient phenotypes of the WS pai1C251Y reporter strain. (A) Representative 2-week-old seedlings of the indicated genotypes photographed under visible (left) and UV (right) light are shown. (B) Representative 4-week-old plants of the indicated genotypes photographed under visible (left) and UV (right) light are shown. The C251Y ddm1 and C251Y met1 plants are from the F5 generation, the same generation used to prepare DNA and RNA for molecular analysis (Figs 1 and 3).
To monitor effects of ddm-induced hypomethylation on PAI expression we crossed the WS pai1C251Y fluorescent reporter strain with WS ddm1 or WS met1 and used the fluorescence phenotype plus polymorphic markers at each methylation mutation to identify double mutant plants. Specifically, to assess the effects of the methylation mutations in the pai1C251Y background without the complication of lingering epigenetic changes inherited from the ddm1 and met1 parents, we identified two F2 individuals from each cross that were phenotypically identical to the parental pai1C251Y mutant and heterozygous for the relevant methylation mutation. We then analyzed segregation of fluorescence phenotype versus ddm1 or met1 genotype in a sampling of F3 progeny of each cross (Fig. 5).
Figure 5.

Genetic pedigrees used to analyze the effects of ddm1 and met1 on the WS pai1C251Y reporter strain. The circled X symbol indicates self-pollination. In the F4 generation phenotypes were assigned based on the average behavior of a population of 24 plants. sf indicates a strong fluorescent phenotype equivalent to that of the parental pai1C251Y strain. wf indicates a weaker fluorescent phenotype than displayed by the parental pai1C251Y strain.
For the pai1C251Y ddm1 cross we found that newly segregated double mutant F3 individuals had phenotypes ranging from strong fluorescence and morphology equivalent to that of the parental pai1C251Y mutant to partial fluorescence with increased plant size and fertility (Fig. 5). In contrast, for the pai1C251Y met1 cross all of the newly segregated double mutant F3 individuals had a strong fluorescent phenotype. These results suggest that the ddm1 mutation has a variable partial effect on PAI expression and that met1 has little or no effect when newly segregated.
To assess effects of the methylation mutations in the second generation of homozygosity, 24 F4 progeny for each of a number of representative F3 double mutant individuals were scored for fluorescence and morphology relative to the parental WS pai1C251Y strain (Fig. 5). For pai1C251Y ddm1 all the members of each F4 family displayed strongly suppressed pai phenotypes, including extremely weak fluorescence and increased size and fertility (Fig. 4), regardless of the suppressed phenotype of the parental F3 plant. For pai1C251Y met1 all the members of each F4 family displayed partially suppressed fluorescence with increased size and fertility. These results indicate that the ddm1 and met1 mutations can progressively suppress the PAI deficiencies of the WS pai1C251Y strain and that after one generation of inbreeding there is uniform suppression. These results also suggest that ddm1 has a stronger effect than met1, consistent with the relative effects of the two mutations on PAI2 methylation density (Figs 1 and 2 and Table 1). Similar progressive effects of the ddm1 mutation on PAI2 silencing were previously observed in a derivative of WS where the PAI1-PAI4 locus is deleted and the PAI2 gene carries reduced residual methylation (16).
Both the pai1C251Y ddm1 and the pai1C251Y met1 lines were inbred for a second generation (F5 generation) to determine if there were additional progressive effects on the fluorescence phenotype (Fig. 4). Double mutant plants in this generation displayed little or no phenotypic difference from the previous generation, indicating that after the first generation of inbreeding the effects of the methylation mutations on PAI silencing are largely complete. This result is consistent with our observation that methylation patterns as monitored from the first generation of inbreeding onwards in the WS ddm1 and WS met1 lines were largely unchanged (see above).
Genomic DNA was prepared from representative WS pai1C251Y ddm1 or met1 homozygous double mutant F5 tissue for methylation analysis of the PAI genes. Southern blot assays with HpaII and MspI revealed that the ddm1 and met1 mutations had the same effects on methylation in the WS pai1C251Y mutant background as when crossed into wild-type WS (Fig. 1A). Northern blot analysis revealed that, as in the WS background, the ddm1 and met1 mutations did not dramatically alter the steady-state levels of PAI transcripts in the WS pai1C251Y background (Fig. 3). Because the changes in pai phenotypes correlate with the degree of PAI2 hypomethylation (stronger in ddm1 than in met1) rather than with the degree of PAI1 hypomethylation (stronger in met1 than in ddm1) (Figs 1, 2 and 4 and Table 1), it is most likely that the phenotypic changes arise primarily from activation of PAI2 expression.
