

Abstract
Background
177Lu-octreotate can be used to treat somatostatin receptor expressing neuroendocrine tumors. It is highly effective in animal models, but clinical studies have so far only demonstrated low cure rates. Hedgehog inhibitors have shown therapeutic effect as monotherapy in neuroendocrine tumor model systems and might be one option to enhance the efficacy of 177Lu-octreotate therapy. The aim of this study was to determine the therapeutic effect of combination therapy using 177Lu-octreotate and the Hedgehog signaling pathway inhibitor sonidegib.
Methods
GOT1-bearing BALB/c nude mice were treated with either sonidegib (80 mg/kg twice a week via oral gavage), a single injection of 30 MBq 177Lu-octreotate i.v., or a combination of both. Untreated animals served as controls. Tumor size was measured twice-weekly using calipers. The animals were killed 41 d after injection followed by excision of the tumors. Total RNA was extracted from each tumor sample and then subjected to gene expression analysis. Gene expression patterns were compared with those of untreated controls using Nexus Expression 3.0, IPA and Gene Ontology terms. Western blot was carried out on total protein extracted from the tumor samples to analyze activation-states of the Hh and PI3K/AKT/mTOR pathways.
Results
Sonidegib monotherapy resulted in inhibition of tumor growth, while a significant reduction in mean tumor volume was observed after 177Lu-octreotate monotherapy and combination therapy. Time to progression was prolonged in the combination therapy group compared with 177Lu-octreotate monotherapy. Gene expression analysis revealed a more pronounced response following combination therapy compared with both monotherapies, regarding the number of regulated genes and biological processes. Several cancer-related signaling pathways (i.e. Wnt/β-catenin, PI3K/AKT/mTOR, G-protein coupled receptor, and Notch) were affected by the combination therapy, but not by either monotherapy. Protein expression analysis revealed an activation of the Hh- and PI3K/AKT/mTOR pathways in tumors exposed to 177Lu-octreotate monotherapy and combination therapy.
Conclusions
A comparative analysis of the different treatment groups showed that combination therapy using sonidegib and 177Lu-octreotate could be beneficial to patients with neuroendocrine tumors. Gene expression analysis revealed a functional interaction between sonidegib and 177Lu-octreotate, i.e. several cancer-related signaling pathways were modulated that were not affected by either monotherapy. Protein expression analysis indicated a possible PI3K/AKT/mTOR-dependent activation of the Hh pathway, independent of SMO.
Electronic supplementary material
The online version of this article (doi:10.1186/s12885-017-3524-x) contains supplementary material, which is available to authorized users.
Keywords: Radionuclide therapy, radiation biology, Odomzo, LDE225, 177Lu-DOTATATE, GEPNET, midgut carcinoid, radiogenomics, radiosensitizer, PRRT
Background
Neuroendocrine tumors (NETs) are the most common malignancies of the small intestine, and incidence rates are increasing [1, 2]. NETs are a heterogeneous group of malignant neoplasms frequently associated with the synthesis and secretion of peptides and amines causing hormone overproduction symptoms (e.g. carcinoid syndrome). However, NETs are slow-proliferating tumors, and symptoms are seldom evident until at a relatively late stage [3]. Surgery is currently the only curative treatment option for patients with localized NET. However, palliative treatment of NET metastases can be achieved by administration of somatostatin analogs to patients having tumors with high expression of somatostatin receptors (SSTR) [3]. Peptide receptor radionuclide therapy (PRRT) with radiolabeled somatostatin analogue 177Lu-[DOTA0, Tyr3]-octreotate (177Lu-octreotate or 177Lu-DOTATATE) is another therapeutic option for patients with SSTR-expressing tumors. This treatment has shown successful results regarding tolerability, tumor regression, increased overall survival, and improved quality of life in patients with inoperable disease [4–7]. However, the treatment is limited by the risk organs bone marrow and kidneys, which restrict the amount of 177Lu-octreotate administered to the patients. Complete tumor remission is rare and attempts to increase treatment effects using 177Lu-octreotate in combination with other systemic treatments (limited by different risk organs), have been performed, with varying success rates [8–10].
The Hedgehog (Hh) pathway is a major developmental signaling pathway, which regulates both proliferation and differentiation of various types of stem cells during embryogenesis [11]. In the absence of Hh ligands, the Hh receptor Patched inhibits activity of the transmembrane protein Smoothened (SMO) [12]. Binding of Hh ligand to Patched results in accumulation of SMO in the primary cilium and activation of transcription factors GLI1, GLI2 (activators) and GLI3 (repressor) [12]. When activated, the GLI proteins translocate into the nucleus and regulate transcription of genes involved in, e.g. cell cycle regulation, cell adhesion, signal transduction, angiogenesis, and apoptosis [13, 14]. Defective Hh signaling has been implicated in various types of human cancers [15], and several components of the Hh pathway have been studied and proposed as targets for cancer treatment [12–14, 16]. Hh signaling has been shown to be activated in NETs, and treatment with Hh inhibitors have resulted in reduced cell viability in vitro [17–19]. Since the Hh pathway is important in cancer initiation and development, it may also be important for tumor radioresistance and regrowth after treatment with ionizing radiation. Preclinically, Hh signaling has been shown to promote radiation resistance, and increased anti-tumor effects have been observed when combining ionizing radiation and Hh inhibitors [2, 13, 20]. Sonidegib (also known as Odomzo®, erismodegib or NVP-LDE225) is a selective and orally bioavailable antagonist of SMO [21], which has previously shown an anti-tumor effect in neuroendocrine tumor models [22]. It has received FDA approval for treatment of basal cell carcinoma, and is currently being investigated as a potential treatment for various cancers (e.g. small cell lung cancer) [23]. Sonidegib treatment is generally well tolerated, but doses are limited by elevations in the concentrations of creatine kinase [16]. Common side effects include neutropenia, anemia and loss of taste sensation [23, 24].
We have previously established a human small intestine NET cell line (GOT1) derived from a surgically removed liver metastasis [25]. The GOT1 cells have retained characteristic properties of NETs, such as expression of SSTR2 and SSTR5, and can be successfully xenotransplanted to nude mice [26]. In addition, it has previously been shown that 177Lu-octreotate induces cell cycle arrest, apoptosis and dose dependent tumor volume reduction in GOT1 tumors [27–29].
Considering the promising results of both Hh pathway inhibitors and 177Lu-octreotate in NET model systems [19, 22, 30], we hypothesized that inhibition of hedgehog signaling in NETs would increase the efficacy of 177Lu-octreotate treatment. The aim of this study was to test this hypothesis by investigating the therapeutic effects of combined treatment with the Hh inhibitor sonidegib and 177Lu-octreotate, compared with those of the two monotherapies consisting of either sonidegib or 177Lu-octreotate, in GOT1 human small intestine NETs in nude mice.
Methods
Tumor and animal model
GOT1 tumor tissue samples were transplanted s.c. in the neck of 4-week-old female BALB/c nude mice (CAnN.Cg-Foxn1nu/Crl, Charles River, Japan and Germany) as previously described [31]. During transplantation, animals were anesthetized using i.p. injection of Domitor® vet. (1 mg/ml injection solution, Orion Pharma Animal Health, Sweden) and Ketaminol® vet. (50 mg/ml injection solution, Intervet AB, Sweden). Antisedan (5 mg/ml injection solution, Orion Pharma Animal Health, Sweden) was injected i.p. after transplantation as antidote. Drinking water and autoclaved food were provided ad libitum.
Pharmaceuticals
Sonidegib was purchased from Active Biochemicals Co., Limited (Hong Kong, China) and dissolved in DMSO as per manufacturer’s instructions.
