

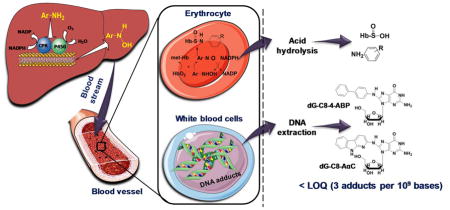
Abstract
Aromatic amines covalently bound to hemoglobin (Hb) as sulfinamide adducts at the cysteine 93 residue of the Hb β chain have served as biomarkers to assess exposure to this class of human carcinogens for the past 30 years. In this study, we report that 2-amino-9H-pyrido[2,3-b]indole (AαC), an abundant carcinogenic heterocyclic aromatic amine formed in tobacco smoke and charred cooked meats, also reacts with Hb to form a sulfinamide adduct. A novel nanoflow liquid chromatography/ion trap multistage mass spectrometry (nanoLC-IT/MS3) method was established to assess exposure to AαC and the tobacco-associated bladder carcinogen 4-aminobiphenyl (4-ABP) through their Hb sulfinamide adducts. Following mild acid hydrolysis of Hb in vitro, the liberated AαC and 4-ABP were derivatized with acetic anhydride to form the N-acetylated amines, which were measured by nanoLC-IT/MS3. The limits of quantification (LOQ) for AαC- and 4-ABP-Hb sulfinamide adducts were ≤ 7.1 picograms per gram Hb. In a pilot study, the mean level of Hb sulfinamide adducts of AαC and 4-ABP, were, respectively, 3.3-fold and 4.8-fold higher in smokers (> 20 cigarettes/day) than nonsmokers. In contrast, the major DNA adducts of 4-ABP, N-(2′-deoxyguanosin-8-yl)-4-aminobiphenyl, and AαC, N-(2′-deoxyguanosin-8-yl)-2-amino-9H-pyrido[2,3-b]indole, were below the LOQ (3 adducts per 109 bases) in white blood cell (WBC) DNA of smokers and nonsmokers. These findings reaffirm that tobacco smoke is a major source of exposure to AαC. Hb sulfinamide adducts are suitable biomarkers to biomonitor 4-ABP and AαC; however, neither carcinogen binds to DNA in WBC, even in heavy smokers, at levels sufficient for biomonitoring.
Keywords: 2-amino-9H-pyrido[2,3-b]indole; 4-aminobiphenyl; heterocyclic aromatic amines; aromatic amines; tobacco smoke; hemoglobin
Graphical Abstract
Introduction
More than 60 carcinogens, including polycyclic aromatic hydrocarbons (PAHs), N-nitrosamines, aromatic amines, and heterocyclic aromatic amines (HAAs) are present in tobacco smoke.1,2 Epidemiological studies have shown that tobacco smoke is a risk factor not only for lung cancer, but also for cancers of the liver, bladder, and the gastrointestinal tract.3,4 PAHs and N-nitrosamines are believed to contribute to lung cancer while aromatic amines, such as 4-aminobiophenyl (4-ABP), are thought to be involved in the pathogenesis of bladder cancer of smokers.3,5 2-Amino-9H-pyrido[2,3-b]indole (AαC) is the most abundant carcinogenic HAA present in mainstream tobacco smoke, with levels ranging from 37 to 258 ng/cigarette.6,7 These amounts are 25–100-fold higher than the levels of 4-ABP and benzo[a]pyrene (B[a]P), and comparable to the levels of 4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone (NNK) that occur in tobacco smoke.8–10 4-ABP, B[a]P, and NNK are recognized as human carcinogens.4
AαC is a liver and blood vessel carcinogen, a lacI transgene colon mutagen, and an inducer of colonic aberrant crypt foci, an early biomarker of colon neoplasia, in mice.11–14 Moreover, high levels of DNA adducts are detected in the liver, colon and other extrahepatic tissues of mice upon AαC treatment.13,15,16 The carcinogenic potential of AαC in humans is unknown, however, AαC is genotoxic to human lymphoblastoid cells (MLC-5) and peripheral blood lymphocytes cells.17,18 We previously reported that primary human hepatocytes also efficiently bioactivate AαC to its N-hydroxylated metabolite, 2-hydroxamino-9H-pyrido[2,3-b]indole (HONH-AαC),19,20 which binds to DNA and can lead to mutations.12,21 Therefore, AαC present in tobacco smoke may contribute to DNA damage of the liver and digestive tract of smokers. Long-lived biomarkers of AαC are required in molecular epidemiology studies to assess the role of AαC in tobacco-associated cancers.
Blood protein adducts of carcinogens have been used to estimate exposures and assess health risks of tobacco-associated carcinogens by mass spectrometry based methods.22–24 Hemoglobin (Hb) adducts have been used to biomonitor environmental and occupational exposures to aromatic amines for the past three decades.22,25–28 In contrast to many DNA adducts that undergo repair, stable blood protein carcinogen adducts are expected to accumulate during the kinetics of the lifetime of the protein and facilitate chemical analysis. Arylamines undergo metabolism by hepatic cytochrome P450 to form the arylhydroxylamine metabolites,29 which undergo systemic circulation through the blood. Once up-taken by the erythrocytes, the arylhydroxylamines can undergo a co-oxidation reaction with oxy-hemolgobin (HbO2) to form methemoglobin (met-Hb) and the arylnitroso intermediates,30,31 which react with the β-Cys93 chain of Hb to form Hb arylsulfinamide adducts (Scheme 1).31–34
Scheme 1.

Metabolism of 4-ABP and AαC and formation of their N-hydroxylated metabolites in liver, followed by uptake of HONH-4-ABP and HONH-AαC in erythrocytes and WBC and reaction with HbO2 and DNA.
Hb arylsulfinamide adducts are easily hydrolyzed by mild acid or base treatment in vitro, leading to the release of the parent arylamine carcinogens and the Hb β-Cys93 sulfinic acid (Scheme 1). 32,33 The arylamines are analyzed, following chemical derivatization with fluorinated anhydrides, by gas chromatography-negative ion chemical ionization mass spectrometry (GC-NICI/MS).25,33,35 The arylamine-Hb adduct biomarker was established thirty years ago 33,35 and it is still used world-wide today.26–28
Peripheral white blood cells (WBC) are another surrogate blood biospecimen, which have been used to assess the exposure to tobacco genotoxicants primarily by screening of DNA adducts with 32P-postlabeling methods.36–39 Some studies have reported elevated levels of putative bulky aromatic type adducts in WBC DNA of smokers compared to nonsmokers.40–42 However, the identities of the chemicals in tobacco smoke that damage DNA are uncertain because of the lack of specificity of the 32P-postlabeling assay. There are several studies reporting positive correlations between the levels of 4-ABP-Hb adduct to the levels of 32P-postabeled DNA lesions in WBC of smokers,41,43,44 but to our knowledge, MS-based methods to measure both arylamine-Hb adducts and their respective WBC DNA adducts have not been reported.
In this study, we have developed a novel approach using nanoflow liquid chromatography-ion trap multistage mass spectrometry (nanoLC-IT/MS3) as a platform to measure the 4-ABP- and AαC-Hb sufinamide adducts in humans. The released amines were derivatized with acetic anhydride to form the N-acetylated derivatives, which were measured by nanoLC-IT/MS3. The nanoLC-IT/MS3 assay has a limit of quantification (LOQ) value of 7.1 picograms of 4-ABP and 6.6 picograms AαC per gram Hb, LOQ values which are comparable to the GC-NICI/MS method for the 4-ABP adduct.33,35 We also employed our ultraperformance liquid chromatography-ion trap/multistage mass spectrometry (UPLC-IT/MS3) method to measure DNA adducts of 4-ABP and AαC in WBC of the same subjects.45,46
Materials and Methods
Caution: AαC and 4-ABP are carcinogens and should be handled with caution in a well ventilated fume hood with the appropriate protective clothing.
Chemicals and Reagents
AαC, 4-ABP and d5-4-ABP (2′,3′,4′,5′,6′-2H5-4-ABP, > 99% isotopic purity) were purchased from Toronto-Research Chemicals, Inc. (Toronto, Ontario, Canada). [13C6]AαC (4b,5,6,7,8,8a-13C6-AαC, 97% isotopic purity) was a gift from Dr. Daniel Doerge, National Center for Toxicological Research (Jefferson, AR). 4-Nitrobiphenyl was obtained from Apolo Scientific (Stockport, UK.) Triethylamine was purchased from Fisher Scientific (Fair Lawn, NJ). 2-Hydroxyamino-9H-pyrido[2,3-b]indole (HONH-AαC) and 4-hydroxyamino-biphenyl (HONH-ABP) were prepared by the reduction of their nitro derivatives as previously described,47,48 dried by vacuum centrifugation, and reconstituted in DMSO:Ethanol (3:1) and stored in liquid nitrogen. N-(2′-Deoxyguanosin-8-yl)-4-aminobiphenyl (dG-C8-4-ABP), [13C10]dG-C8-4-ABP (isotopic purity > 99.5%), N-(2′-deoxyguanosin-8-yl)-2-amino-9H-pyrido[2,3-b]indole (dG-C8-AαC) and [13C10]dG-C8-AαC (isotopic purity > 98.5%) were prepared as previously described.21 Mass cut off filters (Amicon Ultra 0.5 mL Centrifugal Filters, 10 K) were purchased from EMD Millipore (Merck KGaA, Darmstadt, Germany). SOLA HRP cartridges were from Thermo Fisher Scientific (Bellefonte, PA). Pooled human whole blood samples were obtained from Bioreclamation LLC (Hicksville, NY). CT DNA containing known levels of 4-ABP was provided by Dr. Frederick A. Beland from the National Center for Toxicology Research/U.S. FDA.49 De-identified red blood cells and white blood cells of smokers and nonsmokers were obtained from the National University of Singapore.
