

Abstract
Globin-coupled diguanylate cyclases contain globin, middle, and diguanylate cyclase domains that sense O2 to synthesize c-di-GMP and regulate bacterial motility, biofilm formation, and virulence. However, relatively few studies have extensively examined the roles of individual residues and domains of globin-coupled diguanylate cyclases, which can shed light on their signaling mechanisms and provide drug targets. Here, we report the critical residues of two globin-coupled diguanylate cyclases, EcGReg from Escherichia coli and BpeGReg from Bordetella pertussis, and show that their diguanylate cyclase activity requires an intact globin domain. In the distal heme pocket of the globin domain, residues Phe42, Tyr43, Ala68 (EcGReg)/Ser68 (BpeGReg), and Met69 are required to maintain full diguanylate cyclase activity. The highly conserved amino acids His223/His225 and Lys224/Lys226 in the middle domain of EcGReg/BpeGReg are essential to diguanylate cyclase activity. We also identified sixteen important residues (Leu300, Arg306, Asp333, Phe337, Lys338, Asn341, Asp342, Asp350, Leu353, Asp368, Arg372, Gly374, Gly375, Asp376, Glu377, and Phe378) in the active site and inhibitory site of the diguanylate cyclase domain of EcGReg. Moreover, BpeGReg266 (residues 1–266) and BpeGReg296 (residues 1–296), which only contain the globin and middle domains, can inhibit bacterial motility. Our findings suggest that the distal residues of the globin domain affect diguanylate cyclase activity and that BpeGReg may interact with other c-di-GMP-metabolizing proteins to form mixed signaling teams.
Introduction
Globin-coupled diguanylate cyclases (GCDCs) form a subfamily of globin-coupled sensors (GCS) that are heme-binding sensors linked to variable signaling domains [1–5]. The N-terminal globin domains of GCDCs consist of eight alpha helices and display a myoglobin-like topology [6, 7]. The ferrous ion centered in the heme reversely binds O2, and distal residues facilitate O2 migration and stabilization in the hydrophobic heme pocket [6, 8–10]. In the C-terminal diguanylate cyclase (DGC) domains of GCDCs, the highly conserved GGD/EEF (Gly-Gly-Asp/Glu-Glu-Phe) motif serves as the active site to synthesize the second messenger bis-(3'-5')-cyclic diguanosine monophosphate (c-di-GMP) [6, 8].
C-di-GMP-dependent signaling pathways regulate diverse cellular functions including motility, biofilm formation, virulence, differentiation, and the cell cycle [11]. Various c-di-GMP receptors have been identified in bacteria and the mammalian innate immune system [12–20]. C-di-GMP is synthesized by DGCs (including GCDCs) containing a GGDEF domain and degraded by phosphodiesterases (PDEs) with either an EAL or HD-GYP domain [11]. Multiple DGCs and PDEs are found in most bacteria and are often associated with sensory or regulatory domains that allow them to modulate their activities in response to internal and environmental stimuli [11]. Studies on how DGCs and PDEs sense environmental signals to regulate c-di-GMP levels will shed light on the mechanisms of bacterial behavior and provide potential drug targets to attenuate the virulence of pathogens.
Various GCDCs have been characterized from Escherichia coli, Bordetella pertussis, Azotobacter vinelandii, Desulfotalea psychrophila, Shewanella putrefaciens, and Pectobacterium carotovorum [6, 8–10, 21–23]. Two GCDCs, EcGReg (also named DosC) from E. coli and BpeGReg from the whooping cough pathogen B. pertussis, can sense O2 to regulate c-di-GMP synthesis [6, 8]. They differ in respect of O2 affinity and cooperative PDEs [6, 8, 24]. EcGReg and Ec DosP (EAL type) couple to control c-di-GMP homeostasis, whereas BpeGReg may cooperate with Bpe RpfG (HD-GYP type) [8, 24]. To address related questions on how globins regulate DGC activity, here we report the critical residues identified in the three domains of GCDCs and that GCDCs require an intact globin domain for their enzyme activities. We examined Phe42, Tyr43, Ala68 (EcGReg)/Ser68 (BpeGReg), and Met69, which are in the distal heme pocket of the globin domain. We also tested the highly conserved amino acids His223/His225 and Lys224/Lys226 in the middle domain of EcGReg and BpeGReg. In addition, we used EcGReg as a model to examine sixteen conserved residues in the active site and inhibitory site of the DGC domains. We propose that distal globin residues facilitate O2 binding to regulate DGC activity and BpeGReg may interact with other DGCs to form mixed signaling teams.
Materials and methods
Sequence alignment and visualization of predicted critical residues
Protein sequences were aligned using EBI-MAFFT [25, 26]. The alignment was visualized by using EBI-MView [26, 27]. The homology model of BpeGReg was created previously [6]. Visualization of the locations of the critical residues was carried out using UCSF Chimera [28].
Plasmid construction
The ecGReg (GenBank accession no. NP_416007) and bpeGReg (GenBank accession no. NP_882025) genes were cloned into the pTrc99A vector (primers listed in Table A in S1 File). The truncated versions of these genes were also cloned into the pTrc99A vector (primers listed in Tables B and C in S1 File).
The plasmids containing the full-length genes were used as templates for the QuickChange site-directed mutagenesis protocol (Stratagene). Briefly, PfuTurbo DNA polymerase (Stratagene) and the primers listed in Tables B and C in S1 File were used for polymerase chain reactions (PCR). Thermal cycling was carried out with 12 cycles as follows: 94°C for 30 seconds, 55°C for 30 seconds, and 68°C for 12 minutes, with a final extension at 68°C for 7 minutes. One microliter of DpnI (10 U/μl, Promega) was then added to digest the methylated parental DNA template at 37°C for 1 hour. The mutated plasmids were transformed into E. coli TOP10 cells (Invitrogen). The coding regions of the isolated plasmids were verified by Sanger sequencing on ABI 3730xl DNA analyzer.
