

Abstract
Background:
Radix Saposhnikoviae (RS) exerts anti-inflammatory, analgesic, antipyretic, antioxidation effects and has been used in traditional Chinese medicine to treat common colds, headache, and rheumatoid arthritis. Prim-O-glucosylcimifugin (POG) is the highest content chromone and one of the major active constituents in RS.
Objective:
The study was aimed to explore the anti-inflammation effects of POG in lipopolysaccharide (LPS)-stimulated RAW 264.7 macrophages.
Materials and Methods:
Cell viability was detected by Cell Counting Kit-8 assay. Production of nitric oxide (NO), tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β), and IL-6 was assessed by enzyme-linked immunosorbent assay. Real-time polymerase chain reaction and Western blot were performed to analyze mRNA and protein levels, respectively.
Results:
During the whole experiment, 15, 50, and 100 μg/mL of POG had no cytotoxicity on RAW 264.7 cells. POG dose-dependently inhibited the production of NO, TNF-α, IL-1β, and IL-6 that were induced by LPS. POG treatment downregulated the mRNA and protein expression inducible NO synthase (iNOS) and cyclooxygenase 2 (COX-2) in LPS-activated RAW 264.7 macrophages in a concentration-dependent manner. Furthermore, LPS-induced JAK2/STAT3 activation was prevented in RAW 264.7 macrophages by POG treatment. STAT3 overexpression significantly reversed the effects of POG on LPS-activated RAW 264.7 macrophages.
Conclusion:
These results demonstrate that POG exerts anti-inflammatory effects through the inhibition of iNOS and COX-2 expression by inhibiting the phosphorylation of JAK2/STAT3.
SUMMARY
POG exerts anti-inflammatory effects in RAW 264.7 macrophages through the inhibition of iNOS and COX-2 expression by inhibiting JAK2/STAT3 signaling.
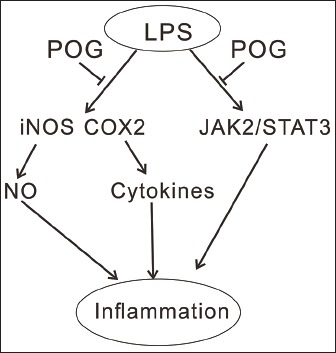
Abbreviations used: LPS: Lipopolyssacharide; NO: Nitric oxide; TNF-α: Tumor necrosis factor-α; IL: Interleukin; RS: Radix Saposhnikoviae; POG: Prim-O-glucosylcimifugin; iNOS: Inducible NO synthase; COX2: Cyclooxygenase; FBS: Fetal bovine serum; DMSO: Dimethylsulfoxide; CCK-8: Cell Counting Kit; RIPA: Radio immunoprecipitation assay buffer; ECL: Enhanced chemiluminescence; SD: Standard deviation; ELISA: Enzyme-Linked immunosorbent assay.
Keywords: Anti-inflammation, cytokines, JAK2/STAT3, nitric oxide, prim-O-glucosylcimifugin
INTRODUCTION
Inflammation is a protective response to local injury. Inflammatory responses mediated by various immune cells can eliminate the initial cause of injury, clear out necrotic cells, and initiate tissue repair. However, deregulated inflammation leads to overproduction of pro-inflammatory cytokines, which can cause exacerbate tissue damage and chronic inflammatory diseases.[1] Macrophages play essential role in inflammation. Compounds that can regulate macrophage activation may be used for the treatment of chronic inflammatory diseases such as asthma, atherosclerosis, and rheumatoid arthritis. Lipopolysaccharide (LPS) treatment of RAW 264.7 murine macrophages induces the production of inflammatory mediators, including nitric oxide (NO), tumor necrosis factor-α (TNF-α), interleukin (IL)-1β, and IL-6[2,3,4] and have been widely used to assess the anti-inflammation effects of compounds.[5,6,7,8]
Radix Saposhnikoviae (RS, Fang-feng in Chinese) is dried root of Saposhnikovia divaricata (Turcz.) Schischek (Umbelliferae), which is mainly distributed in Northeastern and Northern China.[9] RS is a well-known traditional Chinese medicine that has been commonly used in the clinical compound prescription to treat common colds, headache, rheumatoid arthritis, and eczema in China and other Asian countries.[9,10,11] Pharmacological experiments have demonstrated the anti-inflammatory, analgesic, antipyretic, antioxidation properties of RS.[10,11,12,13,14,15,16,17] Several kind of compounds have been isolated from RS, including chromones, coumarins, mannitol, glycoside, and polyacetylenes.[18,19] The major active constituents of RS are considered to be chromones and their glycosides.[10,18,20,21] Prim-O-glucosylcimifugin (POG) [Figure 1] is the highest content chromone of RS.[17] It was reported that POG suppressed LPS-induced nitrite production in RAW 264.7 cells[12] and exhibited significant anti-inflammatory function on mouse ear inflammation.[11]
Figure 1.
