

Abstract
Background:
Plantago major has been reported to have anticancer and anti-inflammatory properties. However, its antiproliferative and anti-inflammatory mechanisms have not been fully elucidated. Moreover, which plant parts are more suitable as starting materials has not been explored.
Objectives:
To investigate the antiproliferative activity of P. major extracts against MCF-7, MDA-MB-231, HeLaS3, A549, and KB cancer cell lines as well as their effects on inflammatory cytokines (tumor necrosis factor [TNF]-α, interleukin [IL]-1β, IL-6, and interferon [IFN]-γ) production by lipopolysaccharide (LPS)-stimulated THP-1 macrophages.
Materials and Methods:
The methanol and aqueous extracts of P. major from different plant parts and its chemical compounds, i.e., ursolic acid (UA), oleanolic acid (OA), and aucubin were tested in this experiment.
Results:
Methanol and aqueous extracts of P. major seeds exhibited the greatest antiproliferative activity. The methanol extracts of seeds also demonstrated the highest inhibition of TNF-α, IL-1β, IL-6, and IFN-γ production. Interestingly, the roots, which were commonly discarded, exhibited comparable activities to those of leaves and petioles. Furthermore, UA exhibited stronger activities than OA and aucubin.
Conclusions:
The seeds are being proposed as the main source for further development of anticancer and anti-inflammatory products, whereas the roots could be included in the preparation of P. major derived products with respect to anti-inflammatory.
SUMMARY
Amongst the parts of Plantago major, seeds exhibited the greatest antiproliferative activity against MCF-7, MDA-MB-231, HeLaS3, A549, and KB cell lines as well as the highest inhibition on TNF-α, IL-1β, IL-6, and IFN-γ production
The roots, which were commonly discarded, exhibited comparable antiproliferative and cytokines inhibition activities to those of leaves and petioles
Ursolic acid, a chemical compound of Plantago major, exhibited stronger activities than oleanolic acid and aucubin
The seeds are being proposed as the main source for further development of anticancer and anti inflammatory products, whereas the roots could be included in the preparation of Plantago major derived products with respect to anti inflammatory.
Abbreviations used: TNF: Tumor Necrosis Factor; IL: Interleukin; IFN: Interferon; HPTLC: High Performance Thin Layer Chromatography; UA: Ursolic Acid; OA: Oleanolic Acid; AUC: Aucubin.
Keywords: Anti-inflammatory, antiproliferative, cytokines, Plantago major, THP-1 macrophages
INTRODUCTION
Plantago L. (Plantaginaceae) is a genus widely distributed all over the world, and more than 275 species have been reported.[1] Plantago major [Figure 1] is an endemic species found in Indonesia. This plant has both perennial and annual growth habit, with ovoid or elliptic leaves. The leaves are normally green, sometimes with purple shading and are hairless or hairy on the surface, with 5–9 veins in parallel venation. Flowering of P. major is a spike which arises up to 30 cm in length. The fruit is a capsule (5 mm in length) which produced on a spike. Every centimeter of spike contains 23–26 capsules, and each capsule carries 4–15 seeds.[2,3,4] It is used traditionally and commercially available.[5] This herb can be found in extremely different geographical origins. Previous studies have indicated a variety of beneficial pharmacological effects of P. major such as anticancer,[6,7,8,9] antioxidant,[1,10,11] and immunomodulatory.[6,12] Concerning its anti-inflammatory activity, several in vivo experiments were carried out to prove this property.[13,14] Inflammatory responses involve critical immune cells, macrophages, which are able to produce various inflammatory mediators including cytokines (tumor necrosis factor [TNF]-α, interleukin [IL]-1αβ, IL-6, IL-8, IL-10, and IL-12), eicosanoid derived from arachidonic acid, and reactive oxygen species.[15] Various in vitro studies demonstrated the anti-inflammatory potency of P. major and its active compounds through inhibition of cyclooxygenases and lipoxygenases, enzymes involved in arachidonic acids pathways.[16,17,18] Anti-inflammatory activity of P. major through modulation of inflammatory cytokines, however, has not been well characterized.
Figure 1.

Plantago major.
In terms of plant parts, leaves of P. major were most frequently used mainly as aqueous preparations, whereas roots were utilized in limited country, especially Iran.[3] In many other countries, including Indonesia, roots are commonly discarded without any utilization. Nevertheless, our previous studies showed that root extracts exerted cytotoxicity on SiHa and HepG2 cell lines as well as possessed free radical scavenging activity which were comparable to those of the other plant parts.[5] To more understand the anti-inflammatory actions of P. major, we investigated the effect on cytokines (TNF-α, IL-1β, IL-6, and interferon [IFN]-γ) production in LPS-activated THP-1 macrophages cells by P. major extracts and its chemical compounds. Their antiproliferative effect against KB, MCF-7, MDA-MB-231, A-549, and HeLaS3 cell lines was determined as well.
MATERIALS AND METHODS
Chemicals
Silica gel, alumina, and high-performance thin-layer chromatography (HPTLC) plates precoated with silica gel 60 F254 were purchased from Merck KGaA (Darmstadt, Germany). Ursolic acid (UA), oleanolic acid (OA), aucubin, 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), sodium pyruvate, glucose, phorbol 12-myristate 13-acetate (PMA), and lipopolysaccharide (LPS, O127:B8) were obtained from Sigma (St. Louis, MO, USA). Dimethyl sulfoxide (DMSO) and cytokine kit (MILLIPLEX® MAP Kit, product code HCYTOMAG-60K-05) were obtained from Merck KGaA (Darmstadt, Germany). RPMI-1640 medium, Modified Eagle Medium (MEM), fetal bovine serum (FBS), 2-mercaptoethanol, phosphate buffer saline, and penicillin-streptomycin solution were purchased from Gibco Life Sciences (GibcoThai, Thailand). All solvents used were analytical grade and obtained from Labscan Asia (Bangkok, Thailand).