Isolation of Red Blood Cells
Pooled human whole blood was centrifuged at 2,000 g for 10 min. The plasma and the white blood cells were collected and stored at −80 °C. The red blood cell pellet was washed with saline buffer (0.9% of NaCl) twice, resuspended in 2 volumes of water and sonicated for 10 min. The red blood cells lysate was then centrifuged at 21,000 g for 30 min to remove cell debris and the supernatant containing Hb was collected. The concentration of Hb was measured by UV/Visible spectroscopy (542 nm, ε(HbO2)= 14.62 mM−1 cm−1; 632 nm, ε(met-Hb)= 4.03 mM−1 cm−1).50
Modification of Hb with HONH-ABP and HONH-AαC in vitro
Hb (100 μg) was diluted in 100 mM sodium phosphate buffer (500 μL, pH 7.4), followed by the addition of a 3-fold molar excess of HONH-4-ABP or HONH-AαC, and incubated for 3 h at 37 °C. The unbound carcinogens were removed from the modified Hb by centrifugation using molecular mass cut-off filters (10K). The Hb was washed twice with 100 mM phosphate buffer (500 μL, pH 7.4) to remove the remaining carcinogens. The efficiency of removal of 4-ABP and AαC was evaluated by spiking experiments, which showed that all the spiked chemicals were removed by this filtration process (data not shown). The levels of Hb modification with carcinogens were 63 ng of Hb-4-ABP sulfinamide and 190 ng Hb-AαC sulfinamide per 100 μg Hb, based on recovery of 4-ABP and AαC from modified Hb, following acid treatment (vide infra).51
Isolation and chemical derivatization of 4-ABP- and AαC-Hb sulfinamide adducts from human blood samples
This study was approved by the Institutional Review Boards of the National University of Singapore, and the University of Minnesota. All blood samples were deidentified. The Hb from packed red blood cells of smokers, non-smokers, and commercial pooled human blood were processed as described above and filtered with molecular mass cut-off filters (10K) to remove low molecular weight compounds. Then, HbO2 (30 mg) was transferred to Eppendorf tubes (2 mL) for acid hydrolysis. [13C6]AαC (3.5 pg) and d5-4-ABP (3.5 pg) were added as internal standards, diluted with LC-MS grade H2O (900 μL), followed by addition of 1N HCl (100 μL). The hydrolysis of the 4-ABP- and AαC-Hb sulfinamide was achieved by incubation for 3 h at 37 °C. Then, 1N NaOH (110 μL) was added to the solution, and the released AαC and ABP were extracted twice with ethyl acetate (900 μL). DMSO (50 μL) was added to the solution, and the extracts were evaporated under a gentle flow of N2 gas, until a volume of ~100 μL remained. The samples were diluted with H2O (1 mL) and applied to a SOLA HRP cartridge, preconditioned with CH3OH (1 mL), followed by H2O (1 mL) The cartridges were washed with 20% (v/v) CH3OH in H2O (1 mL), and the cartridges were dried under vaccum by aspiration (≤ 20 inches of Hg) for several minutes to remove residual water. The 4-ABP and AαC were eluted from the cartridges with ethyl acetate (1 mL), and anhydrous MgSO4 (30 mg) was added to dry the eluent for 30 min. Thereafter, the eluent was centrifuged to remove MgSO4 particles, and the ethyl acetate was transferred to another Eppendorf tube (2 mL) and concentrated by a stream of N2 gas to a final volume of 200 μL. The amines were acetylated by adding freshly prepared triethylamine (10 μL, 3% v/v in ethyl acetate, pre-dried with MgSO4 (0.05 mg/mL)), followed by acetic anhydride (10 μL, 3% v/v in ethyl acetate) and incubated at 45 °C for 3 h. Thereafter, another 10 μL of the triethylamine and acetic anhydride reagents were added and derivatization was continued for another 3 h. After incubation, DMSO (5 μL) was added to the samples, followed by concentration of the residues to a volume of 10 μL under a gentle stream of N2 gas. The samples were reconstituted in 10% DMSO in LC/MS grade water (50 μL).
Calibration curves
HbO2 (30 mg) from Bioreclamation LLC found to contain low amounts of 4-ABP and AαC (below the LOQ values, vide infra) was hydrolyzed with acid, followed by alkaline treatment and extraction with ethyl acetate as described above. The d5-4-ABP and [13C6]AαC (3.5 pg, 117 pg/g Hb) and varying amount ratios of unlabeled 4-ABP to d5-4-ABP (eight calibrants, from 0, 0.05, up to 2.5) and varying amount ratios of unlabeled AαC to [13C6]AαC (eight calibrants, from 0, 0.04, up to 2.0) were added to the extract and derivatized with acetic anhydride as described above. The samples were concentrated under a gentle stream of N2 gas and reconstituted in 10% DMSO in LC/MS grade water (50 μL). Data were fitted to a straight line (area of response of 4-ABP or AαC/internal standard versus the amount of 4-ABP or AαC/internal standard) using ordinary least-squares with equal weightings.
Stability of Hb-4-ABP- and Hb-AαC sulfinamide adducts in vitro
Hb modified with HONH-ABP and HONH-AαC were diluted with 30 mg of unmodified Hb, in 100 mM sodium phosphate buffer (500 μL, pH 7.4, containing 0.9% NaCl). The level of 4-ABP-Hb sulfinamide was 165 pg/g Hb and the level of the AαC-Hb sulfinamide was 85 pg/g Hb. The samples were incubated at 37 °C for different times (0, 8, 24, 48, 72, 96 hours). Mass cut-off filter (10K) was used to terminate the time-dependent incubation and the filtered Hb was hydrolyzed for adducts measurement as described above.
Measurement of cotinine in human plasma
Cotinine was measured in plasma by LC-MS/MS with d3-methyl]-cotinine employed as the internal standard.52 The analysis was performed by the laboratory of Dr. Sharon Murphy, University of Minnesota.
Isolation of DNA from human WBC
The extraction of DNA from WBC was performed with the Gentra® Puregene® kit (Qiagen, Valencia, CA) with modifications. The white blood cell fraction (200 μL) was mixed with of red blood cell lysis solution (600 μL) and incubated for 10 min at room temperature. The mixture then underwent centrifugation for 15 min at 2500 g. The white blood cells pellets were collected, homogenized in TE buffer (300 μL, 50 mM Tris-HCl containing 10 mM EDTA and 10 mM β-mercaptoethanol, pH 8.0) and incubated with RNase T1 (318.75 U) and RNase A (20 μg) for 90 min at 37 °C. Proteinase K (200 μg) and SDS (0.05% final concentration) were added and the mixture was incubated for 2 h at 37 °C. After cooling the mixture on ice for 10 min, protein precipitation solution (100 μL) was added and the samples were incubated for 2 h at −20 °C. The precipitated proteins were removed by centrifugation (16,000 × g, 5 min) and the DNA was precipitated from the supernatant over night at −20 °C, following the addition of 0.1 volume (~ 50 μL) of 5 M NaCl and 2 volume (800 μL) of chilled isopropanol alcohol. The DNA was washed with ethanol (70%), and the DNA was reconstituted in of LC/MS grade water (200 μL). The concentration of DNA was determined using an Agilent 8453 UV/vis spectrophotometer (Agilent Technologies, Santa Clara, CA). Spiking experiments, were conducted by adding 10 μg of modified CT-DNA containing known amounts of dG-C8-4ABP (1 adduct per 108 per bases) to the WBC prior to DNA extraction to assess recovery of the adduct.
Enzymatic digestion of DNA
Each DNA sample (10 μg or 50 μg) was spiked with isotopically labeled internal standards ([13C10]dG-C8-4-ABP and [13C10]dG-C8-AαC) at a level of 5 adduct per 108 bases and digested in 5 mM Bis-Tris-HCl buffer (pH 7.1) as previously described.20 Briefly, DNA (10 or 50 μg) was incubated with DNase I and Nuclease P1 at 37 °C for 3.5 h, followed by incubation with alkaline phosphatase and phosphodiesterase at 37 °C for 18 h. Digested DNA solutions were evaporated to dryness by vacuum centrifugation and re-suspended in 25 μl of 1:1 water/DMSO. After 5 min sonication, the digests were centrifuged for 5 min at 22000 g, and the supernatants were transferred to LC vials.