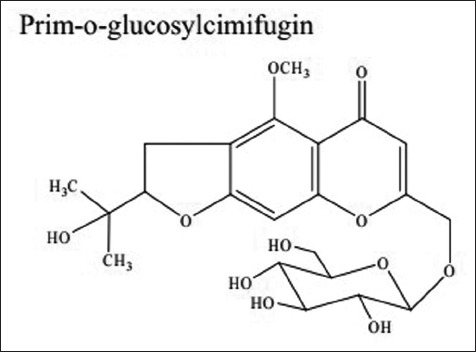
Structure of prim-O-glucosylcimifugin
The present study was aimed to further explore the anti-inflammatory activity of POG in LPS-stimulated RAW 264.7 macrophages and investigated possible mechanisms. We found that POG inhibited LPS-induced production of NO and several cytokines, expression of inducible NO synthase (iNOS) and cyclooxygenase (COX-2) in a concentration-dependent manner. The activation of JAK2/STAT3 in response to LPS was also blocked by POG treatment.
MATERIALS AND METHODS
Cell culture
RAW 264.7 murine macrophages were obtained from American Type Culture Collection (Rockville, MD, USA) and cultured in Dulbecco's modified Eagle's medium (Hyclone, Logan, UT, USA) supplemented with 10% fetal bovine serum (FBS, Gibco® Life Technology, Carlsbad, CA, USA), penicillin (100 units/mL, Solarbio, Beijing, China), and streptomycin (100 μg/mL, Solarbio). The cells were maintained at 37°C in a 5% CO2/95% humidified atmosphere, subjected to no more than 20 cell passages and utilized for experimentation at approximately 60%–80% confluence.
Cell viability
Cell Counting Kit (CCK-8, Dojindo Laboratories, Tokyo, Japan) was used to determine the cytotoxic concentrations of POG. In brief, the Raw 264.7 cells were plated at a density of 1 × 104 cells per well in a 96-well and incubated overnight. Cells were then stimulated with 1 μg/ml LPS (Sigma, St. Louis, MO, USA) and treated with various concentrations of POG (15, 50, and 100 μg/mL; MedChem Express, Princeton, NJ, USA) or dimethyl sulfoxide (DMSO, Sigma; control cells). After incubation at 37°C for 24 h, CCK-8 solution was added to each well and incubated for another 1 h. The absorbance was measured at 450 nm using a microplate reader (Emax, Molecular Devices, Sunnyvale, CA, USA). The cell viability was defined as the percentage of control cells:
Cell viability (%) = Absorbance of POG-treated cells/absorbance of control cells × 100%.
Cell activation and treatment
RAW 264.7 cells were seeded to 6-well plates (5 × 105 cells per well) and incubated overnight. Cells were then stimulated with 1 μg/ml LPS (Sigma) and treated with various concentrations of POG (15, 50, and 100 μg/mL) or DMSO. After an additional 24 h, supernatants were collected for evaluation of cytokine concentration and NO production, and cells were harvested for real-time polymerase chain reaction (PCR) and Western blotting analysis.
Lentiviral infection
The full-length murine STAT3 was cloned into the lentiviral expression vector pLVX-puro (Clontech, Palo Alto, CA, USA) by Genewiz Company (Shanghai, China). RAW 264.7 cells in 6-well plates were infected with STAT3 expressing (STAT3) and control (Vector) lentivirus. After 24 h, cells were stimulated with 1 μg/ml LPS (Sigma) and treated with 100 μg/mL POG or DMSO. After an additional 24 h, supernatants were collected and cells were harvested.
Measurement of nitric oxide production
NO production in the medium was measured according to the Griess reaction method as previously described.[22] In brief, equal volumes of cultured medium from treated cells and Griess reagent (Jiancheng Bioengineering Institute, Nanjing, China) were mixed and incubated in the dark for 10 min. The absorbance at 540 nm was determined with a microplate reader (Molecular Devices, Sunnyvale, CA, USA).