177LuCl3 and [DOTA0, Tyr3]-octreotate were purchased from the Nuclear Research & Consultancy Group (IDB Holland, the Netherlands). Preparation and radiolabeling were conducted per the manufacturer’s instructions. Instant thin layer chromatography (ITLCTM SG, PALL Corporation, USA) was used for quality control, with the mobile phase consisting of 0.1 M sodium citrate (pH 5; VWR International AB, Sweden). The fraction of peptide-bound 177Lu was >98% and the specific activity was approximately 26 MBq/μg octreotate. Saline solution was used to dilute the 177Lu-octreotate stock solution to the desired activity concentration for administration. 177Lu activity in syringes was measured before and after injection using a well-type ionization chamber (CRC-15R; Capintec, IA, USA).
Study design
In total, 21 GOT1 tumor-bearing mice were included in the study (Table 1). Tumor volumes varied between 0.1 and 2.5 ml (measured with slide calipers) at the start of experiments and an effort was made to obtain similar tumor size distributions in all experimental groups. Ten animals were divided into two treatment groups (n = 5/group). One group was treated with sonidegib (80 mg/kg body weight twice a week via oral gavage), while another group received both sonidegib (following the same treatment schedules as the monotherapy group) and an injection of 30 MBq 177Lu-octreotate (a non-curative treatment) into the tail vein. Tumor growth in the treatment groups was compared with that of animals receiving 30 MBq 177Lu-octreotate monotherapy (n = 5) and control animals injected with saline solution (n = 6), which have been characterized in a previous study [29]. During the study period, tumor size measurements were performed twice-weekly using digital slide calipers. Animals were killed 41 days after treatment start using i.p. injection of Pentobarbitalnatrium vet. (60 mg/ml, Apotek Produktion & Laboratorier AB, Sweden), followed by cardiac puncture. Tumor tissue samples were excised and instantly frozen in liquid nitrogen for gene expression analysis.
Table 1.
Number of GOT1-bearing mice used in each analysis after treatment with sonidegib, 177Lu-octreotate, or a combination of both pharmaceuticals, and in control animals
| Sonidegib | 177Lu-octreotate | Sonidegib +177Lu-octreotate | Control | |
|---|---|---|---|---|
| Tumor volume measurements | 5 | 5a | 5 | 6a |
| Dosimetric calculations | - | 5 | 5 | - |
| Gene expression analysis | 3 | 3 | 3 | 3a |
| Protein expression analysis | 3 | 3 | 3 | 3 |
aData has been characterized in a previous publication [29]
Dosimetry
The mean absorbed dose, D(r T, T D), to the target tissue, r T, was calculated according to the Medical Internal Radiation Dose Committee (MIRD) pamphlet 21 formalism [32]:
where is the time-integrated activity in source tissue, r S, over dose-integration period, T D , and M(r T, T D) is the mass of the target tissue, r T. values for 177Lu activity were determined in the tumor samples using activity concentration data presented by Dalmo et al. using GOT1 tumor samples after injection of 15 MBq 177Lu-octreotate [29]. The mean energy emitted per nuclear decay i, ∑iEiYi, was approximated to 147.9 keV/decay [33], including β-particles, Auger and conversion electrons. The absorbed fraction, ϕ(r T ← r S, E i, T D), was set to 1 for all tumors, and r T was set to be the same as r S in all calculations.
RNA extraction and analysis
Gene expression microarray analysis was performed using RNA from three tumor samples per group (treated and control, for a total of 12 animals). Frozen tumor tissue was homogenized with the TissueLyser LT (Qiagen, Hilden, Germany) and total RNA was extracted using the RNeasy Lipid Tissue Mini Kit (Qiagen, Hilden, Germany) per the manufacturer’s instructions.
RNA concentration and purity were determined using an ND-1000 Spectrophotometer (NanoDrop Technologies, Wilmington, DE, USA). RNA integrity was validated with the RNA 6000 Nano LabChip Kit and Agilent 2100 Bioanalyzer (Agilent Technologies, Palo Alto, CA, USA). RNA integrity number (RIN) values higher than 8.1 were used in the present investigation.
Hybridization of the RNA samples was performed at Swegene Center for Integrative Biology (SCIBLU, Lund University, Sweden) on Illumina HumanHT-12 v4 Whole-Genome Expression BeadChips (Illumina, San Diego, CA, USA), containing 47,231 probes per array. The beadchips were analyzed using Illumina iScan N240 microarray scanner (Illumina, San Diego, CA, USA).
Western blot
Western blot was carried out to analyze activation-states of the Hh- and PI3K/AKT/mTOR pathways. Tumor tissue samples from the same animals used in the gene expression analysis were homogenized in RIPA Lysis and Extraction Buffer (Thermo Scientific) using the TissueLyser LT (Qiagen) and Bioruptor® (Diagenode). Cell debris was removed by centrifugation and the protein extract was stored at −20 °C. Protein extracts (100 μg) were run on SDS-PAGE using Mini-PROTEAN® TGX™ Precast Gels (Bio-Rad) and transferred to nitrocellulose membranes using the Trans-Blot® Turbo™ Transfer System (Bio-Rad). Antibodies specific to GLI1 (ab151796, Abcam), GLI2 (LS-C313075, LifeSpan BioSciences), S6 (#2217, Cell Signaling Technology), AKT (#9272, Cell Signaling Technology), p-AKT (#9271, Cell Signaling Technology) and GAPDH (ab9485, Abcam, used as control) were detected using Amersham ECL Rabbit IgG (NA934VS, GE Healthcare Life Sciences). SuperSignal® West Femto Maximum Sensitivity Substrate (Thermo Scientific) was used for detection and digitalized images were acquired using Fujifilm Luminescent Image Analyzer LAS-1000 (Fujifilm, Tokyo, Japan).
Data processing and statistical analysis
All tumor volume measurements for each group were expressed as mean value and standard deviation (SD). Student’s t-test was used to compare data between groups using a two-tailed unpaired t-test, and p < 0.05 was considered statistically significant.
In the transcriptional analysis, data pre-processing and quantile normalization were performed on the raw signal intensities using the web-based BioArray Software Environment (BASE) system. Differentially expressed transcripts (between experimental groups) were identified using Nexus Expression 3.0 (BioDiscovery, El Segundo, CA, USA) as previously described [34, 35]. Transcripts with altered expression ≥1.5 fold (|log2-ratio| ≥ 0.58) and Benjamini-Hochberg-adjusted p-value < 0.01 were considered significantly regulated compared with untreated controls (hereafter referred to as regulated).
Analysis of affected canonical pathways related to human cancer [36] and upstream regulators was conducted using the Ingenuity Pathway Analysis (IPA) software (Ingenuity Systems, Redwood City, USA). The p-value of overlap between the experimental data and the Ingenuity knowledge base was calculated with Fisher’s exact test (significance threshold at p < 0.05). The z-score was used to determine the activation state of the upstream regulators; z > 2 indicates activation, while z < −2 indicates inhibition. The Gene Ontology database was used for analysis of regulated transcripts associated with cell death and cell cycle regulation (significance threshold at p < 0.05 using a modified Fisher’s exact test) [34].
Results
Anti-tumor effect of sonidegib or 177Lu-octreotate monotherapy on GOT1 tumors
Sonidegib monotherapy resulted in significant inhibition of tumor growth (Fig. 1a-b; minimum and maximum mean relative volumes 0.80 (SD = 0.25) and 1.2 (SD = 0.8), found at 7 and 41 d after treatment start, respectively). Statistically significant differences in mean relative volume between sonidegib-treated animals and controls were found at 7, 21, 28 and 35 d after treatment start.
Fig. 1.