NanoLC-IT/MS3 measurement of Hb adducts
N-Acetyl-4-ABP and N-acetyl-4-AαC were measured with a nanoACQUITY UPLC system (Waters Corporation, Milford, MA) interfaced to an Nanoflex source and the Velos Ion-Trap mass spectrometer (Thermo Fisher, San Jose, CA). A Thermo C18 trap column (180 μm x 20 mm, 5 μm particle size) was used for online sample enrichment. Mobile phase A was H2O with 0.05% formic acid, and mobile phase B contained 95% CH3CN with 5% H2O and 0.05% formic acid. The analytical column was a self-packed Luna C18 column (5 μm particle size, Phenomenex Corp., Torrance, CA) prepared with a PicoTip emitter (75 μm × 200 mm, 10 μm tip ID, New Objective Inc., Woburn, MA) was used for LC separation at a flow rate of 0.3 μL/min. The derivatization mixture (5 μL) was injected onto the trap column and washed with mobile phase A at a flow rate of 8 μL/min for 5 min. The sample was then back flushed on to the analytical column. The chromatography commenced at a solvent composition of 5% B and 95% A for 5 min, and then increased to 70% B at 10 min, and 90% B at 18 min, and reached 99% B at 19 min. Then, the flow rate was increased to 0.6 μL/min and held at 99% B for 3 min. Thereafter, the column was re-equilibrated back to the starting solvent conditions of 5% B and 95% A at 24 min, and the flow rate was reduced back to 0.3 μL/min, and held for 4 min. The capillary temperature was 270 °C and ionization voltage was 2.5 kV for positive ion mode. The activation Q value was 0.35, and the activation time was 10 ms. The N-acetylated amines were measured at the MS3 scan stage. The collision energy was set at 34 eV for N-acetyl-4-ABP and 42 eV for N-acetyl-AαC for both MS2 and MS3 scan stages. The isolation width was set at m/z 1 for MS2 and MS3. The transitions used for N-acetyl-4-ABP were: [M+H]+ at m/z 212.1 > 170.1 > 143.1, 153.1, 155.1; N-acetyl-d5-4-ABP: [M+H]+ at m/z 217.1 > 175.1 > 148.1, 155.1, 156.1, 157.1, 158.1; N-acetyl-AαC: [M+H]+ at m/z 226.1 > 208.1 > 167.1, 185.1; and N-acetyl-[13C6]AαC: [M+H]+ at m/z 232.1 > 214.1 > 173.1, 191.1.
UPLC-IT/MS3 measurement of DNA adducts
dG-C8-4-ABP and dG-C8-AαC were measured with the NanoACQUITY UPLC system (Waters Corp, New Milford, MA) equipped with a Thermo C18 trap column (180 μm x 20 mm, 5 μm particle size), a Michrom C18 AQ column (0.1 mm × 150 mm, 3 μm particle size), and a Michrom Captive Spray source interfaced with LTQ Velos. DNA digest (10 μL containing 4 μg DNA) were injected into the trap column and washed with mobile phase A (0.01% HCO2H) for 4 min at a flow rate of 12 μL/min before back-flushing the analytes onto analytical column. The analytes were separated using a linear gradient starting at 1% B (95% CH3CN, 0.01% HCO2H) and increased to 100 B over 15 min with a flow rate of 1 μl/min. the adducts were measured at the MS3 scan stage in the positive ionization mode. The spray voltage was set at 1.95 kV and the capillary temperature was 270 °C. The activation Q value was 0.35, and the activation times was 10 ms. The isolation width was set, respectively, at m/z 4.0 and m/z 1.0 for the MS2 and MS3 scan stages. The collision energy was set at 35 eV for MS2 and 45 eV for MS3. The transitions were: dG-C8-AαC [M+H]+ at m/z 449.1 > 333.1 > 209.1, 291.1, 316.1; [13C10]dG-C8-AαC: [M+H]+ at m/z 459.1 > 338.1 > 210.1, 295.1, 321.1; dG-C8-4-ABP: [M+H]+ at m/z 435.1 > 319.1 > 249.1, 277.1, 302.1 and [13C10]dG-C8-4-ABP: [M+H]+ at m/z 445.1 > 324.1 > 252.1, 281.1, 307.1.
Results
Hb sulfinamide adducts and WBC DNA adducts of 4-ABP and AαC
Aromatic amines and HAAs undergo bioactivation by hepatic cytochrome P450 to form the genotoxic N-hydroxylated metabolites,29,53–55 which can reach the blood stream and be up-taken by the erythrocyte to form sulfinamide adducts with Hb (Scheme 1). The reactive N-hydroxylated metabolites also may form covalent DNA adducts in WBC. We examined the reactivity of HONH-4-ABP and the N-hydroxylated metabolites of AαC, and 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP) and 2-amino-3,8-dimethylimidazo[4,5-f]quinoxaline (MeIQx), two prevalent carcinogenic HAAs formed in cooked meats,14,56 to form adducts with HbO2.51 The N-hydroxylated metabolites of MeIQx and PhIP did not react with the oxy-heme complex and induce met-Hb formation, and as a consequence, the chemical modification of the β-Cys93 residue of Hb was negligible.51 However, HONH-AαC induced methemoglobinemia and formed an Hb sulfinamide adduct in human erythrocytes at greater levels than HONH-4-ABP.51 We also reported that AαC undergoes bioactivation and forms DNA adducts in human T lymphocytes in cell culture at higher levels than 4-ABP, PhIP and MeIQx.57 Therefore, we sought to determine if AαC underwent bioactivation in vivo to form a sulfinamide adduct with Hb using a novel nanoLC-IT/MS3 platform. The major DNA adducts of 4-ABP, N-(2′-deoxyguanosin-8-yl)-4-aminobiphenyl (dG-C8-4-ABP), and AαC, N-(2′-deoxyguanosin-8-yl)-2-amino-9H-pydrido[2,3-b]indole (AαC), were also measured in WBC DNA of the same subjects, by our UPLC-IT/MS3 method.45,46
Optimization of Hb hydrolysis and SPE conditions for isolation of 4-ABP and AαC from blood
The Hb sulfinamide adduct of 4-ABP is quantitatively hydrolyzed in 0.1N NaOH or by 1% HCl in acetone at room temperature for 3 h.32,33 We examined the conditions of hydrolysis of Hb-AαC sulfinamide with AαC-modified Hb diluted to a level of 13 ng of AαC bound per mg Hb (vide supra). The hydrolysis was conducted for 1 or 3 h at 37 or 60 °C using either 0.1N HCl or 0.1N NaOH (Figure S1.) The hydrolysis of the AαC-Hb sulfinamide adduct was also facile, and the recovery of AαC was comparable under all hydrolysis conditions. The acidic hydrolysis for 3 h at 37 °C was employed for all ensuing analyses. During the course of the development of the method, we chose to derivatize 4-ABP and AαC with acetic anhydride. This N-acetylation step decreased the volatility of 4-ABP and minimized its loss during concentration of the solvents prior to LC-MS. Additionally, AαC displayed poor chromatographic properties with severe tailing that was ameliorated by the derivatization step. Moreover, the creation of the acetamide group permitted consecutive reaction monitoring at the MS3 scan stage, and greatly enhanced the selectivity and sensitivity of the assay for both compounds.
The efficiency of SPE resins to recover 4-ABP and AαC, and the purity of the Hb acid hydrolysis extracts were assessed by MS analysis. We had previously employed a mixed mode hydrophobic/ strong cation exchange (SOLA SCX) resin to successfully enrich AαC from human urine.58 However, a strong matrix effect was observed for the transitions of N-acetyl-4-ABP and N-acetyl-d5-4-ABP, and the signals were weak. In contrast, purification of the compounds with the HRP resin provided cleaner extracts and robust MS3 transition signals were observed for N-acetyl-4-ABP, N-acetyl-AαC and their internal standards (Figure S2). On average, the overall recovery of signals for both internal standards was ~50% of the signal of pure standards.
Performance of the analytical method to measure Hb sulfinamide adducts of 4-ABP and AαC
The product ion spectra of N-acetyl-4-ABP, N-acetyl-AαC and their internal standards are shown in Figure 1. N-Acetyl-4-ABP was monitored at the MS3 scan stage using the transitions [M+H]+ at m/z 212.1 > 170.1 > 143.1, 153.1, and 155.1. The fragment ion at m/z 170.1 at the MS2 scan stage is attributed to the loss of ketene, followed by losses of HCN, NH3 and presumably CH3• at the MS3 scan stage. N-Acetyl-d5-4-ABP was monitored at the MS3 scan stage [M+H]+ at m/z 217.1 > 175.1 > 148.1, 155.1, 156.1, 157.1, and 158.1. There was extensive H-D scrambling with prominent fragment ions attributed to the loss of ND3 (m/z 155.1), NHD2 (m/z 156.1), NH2D (m/z 157.1), in addition to the loss of NH3 (m/z 158.1). We had previously observed extensive H-D scrambling of d9-4-ABP under collision induced dissociation (CID) with a triple quadrupole mass spectrometer.59 Here, we observe that the H-D exchange still occurs by CID, when the penta-deuterium label occurs on the phenyl ring of 4-ABP and analyzed by ion trap MS. H-D scrambling of d5-biphenyl under electron impact ionization was previously reported by Cooks, where the deuterium atoms were randomized over both ring systems.60 N-Acetyl-AαC was monitored at the MS3 scan stage [M+H]+ at m/z 224.1 > 208.1 >167.1 and 185.1. The fragment ion at m/z 208.1 at the MS2 scan stage is attributed to loss of H2O, followed by loss of ethenimine (C2H3N) at the MS3 scan stage to form the deaminated charged AαC at m/z 167.1, and its water cluster adduct at m/z 185.1.61,62
Figure 1.

Product ion spectra of N-acetyl-4-ABP, N-acetyl-d5-4-ABP, N-acetyl-AαC, and N-acetyl[13C6]AαC at the MS3 scan stage.
