


Abstract
The ING1 candidate tumor suppressor is downregulated in a variety of primary tumors and established cancer cell lines. Blocking its expression experimentally promotes unregulated growth in vitro and in vivo, using cell and animal models. Alternative splicing products encode proteins that localize to the nucleus, inhibit cell cycle progression and affect apoptosis in different model systems. Here we show that ING1 proteins translocate to the nucleolus 12–48 h after UV-induced DNA damage. When a small 50 amino acid portion of ING1 was fused to green fluorescent protein, the fusion protein was efficiently targeted to the nucleolus, indicating that ING1 possesses an intrinsic nucleolar targeting sequence (NTS). We mapped this activity to two distinct 4 amino acid regions, which individually direct fused heterologous proteins to the nucleolus. Overexpression of ING1 induced apoptosis of primary fibroblasts in the presence and absence of UV exposure. In contrast, NTS mutants of ING1 that were not targeted to the nucleolus did not efficiently induce apoptosis when overexpressed and instead protected cells from UV-induced apoptosis. Taken together, these results indicate that UV induces ING1 to translocate to the nucleolus and that this translocation may facilitate apoptosis.
INTRODUCTION
The ING1 candidate tumor suppressor has been reported to be downregulated in breast (1), gastric (2), lymphoid (3) and squamous cell (4) malignancies. ING1 encodes nuclear proteins involved in cell growth control (5), as well as c-Myc-inducible, p53-independent and -dependent apoptosis (6,7). Several transcripts are generated by alternative splicing from the ING1 gene, all of which encode a common C-terminus (4,8). A yeast ortholog of ING1 (Yng2) interacts with Tra1, a yeast protein with sequence similarity to human TRRAP, and Yng2 immunocomplexes contain abundant histone acetyltransferase (HAT) activity (9). TRRAP recruits at least one HAT, hGCN5, to a protein complex containing both c-Myc and E2F-1 (10). These observations, and our recent evidence of ING1-associated HAT activity in human cells (D.Vieyra, R.Loewith, M.Scott, P.Bonnefin, F.-M.Boisvert, P.Cheema, M.Meijer, D.Bazett-Jones, S.McMahone, M.D.Cole, D.Young and K.Riabowol, manuscript submitted), support a role for ING1 as a modifier of chromatin structure through the regulation of histone, and possibly, transcription factor acetylation.
The nucleolus is a specialized region of the nucleus where transcription and processing of rRNA and partial ribosome assembly occurs. However, there is increasing evidence for the presence of non-ribosomal proteins including growth factors (11–13), death effector proteins (14), viral proteins (15,16) and regulators of apoptosis such as MDM2 and p14ARF (17,18) in the nucleolus under certain conditions. These proteins are involved in processes such as cell cycle and cellular growth control, apoptosis events and viral infection, which implicate the involvement of the nucleolus in diverse cellular processes. Here, we begin the investigation of the role of the p33ING1b candidate tumor suppressor protein in the nucleolus after DNA damage.
The ING1 proteins localize to the nucleoplasm (19), but when overexpressed we noted that they also accumulate in the nucleolus. Here we report that, under conditions that induce cells to undergo DNA repair, ING1 is efficiently targeted to the nucleolus. ING1 possesses a nucleolar targeting sequence (NTS) as previously defined (20), which can direct fused heterologous proteins, such as green fluorescent protein (GFP), to the nucleolus. However, another region of ING1 previously thought to be a nuclear localization signal (NLS) by comparison to other nuclear proteins, can also cause the relocalization of unrelated proteins to the nucleolus. When ING1 translocates to the nucleolus after UV irradiation, we found that polymerase I-mediated RNA transcription continues in the nucleolus but transcription in the nucleoplasm is inhibited. This is consistent with reports indicating a lack of rDNA transcription-coupled repair after UV irradiation (21). While overexpression of p33ING1 efficiently induces apoptosis in different cell lines (6,7), we find that mutants of p33ING1b lacking a functional NTS do not, but instead have a protective effect following UV exposure implying that nucleolar localization of ING1 is required for the efficient induction of apoptosis in primary fibroblast cells.