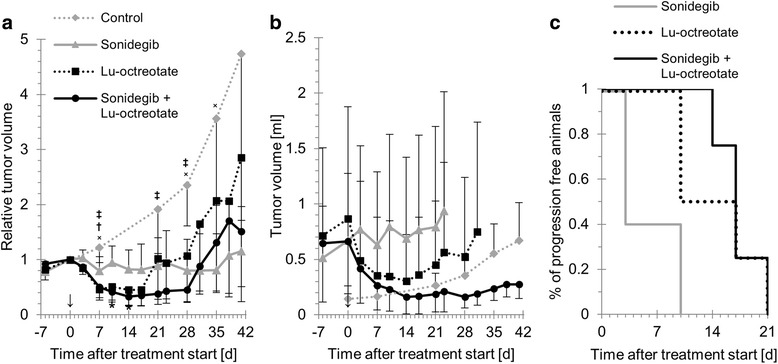
Anti-tumor effects of sonidegib, 177Lu-octreotate and combination treatment on GOT1 tumors in nude mice. a mean relative tumor volume versus time for controls and animals treated with sonidegib, 177Lu-octreotate, or a combination of both. b mean tumor volume versus time, until the first animal had to be killed on account of too high tumor burden, in each treatment group and controls. c percentage of animals in each treatment group without tumor progression. Error bars indicate SD. x, †, ‡ and * indicate time points with statistically significant differences between sonidegib and control, 177Lu-octreotate and control, combination and control, and combination and sonidegib treatment groups, respectively (Student’s t-test, p < 0.05). ↓ indicates the time for treatment start
The mean absorbed dose to the tumors receiving 177Lu-octreotate was 8 Gy at infinity time. Mean tumor volume relative to the day of injection was reduced in the group treated with 177Lu-octreotate monotherapy (Fig. 1a-b). The minimum relative tumor volume (mean = 0.45, SD = 0.29) was reached 14 d after injection. The mean relative tumor volume was below 1 from 3 d to 17 d after injection, after which the tumors began to regrow, resulting in a relative tumor volume of 2.9 at the end of the study. There was a statistically significant difference in relative tumor volumes between animals treated with 177Lu-octreotate and non-treated controls at 7 d after injection.
Sonidegib enhances the anti-tumor effects of 177Lu-octreotate on GOT1 tumors
Combination treatment with sonidegib and 177Lu-octreotate caused a reduction in mean relative tumor volume (Fig. 1a). The minimum relative tumor volume for tumors receiving the combination therapy was lower than in either monotherapy group (mean = 0.33, SD = 0.16 at 14 d after injection). The mean tumor volume in the group treated with a combination of sonidegib and 177Lu-octreotate was also reduced after treatment, and showed the lowest values at all measurement time points after treatment start (Fig. 1b). Furthermore, the combination therapy group had a prolonged time to progression, i.e. time from treatment start to progression of first tumor in the treatment group (Fig. 1c), and the mean tumor volume at study end was smaller compared with the other groups (Fig. 1b). No symptoms of toxic effects were observed in the animals of either group. There was a statistically significant difference between relative tumor volume in animals treated with a combination of sonidegib and 177Lu-octreotate, and non-treated controls at 7, 21 and 28 d after treatment start. In addition, a statistically significant difference between combination treatment and sonidegib monotherapy was found at 10 and 14 d after treatment start.
Different transcriptional responses in GOT1 tumors after monotherapies and combined treatment with sonidegib and 177Lu-octreotate
Genome-wide transcriptional microarray analysis of total RNA revealed diversity in gene regulation between treatment groups, compared with non-treated controls; the distribution and total number of regulated genes differed widely between treatment groups (Fig. 2). Seven, 106 and 496 transcripts were significantly regulated in the sonidegib, 177Lu-octreotate, and combination treatment groups, respectively. Four, seven and 397 transcripts were uniquely regulated in each group, while two genes (corresponding to three transcripts), BCL11A (involved in negative p53-regulation) and CXCR7 (encoding a chemokine receptor), were regulated in all treatment groups (Fig. 2b-d). The EVC2 and PDGFRA genes involved in the Hh pathway were among the four uniquely regulated transcripts in the sonidegib treatment group.
Fig. 2.
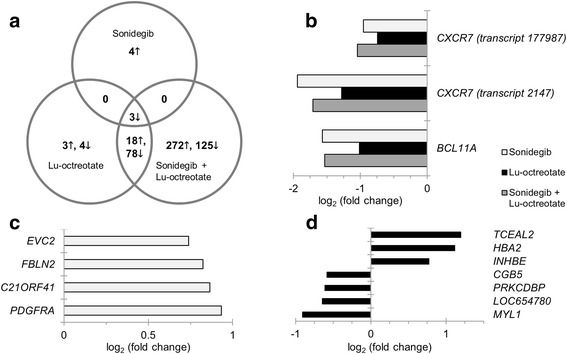
Distribution of differentially expressed genes in GOT1 tumors after sonidegib, 177Lu-octreotate and combination treatment. Differentially expressed transcripts in tumors treated with sonidegib, 177Lu-octreotate, or a combination of both pharmaceutical agents (treated vs. control). a: Venn diagram showing the distribution of up- (↓) and downregulated (↑) transcripts. Four, seven and 397 transcripts were uniquely regulated after sonidegib, 177Lu-octreotate and combination treatment, respectively; 96 transcripts were regulated after both 177Lu-octreotate and combination treatment; three transcripts were regulated in all treatment groups. b, c and d: Regulation patterns for the three commonly regulated transcripts and the four and seven transcripts uniquely regulated after sonidegib and 177Lu-octreotate treatment, respectively, with corresponding Illumina probe IDs. Transcripts with adjusted p-value < 0.01 and |log2(fold change)| ≥ 0.58 were considered significantly regulated. Up- and downregulation is indicated by positive and negative values, respectively
Differential effects on cell signaling in GOT1 tumors after sonidegib and 177Lu-octreotate
IPA analysis based on differentially expressed genes predicted that four upstream regulators (CTBP1, PRKCA, HIC1 and SUZ12) were significantly affected in all treatment groups; five affected upstream regulators were commonly detected in the sonidegib and 177Lu-octreotate treatment groups and three were shared between the sonidegib and combination treatment groups (Table 2A). An analysis of the activation state of the upstream regulators resulted in prediction of two significantly activated (FSH and Lh) and one significantly inhibited (INS) upstream regulators (Table 2B). Gene Ontology analysis revealed regulation of several genes related to apoptotic cell death and cell cycle regulation (including several related to cell cycle arrest or DNA replication) in the 177Lu-octreotate and combination therapy groups (Fig. 3), but none in the sonidegib group.
Table 2.