Plant materials
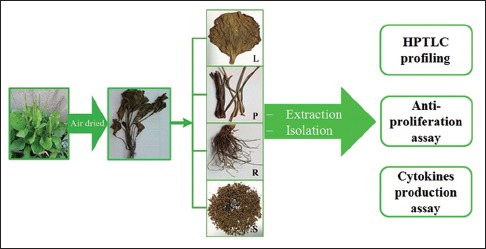
P. major samples were collected from two locations in Indonesia, i.e. Lumajang and Tawangmangu. Whole plant materials were harvested and cleaned with tap water. Lumajang sample was separated into leaves (L1), petioles (P1), roots (R1), and seeds (S1) which were used for bioactivity assays. The aerial part of the sample from Tawangmangu (A1) was used for isolation of the chemical compounds. Authentication of the plants (No. 1101/D.T/XI/2013) was conducted by Prof. Sutarjadi (Center of Information and Development of Herbal Medicine, University of Surabaya, Indonesia). All samples were dried and ground as described in previous work.[5]
Preparation of the extracts
P. major extracts were prepared according to the previous study.[5] Briefly, 50 g of plant materials was extracted twice by boiling in 500 ml distilled water for 1 h each. The extracts were filtered and concentrated in vacuo by a rotary evaporator (Buchi, Switzerland) and then were lyophilized to dryness. For the methanol extracts, 10 g of plant materials was macerated with methanol (3 × 100 ml, 3 × 24 h) at room temperature. The extracts were combined, filtered, and evaporated under vacuum to dryness. The aqueous and methanol extracts of leaves, petioles, roots, and seeds were obtained.
All crude extracts were determined for the antiproliferative activity against several cancer cells in comparison with doxorubicin, an anticancer drug. Moreover, the effects of the methanol extracts on cytokines production of THP-1 macrophages were also investigated.
High-performance thin layer chromatography profiling and isolation of the chemical compounds from Plantago major
High-performance thin layer chromatography profiling
To check the TLC profile of P. major extracts, chromatography was performed with two different solvent systems, i.e., (A) toluene: acetone: formic acid (78:22:0.15) and (B) 1,4 dioxane: xylene: propan-2-ol: 12.5% NH3 (1:2:5:2). Samples were spotted by Camag Linomat 5 sample applicator on the HPTLC plates precoated with silica gel 60 F254. Before development under the first solvent system (A), spotted samples were prederivatized with iodine vapor as described earlier.[5] Development was carried out with 10 ml of each solvent system in a Camag Twin Trough Chamber (10 cm × 10 cm) previously equilibrated with the mobile phase for 20 min at room temperature. After development, the plates were dried under warm air and subjected to derivatization by dipping in 5% sulfuric acid in methanol. TLC plates were further dried in a fume hood and then heated on a Camag TLC Plate Heater III (120°C, 3 min). The photographs were recorded by Camag Reprostar 3 under white light.
Isolation of the chemical compounds
Two kilograms of aerial part of the sample from Tawangmangu (A1) was extracted with methanol (6 × 10 L) by maceration at room temperature for 6 × 24 h. The combined extracts were then evaporated under vacuum to dryness (333 g). A portion (67 g) of this crude extract was subjected to a vacuum silica gel column chromatography eluted with n-hexane, dichloromethane, and methanol by increasing polarity. The like fractions were combined, evaporated in vacuo to dryness, and yielded five fractions (F1–F5). F4 and F5 were combined and further subfractionated in an alumina column and eluted by increasing polarity of ethyl acetate and methanol yielding 100 fractions. The combination of fractions 47–50 (325 mg) was then subjected to another silica gel column and development with ethyl acetate and methanol by increasing polarity afforded 177 subfractions. Subfraction 150 exhibited a single spot on a TLC plate at Rf value of 0.33 in the 1,4-dioxane: xylene: propane-2-ol:12.5%NH3 (1:2:5:2) solvent system. After recrystallization in methanol: ethyl acetate, the white crystalline compound was obtained. The identification of this isolated compound was confirmed by comparison of the spectral data of infrared (IR), proton nuclear magnetic resonance (1H-NMR), Carbon-13 nuclear magnetic resonance (13C-NMR), and electrospray ionization mass spectrometry (ESI-MS) with those in literature.
Antiproliferation assay on cancer cells
Cell culture
Human breast cancer (MCF-7, ATCC® HTB-22™ and MDA-MB-231, ATCC® HTB-26™), human cervical cancer (HeLaS3, ATCC® CCL-2.2™), human lung cancer (A549, ATCC® HTB-22™), and human nasopharynx carcinoma (KB, ATCC® CCL-17™) cell lines were obtained from the American Type Culture Collection (ATCC, Manassas, VA, USA). The cells were maintained in MEM or DMEM, supplemented with 10% heat-inactivated FBS, 100 U/ml penicillin and 100 mg/ml streptomycin, and incubated at 37°C in a humidified atmosphere containing 5% CO2 and 95% air.