MATERIALS AND METHODS
Cells and DNA constructs
Primary normal human diploid fibroblasts (HDFs; Hs68, ATCC CRL#1635) were used for all irradiation studies and both SK-N-SH and Hs68 cells were transfected for overexpression studies. All mutation and deletion constructs were cloned into either pCI-neo vector (Promega) or pEGFP-N in frame with the GFP (Clontech) and sequenced to verify reading frame. The Tat expression construct used is FLAG-Tat-2-exons cloned into pcDNA3.1.
Transfection, irradiation and transcription assays
Hs68 and SK-N-SH cells plated on glass coverslips were transfected with different constructs using Lipofectamine 2000 as recommended by the supplier (Gibco BRL) and were left to recover 24 h before fixing and labeling. For the study of localization of ING1 and transcription after UV irradiation, cells plated on glass coverslips were irradiated with several doses of UV ranging from 10 to 100 J/m2 in initial dose-response assays and 60–70 J/m2 was found to be optimal. Irradiation was carried out in the absence of media and cells were left for 48 h in complete medium at 37°C to recover. They were then incubated with 2 mM 5-fluorouridine (5-FU) in complete medium for 30 min at 37°C, fixed and labeled with antibodies specific for ING1 protein or 5-FU as described below.
Inhibition of RNA polymerases
Cells plated on glass coverslips were UV irradiated as described above and left to recover at 37°C for 45 h. Cells were then treated with RNA polymerase inhibitors for 3 h before fixation, as follows: 0.5 µg/ml of actinomycin D (from a 2 mg/ml stock solution in methanol), 75 µg/ml of DRB (from a 25 mg/ml stock solution in DMSO) or 3 µg/ml of α-amanitin (from a 2 mg/ml stock solution in water). After 2.5 h, 5-FU was added to the media as described above, for 30 min before fixation.
Apoptosis assays
Cells were assayed for apoptosis by propidium iodide staining followed by FACS and by detection of DNA breaks by TUNEL as described (22) using a CMV-GFP plasmid as a transfection control and normalizing to vector-transfected cells assayed in parallel.
Indirect immunofluorescence and confocal microscopy
Hs68 HDFs were analyzed as described previously (23). Briefly, cells were fixed for 5 min in 1.0% paraformaldehyde in PBS pH 7.5 and then permeabilized for 5 min in 0.5% Triton X-100 in PBS. ING1 proteins were labeled with a cocktail of four mouse monoclonal antibodies (CAb1–4; 24) or a rabbit polyclonal anti-ING1 antibody (19). Tat proteins were labeled with a mouse monoclonal anti-FLAG or rabbit polyclonal anti-FLAG antibody and 5-FU was visualized by incubating with a mouse monoclonal anti-bromodeoxyuridine antibody (Sigma # B-2531), for at least 30 min. Cells were then washed with 0.1% Triton X-100 in PBS, followed by PBS and were then incubated with secondary antibody (goat anti-rabbit Cy3, goat anti-mouse Cy3 or goat anti-mouse Alexa 488 from Chemicon) for 30 min. Cells were rinsed with 0.1% Triton X-100 in PBS, in PBS alone and then mounted in 1 mg/ml paraphenylenediamine in PBS/90% glycerol that also contained DAPI at 1 µg/ml. Digital imaging was performed using a 14-bit cooled CCD camera (Princeton Instruments) mounted on a Leica DMRE immunofluorescence microscope.
RESULTS
p33ING1b is targeted to the nucleolus when overexpressed and after UV irradiation
Visualization of endogenous ING1 using a cocktail of monoclonal antibodies shows staining of the nucleoplasm with no staining of the nucleolus (Fig. 1A–C). However, overexpression in cells transfected with an expression construct of p33ING1b produces a nuclear as well as a very prominent nucleolar staining pattern by immunofluorescence microscopy (Fig. 1D–F).
Figure 1.