The calibration curves were constructed with commercial blood containing low amounts of 4-ABP and AαC (below the LOQ values). The curves are shown in Figure S3. The slopes of the curves and the goodness-of-fit linear regression values are (y = 0.981x + 0.038, r2 = 0.9998) for 4-ABP and (y = 0.909x + 0.039, r2 = 0.9998) for AαC. The positive intercept for 4-ABP is attributed to trace levels of 4-ABP in the unspiked sample, whereas the positive intercept for AαC is largely attributed to the impurity of [13C6]AαC, which contained 3% unlabeled AαC. The limit of detection (LOD) and LOQ values were determined with blank samples containing d5-4-ABP and [13C6]AαC. The blank samples were processed with the same buffers, SPE resin, acid hydrolysis, and chemical derivatization conditions as used for the Hb samples. The recommended guidelines for data acquisition and quality evaluation in environmental chemistry were used as criteria, and the LOD and LOQ values for the 4-ABP and AαC were set at 3σ and 10σ SD units above the background level signals.63 The LOD and LOQ values were, respectively, 2.4 and 7.1 pg 4-ABP and 2.1 and 6.6 pg of AαC when normalized per g equivalent of Hb assayed. The amounts of 4-ABP and AαC present in the unspiked commercial blood sample were estimated at ≤ 2 pg of carcinogen per g Hb, when calculated by the linear regression curves. If the Y intercept values were omitted from the regression curves, the amounts of each carcinogen in blood were estimated at 4.8 ± 1.0 pg of 4-ABP per g of Hb and 6.2 ± 0.4 pg of AαC per g of Hb.
The accuracy and precision of the method were determined with four independent measurements conducted on three different days over a time period of one week. HbO2 was modified with HONH-4-ABP and HONH-AαC as described in Materials and Methods, and the adducted proteins were diluted with non-modified HbO2 to arrive at target levels of 42.0 pg 4-ABP/g Hb and 28.0 pg AαC per g Hb. The data are summarized in Table 1. The estimated levels of 4-ABP were within 4% of the target value, whereas the estimated levels of AαC were within 14% of the target value. The intra-day and inter-day % coefficient of variation (% CV) were, respectively, ≤17.0% and ≤22.4%, which are acceptable levels of precision for an ion trap MS instrument, particularly at these low levels of Hb modification.
Table 1.
Within-Day and Between-Day Estimates of 4-ABP-Hb and AαC-Hb Adducts in Human Blooda.
| Hb adduct | Targeted value | Mean (pg/g Hb) | Intra-day %CV | Inter-day %CV | LODb (pg/g Hb) | LOQb (pg/g Hb) |
|---|---|---|---|---|---|---|
| ABP-Hb | 42.0 | 43.8 | 9.9 | 13.4 | 2.4 | 7.1 |
| AαC-Hb | 28.0 | 24.2 | 17.0 | 22.4 | 2.1 | 6.6 |
Data were acquired on four independent measurements conducted on three different days.
The LOD and LOQ values were determined with the blank matrix sample work-up.
Hb-adducts of ABP and AαC in smokers and nonsmokers
The levels of 4-ABP- and AαC-Hb adducts were quantified in another batch of commercial blood obtained and measured at 22.9 ± 0.2 pg 4-ABP/g Hb and 4.3 ± 0.3 pg AαC/g Hb. The levels of 4-ABP- and AαC-Hb adducts were subsequently measured in three nonsmokers and seven current smokers. The Hb adduct levels, along with number of cigarettes smoked per day, pack years, and plasma cotinine levels are reported in Table 2. Representative reconstructed ion chromatograms of N-acetyl-4-ABP and N-acetyl-AαC obtained from blood samples of a nonsmoker and smoker are presented in Figure 2. The product ion spectra of the analytes at the MS3 scan stage confirm the structures of N-acetyl-4-ABP and N-acetyl-AαC.
Table 2.
Hb Adducts of 4-ABP, AαC, Cotinine Levels, and Smoking Status.
| ABP-Hb (pg/g Hb) | AαC-Hb (pg/g Hb) | Cotinine (ng/mL) | Cigarettes per day | Smoking years | |
|---|---|---|---|---|---|
|
| |||||
| Commercial Blood | 22.9 ± 0.2 | 4.3 ± 2.4 | n.aa | n.aa | n.aa |
| R01 | 29.8 ± 6.9 | 5.6 ± 0.8 | 0.13 | 0 | 0 |
| R03 | 23.8 ± 6.9 | 4.6 ± 0.6 | 0.13 | 0 | 0 |
| R05 | 21.8 ± 2.8 | 5.6 ± 1.1 | 0.13 | 0 | 0 |
| R02 | 189 ± 8.5 | 16.2 ± 1.9 | 164 | 30 | 42 |
| R04 | 71.4 ± 8.9 | 16.1 ± 1.0 | 224 | 20 | 25 |
| R06 | 125 ± 6.8 | 26.8 ± 2.5 | 228 | 21 | 33 |
| R07 | 134 ± 7.5 | 24.5 ± 0.8 | 210 | 30 | 32 |
| R08 | 71.3 ± 1.6 | 17.3 ± 0.7 | 86 | 25 | 30 |
| R09 | 162 ± 7.8 | 15.5 ± 0.9 | 108 | 70 | 53 |
| R10 | 90.4 ± 3.7 | 6.6 ± 0.6 | 55 | 25 | 30 |
n.a., not analyzed, mean ± SD (n = 3)
Figure 2.

The reconstructed ion chromatograms at the MS3 scan stage of acid-labile 4-ABP- and AαC-Hb adducts in (A) nonsmoker (R01) and (B) smoker (R07), following derivatization with acetic anhydride. (C) Product ion spectra of N-acetyl-4-ABP and N-acetyl-AαC and their internal standards obtained from the smoker’s blood sample. The * signifies that the ion at m/z 147.1 in the product ion spectrum of N-acetyl-d5-4-ABP contains an isobaric interferent.
The amounts of 4-ABP-Hb sulfinamide were about 4-fold above the LOQ value in nonsmokers, whereas the levels of AαC-Hb sulfinamide hovered close to the LOQ value. The mean level of the 4-ABP-Hb adduct in smokers (120 ± 45 pg/g Hb, N = 7, mean ± SD was 4.8-fold higher than in nonsmokers (25 ± 4.2 pg/g Hb, N = 3, mean ± SD), a magnitude consistent with values previously reported.25,27,28,33 The mean level of the AαC-Hb adduct was 3.4-fold higher in smokers (mean 18 ± 6.6 pg/g Hb, N = 7, mean ± SD) than nonsmokers (mean 5.3 pg/g Hb, N = 3, mean ± SD). However, the levels of 4-ABP-Hb adducts were higher than AαC-Hb levels in all blood samples. The mean level of the 4-ABP-Hb adduct was 7.4-fold higher than level of AαC-Hb adduct in smokers, when expressed as pmol of carcinogen bound per mg of Hb.
Stability of Hb-adducts of 4-ABP and AαC in vitro
The kinetics of removal of 4-ABP-Hb sulfinamide in humans was reported in a tobacco cessation study, and the adduct levels decreased to those levels seen in nonsmokers 6 to 8 weeks after the cessation of tobacco usage.33 The kinetics data signify that the adduct is removed more rapidly than the life span of Hb, which is about 4 months.24 To our knowledge, the chemical stability of the 4-ABP-Hb sulfinamide has not been reported in vitro. We examined the chemical stabilities of the Hb sulfinamides of 4-ABP and AαC in vitro to determine if a more rapid rate of hydrolysis of the Hb sulfinamide of AαC may explain the large differences observed between the levels of 4-ABP- and AαC-Hb adducts formed in vivo. The Hb modified with HONH-ABP and HONH-AαC in vitro was diluted to 30 mg of unmodified Hb in 100 mM sodium phosphate buffer (500 μL, pH 7.4, containing 0.9% NaCl) to a final level of 4-ABP-Hb sulfinamide of 165 pg/g Hb and AαC-Hb sulfinamide at 85 pg/g Hb. The samples were incubated at 37 °C for different times (0, 8, 24, 48, 72, 96 h). The rates of decay of both adducts are shown in Figure 3. Linear regression of the slopes of decay show that the slopes are not significantly different (P = 0.1741). Thus, we conclude that the rates of hydrolysis of the 4-ABP- and AαC-Hb sulfinamide linkages are comparable. However, the rate of hydrolysis of the 4-ABP-Hb sulfinamide in vitro is considerable more rapid that that reported in vivo.33 We observed that the amounts of met-Hb increased by 13-fold during the 96 h kinetics study. The reactive oxygen species generated as a by-product of met-Hb formation may have oxidized a portion of the Hb sulfinamide linkages to the acid-stable sulfonamide linked adducts and explain some of the rapid loss of the Hb sulfinamide adducts in vitro over time.51,64
Figure 3.

Stability of AαC-Hb and 4-ABP-Hb sulfinamide adducts and formation of met-Hb over time at 37 °C in phosphate buffer (pH 7.4 with 0.9% NaCl). The 95% confidence interval of the slopes of the regression curves are shown as dotted lines.