For protein expression and purification, full-length and truncated BpeGReg proteins were engineered with an N-terminal hexahistidine tag by PCR. Primers are listed in Table D in S1 File. The PCR products were cloned into the pCR4Blunt-TOPO vector (Invitrogen) and then subcloned into the pET-3a expression vector (Novagen).
The laboratory protocol of the molecular cloning has been deposited in protocols.io (DOI: http://dx.doi.org/10.17504/protocols.io.ijxccpn).
Phenotypic assays
Plasmids were transformed into Salmonella typhimurium ATCC 14028 by electroporation and maintained in LB broth or plates with ampicillin (100 μg/ml) at 37°C. For biofilm formation, cells were grown on LB without salt plates containing Congo red (40 μg/ml) for 40 hours at 37°C. Swimming motility was assayed on 0.3% agar plates (1% tryptone, 0.5% NaCl, 1 μM thiamine) at 28°C for 6 hours. Due to leaky expression of the pTrc99A vector, isopropyl β-D-thiogalactopyranoside (IPTG) was not used to induce protein expression during these assays.
Overexpression, purification and absorption spectra
His-tagged BpeGReg, BpeGReg155, BpeGReg266, and BpeGReg296 were overexpressed in E. coli Rosetta2(DE3)pLysS cells for 6–8 h at room temperature. Protein expression was induced with 0.05 mM isopropyl-β-D-thiogalactopyranoside. Proteins were purified by Co2+-affinity chromatography according to Piatibratov et al. [29]. The detailed protocol has been deposited in protocols.io (DOI: http://dx.doi.org/10.17504/protocols.io.ikhcct6). Spectra were measured using a Cary 1E UV-Visible spectrophotometer (Varian).
Results and discussion
Prediction of critical residues of globin-coupled diguanylate cyclases
The open reading frames of ecGReg and bpeGReg encode multidomain proteins containing 460 and 475 amino acids, respectively (Fig 1A). The region linking the N-terminal globin domain and C-terminal DGC domain could not be classified but is highly conserved in many GCDCs, including EcGReg and BpeGReg [3]. We refer to this region as the middle domain (Fig 1A). Fig 1B shows the protein sequence alignment of EcGReg, BpeGReg, and other GCDCs with the same domain architecture. We predicted a series of residues in the globin, middle, and DGC domains of EcGReg and BpeGReg that may affect their enzymatic activities, based on amino acid sequence alignment and the homology model of BpeGReg [6]. The crystal structure of EcGReg [7] was not available at the time this study was performed, so it was only used for subsequently visualizing the positions of the residues we tested rather than aiding our predictions (Fig 2).
Fig 1. Two globin-coupled diguanylate cyclases consist of three domains.

(A) Domain structures of EcGReg and BpeGReg. BLASTP searches against the non-redundant protein sequences database at the National Center for Biotechnology Information showed that EcGReg residues 5–153 represent a globin sensor domain and residues 268–458 represent a DGC domain, while BpeGReg residues 5–155 and residues 297–457 represent a globin sensor domain and a DGC domain, respectively. (B) Amino acid sequence alignment of five globin-coupled diguanylate cyclases: EcGReg, BpeGReg, AvGReg (Azotobacter vinelandii), SpGReg (Shewanella putrefaciens), and PccGReg (Pectobacterium carotovorum). The sequence order is based on the alignment. The residues highlighted in red are highly conserved and discussed in this work. The residues highlighted in blue are not highly conserved and discussed in this work. The line colors indicate the globin (orange), middle (green), and DGC (blue) domains.
Fig 2. Positions of predicted critical amino acids in the crystal structure of EcGReg/DosC.
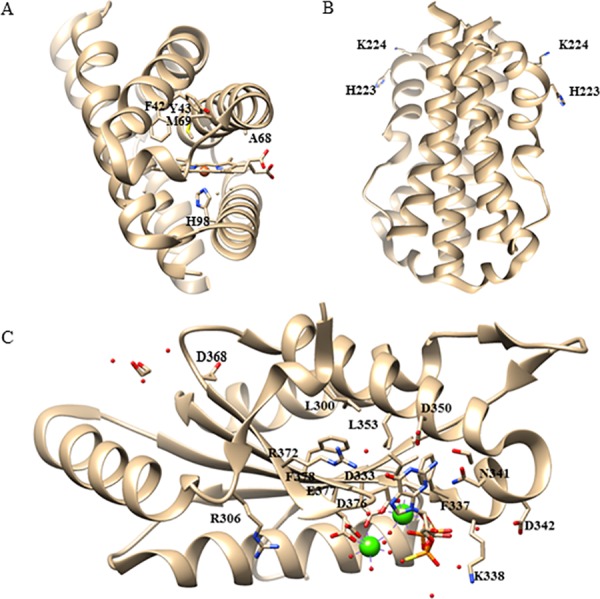
(A) Crystal structure of the globin domain of EcGReg/DosC (pdb: 4zvb, ferrous form) shows the locations of the critical residues in the heme pocket. H98 is the proximal histidine which binds the heme. The side chains of F42, Y43, and M69 surround the heme center. The side chain of A68 points away from the heme center. (B) Crystal structure of the middle domain of EcGReg/DosC (pdb: 4zvc, form I) shows the locations of critical residues H223 and K224, (C) Crystal structure of the DGC domain of EcGReg/DosC (pdb: 4zvf, GTPαS-bound) shows the locations of mutated residues in this work.
In the EcGReg globin domain, the distal residue Tyr43 stabilizes O2 to ferrous ion [10]. Our unpublished data suggests a similar role for Tyr43 in BpeGReg. We selected Tyr43 and three other residues in the distal heme pocket of both proteins for site-directed mutagenesis. In the middle domain, BpeGReg His225 was examined previously [6]. Here we selected His223 in EcGReg and nearby conserved residues Lys224 (EcGReg)/Lys226 (BpeGReg) for mutagenesis. To analyze the conserved residues in the DGC domain, we used EcGReg as a model and selected 16 residues in the active and inhibitory sites.