Hs68 fibroblasts were fixed and stained for ING1 (B,E and H) and DNA (A,D and G). Endogenous ING1 (B) is found in the nucleus but not in the nucleolus [which is shown in the differential interference contrast (DIC) images (C,F and I)]. However, when these proteins are overexpressed, they localize mainly to the nucleolus and can also be seen in the nucleoplasm (E). Endogenous ING1 proteins translocate to, and localize within, the nucleolus 48 h after UV-induced DNA damage as shown in (H).
Since our studies and the studies of others (25) indicated that the ING1 gene is inducible in response to some forms of DNA damage, we tested whether UV irradiation could also alter endogenous ING1 distribution. Normal diploid fibroblasts irradiated with relatively high doses of UV light (60–70 J/m2), which causes DNA damage and leads to apoptosis in most cells (26), showed nucleolar ING1 staining from 12–48 h after UV irradiation (Fig. 1G–I). Nucleolar targeting of ING1 did not occur before 12 h after UV irradiation of cells, indicating that further cellular events are necessary to allow this translocation. p53, which has been reported by others to interact with p33ING1b when ectopically overexpressed (27), did not translocate with p33ING1b to the nucleolus under the same conditions (data not shown).
p33ING1b contains a functional NTS
We next examined whether ING1 encoded a potential NTS. Amino acid sequence analyses revealed the presence of a complete NTS as shown in Figure 2A, compared to previously defined NTS sequences. To determine if it was responsible for targeting p33ING1b to the nucleolus, a construct of 156 bp (amino acids 142–194 of the protein) consisting of the putative NTS as well as the NLS found in p33ING1b was cloned into pEGFP-N2 in frame with the cDNA encoding GFP (Fig. 2B). An ATG codon was added 5′ of the 156 bp sequence to allow proper translation of the fusion protein. This fusion protein localized almost exclusively in the nucleolus (data not shown) and additional fusion proteins (Fig. 2B) confirmed the regions of ING1 required for nucleolar targeting.
Figure 2.

Examination of the amino acid sequence of p33ING1b reveals the presence of a complete NTS in the second half of the protein. Several known cellular and viral proteins, which can be found in the nucleolus, at least under certain conditions, contain this targeting sequence (A). Cells were transfected with the ING1 deletion constructs shown in (B) and examined for subcellular distribution by indirect immunofluorescence, which confirmed a requirement for the region between amino acids 142 and 194.
Constructs encoding fusion proteins containing different mutations in the NTS were transfected into cells, which were fixed and stained for expression 24 h later by indirect immunofluorescence. Since previous reports have indicated that some nucleolar proteins do not contain an NTS or contain an NTS but depend on other sequences for nucleolar targeting (28,29), we also constructed additional fusions of p33ING1b containing other key residues. As shown in Figure 3, amino acids 142–159 (construct N1N2 containing the -RRQR- consensus) alone could target GFP to the nucleolus. Surprisingly, amino acids 174–194 (construct N3N4), which was believed to contain a unique NLS based upon sequence, could also target GFP to the nucleolus. This result is similar to the finding that a putative NLS (one of three found in the nucleolar protein Rpp38) is an efficient NTS (29). Mutational analysis of p33ING1b showed that the targeting of this protein to the nucleolus requires either the amino acids RRQR within the NTS or the amino acids KKKK within the region previously believed to be an NLS (Fig. 3A and B). Mutation of these 8 amino acids (mut3-4p) completely abrogated the translocation of p33ING1b to the nucleolus. Mutation of other residues did not inhibit the nucleolar targeting of p33ING1b (Fig. 3A and data not shown). In addition, mutation of the amino acids RRQR or KKKK separately (mut3p and mut4p in Fig. 3A) did not prevent nucleolar localization of the full-length GFP fusion protein. Mutation of RRQR in a portion of the protein that does not contain KKKK (mut3G) and vice versa (mut4G) did, however, block the nucleolar targeting of these portions of p33ING1b, although they still localized to the nucleus. Interestingly, the amino acids KEKK (which form the other half of the putative NLS with amino acids KKKK) are not responsible for nucleolar targeting (mut5G2 in Fig. 3A). Taken together, these results indicate that this protein contains two regions of 4 amino acids that function as NTSs. Each one is individually capable of localizing ING1 and heterologous fusion proteins to the nucleolus.
