

Abstract
Mammalian physiology and behavior are regulated by an internal time-keeping system, referred to as circadian rhythm. The circadian timing system has a hierarchical organization composed of the master clock in the suprachiasmatic nucleus (SCN) and local clocks in extra-SCN brain regions and peripheral organs. The circadian clock molecular mechanism involves a network of transcription-translation feedback loops. In addition to the clinical association between circadian rhythm disruption and mood disorders, recent studies have suggested a molecular link between mood regulation and circadian rhythm. Specifically, genetic deletion of the circadian nuclear receptor Rev-erbα induces mania-like behavior caused by increased midbrain dopaminergic (DAergic) tone at dusk. The association between circadian rhythm and emotion-related behaviors can be applied to pathological conditions, including neurodegenerative diseases. In Parkinson’s disease (PD), DAergic neurons in the substantia nigra pars compacta progressively degenerate leading to motor dysfunction. Patients with PD also exhibit non-motor symptoms, including sleep disorder and neuropsychiatric disorders. Thus, it is important to understand the mechanisms that link the molecular circadian clock and brain machinery in the regulation of emotional behaviors and related midbrain DAergic neuronal circuits in healthy and pathological states. This review summarizes the current literature regarding the association between circadian rhythm and mood regulation from a chronobiological perspective, and may provide insight into therapeutic approaches to target psychiatric symptoms in neurodegenerative diseases involving circadian rhythm dysfunction.
Keywords: circadian rhythm, dopaminergic system, mood disorder, Parkinson’s disease, REV-ERBα
INTRODUCTION
The circadian time-keeping system evolved from cyanobacteria to humans, and drives circadian rhythm over a 24-h period to anticipate and respond to environmental changes in accordance with sunrise and sunset. Molecular clocks, found in nearly all tissues, are organized in a hierarchical system, with the master clock located in the hypothalamic suprachiasmatic nucleus (SCN) of the anterior hypothalamus and local clocks located in both extra-SCN brain regions and peripheral organs. The master clock synchronizes the internal timing of peripheral clocks to drive circadian control of physiology and behavior.
Chronic disturbances in circadian rhythmicity in patients with mood disorders were noted over 50 years ago (Wirz-Justice, 2006). Patients with mood disorders, including major depressive disorder (MDD), bipolar disorder (BPD), and seasonal affective disorder (SAD), exhibit disrupted circadian rhythmicity in body temperature, hormone secretions, e.g., cortisol and melatonin, blood pressure, and sleep-wake cycles (Albrecht, 2013; Wirz-Justice, 2006). Human genetic and animal studies have shown molecular links between circadian rhythm and mood disorders (McCarthy and Welsh, 2012). The central neurotransmitter system has been implicated in mood regulation via a variety of biochemical and signal transduction processes (McClung, 2007; Russo and Nestler, 2013). Identification of molecular clockwork components has provided a link between the circadian clock and the monoamine system (Albrecht, 2017).
There is an association between circadian rhythm and emotional regulation in neurodegenerative diseases, such as Parkinson’s disease (PD) (Wulff et al., 2010). PD is characterized by progressive degeneration of dopaminergic (DAergic) neurons in the substantia nigra pars compacta (SNpc), leading to motor dysfunction (Dauer and Przedborski, 2003). Patients with PD also display non-motor and circadian rhythm-related symptoms, such as mood dysregulation, specifically depression, anxiety, and apathy (Chaudhuri and Schapira, 2009; Videnovic et al., 2014). However, the etiology of mood disorders in PD has not been elucidated, and drug development has been limited (Aarsland et al., 2011). We review the literature concerning mood regulation in healthy and PD states from a chronobiological view.
MAMMALIAN CIRCADIAN CLOCK
The mammalian circadian clock is a hierarchical time-keeping system. The master clock, which acts as the circadian pacemaker, is located in the suprachiasmatic nucleus (SCN) of the anterior hypothalamus. Local clocks exist in extra-SCN brain regions and peripheral organs (Balsalobre et al., 1998; Yamazaki et al., 2000). The master clock synchronizes internal clock timing to external photic zeitgebers from light input via the retina, and peripheral clocks mediate circadian control of physiology and behavior by adjustment from the SCN via endocrine and systemic cues (Dibner et al., 2010).
The daily timing of physiological processes is influenced by peripheral oscillators. Transcriptome-profiling supports this notion; more than 10% of total mRNA shows circadian expression patterns in the liver (Akhtar et al., 2002; Storch et al., 2002; Zhang et al., 2014). By comparing the degree of circadian regulation in different tissues, it has been shown that most circadian gene transcripts are expressed in a tissue-specific manner, and that the circadian phase of gene transcripts is distinct (Panda et al., 2002; Storch et al., 2002).
The circadian clock consists of a network of transcription-translation feedback loops that generate endogenous circadian rhythm. In the core feedback loop, the positive elements include members of the basic helix-loop-helix (bHLH)-PAS (Period-ARNT-Single-minded) domain-containing transcription factor family, CLOCK (or NPAS), and BMAL1 (Bunger et al., 2000; King et al., 1997). CLOCK and BMAL1 heterodimerize and activate transcription of target genes containing E-box cis-regulatory enhancer sequences in their promoter regions, such as Periods (Per1, Per2, and Per3) and Cryptochromes (Cry1 and Cry2) (Gekakis et al., 1998). Negative feedback is achieved by PER:CRY heterodimers that translocate to the nucleus to repress their own expression by inhibiting CLOCK:BMAL1 heterodimer activity (Kume et al., 1999; Shearman et al., 2000).
Another regulatory loop controls Bmal1 expression through competitive binding of two retinoic acid-related orphan nuclear receptors, REV-ERBα and RORα, to retinoic acid-related orphan receptor response elements (ROREs) in the Bmal1 promoter. While RORα induces Bmal1 transcription, REV-ERBα represses Bmal1 expression (Guillaumond et al., 2005; Preitner et al., 2002) through the recruitment of co-repressor N-CoR and histone deacetylase3 (HDAC3) (Yin and Lazar, 2005). Extensive post-translational modifications of core clock proteins also provide fine tuning of circadian clock system by controlling their stability, nuclear localization, and activity (Gallego and Virshup, 2007).