DISCUSSION
Studies of the methylation-deficient mutations ddm1 and met1 in Arabidopsis indicated that these mutations confer strongly reduced methylation of most genomic sequences (5,8–10). In contrast, we found that these mutations, either singly or combined, confer only partial hypomethylation of the methylated PAI gene family. A unique feature of the PAI family is that two of the genes, PAI1-PAI4, are arranged as an inverted repeat. In previous work we showed that the PAI1-PAI4 locus triggers de novo methylation of PAI sequences and that removal of this locus from the genome results in reduced methylation on unlinked PAI genes (13,15,16). Thus, we favor the view that the PAI genes resist hypomethylation by the ddm1 and met1 mutations because of an opposing methylation-stimulatory effect from the inverted repeat locus.
The residual partial methylation on the PAI genes in the ddm1 and met1 strains gave us a unique opportunity to examine patterns of cytosine methylation promoted by other methylation activities. For both mutations we found that CG and non-CG methylation persist, indicating that secondary methylation systems have both types of substrate specificities. To date no other Arabidopsis methylation functions have been described based on genetic analysis, but several candidate methyltransferase genes have been identified in the genome sequence, including close homologs of MET1 (22,23), chromomethylase genes carrying a chromo domain motif in the predicted catalytic region of the protein (22–25) and domain-rearranged methyltransferases with an altered order of catalytic motifs in the primary amino acid sequence and homology to the mammalian Dnmt3 family (26).
Our analysis showed that each methylation mutation had a different effect on PAI1-PAI4 inverted repeat versus singlet PAI gene methylation (Figs 1 and 6). The ddm1 mutation, in a chromatin remodeling factor-related protein (6), had a strong effect on singlet gene methylation but a weaker effect on inverted repeat methylation. The met1 mutation, in a cytosine methyltransferase (E.Richards, personal communication), had a stronger effect on the inverted repeat than on the singlet PAI genes. Thus, even though all three wild-type WS PAI loci carry uniform dense methylation (15), the three loci are differentially demethylated by ddm1 and met1. This observation suggests that methylation determinants for residual methylation systems in each mutant background vary with each PAI locus. Methylation determinants are likely to include specific chromatin features unique to each locus that contribute to targeting of methylation proteins and/or accessibility of the substrate DNA. Our analysis also showed that the overall PAI methylation patterns conferred by the two mutations are different from each other. This observation suggests that DDM1 and MET1 act at least partially independently.
Figure 6.

A summary of methylation changes on the PAI genes in various mutant backgrounds. The organization of the four PAI genes in WS is shown. The arrows depict the direction of transcription for each gene, with the hatch marks between the PAI1-PAI4 locus and the PAI3 locus indicating that these loci are on the same chromosome but genetically unlinked. The Δ symbol represents deletion of the PAI1-PAI4 genes. The boxes around each gene locus indicate cytosine methylation. The approximate density of methylation at each locus is indicated as follows: a solid bold line indicates parental WS levels of methylation, a bold dashed line indicates a <50% reduction in methylation, a fine dashed line indicates a >50% reduction in methylation and no line indicates only trace levels or no detectable methylation. Density indications are based on combined results from HpaII/MspI Southern blot and promoter bisulfite sequencing analyses, except for the WS ddm1 met1 strain, which is based solely on Southern blot analysis.
For two WS ddm1 met1 double mutant lineages that we examined we found residual PAI methylation by Southern blot assay of second generation DNA (Figs 1 and 6). This result indicates that at least some of the residual methylation systems in Arabidopsis can act independently of ddm1 and met1. We also found that the pattern of partial methylation in both lineages was similar to that of the ddm1 single mutant, with a high level of residual methylation on the inverted repeat and much lower levels on the singlet PAI genes. This result suggests a complex epistatic relationship between ddm1 and met1 rather than a simple additive effect of the two mutations, which would have been expected to yield plants with low residual methylation on all three PAI loci. For example, because methylation of the inverted repeat is apparently higher in the ddm1 met1 double mutant than in the met1 single mutant, it could be postulated that a secondary methylation system with specificity for the inverted repeat is selectively activated in the presence of the ddm1 mutation. However, there are two alternative explanations for the apparently epistatic pattern of double mutant methylation. First, the Southern blot assay, which only monitors one site in each PAI locus, might not reflect methylation changes over the rest of the PAI sequences. Secondly, it is possible that the two double mutant lineages we examined happened to inherit and maintain ddm1-modified PAI loci from their ddm1 parent.