Predicted upstream regulators in GOT1 tumors after sonidegib, 177Lu-octreotate, or a combination of both treatments
| Upstream Regulator | Molecule Type | p-value: | Targets from data | ||
|---|---|---|---|---|---|
| Sonidegib | 177Lu-octreotate | Combination | |||
| A | |||||
| SUZ12 | Enzyme | 2.0·10−2 | 4.6·10−3 | 1.3·10−4 | UP: CDKN2A,
HSD17B11, LITAF
DOWN: CXCR7, KCNA1, RBMS1 |
| CTBP1 | Enzyme | 2.4·10−3 | 3.9·10−2 | 1.3·10−2 | UP: CDKN1A
DOWN: CXCR7 |
| PRKCA | Kinase | 1.2·10−2 | 1.9·10−2 | 2.5·10−3 | DOWN: BCL11A, EGR1, ETS1, HSPA1B, PVR |
| HIC1 | Transcription regulator | 1.6·10−2 | 2.9·10−2 | 3.3·10−2 | UP: CDO1
DOWN: CXCR7, ID4, KCNJ6 |
| miR-292b-5p | Mature microRNA | 1.1·10−3 | 1.8·10−2 | - | DOWN: BCL11A |
| LIN28B | Other | 1.3·10−3 | 2.2·10−2 | - | DOWN: BCL11A |
| CHD4 | Enzyme | 2.7·10−3 | 4.4·10−2 | - | DOWN: BCL11A |
| EZH2 | Transcription regulator | 2.4·10−2 | 8.1·10−3 | - | DOWN: CXCR7, KCNA1, NFKBIA |
| MAPK1 | Kinase | 4.2·10−2 | 3.5·10−2 | - | UP: HBA2
DOWN: BCL11A, CTNNA2 |
| Histone h3 | Group | 3.9·10−2 | - | 2.1·10−4 | UP: BTG3, CDKN1A, CDKN2A, ENO3, PCNA, RGS10, STOM
DOWN: CXCR7, NFKBIA, SOD2 |
| Ctbp | Group | 3.2·10−3 | - | 2.3·10−2 | UP: CDKN2A
DOWN: CXCR7 |
| ETS1 | Transcription regulator | 1.7·10−2 | - | 4.2·10−2 | DOWN: BCL11A, CTGF, HSPA1B, PVR |
| B | |||||
| Upstream Regulator | Molecule Type | z | p-value | Targets from data | |
| INS | Other | −2.1 | 2.6·10−4 | UP: CBS, LPL
DOWN: EGR1, OGT, SREBF1 |
|
| FSH | Complex | 2.2 | 7.5·10−3 | UP: DLX5, GNAS, PMAIP1, RGS16, SC4MOL
DOWN: BCL11A, DUSP3, FILIP1L, FLNC, ILK, ITGA3, TPM2 |
|
| Lh | Complex | 2.2 | 2.4·10−2 | UP: GNAS, PMAIP1, RGS16, SC4MOL
DOWN: DUSP3, FLNC, ILK, ITGA3, TPM2 |
|
A) Upstream regulators with statistically significant p-values (p < 0.05) in at least two treatment groups. B) Upstream regulators with activation |z-score| > 2, identified in tumor samples treated with a combination of sonidegib and 177Lu-octreotate. No upstream regulators with |z-score| > 2 were found in either monotherapy group. z > 2 indicates activation, z < −2 indicates inhibition. UP and DOWN indicate up- and downregulation, respectively
Fig. 3.
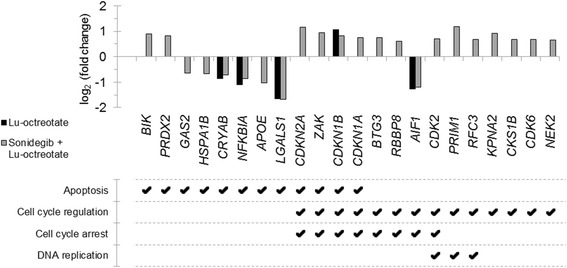
Expression of genes associated to cell death and/or cell cycle regulation in GOT1 tumors after 177Lu-octreotate and combination treatment. Differential expression (treated vs. control) of transcripts associated with cell death- and/or cell cycle regulation-processes in tumor samples from animals treated with 177Lu-octreotate (black), or a combination of sonidegib and 177Lu-octreotate (gray). No significant association with cell death or cell cycle regulation was found in tumor samples from animals treated with sonidegib monotherapy. Transcripts with |log2(fold change)| ≥ 0.58 and adjusted p < 0.01 were considered significantly regulated, association to biological processes was performed using the Gene Ontology database (threshold p < 0.05). Up- and downregulation is indicated by positive and negative values, respectively
Impact on cancer-related signaling pathways in GOT1 tumors after sonidegib and 177Lu-octreotate
The gene expression data was further analyzed by IPA. The affected signaling pathways in GOT1 tumors after treatment were identified by IPA analysis using differentially regulated genes. The signaling pathways in human cancer generated by IPA are presented in Table 3. The NF-ΚB signaling pathway was significantly affected in the group treated with sonidegib monotherapy – owing to the unique upregulation of the PDGFRA gene (encoding the platelet-derived growth factor receptor, alpha polypeptide). Interestingly, the gene encoding the ligand for the PDGFRA receptor (PDGFA) was downregulated in both the 177Lu-octreotate and combination therapy groups (log2-ratios of −1.5 and −1.7, respectively). The Wnt/β-catenin signaling pathway was significantly affected in both the 177Lu-octreotate and combination therapy groups, where the SOX2, TLE4, and WNT11 genes were downregulated in both groups. However, a larger number of genes in the Wnt/β-catenin pathway were affected in the combination therapy group. The PI3K/AKT/mTOR-, G-protein coupled receptor-, and Notch-signaling pathways were also affected in the combination therapy group.
Table 3.
Affected cancer-related canonical pathways in GOT1 tumors after sonidegib, 177Lu-octreotate, or combination of both treatments
| Canonical pathway | Treatment | p-value | Targets from data |
|---|---|---|---|
| Wnt/β-catenin | Sonidegib | - | - |
| signaling | 177Lu-octreotate | 0.045 | DOWN: SOX2, TLE4, WNT11 |
| Combination | 0.003 | UP: CDKN2A, PPP2R2C
DOWN: FZD9, GNAQ, ILK, SOX2, TCF4, TGFB3, TLE4, WNT11 |
|
| PI3K/AKT/mTOR signaling | Sonidegib | - | - |
| 177Lu-octreotate | 0.110 | UP: CDKN1B
DOWN: NFKBIA |
|
| Combination | 0.013 | UP: CDKN1A, CDKN1B, PPP2R2C, PTEN
DOWN: ILK, ITGA3, NFKBIA |
|
| G-protein coupled | Sonidegib | - | - |
| receptor signaling | 177Lu-octreotate | - | - |
| Combination | 0.043 | UP: GNAQ, GNAS, GRM8, PDE5A, RGS10, RGS16
DOWN: ADCY3, CAMK2B, GNAI2, NFKBIA |
|
| Notch signaling | Sonidegib | - | - |
| 177Lu-octreotate | - | - | |
| Combination | 0.043 | DOWN: DLL1, LFNG, PSENEN | |
| NF-ΚB signaling | Sonidegib | 0.048 | UP: PDGFRA |
| 177Lu-octreotate | 0.181 | DOWN: GHR, NFKBIA | |
| Combination | - | - |
Selection of Ingenuity canonical pathways (with statistical significance (p < 0.05) for at least one treatment group) related to human cancer [36], enriched by differentially expressed genes in each treatment group. UP and DOWN indicate up- and downregulation, respectively
Protein expression analysis reveals Hh-activation downstream of SMO, and activation of PI3K/AKT/mTOR
Western blotting showed increased amounts of GLI1 in tumors from animals treated with 177Lu-octreotate monotherapy and combination treatment, and increased amounts of GLI2 in tumors from the combination therapy group, compared with controls (Fig. 4). This indicates an activation of the Hh pathway in these tumors. Protein levels of AKT and p-AKT were elevated in all three treatment groups, while S6 was elevated in tumors from the 177Lu-octreotate monotherapy and combination treatment groups (Fig. 4).
Fig. 4.