RNA extraction and real-time polymerase chain reaction
Total RNA was extracted from treated cells using TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and reverse transcribed with cDNA synthesis kit (Thermo Fisher Scientific, Rockford, IL, USA), according to the manufacturer's instructions. Real-time PCR was conducted on an ABI 7300 real-time PCR machine (Applied Biosystems, Foster City, CA, USA) using qPCR SYBR Green Mix (Thermo Fisher Scientific, Rockford, IL, USA) with GAPDH as internal control. The following cycling parameters were used: 95°C for 10 min, followed by 40 cycles of 95°C for 15 s, 60°C for 45 s. The gene expression was calculated using the ΔΔ Ct method. The primers used were list as follows: iNOS (NM_008713.4), 5’-TTGTCTGCGGCGATGTCAC-3’, and 5’-TGCGTATGCGGCTTGTCAC-3’; COX-2 (NM_011198.3), 5’-CCTGGTCTGATGATGTATGC-3’, and 5’-GTATGAGTCTGCTGGTTTGG-3’; GAPDH (NM_008084.2), 5’-ATCACTGCCACCCAGAAG-3’, and 5’-TCCACGACGGACACATTG-3’.
Western blotting and antibodies
Treated RAW 264.7 macrophages were washed twice with ice-cold PBS and lysed in ice-cold radioimmunoprecipitation assay buffer (RIPA, Beyotime, Shanghai, China). 25 μg of total proteins was separated on 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto nitrocellulose membrane (Millipore, Bedford, USA), and the nonspecific binding of antibodies was blocked with 5% skim milk. The membranes were then incubated with primary antibodies and corresponding secondary antibodies (Beyotime). The signals were detected with enhanced chemiluminescence (ECL, Millipore). The target bands were quantified using Image J software (http://rsb.info.nih.gov/ij/, Bethesda, MD, USA) with GAPDH as internal control.
Antibodies against COX-2, GAPDH, JAK2, p-JAK2, and STAT3 were obtained from Cell Signaling Technology (Danvers, MA, USA) while anti-p-STAT3 and anti-iNOS were from Abcam (Cambridge, MA, USA).
Enzyme-linked immunosorbent assay analysis
Concentrations of TNF-α, IL-6, and IL-1β in the culture medium were assessed by enzyme-linked immunosorbent assay (ELISA). Assays were performed according to the instructions of the manufacturer (Bio-Swamp Life Science, Shanghai, China). Plates were read at 450 nm (correction wavelength 570 nm) using a microplate reader (Molecular Devices, Sunnyvale, CA, USA).
Data analysis
The results were presented as means ± standard deviation from three independent experiments. One-way ANOVA was applied to analyze the statistical significance of the differences between the study groups. Differences were considered to be statistically significant at P < 0.05.
RESULTS
Effects of prim-O-glucosylcimifugin on cell viability of RAW 264.7 macrophages
We first measured the cytotoxicity of POG to LPS-activated Raw 264.7 macrophages. Raw 264.7 macrophages were treated with LPS (1 μg/mL) and increasing concentrations of POG (15, 50, and 100 μg/mL) for 24 h and cell viability was evaluated by CCK-8 assay. As shown in Figure 2, cell viability was not significantly affected after 24 h and exposure to 15–100 μg/mL POG as compared with DMSO-treated cells (control).
Figure 2.
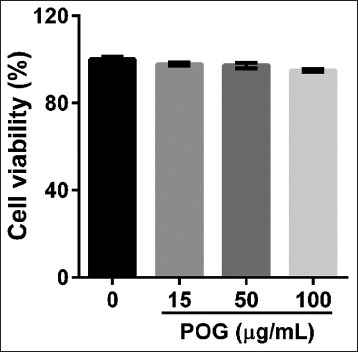
Effects of prim-O-glucosylcimifugin on cell viability with a Cell Counting Kit. RAW 264.7 cells were exposed with lipopolyssacharide (1 μg/mL) and various concentrations of prim-O-glucosylcimifugin (15, 50 and 100 μg/mL) or dimethyl sulfoxide alone. Cell viability was assessed 24 h after treatment and expressed as percentage of the dimethyl sulfoxide control. All values are means ± standard deviation (n = 3)
Prim-O-glucosylcimifugin treatment inhibits lipopolysaccharide-induced nitric oxide production in RAW 264.7 macrophages
To investigate the anti-inflammatory effect of POG, we examined whether POG could affect NO synthesis in LPS-activated RAW 264.7 cells. Macrophages were treated with LPS (1 μg/mL) and various concentrations of POG (15, 50, and 100 μg/mL) for 24 h. No concentrations were measured in the culture supernatants by Griess reaction. As shown in Figure 3a, the concentrations of NO in the culture supernatants were markedly increased in response to LPS exposure, and POG significantly inhibited LPS-induced NO production in a concentration-dependent manner.
Figure 3.