The WS pai1C251Y blue fluorescent mutant strain described here provides a facile reporter for methylation and transcriptional silencing of the PAI2 gene. Using this reporter strain we showed that loss of PAI2 silencing is progressive for both the ddm1 and met1 mutations, so that newly segregated homozygous mutants have stronger fluorescence phenotypes than their first generation progeny plants (Figs 4 and 5). In both cases progressive loss of PAI2 silencing is likely to reflect impaired but not abolished maintenance methylation activity. For the ddm1 mutation maintenance methylation might be impaired but not abolished as an indirect consequence of chromatin changes induced by loss of DDM1 function; for the met1 mutation, maintenance methylation might be impaired but not abolished due to a partial loss-of-function allele and/or due to redundant methyltransferases. The progressive relief of PAI2 silencing by ddm1 in the presence of the PAI1-PAI4 inverted repeat is similar to the effect we previously observed in the Δpai1-pai4 background. However, ddm1 ultimately has a stronger effect in the deletion mutant strain, where it confers almost complete loss of PAI2 promoter methylation after just two generations of inbreeding (16), compared to 32% residual methylation after four generations of inbreeding (Table 1 and Fig. 6). This difference is likely to reflect the methylation-stimulatory effects of the PAI1-PAI4 locus acting in opposition to ddm1-induced hypomethylation, as discussed above.
In the WS pai1C251Y background the density of methylation on PAI2 (Fig. 1) corresponds to the severity of the PAI-deficient blue fluorescent phenotype (Fig. 4). Specifically, the strong hypomethylation of PAI2 by ddm1 is reflected in a weak fluorescent phenotype and the partial hypomethylation of PAI2 by met1 is reflected in an intermediate fluorescent phenotype. Beyond the analysis of ddm1 and met1 mutations the pai1C251Y strain promises to be a powerful tool in explicit analysis of other Arabidopsis mutations that are implicated in methylation, chromatin structure and gene silencing. Furthermore, the pai1C251Y strain provides a genetic screening system for novel mutations that disrupt PAI2 methylation and silencing.
Acknowledgments
ACKNOWLEDGEMENTS
We thank Dr Eric Richards for providing the ddm1-2 and ddm2-1 mutant strains, the pARR20-1 centromere repeat probe plasmid and information regarding detection of the sequence alterations in the ddm1-2 and ddm2-1 alleles. We also thank Dr E. Jean Finnegan for providing the antisense MET1 transgenic strain. This work was supported by March of Dimes grant 1-FY00-418 and a Searle Scholars Award 97-E-103 to J.B. L.B. was supported by a training grant from the National Cancer Institute, 5 T32 CA09110.
References
- 1.Yoder J.A., Walsh,C.P. and Bestor,T.H. (1997) Cytosine methylation and the ecology of intragenomic parasites. Trends Genet., 13, 335–340. [DOI] [PubMed] [Google Scholar]
- 2.Finnegan E.J., Genger,R.K., Peacock,W.J. and Dennis,E.S. (1998) DNA methylation in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol., 49, 223–247. [DOI] [PubMed] [Google Scholar]
- 3.Lei H., Oh,S.P., Okano,M., Juttermann,R., Goss,K.A., Jaenisch,R. and Li,E. (1996) De novo cytosine methyltransferase activities in mouse embryonic stem cells. Development, 122, 3195–3205. [DOI] [PubMed] [Google Scholar]
- 4.Okano M., Bell,D.W., Haber,D.A. and Li,E. (1999) DNA methyltransferases Dnmt3a and Dnmt3b are essential for de novo methylation and mammalian development. Cell, 99, 247–257. [DOI] [PubMed] [Google Scholar]
- 5.Vongs A., Kakutani,T., Martienssen,R.A. and Richards,E.J. (1993) Arabidopsis thaliana DNA methylation mutants. Science, 260, 1926–1928. [DOI] [PubMed] [Google Scholar]
- 6.Jeddeloh J.A., Stokes,T.L. and Richards,E.J. (1999) Maintenance of genomic methylation requires a SWI2/SNF2-like protein. Nat. Genet., 22, 94–97. [DOI] [PubMed] [Google Scholar]