Mutated proteins were expressed in Salmonella typhimurium ATCC 14028 to examine their in vivo DGC activities by rdar (red, dry, and rough) formation and motility assays (Figs 3–6, Tables 1 and 2). High levels of intracellular c-di-GMP can inhibit motility, increase the production of exopolysaccharides (EPS) (e.g., cellulose) and adhesion factors (e.g., curli fimbriae), and enhance biofilm formation [30]. S. typhimurium develops an rdar morphotype on Congo red agar plates at 28°C but not at 37°C, indicating the expression of cellulose and curli fimbriae [31, 32]. At 37°C, the temperature regulation of rdar morphotype development can be overcome by overexpression of the DGC AdrA as well as heterologous expression of other DGCs such as EcGReg and BpeGReg [6, 30, 33].
Fig 3. Rdar morphotype development of EcGReg mutants on Congo red agar plates.
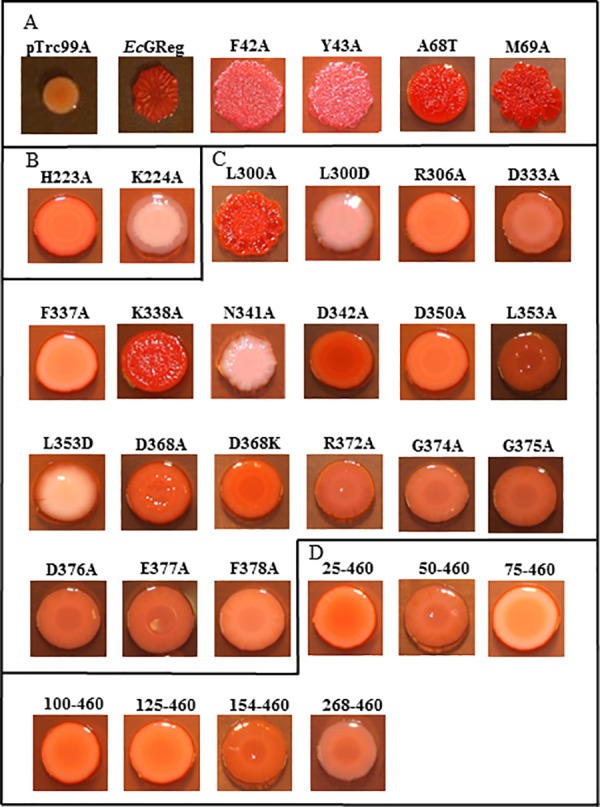
(A) Rdar morphotypes of EcGReg globin domain mutants. (B) Rdar morphotypes of EcGReg middle domain mutants. (C) Rdar morphotypes of EcGReg DGC domain mutants. (D) Rdar morphotypes of truncated EcGReg proteins.
Fig 6. Swimming motility of BpeGReg mutants on 0.3% tryptone agar plates.

(A) Swimming motility of pTrc99A vector control, BpeGReg, and BpeGReg globin domain mutants. (B) Swimming motility of BpeGReg middle domain mutant. (C) Swimming motility of truncated BpeGReg proteins.
Table 1. Summary of phenotypes of S. typhimurium expressing EcGReg mutants.
| Residue | Mutant | RDAR | Motility inhibition | Group |
|---|---|---|---|---|
| Globin domain | ||||
| F42 | A | + | - | 2 |
| Y43 | A | + | - | 2 |
| A68 | T | partial | - | 2 |
| M69 | A | + | - | 2 |
| Middle domain | ||||
| H223 | A | - | - | 1 |
| K224 | A | - | - | 1 |
| DGC domain | ||||
| L300 | A | partial | - | 2 |
| D | - | + | 3 | |
| R306 | A | - | - | 1 |
| D333 | A | - | - | 1 |
| F337 | A | - | - | 1 |
| K338 | A | partial | - | 2 |
| N341 | A | - | partial | 3 |
| D342 | A | - | - | 1 |
| D350 | A | - | - | 1 |
| L353 | A | - | - | 1 |
| D | - | + | 3 | |
| D368 | A | - | - | 1 |
| K | - | - | 1 | |
| R372 | A | - | + | 3 |
| G374 | A | - | - | 1 |
| G375 | A | - | - | 1 |
| D376 | A | - | - | 1 |
| E377 | A | - | - | 1 |
| F378 | A | - | - | 1 |
Table 2. Summary of phenotypes of S. typhimurium expressing BpeGReg mutants.
| Residue | Mutant | RDAR | Motility inhibition | Group |
|---|---|---|---|---|
| Globin domain | ||||
| F42 | A | + | - | 2 |
| Y43 | A | partial | + | 3 |
| S68 | A | - | - | 1 |
| M69 | A | + | - | 2 |
| Middle domain | ||||
| K226 | A | - | - | 1 |
Phe42, Tyr43, Ala68/Ser68 and Met69 in the globin distal heme pockets of EcGReg and BpeGReg are required for DGC activity
In the N-terminal globin domains, we selected residues because they were located near the distal heme pocket, affecting the orientation and migration of ligands. Expression of the EcGReg distal residue mutants, F42A, Y43A, A68T, and M69A, produced the rdar morphotype but failed to inhibit S. typhimurium motility (Figs 3A and 4A). Similar results were also observed for the BpeGReg mutants F42A and M69A (Figs 5A and 6A). On the other hand, the BpeGReg Y43A mutation severely impaired rdar development but did not affect the ability to inhibit motility (Figs 5A and 6A). Only the BpeGReg S68A mutant could neither confer the rdar morphotype nor inhibit motility (Figs 5A and 6A). These results indicate that residues in the heme pocket of the globin domain are required for EcGReg and BpeGReg to be fully active. Particularly, in BpeGReg, the hydroxyl group of Ser68 is required for full protein function.
Fig 4. Swimming motility of EcGReg mutants on 0.3% tryptone agar plates.