Under pathological conditions, decreased circadian amplitude – the difference between peaks and troughs of circadian rhythms – is often observed (Gloston et al., 2017). Rev-erb α is a key molecule for determining amplitude. F-box protein FBX7 ubiquitinates and degrades phosphorylated REV-ERBα by cyclin-dependent kinase 1 (CDK1) to regulate clock amplitude (Zhao et al., 2016). ROR promotes chromatin decondensation during the activation phase of the circadian cycle to facilitate REV-ERB binding to open chromatin during the inactivation phase to maintain circadian amplitude (Zhu et al., 2015).
CIRCADIAN DYSFUNCTION AND MOOD DISORDERS
Patients with mood disorders often display abnormal rhythmicity of body temperature, cortisol and melatonin levels, blood pressure, and sleep/wake cycles suggesting circadian rhythm disruption (Wirz-Justice, 2006). Sundown syndrome, also referred to as “nocturnal delirium,” is characterized by worsening of behaviors such as agitation, aggression, restlessness, and delirium, particularly during the late afternoon/early evening, implying a strong association between circadian rhythm and mood regulation (Bedrosian and Nelson, 2013). Genome-wide association studies have identified circadian gene polymorphisms that influence psychiatric disease susceptibility. These circadian gene variants include CLOCK, BMAL1, PER3, and REV-ERBα, which are associated with BPD, MDD, and SAD (Kripke et al., 2009; Mansour et al., 2006; Soria et al., 2010). Moreover, a transcriptome-wide analysis of postmortem brains from patients with MDD revealed weaker circadian gene expression and disrupted phase relationships between individual clock genes (Li et al., 2013).
Several animal studies support the influence of circadian clock genes on mood regulation in brain regions implicated in emotion. ClockΔ19 mice, which harbor Clock gene mutations, exhibit mania-like behavioral patterns, similar to those in BPD, including hyperactivity, increased cocaine sensitization, decreased depression, anxiety-like behaviors, and decreased sleep latency (Easton et al., 2003; McClung et al., 2005; Roybal et al., 2007). ClockΔ19 mice display enhanced ventral tegmental area (VTA) DAergic cell firing and bursting (McClung et al., 2005). Viral delivery of a functional Clock gene into the VTA rescues mania-like behaviors (Roybal et al., 2007), suggesting that manipulation of CLOCK activity in the VTA DAergic system regulates mood. Per1 knockout mice (Zheng et al., 2001) and Per2Brdm1 mice (Per2 mutant mice harboring a deletion in the PAS domain) (Zheng et al., 1999) exhibit different sensitivities to cocaine administration, indicating abnormalities in the reward circuit. Per1 mutant mice show lack of sensitization, while Per2 mutant mice display hypersensitization to cocaine compared to wild-type mice (Abarca et al., 2002). Per2Brdm1 mice exhibit a less depressive-like phenotype in despair-based tests. The phenotype is associated with increased DA levels in the mesolimbic DAergic circuit via reduced monoamine oxidase A (MAOA) activity, which plays an important role in dopamine metabolism (Hampp et al., 2008).
In the midbrain, the circadian nuclear receptor REV-ERBα is a crucial modulator of mood-related behaviors (Chung et al., 2014). Rev-erbα-deficient mice exhibit mania-like behaviors characterized by decreased anxiety, a depressive-like phenotype, hyperactivity, increased risk-taking, and augmented aggression. Pharmacological inhibition of REV-ERB activity, specifically in the ventral midbrain, induces similar behavioral patterns and hyperdopaminergic states. REV-ERB α directly represses tyrosine hydroxylase (TH) expression, which is a rate-limiting enzyme for DA biosynthesis, through competition with nuclear receptor-related 1 protein (NURR1; an essential nuclear receptor for DAergic neuronal function). This interaction produces the circadian rhythmicity of the DAergic system. REV-ERBα inhibits TH gene expression through HDAC3 recruitment and histone acetylation regulation according to the circadian rhythm. These mechanisms contribute to the circadian nature of the DAergic system and mood regulation (Fig. 1).
Fig. 1. Circadian rhythm molecular networks in midbrain DAergic neurons and mood regulation.
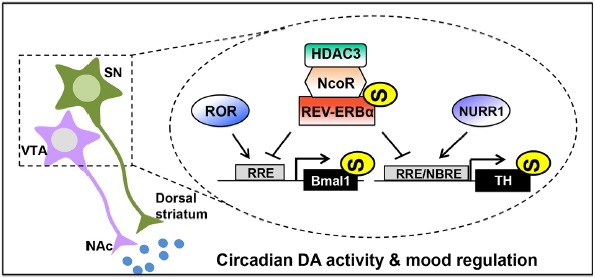
Circadian Bmal1 expression is regulated by competitive binding of REV-ERBs and RORs to RRE in the Bmal1 promoter. RRE, the cis-element recognized by the circadian nuclear receptor REV-ERBα, overlaps with NBRE, which is recognized by the nuclear receptor NURR1 in the TH promoter. REV-ERBα antagonizes NURR1-induced activation of the TH promoter via binding to RRE/NBRE, thereby contributing to the circadian rhythmicity of the DAergic system. REV-ERBα controls circadian TH gene transcription by recruiting the corepressors N-CoR and HDAC3, which suppress TH gene transcription by negatively modifying permissive histone acetylation. SN, substantia nigra; VTA, ventral tegmental area; NAc, nucleus accumbens; DA, dopamine; TH, tyrosine hydroxylase; RORs, retinoic acid receptor-related to orphan nuclear receptors; RRE, ROR/REV-ERB response element; NBRE, NGFI-B response element; N-CoR, nuclear receptor co-repressor 1; HDAC3, histone deacetylase 3; NURR1, nuclear receptor related 1 protein.
MIDBRAIN DOPAMINERGIC NEURONAL CIRCUIT AND FUNCTION
Mood is controlled by complex neural circuits and various neurotransmitters. Many brain regions that contribute to mood, including the hippocampus, prefrontal cortex (PFC), VTA, nucleus accumbens (NAc), amygdala, hypothalamus, and lateral habenula, interact with each other via circuits in the DAergic, noradrenergic, serotonergic, glutamatergic, and GABAergic pathways (Nestler and Carlezon, 2006).