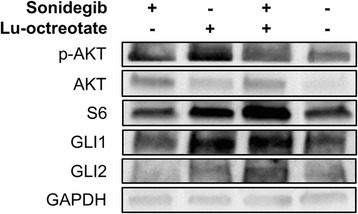
Expression of proteins in the Hh- and PI3K/AKT/mTOR pathways in GOT1 tumors after sonidegib, 177Lu-octreotate and combination treatment. Expression of Hh-related (GLI1, GLI2) and PI3K/AKT/mTOR-related (AKT, p-AKT, S6) proteins in tumors treated with sonidegib, 177Lu-octreotate, or a combination of both pharmaceutical agents, and untreated controls, measured using western blot analysis. Representative immunoblots from each group are shown. + and - indicates treated and untreated, respectively
Discussion
PRRT using 177Lu-octreotate is a promising treatment option for patients with NETs, with longer progression-free survival and higher response rates than alternative treatments [7]. However, due to dose limiting risk organs, curative treatment is still rare. This study describes the first combination treatment for neuroendocrine tumors with Hh pathway inhibition and PRRT. We show that, given as monotherapy, both sonidegib and 177Lu-octreotate have anti-tumor effects on GOT1 tumors in nude mice, with sonidegib resulting in inhibition of tumor growth over time and 177Lu-octreotate resulting in initial tumor volume regression followed by regrowth, in agreement with previous studies [16, 22, 27, 28]. The initial tumor volume response in the animals treated with a combination of sonidegib and 177Lu-octreotate mimicked that of the 177Lu-octreotate monotherapy. However, the time to progression was longer in the combination therapy group, resulting in the lowest mean tumor volume at the time of study end. This indicates a potential benefit when using Hh inhibitors in combination with 177Lu-octreotate for treatment of small intestine neuroendocrine tumors. However, further studies on the difference in adverse effects between different treatment schedules are needed, especially concerning adverse effects on risk organs (e.g. kidneys and bone marrow).
The tumor absorbed dose in animals receiving 30 MBq 177Lu-octreotate was estimated to 8 Gy at infinity time, assuming homogeneous activity distribution and based on the biokinetics of 15 MBq 177Lu-octreotate [29]. However, saturation of the SSTR is an issue that must be considered when using radiolabeled somatostatin analogs and it is possible that the 30 MBq used in this study may have resulted in a lower mean absorbed dose to the tumor [37]. In the present study, it was not possible to define the level of potential saturation.
A substantially higher number of genes were regulated in the combination therapy group compared with the two monotherapy groups. CXCR7 and BCL11A were regulated in all groups. The CXCR7 gene has been studied in human breast cancer models, where treatment with a CXCR7 antagonist has been shown to delay tumor growth and increase survival rates [38]. CXCR7 has also been identified as a possible downstream target of Hh pathway members GLI1 and GLI2 [14]. The BCL11A gene was downregulated in all three treatment groups. It negatively regulates p53 by directly regulating the BCL2, BCL- x L, MDM2 and MDM4 genes. Consequently, downregulation of the BCL11A gene might result in apoptotic and proliferative defects [39]. Ninety-six transcripts were regulated in both the 177Lu-octreotate and combination therapy groups. In similarity with the two commonly regulated genes, these were all regulated in the same direction (i.e either downregulated in both groups, or upregulated in both groups), and the expression levels were roughly similar (see Additional file 1). These results indicate that the combination therapy increased the diversity of transcriptional regulation, while having minor effects on the extent of regulation.
Several of the uniquely regulated genes have been associated with Hh signaling, namely the EVC2 and PDGFRA genes in the sonidegib group and the GNAS gene in the combination treatment group. The EVC2 gene has been identified as a tissue-specific regulator of Hh signaling: The EVC2 protein binds to SMO after it accumulates in cilia in response to Hh ligands, and upregulation of the EVC2 gene can activate the Hh pathway downstream of SMO, but upstream of GLI transcription factors [12]. The PDGFRA gene is a transcriptional target of GLI1, and downregulation of the PDGFRA gene has previously been associated with decreased GLI1 levels despite Hh pathway activation [40]. In the present study, upregulation of the EVC2 and PDGFRA genes in the sonidegib group may therefore correspond to activation of the Hh pathway downstream of SMO, countering the effect of the SMO antagonist.
Several of the regulated genes were associated with apoptotic cell death and cell cycle regulation. Among these, the CDK2 and CDK6 genes involved in cell cycle checkpoint activity have been found to be activated by GLI1, independent of SMO activation status [41, 42]. In addition, several genes involved in the TP53-signaling pathway were regulated in the present study, corresponding to both growth arrest (e.g. BTG3, CDK6, CDKN1A (p21), CDKN1B (p27), and CDKN2A (p16)) and apoptosis (e.g. APOE and BIK) [43–47].
The IPA pathway analysis resulted in the prediction of several cancer-related signaling pathways. The Wnt/β-catenin signaling is important in regulating cancer cell invasiveness, and has been found to be implicated in the acquisition of radioresistance and radiation-induced cell invasion in glioblastomas [48]. Downregulation of several key components of the Wnt/β-catenin pathway (e.g. FZD9 and WNT11) in the combination therapy group suggests that evasive radioresistance may be reduced following this treatment regimen. G-protein coupled receptor signaling was also found to be affected by the combination treatment. Out of the molecular targets for the treatments used in the present study, SSTR are G-protein coupled receptors, and SMO has been classified as a G-protein coupled receptor or a G-protein coupled receptor-like receptor. G-protein coupled receptor signaling is a major factor in many cellular functions in cancers [49]. These diverse biological functions complicate an interpretation of the predicted effect on G-protein coupled receptor signaling. However, the unique upregulation of the GNAS gene in the combination therapy group indicates a possible inhibition of the Hh pathway. The GNAS gene encodes the heterotrimeric Gs-protein α subunit (Gαs), which transmits various G-protein coupled receptor signals regulating, e.g. cell growth and survival. Previous in vivo studies have shown that the GNAS gene can act as a tumor suppressor in Hh-driven medulloblastomas [50]. The Notch signaling pathway was also predicted to be affected in the group receiving a combination of sonidegib and 177Lu-octreotate. Notch has a direct role in DNA damage response and Notch inhibitors have been considered for treatment of various cancers in combination with radiotherapy [51]. Inhibition of Notch has been shown to prevent upregulation of Notch ligands, e.g. DLL1, after radiotherapy in breast cancer cells, and the downregulation of DLL1 in the combination therapy group in the present study may indicate a possible explanation of the mechanism involved in the enhanced anti-tumor effects in this treatment group [52].
The PI3K/AKT/mTOR signaling pathway was predicted to be activated in the combination therapy group. This pathway has previously been recognized as a possible candidate for combination therapy with PRRT, since the mTOR signaling pathway is often upregulated in NETs and the mTOR inhibitor everolimus has shown promising anti-NET results [9]. However, a previous study found that a combination treatment with everolimus and 177Lu-octreotate promotes metastasis in a pancreatic NET model in rats [9]. The mTOR target S6 (a serine/threonine kinase) has previously been shown to activate GLI1 in multiple cancer types, independent of SMO, indicating a crosstalk between the PI3K/AKT/mTOR- and Hh pathways [16, 53]. Furthermore, a combination of PI3K/AKT/mTOR- and Hh inhibitors have been shown to have more potent anti-tumor effects than either monotherapy [53, 54]. Our western blot data showed elevated levels of GLI1, GLI2 and S6 in both the 177Lu-octerotate monotherapy and combination therapy groups. This suggests that 177Lu-octerotate may lead to SMO-independent Hh-activation via the PI3K/AKT/mTOR pathway, indicating a possibility for further increased therapeutic results from a triple-combination of 177Lu-octerotate, sonidegib and a PI3K/AKT/mTOR inhibitor.