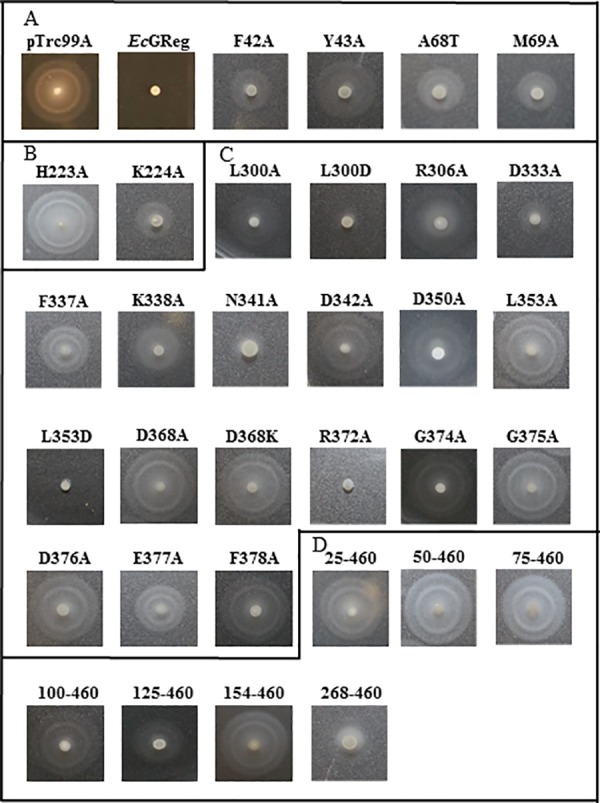
(A) Swimming motility of EcGReg globin domain mutants. (B) Swimming motility of EcGReg middle domain mutants. (C) Swimming motility of EcGReg DGC domain mutants. (D) Swimming motility of truncated EcGReg proteins.
Fig 5. Rdar morphotype development of BpeGReg mutants on Congo red agar plates.
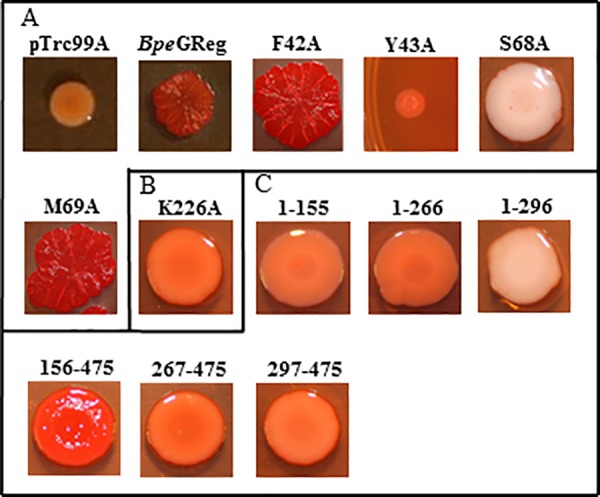
(A) Rdar morphotypes of pTrc99A vector control, BpeGReg, and BpeGReg globin domain mutants. (B) Rdar morphotypes of BpeGReg middle domain mutant. (C) Rdar morphotypes of truncated BpeGReg proteins.
His223/His225 and Lys224/Lys226 in the middle domains of EcGReg and BpeGReg are required for DGC activity
We previously demonstrated that BpeGReg could be inactivated by an H225A mutation in the middle domain, as no detectable c-di-GMP was produced in vitro and no c-di-GMP-dependent physiological effects were observed when expressed in S. typhimurium [6]. When the corresponding residue in EcGReg was mutated (H223A), this protein was also unable to induce the rdar morphotype or inhibit motility in S. typhimurium (Figs 3B and 4B). Mutation of another highly conserved residue, K224A (EcGReg)/K226A (BpeGReg), led to inactive phenotypes as well (Figs 3B–6B). Taken together, these results suggest an essential role of the middle domain for DGC activation. EcGReg His223 and Lys224 locate in a π-helix (residues 221–225) that constitutes the middle region of the α-helix B (residues 216–228) of the middle domain [7]. Although the α-helices-associated π-helices are overlooked and rarely annotated, they are present in ~15% of all proteins and tend to be associated with protein function, e.g. peristaltic-like shifts to extend the binding cavity of a substrate [34]. In the homodimer structure of the EcGReg middle domain (Fig 2B) [7], the side chains of His223 and Lys224 point towards the surrounding environment, contributing to the positive charges on the protein surface. Charged side chains on protein surfaces may play roles in allosteric regulation, protein-protein interaction, or folding/stability of protein. Further investigation will help us to understand the specific roles of the middle domain and the conserved π-helix of GCDCs.
Critical residues in the A-site and I-site of the DGC domain are required for enzyme activity
DGCs function as homodimers, with the two monomers forming the active site (A-site) at the dimer interface [35, 36]. The signature GG(D/E)EF motif constitutes part of the A-site, and several studies have suggested that absolute conservation of all five residues in this motif is required for catalysis [37, 38]. However, the first residue of the GG(D/E)EF motif appears to be flexible to substitutions in some DGCs. A DGC from Pectobacterium atrosepticum was observed to be active with Gly, Ser, or Ala in the first position, while a DGC from Vibrio cholerae was active with Gly, Ala, Met, or His [39, 40]. Using EcGReg as a model, we wanted to determine which residues of the GGDEF motif were essential for its catalytic function. We therefore mutated each residue of the GGDEF motif to Ala (G374A, G375A, D376A, E377A, and F378A). All of these mutants failed in conferring the rdar morphotype or inhibiting motility in S. typhimurium (Figs 3C and 4C), indicating that the canonical GGDEF motif is required for the DGC activity of EcGReg. We further examined the roles of other residues within the A-site. Six mutations (R306A, D333A, F337A, D342A, D350A, and L353A) resulted in a lack of in vivo function (Figs 3C and 4C). Six other mutants retained partial function: cells harboring the L300A and K338A mutants formed a partial rdar morphotype (Fig 3C), while the L300D, N341A, L353D, and R372A mutants were able to inhibit or partially inhibit motility (Fig 4C). These results highlight the requirement of additional A-site residues for DGC activity, besides those of the GGDEF motif. Many of the residues (Asp333, Phe337, Lys338, Asn341, and Asp350) directly interact with either the substrate or metal ions in the A-site [7]. The other residues may help maintain the functional structure of the DGC domain.
