

Abstract
The nuclear receptors (NRs) are ligand-dependent transcription factors that respond to various internal as well as external cues such as nutrients, pheromones, and steroid hormones that play crucial roles in regulation and maintenance of homeostasis and orchestrating the physiological and stress responses of an organism. We annotated the Fundulus heteroclitus (mummichog; Atlantic killifish) nuclear receptors. Mummichog are a non-migratory, estuarine fish with a limited home range often used in environmental research as a field model for studying ecological and evolutionary responses to variable environmental conditions such as salinity, oxygen, temperature, pH, and toxic compounds because of their hardiness. F. heteroclitus have at least 74 NRs spanning all seven gene subfamilies. F. heteroclitus is unique in that no RXRα member was found within the genome. Interestingly, some of the NRs are highly conserved between species, while others show a higher degree of divergence such as PXR, SF1, and ARα. Fundulus like other fish species show expansion of the RAR (NR1B), Rev-erb (NR1D), ROR (NR1F), COUPTF (NR2F), ERR (NR3B), RXR (NR2B), and to a lesser extent the NGF (NR4A), and NR3C steroid receptors (GR/AR). Of particular interest is the co-expansion of opposing NRs, Reverb-ROR, and RAR/RXR-COUPTF.
Keywords: RXR, phylogenetics, nuclear receptors, stress, endocrinology
Introduction
Fundulus heteroclitus (mummichogs) are an atlantic killifish species found from Nova Scotia, Canada to northern Florida, USA. In some estuaries, they can account for as much as 25% of the total macrofauna (1). Mummichog are noted for their hardiness, which includes the ability to survive a wide variety of temperatures, large salinity fluctuations, low dissolved oxygen, and heavily polluted ecosystems (1–7). The ability to adapt and acclimate to these different conditions has made mummichogs a popular research subject in a number of fields, including ecology, evolution, physiology, and toxicology (1,3,8–12).
Coastal population growth, urban runoff from increased impervious cover, and industrial pollution results in significant anthropogenic stress in estuaries. The abundance of F. heteroclitus in estuaries, their association with sediments due to hiding from predators or searching for food, and their non-migratory nature makes them an excellent bioindicator species. Fundulus species have been used as bioindicators at contaminated sites (1,2,5,13,14) and to follow the evolution of pollution resistance in a vertebrate (3,10,11,15–17). Thus, mummichogs are an excellent vertebrate bioindicator species for understanding rapid adaptation to environmental change in feral populations (5,8), including adaptations in transcription factors such as estrogen receptor alpha (ERα) splice variants (18), and evolutionary convergence in the aryl hydrocarbon receptor pathway (AHR1a, AHR2a, CYP1A) (5,8).
NRs consist of five modules: A/B, C, D, E, and F. The A/B module contains activation function-1 (AF-1) sites crucial in binding coactivators. The C module contains the DNA-binding domain (DBD) that encompasses the zinc-fingers necessary for binding DNA at response elements and mediating the basic transcriptional functions of a NR. The DBD is also highly conserved between orthologs and in turn is used in phylogenetic analysis (19,20). The D module contains the hinge region and nuclear localization sites. The E module contains the ligand-binding domain (LBD) domain and AF-2 function crucial in binding coactivators. This module mediates receptor activity by sensing the cellular environment, binding ligand and responding by activating transcription (21,22). The LBD is larger than the DBD, moderately conserved among the orthologs of different species, and therefore used in phylogenetic analysis (20,23). The F module is of unknown function and is missing in some NRs (21,24).
NRs regulate multiple physiological pathways such as cell differentiation, resource allocation, reproduction, development, and maintenance of homeostasis. They regulate these diverse physiological conditions by responding to both internal or external cues such as nutrients, steroids, heme, or xenobiotics (25–29). Thus, NRs are considered conduits that help in the maintenance of homeostatic conditions by responding to various internal and external cues. Thus, NRs occupy crucial roles in the disciplines of environmental physiology, endocrinology, pharmacology, toxicology, nutrition, biochemistry, gene regulation, ecology, chemistry, and other fields of study (30).
In this study, we annotated the F. heteroclitus NRs through phylogenetic analysis. F. heteroclitus are keystone species in the estuaries of the eastern seaboard of North America that are hardy and able to thrive in various conditions of toxic insult, salinity, and temperature (1). Therefore, we anticipate that analysis of the NRs will ultimately be useful in providing novel insight regarding their unique abilities to withstand diverse stressors and the subsequent transcriptional and physiological responses of these fish.
METHODS
Identification of Fundulus heteroclitus nuclear receptors
F. heteroclitus NRs were identified using a Basic Local Alignment Search Tool (BLAST) (31) with NR DNA binding domains (DBDs) from human, Takifugu rubripes, or D. magna (20) against the assembled F. heteroclitus genome (https://my.mdibl.org/display/FGP/Home) as described previously (22). Positive BLAST hits were confirmed as nuclear receptors with BLASTp on the NCBI database as were percent identity determinations between orthologous NRs (32).
Phylogenetics
Phylogenetic analysis was performed using methods described previously (20,33). The F. heteroclitus sequences are predicted protein sequences from the genome browser, and non-killifish sequences used for phylogenetic analysis were derived from the NCBI database, or the Takifugu genome brower (http://www.fugu-sg.org/). Phylogenetic comparisons included F. heteroclitus, Takifugu rubripes, Danio rerio, and Homo sapiens. A list of nuclear receptors used in comparisons to F. heteroclitus is provided in Additional File 1.