Conclusions
In summary, combination therapy of GOT1 tumors in nude mice using sonidegib and 177Lu-octreotate resulted in a profound reduction in tumor volume shortly after treatment start, similar to the effect of 177Lu-octreotate monotherapy. In contrast to the 177Lu-octreotate monotherapy, a prolonged time to progression (tumor regrowth) was observed in the combination therapy group. These results show that combination therapy using sonidegib and 177Lu-octreotate could be beneficial to patients with NE-tumors, but further studies are needed to determine the optimal dose of sonidegib and 177Lu-octreotate, regarding anti-tumor and toxic effects.
Gene expression analysis revealed an interaction between sonidegib and 177Lu-octreotate, affecting several cancer-related signaling pathways (i.e. Wnt/β-catenin, PI3K/AKT/mTOR, G-protein coupled receptor, and Notch) not affected by either monotherapy. This may explain the underlying mechanisms of the enhanced anti-tumor effects from combination treatment with sonidegib and 177Lu-octreotate. Protein expression analysis indicated a possible PI3K/AKT/mTOR-dependent activation of GLI1 and GLI2, independent of SMO. This indicates that future studies of combination therapy using 177Lu-octerotate, sonidegib and a PI3K/AKT/mTOR inhibitor are warranted.
Acknowledgements
The authors thank Ann Wikström and Lilian Karlsson for their skilled technical work with the animals and the sample collection. Thanks also to the staff at Central Radiopharmacy at Sahlgrenska University Hospital, especially to Petra Bergström, PhLic, and Barbro Fredriksson who performed the labeling of 177Lu-octreotate.
Funding
This study was supported by grants from the Swedish Research Council, the Swedish Cancer Society, BioCARE – a National Strategic Research Program at University of Gothenburg, the King Gustav V Jubilee Clinic Cancer Research Foundation, the Wilhelm and Martina Lundgren Research Foundation, the Assar Gabrielsson Cancer Research Foundation, the Adlerbertska Research Fund, and Sahlgrenska University Hospital Research Funds. The funders had no role in the study design, or collection, analysis or interpretation of data, or writing the manuscript.
Availability of data and materials
The gene expression datasets generated during and/or analysed during the current study are available in the NCBI’s Gene Expression Omnibus repository [55].
Abbreviations
- 177Lu-octreotate
177Lu-[DOTA0, Tyr3]-octreotate
- BASE
BioArray Software Environment
- Hh
Hedgehog
- IPA
Ingenuity Pathway Analysis
- MIRD
Medical Internal Radiation Dose Committee
- NET
Neuroendocrine tumor
- PRRT
Peptide receptor radionuclide therapy
- RIN
RNA Integrity Number
- SCIBLU
Swegene Centre for Integrative Biology at Lund University
- SD
Standard deviation
- SMO
Smoothened
- SSTR
Somatostatin receptor
Additional file
Regulation patterns for the 96 transcripts regulated both after 177Lu-octreotate monotherapy and after combination therapy with sonidegib and 177Lu-octreotate. (XLS 40 kb)
Authors’ contributions
JS, BL, KH, EFA conceived and designed the experiment. JS, BL, NR performed the experiments. JS, BL, NR, TZP, KH, ON, EFA analyzed the data. KH, ON, EFA contributed reagents/materials/analysis tools. JS, KH, ON and EFA drafted the manuscript. All authors contributed to writing the manuscript, commented on this manuscript, and approved the final manuscript.
Ethics approval and consent to participate
All animal procedures were approved by the Ethical Committee on Animal Experiments in Gothenburg (reference number 107-2015).
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Footnotes
Electronic supplementary material
The online version of this article (doi:10.1186/s12885-017-3524-x) contains supplementary material, which is available to authorized users.
Contributor Information
Johan Spetz, Phone: +46-31-342 96 95, Email: spetz@hsph.harvard.edu.
Britta Langen, Email: britta.langen@gu.se.
Nils Rudqvist, Email: nils.rudqvist@gmail.com.
Toshima Z. Parris, Email: toshima.parris@oncology.gu.se
Khalil Helou, Email: khalil.helou@oncology.gu.se.
Ola Nilsson, Email: ola.nilsson@llcr.med.gu.se.
Eva Forssell-Aronsson, Email: eva.forssell_aronsson@radfys.gu.se.
References
- 1.Yao JC, Hassan M, Phan A, Dagohoy C, Leary C, Mares JE, Abdalla EK, Fleming JB, Vauthey JN, Rashid A, Evans DB. One hundred years after “carcinoid”: epidemiology of and prognostic factors for neuroendocrine tumors in 35,825 cases in the United States. J Clin Oncol. 2008;26:3063–3072. doi: 10.1200/JCO.2007.15.4377. [DOI] [PubMed] [Google Scholar]
- 2.Gu D, Liu H, Su GH, Zhang X, Chin-Sinex H, Hanenberg H, Mendonca MS, Shannon HE, Chiorean EG, Xie J. Combining hedgehog signaling inhibition with focal irradiation on reduction of pancreatic cancer metastasis. Mol Cancer Ther. 2013;12:1038–1048. doi: 10.1158/1535-7163.MCT-12-1030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Modlin IM, Öberg K, Chung DC, Jensen RT, de Herder WW, Thakker RV, Caplin M, Delle Fave G, Kaltsas GA, Krenning EP, et al. Gastroenteropancreatic neuroendocrine tumours. Lancet Oncol. 2008;9:61–72. doi: 10.1016/S1470-2045(07)70410-2. [DOI] [PubMed] [Google Scholar]
- 4.Delpassand ES, Samarghandi A, Zamanian S, Wolin EM, Hamiditabar M, Espenan GD, Erion JL, O'Dorisio TM, Kvols LK, Simon J, et al. Peptide receptor radionuclide therapy with 177Lu-DOTATATE for patients with somatostatin receptor-expressing neuroendocrine tumors: the first US phase 2 experience. Pancreas. 2014;43:518–525. doi: 10.1097/MPA.0000000000000113. [DOI] [PubMed] [Google Scholar]
- 5.Sward C, Bernhardt P, Ahlman H, Wangberg B, Forssell-Aronsson E, Larsson M, Svensson J, Rossi-Norrlund R, Kolby L. [177Lu-DOTA 0-Tyr 3]-octreotate treatment in patients with disseminated gastroenteropancreatic neuroendocrine tumors: the value of measuring absorbed dose to the kidney. World J Surg. 2010;34:1368–1372. doi: 10.1007/s00268-009-0387-6. [DOI] [PubMed] [Google Scholar]
- 6.Bodei L, Cremonesi M, Kidd M, Grana CM, Severi S, Modlin IM, Paganelli G. Peptide receptor radionuclide therapy for advanced neuroendocrine tumors. Thorac Surg Clin. 2014;24:333–349. doi: 10.1016/j.thorsurg.2014.04.005. [DOI] [PubMed] [Google Scholar]
- 7.Strosberg J, El-Haddad G, Wolin E, Hendifar A, Yao J, Chasen B, Mittra E, Kunz PL, Kulke MH, Jacene H, et al. Phase 3 trial of 177Lu-Dotatate for Midgut Neuroendocrine tumors. N Engl J Med. 2017;376:125–135. doi: 10.1056/NEJMoa1607427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 8.Forssell-Aronsson E, Spetz J, Ahlman H. Radionuclide therapy via SSTR: future aspects from experimental animal studies. Neuroendocrinology. 2013;97:86–98. doi: 10.1159/000336086. [DOI] [PubMed] [Google Scholar]
- 9.Pool SE, Bison S, Koelewijn SJ, van der Graaf LM, Melis M, Krenning EP, de Jong M. mTOR inhibitor RAD001 promotes metastasis in a rat model of pancreatic neuroendocrine cancer. Cancer Res. 2013;73:12–18. doi: 10.1158/0008-5472.CAN-11-2089. [DOI] [PubMed] [Google Scholar]
- 10.Claringbold PG, Price RA, Turner JH. Phase I-II study of radiopeptide 177Lu-octreotate in combination with capecitabine and temozolomide in advanced low-grade neuroendocrine tumors. Cancer Biother Radiopharm. 2012;27:561–569. doi: 10.1089/cbr.2012.1276. [DOI] [PubMed] [Google Scholar]