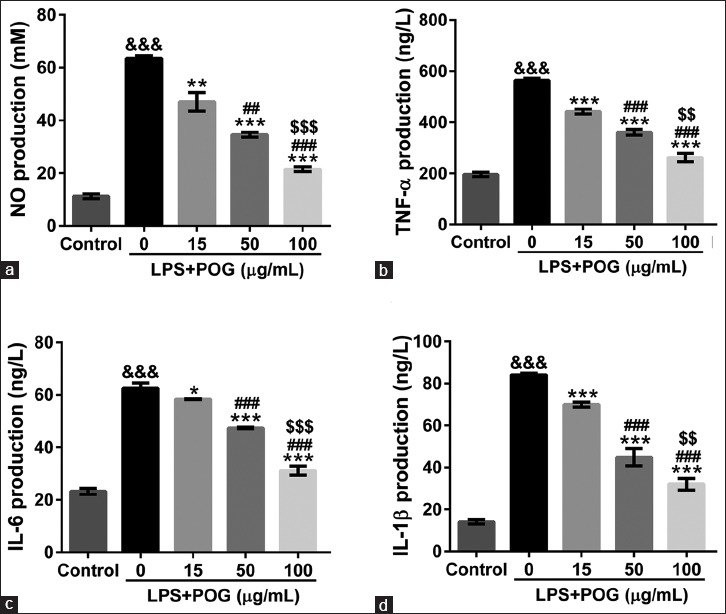
Effects of prim-O-glucosylcimifugin on lipopolyssacharide-induced NO and cytokine production. Raw 264.7 cells were incubated in a medium containing lipopolysaccharide (1 μg/mL) and various concentrations of prim-O-glucosylcimifugin (15, 50, and 100 μg/mL). Cells treated with dimethyl sulfoxide were set as control. The amount of nitrite (a), tumor necrosis factor-α (b), interleukin-6 (c), and interleukin-1β (d) in the medium was monitored at 24 h after exposure as described in Materials and Methods. All values are means ± standard deviation (n = 3). &&&P < 0.001 versus control; *P < 0.05, **P < 0.01, and ***P < 0.001 versus lipopolysaccharide-treated cells; #P < 0.05, ##P < 0.01, and ###P < 0.001 versus lipopolysaccharide and 15 μg/mL prim-O-glucosylcimifugin-treated cells; $$P < 0.01 and $$$P < 0.001 versus lipopolysaccharide and 50 μg/mL prim-O-glucosylcimifugin-treated cells
Prim-O-glucosylcimifugin inhibits cytokine production in lipopolysaccharide-treated macrophage
TNF-α, IL-6, and IL-1β are crucial cytokines involved in response to LPS.[3,4] To explore the effects of POG on TNF-α, IL-6, and IL-1β secretion by activated macrophages, cytokine concentrations in the cultured supernatants of treated RAW 264.7 cells were assessed by ELISA. LPS stimulation significantly increased cytokine production of RAW 264.7 cells, which was notably suppressed by POG exposure (15, 50, and 100 μg/mL) dose-dependently [Figure 3b–d].
Prim-O-glucosylcimifugin inhibits lipopolysaccharide-induced expression of inducible nitric oxide synthase and cyclooxygenase 2
iNOS and COX-2 serve as key mediators of inflammation[23,24] and could be induced by LPS and several cytokines.[25] We then assessed the inhibitory effects of different concentrations of POG on the LPS-induced expression of iNOS and COX-2. Stimulation of the RAW 264.7 cells with LPS evidently upregulated the mRNA and protein levels of iNOS and COX-2 as determined by real-time PCR and Western blotting, respectively [Figure 4]. POG inhibited the expression of both proteins in a dose-dependent manner.
Figure 4.