The DAergic system is implicated in mood regulation (Chung et al., 2014; Hampp et al., 2008; Roybal et al., 2007). DAergic neurons in the VTA innervate the PFC and the NAc, which are referred to as the mesocortical and mesolimbic pathways, respectively. The mesocorticolimbic pathway is important for the control of motivation, emotion, and reward functions (Renard et al., 2001; Wise, 1998; Yadid et al., 2001). Abnormalities in this circuit induce addiction, affective disorders, schizophrenia, and attention-deficit hyperactive disorder (Grace, 2016; Hyman and Malenka, 2001; Volkow et al., 2009). The nigrostriatal pathway from the substantia nigra (SN) to dorsal striatum (also known as the caudate-putamen) is another DAergic pathway associated with motor function; degeneration of this circuit can induce PD (Cheng et al., 2010).
DAergic neurons in the SN and VTA have distinct anatomical, molecular, and electrophysiological characteristics, though some projection patterns have been shown to overlap (Brichta and Greengard, 2014). Few DAergic projections originating from the SN innervate the ventral striatum, while some VTA projections innervate the dorsal striatum. DAergic projections to the amygdala and PFC come from both SN and VTA (Björklund and Dunnett, 2007). The SN and VTA receive afferent inputs from brain regions involved in mood regulation, such as the central amygdala and dorsal raphe (Watabe-Uchida et al., 2012). Based on the partially shared input-output networks, we speculate that the functions of the SN and VTA may not be mutually exclusive.
Notably, both the SN and VTA harbor functional clockwork, and the key activities of these regions oscillate in a circadian manner. Core clock genes, including Bmal1, Per2 and Rev-erbα, in the ventral midbrain exhibit robust circadian oscillations (Chung et al., 2014). TH expression is also circadian (Chung et al., 2014; Webb et al., 2009; Weber et al., 2004). Additionally, the electrophysiological activities of the VTA demonstrate a robust diurnal pattern (Dominguez-Lopez et al., 2014; Luo and Aston-Jones, 2009). These suggest that SN and VTA function are under circadian rhythm control.
CIRCADIAN DISRUPTION AND MOOD DISORDERS IN PARKINSON’S DISEASE
PD is a neurodegenerative disorder that results from DAergic neuronal death in the SNpc (Dauer and Przedborski, 2003), resulting in motor deficits as well as non-motor symptoms, such as mood dysregulation. DAergic neuron degeneration is more concentrated in the SNpc ventral tier than the SNpc dorsal tier and VTA, suggesting that SN and VTA DAergic neurons have different susceptibilities to degeneration in PD (Brichta and Greengard, 2014). Ventral SNpc DAergic neurons innervate not only the striatum (Cebrián and Prensa, 2010) but also the PFC (Björklund and Dunnett, 2007), which is implicated in PD with depression (Aarsland et al., 2011). Thus, the SNpc ventral tier may be implicated in both the motor and non-motor symptoms of PD. Approximately 45% of patients with PD experience depression (Burn, 2002; Chaudhuri and Schapira, 2009), and 50% have comorbid anxiety (Brown et al., 2011). Although the etiology of mood disorders in PD is unclear, dysfunctional monoaminergic neurotransmission is widely observed (Braak et al., 2003; Halliday et al., 1990; Zarow et al., 2003). However, direct evidence for the contribution of serotonin to mood disturbances in PD is lacking (Leentjens et al., 2006). Selective serotonin reuptake inhibitors (SSRIs), classic antidepressants, lack efficacy in patients with PD (Weintraub et al., 2005). Levodopa (L-DOPA) treatment, a well-known antiparkinsonian drug, does not improve anxiety and depression in patients with PD (Kim et al., 2009; Richard et al., 2004). Pramipexole (DA agonist) and nortriptyline (tricyclic antidepressant that acts as a serotonin and norepinephrine reuptake inhibitor but has side effects) are the only effective drugs for depression in PD (Aarsland et al., 2011). Therefore, therapeutic approaches that control the upstream regulators of the DA system, such as REV-ERBα, may hold promise.
Local inhibition of REV-ERBα activity by injection of a potent REV-ERBα antagonist SR8278 (Kojetin et al., 2011) into the ventral midbrain induces a hyperdopaminergic state, caused by disappearance of REV-ERBα repression of TH expression. SR8278-injected mice also exhibit mania-like behaviors, including decreased depressive- and anxiety-like phenotypes at dusk, leading to disappearance of the circadian pattern of mood-related behaviors (Chung et al., 2014; Fig. 2A). Conversely, 6-hydroxydopamine (6-OHDA)-injected mice (animal model of PD) exhibit a hypodopaminergic state owing to DAergic neuronal death in the SNpc and increased depressive and anxiety-like phenotypes at dawn, resulting in disruption of the circadian pattern of mood-related behaviors (unpublished data: Fig. 2A). Changes in mood-related behaviors in SR8278- and 6-OHDA-injected animals can be dissociated from locomotor dysfunction. SR8278-infused mice show an array of mania-like behaviors that cannot be attributed to increased locomotion (Chung et al., 2014). Mice injected unilaterally with 6-OHDA intrastriatally exhibit motor coordination problems, though not regarding voluntary locomotion (Heuer et al., 2012; Roedter et al., 2001). These suggest that appropriate circadian oscillation of DA levels is necessary to elicit circadian mood-related behaviors (Fig. 2B).
Fig. 2. Comparison of mood-related behaviors and DA levels in vehicle (VEH)- and SR8278 (REV-ERBα antagonist)-injected mice, and VEH- and 6-hydroxydopamine (6-OHDA)-treated mice.