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide cell proliferation assay
Cellular viability in the presence or absence of P. major extracts and three pure compounds, i.e. UA, OA, and aucubin, was examined using the MTT colorimetric assay as described previously.[19] Briefly, cancer cells under log phase of growth at the density of 1 × 103 cells/ml in 100 ml of culture medium were seeded onto 96-well culture plate (Costar, Cambridge, MA, USA). After 24 h of preincubation, various concentrations of the tested extracts or pure compounds were added and then incubated for a further 72 h. At the end of incubation, 20 μl of MTT solution (5 mg/ml) was added to each well and further incubated at 37°C for 3 h. The MTT formazan product was dissolved in DMSO and evaluated by measuring the absorbance at 550 nm using a microplate reader (Benchmark 550, Bio-Rad, USA).
Quantification of pro-inflammatory cytokines production in lipopolysaccharide-activated THP-1 macrophages
Cell culture and differentiation
THP-1 (human monocytic leukemia, ATCC® TIB-202™) cell line was grown in suspension in RPMI-1640 medium, supplemented with 0.05 mM 2-mercaptoethanol, 2 mM glutamine, 1 mM sodium pyruvate, 990 mg/L glucose, 10% FBS, and 100 U/ml penicillin-100 μg/ml streptomycin. The cells were maintained in a humidified incubator (5% CO2, at 37°C). Media were changed once every 3–4 days. Cells were disposed and changed by frozen cells every 20 passages.
The macrophage-like state was obtained by treating THP-1 monocytes for 48 h with 40 nM PMA in FBS-free medium at a density of 5 × 105 cells/ml in 96- and 24-well plates (100 μl and 1 ml, respectively). Furthermore, PMA-stimulated THP-1 cells (referred to as THP-1 macrophages) were washed with FBS-free medium and rested for another 24 h in the FBS-free medium and treatments were then initiated.
Cytotoxicity test
Cytotoxicity of P. major extracts and its pure compounds on THP-1 macrophages was assessed using the MTT method. THP-1 macrophages were incubated with a series concentration of extract (1–1000 μg/ml) in 96-well plate for 48 h. Cells were subsequently incubated with 100 μl of 1 mg/ml MTT in FBS-free medium for 3 h. After centrifugation (1200 rpm) at 4°C for 5 min, the medium was aspirated. The formazan product was dissolved in 100 μl DMSO, and the absorbance was further measured at 550 nm using a microplate reader. The absorbance of formazan formed in control cells (without extracts or pure compounds) was referred as 100% viability.
Multiplex quantification of pro-inflammatory cytokines production in lipopolysaccharide-activated THP-1 macrophages
THP-1 monocyte cells were plated at a density of 5 × 105 cells/ml in 24-well plate, differentiated with PMA, and subsequently incubated with the corresponding extracts or pure compounds in appropriate concentrations. One hour later, LPS was added to each well at the final concentration of 5 μg/ml and incubated for another 47 h. At the end of incubation, the plates were centrifuged (1200 rpm) for 5 min at 4°C. The supernatant in each well was aspirated and kept in −80°C until cytokines determination. TNF-α, IL-1β, IL-6, and IFN-γ concentrations in the supernatants were quantified simultaneously using MILLIPLEX® MAP kit according to the manufacturer's instruction (Merck KGaA, Darmstadt, Germany).
Statistical analysis
All values were presented as mean ± standard error mean or mean ± standard deviation, n = 2–3. For multiple variables comparison, data were analyzed by ANOVA followed by Dunnett's multiple comparison and Tukey's test when necessary using GraphPad Prism statistical software (GraphPad Software Inc., Sandiego, California, Windows Version 5.01). Differences were considered statistically significant at P < 0.05.
RESULTS
High-performance thin layer chromatography profiling and chemical structure of the isolated compound
To evaluate the chemical composition of P. major, HPTLC profiling of each extract was carried out using two different systems [Figure 2]. It was shown that all extracts contain UA (Rf value: 0.53), whereas OA (Rf value: 0.71) could be found in petioles and seeds only. Isolation of UA from the aerial part as well as determination of UA and OA concentration in different parts of P. major has been reported in our previous study.[5] Contrastingly, aucubin (Rf value: 0.39) was detected in all extracts [Figure 2b].
Figure 2.
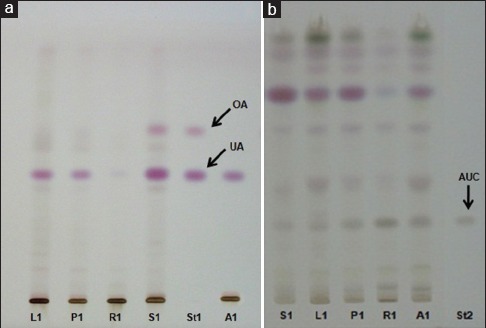
High-performance thin layer chromatography profile of Plantago major extracts. Solvent system (a) toluene: acetone: formic acid (78:22:0.15) and (b) 1,4 dioxane: xylene: propan-2-ol: 12.5% NH3 (1:2:5:2). L1, P1, R1, and S1 are methanol extracts of leaves, petioles, roots, and seeds of Plantago major from Lumajang, respectively. A1 is methanol extract of aerial part of Plantago major from Tawangmangu. St1 and St2 are ursolic acid-oleanolic acid and aucubin (AUC) standard, respectively
The isolated compound from this study was a white crystal with melting point 174–177°C, UV λmax (MeOH) 225.5 nm. IR nmax(KBr) 3289.83, 2914.38, 2882.50, 2359.80, 1652.67, and 1480.94 cm−1; ESI-MS m/z 369.1169 [M + Na]+; 1H NMR (DMSO-d5) d 4.99 (1H, m, H-1), 6.27 (1H, dd, H-3), 4.81 (1H, d, H-4), 2.49 (1H, m, H-5), 4.27 (1H, br s, H-6), 5.62 (1H, br s, H-7), 2.71 (1H, br t, H-9), 3.93 (1H, d, H-10A), 4.13 (1H, d, H-10B), 4.48 (1H, d, H-1’), 2.94–3.65 (1H, H-2’-6’B); 13C NMR (DMSO-d5) d 95.4 (C-1), 140.1 (C-3), 105.1 (C-4), 44.6 (C-5), 80.5 (C-6), 129.2 (C-7), 146.2 (C-8), 46.5 (C-9), 59.6 (C-10), 98.2 (C-1’), 73.5 (C-2’), 76.2 (C-3’), 70.2 (C-4’), 77.2 (C-5’), 61.1 (C-6’). These results were in concordance with those previously reported for aucubin.[20] Therefore, it can be concluded that the isolated compound was aucubin [Figure 3].