Phylogenetic analysis was performed using only the highly conserved DBD and moderately conserved LBD of each receptor. These domains were identified and isolated using the pfam00105 (Zf-C4) and pfam00104 (hormone receptor) designations on the conserved domain database CDD (34) as described previously (20,23,35). The DBD and LBD from 246 NRs from three fish species and humans were aligned using ClustalX default parameters (36) (Additional File 2). Phylogenetic analysis was performed with Maximum Likelihood (ML) using MEGA 6.0 (37). “Find Best Model” was used to determine the parameters for Maximum Likelihood. Further analysis was performed using the Bootstrap method with 500 replications, the JTT model with Gamma distributed rates among sites (3). Tree inference options included SPR level 3, BIONJ with a very strong branch filter.
Maximum parsimony and distance parameters were used to provide additional support for the phylogenetic relationships observed. Distance parameters were measured using PAUP 4.0b10 with default characteristics (mean character difference and among site rate variation), and full heuristic searches. Branch support was measured by bootstrap analysis with 1000 replicates. Parsimony was constructed using PAUP version 4.0b10 with heuristic searches, tree-bisection-reconnection, topological constraints not enforced, and multiple tree option in effect with an initial maximum tree setting at 100,000. Branch support was measured by bootstrapping with 10,000 replicates (20,38). Maximum Parsimony trees were visualized with PAUP 4.0b10 and Neighbor-Joining trees were visualized with FigTree (http://tree.bio.ed.ac.uk/software).
RESULTS and DISCUSSION
Nuclear receptor isolation from Fundulus
Analysis of the F. heteroclitus genome found 74 NRs, which include representatives of all seven subfamily members. Additional File 3 provides links to the scaffold for each NR, its cDNA and its protein sequence. Seventy-four NRs are in range of what has been found in other fish species as 68 NRs were found in Takifugu rubripes (39) and 72 NRs in Tetraodon nigriviridis (40) although recent improvements in the Takifugu genome indicates 73 NRs (23). The number of NRs in teleost genomes is significantly greater than those found in invertebrates and mammals because of whole genome duplication events (41). The number of NRs in the common carp (Cyprinus carpio) is even greater (137 receptors) than other fish species because of an additional genome duplication event (23). Genome duplication is often followed by loss of some genes, gain of function of some genes (neofunctionalization) including differences in spatial or temporal gene expression between paralogs, or partitioning of ancient functions on duplicated genes (subfunctionalization) (42).
Phylogenetics
Phylogenetic analysis by Maximum Likelihood (ML) confirms the presence of all seven NR subfamilies in F. heteroclitus, and demonstrates that there are 2 NR0 members, 30 NR1 members, 17 NR2 members, 15 NR3 members, 5 NR4 members, 4 NR5 members, and 1 NR6 member (Fig. 1). This file is also available as an expandable pdf (Additional File 4). The percentage of each subfamily of NRs is relatively similar between F. heteroclitus, the fish species examined and humans with minor exceptions. The three different phylogenetic models used (Maximum Likelihood, Maximum Parsimony, Neighbor-Joining) agreed at the group level and often at the subfamily level, but there are differences at the base of the phylogram; primarily the relationship of the 0-subfamily to the 5 and 6 subfamilies. However, the ancient bootstrap values are typically not significant using any of the analysis with Maximum Parsimony rarely being able to resolve distinct clades on the left hand side of the tree. The Neighbor-Joining and Maximum Parsimony phylogenetic trees are provided as additional files (Additional Files 5, 6).
Figure 1. Phylogenetic relationship of nuclear receptors as determined by ML.
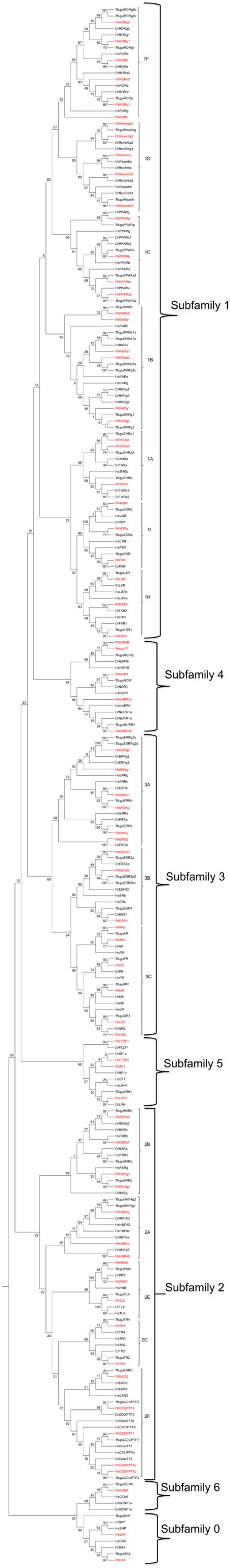
The phylogenetic tree is shown with bootstrap support values (frequency of occurrence) from ML at each node. Species included are Takifugu rubripes (Tfugu), H. sapiens (Hs), Danio rerio (Dr), and Fundulus heteroclitus (Fh). All F. heteroclitus sequences are in red.
NR0 Subfamily
The NR0 subfamily contains two groups, NR0A and NR0B. NR0A receptors lack a ligand-binding domain (LBD) and NR0B receptors lack a DNA-binding domain (DBD). Similar to other vertebrates, there are no NR0A members (knirps) in F. heteroclitus (20,43). Mummichogs contain two NR0B members, SHP and DAX (Fig. 1; Table 1). We found that DAX/SHP fall within the 6 subfamily based on Maximum Likelihood but with low posterior probabilities (50%)(Fig. 1). Maximum parsimony resolved the 2, 6, and 0 subfamilies as separate subfamilies but without resolving their evolutionary relationship to each other (Additional File 5). Previous work using parsimony and NJ suggested with 73% confidence that the NR0B group is evolutionarily related to 2 subfamily (44). Bayesian Inference also indicated that these NRs evolved as part of the 2 subfamily (22). However, most studies performed do not include them as part of a phylogenetic tree because of the reduced molecular information due to the lack of the LBD and subsequent uncertainty in the analysis (20,45,46).