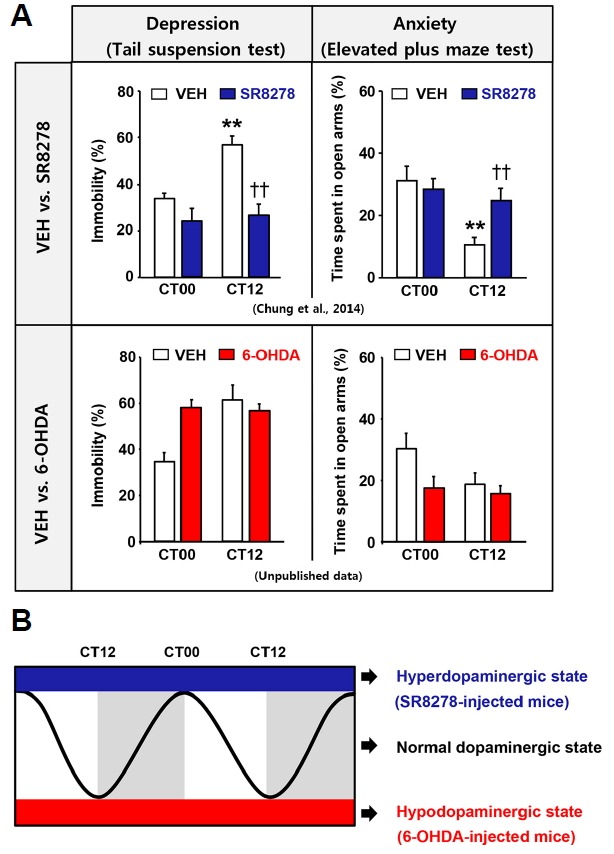
(A) Pharmacological REV-ERBα activity inhibition by microinfusion of SR8278 into the ventral midbrain (VMB) of wild type (WT) mice induced mania-like behaviors, such as decreased depression-and anxiety-like phenotypes at dusk (circadian time 12 h [CT12]) with disappearance of daily variation of mood-related behaviors in the tail suspension test (TST) and elevated plus maze test (EPM) (Chung et al., 2014). 6-OHDA-treated mice exhibited disruption of daily rhythmic mood-related behaviors toward increased depression- and anxiety-like behaviors at dawn (CT00) in the TST and EPM (unpublished data). **p < 0.01, CT00 vs. CT12; ††p < 0.01, VEH vs. SR8278. (B) Microinfusion of SR8278 into the VMB induced a hyperdopaminergic state without daily oscillation of DA, which was caused by loss of the repressive function of REV-ERBα on TH expression. Conversely, 6-OHDA-treated mice showed a hypodopaminergic state owing to DAergic neuronal death in the SNpc. Hyperdopaminergic and hypodopaminergic states can induce mood dysfunction. Therefore, appropriate levels of DA with circadian oscillation may be important to sustain circadian mood-related behaviors.
Mood disorder, which is often accompanied by sundown syndrome, and disruption of circadian rhythmicity are both common in neurodegenerative diseases, including Alzheimer’s disease (AD) and PD (Bliwise et al., 1995; Wulff et al., 2010). Clinical studies have shown disrupted rest-activity cycles, changes in blood pressure and heart rate rhythm, abnormal hormone secretion, and non-motor symptoms in PD (Videnovic and Willis, 2016). Conversely, circadian abnormalities in PD may also influence pathological processes, e.g., circadian disruption exacerbates motor deficits and DAergic neuronal loss in the MPTP (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine) model of PD (Lauretti et al., 2017). Circadian clock protein deficiency leads to vulnerability to oxidative injury and neurodegeneration in various animal models (Musiek, 2015). Thus, the mechanisms linking molecular clocks and various non-motor symptoms and neurodegeneration in PD should be investigated further. Clock-targeted therapeutics might be beneficial for treating neurodegenerative diseases.
CONCLUSIONS
We reviewed the role of the circadian clock in mood-related behaviors in healthy and pathological states. Pharmacological treatments to improve mood disorders, such as SSRIs, lithium, or valproic acid, shift the phase or modulate the circadian rhythm period (Johnsson et al, 1983; McClung et al., 2007; Sprouse et al., 2006). Lithium, a mood stabilizer used to treat BPD, inhibits glycogen synthase kinase-3 beta (GSK3β), which phosphorylates and stabilizes REV-ERBα (Yin et al., 2006). REV-ERBα influences mood regulation through circadian control of the DAergic system (Chung et al., 2014), suggesting that REV-ERBα is a potential therapeutic target for PD with comorbid mood disorders. Small molecules targeting the molecular clock, e.g., a synthetic antagonist of REV-ERB (Kojetin et al., 2011), are being evaluated. REV-ERB agonists alter circadian gene expression in various peripheral tissues and brain regions, and cause diverse physiological changes, especially in sleep architecture and emotional behaviors in mice (Banerjee et al., 2014; Solt et al., 2012). Future studies should characterize the mechanisms of diverse clock-targeting molecules that have therapeutic potential for treating clock-related diseases.
While we mainly highlighted the contribution of the DAergic system to the circadian rhythm of mood regulation and its implication in mood disorders accompanied with PD, we cannot rule out the contribution of other brain circuits. For example, the lateral preoptic area and paraventricular nucleus are enriched with DAergic neurons regulated by photoperiods (Dulcis et al., 2013). The lateral habenula may act as a link between the circadian DAergic system and daily mood regulation in terms of regulating DA release through projections to the SN and VTA (Hikosaka, 2010). Other candidates include the bed nucleus of the stria terminalis, central amygdala, basolateral amygdala, dentate gyrus, and paraventricular thalamic nucleus (Amir and Stewart, 2009; Colavito et al., 2015). Therefore, it is important to understand the role of circadian rhythm in other neuronal circuits and brain regions involved in mood regulation.
ACKNOWLEDGMENTS
This work was supported by the National Research Foundation of Korea (NRF) funded by the Ministry of Science, ICT & Future Planning (NRF-2017R1A2A1A05001351); and the DGIST MIREBraiN Program of the Ministry of Science, ICT & Future Planning (2017010095 and 2017040002).