Figure 3.
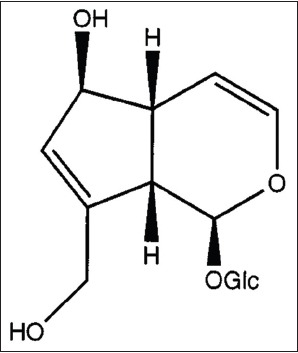
Chemical structure of aucubin
Effects of Plantago major extracts and its chemical compounds on cancer cells proliferation
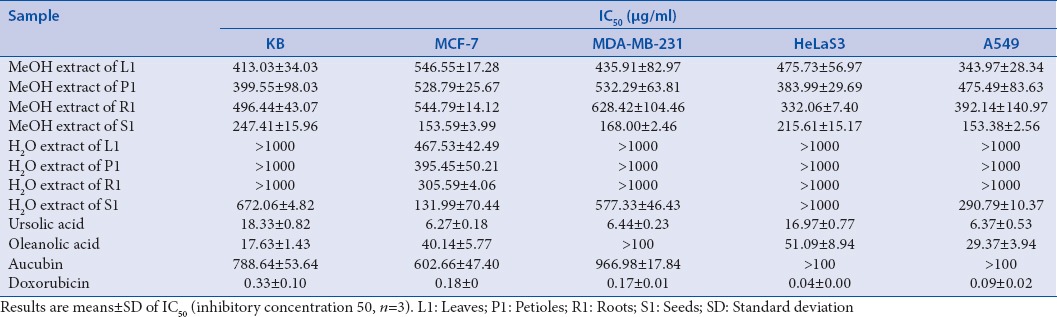
Antiproliferative activity of P. major extracts and its chemical compounds against various cancer cells [Table 1] was evaluated in vitro by the MTT colorimetric assay. Among the extracts tested, methanol extracts of P. major seeds exhibited the most potent antiproliferative activity against five cancer cell lines used with IC50 values between 153.38 and 247.41 μg/ml. In terms of pure compounds, UA showed the strongest antiproliferative activity against all cancer cell lines used with IC50 values between 6.27 and 18.33 μg/ml, followed by OA (IC50 values between 17.63 and >100 μg/ml), whereas aucubin had the lowest effects.
Table 1.
Antiproliferative effect of Plantago major extracts and its chemical compounds
Effects of Plantago major extracts and its chemical compounds on pro-inflammatory cytokines production in lipopolysaccharide-activated THP-1 macrophages
Cytotoxicity against THP-1 macrophage cells
In this study, methanol extracts of leaves, petioles, roots, and seeds of P. major were used for cytokines production assay. Before cytokines assay, potential cytotoxicity of the extracts and pure constituents on THP-1 macrophages was evaluated. Leaves, petioles, and seeds extracts reduced significantly cell viability at concentrations ≥300, 1000, and 100 μg/ml, respectively. However, root extracts did not show any cytotoxic effect under the concentrations applied. Both UA and OA reduced significantly cells viability at concentrations ≥30 μg/ml, whereas cytotoxicity due to aucubin was observed at 1000 μg/ml [Figure 4]. Considering this cytotoxic effect, the cytokines production assay was conducted at the concentrations having noncytotoxic effects.
Figure 4.
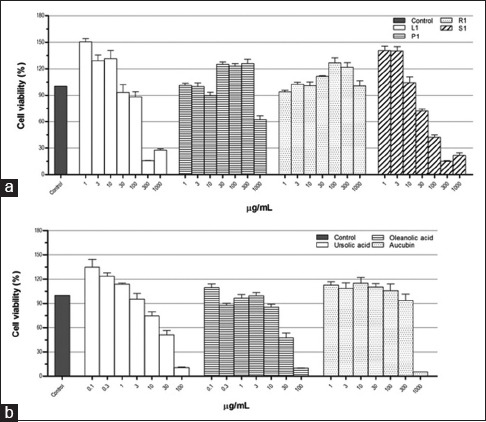
Cytotoxicity of Plantago major methanol extracts (a) and its chemical compounds (b) on THP-1 macrophages. Data represent the means ± standard error mean of three replications
Pro-inflammatory cytokines production in lipopolysaccharide-activated THP-1 macrophages
THP-1 macrophage cells were incubated with various concentrations of either extracts or pure compounds for 48 h in the presence of LPS. In these exposed cells, secretion of cytokines was analyzed and compared to LPS-stimulated cells. Figure 5 shows that treatment of leaves, petioles, and roots extracts at low concentrations increased the level of TNF-α; however, decreased it at high concentrations. Seeds extract slightly suppressed TNF-α production at both concentrations. UA, OA, and aucubin suppressed TNF-α production at all concentrations used, in which the effect of UA was in a dose-dependent manner.