- 11.Villavicencio EH, Walterhouse DO, Iannaccone PM. The sonic hedgehog-patched-gli pathway in human development and disease. Am J Hum Genet. 2000;67:1047–1054. doi: 10.1016/S0002-9297(07)62934-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Yang C, Chen W, Chen Y, Jiang J. Smoothened transduces hedgehog signal by forming a complex with Evc/Evc2. Cell Res. 2012;22:1593–1604. doi: 10.1038/cr.2012.134. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 13.Chen YJ, Lin CP, Hsu ML, Shieh HR, Chao NK, Chao KS. Sonic hedgehog signaling protects human hepatocellular carcinoma cells against ionizing radiation in an autocrine manner. Int J Radiat Oncol Biol Phys. 2011;80:851–859. doi: 10.1016/j.ijrobp.2011.01.003. [DOI] [PubMed] [Google Scholar]
- 14.Inaguma S, Riku M, Ito H, Tsunoda T, Ikeda H, Kasai K. GLI1 orchestrates CXCR4/CXCR7 signaling to enhance migration and metastasis of breast cancer cells. Oncotarget. 2015;6:33648–33657. doi: 10.18632/oncotarget.5203. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Jiang J, Hui CC. Hedgehog signaling in development and cancer. Dev Cell. 2008;15:801–812. doi: 10.1016/j.devcel.2008.11.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Amakye D, Jagani Z, Dorsch M. Unraveling the therapeutic potential of the hedgehog pathway in cancer. Nat Med. 2013;19:1410–1422. doi: 10.1038/nm.3389. [DOI] [PubMed] [Google Scholar]
- 17.Fendrich V, Waldmann J, Esni F, Ramaswamy A, Mullendore M, Buchholz M, Maitra A, Feldmann G. Snail and sonic hedgehog activation in neuroendocrine tumors of the ileum. Endocr Relat Cancer. 2007;14:865–874. doi: 10.1677/ERC-07-0108. [DOI] [PubMed] [Google Scholar]
- 18.Shida T, Furuya M, Nikaido T, Hasegawa M, Koda K, Oda K, Miyazaki M, Kishimoto T, Nakatani Y, Ishikura H. Sonic hedgehog-Gli1 signaling pathway might become an effective therapeutic target in gastrointestinal neuroendocrine carcinomas. Cancer Biol Ther. 2006;5:1530–1538. doi: 10.4161/cbt.5.11.3458. [DOI] [PubMed] [Google Scholar]
- 19.Bohinc B, Michelotti G, Diehl AM. Hedgehog signaling in human medullary thyroid carcinoma: a novel signaling pathway. Thyroid. 2013;23:1119–1126. doi: 10.1089/thy.2012.0474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Sims-Mourtada J, Izzo JG, Apisarnthanarax S, Wu TT, Malhotra U, Luthra R, Liao Z, Komaki R, van der Kogel A, Ajani J, Chao KS. Hedgehog: an attribute to tumor regrowth after chemoradiotherapy and a target to improve radiation response. Clin Cancer Res. 2006;12:6565–6572. doi: 10.1158/1078-0432.CCR-06-0176. [DOI] [PubMed] [Google Scholar]
- 21.Pan S, Wu X, Jiang J, Gao W, Wan Y, Cheng D, Han D, Liu J, Englund NP, Wang Y, et al. Discovery of NVP-LDE225, a potent and selective smoothened antagonist. ACS Med Chem Lett. 2010;1:130–134. doi: 10.1021/ml1000307. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Fendrich V, Wiese D, Waldmann J, Lauth M, Heverhagen AE, Rehm J, Bartsch DK. Hedgehog inhibition with the orally bioavailable Smo antagonist LDE225 represses tumor growth and prolongs survival in a transgenic mouse model of islet cell neoplasms. Ann Surg. 2011;254:818–823. doi: 10.1097/SLA.0b013e318236bc0f. [DOI] [PubMed] [Google Scholar]
- 23.Pietanza MC, Litvak AM, Varghese AM, Krug LM, Fleisher M, Teitcher JB, Holodny AI, Sima CS, Woo KM, Ng KK, et al. A phase I trial of the hedgehog inhibitor, sonidegib (LDE225), in combination with etoposide and cisplatin for the initial treatment of extensive stage small cell lung cancer. Lung Cancer. 2016;99:23–30. doi: 10.1016/j.lungcan.2016.04.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Kumari A, Ermilov AN, Allen BL, Bradley RM, Dlugosz AA, Mistretta CM. Hedgehog pathway blockade with the cancer drug LDE225 disrupts taste organs and taste sensation. J Neurophysiol. 2015;113:1034–1040. doi: 10.1152/jn.00822.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Kölby L, Bernhardt P, Ahlman H, Wängberg B, Johanson V, Wigander A, Forssell-Aronsson E, Karlsson S, Ahren B, Stenman G, Nilsson O. A transplantable human carcinoid as model for somatostatin receptor-mediated and amine transporter-mediated radionuclide uptake. Am J Pathol. 2001;158:745–755. doi: 10.1016/S0002-9440(10)64017-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Nilsson O, Kölby L, Bernhardt P, Forssell-Aronsson E, Johanson V, Ahlman H. GOT1 xenografted to nude mice: a unique model for in vivo studies on SSTR-mediated radiation therapy of carcinoid tumors. Ann N Y Acad Sci. 2004;1014:275–279. doi: 10.1196/annals.1294.031. [DOI] [PubMed] [Google Scholar]
- 27.Kölby L, Bernhardt P, Johanson V, Schmitt A, Ahlman H, Forssell-Aronsson E, Macke H, Nilsson O. Successful receptor-mediated radiation therapy of xenografted human midgut carcinoid tumour. Br J Cancer. 2005;93:1144–1151. doi: 10.1038/sj.bjc.6602845. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Swärd C, Bernhardt P, Johanson V, Schmitt A, Ahlman H, Stridsberg M, Forssell-Aronsson E, Nilsson O, Kölby L. Comparison of [177Lu-DOTA0,Tyr3]-octreotate and [177Lu-DOTA0,Tyr3]-octreotide for receptor-mediated radiation therapy of the xenografted human midgut carcinoid tumor GOT1. Cancer Biother Radiopharm. 2008;23:114–120. doi: 10.1089/cbr.2007.0421. [DOI] [PubMed] [Google Scholar]
- 29.Dalmo J, Spetz J, Montelius M, Langen B, Arvidsson Y, Johansson H, Parris TZ, Helou K, Wangberg B, Nilsson O, et al. Priming increases the anti-tumor effect and therapeutic window of 177Lu-octreotate in nude mice bearing human small intestine neuroendocrine tumor GOT1. EJNMMI Res. 2017;7:6. doi: 10.1186/s13550-016-0247-y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.de Jong M, Breeman WA, Bernard BF, Bakker WH, Schaar M, van Gameren A, Bugaj JE, Erion J, Schmidt M, Srinivasan A, Krenning EP. [177Lu-DOTA(0),Tyr3] octreotate for somatostatin receptor-targeted radionuclide therapy. Int J Cancer. 2001;92:628–633. doi: 10.1002/1097-0215(20010601)92:5<628::AID-IJC1244>3.0.CO;2-L. [DOI] [PubMed] [Google Scholar]
- 31.Kolby L, Bernhardt P, Ahlman H, Wangberg B, Johanson V, Wigander A, Forssell-Aronsson E, Karlsson S, Ahren B, Stenman G, Nilsson O. A transplantable human carcinoid as model for somatostatin receptor-mediated and amine transporter-mediated radionuclide uptake. Am J Pathol. 2001;158:745–755. doi: 10.1016/S0002-9440(10)64017-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Bolch WE, Eckerman KF, Sgouros G, Thomas SR. MIRD pamphlet no. 21: a generalized schema for radiopharmaceutical dosimetry--standardization of nomenclature. J Nucl Med. 2009;50:477–484. doi: 10.2967/jnumed.108.056036. [DOI] [PubMed] [Google Scholar]
- 33.Eckerman K, Endo A. ICRP publication 107. Nuclear decay data for dosimetric calculations. Ann ICRP. 2008;38:7–96. doi: 10.1016/j.icrp.2008.10.001. [DOI] [PubMed] [Google Scholar]
- 34.Ashburner M, Ball CA, Blake JA, Botstein D, Butler H, Cherry JM, Davis AP, Dolinski K, Dwight SS, Eppig JT, et al. Gene ontology: tool for the unification of biology. The gene ontology consortium. Nat Genet. 2000;25:25–29. doi: 10.1038/75556. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Parris TZ, Danielsson A, Nemes S, Kovacs A, Delle U, Fallenius G, Mollerstrom E, Karlsson P, Helou K. Clinical implications of gene dosage and gene expression patterns in diploid breast carcinoma. Clin Cancer Res. 2010;16:3860–3874. doi: 10.1158/1078-0432.CCR-10-0889. [DOI] [PubMed] [Google Scholar]
- 36.Weinberg RA: The biology of cancer. Second edition. edn; 2013.