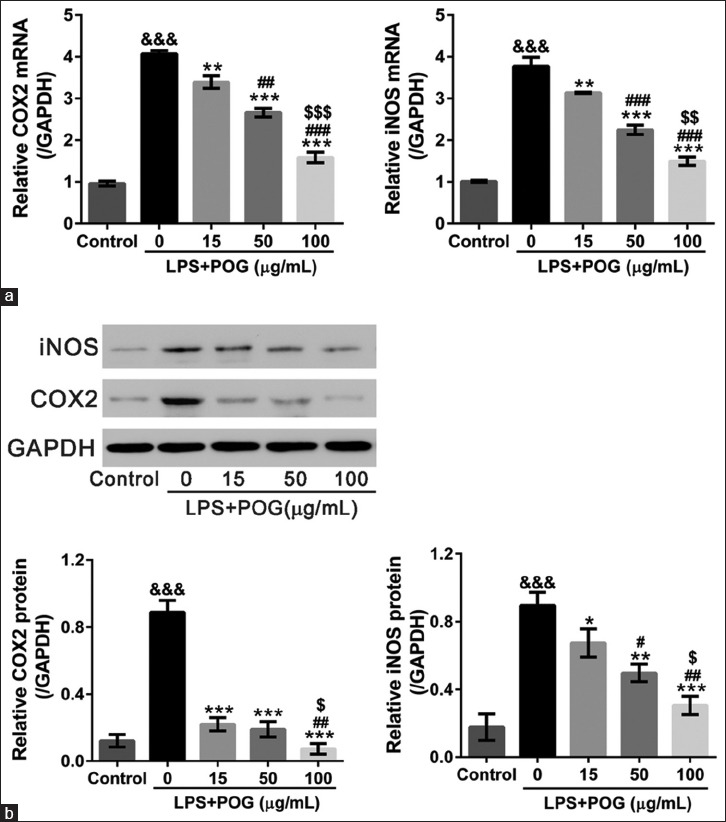
Effects of prim-O-glucosylcimifugin on the inducible nitric oxide synthase and cyclooxygenase 2 expressions of lipopolysaccharide-stimulated RAW 264.7 cells. RAW 264.7 cells were treated with lipopolysaccharide (1 μg/mL) and various concentrations of prim-O-glucosylcimifugin (15, 50, and 100 μg/mL) and harvest at 24 h posttreatment. Cells treated with dimethyl sulfoxide only were set as control. (a) mRNA expression of inducible nitric oxide synthase and cyclooxygenase 2 was detected by real-time polymerase chain reaction with GAPDH as internal control; All values are means ± standard deviation (n = 3). (b) Protein expression of inducible nitric oxide synthase and cyclooxygenase 2 was detected by Western blotting. Representative Western blots (upper panel) and quantitative results (lower panel) were shown. All values are means ± standard deviation (n = 3). &&&P < 0.001 versus control; *P < 0.05, **P < 0.01, and ***P < 0.001 versus lipopolysaccharide-treated cells; #P < 0.05, ##P < 0.01, and ###P < 0.001 versus lipopolysaccharide and 15 μg/mL prim-O-glucosylcimifugin-treated cells; $P < 0.05, $$P < 0.01, and $$$P < 0.001 versus lipopolysaccharide and 50 μg/mL prim-O-glucosylcimifugin-treated cells
Involvement of JAK2/STAT3 signaling
JAK/STAT pathway is considered to be involved in the upregulation of iNOS and COX-2 in H2O2 preconditioned cells.[26] Figure 5a showed that LPS-activated JAK2/STAT3 phosphorylation was blocked by POG treatment in a concentration-dependent manner.
Figure 5.
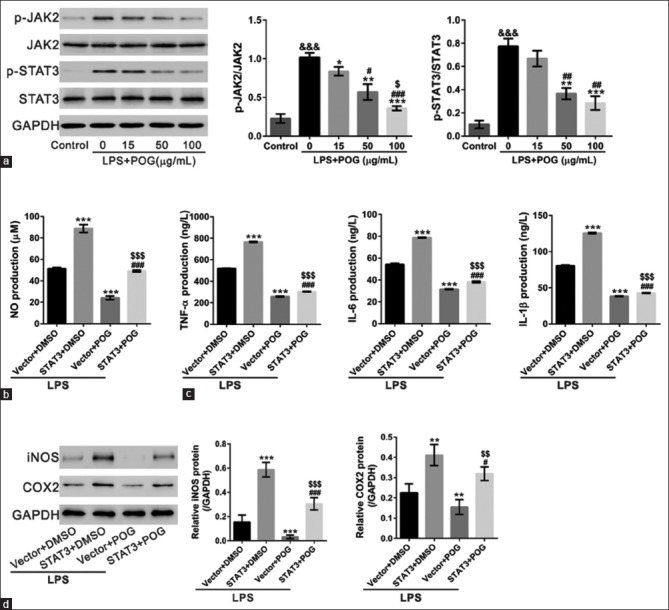
Involvement of JAK2/STAT3 signaling. (a) RAW 264.7 cells were treated with lipopolysaccharide (1 μg/mL) and various concentrations of prim-O-glucosylcimifugin (15, 50 and 100 μg/mL) and harvest at 4 h posttreatment. Cells treated with dimethyl sulfoxide were set as control. p-JAK2, JAK2, p-STAT3, and STAT3 were detected by Western blotting. Representative Western blots (left panel) and quantitative results (right panel) were shown. All values are means ± standard deviation (n = 3). &&&P < 0.001 versus control; *P < 0.05, **P < 0.01, and ***P < 0.001 versus lipopolysaccharide-treated cells; #P < 0.05 and ##P < 0.01 versus lipopolysaccharide and 15 μg/mL prim-O-glucosylcimifugin-treated cells;$P < 0.05 versus lipopolysaccharide and 50 μg/mL prim-O-glucosylcimifugin-treated cells. (b-d) RAW 264.7 cells were infected with STAT3 expression lentivirus or control vector lentivirus (Vector). After 24 h, cells were treated with lipopolysaccharide (1 μg/mL), and dimethyl sulfoxide or 100 μg/mL prim-O-glucosylcimifugin. The amount of nitrite (b) and cytokines (c) in the medium was monitored at 24 h after exposure. Protein expression of inducible nitric oxide synthase and cyclooxygenase 2 (d) in RAW 264.7 cells was detected by Western blotting. **P < 0.01 and ***P < 0.001 versus Vector + lipopolysaccharide; #P < 0.05 and ###P < 0.001 versus STAT3 + lipopolysaccharide;$P < 0.05,$$P < 0.01, and $$$P < 0.001 versus POS + lipopolysaccharide
To further investigate the involvement of STAT3, RAW 264.7 cells were infected with STAT3 expressing or Vector lentivirus, and then treated with LPS and 100 μg/mL POG or DMSO. As shown in Figure 5b–d, STAT3 overexpression partially reversed the effects of POG on NO production, cytokine production, and iNOS and COX-2 expression. These data suggested that POG exerted anti-inflammatory effects through suppressing the activation of JAK2/STAT3 pathway.