REFERENCES
- Aarsland D., Påhlhagen S., Ballard C.G., Ehrt U., Svenningsson P. Depression in Parkinson disease--epidemiology, mechanisms and management. Nat Rev Neurol. 2011;8:35–47. doi: 10.1038/nrneurol.2011.189. [DOI] [PubMed] [Google Scholar]
- Abarca C., Albrecht U., Spanagel R. Cocaine sensitization and reward are under the influence of circadian genes and rhythm. Proc Natl Acad Sci USA. 2002;99:9026–9030. doi: 10.1073/pnas.142039099. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Akhtar R.A., Reddy A.B., Maywood E.S., Clayton J.D., King V.M., Smith A.G., Gant T.W., Hastings M.H., Kyriacou C.P. Circadian cycling of the mouse liver transcriptome, as revealed by cDNA microarray, is driven by the suprachiasmatic nucleus. Curr Biol. 2002;12:540–550. doi: 10.1016/s0960-9822(02)00759-5. [DOI] [PubMed] [Google Scholar]
- Albrecht U. Circadian clocks and mood-related behaviors. Handb Exp Pharmacol. 2013;217:227–239. doi: 10.1007/978-3-642-25950-0_9. [DOI] [PubMed] [Google Scholar]
- Albrecht U. Molecular mechanisms in mood regulation involving the circadian clock. Front Neurol. 2017;8:30. doi: 10.3389/fneur.2017.00030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Amir S., Stewart J. Motivational modulation of rhythms of the expression of the clock protein PER2 in the limbic forebrain. Biol Psychiatry. 2009;65:829–834. doi: 10.1016/j.biopsych.2008.12.019. [DOI] [PubMed] [Google Scholar]
- Balsalobre A., Damiola F., Schibler U. A serum shock induces circadian gene expression in mammalian tissue culture cells. Cell. 1998;93:929–937. doi: 10.1016/s0092-8674(00)81199-x. [DOI] [PubMed] [Google Scholar]
- Banerjee S., Wang Y., Solt L.A., Griffett K., Kazantzis M., Amador A., El-Gendy B.M., Huitron-Resendiz S., Roberts A.J., Shin Y., et al. Pharmacological targeting of the mammalian clock regulates sleep architecture and emotional behaviour. Nat Commun. 2014;5:5759. doi: 10.1038/ncomms6759. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bedrosian T.A., Nelson R.J. Sundowning syndrome in aging and dementia: research in mouse models. Exp Neurol. 2013;243:67–73. doi: 10.1016/j.expneurol.2012.05.005. [DOI] [PubMed] [Google Scholar]
- Björklund A., Dunnett S.B. Dopamine neuron systems in the brain: an update. Trends Neurosci. 2007;30:194–202. doi: 10.1016/j.tins.2007.03.006. [DOI] [PubMed] [Google Scholar]
- Bliwise D.L., Watts R.L., Watts N., Rye D.B., Irbe D., Hughes M. Disruptive nocturnal behavior in Parkinson’s disease and Alzheimer’s disease. J Geriatr Psychiatry Neurol. 1995;8:107–110. doi: 10.1177/089198879500800206. [DOI] [PubMed] [Google Scholar]
- Braak H., Del Tredici K., Rüb U., de Vos R.A., Jansen Steur E.N., Braak E. Staging of brain pathology related to sporadic Parkinson’s disease. Neurobiol Aging. 2003;24:197–211. doi: 10.1016/s0197-4580(02)00065-9. [DOI] [PubMed] [Google Scholar]
- Brichta L., Greengard P. Molecular determinants of selective dopaminergic vulnerability in Parkinson’s disease: an update. Front Neuroanat. 2014;8:152. doi: 10.3389/fnana.2014.00152. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Brown R.G., Landau S., Hindle J.V., Playfer J., Samuel M., Wilson K.C., Hurt C.S., Anderson R.J., Carnell J., Dickinson L., et al. Depression and anxiety related subtypes in Parkinson’s disease. J Neurol Neurosurg Psychiatry. 2011;82:803–809. doi: 10.1136/jnnp.2010.213652. [DOI] [PubMed] [Google Scholar]
- Bunger M.K., Wilsbacher L.D., Moran S.M., Clendenin C., Radcliffe L.A., Hogenesch J.B., Simon M.C., Takahashi J.S., Bradfield C.A. Mop3 is an essential component of the master circadian pacemaker in mammals. Cell. 2000;103:1009–1017. doi: 10.1016/s0092-8674(00)00205-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Burn D.J. Beyond the iron mask; towards better recognition and treatment of depression associated with Parkinson’s disease. Mov Disord. 2002;17:445–454. doi: 10.1002/mds.10114. [DOI] [PubMed] [Google Scholar]
- Cebrián C., Prensa L. Basal ganglia and thalamic input from neurons located within the ventral tier cell cluster region of the substantia nigra pars compacta in the rat. J Comp Neurol. 2010;518:1283–1300. doi: 10.1002/cne.22275. [DOI] [PubMed] [Google Scholar]
- Chaudhuri K.R., Schapira A.H. Non-motor symptoms of Parkinson’s disease: dopaminergic pathophysiology and treatment. Lancet Neurol. 2009;8:464–474. doi: 10.1016/S1474-4422(09)70068-7. [DOI] [PubMed] [Google Scholar]
- Cheng H.C., Ulane C.M., Burke R.E. Clinical progression in Parkinson disease and the neurobiology of axons. Ann Neurol. 2010;67:715–725. doi: 10.1002/ana.21995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chung S., Lee E.J., Yun S., Choe H.K., Park S.B., Son H.J., Kim K.S., Dluzen D.E., Lee I., Hwang O., et al. Impact of circadian nuclear receptor REV-ERBα on midbrain dopamine production and mood regulation. Cell. 2014;157:858–868. doi: 10.1016/j.cell.2014.03.039. [DOI] [PubMed] [Google Scholar]
- Colavito V., Tesoriero C., Wirtu A.T., Grassi-Zucconi G., Bentivoglio M. Limbic thalamus and state-dependent behavior: The paraventricular nucleus of the thalamic midline as a node in circadian timing and sleep/wake-regulatory networks. Neurosci Biobehav Rev. 2015;54:3–17. doi: 10.1016/j.neubiorev.2014.11.021. [DOI] [PubMed] [Google Scholar]
- Dauer W., Przedborski S. Parkinson’s disease: mechanisms and models. Neuron. 2003;39:889–909. doi: 10.1016/s0896-6273(03)00568-3. [DOI] [PubMed] [Google Scholar]