Figure 3.

Mutated versions of p33ING1b (A) were generated and transfected into Hs68 fibroblasts. Cells were visualized (B) by staining for DNA (A,D,G and J), ING1 proteins (B,E,H and K) and by DIC (C,F,I and L).
ING1 is not retained in the nucleoplasm by a limiting factor that binds a consensus NTS
Since no detectable endogenous ING1 is present in the nucleolus of untreated cells (Fig. 1), there may be a common mechanism to retain proteins that contain NTSs in the nucleoplasm. For example, ING1 might be bound in either the nucleoplasm or nucleolus by a titratable factor. Alternatively, translocation could occur as a result of ING1 undergoing a conformational change. To address these possibilities, we transfected cells with a construct encoding the viral protein Tat, which contains an NTS very similar to one of the NTS elements present in ING1 (Fig. 2A) and which has been shown previously to localize to the nucleoplasm when present at low levels and to the nucleolus when overexpressed (16). When Tat was overexpressed, it localized mainly to the nucleolus but endogenous ING1 remained in the nucleoplasm (Fig. 4A–D). This indicated that ING1 was not retained in the nucleoplasm by a limiting factor that is titratable by Tat in this experimental system. If it were, the limiting factor would also bind the NTS present on Tat and would not be present in amounts large enough to keep all the ING1 in the nucleoplasm. Also, coexpression of ING1 and Tat resulted in targeting of both proteins to the nucleolus to the same degree as observed when only one of the proteins was transfected (Fig. 4E–H). This implies that they do not compete for the binding of a limiting factor in the nucleolus to remain in that organelle.
Figure 4.
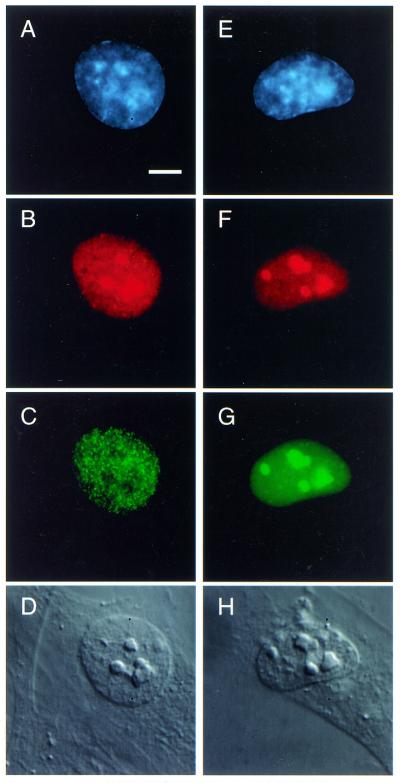
Hs68 fibroblasts were transfected with Tat (A–D) or with Tat and GFP–ING1b (E–H). Cells were stained for DNA (A and E), Tat (B and F) and ING1 (C andG). When overexpressed, Tat localizes to the nucleolus (shown by DIC in D) but does not cause relocation of ING1 to the nucleolus (C).