Table 1.
Nuclear receptors from Fundulus compared to those described in other species.
| Group | F. heteroclitus | H. sapiens | D. rerio | T. rubripes | T. nigroviridis | D. pulex | D. melanogaster |
|---|---|---|---|---|---|---|---|
|
| |||||||
| 0A | KNR-R1 | KNI | |||||
| KNR-R2 | KNRL | ||||||
| EGON | |||||||
|
| |||||||
| 0B | DAX | DAX | DAX | DAX | |||
| SHP | SHP | SHP | SHP | ||||
|
| |||||||
| 1A | THRα1 | THRα | THRα | THRα1 | THRα1 | THRL11 | |
| THRα2 | THRα2 | THRα2 | |||||
| THRβ | THRβ | THRβ1 | THRβ | THRβ | |||
| THRβ2 | |||||||
|
| |||||||
| 1B | RARα1 | RARα | RARα | RARα1a | RARα1 | RARL10 (unresolve d; may be member of 1M family) | |
| RARα2 | RARα1b | RARα2 | |||||
| RARα2a | |||||||
| RARα2b | |||||||
| RARβ1 | RARβ | RARβ | |||||
| RARβ2 | |||||||
| RARγ1 | RARγ | RARγ1 | RARγ1 | RARγ | |||
| RARγ2 | RARγ2 | RARγ2 | |||||
| RARγ3 | |||||||
|
| |||||||
| 1C | PPARα1 | PPARα | PPARα1 | PPARα1 | |||
| PPARα2 | PPARα2 | PPARα2 | PPARα2 | ||||
| PPARβ | PPARβ | PPARβ1 | PPARβ | PPARβ | |||
| PPARβ2 | |||||||
| PPARγ | PPARγ | PPARγ | PPARγ | PPARγ | |||
|
| |||||||
| 1D | Rev-erb-α | Rev-erb-α | Rev-erb-α | Reb-erb-α | Rev-Erb-α1 | E75 | E75 |
| Rev-Erb-α2 | |||||||
| Rev-erb-β1 | Rev-erb-β | Rev-erb-β1 | Rev-erb-β | Rev-erb-β1 | |||
| Rev-erb-β2 | Rev-erb-β2 | Rev-erb-β2 | |||||
| Rev-erb-γ1 | Rev-erb-γ1 | Rev-erb-γ | Rev-erb-γ | ||||
| Rev-erb-γ2 | Rev-erb-γ2 | ||||||
|
| |||||||
| 1E | E78 | E78 | |||||
|
| |||||||
| 1F | RORα1 | RORα | RORα1 | RORα | RORα1 | HR3 | DHR3 |
| RORα2 | RORα2 | RORα2 | |||||
| RORβ | RORβ | RORβ | RORβ | RORβ | |||
| RORβ | |||||||
| RORγ1 | RORγ | RORγ1 | RORγ1 | RORβ | |||
| RORγ2 | RORγ1b | RORγ2a | |||||
| RORc | RORγ2b | ||||||
|
| |||||||
| 1H | LXRα | LXRα | LXRα | LXRα | LXRα | EcRα | EcR |
| LXRβ | EcRβ | ||||||
| FXR1 | FXR | FXR1 | FXR1 | FXRa | |||
| FXR2 | FXR2 | FXR2 | FXRa | ||||
| FXRb | |||||||
|
| |||||||
| 1I | VDR-A | VDR | VDR-A | VDR-A | VDR-A1 | ||
| VDR-B | VDR-B | VDR-A2 | |||||
| VDR-B | |||||||
| PXR | PXR | PXR | PXR | PXR | |||
| CAR | |||||||
|
| |||||||
| 1J | HR96 | DHR96 | |||||
|
| |||||||
| 1L | HR97a | ||||||
| HR97b | |||||||
| HR97g | |||||||
|
| |||||||
| 2A | HNF4α | HNF4α | HNF4α | HNF4α | HNF4α | HNF4 | HNF4 |
| HNF4β | HNF4β | ||||||
| HNF4γ | HNF4γ | HNF4γ | HNF4γ1 | HNF4γ | |||
| HNF4γ2 | |||||||
|
| |||||||
| 2B | RXRα | RXRα | RXRα | RXRα | RXR | USP | |
| RXRβ-p | RXRα | ||||||
| RXRβ1 | RXRβ | RXRβ1 | RXRβ | RXRβ | |||
| RXRβ2 | RXRβ2 | RXRβ | |||||
| RXRγ1 | RXRγ | RXRγ | RXRγ | RXRβ | |||
| RXRγ2 | |||||||
|
| |||||||
| 2C | TR2 | TR2 | TR2 | TR2 | TR2 | ||
| TR4 | TR4 | TR4 | TR4 | TR4 | |||
|
| |||||||
| 2D | HR78 | DHR78 | |||||
|
| |||||||
| 2E | TLX | TLX | TLX | TLX | TLX | TLL | TLL |
| PNR 1 | PNR | PNR | PNR | PNR | PNR | PNR | |
| PNR 2 | DSF | DSF | |||||
| NR2E6 | FAX-1 | ||||||
|
| |||||||
| 2F | COUP-TF1 | COUP-TFa | COUP-TF1a | COUP-TF1 | COUP-TFa | SVP | SVP |
| COUP-TF1b | COUP-TF1 | COUP-TFa | |||||
| COUP-TF2a | COUP-TFb | COUP-TF2 | COUP-TF2 | COUP-TFb | |||
| COUP-TF2b | COUP-TFb | ||||||
| COUP-TF3 | COUP-TF3 | COUP-TF3 | COUP-TFg | ||||
| EAR2 | EAR2 | EAR2 | EAR2 | ||||
| EAR2 | EAR2 | EAR2 | |||||
| EAR2 | |||||||
|
| |||||||
| 3A | ESR1 | ESR1 | ESR1 | ER1 | ESR1 | ||
| ESR2a | ESR2 | ESR2a | ERb | ESR2a | |||
| ESR2b | ESR2b | ER2a | ESR2b | ||||
| ER2b | |||||||
|
| |||||||
| 3B | ERRα | ERRα | ERRα | ERRα | ERRα | ERR | ERR |
| ERRβ1 | ERRβ | ERRβ | ERRβ | ERRβ1 | |||
| ERRβ2 | ERRβ2 | ||||||
| ERRδ | ERRδ | ||||||
| ERRγ1 | ERRγ | ERRγ1 | ERRγ2a | ERRγ | |||
| ERRγ2 | ERRγ2 | ERRγ2b | ERRγ | ||||
| ERRγ | ERRγ | ||||||
| ERRγ | |||||||
|
| |||||||
| 3C | GR | GR | GR | GR | GR | ||
| GR2 | GR2 | GR2 | |||||
| MR | MR | MR | MR | MR | |||
| PR | PR | PR | PR | PR | |||
| ARα | AR | AR | ARα | ARα | |||
| ARβ | ARβ | ARβ | |||||
|
| |||||||
| 4A | NGF1B | NGFIB | NGFIB | NGFIBa | NGF1B | HR38 | DHR38 |
| Nur77 | NGF1B | ||||||
| NURR1a | NURR1 | NURR1a | NURR1 | NURR1 | |||
| NURR1b | NURR1b | ||||||
| NOR1 | NOR1 | NOR1 | NOR1 | NOR1 | |||
|
| |||||||
| 5A | SF1 | SF1 | SF1a | FTZ-F2 | SF1 | FTZ-F1 | FTZ-F1 |
| FTZ-F2/SF1b | SF1b | ||||||
| LHR1 | LRH1 | LRH1 | LRH1 | LRH1 | |||
| FTZ-F1 | FTZF1 | FTZF1 | FTZ-F1 | ||||
|
| |||||||