An inhibitory site (I-site), consisting of an RXXD motif, is found in DGC domains, including those of EcGReg and BpeGReg [41]. The I-site is located five residues upstream of the GG(D/E)EF motif (Fig 1B). Feedback inhibition occurs when a c-di-GMP dimer binds to this allosteric site, thus decreasing DGC activity [35, 41]. We tested the effect of mutating the conserved Asp in the R365XXD368 motif of EcGReg. The D368A and D368K variants both lacked the ability to produce the rdar morphotype or inhibit motility in S. typhimurium (Figs 3C and 4C). This is consistent with the findings of Kitanishi et al. [10], who observed that EcGReg D368A did not have biofilm formation activity when expressed in E. coli. Furthermore, Burns et al. [42] found that mutation of the first Arg in the BpeGReg I-site (R364A) affected oligomerization and decreased catalytic activity, leading to the conclusion that the I-site plays a role in controlling the conformation/dynamics of DGC domains, in addition to product inhibition.
The mutations characterized in this study can be classified into three groups: 1) mutants that could no longer confer the rdar morphotype or inhibit motility; 2) mutants that could confer the rdar morphotype but not inhibit motility; and 3) mutants that could inhibit motility but not confer the rdar morphotype (Tables 1 and 2). We conclude that the group 1 mutants, which are mainly located within the middle and DGC domains, include the most crucial residues required for DGC activity. The group 2 and 3 mutants are found in the globin and DGC domains. Particularly, the globin distal residue mutants Y43A are classified into different groups (EcGReg in group 2 versus BpeGReg in group 3). The differential effects of the group 2 and 3 mutants on rdar formation and motility could be due to signaling specificity. Rdar formation and motility inhibition involve a number of processes that are regulated at multiple levels, and particular DGCs and/or PDEs are often attributed to regulating specific processes [11]. The molecular mechanisms by which EcGReg and BpeGReg exert their specificities are not yet clearly understood. We presume that the group 2 and 3 mutations led to decreased c-di-GMP production, resulting in threshold levels sufficient for activating one phenotype over the other. This could involve c-di-GMP receptors with different binding affinities [43]. However, we cannot exclude the possibility that proximity to the appropriate targets was disrupted, as some of the mutations may have affected localization of the GCDC proteins in the cell or their association into protein complexes. It was shown that EcGReg, along with Ec DosP, associates with PNPase in a ribonucleoprotein complex to regulate RNA turnover [15], but it is unknown how this may be connected to biofilm formation or motility.
The biofilm formation (rdar) activities of a few EcGReg mutant variants have been previously evaluated in E. coli [10, 44], and the results were, for the most part, in agreement with what we observed for similar mutant proteins expressed in S. typhimurium. However, there was one discrepancy. While the H223A mutant displayed a wild-type phenotype in E. coli [10], our result indicated that the activity of the mutant was impaired. Differences in the c-di-GMP signaling networks of E. coli and S. typhimurium may account for the different phenotypic outcomes [45]. Contributions of species-specific DGCs/PDEs, interactions with distinct sets of proteins, or differences in the c-di-GMP thresholds needed to elicit a phenotypic response are among the possible factors that could lead to the variable effects seen in these organisms.
S. typhimurium expressing individual domains of BpeGReg and EcGReg cannot form the full rdar morphotype, while BpeGReg266 and BpeGReg296 can inhibit motility
Individual DGC domains have been shown to possess low-level enzymatic activity in vitro, which is several orders of magnitude lower than that of the full-length proteins [46]. A construct containing the middle and DGC domains of EcGReg (residues 173–460) exhibited four-fold less activity than the full-length protein, whereas the DGC domain alone (residues 297–460) did not have any detectable activity due to it being monomeric in solution [7]. In order to assess whether truncated forms of EcGReg and BpeGReg have DGC activity in vivo, we tested the functions of various constructs in S. typhimurium (Fig 7). We used EcGReg as a model to examine whether it could maintain its function when the globin domain was gradually truncated. All of the EcGReg truncated proteins could not confer a full rdar morphotype (Fig 3D), and were unable to inhibit motility as well (Fig 4D). We also tested the individual or combined domains of BpeGReg. Intriguingly, two of the BpeGReg constructs, BpeGReg266 and BpeGReg296 (residues 1–266 and 1–296), which only contain the globin and middle domains, were able to inhibit motility (Fig 6C). Compared to wild-type BpeGReg, the absorption spectra of BpeGReg155, BpeGReg266 and BpeGReg296 showed that they were able to bind heme (Fig A in S1 File), indicating that the heme-based globin was present. The presence of the globin domain alone (BpeGReg155), however, was not sufficient for eliciting a physiological response. Our data suggest that both the heme-based globin and middle domain are required for signaling and they may interact with other DGCs to form signaling clusters.
Fig 7.

Schematic diagram of truncated EcGReg (A) and BpeGReg (B).
Our analysis of the truncated proteins suggests that the complete structures of EcGReg and BpeGReg are required for full DGC activity. Previous studies have demonstrated that the cyclase activity of GCDCs is regulated by O2 binding to the globin sensor domain [6, 8, 21–23], so it is not surprising that the presence of this domain would be necessary for optimal function. However, it is unexpected to find that expression of two BpeGReg constructs lacking the DGC domain resulted in motility inhibition. As these proteins did not have the ability to synthesize c-di-GMP, we postulate that the globin and middle domains of BpeGReg may be able to interact with other DGCs, PDEs, or effector proteins to coordinate c-di-GMP signaling. Thus, the function of the truncated BpeGReg proteins may be partially rescued by its partners in mixed signaling teams. There is growing evidence for the involvement of protein-protein interactions in regulating c-di-GMP networks. For example, EcGReg forms a complex with the PDE DosP [8]; the stand-alone NO sensor H-NOX binds to HaCE, a dual-functioning DGC and PDE in Shewanella woodyi [47, 48]; and the DGC GcbC associates with its target receptor LapD in Pseudomonas fluorescens [49, 50].