Figure 5.
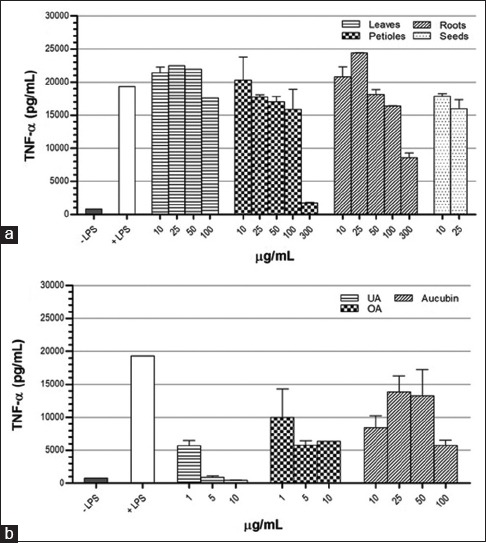
Effect of Plantago major methanol extracts (a) and its chemical compounds (b) on the production of tumor necrosis factor-α in THP-1 macrophages cells. Data represent the means ± standard error mean of two replications
Regarding IL-6 secretion [Figure 6], treatment of activated cells with any of P. major extracts increased this cytokine at low concentrations but inhibited its production at high concentrations. IL-6 secretion was reduced significantly by UA at all concentrations, and treatment with high concentration reached the basal production of nonactivated cells. In addition, the inhibition effect of UA was in a dose-dependent manner. OA and aucubin slightly decreased IL-6 secretion.
Figure 6.
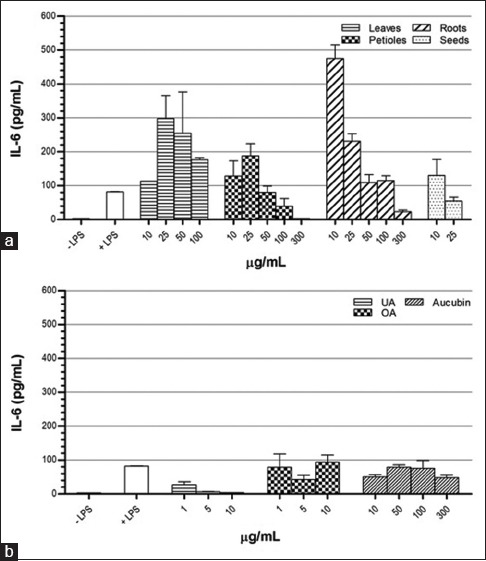
Effect of Plantago major methanol extracts (a) and its chemical compounds (b) on the production of interleukin-6 in THP-1 macrophages cells. Data represent the means ± standard error mean of two replications
IL-1β secretion [Figure 7] was stimulated when activated cells were incubated with leaves, petioles, and roots extracts. On the contrary, seeds extract slightly inhibited the production of IL-1β. All pure compounds tested decreased significantly IL-1β production.
Figure 7.
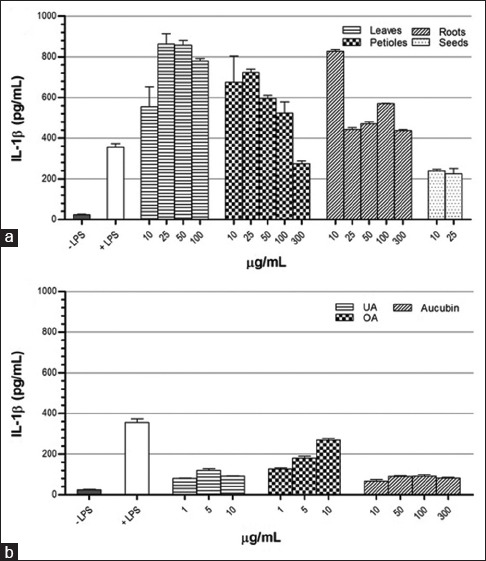
Effect of Plantago major methanol extracts (a) and its chemical compounds (b) on the production of interleukin-1β in THP-1 macrophage cells. Data represent the means ± standard error mean of two replications
In general, all of P. major extracts decreased the IFN-γ production [Figure 8]. The only one sample had a contrary effect, i.e., roots extract at the concentration of 100 μg/ml. Inhibition effect was observed as well as in UA, OA, and aucubin.
Figure 8.
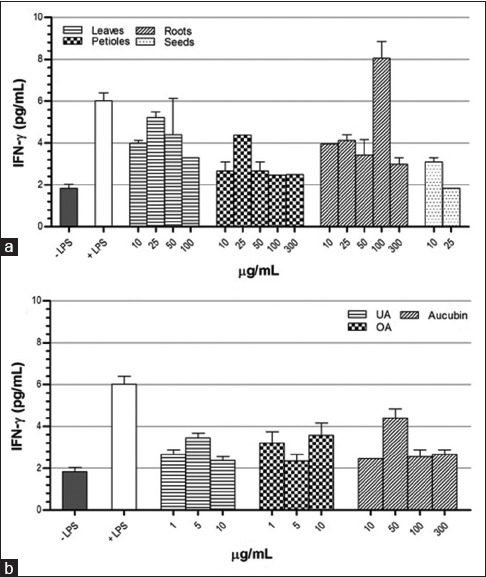
Effect of Plantago major methanol extracts (a) and its chemical compounds (b) on the production of interferon-γ. in THP-1 macrophage cells. Data represent the means ± standard error mean of two replications
DISCUSSION
P. major is one of the herbs widely used in cancer and pain relief management.[3] The quality of herbs may fluctuate depending on plant parts, growing environment, drying processes, solvents and methods of extraction, as well as many other postharvest factors.[21] In this study, we evaluated antiproliferative activity of P. major extracts and its chemical compounds against various cancer cell lines. We investigated as well as their effect on the production of inflammatory cytokines. P. major extracts were prepared from different plant parts using methanol and water as extracting solvents.