RNA polymerase I (RNA pol I) is not inhibited under conditions of DNA damage that induce translocation of p33ING1b to the nucleolus
Since the nucleolus is the site of rRNA transcription by RNA pol I, we asked whether the higher concentrations of p33ING1b found in some cells might influence transcription by RNA pol I compared to transcription by RNA polymerases II and III. We therefore exposed normal diploid fibroblasts to UV irradiation, let them recover for 48 h and added 5-FU to the cells for 30 min. Cells were then labeled with both polyclonal antibodies against ING1 proteins and monoclonal antibodies against BrdU as described previously (23). 5-FU is a uridine analog that is incorporated into nascent RNA transcribed by RNA polymerases I, II and III. The antibodies against BrdU also recognize 5-FU, which allows visualization of transcriptional activity by immunofluorescence microscopy. As expected, in cells treated with UV, ING1 localized to the nucleolus (Fig. 5G), whereas in untreated cells, ING1 was detected only in the nucleoplasm (Fig. 5C). The large size of the nuclei of cells irradiated with UV (Fig. 5E) and their DNA staining pattern is consistent at this dose, with UV inducing apoptosis in these cells. Transcription was detected throughout the nucleus of all untreated cells, with higher amounts in the nucleolus (Fig. 5B). However, in cells irradiated with UV, transcription was nearly exclusively nucleolar (Fig. 5F). Treatment of cells with the RNA polymerase inhibitors DRB, α-amanitin or actinomycin D confirmed that RNA polymerases II and III are efficiently inhibited after cells are UV-irradiated (data not shown). In contrast, RNA pol I was not inhibited and continued to transcribe RNA under conditions of DNA damage caused by UV irradiation, during the same time period that the candidate tumor suppressor ING1b translocates to the nucleolus. Causal links, however, if they exist, remain to be established. The inhibition of nucleoplasmic, but not nucleolar transcription is consistent with studies demonstrating that rDNA in the nucleolus does not undergo transcription-coupled repair (21) or repair of cyclobutane pyrimidine dimers (30) after UV treatment.
Figure 5.

UV-irradiated HS68 cells were allowed to recover for 48 h and were incubated for 30 min with 5-FU. In cells treated with UV (E–H), ING1 localized to the nucleolus (G) whereas in untreated cells (A–D), ING1 was detected only in the nucleoplasm (C). Transcription was detected using anti-BrdU antibodies throughout the nucleus of all untreated cells with higher amounts of transcription found in the nucleolus (B). However, in cells treated with UV irradiation, transcription was found nearly exclusively in the nucleolus (F).
NTS mutants that do not translocate to the nucleolus do not efficiently induce apoptosis
Since exposure to UV caused endogenous ING1 to translocate to the nucleolus and overexpression of p33ING1b had been previously reported to induce apoptosis in different primary and established cell types (6,7), we wished to determine if nucleolar translocation affected the induction of apoptosis in primary human diploid fibroblasts. Plasmids containing empty vector, wild-type p33ING1b or p33ING1b lacking functional NTSs were transfected into cells and 24 h later one set of the transfected cells was exposed to 70 J/m2 of UV. Forty-eight hours after transfection, cells were analyzed for DNA content by FACS analysis of propidium iodide-stained cells or for DNA strand breaks by TUNEL. Results from three independent experiments are shown in Table 1. Results were normalized to values using vector, in the absence or presence of UV to control for electroporation trauma and the accumulation of necrotic cells that can contribute to measures of both sub-G1 DNA content and DNA strand breaks. p33ING1 induced apoptosis when overexpressed and this effect was more pronounced following exposure to UV. In contrast, overexpression of the NTS mutant did not induce apoptosis as efficiently as wild-type p33ING1 in the absence of UV. In populations of cells transfected with the NTS mutant and exposed to UV light, the number of apoptotic cells was reproducibly reduced compared to UV-exposed cells harboring vector, suggesting that the translocation mutant was capable of showing a dominant negative (protective) effect following UV-induced DNA damage.
Table 1. Effects of wild-type and NTS mutant forms of p33ING1b on apoptosis.
| Percentage apoptosis | ||
| Treatment |
p33ING1b |
p33ING1b NTS mutant |
| –UV |
27.8 ± 3.1 |
14.1 ± 2.5 |
| +UV | 61.2 ± 6.2 | –7.5 ± 1.2 |
Wild-type or an NTS mutant of p33ING1b in expression constructs driven by cytomegalovirus (CMV) promoters were cotransfected with a CMV–GFP expression construct into primary human Hs68 fibroblasts. After 48 h of incubation, cells were harvested and stained using the TUNEL procedure. FACS analyses were then performed to identify the percentage of cells expressing GFP that also stained by TUNEL.