- Dibner C., Schibler U., Albrecht U. The mammalian circadian timing system: organization and coordination of central and peripheral clocks. Annu Rev Physiol. 2010;72:517–549. doi: 10.1146/annurev-physiol-021909-135821. [DOI] [PubMed] [Google Scholar]
- Domínguez-López S., Howell R.D., López-Canúl M.G., Leyton M., Gobbi G. Electrophysiological characterization of dopamine neuronal activity in the ventral tegmental area across the light-dark cycle. Synapse. 2014;68:454–467. doi: 10.1002/syn.21757. [DOI] [PubMed] [Google Scholar]
- Dulcis D., Jamshidi P., Leutgeb S., Spitzer N.C. Neurotransmitter switching in the adult brain regulates behavior. Science. 2013;340:449–453. doi: 10.1126/science.1234152. [DOI] [PubMed] [Google Scholar]
- Easton A., Arbuzova J., Turek F.W. The circadian Clock mutation increases exploratory activity and escape-seeking behavior. Genes Brain Behav. 2003;2:11–19. doi: 10.1034/j.1601-183x.2003.00002.x. [DOI] [PubMed] [Google Scholar]
- Gallego M., Virshup D.M. Post-translational modifications regulate the ticking of the circadian clock. Nat Rev Mol Cell Biol. 2007;8:139–148. doi: 10.1038/nrm2106. [DOI] [PubMed] [Google Scholar]
- Gekakis N., Staknis D., Nguyen H.B., Davis F.C., Wilsbacher L.D., King D.P., Takahashi J.S., Weitz C.J. Role of the CLOCK protein in the mammalian circadian mechanism. Science. 1998;280:1564–1569. doi: 10.1126/science.280.5369.1564. [DOI] [PubMed] [Google Scholar]
- Gloston G.F., Yoo S.H., Chen Z. Clock-enhancing small molecules and potential applications in chronic diseases and aging. Front Neurol. 2017;8:100. doi: 10.3389/fneur.2017.00100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Grace A.A. Dysregulation of the dopamine system in the pathophysiology of schizophrenia and depression. Nat Rev Neurosci. 2016;17:524–532. doi: 10.1038/nrn.2016.57. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Guillaumond F., Dardente H., Giguère V., Cermakian N. Differential control of Bmal1 circadian transcription by REV-ERB and ROR nuclear receptors. J Biol Rhythms. 2005;20:391–403. doi: 10.1177/0748730405277232. [DOI] [PubMed] [Google Scholar]
- Halliday G.M., Li Y.W., Blumbergs P.C., Joh T.H., Cotton R.G., Howe P.R., Blessing W.W., Geffen L.B. Neuropathology of immunohistochemically identified brainstem neurons in Parkinson’s disease. Ann Neurol. 1990;27:373–385. doi: 10.1002/ana.410270405. [DOI] [PubMed] [Google Scholar]
- Hampp G., Ripperger J.A., Houben T., Schmutz I., Blex C., Perreau-Lenz S., Brunk I., Spanagel R., Ahnert-Hilger G., Meijer J.H., et al. Regulation of monoamine oxidase A by circadian-clock components implies clock influence on mood. Curr Biol. 2008;18:678–683. doi: 10.1016/j.cub.2008.04.012. [DOI] [PubMed] [Google Scholar]
- Heuer A., Smith G.A., Lelos M.J., Lane E.L., Dunnett S.B. Unilateral nigrostriatal 6-hydroxydopamine lesions in mice I: motor impairments identify extent of dopamine depletion at three different lesion sites. Behav Brain Res. 2012;228:30–43. doi: 10.1016/j.bbr.2011.11.027. [DOI] [PubMed] [Google Scholar]
- Hikosaka O. The habenula: from stress evasion to valuebased decision-making. Nat Rev Neurosci. 2010;11:503–513. doi: 10.1038/nrn2866. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hyman S.E., Malenka R.C. Addiction and the brain: the neurobiology of compulsion and its persistence. Nat Rev Neurosci. 2001;2:695–703. doi: 10.1038/35094560. [DOI] [PubMed] [Google Scholar]
- Johnsson A., Engelmann W., Pflug B., Klemke W. Period lengthening of human circadian rhythms by lithium carbonate, a prophylactic for depressive disorders. Int J Chronobiol. 1983;8:129–147. [PubMed] [Google Scholar]
- Kim H.J., Park S.Y., Cho Y.J., Hong K.S., Cho J.Y., Seo S.Y., Lee D.H., Jeon B.S. Nonmotor symptoms in de novo Parkinson disease before and after dopaminergic treatment. J Neurol Sci. 2009;287:200–204. doi: 10.1016/j.jns.2009.07.026. [DOI] [PubMed] [Google Scholar]
- King D.P., Zhao Y., Sangoram A.M., Wilsbacher L.D., Tanaka M., Antoch M.P., Steeves T.D., Vitaterna M.H., Kornhauser J.M., Lowrey P.L., et al. Positional cloning of the mouse circadian clock gene. Cell. 1997;89:641–653. doi: 10.1016/s0092-8674(00)80245-7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kojetin D., Wang Y., Kamenecka T.M., Burris T.P. Identification of SR8278, a synthetic antagonist of the nuclear heme receptor REV-ERB. ACS Chem Biol. 2011;6:131–134. doi: 10.1021/cb1002575. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kripke D.F., Nievergelt C.M., Joo E., Shekhtman T., Kelsoe J.R. Circadian polymorphisms associated with affective disorders. J Circadian Rhythms. 2009;7:2. doi: 10.1186/1740-3391-7-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kume K., Zylka M.J., Sriram S., Shearman L.P., Weaver D.R., Jin X., Maywood E.S., Hastings M.H., Reppert S.M. mCRY1 and mCRY2 are essential components of the negative limb of the circadian clock feedback loop. Cell. 1999;98:193–205. doi: 10.1016/s0092-8674(00)81014-4. [DOI] [PubMed] [Google Scholar]
- Lauretti E., Di Meco A., Merali S., Praticò D. Circadian rhythm dysfunction: a novel environmental risk factor for Parkinson’s disease. Mol Psychiatry. 2017;22:280–286. doi: 10.1038/mp.2016.47. [DOI] [PubMed] [Google Scholar]