| 5B | DpHR39 | DHR39 | |||||
|
| |||||||
| 6A | GCNF | GCNF | GCNF1a | GCNF | GCNF | DpHR4 | DHR4 |
| GCNF1b | |||||||
Overall, the resolution of the 0 subfamily is poor. NR0B members clearly form their own group based on the lack of a C domain; however, their evolutionary relationship to other NRs is questionable. In turn, unlike other receptors, NR0 members are not named for their phylogenetic position but instead for their lack of a key domain (47). The NR0B receptors SHP and DAX primarily work as co-regulators as they both contain LxxLL domains typically found in coregulators (48–50). DAX is involved in reproductive development and steroidogenesis; SHP regulates cholesterol and glucose homeostasis (51).
NR1 Subfamily
The NR1 subfamily is the largest subfamily in F. heteroclitus with 30 members. 40.5 Percent of mummichog NRs are in the 1-subfamily; 36–40% of NRs are in the 1-subfamily of other fish species and 40% of NRs are 1 subfamily members in humans. The 1-subfamily separates into two distinct clades; one clade includes the NR1B (RAR), NR1C (PPAR), NR1D (Reverb), and NR1F (ROR) groups, and the other one includes the NR1A (THR), NR1H (LXR/FXR), and NR1I (VDR) groups (Fig. 1). There is disagreement in some of our different phylogenetic analysis at the extreme left of the trees where bootstrap values may drop as low as 23 indicating poor resolution.
The fish species, including F. heteroclitus show significant expansion of NR1B (RAR), NR1D (Rev-erb), and NR1F (ROR) groups relative to humans (Table 1). Interestingly, all of these NRs are found in the same NR1 clade. There are 6 members of the F. heteroclitus RAR group. Humans have 3 members and there are 3 – 7 RAR members in the other fish species investigated. There are 5 members of the Rev-erb clade and 6 members of the ROR groups in F. heteroclitus. Five members in the Rev-erb group are typical in fish; however, the other fish species examined only have 5 ROR members. In comparison, humans have 2 Rev-erb and 3 ROR members (Table 1).
The RAR, Rev-erb, and ROR groups are important in lipid and glucose metabolism, gas-response, development, promoting T-cell differentiation, inflammation, and circadian rhythms (52–54). Several genes are coordinately regulated by Rev-erbα and RORα as they share the same response elements but exert opposing effects on transcription. Significant expansion of these two opposing sets of NRs in fish species including F. heteroclitus appears synchronous. It is thought that the crosstalk between the two receptor groups helps regulate their transcriptional and physiological networks, including circadian rhythms, lipid and glucose metabolism, and inflammation (55,56).
In contrast, the NR1C (PPAR) group shows little expansion in F. heteroclitus or other fish species. There are three members in humans and four in each of the fish species investigated (Table 1). PPARs are crucial in the regulation of lipids (27,57). The pufferfish (Takifugu and Tetraodon) genomes both show two PPARα NRs similar to F. heteroclitus, while the Atlantic salmon (Salmo salar) and zebrafish (D. rerio) genomes have two members of the PPARβ/δ group (41,58,59). Danio are ancient Ostariophysi, Salmo are Salmoniformes, and Fundulus and Takifugu are both modern Percomorphs (60). Because of whole genome duplication in teleosts (42), it is most likely that an ancient relative lost PPARβ/δ in the Percomorphs and PPARα in the early teleosts. However, it cannot be completely ruled out that separate duplication events occurred in the Percomorphs and early teleosts as individual gene duplication events have occurred multiple times including cytochrome P450s, opsins, and NRs (20,35,61–63). The organ distribution of PPARs differs from mammals. For example, pufferfish show wide tissue distribution of the PPARs; in mammals only PPARβ/δ is widely distributed (58). Changes in organ distribution or domain structure and function is common in neofunctional retained duplicated receptors. Therefore, we performed pairwise comparisons between human NRs and their F. heteroclitus orthologs with duplications (Table 2). Mummichog PPARα2 shows greater differences in its LBD than PPARα1 compared to human PPARα suggesting neofunctionalization (Table 2).