Conclusion
Characterization of EcGReg and BpeGReg will provide general insights into the structure and function of GCSs. Here we provided evidence that residues Phe42, Tyr43, Ala68/Ser68 and Met69 in the distal heme pockets of EcGReg and BpeGReg could affect C-terminal DGC activity. His223/His225 and Lys224/Lys226 in the middle domains of EcGReg and BpeGReg were also required, and a number of critical residues in the A-site and I-site of the EcGReg DGC domain were identified. In addition, the full globin fold is required for GCDC activity. We further hypothesize that BpeGReg, via its globin and middle domains, may be able to form clusters with other c-di-GMP-metabolizing proteins. This may shed light on the functions of other globins, especially those single-domain proteins with unknown functions.
Supporting information
Table A. Primers used for cloning EcGReg and BpeGReg in pTrc99A. Table B. Primers used for the construction of EcGReg mutants and truncated EcGReg. Table C. Primers used for the construction of BpeGReg mutants and truncated BpeGReg. Table D. Primers used for expression of truncated BpeGReg in pET-3a. Figure A. Absorption spectra of truncated BpeGReg proteins. a) Wild-type BpeGReg (solid red line) showed heme-bound absorption spectra. b) BpeGReg155, BpeGReg266, and BpeGReg296 showed heme-bound absorption spectra, similar to that of wild-type BpeGReg.
(PDF)
Acknowledgments
Dr. Maqsudul Alam passed away before the submission of the final version of this manuscript. Dr. Xuehua Wan accepts responsibility for the integrity and validity of the data collected and analyzed.
Data Availability
All relevant data are within the paper and its Supporting Information files.
Funding Statement
This work was supported by the National Science Foundation (grant no. MCB0446431) (MA). The funder had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
References
- 1.Hou S, Larsen RW, Boudko D, Riley CW, Karatan E, Zimmer M, et al. 2000. Myoglobin-like aerotaxis transducers in Archaea and Bacteria. Nature 403:540–544 doi: 10.1038/35000570 [DOI] [PubMed] [Google Scholar]
- 2.Hou S, Freitas T, Larsen RW, Piatibratov M, Sivozhelezov V, Yamamoto A, et al. 2001. Globin-coupled sensors: a class of heme-containing sensors in Archaea and Bacteria. Proceedings of the National Academy of Sciences of the United States of America 98:9353–9358 doi: 10.1073/pnas.161185598 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 3.Freitas TAK, Hou S, Alam M. 2003. The diversity of globin-coupled sensors. FEBS Letters 552: 99–104 [DOI] [PubMed] [Google Scholar]
- 4.Freitas TAK, Saito JA, Wan X, Hou S, Alam M. 2008. Protoglobin and globin-coupled sensors In: Ghosh A, ed. The smallest biomolecules: diatomics and their interactions with heme proteins. Amsterdam London: Elsevier, 175–202 [Google Scholar]
- 5.Kitanishi K, Kobayashi K, Uchida T, Ishimori K, Igarashi J, Shimizu T. 2011. Identification and functional and spectral characterization of a globin-coupled histidine kinase from Anaeromyxobacter sp. Fw109-5. Journal of Biological Chemistry 286:35522–35534 doi: 10.1074/jbc.M111.274811 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Wan X, Tuckerman JR, Saito JA, Freitas TAK, Newhouse JS, Denery JR, et al. 2009. Globins synthesize the second messenger c-di-GMP in bacteria. Journal of Molecular Biology 388:262–270 doi: 10.1016/j.jmb.2009.03.015 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Tarnawski M, Barends TR, Schlichting I. 2015. Structural analysis of an oxygen-regulated diguanylate cyclase. Acta Crystallographica D Biological Crystallography 71:2158–2177 doi: 10.1107/S139900471501545X [DOI] [PubMed] [Google Scholar]
- 8.Tuckerman JR, Gonzalez G, Sousa EH, Wan X, Saito JA, Alam M, et al. 2009. An oxygen-sensing diguanylate cyclase and phosphodiesterase couple for c-di-GMP control. Biochemistry 48:9764–9774 doi: 10.1021/bi901409g [DOI] [PubMed] [Google Scholar]
- 9.Thijs L, Vinck E, Bolli A, Trandafir F, Wan X, Hoogewijs D, et al. 2007. Characterization of a globin-coupled oxygen sensor with a gene-regulating function. Journal of Biological Chemistry 282:37325–37340 doi: 10.1074/jbc.M705541200 [DOI] [PubMed] [Google Scholar]
- 10.Kitanishi K, Kobayashi K, Kawamura Y, Ishigami I, Ogura T, Nakajima K, et al. 2010. Important roles of Tyr43 at the putative heme distal side in the oxygen recognition and stability of the Fe(II)-O2 complex of YddV, a globin-coupled heme-based oxygen sensor diguanylate cyclase. Biochemistry 49:10381–10393 doi: 10.1021/bi100733q [DOI] [PubMed] [Google Scholar]
- 11.Römling U, Galperin MY, Gomelsky M. 2013. Cyclic di-GMP: the first 25 years of a universal bacterial second messenger. Microbiology and Molecular Biology Reviews 77:1–52 doi: 10.1128/MMBR.00043-12 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Ryan RP, Tolker-Nielsen T, Dow JM. 2012. When the PilZ don’t work: effectors for cyclic di-GMP action in bacteria. Trends in Microbiology 20:235–242 doi: 10.1016/j.tim.2012.02.008 [DOI] [PubMed] [Google Scholar]
- 13.Amikam D, Galperin MY. 2006. PilZ domain is part of the bacterial c-di-GMP binding protein. Bioinformatics 22:3–6 doi: 10.1093/bioinformatics/bti739 [DOI] [PubMed] [Google Scholar]