- Leentjens A.F., Scholtissen B., Vreeling F.W., Verhey F.R. The serotonergic hypothesis for depression in Parkinson’s disease: an experimental approach. Neuropsychopharmacology. 2006;31:1009–1015. doi: 10.1038/sj.npp.1300914. [DOI] [PubMed] [Google Scholar]
- Li J.Z., Bunney B.G., Meng F., Hagenauer M.H., Walsh D.M., Vawter M.P., Evans S.J., Choudary P.V., Cartagena P., Barchas J.D., et al. Circadian patterns of gene expression in the human brain and disruption in major depressive disorder. Proc Natl Acad Sci USA. 2013;110:9950–9955. doi: 10.1073/pnas.1305814110. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Luo A.H., Aston-Jones G. Circuit projection from suprachiasmatic nucleus to ventral tegmental area: a novel circadian output pathway. Eur J Neurosci. 2009;29:748–760. doi: 10.1111/j.1460-9568.2008.06606.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Mansour H.A., Wood J., Logue T., Chowdari K.V., Dayal M., Kupfer D.J., Monk T.H., Devlin B., Nimgaonkar V.L. Association study of eight circadian genes with bipolar I disorder, schizoaffective disorder and schizophrenia. Genes Brain Behav. 2006;5:150–157. doi: 10.1111/j.1601-183X.2005.00147.x. [DOI] [PubMed] [Google Scholar]
- McCarthy M.J., Welsh D.K. Cellular circadian clocks in mood disorders. J Biol Rhythms. 2012;27:339–352. doi: 10.1177/0748730412456367. [DOI] [PubMed] [Google Scholar]
- McClung C.A. Circadian genes, rhythms and the biology of mood disorders. Pharmacol Ther. 2007;114:222–232. doi: 10.1016/j.pharmthera.2007.02.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McClung C.A., Sidiropoulou K., Vitaterna M., Takahashi J.S., White F.J., Cooper D.C., Nestler E.J. Regulation of dopaminergic transmission and cocaine reward by the Clock gene. Proc Natl Acad Sci USA. 2005;102:9377–9381. doi: 10.1073/pnas.0503584102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Musiek E.S. Circadian clock disruption in neurodegenerative diseases: cause and effect? Front Pharmacol. 2015;27:6–29. doi: 10.3389/fphar.2015.00029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Nestler E.J., Carlezon W.A., Jr The mesolimbic dopamine reward circuit in depression. Biol Psychiatry. 2006;59:1151–1159. doi: 10.1016/j.biopsych.2005.09.018. [DOI] [PubMed] [Google Scholar]
- Panda S., Antoch M.P., Miller B.H., Su A.I., Schook A.B., Straume M., Schultz P.G., Kay S.A., Takahashi J.S., Hogenesch J.B. Coordinated transcription of key pathways in the mouse by the circadian clock. Cell. 2002;109:307–320. doi: 10.1016/s0092-8674(02)00722-5. [DOI] [PubMed] [Google Scholar]
- Preitner N., Damiola F., Lopez-Molina L., Zakany J., Duboule D., Albrecht U., Schibler U. The orphan nuclear receptor REV-ERBalpha controls circadian transcription within the positive limb of the mammalian circadian oscillator. Cell. 2002;110:251–260. doi: 10.1016/s0092-8674(02)00825-5. [DOI] [PubMed] [Google Scholar]
- Renard C.E., Fiocco A.J., Clenet F., Hascoet M., Bourin M. Is dopamine implicated in the antidepressant-like effects of selective serotonin reuptake inhibitors in the mouse forced swimming test? Psychopharmacology. 2001;159:42–50. doi: 10.1007/s002130100836. [DOI] [PubMed] [Google Scholar]
- Richard I.H., Frank S., McDermott M.P., Wang H., Justus A.W., LaDonna K.A., Kurlan R. The ups and downs of Parkinson disease: a prospective study of mood and anxiety fluctuations. Cogn Behav Neurol. 2004;17:201–207. [PubMed] [Google Scholar]
- Roedter A., Winkler C., Samii M., Walter G.F., Brandis A., Nikkhah G. Comparison of unilateral and bilateral intrastriatal 6-hydroxydopamine-induced axon terminal lesions: evidence for interhemispheric functional coupling of the two nigrostriatal pathways. J Comp Neurol. 2001;432:217–229. doi: 10.1002/cne.1098. [DOI] [PubMed] [Google Scholar]
- Roybal K., Theobold D., Graham A., DiNieri J.A., Russo S.J., Krishnan V., Chakravarty S., Peevey J., Oehrlein N., Birnbaum S., et al. Mania-like behavior induced by disruption of CLOCK. Proc Natl Acad Sci USA. 2007;104:6406–6411. doi: 10.1073/pnas.0609625104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Russo S.J., Nestler E.J. The brain reward circuitry in mood disorders. Nat Rev Neurosci. 2013;14:609–625. doi: 10.1038/nrn3381. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Shearman L.P., Sriram S., Weaver D.R., Maywood E.S., Chaves I., Zheng B., Kume K., Lee C.C., van der Horst G.T., Hastings M.H., et al. Interacting molecular loops in the mammalian circadian clock. Science. 2000;288:1013–1019. doi: 10.1126/science.288.5468.1013. [DOI] [PubMed] [Google Scholar]
- Solt L.A., Wang Y., Banerjee S., Hughes T., Kojetin D.J., Lundasen T., Shin Y., Liu J., Cameron M.D., Noel R., et al. Regulation of circadian behaviour and metabolism by synthetic REV-ERB agonists. Nature. 2012;485:62–68. doi: 10.1038/nature11030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Soria V., Martínez-Amorós E., Escaramís G., Valero J., Pérez-Egea R., García C., Gutiérrez-Zotes A., Puigdemont D., Bayés M., Crespo J.M., et al. Differential association of circadian genes with mood disorders: CRY1 and NPAS2 are associated with unipolar major depression and CLOCK and VIP with bipolar disorder. Neuropsychopharmacology. 2010;35:1279–1289. doi: 10.1038/npp.2009.230. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sprouse J., Braselton J., Reynolds L. Fluoxetine modulates the circadian biological clock via phase advances of suprachiasmatic nucleus neuronal firing. Biol Psychiatry. 2006;60:896–899. doi: 10.1016/j.biopsych.2006.03.003. [DOI] [PubMed] [Google Scholar]