WBC DNA adducts of ABP and AαC in smokers and nonsmokers
The levels dG-C8-AαC and dG-C8-4-ABP, the major adducts formed with AαC and 4-ABP,5,21,65 were also measured in WBC of the same subjects. Both dG-C8-AαC and dG-C8-4-ABP were below the LOQ value in all samples (~ 3 adducts per 109 DNA bases with 4 μg assayed on column).19,20 The effect of the WBC matrix on DNA adduct measurements was assessed by spiking 10 μg of modified CT DNA containing dG-C8-4-ABP at a level of 1 adduct per 108 bases into the DNA samples. The dilution with WBC DNA decreased the level of dG-C8-4-ABP to levels of 2 – 5 adducts per 109 bases. The recovery average of the spiked CT DNA containing dG-C8-4-ABP in the 10 WBC samples prior to processing of DNA was 84.9 ± 7.0 (Mean ± SD) proving that the WBC matrix had a negligible effect on the recovery and the response of dG-C8-4-ABP adduct. DNA containing a known, verified level dG-C8-AαC was not available for study. However, we had previously shown that dG-C8-AαC was recovered from human hepatocytes treated with AαC in a dose-dependent manner down to a concentration as low as 1 nM of AαC, a dosage where dG-C8-AαC adduct formation was measured at 3 adducts per 109 DNA bases.19 This successful measurement demonstrated the high efficiency in enzymatic digestion and recovery of the dG-C8-AαC DNA adduct.19 Moreover, we pooled WBC DNA from three nonsmokers and three smokers for a final amount of 50 μg DNA for each specimen. The spiking dG-C8-AαC and dG-C8-4-ABP at a level of 1 adduct per 108 DNA bases prior to digestion to DNA. Adducts were not detected in the unspiked sample, but both adducts are clearly visible in the reconstructed ion chromatograms with 20 μg DNA assayed on column (Figure S4). The levels of adducts are 10-fold or greater above the background level. A representative UPLC-ESI/MS3 reconstructed ion chromatograms are shown in Figure 4.
Figure 4.

(A) The reconstructed ion chromatograms at the MS3 scan stage of dG-C8-ABP and dG-C8-AαC in nonspiked and spiked WBC DNA of a nonsmoker (R01) and smoker (R07). The level of spiking of CT DNA containing known levels of dG-C8-4-ABP in the WBC was 4 adducts per 109 bases in the nonsmoker and 3 adducts per 109 bases in the smoker’s DNA. Peaks were integrated at or near the tR for dG-C8-4-ABP and dG-C8-AαC to show the background signals were negligible.
Discussion
We have established a robust nanoLC-IT/MS3 method to measure Hb sulfinamide adducts formed with 4-ABP and AαC. The chemical derivatization step with acetic anhydride improved the chromatography of the analytes, and the introduction of the acetamide group permitted consecutive reaction monitoring at the MS3 scan stage, which greatly improved the sensitivity of the assay. Moreover, the rapid scanning features of the linear ion trap permitted the acquisition of the MS3 scan stage product ion spectra and provided confirmatory data on the identities of the analytes. The LOQ values for both 4-ABP and AαC are comparable to those values reported for 4-ABP using GC-NICI/MS,27,33,35 or GC/MS with electron impact in the positive-ion mode using multiple-reaction-monitoring.28 However, our nanoLC-IT/MS3 method requires only 30 mg of HbO2, or approximately 0.2 mL of blood, compared to several mL of blood required for the GC/MS based assays. Our analysis focused on 4-ABP and AαC, but the method should be amenable to measure Hb adducts formed with other aromatic amines or HAAs found in tobacco smoke or the environment.25,33
Even though AαC is present in mainstream tobacco smoke at 25–100-fold higher amounts than 4-ABP,6–9 the mean level of the 4-ABP-Hb adduct is 7.4-fold greater than the mean level of the AαC-Hb adduct in blood of smokers. The formation of Hb-arylsulfinamide adduct is thought to occur by N-hydroxylation of the aromatic amine, by hepatic cytochrome P450, followed by transport of the arylhydroxylamine to the blood, and uptake by the erythrocytes to form Hb arylsulfinamide adduct (Scheme 1).31–33 The higher level of 4-ABP-Hb adduct formation is not attributed to the superior reactivity of the HONH-4-ABP with HbO2 since HNOH-AαC induces more methemoglobinemia in erythrocytes and forms higher levels of the Hb-sulfinamide than does HONH-4-ABP.51 Higher rates of N-oxidation of 4-ABP also do not appear to explain the elevated levels of 4-ABP-Hb formation. The rates of N-oxidation of 4-ABP and AαC are similar with human liver microsomes,29,55 and AαC and 4-ABP form comparable levels of DNA adducts in primary human hepatocytes.19,20 These results signify that both AαC and 4-ABP are efficiently N-hydroxylated in human liver. AαC undergoes extensive metabolism by primary human hepatocytes to produce multiple phase I and phase II metabolites,66 whereas the pathways of metabolism of 4-ABP in human hepatocytes are not well characterized.20 On the basis of our understanding of 4-ABP metabolism in rodent and canine models,32,67–69 we surmise that the amount of HONH-4-ABP circulating in blood and delivered to the erythrocyte is far greater than that of HONH-AαC, resulting in proportionately higher levels of 4-ABP-Hb adducts.25,33 The N-hydroxylated metabolites of 4-ABP and AαC form adducts with human or rat serum albumin64,70–72 In the case of 4-ABP, the binding of its N-oxidized metabolite is 250 fold-greater for Hb than for albumin in the rat model;70,73 however, the comparative levels of HONH-AαC binding to Hb and albumin have not been measured in vivo in rodents or humans. The covalent (or non-covalent) binding of HONH-AαC to drug binding sites of albumin may be favored because of the planar nature of its fused 3-ring system, as opposed to the non-planar 4-ABP, and impact the relative levels of adduct formation between these two blood proteins.74,75 Further studies on the relative covalent binding of N-oxidized AαC to Hb and albumin are warranted.
We examined the capacity of 4-ABP and AαC to form DNA adducts in peripheral WBC. The LOQ values for both dG-C8-4-ABP and dG-C8-AαC, by our UPLC-IT/MS3 method, are ~ 3 adducts per 109 DNA bases, with 4 μg of DNA assayed on column.19,20 This level of sensitivity rivals that of the highly sensitive 32P-postlabeling method.49 Even though volunteers were heavy smokers for 25 years or more, we were unable to detect WBC DNA adducts for either carcinogen. The inability to detect DNA adducts may be attributed to the short life-span of some WBC, including monocytes (5–10% of total WBC), and granulocytes (40–75%), with half-lives of hours to days, and thus, DNA adducts are rapidly removed due to cell turnover.37 We previously reported that DNA adducts of AαC are formed at higher levels than the adducts formed with 4-ABP, PhIP and MeIQx in human T lymphocytes in vitro; however, the DNA adducts are only detected in lymphocytes pre-activated with PMA/Ionomycin or CD3/CD28, which induce the expression of functional CYP1 activity for bioactivation.76 Therefore, the capacity of cytochrome P450 enzymes expressed in peripheral WBC may be insufficient to N-hydroxylate 4-ABP and AαC. Alternatively, the phase II enzymes such as N-acetyltransferases (NAT) and sulfotransferases (SULT), which bioactivate HONH-4-ABP and HONH-AαC to the penultimate species that form DNA adducts,48,77 may be expressed at insufficient levels in WBC to catalyze DNA binding of the N-hydroxylated metabolites. The data reported in the literature on the expression of Phase II enzymes in human WBC are restricted to few reports on the detection of SULT and UDP-glucuronosyltransferase activities in human lymphocytes.78,79 However, to the best of knowledge, there is no report on the measurement of NAT activities in human WBC. The failure to detect DNA adducts or 4-ABP and AαC in WBC also could be due, in part, to the large difference in the amount of Hb compared to WBC DNA. There is approximately 150 mg Hb/mL blood,24 an amount that corresponds to ~9.30 mM of Hb subunit or ~4.65 mM of Hb β-Cys93 residues 80 For comparison, the amount of WBC DNA is ~6 μg/mL blood. Assuming an average molecular weight of 330 Da for a deoxynucleotide, the concentration of WBC deoxynucleoside is 18.2 μM, and the concentration of dG is about 4.55 μM. This molar value of dG is about 1,000-fold lower than that of the Hb β-Cys93 residue in blood. The vast excess of nucleophilic β-Cys93 in blood may help to explain the success in detection of Hb adducts over DNA adducts formed with WBC. In summary, both 4-ABP and AαC undergo N-oxidation and bioactivation in humans to form sulfinamide adducts with Hb, but neither compound binds to DNA in WBC at quantifiable levels. The striking differences in the yield of Hb adducts highlight the important differences in the metabolic processing of these structurally related carcinogens, which impact the levels of Hb adduct formation and possibly differences in DNA adduct formation in target organs of tobacco-associated cancers.
Supplementary Material
Acknowledgments
Funding
This work was supported by NIH grants R01CA134700 and R01CA134700-03S1 (to R.J.T.) of the Family Smoking Prevention and Tobacco Control Act, and Cancer Center Support grant no. CA-77598 (to R.J.T.).
Abbreviations
- 4-ABP
4-aminobiphenyl
- AαC
2-amino-9H-pyrido[2,3-b]indole
- B[a]P
benzo[a]pyrene
- ESI
electrospray ionization
- GC-MS
gas chromatography mass spectrometry
- GC-NICI/MS
gas chromatography-negative ion chemical ionization mass spectrometry
- HAA
heterocyclic aromatic amines
- Hb
hemoglobin
- met-Hb
methemoglobin
- HbO2
oxy-hemoglobin
- nanoLC-IT/MS3
nanoflow liquid chromatography/ion trap multistage mass spectrometry
- NNK
4-(methylnitrosamino)-1-(3-pyridyl)-1-butanone
- PAH
polycyclic aromatic hydrocarbons
- UPLC-IT/MS3
ultraperformance liquid chromatography-ion trap multistage mass spectrometry
- WBC
white blood cells
Footnotes
Notes
The authors declare no competing financial interest.