HPTLC profiles, Figure 2, exhibited that UA, OA, and aucubin are clearly found in P. major. Anticancer activity of UA and OA has been actively investigated in recent years.[22,23,24,25] Moreover, these two compounds along with aucubin have been known as anti-inflammatory agents.[26,27,28,29,30] In this recent work, we isolated aucubin from the aerial part of P. major, whereas the isolation of UA was reported in our previous study.[5]
In terms of antiproliferative activity, methanol extracts of all parts of P. major demonstrated cytotoxicities on five cancer cell lines in various potencies [Table 1], and the strongest activity was shown by the seeds extract (IC50 value was between 153.38 and 247.41 μg/ml). Water extract of P. major seed was active against KB, MCF-7, MDA-MB-231, and A549 cell lines, whereas the aqueous extracts of leaves, petioles, and root were active on MCF-7 only. These results were in concordance with the observations of Chiang et al. whose findings showed that aqueous extract of the whole plant of P. major showed cytotoxicities against various carcinoma cells with IC50 values ranged from 283 to 1809 μg/ml.[6] We found also that P. major roots had comparable cytotoxic effect with leaves and petioles. This finding supported our previous report[5] that roots could be included in the preparation of P. major derived herbal medicines. Further, toxicity and bioactivity tests are required to ensure its safety and efficacy.
Among the three pure chemicals utilized, UA showed the strongest cytotoxicity against five cancer cells tested, followed by OA. This finding confirmed our previous work which proposed UA as one of the active markers for quality assessment of P. major.[5] Contrastingly, aucubin did not express cytotoxic effect (IC50 >100 μg/ml). This finding was in agreement to Isiguro et al. whose investigation showed that aucubin did not have a cytotoxic effect toward leukemia P388, but its aglycone (i.e., aucubigenin) did.[31]
Cytokines are soluble messengers having a fundamental role for cell–cell communication and regulation within the immune system. Macrophages release cytokines to activate and recruit other cells during inflammation or as direct killing agents.[32] However, excessive production of this mediator in an inflammatory site may lead to chronic diseases such as cancer.[26] The effects of P. major extracts on the production of pro-inflammatory cytokines (TNF-α, IL-1 β, IL-6, and IFN-γ) were determined.
Studies have shown that P. major extracts were able to alter the production of inflammatory cytokines. Levels and patterns of inflammatory cytokines production were found to be different for different parts of P. major, which may be due to the content of chemical compounds. Leaves, petioles, and roots extracts showed a similar effect on TNF-α and IFN-γ secretions [Figures 5 and 8]. These samples enhanced the production of TNF-α at low concentrations but decreased it at high concentrations and decreased the level of IFN-γ at any concentrations. Leaves extract increased IL-6 at all concentrations, whereas petioles and roots increased this cytokine at low concentrations and decreased it at high concentrations [Figure 6]. Leaves and roots extracts induced production of IL-1β at all tested concentrations, whereas petioles extract increased its production at low concentrations and decreased it at high concentrations [Figure 7]. Our results seemed to be in concordance with the observations of Chiang et al. whose results revealed that P. major extracts enhanced the immune components at low concentrations, whereas inhibited these effect at high concentrations. Our findings support the concept of dual immunomodulatory activity owned by P. major.[6]
Seeds extract exhibited different patterns. It inhibited secretion of TNF-α, IL-1β, and IFN-γ in the range of concentrations used; however, it slightly increased IL-6 at low concentrations, whereas decreased it at high concentrations. If we compare among P. major parts at equivalent concentrations, seeds caused the lowest production of TNF-α, IL-1 β, IL-6, and IFN-γ. This effectiveness might be caused by its high content in UA[5] which showed the greatest inhibition activity among the chemical compounds tested.
Various trends in the modulation of cytokines levels were exhibited by the pure compounds. UA was capable of inhibiting the production of TNF-α, IL-6, IL-1 β, and IFN-γ. UA is a unique triterpenoid, having contrary anti- and pro-inflammatory activities that are dependent on the biological status of cells and tissues.[26] Our recent results, UA inhibited pro-inflammatory cytokines production in LPS-induced THP-1 macrophages, were in concordance with the previous study.[26]
We observed that OA and aucubin inhibited TNF-α, IL-1 β, and IFN-γ; however, they did not exhibit any effect on IL-6. These findings were in agreement with the previous work which showed that a series of OA analogs had weak inhibition toward IL-1β production in LPS-induced RAW 264.7 and J774A.1 cells.[33]
From this study, we observed that roots extracts possessed comparable antiproliferative and cytokines production activities to the other parts (leaves and petioles). This indicated that the roots could be included in the preparation of P. major derived herbal medicines. Seeds extract showed the highest antiproliferative effect as well as inhibition of inflammatory cytokines production. Among the pure constituents investigated, UA exhibited the strongest activities. Our previous study showed that UA was found abundantly in seeds, whereas very low in roots. Figure 2b showed that roots contain a high amount of aucubin. Therefore, in addition to UA,[5] we proposed aucubin as another marker for standardization of P. major.