- 14.Fang X, Ahmad I, Blanka A, Schottkowski M, Cimdins A, Galperin MY, et al. 2014. GIL, a new c-di-GMP-binding protein domain involved in regulation of cellulose synthesis in enterobacteria. Molecular Microbiology 93:439–452 doi: 10.1111/mmi.12672 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Tuckerman JR, Gonzalez G, Gilles-Gonzalez MA. 2011. Cyclic di-GMP activation of polynucleotide phosphorylase signal-dependent RNA processing. Journal of Molecular Biology 407:633–639 doi: 10.1016/j.jmb.2011.02.019 [DOI] [PubMed] [Google Scholar]
- 16.Lori C, Ozaki S, Steiner S, Böhm R, Abel S, Dubey BN, et al. 2015. Cyclic di-GMP acts as a cell cycle oscillator to drive chromosome replication. Nature 523:236–239 doi: 10.1038/nature14473 [DOI] [PubMed] [Google Scholar]
- 17.Sudarsan N, Lee ER, Weinberg Z, Moy R H, Kim JN, Link KH, et al. 2008. Riboswitches in eubacteria sense the second messenger cyclic di-GMP. Science 321:411–413 doi: 10.1126/science.1159519 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Burdette DL, Monroe KM, Sotelo-Troha K, Iwig JS, Eckert B, Hyodo M, et al. 2011. STING is a direct innate immune sensor of cyclic di-GMP. Nature 478:515–518 doi: 10.1038/nature10429 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Parvatiyar K, Zhang Z, Teles RM, Ouyang S, Jiang Y, Iyer SS, et al. 2012. The helicase DDX41 recognizes the bacterial secondary messengers cyclic di-GMP and cyclic di-AMP to activate a type I interferon immune response. Nature Immunology 13:1155–1161 doi: 10.1038/ni.2460 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Li W, Cui T, Hu L, Wang Z, Li Z, He Z-G. 2015b. Cyclic diguanylate monophosphate directly binds to human siderocalin and inhibits its antibacterial activity. Nature Communications 6:8330. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Sawai H, Yoshioka S, Uchida T, Hyodo M, Hayakawa Y, Ishimori K, et al. 2010. Molecular oxygen regulates the enzymatic activity of a heme-containing diguanylate cyclase (HemDGC) for the synthesis of cyclic di-GMP. Biochimica et Biophysica Acta 1804:166–172 doi: 10.1016/j.bbapap.2009.09.028 [DOI] [PubMed] [Google Scholar]
- 22.Wu C, Cheng YY, Yin H, Song XN, Li WW, Zhou XX, et al. 2013. Oxygen promotes biofilm formation of Shewanella putrefaciens CN32 through a diguanylate cyclase and an adhesin. Scientific Reports 3:1945 doi: 10.1038/srep01945 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Burns JL, Deer DD, Weinert EE. 2014. Oligomeric state affects oxygen dissociation and diguanylate cyclase activity of globin coupled sensors. Molecular BioSystems 10:2823–2826 doi: 10.1039/c4mb00366g [DOI] [PubMed] [Google Scholar]
- 24.Méndez-Ortiz MM, Hyodo M, Hayakawa Y, Membrillo-Hernández J. 2006. Genome-wide transcriptional profile of Escherichia coli in response to high levels of the second messenger 3',5'-cyclic diguanylic acid. Journal of Biological Chemistry 281:8090–8099 doi: 10.1074/jbc.M510701200 [DOI] [PubMed] [Google Scholar]
- 25.Katoh K, Standley DM. 2013. MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Molecular Biology and Evolution 30:772–780 doi: 10.1093/molbev/mst010 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Li W, Cowley A, Uludag M, Gur T, McWilliam H, Squizzato S, et al. 2015a. The EMBL-EBI bioinformatics web and programmatic tools framework. Nucleic Acids Research 43:W580–W584 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Brown NP, Leroy C, Sander C. 1998. MView: a web-compatible database search or multiple alignment viewer. Bioinformatics 14:380–381 [DOI] [PubMed] [Google Scholar]
- 28.Pettersen EF, Goddard TD, Huang CC, Couch GS, Greenblatt DM, Meng EC, et al. 2004. UCSF Chimera—a visualization system for exploratory research and analysis. Journal of Computational Chemistry 25:1605–1612 doi: 10.1002/jcc.20084 [DOI] [PubMed] [Google Scholar]
- 29.Piatibratov M, Hou S, Brooun A, Yang J, Chen H, Alam M. (2000). Expression and fast-flow purifica- tion of a polyhistidine-tagged myoglobin-like aero- taxis transducer. Biochim. Biophys. Acta 1524: 149–154. [DOI] [PubMed] [Google Scholar]
- 30.Simm R, Morr M, Kader A, Nimtz M, Römling U. 2004. GGDEF and EAL domains inversely regulate cyclic di-GMP levels and transition from sessility to motility. Molecular Microbiology 53:1123–1134 doi: 10.1111/j.1365-2958.2004.04206.x [DOI] [PubMed] [Google Scholar]
- 31.Römling U, Bian Z, Hammar M, Sierralta WD, Normark S. 1998. Curli fibers are highly conserved between Salmonella typhimurium and Escherichia coli with respect to operon structure and regulation. Journal of Bacteriology 180:722–731 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Zogaj X, Nimtz M, Rohde M, Bokranz W, Romling U. 2001. The multicellular morphotypes of Salmonella typhimurium and Escherichia coli produce cellulose as the second component of the extracellular matrix. Molecular Microbiology 39:1452–1463 [DOI] [PubMed] [Google Scholar]
- 33.Kader A, Simm R, Gerstel U, Morr M, Römling U. 2006. Hierarchical involvement of various GGDEF domain proteins in rdar morphotype development of Salmonella enterica serovar Typhimurium. Molecular Microbiology 60:602–616 doi: 10.1111/j.1365-2958.2006.05123.x [DOI] [PubMed] [Google Scholar]