DISCUSSION
In this study, we have shown that overexpression of p33ING1b causes this protein to be highly enriched in the nucleolus and that endogenous p33ING1b also translocates to the nucleolus in response to UV exposure that causes DNA damage. The nucleolar translocation of p33ING1b depends upon two stretches of 4 amino acids each, which act as potent NTSs and which are also capable of targeting heterologous fusion proteins to the nucleolus. p33ING1b mutants of the NTS are less effective in inducing apoptosis suggesting that such localization contributes to ING1 promoting programmed cell death. Unlike the case for ARF [which impinges upon apoptosis through interacting with MDM2 in the nucleolus, subsequently affecting p53 activity (17,18)], the mechanism by which ING1 affects the apoptosis of primary cells, which occurs in a protracted time frame (31), is presently unknown. It is clear, however, that the NTS mutant is capable of weakly inducing apoptosis in the absence of DNA damage but that it has a protective effect following UV exposure. These observations suggest that p33ING1b affects at least two different cellular processes. It is also interesting to note that the sequences responsible for the translocation of p33ING1b to the nucleolus are located in a region common to all splicing isoforms of the ING1 gene. This implies that the other two known isoforms of the ING1 gene, p47ING1a and p24ING1c (4,32,33), also possess the NTS and should be able to localize to the nucleolus under some circumstances. Further studies using isoform-specific reagents will be necessary to address this question.
Other studies have shown that proteins that are non-nucleolar under normal cell growth conditions translocate to the nucleolus under conditions of DNA damage and apoptosis. For example, the death effector domain containing DNA-binding protein (DEDD) is cytoplasmic but translocates to the nucleolus when CD95 is stimulated in 293T cells. DEDD is thought to be involved in the CD95 apoptotic pathway as an effector molecule (14). Additionally, the parathyroid hormone-related peptide (PTHrP) has been shown to be a secreted protein also found in the cytoplasm of normal murine cells, but can be present in the nucleolus of a subpopulation of these cells. PTHrP can enhance the survival of chondrocytes undergoing apoptosis, but only when the NTS of the protein is intact and the protein is found in the nucleolus (20). Such inducibly targeted proteins may specify a new functional class of proteins that are involved in apoptotic events. Overexpression of wild-type p33ING1b promotes apoptosis as shown by several groups. The fact that nucleolar translocation mutants of p33ING1b do not efficiently promote programmed cell death and can protect against apoptosis under some circumstances provides direct evidence for a nucleolus-sensitive function.
Acknowledgments
ACKNOWLEDGEMENTS
We thank D.Jackson for help with fluorouridine labeling, L.Robertson for FACS analyses, the SACRC Hybridoma Facility for immunological reagents and E.Verdin for the Tat expression construct. K.R. is a scientist of the Alberta Heritage Foundation for Medical Research (AHFMR) and the Canadian Institutes for Health. M.S. is supported by the National Science and Engineering Council of Canada and M.S. and F.-M.B. are recipients of AHFMR studentships. This work was supported by grants to K.R. from the Canadian Institutes of Health Research (CIHR) and the National Cancer Institute of Canada, to D.P.B.-J. from the Cancer Research Society Inc. and to R.N.J., D.P.B.-J. and K.R. from the Alberta Cancer Board.