The Supporting Information is available free of charge on the ACS Publications website at DOI:…… at http://pubs.acs.org
Optimization of Hb hydrolysis conditions of AαC-Hb; effect of SPE resins and conditions on the analysis of N-actetyl-4-ABP and N-acetyl-AαC derivatives in blood sample workups; calibration curves of 4-ABP and AαC generated in 30 mg of Hb, following chemical derivatization with acetic anhydride; reconstructed ion chromatograms at the MS3 scan stage of dG-C8-ABP and dG-C8-AαC in non-spiked and spiked pooled WBC DNA from three nonsmokers and smokers with 20 μg DNA injected on column and MS3 scan stage product ion spectra.
References
- 1.Hecht SS. Tobacco carcinogens, their biomarkers and tobacco-induced cancer. Nat Rev Cancer. 2003;3:733–744. doi: 10.1038/nrc1190. [DOI] [PubMed] [Google Scholar]
- 2.Hoffmann D. Letters to the editor: Tobacco smoke components. Beiträge zur Tabakforschung Int. 1998;18:49–52. [Google Scholar]
- 3.International Agency for Research on Cancer. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans: Tobacco smoking. Vol. 38. International Agency for Research on Cancer; Lyon, France: 1986. [Google Scholar]
- 4.International Agency for Research on Cancer. Personal habits and indoor combustions. 100E. Lyon, France: 2012. IARC Monographs on the Evaluation of Carcinogenic Risks to Humans. [PMC free article] [PubMed] [Google Scholar]
- 5.Beland FA, Kadlubar FF. Formation and persistence of arylamine DNA adducts in vivo. Environ Health Perspect. 1985;62:19–30. doi: 10.1289/ehp.856219. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Yoshida Daisuke, Matsumoto Takashi. Amino-α-carbolines as mutagenic agents in cigarette smoke condensate. Cancer Letters. 1980;10:141–149. doi: 10.1016/0304-3835(80)90037-3. [DOI] [PubMed] [Google Scholar]
- 7.Zhang L, Ashley DL, Watson CH. Quantitative analysis of six heterocyclic aromatic amines in mainstream cigarette smoke condensate using isotope dilution liquid chromatography-electrospray ionization tandem mass spectrometry. Nicotine Tob Res. 2011;13:120–126. doi: 10.1093/ntr/ntq219. [DOI] [PubMed] [Google Scholar]
- 8.Patrianakos Constantin, Hoffmann Dietrich. Chemical Studies on Tobacco Smoke LXIV. On the Analysis of Aromatic Amines in Cigarette Smoke. J Anal Toxicol. 1979;3:150–154. [Google Scholar]
- 9.Zha Q, Qian NX, Moldoveanu SC. Analysis of polycyclic aromatic hydrocarbons in the particulate phase of cigarette smoke using a gas chromatographic-high-resolution mass spectrometric technique. J Chromatogr Sci. 2002;40:403–408. doi: 10.1093/chromsci/40.7.403. [DOI] [PubMed] [Google Scholar]
- 10.Hecht SS. Research opportunities related to establishing standards for tobacco products under the Family Smoking Prevention and Tobacco Control Act. Nicotine Tob Res. 2012;14:18–28. doi: 10.1093/ntr/ntq216. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Okonogi H, Ushijima T, Shimizu H, Sugimura T, Nagao M. Induction of aberrant crypt foci in C57BL/6N mice by 2-amino-9H-pyrido[2,3-b]indole (AalphaC) and 2-amino-3,8-dimethylimidazo[4,5-f]quinoxaline (MeIQx) Cancer Lett. 1997;111:105–109. doi: 10.1016/s0304-3835(96)04505-3. [DOI] [PubMed] [Google Scholar]
- 12.Okonogi H, Ushijima T, Zhang XB, Heddle JA, Suzuki T, Sofuni T, Felton JS, Tucker JD, Sugimura T, Nagao M. Agreement of mutational characteristics of heterocyclic amines in lacI of the Big Blue mouse with those in tumor related genes in rodents. Carcinogenesis. 1997;18:745–748. doi: 10.1093/carcin/18.4.745. [DOI] [PubMed] [Google Scholar]
- 13.Kim S, Guo J, O’Sullivan MG, Gallaher DD, Turesky RJ. Comparative DNA adduct formation and induction of colonic aberrant crypt foci in mice exposed to 2-amino-9H-pyrido[2,3-b]indole, 2-amino-3,4-dimethylimidazo[4,5-f]quinoline, and azoxymethane. Environ Mol Mutagen. 2016;57:125–136. doi: 10.1002/em.21993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Sugimura T, Wakabayashi K, Nakagama H, Nagao M. Heterocyclic amines: Mutagens/carcinogens produced during cooking of meat and fish. Cancer Sci. 2004;95:290–299. doi: 10.1111/j.1349-7006.2004.tb03205.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Turesky RJ, Konorev D, Fan X, Tang Y, Yao L, Ding X, Xie F, Zhu Y, Zhang QY. Effect of Cytochrome P450 Reductase Deficiency on 2-Amino-9H-pyrido[2,3-b]indole Metabolism and DNA Adduct Formation in Liver and Extrahepatic Tissues of Mice. Chem Res Toxicol. 2015;28:2400–2410. doi: 10.1021/acs.chemrestox.5b00405. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Tang Y, Kassie F, Qian X, Ansha B, Turesky RJ. DNA adduct formation of 2-amino-9H-pyrido[2,3-b]indole and 2-amino-3,4-dimethylimidazo[4,5-f]quinoline in mouse liver and extrahepatic tissues during a subchronic feeding study. Toxicol Sci. 2013;133:248–258. doi: 10.1093/toxsci/kft077. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Majer BJ, Kassie F, Sasaki Y, Pfau W, Glatt H, Meinl W, Darroudi F, Knasmuller S. Investigation of the genotoxic effects of 2-amino-9H-pyrido[2,3-b]indole in different organs of rodents and in human derived cells. J Chromatogr B Analyt Technol Biomed Life Sci. 2004;802:167–173. doi: 10.1016/j.jchromb.2003.10.042. [DOI] [PubMed] [Google Scholar]
- 18.Pfau W, Martin FL, Cole KJ, Venitt S, Phillips DH, Grover PL, Marquardt H. Heterocyclic aromatic amines induce DNA strand breaks and cell transformation. Carcinogenesis. 1999;20:545–551. doi: 10.1093/carcin/20.4.545. [DOI] [PubMed] [Google Scholar]
- 19.Nauwelaers G, Bellamri M, Fessard V, Turesky RJ, Langouet S. DNA adducts of the tobacco carcinogens 2-amino-9H-pyrido[2,3-b]indole and 4-aminobiphenyl are formed at environmental exposure levels and persist in human hepatocytes. Chem Res Toxicol. 2013;26:1367–1377. doi: 10.1021/tx4002226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Nauwelaers G, Bessette EE, Gu D, Tang Y, Rageul J, Fessard V, Yuan JM, Yu MC, Langouet S, Turesky RJ. DNA adduct formation of 4-aminobiphenyl and heterocyclic aromatic amines in human hepatocytes. Chem Res Toxicol. 2011;24:913–925. doi: 10.1021/tx200091y. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Turesky RJ, Bendaly J, Yasa I, Doll MA, Hein DW. The impact of NAT2 acetylator genotype on mutagenesis and DNA adducts from 2-amino-9H-pyrido[2,3-b]indole. Chem Res Toxicol. 2009;22:726–733. doi: 10.1021/tx800473w. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Skipper PL, Tannenbaum SR. Protein adducts in the molecular dosimetry of chemical carcinogens. Carcinogenesis. 1990;11:507–518. doi: 10.1093/carcin/11.4.507. [DOI] [PubMed] [Google Scholar]
- 23.Hecht SS. Human urinary carcinogen metabolites: biomarkers for investigating tobacco and cancer. Carcinogenesis. 2002;23:907–922. doi: 10.1093/carcin/23.6.907. [DOI] [PubMed] [Google Scholar]
- 24.Tornqvist M, Fred C, Haglund J, Helleberg H, Paulsson B, Rydberg P. Protein adducts: quantitative and qualitative aspects of their formation, analysis and applications. J Chromatogr B Analyt Technol Biomed Life Sci. 2002;778:279–308. doi: 10.1016/s1570-0232(02)00172-1. [DOI] [PubMed] [Google Scholar]
- 25.Bryant MS, Vineis P, Skipper PL, Tannenbaum SR. Hemoglobin adducts of aromatic amines: associations with smoking status and type of tobacco. Proc Natl Acad Sci USA. 1988;85:9788–9791. doi: 10.1073/pnas.85.24.9788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Ward EM, Sabbioni G, DeBord DG, Teass AW, Brown KK, Talaska GG, Roberts DR, Ruder AM, Streicher RP. Monitoring of aromatic amine exposures in workers at a chemical plant with a known bladder cancer excess. J Natl Cancer Inst. 1996;88:1046–1052. doi: 10.1093/jnci/88.15.1046. [DOI] [PubMed] [Google Scholar]