DISCUSSION
For many years, RS has been commonly used to treat common colds, headache, rheumatoid arthritis, and eczema.[9,10,11] Chromones and their glycosides are major active constituents of RS.[10,18,20,21] POG is the highest content chromone of RS[17] and known to exhibited anti-inflammatory function.[11,12] TNF-α, as the primary mediator of the systematic toxicity of LPS,[27,28] can induce the secretion of cytokines such as IL-6 and IL-1β.[29] These cytokines are mainly produced by macrophages and play a key role in inflammatory conditions. NO is a free radical generated by NO synthase and serves as an important cellular second messenger. NO induces tissue injury at the inflammatory site.[30] In the current study, we demonstrated the potential of POG to decrease the LPS-induced production or expression of NO, TNF-α, IL-6, and IL-1β [Figure 3] and further confirmed its anti-inflammatory function.
Previous studies have shown that COX and iNOS are involved in chronic inflammatory disease. LPS and inflammatory cytokines in the macrophages have been found to induce both iNOS and COX-2.[23,24,25] Here, the mRNA and protein expression of iNOS and COX-2 were induced by LPS exposure while significantly decreased by additional POG treatment [Figure 4]. Our data suggest that POG modulates iNOS and COX-2 expression at the transcriptional level.
JAK/STAT signal pathway plays a pivotal role in mediating the biological response for inflammation. It has been reported that JAK/STAT signaling is involved in LPS-mediated iNOS production.[31,32] In H2O2 preconditioned cells, JAK/STAT pathway contributes to the upregulation of iNOS and COX-2.[26] In this study, POG inhibited the phosphorylation of JAK2 and STAT3. STAT3 overexpression partially reversed the inhibitory effects of POG on iNOS and COX-2 expression [Figure 5]. Thus, it seems that POG modulates iNOS and COX-2 expression through regulating JAK2/STAT3 signaling although the mechanism is needed further study.
CONCLUSION
We determined that the POG has anti-inflammatory potential in LPS-activated Raw 264.7 cells through suppressing the activation of JAK2/STAT3 signaling and through inhibiting the expression of iNOS and COX-2.
Financial support and sponsorship
This work was supported by funds from the Special Development of Traditional Chinese Medicine of Pudong New Area of Shanghai (PDYNZJ2015-03), Outstanding Young Talents Training Program of Pudong Health Bureau of Shanghai (No. PWRq2014-09), and Key Discipline Construction Project of Pudong Health Bureau of Shanghai (No. PWZxq2014-16).
Conflicts of interest
There are no conflicts of interest.