References
- 1.Toyama T., Iwase,H., Watson,P., Muzik,H., Saettler,E., Magliocco,A., DiFrancesco,L., Forsyth,P., Garkavtsev,I., Kobayashi,S. and Riabowol,K. (1999) Suppression of ING1 expression in sporadic breast cancer. Oncogene, 18, 5187–5193. [DOI] [PubMed] [Google Scholar]
- 2.Oki E., Maehara,Y., Tokunaga,E., Kakegi,Y. and Sugimachi,K. (1999) Reduced expression of p33ING1 and the relationship with p53 expression in human gastric cancer. Cancer Lett., 62, 157–162. [DOI] [PubMed] [Google Scholar]
- 3.Ohmori M., Nagai,M., Tasaka,T., Koeffler,H.P., Riabowol,K. and Takahara,K. (1999) Decreased expression of p33ING1 mRNA in lymphoid malignancies. Am. J. Hematol., 62, 118–119. [DOI] [PubMed] [Google Scholar]
- 4.Gunduz M., Ouchida,M., Fukushima,K., Hanafusa,H., Etani,T., Nishioka,S., Nishizaki,K. and Shimizu,K. (2000) Genomic structure of the human ING1 gene and tumor-specific mutations detected in head and neck squamous cell carcinomas. Cancer Res., 60, 3143–3146. [PubMed] [Google Scholar]
- 5.Garkavtsev I., Karazov,A., Gudkov,A. and Riabowol,K. (1996) Suppression of the novel growth inhibitor p33ING1 promotes neoplastic transformation. Nat. Genet., 14, 415–420. [DOI] [PubMed] [Google Scholar]
- 6.Helbing C.C., Veillette,C., Riabowol,K., Johnston,R.N. and Garkavtsev,I. (1997) A novel candidate tumor suppressor, ING1, is involved in the regulation of apoptosis. Cancer Res., 57, 1255–1258. [PubMed] [Google Scholar]
- 7.Shinoura N., Muramatsu,Y., Nishimura,M., Yoshida,Y., Saito,A., Yokoyama,T., Furukawa,T., Horii,A., Hashimoto,M., Asai,A., Kirino,T. and Hamada,H. (1999) Adenovirus-mediated transfer of p33ING1 with p53 drastically augments apoptosis in gliomas. Cancer Res., 59, 5521–5528. [PubMed] [Google Scholar]
- 8.Jager D., Stockert,E., Scanlan,M., Gure,A.O., Jager,E., Knuth,A., Old,L.J. and Chen,Y.-T. (1999) Cancer-testis antigens and ING1 tumor suppressor gene product are breast cancer antigens: characterization of tissue-specific ING1 transcripts and a homologue gene. Cancer Res., 59, 6197–6204. [PubMed] [Google Scholar]
- 9.Loewith R., Meijer,M., Lees-Miller,S.P., Riabowol,K. and Young,D. (2000) A family of yeast proteins related to the candidate human tumor suppressor p33ING1 associated with histone acetyl transferase in vivo. Mol. Cell. Biol., 20, 3807–3816. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.McMahon S.B., Van Buskirk,H.A., Dugan,K.A., Copeland,T.D. and Cole,M.D. (1998) The novel ATM-related protein TRRAP is an essential cofactor for the c-Myc and E2F oncoproteins. Cell, 94, 363–374. [DOI] [PubMed] [Google Scholar]
- 11.Antoine M., Reimers,K., Dickson,C. and Kiefer,P. (1997) Fibroblast growth factor 3, a protein with dual subcellular localization, is targeted to the nucleus and nucleolus by the concerted action of two nuclear localization signals and a nucleolar retention signal. J. Biol. Chem., 272, 29475–29481. [DOI] [PubMed] [Google Scholar]
- 12.Galcheva-Gargova Z., Gangwani,L., Konstantinov,K.N., Mikrut,M., Theroux,S.J., Enoch,T. and Davis,R.J. (1998) The cytoplasmic zinc finger protein ZPR1 accumulates in the nucleolus of proliferating cells. Mol. Biol. Cell, 9, 2963–2971. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Pederson T. (1998) Growth factors in the nucleolus? J. Cell Biol., 143, 279–281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Stegh A., Schickling,O., Ehret,A., Scaffidi,C., Peterhansel,C., Hofmann,T.G., Grummt,I., Krammer,P.H. and Peter,M. (1998) DEDD, a novel death effector domain-containing protein, targeted to the nucleolus. EMBO J., 17, 5974–5986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Hatanaka M. (1990) Discovery of the nucleolar targeting signal. Bioessays, 12, 143–148. [DOI] [PubMed] [Google Scholar]