CONCLUSIONS
The results of this study support the existing data and the empirical uses of P. major. Five cancer cell lines were used in antiproliferative assay to strengthen the usage of P. major in cancer management. Anti-inflammatory assay of P. major through modulation of cytokines production was carried out to afford its comprehensive mechanism. We have demonstrated that methanol extract of seeds possessed the greatest activity both on antiproliferative and cytokines production assay. Roots extract exhibited comparable activities to leaves and petioles. The seeds are being proposed as the main source for further development of anticancer and anti-inflammatory products. The roots could be included in the preparation of P. major derived products with respect to anti-inflammatory. It could also be considered as the source of aucubin.
Financial support and sponsorship
This study was financially supported by Directorate of Higher Education of Indonesia, contract number 373.11/E4.4/K/2012.
Conflicts of interest
There are no conflicts of interest.
Acknowledgement
This research is part of Ph.D. thesis of Mahidol University, Bangkok, Thailand, which was funded by the Directorate of Higher Education of Indonesia, contract number 373.11/E4.4/K/2012. The authors acknowledged the Natural Products Research Section, Research Division, National Cancer Institute, Bangkok, Thailand, for providing the facilities and equipment for doing throughout the research work.
REFERENCES
- 1.Beara IN, Lesjak MM, Jovin ED, Balog KJ, Anackov GT, Orcic DZ, et al. Plantain (Plantago L.) species as novel sources of flavonoid antioxidants. J Agric Food Chem. 2009;57:9268–73. doi: 10.1021/jf902205m. [DOI] [PubMed] [Google Scholar]
- 2.Sagar GR, Harper JL. Plantago major L., P. Media L and P. Lanceolata L. J Ecol. 1964;52:189–221. [Google Scholar]
- 3.Samuelsen AB. The traditional uses, chemical constituents and biological activities of Plantago major L. A review. J Ethnopharmacol. 2000;71:1–21. doi: 10.1016/S0378-8741(00)00212-9. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Warwick SI, Briggs D. The genecology of lawn weeds. V. The adaptive significance of different growth habit in lawn and roadside populations of Plantago major L. New Phytol. 1980;85:289–300. [Google Scholar]
- 5.Kartini, Piyaviriyakul S, Siripong P, Vallisuta O. HPTLC simultaneous quantification of triterpene acids for quality control of Plantago major L.and evaluation of their cytotoxic and antioxidant activities. Ind Crops Prod. 2014;60:239–46. [Google Scholar]
- 6.Chiang LC, Chiang W, Chang MY, Lin CC. In vitro cytotoxic, antiviral and immunomodulatory effects of Plantago major and Plantago asiatica. Am J Chin Med. 2003;31:225–34. doi: 10.1142/S0192415X03000874. [DOI] [PubMed] [Google Scholar]
- 7.Gálvez M, Martín-Cordero C, López-Lázaro M, Cortés F, Ayuso MJ. Cytotoxic effect of Plantago spp. on cancer cell lines. J Ethnopharmacol. 2003;88:125–30. doi: 10.1016/s0378-8741(03)00192-2. [DOI] [PubMed] [Google Scholar]
- 8.Lithander A. Intracellular fluid of waybread (Plantago major) as a prophylactic for mammary cancer in mice. Tumour Biol. 1992;13:138–41. doi: 10.1159/000217757. [DOI] [PubMed] [Google Scholar]
- 9.Ruffa MJ, Ferraro G, Wagner ML, Calcagno ML, Campos RH, Cavallaro L. Cytotoxic effect of Argentine medicinal plant extracts on human hepatocellular carcinoma cell line. J Ethnopharmacol. 2002;79:335–9. doi: 10.1016/s0378-8741(01)00400-7. [DOI] [PubMed] [Google Scholar]
- 10.Kolak U, Boǧa M, Uruşak EA, Ulubelen A. Constituents of Plantago major subsp. intermedia with antioxidant and anticholinesterase capacities. Turk J Chem. 2011;35:637–45. [Google Scholar]
- 11.Stanisavljević IT, Stojičević SS, Veličković DT, Lazić ML, Veljković VB. Screening the antioxidant and antimicrobial properties of the extracts from plantain (Plantago major L.) leaves. Sep Sci Technol. 2008;43:3652–62. [Google Scholar]
- 12.Gomez-Flores R, Calderon CL, Scheibel LW, Tamez-Guerra P, Rodriguez-Padilla C, Tamez-Guerra R, et al. Immunoenhancing properties of Plantago major leaf extract. Phytother Res. 2000;14:617–22. doi: 10.1002/1099-1573(200012)14:8<617::aid-ptr674>3.0.co;2-n. [DOI] [PubMed] [Google Scholar]
- 13.Núñez Guillén ME, da Silva Emim JA, Souccar C, Lapa AJ. Analgesic and anti-inflammatory activities of the aqueous extract of Plantago major L. Pharm Biol. 1997;35:99–104. [Google Scholar]