REFERENCES
- 1.Morson BC. Pathology of inflammatory diseases. Proc R Soc Med. 1970;63(Suppl (1)):63. [PMC free article] [PubMed] [Google Scholar]
- 2.Hewett JA, Roth RA. Hepatic and extrahepatic pathobiology of bacterial lipopolysaccharides. Pharmacol Rev. 1993;45:382–411. [PubMed] [Google Scholar]
- 3.Rossol M, Heine H, Meusch U, Quandt D, Klein C, Sweet MJ, et al. LPS-induced cytokine production in human monocytes and macrophages. Crit Rev Immunol. 2011;31:379–446. doi: 10.1615/critrevimmunol.v31.i5.20. [DOI] [PubMed] [Google Scholar]
- 4.Schletter J, Heine H, Ulmer AJ, Rietschel ET. Molecular mechanisms of endotoxin activity. Arch Microbiol. 1995;164:383–9. doi: 10.1007/BF02529735. [DOI] [PubMed] [Google Scholar]
- 5.Kang SM, Kim KN, Lee SH, Ahn G, Cha SH, Kim AD, et al. Anti-inflammatory activity of polysaccharide purified from AMG-assistant extract of Ecklonia cava in LPS-stimulated RAW 264.7 macrophages. Carbohydr Polym. 2011;85:80–5. [Google Scholar]
- 6.Kim HG, Shrestha B, Lim SY, Yoon DH, Chang WC, Shin DJ, et al. Cordycepin inhibits lipopolysaccharide-induced inflammation by the suppression of NF-κB through Akt and p38 inhibition in RAW 264.7 macrophage cells. Eur J Pharmacol. 2006;545:192–9. doi: 10.1016/j.ejphar.2006.06.047. [DOI] [PubMed] [Google Scholar]
- 7.Ryu M, Kim EH, Chun M, Kang S, Shim B, Yu YB, et al. Astragali Radix elicits anti-inflammation via activation of MKP-1, concomitant with attenuation of p38 and Erk. J Ethnopharmacol. 2008;115:184–93. doi: 10.1016/j.jep.2007.09.027. [DOI] [PubMed] [Google Scholar]
- 8.Heo SJ, Yoon WJ, Kim KN, Ahn GN, Kang SM, Kang DH, et al. Evaluation of anti-inflammatory effect of fucoxanthin isolated from brown algae in lipopolysaccharide-stimulated RAW 264.7 macrophages. Food Chem Toxicol. 2010;48:2045–51. doi: 10.1016/j.fct.2010.05.003. [DOI] [PubMed] [Google Scholar]
- 9.Shao ML. Pharmacopoeia of the People's Republic of China. Beijing: Chemical Industry Press; 2010. [Google Scholar]
- 10.Okuyama E, Hasegawa T, Matsushita T, Fujimoto H, Ishibashi M, Yamazaki M. Analgesic components of saposhnikovia root (Saposhnikovia divaricata) Chem Pharm Bull (Tokyo) 2001;49:154–60. doi: 10.1248/cpb.49.154. [DOI] [PubMed] [Google Scholar]
- 11.Xue BY, Li W, Li L, Xiao YQ. A pharmacodynamic research on chromone glucosides of fangfeng. Zhongguo Zhong Yao Za Zhi. 2000;25:297–9. [PubMed] [Google Scholar]
- 12.Wang CC, Chen LG, Yang LL. Inducible nitric oxide synthase inhibitor of the Chinese herb I. Saposhnikovia divaricata (Turcz.) Schischk. Cancer Lett. 1999;145:151–7. doi: 10.1016/s0304-3835(99)00248-7. [DOI] [PubMed] [Google Scholar]
- 13.Wang CN, Shiao YJ, Kuo YH, Chen CC, Lin YL. Inducible nitric oxide synthase inhibitors from Saposhnikovia divaricata and Panax quinquefolium. Planta Med. 2000;66:644–7. doi: 10.1055/s-2000-8624. [DOI] [PubMed] [Google Scholar]
- 14.Ge WH, Guo JY, Shen YJ, Chen ML, Shi SL, Han YH, et al. Effects of volatie oil of Schizonepeta tenuifolia Briq herb and Saposhnikovia divaricata Schischke root on proinflammatory cytokine expression and regulation. Zhongguo Zhong Yao Za Zhi. 2007;32:1777–9. [PubMed] [Google Scholar]
- 15.Tai J, Cheung S. Anti-proliferative and antioxidant activities of Saposhnikovia divaricata. Oncol Rep. 2007;18:227–34. [PubMed] [Google Scholar]
- 16.Guo LQ, Taniguchi M, Chen QY, Baba K, Yamazoe Y. Inhibitory potential of herbal medicines on human cytochrome P450-mediated oxidation: Properties of umbelliferous or citrus crude drugs and their relative prescriptions. Jpn J Pharmacol. 2001;85:399–408. doi: 10.1254/jjp.85.399. [DOI] [PubMed] [Google Scholar]
- 17.Kong X, Liu C, Zhang C, Zhao J, Wang J, Wan H, et al. The suppressive effects of Saposhnikovia divaricata (Fangfeng) chromone extract on rheumatoid arthritis via inhibition of nuclear factor-kappaB and mitogen activated proteinkinases activation on collagen-induced arthritis model. J Ethnopharmacol. 2013;148:842–50. doi: 10.1016/j.jep.2013.05.023. [DOI] [PubMed] [Google Scholar]