- 27.Myers Steven R. Characterization of 4-aminobiphenyl-hemoglobin adducts in maternal and fetal blood samples. Jf Toxicol and Environ Health. 1996;47:553–566. doi: 10.1080/009841096161537. [DOI] [PubMed] [Google Scholar]
- 28.Seyler Tiffany H, Levi-Rose Reyes, Bernert John T. Analysis of 4-Aminobiphenyl Hemoglobin Adducts in Smokers and Nonsmokers by Pseudo Capillary On-Column Gas Chromatography-Tandem Mass Spectrometry. J Anal Toxicol. 2010;34:304–311. doi: 10.1093/jat/34.6.304. [DOI] [PubMed] [Google Scholar]
- 29.Butler MA, Iwasaki M, Guengerich FP, Kadlubar FF. Human cytochrome P-450 PA (P450IA2), the phenacetin O-deethylase, is primarily responsible for the hepatic 3-demethylation of caffeine and N-oxidation of carcinogenic arylamines. Proc Natl Acad Sci USA. 1989;86:7696–7700. doi: 10.1073/pnas.86.20.7696. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Kiese M. The biochemical production of ferrihemoglobin-forming derivatives from aromatic amines, and mechanisms of ferrihemoglobin formation. Pharmacol Rev. 1966;18:1091–1161. [PubMed] [Google Scholar]
- 31.Kiese M, Taeger K. The fate of phenylhydroxylamine in human red cells. Naunyn Schmiedebergs Arch Pharmacol. 1976;292:59–66. doi: 10.1007/BF00506490. [DOI] [PubMed] [Google Scholar]
- 32.Green LC, Skipper PL, Turesky RJ, Bryant MS, Tannenbaum SR. In vivo dosimetry of 4-aminobiphenyl in rats via a cysteine adduct in hemoglobin. Cancer Res. 1984;44:4254–4259. [PubMed] [Google Scholar]
- 33.Bryant MS, Skipper PL, Tannenbaum SR, Maclure M. Hemoglobin adducts of 4-aminobiphenyl in smokers and nonsmokers. Cancer Res. 1987;47:602–608. [PubMed] [Google Scholar]
- 34.Ringe D, Turesky RJ, Skipper PL, Tannenbaum SR. Structure of the single stable hemoglobin adduct formed by 4-aminobiphenyl in vivo. Chem Res Toxicol. 1988;1:22–24. doi: 10.1021/tx00001a003. [DOI] [PubMed] [Google Scholar]
- 35.Stillwell WG, Bryant MS, Wishnok JS. GC/MS analysis of biologically important aromatic amines. Application to human dosimetry. Biomed Environ Mass Spectrom. 1987;14:221–227. doi: 10.1002/bms.1200140505. [DOI] [PubMed] [Google Scholar]
- 36.Randerath K, Reddy MV, Gupta RC. 32P-labeling test for DNA damage. Proc Natl Acad Sci USA. 1981;78:6162–6129. doi: 10.1073/pnas.78.10.6126. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Godschalk RW, Van Schooten FJ, Bartsch H. A critical evaluation of DNA adducts as biological markers for human exposure to polycyclic aromatic compounds. J Biochem Mol Biol. 2003;36:1–11. doi: 10.5483/bmbrep.2003.36.1.001. [DOI] [PubMed] [Google Scholar]
- 38.Peluso M, Airoldi L, Magagnotti C, Fiorini L, Munnia A, Hautefeuille A, Malaveille C, Vineis P. White blood cell DNA adducts and fruit and vegetable consumption in bladder cancer. Carcinogenesis. 2000;21:183–187. doi: 10.1093/carcin/21.2.183. [DOI] [PubMed] [Google Scholar]
- 39.Ricceri F, Godschalk RW, Peluso M, Phillips DH, Agudo A, Georgiadis P, Loft S, Tjonneland A, Raaschou-Nielsen O, Palli D, Perera F, Vermeulen R, Taioli E, Sram RJ, Munnia A, Rosa F, Allione A, Matullo G, Vineis P. Bulky DNA adducts in white blood cells: a pooled analysis of 3,600 subjects. Cancer Epidemiol Biomarkers Prev. 2010;19:3174–3181. doi: 10.1158/1055-9965.EPI-10-0314. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Tang D, Phillips DH, Stampfer M, Mooney LA, Hsu Y, Cho S, Tsai WY, Ma J, Cole KJ, She MN, Perera FP. Association between carcinogen-DNA adducts in white blood cells and lung cancer risk in the physicians health study. Cancer Res. 2001;61:6708–6712. [PubMed] [Google Scholar]
- 41.Mooney LA, Santella RM, Covey L, Jeffrey AM, Bigbee W, Randall MC, Cooper TB, Ottman R, Tsai WY, Wazneh L, et al. Decline of DNA damage and other biomarkers in peripheral blood following smoking cessation. Cancer Epidemiol Biomarkers Prev. 1995;4:627–634. [PubMed] [Google Scholar]
- 42.Phillips DH, Venitt S. DNA and protein adducts in human tissues resulting from exposure to tobacco smoke. Int J Cancer. 2012;131:2733–2753. doi: 10.1002/ijc.27827. [DOI] [PubMed] [Google Scholar]
- 43.Peluso M, Airoldi L, Munnia A, Colombi A, Veglia F, Autrup H, Dunning A, Garte S, Gormally E, Malaveille C, Matullo G, Overvad K, Raaschou-Nielsen O, Clavel-Chapelon F, Linseisen J, Boeing H, Trichopoulou A, Palli D, Krogh V, Tumino R, Panico S, Bueno-De-Mesquita BH, Peeters PH, Kumle M, Agudo A, Martinez C, Dorronsoro M, Barricarte A, Tormo MJ, Quiros JR, Berglund G, Jarvholm B, Day NE, Key TJ, Saracci R, Kaaks R, Riboli E, Bingham S, Vineis P. Bulky DNA adducts, 4-aminobiphenyl-haemoglobin adducts and diet in the European Prospective Investigation into Cancer and Nutrition (EPIC) prospective study. Br J Nutr. 2008;100:489–495. doi: 10.1017/S0007114508911600. [DOI] [PubMed] [Google Scholar]
- 44.Dallinga JW, Pachen DM, Wijnhoven SW, Breedijk A, van ‘t Veer L, Wigbout G, van Zandwijk N, Maas LM, van Agen E, Kleinjans JC, van Schooten FJ. The use of 4-aminobiphenyl hemoglobin adducts and aromatic DNA adducts in lymphocytes of smokers as biomarkers of exposure. Cancer Epidemiol Biomarkers Prev. 1998;7:571–577. [PubMed] [Google Scholar]
- 45.Guo J, Yun BH, Upadhyaya P, Yao L, Krishnamachari S, Rosenquist TA, Grollman AP, Turesky RJ. Multiclass Carcinogenic DNA Adduct Quantification in Formalin-Fixed Paraffin-Embedded Tissues by Ultraperformance Liquid Chromatography-Tandem Mass Spectrometry. Anal Chem. 2016;88:4780–4787. doi: 10.1021/acs.analchem.6b00124. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Goodenough AK, Schut HA, Turesky RJ. Novel LC-ESI/MS/MS n method for the characterization and quantification of 2′-deoxyguanosine adducts of the dietary carcinogen 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine by 2-D linear quadrupole ion trap mass spectrometry. Chem Res Toxicol. 2007;20:263–276. doi: 10.1021/tx0601713. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Westra JG. A rapid and simple synthesis of reactive metabolites of carcinogenic aromatic amines in high yield. Carcinogenesis. 1981;2:355–357. doi: 10.1093/carcin/2.4.355. [DOI] [PubMed] [Google Scholar]
- 48.King RS, Teitel CH, Kadlubar FF. In vitro bioactivation of N-hydroxy-2-amino-alpha-carboline. Carcinogenesis. 2000;21:1347–1354. [PubMed] [Google Scholar]
- 49.Beland FA, Doerge DR, Churchwell MI, Poirier MC, Schoket B, Marques MM. Synthesis, characterization, and quantitation of a 4-aminobiphenyl-DNA adduct standard. Chem Res Toxicol. 1999;12:68–77. doi: 10.1021/tx980172y. [DOI] [PubMed] [Google Scholar]
- 50.Zijlstra WG, Buursma A, Meeuwsen-van der Roest WP. Absorption spectra of human fetal and adult oxyhemoglobin, de-oxyhemoglobin, carboxyhemoglobin, and methemoglobin. Clin Chem. 1991;37:1633–1638. [PubMed] [Google Scholar]
- 51.Pathak KV, Chiu TL, Amin EA, Turesky RJ. Methemoglobin Formation and Characterization of Hemoglobin Adducts of Carcinogenic Aromatic Amines and Heterocyclic Aromatic Amines. Chem Res Toxicol. 2016;29:255–269. doi: 10.1021/acs.chemrestox.5b00418. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Murphy SE, Wickham KM, Lindgren BR, Spector LG, Joseph A. Cotinine and trans 3′-hydroxycotinine in dried blood spots as biomarkers of tobacco exposure and nicotine metabolism. J Expo Sci Environ Epidemiol. 2013;23:513–518. doi: 10.1038/jes.2013.7. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Rich KJ, Murray BP, Lewis I, Rendell NB, Davies DS, Gooderham NJ, Boobis AR. N-Hydroxy-MeIQx is the major microsomal oxidation product of the dietary carcinogen MeIQx with human liver. Carcinogenesis. 1992;13:2221–2226. doi: 10.1093/carcin/13.12.2221. [DOI] [PubMed] [Google Scholar]