- 14.Türel I, Ozbek H, Erten R, Oner AC, Cengiz N, Yilmaz O. Hepatoprotective and anti-inflammatory activities of Plantago major L. Indian J Pharmacol. 2009;41:120–4. doi: 10.4103/0253-7613.55211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Dudhgaonkar S, Thyagarajan A, Sliva D. Suppression of the inflammatory response by triterpenes isolated from the mushroom Ganoderma lucidum. Int Immunopharmacol. 2009;9:1272–80. doi: 10.1016/j.intimp.2009.07.011. [DOI] [PubMed] [Google Scholar]
- 16.Beara IN, Orcic DZ, Lesjak MM, Mimica-Dukic NM, Pekovic BA, Popovic MR. Liquid chromatography/tandem mass spectrometry study of anti-inflammatory activity of plantain (Plantago L.) species. J Pharm Biomed Anal. 2010;52:701–6. doi: 10.1016/j.jpba.2010.02.014. [DOI] [PubMed] [Google Scholar]
- 17.Ringbom T, Segura L, Noreen Y, Perera P, Bohlin L. Ursolic acid from Plantago major, a selective inhibitor of cyclooxygenase-2 catalyzed prostaglandin biosynthesis. J Nat Prod. 1998;61:1212–5. doi: 10.1021/np980088i. [DOI] [PubMed] [Google Scholar]
- 18.Stenholm A, Göransson U, Bohlin L. Bioassay-guided supercritical fluid extraction of cyclooxygenase-2 inhibiting substances in Plantago major L. Phytochem Anal. 2013;24:176–83. doi: 10.1002/pca.2398. [DOI] [PubMed] [Google Scholar]
- 19.Buckberry LD. Cytotoxicity testing using cell lines. In: Jenkins N, editor. Animal cell biotechnology: Methods and protocols. Totowa, NJ: Humana Press; 1999. pp. 239–52. [Google Scholar]
- 20.Akdemir ZŞ, Tatli İİ, Bedir E, Khan IA. Acylated iridoid glycosides from Verbascum lasianthum. Turk J Chem. 2009;28:101–10. [Google Scholar]
- 21.Gad HA, El-Ahmady SH, Abou-Shoer MI, Al-Azizi MM. Application of chemometrics in authentication of herbal medicines: A review. Phytochem Anal. 2013;24:1–24. doi: 10.1002/pca.2378. [DOI] [PubMed] [Google Scholar]
- 22.Sun H, Fang WS, Wang WZ, Hu C. Structure-activity relationships of oleanane-and ursane-type triterpenoids. Bot Stud. 2006;47:339–68. [Google Scholar]
- 23.Liu J. Pharmacology of oleanolic acid and ursolic acid. J Ethnopharmacol. 1995;49:57–68. doi: 10.1016/0378-8741(95)90032-2. [DOI] [PubMed] [Google Scholar]
- 24.Liu J. Oleanolic acid and ursolic acid: research perspectives. J Ethnopharmacol. 2005;100:92–4. doi: 10.1016/j.jep.2005.05.024. [DOI] [PubMed] [Google Scholar]
- 25.Shanmugam MK, Dai X, Kumar AP, Tan BK, Sethi G, Bishayee A. Ursolic acid in cancer prevention and treatment: Molecular targets, pharmacokinetics and clinical studies. Biochem Pharmacol. 2013;85:1579–87. doi: 10.1016/j.bcp.2013.03.006. [DOI] [PubMed] [Google Scholar]
- 26.Ikeda Y, Murakami A, Ohigashi H. Ursolic acid: An anti- and pro-inflammatory triterpenoid. Mol Nutr Food Res. 2008;52:26–42. doi: 10.1002/mnfr.200700389. [DOI] [PubMed] [Google Scholar]
- 27.Jeong HJ, Koo HN, Na HJ, Kim MS, Hong SH, Eom JW, et al. Inhibition of TNF-alpha and IL-6 production by aucubin through blockade of NF-kappaB activation RBL-2H3 mast cells. Cytokine. 2002;18:252–9. doi: 10.1006/cyto.2002.0894. [DOI] [PubMed] [Google Scholar]
- 28.Park KS. Aucubin, a naturally occurring iridoid glycoside inhibits TNF-α-induced inflammatory responses through suppression of NF-κB activation in 3T3-L1 adipocytes. Cytokine. 2013;62:407–12. doi: 10.1016/j.cyto.2013.04.005. [DOI] [PubMed] [Google Scholar]
- 29.Park KS, Chang IM. Anti-inflammatory activity of aucubin by inhibition of tumor necrosis factor-alpha production in RAW 264.7 cells. Planta Med. 2004;70:778–9. doi: 10.1055/s-2004-827211. [DOI] [PubMed] [Google Scholar]
- 30.Park KS, Kim BH, Chang IM. Inhibitory potencies of several iridoids on cyclooxygenase-1, cyclooxygnase-2 enzymes activities, tumor necrosis factor-a and nitric oxide production in vitro. Evid Based Complement Alternat Med. 2010;7:41–5. doi: 10.1093/ecam/nem129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Isiguro K, Yamaki M, Takagi S, Ikeda Y, Kawakami K, Ito K, et al. Studies on iridoid-related compounds. IV. Antitumor activity of iridoid aglycones. Chem Pharm Bull (Tokyo) 1986;34:2375–9. doi: 10.1248/cpb.34.2375. [DOI] [PubMed] [Google Scholar]
- 32.Stow JL, Low PC, Offenhäuser C, Sangermani D. Cytokine secretion in macrophages and other cells: Pathways and mediators. Immunobiology. 2009;214:601–12. doi: 10.1016/j.imbio.2008.11.005. [DOI] [PubMed] [Google Scholar]
- 33.Bhandari P, Patel NK, Gangwal RP, Sangamwar AT, Bhutani KK. Oleanolic acid analogs as NO, TNF-α and IL-1ß inhibitors: Synthesis, biological evaluation and docking studies. Bioorg Med Chem Lett. 2014;24:4114–9. doi: 10.1016/j.bmcl.2014.07.056. [DOI] [PubMed] [Google Scholar]