- 18.Gui Y, Tsao R, Li L, Liu CM, Wang J, Zong X. Preparative separation of chromones in plant extract of Saposhnikovia divaricata by high-performance counter-current chromatography. J Sep Sci. 2011;34:520–6. doi: 10.1002/jssc.201000721. [DOI] [PubMed] [Google Scholar]
- 19.Kang J, Sun JH, Zhou L, Ye M, Han J, Wang BR, et al. Characterization of compounds from the roots of Saposhnikovia divaricata by high-performance liquid chromatography coupled with electrospray ionization tandem mass spectrometry. Rapid Commun Mass Spectrom. 2008;22:1899–911. doi: 10.1002/rcm.3559. [DOI] [PubMed] [Google Scholar]
- 20.Sasaki H, Taguchi H, Endo T, Yosioka I. The constituents of Ledebouriella seseloides wolff. I. Structures of three new chromones. Chem Pharm Bull. 1982;30:3555–62. [Google Scholar]
- 21.Dai J, Chen X, Cheng W, Liu X, Fan X, Shen Z, et al. A sensitive liquid chromatography-mass spectrometry method for simultaneous determination of two active chromones from saposhnikovia root in rat plasma and urine. J Chromatogr B Analyt Technol Biomed Life Sci. 2008;868:13–9. doi: 10.1016/j.jchromb.2008.03.031. [DOI] [PubMed] [Google Scholar]
- 22.Guevara I, Iwanejko J, Dembinska-Kiec A, Pankiewicz J, Wanat A, Anna P, et al. Determination of nitrite/nitrate in human biological material by the simple Griess reaction. Clin Chim Acta. 1998;274:177–88. doi: 10.1016/s0009-8981(98)00060-6. [DOI] [PubMed] [Google Scholar]
- 23.Moncada S, Palmer RM, Higgs EA. Nitric oxide: Physiology, pathophysiology, and pharmacology. Pharmacol Rev. 1991;43:109–42. [PubMed] [Google Scholar]
- 24.Seibert K, Masferrer J, Zhang Y, Gregory S, Olson G, Hauser S, et al. Mediation of inflammation by cyclooxygenase-2. Agents Actions Suppl. 1995;46:41–50. doi: 10.1007/978-3-0348-7276-8_5. [DOI] [PubMed] [Google Scholar]
- 25.Di Rosa M, Ialenti A, Ianaro A, Sautebin L. Interaction between nitric oxide and cyclooxygenase pathways. Prostaglandins Leukot Essent Fatty Acids. 1996;54:229–38. doi: 10.1016/s0952-3278(96)90053-8. [DOI] [PubMed] [Google Scholar]
- 26.Yu H, Liu Z, Zhou H, Dai W, Chen S, Shu Y, et al. JAK-STAT pathway modulates the roles of iNOS and COX-2 in the cytoprotection of early phase of hydrogen peroxide preconditioning against apoptosis induced by oxidative stress. Neurosci Lett. 2012;529:166–71. doi: 10.1016/j.neulet.2012.09.013. [DOI] [PubMed] [Google Scholar]
- 27.Cerami A, Beutler B. The role of cachectin/TNF in endotoxic shock and cachexia. Immunol Today. 1988;9:28–31. doi: 10.1016/0167-5699(88)91353-9. [DOI] [PubMed] [Google Scholar]
- 28.Heumann D, Glauser M. Anticytokine strategies for the treatment of septic shock: Relevance of animal models. Pathology of Septic Shock. Berlin Heidelberg: Springer; 1996. pp. 299–311. [DOI] [PubMed] [Google Scholar]
- 29.Woolf CJ, Allchorne A, Safieh-Garabedian B, Poole S. Cytokines, nerve growth factor and inflammatory hyperalgesia: The contribution of tumour necrosis factor alpha. Br J Pharmacol. 1997;121:417–24. doi: 10.1038/sj.bjp.0701148. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Nathan C. Nitric oxide as a secretory product of mammalian cells. FASEB J. 1992;6:3051–64. [PubMed] [Google Scholar]
- 31.Hong J, Sang S, Park HJ, Kwon SJ, Suh N, Huang MT, et al. Modulation of arachidonic acid metabolism and nitric oxide synthesis by garcinol and its derivatives. Carcinogenesis. 2006;27:278–86. doi: 10.1093/carcin/bgi208. [DOI] [PubMed] [Google Scholar]
- 32.Jacobs AT, Ignarro LJ. Lipopolysaccharide-induced expression of interferon-beta mediates the timing of inducible nitric-oxide synthase induction in RAW 264.7 macrophages. J Biol Chem. 2001;276:47950–7. doi: 10.1074/jbc.M106639200. [DOI] [PubMed] [Google Scholar]