- 7.Finnegan E.J. and Dennis,E.S. (1993) Isolation and identification by sequence homology of a putative cytosine methyltransferase from Arabidopsis thaliana. Nucleic Acids Res., 21, 2383–2388. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Finnegan E.J., Peacock,W.J. and Dennis,E.S. (1996) Reduced DNA methylation in Arabidopsis thaliana results in abnormal plant development. Proc. Natl Acad. Sci. USA, 93, 8449–8454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Ronemus M.J., Galbiati,M., Ticknor,C., Chen,J. and Dellaporta,S.L. (1996) Demethylation-induced developmental pleiotropy in Arabidopsis. Science, 273, 654–657. [DOI] [PubMed] [Google Scholar]
- 10.Kakutani T., Jeddeloh,J.A., Flowers,S.K., Munakata,K. and Richards,E.J. (1996) Developmental abnormalities and epimutations associated with DNA hypomethylation mutations. Proc. Natl Acad. Sci. USA, 93, 12406–12411. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Richards E.J. (1997) DNA methylation and plant development. Trends Genet., 13, 319–323. [DOI] [PubMed] [Google Scholar]
- 12.Jacobsen S.E., Sakai,H., Finnegan,E.J., Cao,X. and Meyerowitz,E.M. (2000) Ectopic hypermethylation of flower-specific genes in Arabidopsis. Curr. Biol., 10, 179–186. [DOI] [PubMed] [Google Scholar]
- 13.Bender J. and Fink,G.R. (1995) Epigenetic control of an endogenous gene family is revealed by a novel blue fluorescent mutant of Arabidopsis. Cell, 83, 725–734. [DOI] [PubMed] [Google Scholar]
- 14.Melquist S., Luff,B. and Bender,J. (1999) Arabidopsis PAI gene arrangements, cytosine methylation and expression. Genetics, 153, 401–413. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Luff B., Pawlowski,L. and Bender,J. (1999) An inverted repeat triggers cytosine methylation of identical sequences in Arabidopsis. Mol. Cell, 3, 505–511. [DOI] [PubMed] [Google Scholar]
- 16.Jeddeloh J.A., Bender,J. and Richards,E.J. (1998) The DNA methylation locus DDM1 is required for maintenance of gene silencing in Arabidopsis. Genes Dev., 12, 1714–1725. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Niyogi K.K., Last,R.L., Fink,G.R. and Keith,B. (1993) Suppressors of trp1 fluorescence identify a new Arabidopsis gene, TRP4, encoding the anthranilate synthase β subunit. Plant Cell, 5, 1011–1027. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Haughn G.W. and Somerville,C. (1986) Sulfonylurea-resistant mutants of Arabidopsis thaliana. Mol. Gen. Genet., 204, 430–434. [Google Scholar]
- 19.Frommer M., McDonald,L.E., Millar,D.S., Collis,C.M., Watt,F., Grigg,G.W., Molloy,P.L. and Paul,C.L. (1992) A genomic sequencing protocol that yields a positive display of 5-methylcytosine residues in individual DNA strands. Proc. Natl Acad. Sci. USA, 89, 1827–1831. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Last R.L. and Fink,G.R. (1988) Tryptophan-requiring mutants of the plant Arabidopsis thaliana. Science, 240, 305–310. [DOI] [PubMed] [Google Scholar]
- 21.Li J., Zhao,J., Rose,A.B., Schmidt,R. and Last,R.L. (1995) Arabidopsis phosphoribosylanthranilate isomerase: molecular genetic analysis of triplicate tryptophan pathway genes. Plant Cell, 7, 447–461. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Genger R.K., Kovac,K.A., Dennis,E.S., Peacock,W.J. and Finnegan,E.J. (1999) Multiple DNA methyltransferase genes in Arabidopsis thaliana. Plant Mol. Biol., 41, 269–278. [DOI] [PubMed] [Google Scholar]
- 23.Finnegan E.J. and Kovac,K.A. (2000) Plant DNA methyltransferases. Plant Mol. Biol., 43, 189–201. [DOI] [PubMed] [Google Scholar]
- 24.Henikoff S. and Comai,L. (1998) A DNA methyltransferase homolog with a chromodomain exists in multiple polymorphic forms in Arabidopsis. Genetics, 149, 307–318. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.McCallum C.M., Comai,L., Greene,E.A. and Henikoff,S. (2000) Targeted screening for induced mutations. Nat. Biotechnol., 18, 455–457. [DOI] [PubMed] [Google Scholar]
- 26.Cao X., Springer,N.M., Muszynski,M.G., Phillips,R.L., Kaeppler,S. and Jacobsen,S.E. (2000) Conserved plant genes with similarity to mammalian de novo DNA methyltransferases. Proc. Natl Acad. Sci. USA, 97, 4979–4984. [DOI] [PMC free article] [PubMed] [Google Scholar]