- 16.Stauber R.H. and Pavlakis,G.N. (1998) Intracellular trafficking and interactions of the HIV-1 Tat protein. Virology, 252, 126–136. [DOI] [PubMed] [Google Scholar]
- 17.Zhang Y. and Xiong,Y. (1999) Mutations in human ARF exon 2 disrupt its nucleolar localization and impair its ability to block nuclear export of MDM2 and p53. Mol. Cell, 3, 579–591. [DOI] [PubMed] [Google Scholar]
- 18.Weber J.D., Taylor,L.J., Roussel,M.F., Sherr,C.J. and Bar-Sagi,D. (1999) Nucleolar Arf sequesters Mdm2 and activates p53. Nat. Cell Biol., 1, 20–26. [DOI] [PubMed] [Google Scholar]
- 19.Garkavtsev I., Demetrick,D. and Riabowol,K. (1997) Cellular localization and chromosome mapping of a novel candidate tumor suppressor gene (ING1). Cytogenet. Cell Genet., 76, 176–178. [DOI] [PubMed] [Google Scholar]
- 20.Henderson J.E., Amizuka,N., Warshawshy,H., Biasotto,D., Lanske,B.M.K., Goltzman,D. and Karaplis,A.C. (1995) Nucleolar localization of parathyroid hormone-related peptide enhances survival of chondrocytes under conditions that promote apoptotic cell death. Mol. Cell. Biol., 15, 4064–4075. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Christians F.C. and Hanawalt,P.C. (1993) Lack of transcription-coupled repair in mammalian ribosomal RNA genes. Biochemistry, 32, 10512–10518. [DOI] [PubMed] [Google Scholar]
- 22.Kaufmann S., Mesner,P., Samejima,K., Tone,S. and Earnshaw,W. (2000) Measuring apoptosis and apoptosis-induced endonucleases. Methods Enzymol., 322, 3–47. [DOI] [PubMed] [Google Scholar]
- 23.Boisvert F.-M., Hendzel,M.H. and Bazett-Jones,D.P. (2000) Promyelocytic leukemia (PML) nuclear bodies are protein structures that do not accumulate RNA. J. Cell Biol., 148, 283–292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Boland D., Olineck,V., Bonnefin,P., Vieyra,D., Parr,E. and Riabowol,K. (2000) A panel of Cab antibodies recognize endogenous and ectopically expressed ING1 protein. Hybridoma, 19, 161–166. [DOI] [PubMed] [Google Scholar]
- 25.Cheung K.-J.,Jr, Bush,J.A and Li,G. (2000) Expression of the novel tumour suppressor p33ING1 is independent of p53. Br. J. Cancer, 83, 1468–1472. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Lu X. and Lane,D.P. (1993) Differential induction of transcriptionally active p53 following UV or ionizing radiation: defects in chromosome instability syndrome? Cell, 75, 765–778. [DOI] [PubMed] [Google Scholar]
- 27.Garkavtsev I., Grigorian,I.A., Ossovskaya,V.S., Chernov,M.V., Chumakov,P.M. and Gudkov,A.V. (1998) The candidate tumour suppressor p33ing1 cooperates with p53 in cell growth control. Nature, 391, 295–298. [DOI] [PubMed] [Google Scholar]
- 28.Schmidt-Zachmann M.S. and Nigg,E.A. (1993) Protein localization to the nucleolus: a search for targeting domains in nucleolin. J. Cell Sci., 105, 799–806. [DOI] [PubMed] [Google Scholar]
- 29.Jarrous N., Wolenski,J.S., Wesolowski,D., Lee,C. and Altman,S. (1999) Localization in the nucleolus and coiled bodies of protein subunits of the ribonucleoprotein ribonuclease P.J. J. Cell Biol., 146, 559–571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Balajee A.S., May,A. and Bohr,V.A. (1999) DNA repair of pyrimidine dimers and 6-4 photoproducts in the ribosomal DNA. Nucleic Acids Res., 27, 2511–2520. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Andera L. and Wasylyk,B. (1997) Transcription abnormalities potentiate apoptosis of normal human fibroblasts. Mol. Med., 3, 852–863. [PMC free article] [PubMed] [Google Scholar]
- 32.Ma D., Lawless,D. and Riabowol,K. (1999) Correction and addition to ING1 isoform sequences. Nat. Genet., 23, 73. [Google Scholar]
- 33.Saito A., Furukawa,T., Fukushige,S., Koyama,S., Hoshi,M., Hayashi,Y. and Horii,A. (2000) p24/ING1-ALT1 and p47/ING1-ALT2, distinct alternative transcripts of p33/ING1. J. Hum. Genet., 45, 177–181. [DOI] [PubMed] [Google Scholar]