Table 2.
Percent Identity comparisons of the DNA binding domain (DBD) and ligand binding domain (LBD) of orthologous human and F. heteroclitus nuclear receptors with Blastp.
| Nuclear Receptora | DBD | LBD | Ligandsb |
|---|---|---|---|
| SHP | 51 | ||
| DAX | 51 | ||
| THRα1-110 | 92 | 84 | Thyroid hormones (114) |
| THRα2-10098 | 96 | 91 | |
| PPARα1 | 86 | 85 | Leukotriene B4, 1-palmityl-2-oleolyl-sn-glycerol-3-phosphocholine, Fatty acids (115) |
| PPARα2 | 86 | 73 | |
| Rev-Erb-β1-10090 | 94 | 83 | Heme (114) |
| Rev-Erb-β2-10062 | 93 | 77 | |
| RORα-9958 | 99 | 93 | |
| RORα-369 | 91 | 75 | |
| RORγ-9885 | 90 | 43 | |
| RORγ-10005 | 90 | 47 | |
| FXR1-10028 | 96 | 72 | 5β-bile acids (83) |
| FXR2-9861 | 84 | 35 | |
| VDR-A | 93 | 79 | 1,25-OH vitamin D3, bile salts (116,117) |
| VDR-B | 94 | 80 | |
| PXR | 72 | 58 | Xenobiotics, bile salts (73,80) |
| RXRβ1-483 | 94 | 88 | 9-cis retinoic acid, rexinoids (114,118) |
| RXRβ2-9867 | 90 | 79 | |
| RXRγ1-9885 | 94 | 95 | |
| RXRγ2-9880 | 94 | 95 | |
| PNR1-9949 | 93 | 74 | |
| PNR2-195 | 91 | 59 | |
| ESR1 | 96 | 66 | 17β-estradiol (119) |
| ESR2α – 10024 | 96 | 68 | |
| ESR2β – 10026 | 96 | 65 | |
| ERRβ-9910 | 100 | 76 | |
| ERRβ-3 | 100 | 84 | |
| ERRγ _10140 | 97 | 94 | |
| ERRγ2 _9925 | 91 | 81 | |
| GR - 0 | 96 | 74 | Cortisol (120) |
| GR2 -10031 | 96 | 74 | |
| ARα-9941 | 65 | 59 | Testosterone, 11-ketotestosterone, 17α-methyltestosterone (121) |
| ARβ-9884 | 91 | 70 | |
| NGF1B | 99 | 69 | |
| Nurr77 | 93 | 66 | |
| NURR1a-9995 | 96 | 95 | |
| NURR1b-114 | 97 | 58 | |
| SF1a | 72 | 58 | Phosphatidylinositol, phosphatidylcholine (122) |
| FTZ-F2/SF1b-224 | 84 | nd |
Scaffold is often provided after the receptor to aid in determining the receptor in question.
Known or putative endogenous physiological ligands for fish (or human if fish unknown) are included for the receptors that show show high divergence from humans or have a duplicated receptor with potential neofunctionalization.
The presence of PPARs in fish may make them sensitive to peroxisome proliferation or perturbations in lipid allocation and homeostasis. For example, peroxisome proliferation or induction of biomarkers of peroxisome proliferation has been measured in fish exposed to PAHs, pharmaceuticals, phthalates, alkylphenols, and pesticides (64–69). Peroxisome proliferation has been measured in mummichogs following 2,4-D exposure (70).
Furthermore, pharmaceuticals such as the PPARα activator, gemfibrozil reduced n-3 fatty acids in rainbow trout (Oncorhynchus mykiss), which may reduce the nutritional quality of the fish, perturb their ability to acclimate to changes in water temperature, and repress their ability to reproduce following migration (64). The PPARγ activator, TBT activated an obesogen response by increasing body weight and whole-lipid content in Chinook salmon (Oncorhynchus tshawytscha) and condition factor, triglycerides, and hepatosomatic index in zebrafish (71). Interestingly, toxicants that activate PPARs may enhance condition factor while also increasing other stress responses typically associated with poor physiological conditions (72).
The other NR1 clade includes the NR1A (THR), NR1H (LXR/FXR), and NR1I (VDR/PXR) groups. THR is split into two subgroups, THRα and THRβ. There are three THR members in each of the fish species examined compared to two in humans that only contain one THRα and THRβ member. Pufferfish and mummichogs contain two THRα members and one THRβ members; D. rerio contains one THRα member and two THRβ members (Table 1).
The NR1I group in fish that contains the vitamin D receptor (VDR), PXR, and CAR, has been relatively well studied because of the roles of PXR and CAR in acclimation to foreign chemicals (21,73). PXR was previously cloned in F. heteroclitus from a PCB polluted site and this PXR is nearly identical to the PXR sequenced during the genome project with the exception of a three amino acid region at amino acid 190 that is missing in the F. heteroclitus genome sequence (74).