- 37.Bernhardt P, Kolby L, Johanson V, Nilsson O, Ahlman H, Forssell-Aronsson E. Biodistribution of 111in-DTPA-D-Phe1-octreotide in tumor-bearing nude mice: influence of amount injected and route of administration. Nucl Med Biol. 2003;30:253–260. doi: 10.1016/S0969-8051(02)00417-1. [DOI] [PubMed] [Google Scholar]
- 38.Burns JM, Summers BC, Wang Y, Melikian A, Berahovich R, Miao Z, Penfold ME, Sunshine MJ, Littman DR, Kuo CJ, et al. A novel chemokine receptor for SDF-1 and I-TAC involved in cell survival, cell adhesion, and tumor development. J Exp Med. 2006;203:2201–2213. doi: 10.1084/jem.20052144. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Yu Y, Wang J, Khaled W, Burke S, Li P, Chen X, Yang W, Jenkins NA, Copeland NG, Zhang S, Liu P. Bcl11a is essential for lymphoid development and negatively regulates p53. J Exp Med. 2012;209:2467–2483. doi: 10.1084/jem.20121846. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Lin C, Chen MH, Yao E, Song H, Gacayan R, Hui CC, Chuang PT. Differential regulation of Gli proteins by Sufu in the lung affects PDGF signaling and myofibroblast development. Dev Biol. 2014;392:324–333. doi: 10.1016/j.ydbio.2014.05.014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Rizvi S, Demars CJ, Comba A, Gainullin VG, Rizvi Z, Almada LL, Wang K, Lomberk G, Fernandez-Zapico ME, Buttar NS. Combinatorial chemoprevention reveals a novel smoothened-independent role of GLI1 in esophageal carcinogenesis. Cancer Res. 2010;70:6787–6796. doi: 10.1158/0008-5472.CAN-10-0197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Chung JH, Bunz F. Cdk2 is required for p53-independent G2/M checkpoint control. PLoS Genet. 2010;6:e1000863. doi: 10.1371/journal.pgen.1000863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Riley T, Sontag E, Chen P, Levine A. Transcriptional control of human p53-regulated genes. Nat Rev Mol Cell Biol. 2008;9:402–412. doi: 10.1038/nrm2395. [DOI] [PubMed] [Google Scholar]
- 44.Gillissen B, Essmann F, Graupner V, Starck L, Radetzki S, Dorken B, Schulze-Osthoff K, Daniel PT. Induction of cell death by the BH3-only Bcl-2 homolog Nbk/Bik is mediated by an entirely Bax-dependent mitochondrial pathway. EMBO J. 2003;22:3580–3590. doi: 10.1093/emboj/cdg343. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Ou YH, Chung PH, Hsu FF, Sun TP, Chang WY, Shieh SY. The candidate tumor suppressor BTG3 is a transcriptional target of p53 that inhibits E2F1. EMBO J. 2007;26:3968–3980. doi: 10.1038/sj.emboj.7601825. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Li J, Yang XK, Yu XX, Ge ML, Wang WL, Zhang J, Hou YD. Overexpression of p27(KIP1) induced cell cycle arrest in G(1) phase and subsequent apoptosis in HCC-9204 cell line. World J Gastroenterol. 2000;6:513–521. doi: 10.3748/wjg.v6.i4.513. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Shapiro GI, Edwards CD, Rollins BJ. The physiology of p16(INK4A)-mediated G1 proliferative arrest. Cell Biochem Biophys. 2000;33:189–197. doi: 10.1385/CBB:33:2:189. [DOI] [PubMed] [Google Scholar]
- 48.Dong Z, Zhou L, Han N, Zhang M, Lyu X. Wnt/beta-catenin pathway involvement in ionizing radiation-induced invasion of U87 glioblastoma cells. Strahlenther Onkol. 2015;191:672–680. doi: 10.1007/s00066-015-0858-7. [DOI] [PubMed] [Google Scholar]
- 49.O'Hayre M, Degese MS, Gutkind JS. Novel insights into G protein and G protein-coupled receptor signaling in cancer. Curr Opin Cell Biol. 2014;27:126–135. doi: 10.1016/j.ceb.2014.01.005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.He X, Zhang L, Chen Y, Remke M, Shih D, Lu F, Wang H, Deng Y, Yu Y, Xia Y, et al. The G protein alpha subunit Galphas is a tumor suppressor in sonic hedgehog-driven medulloblastoma. Nat Med. 2014;20:1035–1042. doi: 10.1038/nm.3666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Yahyanejad S, Theys J, Vooijs M. Targeting notch to overcome radiation resistance. Oncotarget. 2016;7:7610–7628. doi: 10.18632/oncotarget.6714. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Lagadec C, Vlashi E, Alhiyari Y, Phillips TM, Bochkur Dratver M, Pajonk F. Radiation-induced notch signaling in breast cancer stem cells. Int J Radiat Oncol Biol Phys. 2013;87:609–618. doi: 10.1016/j.ijrobp.2013.06.2064. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Wang Y, Ding Q, Yen CJ, Xia W, Izzo JG, Lang JY, Li CW, Hsu JL, Miller SA, Wang X, et al. The crosstalk of mTOR/S6K1 and hedgehog pathways. Cancer Cell. 2012;21:374–387. doi: 10.1016/j.ccr.2011.12.028. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Sharma N, Nanta R, Sharma J, Gunewardena S, Singh KP, Shankar S, Srivastava RK. PI3K/AKT/mTOR and sonic hedgehog pathways cooperate together to inhibit human pancreatic cancer stem cell characteristics and tumor growth. Oncotarget. 2015;6:32039–32060. doi: 10.18632/oncotarget.5055. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.GEO Accession viewer. https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE80022. Accessed 27 Sept 2016.
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Data Availability Statement
The gene expression datasets generated during and/or analysed during the current study are available in the NCBI’s Gene Expression Omnibus repository [55].