- 34.Cooley RB, Arp DJ, Karplus PA. 2010. Evolutionary origin of a secondary structure: π-helices as cryptic but widespread insertional variations of α-helices enhancing protein functionality. Journal of Molecular Biology 404:232–246 doi: 10.1016/j.jmb.2010.09.034 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Chan C, Paul R, Samoray D, Amiot NC, Giese B, Jenal U, et al. 2004. Structural basis of activity and allosteric control of diguanylate cyclase. Proceedings of the National Academy of Sciences of the United States of America 101:17084–17089 doi: 10.1073/pnas.0406134101 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Paul R, Abel S, Wassmann P, Beck A, Heerklotz H, Jenal U. 2007. Activation of the diguanylate cyclase PleD by phosphorylation-mediated dimerization. Journal of Biological Chemistry 282:29170–29177 doi: 10.1074/jbc.M704702200 [DOI] [PubMed] [Google Scholar]
- 37.Kirillina O, Fetherston JD, Bobrov AG, Abney J, Perry RD. 2004. HmsP, a putative phosphodiesterase, and HmsT, a putative diguanylate cyclase, control Hms-dependent biofilm formation in Yersinia pestis. Molecular Microbiology 54:75–88 doi: 10.1111/j.1365-2958.2004.04253.x [DOI] [PubMed] [Google Scholar]
- 38.Malone JG, Williams R, Christen M, Jenal U, Spiers AJ, Rainey PB. 2007. The structure-function relationship of WspR, a Pseudomonas fluorescens response regulator with a GGDEF output domain. Microbiology 153:980–994 doi: 10.1099/mic.0.2006/002824-0 [DOI] [PubMed] [Google Scholar]
- 39.Perez-Mendoza D, Coulthurst SJ, Humphris S, Campbell E, Welch M, Toth IK, et al. 2011. A multi-repeat adhesin of the phytopathogen, Pectobacterium atrosepticum, is secreted by a type I pathway and is subject to complex regulation involving a non-canonical diguanylate cyclase. Molecular Microbiology 82:719–733 doi: 10.1111/j.1365-2958.2011.07849.x [DOI] [PubMed] [Google Scholar]
- 40.Hunter JL, Severin GB, Koestler BJ, Waters CM. 2014. The Vibrio cholerae diguanylate cyclase VCA0965 has an AGDEF active site and synthesizes cyclic di-GMP. BMC Microbiology 14:22 doi: 10.1186/1471-2180-14-22 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Christen B, Christen M, Paul R, Schmid F, Folcher M, Jenoe P, et al. 2006. Allosteric control of cyclic di-GMP signaling. Journal of Biological Chemistry 281:32015–32024 doi: 10.1074/jbc.M603589200 [DOI] [PubMed] [Google Scholar]
- 42.Burns JL, Rivera S, Deer DD, Joynt SC, Dvorak D, Weinert EE. 2016. Oxygen and bis(3',5')-cyclic dimeric guanosine monophosphate binding control oligomerization state equilibria of diguanylate cyclase-containing globin coupled sensors. Biochemistry 55:6642–6651 doi: 10.1021/acs.biochem.6b00526 [DOI] [PubMed] [Google Scholar]
- 43.Pultz IS, Christen M, Kulasekara HD, Kennard A, Kulasekara B, Miller SI. 2012. The response threshold of Salmonella PilZ domain proteins is determined by their binding affinities for c-di-GMP. Molecular Microbiology 86:1424–1440 doi: 10.1111/mmi.12066 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Nakajima K, Kitanishi K, Kobayashi K, Kobayashi N, Igarashi J, Shimizu T. 2012. Leu65 in the heme distal side is critical for the stability of the Fe(II)-O2 complex of YddV, a globin-coupled oxygen sensor diguanylate cyclase. Journal of Inorganic Biochemistry 108:163–170 doi: 10.1016/j.jinorgbio.2011.09.019 [DOI] [PubMed] [Google Scholar]
- 45.Sommerfeldt N, Possling A, Becker G, Pesavento C, Tschowri N, Hengge R. 2009. Gene expression patterns and differential input into curli fimbriae regulation of all GGDEF/EAL domain proteins in Escherichia coli. Microbiology 155:1318–1331 doi: 10.1099/mic.0.024257-0 [DOI] [PubMed] [Google Scholar]
- 46.Ryjenkov DA, Tarutina M, Moskvin OV, Gomelsky M. 2005. Cyclic diguanylate is a ubiquitous signaling molecule in bacteria: insights into biochemistry of the GGDEF protein domain. Journal of Bacteriology 187:1792–1798 doi: 10.1128/JB.187.5.1792-1798.2005 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Liu N, Xu Y, Hossain S, Huang N, Coursolle D, Gralnick JA, et al. 2012. Nitric oxide regulation of cyclic di-GMP synthesis and hydrolysis in Shewanella woodyi. Biochemistry 51:2087–2099 doi: 10.1021/bi201753f [DOI] [PubMed] [Google Scholar]
- 48.Lahiri T, Luan B, Raleigh DP, Boon EM. 2014. A structural basis for the regulation of an H-NOX-associated cyclic-di-GMP synthase/phosphodiesterase enzyme by nitric oxide-bound H-NOX. Biochemistry 53:2126–2135 doi: 10.1021/bi401597m [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Dahlstrom KM, Giglio KM, Collins AJ, Sondermann H, O’Toole GA. 2015. Contribution of physical interactions to signaling specificity between a diguanylate cyclase and its effector. mBio 6:e01978–15 doi: 10.1128/mBio.01978-15 [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Dahlstrom KM, Giglio KM, Sondermann H, O’Toole GA. 2016. The inhibitory site of a diguanylate cyclase is a necessary element for interaction and signaling with an effector protein. Journal of Bacteriology 198:1595–1603 doi: 10.1128/JB.00090-16 [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Table A. Primers used for cloning EcGReg and BpeGReg in pTrc99A. Table B. Primers used for the construction of EcGReg mutants and truncated EcGReg. Table C. Primers used for the construction of BpeGReg mutants and truncated BpeGReg. Table D. Primers used for expression of truncated BpeGReg in pET-3a. Figure A. Absorption spectra of truncated BpeGReg proteins. a) Wild-type BpeGReg (solid red line) showed heme-bound absorption spectra. b) BpeGReg155, BpeGReg266, and BpeGReg296 showed heme-bound absorption spectra, similar to that of wild-type BpeGReg.
(PDF)
Data Availability Statement
All relevant data are within the paper and its Supporting Information files.