PXR, but not CAR, has been identified in teleost species (39,75). Originally it was thought that CAR diverged from PXR at some later evolutionary point because CAR had only been found in mammals. However, recent phylogenetic data, examination of Saurapsid genomes such as reptiles and birds (76) and the lobe-finned fish Coelocanth indicates that CAR was lost in teleosts and PXR was lost in reptiles and birds (77). Therefore, it is thought that the lobe-finned fish that arose 400 million years ago during the Devonian period and are the ancestors of all tetrapods, contained CAR and PXR (78,79).
The DBD and LBD of F. heteroclitus PXR are highly diverged from humans with pairwise comparisons showing 72% identity to human DBD and 58% to human LBD (Table 2). This is typical of fish PXRs as previous comparisons to human PXR show 61–73% sequence identity to the DBD and 52–57% identity to the LBD (78). Only ARα (65%) and SF1a (72%) show as much divergence in the DBD as PXR; only RORγ, SF1, FXR2, SHP, and DAX show similar divergence within the LBD when compared to mammals (78). However, with the exception of the NR0 members, SHP and DAX, all the other receptors showing high divergence have duplicated members and at least one of the duplicates appears to show neofunctionalization based on the large change in amino acid identity (Table 2).
The large differences in LBD sequences for PXR provides biochemical support for the significant differences in activation profiles between different fish species and between fish and humans, especially for fenvalerate and several organochlorine insecticides (73). Ligand activation of zebrafish PXR matched only 30% of the representative human PXR ligands further indicating the divergence of fish and human PXR (75). Ligands tested include primarily bile acids, steroids and pharmaceuticals. Of these, Phenobarbital, clotrimazole, dihydrotestosterone, androstanol and 5β-pregnane-3,20-dione all activated the zebrafish PXR. In general, human PXR can be activated by many bile salts, including both C24 and C27 bile salts. However, the zebrafish PXR was only activated by C27 bile salts with a strong preference for sulfated bile alcohols (80). The C27 pathway is an ancient pathway found in early fish and amphibians and it has been hypothesized that PXR has evolved to deal with the increasingly complex bile salt synthetic pathways (76). This is in contrast to other theories that suggest that the reason for the great divergence in the LBD of PXR between species, including zebrafish and Fugu, is in response to the different sets of xenobiotic/environmental challenges encountered by the different species (81).
The FXR/LXR group (NR1H) in F. heteroclitus contains one LXR member, LXRa, and two FXR members. The FXR/LXR group is related to the ecdysone receptors in invertebrates (Table 1)(20,82). Humans have two LXR members and one FXR that is more closely aligned with FXR1 (Fig. 1). The existence of two or more FXR members is common among fish species. The green pufferfish (T. nigroviridis) and Japanese medaka (Oryzias latipes) have at least 3 FXR members because they have two similar FXRa members (Table 1; Fig. 1). In medaka, FXRa2 is activated by C24 bile acids and GW4064, but FXRa1 is not activated by these common ligands probably due to differences in the A/B domain (83). Interestingly, in Fundulus, FXR2’s LBD has the lowest percent identity when compared to humans (Table 2), indicating neofunctionalization and a different activation profile than humans.
In addition, there is significant diversity in FXR ligand responses in fish species (84). Tetraodon FXR has a ligand selectivity profile very similar to human FXR, with activation by the synthetic ligand GW4064 and the bile acid, chenodeoxycholic acid. Furthermore, modeling and docking studies suggest that Tetraodon’s ligand-binding pocket more similar to mammalian FXRs than to lamprey or zebrafish FXRs (85), which are activated by 5α-bile alcohols but not by the evolutionarily more recent 5β-bile acids (86). Based on phylogenetics, Fundulus FXR is more like Tetraodon and medaka than zebrafish (Fig. 1)(78).
NR2 Subfamily
The NR2 subfamily is the second largest subfamily of NRs in F. heteroclitus with 18 NRs. NR2 is divided into 5 groups A, B, C/D, E and F. Of these, Group 2B (RXR) has expanded the most of the fish species investigated relative to humans, and group 2F (COUP) has expanded in all of the fish species investigated relative to human NRs (Table 1). In addition, F. heteroclitus has three Group 2E members, which are involved in eye development in zebrafish (87,88), because it has two PNR receptors (Table 1). To our knowledge, other fish species only have one PNR with the recent exception of Nile tilapia (Oreochromis niloticus) and medaka (23,78). Considering the phylogenetic distance of the receptors it is unlikely that this is a recent event (Fig. 1) and instead it is more likely that the second PNR was not lost following the genome wide duplication in fishes (41). Interestingly, invertebrate species often contain more group 2E members than vertebrates (Table 1)(20).
RXRs bind to retinoids, are crucial for growth and development, and are key heterodimeric partners with several other NRs (89,90). COUP-TFs are necessary for growth and development, including venous and lymphatic development in zebrafish (91,92). Interestingly, COUP-TF members have also been shown to interact with a few other NRs including RXRα, RAR, THR, ERRγ, ERα, and other COUP-TF members as hetero- and homodimers (90); however they typically repress transcription including RAR/RXR responses (93). Thus, the NRs that have shown expansion may have a repressive counterpart that also expanded.