- 54.Zhao K, Murray S, Davies DS, Boobis AR, Gooderham NJ. Metabolism of the food derived mutagen and carcinogen 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP) by human liver microsomes. Carcinogenesis. 1994;15:1285–1288. doi: 10.1093/carcin/15.6.1285. [DOI] [PubMed] [Google Scholar]
- 55.Raza H, King RS, Squires RB, Guengerich FP, Miller DW, Freeman JP, Lang NP, Kadlubar FF. Metabolism of 2-amino-alpha-carboline. A food-borne heterocyclic amine mutagen and carcinogen by human and rodent liver microsomes and by human cytochrome P4501A2. Drug Metab Dispos. 1996;24:395–400. [PubMed] [Google Scholar]
- 56.Felton JS, Jagerstad M, Knize MG, Skog K, Wakabayashi K. Contents in foods, beverages and tobacco. In: Nagao M, Sugimura T, editors. Food Borne Carcinogens Heterocyclic Amines. John Wiley & Sons Ltd; Chichester, England: 2000. pp. 31–71. [Google Scholar]
- 57.Bellamri M, Le Hegarat L, Vernhet L, Baffet G, Turesky RJ, Langouët S. Human T lymphocytes bioactivate heterocyclic aromatic amines by forming DNA adducts. Environ Mol Mutagen. 2016;57:656–667. doi: 10.1002/em.22059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Konorev D, Koopmeiners JS, Tang Y, Franck Thompson EA, Jensen JA, Hatsukami DK, Turesky RJ. Measurement of the Heterocyclic Amines 2-Amino-9H-pyrido[2,3-b]indole and 2-Amino-1-methyl-6-phenylimidazo[4,5-b]pyridine in Urine: Effects of Cigarette Smoking. Chem Res Toxicol. 2015;28:2390–2399. doi: 10.1021/acs.chemrestox.5b00401. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Turesky RJ, Freeman JP, Holland RD, Nestorick DM, Miller DW, Ratnasinghe DL, Kadlubar FF. Identification of aminobiphenyl derivatives in commercial hair dyes. Chem Res Toxicol. 2003;16:1162–1173. doi: 10.1021/tx030029r. [DOI] [PubMed] [Google Scholar]
- 60.Cooks R Graham, Howe Ian, Tam SW, Williams Dudley H. Studies in mass spectrometry. XXIX. Hydrogen scrambling in some bicyclic aromatic systems. Randomization over two rings. J American Chemical Society. 1968;90:4064–4069. [Google Scholar]
- 61.Holland RD, Taylor J, Schoenbachler L, Jones RC, Freeman JP, Miller DW, Lake BG, Gooderham NJ, Turesky RJ. Rapid biomonitoring of heterocyclic aromatic amines in human urine by tandem solvent solid phase extraction liquid chromatography electrospray ionization mass spectrometry. Chem Res Toxicol. 2004;17:1121–1136. doi: 10.1021/tx049910a. [DOI] [PubMed] [Google Scholar]
- 62.Toribio F, Moyano E, Puignou L, Galceran MT. Multistep mass spectrometry of heterocyclic amines in a quadrupole ion trap mass analyser. J Mass Spectrom. 2002;37:812–828. doi: 10.1002/jms.340. [DOI] [PubMed] [Google Scholar]
- 63.MacDougall D, Amore FJ, Cox GV, Crosby DG, Estes FL, Freeman DH, Gibbs WE, Gordon GE, Keith LH, Lal J, Langner RR, McClelland NI, Phillips WF, Pojasek RB, Sievers RE. Guidelines for data acquistion and data quality evaluation in environmental chemistry. Anal Chem. 1980;52:2242–2249. [Google Scholar]
- 64.Peng L, Turesky RJ. Capturing labile sulfenamide and sulfinamide serum albumin adducts of carcinogenic arylamines by chemical oxidation. Anal Chem. 2013;85:1065–1072. doi: 10.1021/ac3028273. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 65.Pfau W, Schulze C, Shirai T, Hasegawa R, Brockstedt U. Identification of the major hepatic DNA adduct formed by the food mutagen 2-amino-9H-pyrido[2,3-b]indole (AaC) Chem Res Toxicol. 1997;10:1192–1197. doi: 10.1021/tx9701182. [DOI] [PubMed] [Google Scholar]
- 66.Bellamri M, Le Hegarat L, Turesky RJ, Langouet S. Metabolism of the Tobacco Carcinogen 2-Amino-9H-pyrido[2,3-b]indole (AalphaC) in Primary Human Hepatocytes. Chemical research in toxicology. 2017;30:657–668. doi: 10.1021/acs.chemrestox.6b00394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Radomski JL, Brill E. Bladder cancer induction by aromatic amines: role of N-hydroxy metabolites. Science. 1970;167:992–993. doi: 10.1126/science.167.3920.992. [DOI] [PubMed] [Google Scholar]
- 68.Heilmair R, Karreth S, Lenk W. The metabolism of 4-aminobiphenyl in rat. II. Reaction of N-hydroxy-4-aminobiphenyl with rat blood in vitro. Xenobiotica. 1991;21:805–815. doi: 10.3109/00498259109039519. [DOI] [PubMed] [Google Scholar]
- 69.Kadlubar FF, Beland FA. Chemical properties of ultimate carcinogenic metabolites of arylamines and arylamides. In: Harvey RG, editor. Polycyclic Hydrocarbons and Carcinogenesis. American Chemical Society; Washington, D.C: 1985. pp. 332–370. [Google Scholar]
- 70.Skipper PL, Obiedzinski MW, Tannenbaum SR, Miller DW, Mitchum RK, Kadlubar FF. Identification of the major serum albumin adduct formed by 4-aminobiphenyl in vivo in rats. Cancer Res. 1985;45:5122–5127. [PubMed] [Google Scholar]
- 71.Tannenbaum SR, Skipper PL, Wishnok JS, Stillwell WG, Day BW, Taghizadeh K. Characterization of various classes of protein adducts. Environ Health Perspect. 1993;99:51–55. doi: 10.1289/ehp.939951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Pathak KV, Bellamri M, Wang Y, Langouet S, Turesky RJ. 2-Amino-9H-pyrido[2,3-b]indole (AalphaC) Adducts and Thiol Oxidation of Serum Albumin as Potential Biomarkers of Tobacco Smoke. J Bio Chem. 2015;290:16304–16318. doi: 10.1074/jbc.M115.646539. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Skipper PL, Green LC, Bryant MS, Tannenbaum SR, Kadlubar FF. Monitoring exposure to 4-aminobiphenyl via blood protein adducts. IARC Sci Publ. 1984:143–150. [PubMed] [Google Scholar]
- 74.Skipper PL. Influence of tertiary structure on nucleophilic substitution reactions of proteins. Chem Res Toxicol. 1996;9:918–923. doi: 10.1021/tx960028h. [DOI] [PubMed] [Google Scholar]
- 75.Ghuman J, Zunszain PA, Petitpas I, Bhattacharya AA, Otagiri M, Curry S. Structural basis of the drug-binding specificity of human serum albumin. J Mol Biol. 2005;353:38–52. doi: 10.1016/j.jmb.2005.07.075. [DOI] [PubMed] [Google Scholar]
- 76.Bellamri M, Le Hegarat L, Turesky RJ, Langouët S. Metabolism of the Tobacco Carcinogen 2-Amino-9H-pyrido[2,3-b]indole (AalphaC) in Primary Human Hepatocytes. Chem Res Toxicol. 2016 doi: 10.1021/acs.chemrestox.6b00394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Turesky RJ, Lang NP, Butler MA, Teitel CH, Kadlubar FF. Metabolic activation of carcinogenic heterocyclic aromatic amines by human liver and colon. Carcinogenesis. 1991;12:1839–1845. doi: 10.1093/carcin/12.10.1839. [DOI] [PubMed] [Google Scholar]
- 78.Hoelzl C, Glatt H, Meinl W, Sontag G, Haidinger G, Kundi M, Simic T, Chakraborty A, Bichler J, Ferk F, Angelis K, Nersesyan A, Knasmuller S. Consumption of Brussels sprouts protects peripheral human lymphocytes against 2-amino-1-methyl-6-phenylimidazo[4,5-b]pyridine (PhIP) and oxidative DNA-damage: results of a controlled human intervention trial. Mol Nutr Food Res. 2008;52:330–341. doi: 10.1002/mnfr.200700406. [DOI] [PubMed] [Google Scholar]
- 79.Wells PG, Mackenzie PI, Chowdhury JR, Guillemette C, Gregory PA, Ishii Y, Hansen AJ, Kessler FK, Kim PM, Chowdhury NR, Ritter JK. Glucuronidation and the UDP-glucuronosyltransferases in health and disease. Drug Metab Dispos. 2004;32:281–290. doi: 10.1124/dmd.32.3.281. [DOI] [PubMed] [Google Scholar]
- 80.Lodemann P, Schorer G, Frey BM. Wrong molar hemoglobin reference values-a longstanding error that should be corrected. Ann Hematol. 2010;89:209. doi: 10.1007/s00277-009-0791-x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.