- Storch K.F., Lipan O., Leykin I., Viswanathan N., Davis F.C., Wong W.H., Weitz C.J. Extensive and divergent circadian gene expression in liver and heart. Nature. 2002;417:78–83. doi: 10.1038/nature744. [DOI] [PubMed] [Google Scholar]
- Videnovic A., Willis G.L. Circadian system - A novel diagnostic and therapeutic target in Parkinson’s disease? Mov. Disord. 2016;31:260–269. doi: 10.1002/mds.26509. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Videnovic A., Lazar A.S., Barker R.A., Overeem S. ‘The clocks that time us’--circadian rhythms in neurodegenerative disorders. Nat Rev Neurol. 2014;10:683–693. doi: 10.1038/nrneurol.2014.206. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Volkow N.D., Wang G.J., Kollins S.H., Wigal T.L., Newcorn J.H., Telang F., Fowler J.S., Zhu W., Logan J., Ma Y. Evaluating dopamine reward pathway in ADHD: clinical implications. JAMA. 2009;302:1084–1091. doi: 10.1001/jama.2009.1308. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Watabe-Uchida M., Zhu L., Ogawa S.K., Vamanrao A., Uchida N. Whole-brain mapping of direct inputs to midbrain dopamine neurons. Neuron. 2012;74:858–873. doi: 10.1016/j.neuron.2012.03.017. [DOI] [PubMed] [Google Scholar]
- Webb I.C., Baltazar R.M., Wang X., Pitchers K.K., Coolen L.M., Lehman M.N. Diurnal variations in natural and drug reward, mesolimbic tyrosine hydroxylase, and clock gene expression in the male rat. J Biol Rhythms. 2009;24:465–476. doi: 10.1177/0748730409346657. [DOI] [PubMed] [Google Scholar]
- Weber M., Lauterburg T., Tobler I., Burgunder J.M. Circadian patterns of neurotransmitter related gene expression in motor regions of the rat brain. Neurosci Lett. 2004;358:17–20. doi: 10.1016/j.neulet.2003.12.053. [DOI] [PubMed] [Google Scholar]
- Weintraub D., Morales K.H., Moberg P.J., Bilker W.B., Balderston C., Duda J.E., Katz I.R., Stern M.B. Antidepressant studies in Parkinson’s disease: a review and meta-analysis. Mov Disord. 2005;20:1161–1169. doi: 10.1002/mds.20555. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wirz-Justice A. Biological rhythm disturbances in mood disorders. Int Clin Psychopharmacol. 2006;21(Suppl 4):S11–S15. doi: 10.1097/01.yic.0000195660.37267.cf. [DOI] [PubMed] [Google Scholar]
- Wise R.A. Drug-activation of brain reward pathways. Drug Alcohol Depend. 1998;51:13–22. doi: 10.1016/s0376-8716(98)00063-5. [DOI] [PubMed] [Google Scholar]
- Wulff K., Gatti S., Wettstein J.G., Foster R.G. Sleep and circadian rhythm disruption in psychiatric and neurodegenerative disease. Nat Rev Neurosci. 2010;11:589–599. doi: 10.1038/nrn2868. [DOI] [PubMed] [Google Scholar]
- Yadid G., Overstreet D.H., Zangen A. Limbic dopaminergic adaptation to a stressful stimulus in a rat model of depression. Brain Res. 2001;896:43–47. doi: 10.1016/s0006-8993(00)03248-0. [DOI] [PubMed] [Google Scholar]
- Yamazaki S., Numano R., Abe M., Hida A., Takahashi R., Ueda M., Block G.D., Sakaki Y., Menaker M., Tei H. Resetting central and peripheral circadian oscillators in transgenic rats. Science. 2000;288:682–685. doi: 10.1126/science.288.5466.682. [DOI] [PubMed] [Google Scholar]
- Yin L., Lazar M.A. The orphan nuclear receptor Rev-erbα recruits the N-CoR/histone Deacetylase 3 corepressor to regulate the circadian Bmal1 gene. Mol Endocrinol. 2005;19:1452–1459. doi: 10.1210/me.2005-0057. [DOI] [PubMed] [Google Scholar]
- Yin L., Wang J., Klein P.S., Lazar M.A. Nuclear receptor Rev-erbalpha is a critical lithium-sensitive component of the circadian clock. Science. 2006;311:1002–1005. doi: 10.1126/science.1121613. [DOI] [PubMed] [Google Scholar]
- Zarow C, Lyness S.A., Mortimer J.A., Chui H.C. Neuronal loss is greater in the locus coeruleus than nucleus basalis and substantia nigra in Alzheimer and Parkinson diseases. Arch Neurol. 2003;60:337–341. doi: 10.1001/archneur.60.3.337. [DOI] [PubMed] [Google Scholar]
- Zhang R., Lahens N.F., Balance H.I., Hughes M.E., Hogenesch J.B. A circadian gene expression atlas in mammals: implications for biology and medicine. Proc Natl Acad Sci USA. 2014;111:16219–16224. doi: 10.1073/pnas.1408886111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zhao X., Hirota T., Han X., Cho H., Chong L.W., Lamia K., Liu S., Atkins A.R., Banayo E., Liddle C., et al. Circadian amplitude regulation via FBXW7-targeted REV-ERBα degradation. Cell. 2016;165:1644–1657. doi: 10.1016/j.cell.2016.05.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Zheng B., Larkin D.W., Albrecht U., Sun Z.S., Sage M., Eichele G., Lee C.C., Bradley A. The mPer2 gene encodes a functional component of the mammalian circadian clock. Nature. 1999;400:169–173. doi: 10.1038/22118. [DOI] [PubMed] [Google Scholar]
- Zheng B., Albrecht U., Kaasik K., Sage M., Lu W., Vaishnav S., Li Q., Sun Z.S., Eichele G., Bradley A., et al. Nonredundant roles of the mPer1 and mPer2 genes in the mammalian circadian clock. Cell. 2001;105:683–694. doi: 10.1016/s0092-8674(01)00380-4. [DOI] [PubMed] [Google Scholar]
- Zhu B., Gates L.A., Stashi E., Dasgupta S., Gonzales N., Dean A., Dacso C.C., York B., O’Malley B.W. Coactivatordependent oscillation of chromatin accessibility dictates circadian gene amplitude via REV-ERB loading. Mol Cell. 2015;60:769–783. doi: 10.1016/j.molcel.2015.10.024. [DOI] [PMC free article] [PubMed] [Google Scholar]