There are three RXR members in humans and 4–5 members in F. heteroclitus of which none appear to be RXRα members. The exact number of RXR members is unknown because of the brevity of scaffold 2083 (Additional File 3), which ends at 17,995 bp and in turn cuts off a potential RXR member at the AB domain (AF-1). Therefore, this RXR member appears to be a pseudogene but may not be. This partial gene contains a start site, two introns, and a stop codon. It is also structurally different than the other RXRβ genes that have two relatively close exons near the 5′-end. However given that the scaffold ends shortly after the stop codon it cannot be completely ruled out that there might be other splice sites and exons following. Interestingly, scaffold’s 2083 gene fragment aligns well with RXRβ members from several fish species. If it is a RXRβ member than phylogenetic analysis would indicate that F. heteroclitus has 3 RXRβ, 2 RXRγ, and 0 RXRα members (Table 1; Fig 1). Initial examination of the genome by BLAST suggested that the RXR at scaffold 9880 may be a RXRα member; however phylogenetic analysis indicates that this NR is an RXRγ member with a weak posterior probability of 37. While the posterior probability is weak, Maximum Likelihood (Fig. 1), Maximum Parsimony (Additional File 5), and Neighbor-Joining (Additional File 6) all agree that complete RXR members are found within RXRβ or RXRγ subgroups and therefore at this time no RXRα member was found in F. heteroclitus. If the RXR at scaffold 2380 is a complete RXRα member than the F. heteroclitus genome will have at least 75 NRs. The lack of an RXRα member would be surprising if not unprecedented as we did not find other fish or mammalian species lacking RXRα.
Group 2A (HNF) is an ancient group (46) that expanded greatly in C. elegans (94). HNF4 contains 3 members, which is typical of most fish species (Table 1). The HNF group of receptors has been highly studied in fish species, but what work has been done indicates that similar to mammals the HNF4 receptors are enrich in the liver and regulate liver enriched gene expression (95) sometimes in conjunction with other HNFs including HNF1 and COUP-TF1 (95,96).
NR3 subfamily
There are 15 NR3 subfamily members in F. heteroclitus. In comparison, there are 15 in T. rubripes, 16 in T. nigroviridis, and 9 in humans. There are 6 estrogen related receptors (ERRs; NR3B) in F. heteroclitus, while most fish species contain 5–7 and humans contain 3 (Table 1). There are 3 estrogen receptors (ERs)(NR3A) in F. heteroclitus, while most fish species contain 3–4 receptors and humans contain 2 (Table 1). Overall, there is a significant expansion of the 3A and 3B group in F. heteroclitus and other fish species relative to humans. Interestingly, Fundulus chronically exposed to estrogenic pollutants developed heritable splicing variants that show different transcriptional responses than wild-type ER potentially as an adaptive mechanism to the estrogens found in the polluted environment (11,18).
There are 6 NR3C (glucocorticoid, mineralcorticoid, androgen, and progesterone receptors) members in F. heteroclitus. There are also six members in Tetraodon and Takifugu, but only 4 NR3C members in zebrafish and humans (Table 1). F. heteroclitus has two glucocorticoid receptors (GR), two androgen receptors (AR), and one mineralocorticoid (MR) and one progesterone receptor (PR) within the NR3C group. This is consistent with several other fish species such as Tetraodon and Takifugu (Fig 1; Table 1) that have two GRs and two ARs (39,40).
In addition, medaka, stickleback (Gasterosteus aculeatus), common carp (Cyprinus carpio) and rainbow trout (Oncorynchus mykiss) all have two GRs (97,98); however, rainbow trout and common carp have undergone an additional round of genome duplication that may explain why these species have two GRs. Zebrafish (Danio rerio), only has one GR, GR2, that is missing a 9 amino acid sequence between the zinc fingers of the DBD (98,99). It is interesting to speculate that the increase in GRs are crucial in salt balance, especially in marine or estuarine species (12,100,101) as GR regulates salt balance in part by regulating the transcription of the sodium-chloride cotransporter and sodium-potassium ATPase (102,103). Carp, like zebrafish, are Ostariophysi and they contain two GRs. Thus, it appears that zebrafish unlike most other teleosts, lost a GR after the genome wide duplication in fish (104,105) and carp (and rainbow trout) have two GR because of the second genome duplication.
The loss of the second AR occurred in several fish species unlike the loss of GR2, which occurred in only a few species. F. heteroclitus, Takifugu and Tetraodon all have two ARs. However, Cypriniformes such as zebrafish, Siluriformes such as catfish, and Salmoniformes, such as salmon and trout all have one AR. Interestingly, early teleosts such as Osteoglossiformes (arowana, knifefish) and Anguilliformes (eels) contain two very similar ARs, while Percomorphs such as pufferfish and Fundulus show divergence between their ARs (106) with different binding affinities for steroids and xenobiotics (107). ARα which shows high divergence relative to humans and ARβ (Table 2), also has higher transactivation activity confirming neofunctionalization (108).
NR subfamilies 4–6
In F. heteroclitus, subfamilies 4–6 account for nine NRs; one more than T. nigroviridis because F. heteroclitus contains 5 NR4 members. Overall, there is a small expansion of the 4-subfamily as most fish species contain 4 members; however F. heteroclitus and Nile tilapia (O. niloticus) contain 5 members. Humans contain three NR4 members and most invertebrates contain one (20,22). Some NR4 members are crucial in brain differentiation, cell cycle, inflammation, and atherosclerosis (109–111). The NR4 subfamily is related to the NR1 subfamily, and is an ancient family that is ligand independent (46,112). Several of the NR4 members show relatively large differences between mummichog and human LBDs. This divergence is unique to the duplicated NR4A2 member, Nurr1b, but occurs is both NR4A1 member (Table 2).
The NR5 subfamiy contains FTZ members that regulate development. There are four members of the NR5 subfamily. SF-1 members are crucial in sex determination and down-regulated upon exposure to anthropogenic estrogenic chemicals during sex reversal (113). SF1a and FTZ-F2 show very high divergence relatively to human SF1 (Table 2). GCNF, which is the only NR6 subfamily member, is involved in growth and maturation (112). Zebrafish have two GCNF members but other fish species, Daphnia, and humans have one (Table 1)(23).
Conclusions
There are at least 74 full length NRs in the F. heteroclitus genome spanning all seven gene subfamilies of which 40% are in the NR1 subfamily often involved in circadian rhythms, development, energy metabolism and resource allocation. Fish species show expansion of the RAR (NR1B), Reverb (NR1D), ROR (NR1F), COUPTF (NR2F), ERR (NR3B), RXR (NR2B), and to a lesser extent the NGF (NR4A), and NR3C steroid receptors (GR/AR). Of particular interest is the co-expansion of opposing NRs, Reverb-ROR, and RAR/RXR-COUPTF, and the potential lack of an RXRα member in the F. heteroclitus genome.
Supplementary Material
Acknowledgments
Funds for this project were provided through a Salisbury Cove Research Fund Fellowship through Mount Desert Island Biological Laboratory (MDIBL) and NIH R15ES017321. We would also like to thank the Fundulus genomics consortium and NSF grant 1120333 for the genomic resources.
Footnotes
The authors have no conflicts of interest to report.
References
- 1.Burnett KG, Bain LJ, Baldwin WS, Callard GV, Cohen S, Di Giulio RT, Evans DH, Gomez-Chiarri M, Hahn ME, Hoover CA, Karchner SI, Katoh F, MacLatchy DL, Marshall WS, Meyer JN, Nacci DE, Oleksiak MF, Rees BB, Singer TD, Stegeman JJ, Towle DW, Van Veld PA, Vogelbein WK, Whitehead A, Winn RN, Crawford DL. Fundulus as the premier teleost model in environmental biology: Opportunities for new insights using genomics. Comp Biochem Physiol D. 2007;2:257–286. doi: 10.1016/j.cbd.2007.09.001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 2.Roling JA, Bain LJ, Gardea-Torresdey J, Key PB, Baldwin WS. Using mummichog (Fundulus heteroclitus) arrays to monitor the effectiveness of remediation at a superfund site in Charleston, South Carolina, USA. Environ Toxicol Chem. 2007;26:1205–1213. doi: 10.1897/06-421r.1. [DOI] [PubMed] [Google Scholar]
- 3.Reitzel AM, Karchner SI, Franks DG, Evans BR, Nacci D, Champlin D, Vieira VM, Hahn ME. Genetic variation at aryl hydrocarbon receptor (AHR) loci in populations of Atlantic killifish (Fundulus heteroclitus) inhabiting polluted and reference habitats. BMC Evol Biol. 2014;14:6. doi: 10.1186/1471-2148-14-6. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Roling JA, Bain LJ, Baldwin WS. Differential gene expression in mummichogs (Fundulus heteroclitus) following treatment with pyrene: comparison to a creosote contaminated site. Mar Environ Res. 2004;57:377–395. doi: 10.1016/j.marenvres.2003.11.001. [DOI] [PubMed] [Google Scholar]
- 5.Reid NM, Proestou DA, Clark BW, Warren WC, Colbourne JK, Shaw JR, Karchner SI, Crawford DL, Oleksiak MF, Hahn ME, Nacci D, Whitehead A. The genomic landscape of rapid repeated evolutionary adaptation to toxic pollution in wild fish. Science. 2016;354:1305–1308. doi: 10.1126/science.aah4993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Mancera JM, McCormick SD. Influence of cortisol, growth hormone, insulin-like growth factor I and 3,3′,5 triiodo-L-thyronine on hypoosmoregulatory ability in the euryhaline teleost Fundulus heteroclitus. Fish Physiol Biochem. 1999;21:25–33. [Google Scholar]
- 7.Podrabsky JE, Somero GN. Changes in gene expression associated with acclimation to constant temperatures and fluctuating daily temperatures in an annual killifish Austrofundulus limnaeus. J Exp Biol. 2004;207:2237–2254. doi: 10.1242/jeb.01016. [DOI] [PubMed] [Google Scholar]
- 8.Reid NM, Jackson CE, Gilbert D, Minx P, Montague MJ, Hampton TH, Helfrich LW, King BL, Nacci D, Aluru N, Karchner SI, Colbourne JK, Hahn ME, Shaw JR, Oleksiak MF, Crawford DL, Warren WC, Whitehead A. The Atlantic killifish (Fundulus heteroclitus) genome and the landscape of genome variation within a population. Genome Biol Evol. 2017;9:659–767. doi: 10.1093/gbe/evx023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Roling JA, Bain LJ, Gardea-Torresdey J, Bader J, Baldwin WS. Hexavalent chromium reduces juvenile growth and alters gene expression in Fundulus heteroclitus. Environ Toxicol Chem. 2006;25:62–70. doi: 10.1897/05-659r.1. [DOI] [PubMed] [Google Scholar]
- 10.Hahn ME, Karchner SI, Franks DG, Merson RR. Aryl hydrocarbon receptor polymorphisms and dioxin resistance to Atlantic killifish (Fundulus heroclitus) Pharmacogenetics. 2004;14:131–143. doi: 10.1097/00008571-200402000-00007. [DOI] [PubMed] [Google Scholar]
- 11.Cotter KA, Nacci D, Champlin D, Chuprin J, Callard GV. Cloning of multiple ERα mRNA variants in killifish (Fundulus heteroclitus), and differential expression by tissue type, stage of reproduction, and estrogen exposure in fish from polluted and unpolluted environments. Aquat Toxicol. 2015;159:184–197. doi: 10.1016/j.aquatox.2014.12.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Shaw JR, Gabor K, Hand E, Lankowski A, Durant L, Thibodeau R, Stanton CR, Barnaby R, Coutermarsh B, Karlson KH, Sato JD, Hamilton JW, Stanton BA. Role of glucocorticoid receptor in acclimation of killifish (Fundulus heteroclitus) to seawater and effects of arsenic. Am J Physiol Regul Integr Comp Physiol. 2007;292:R1052–R1060. doi: 10.1152/ajpregu.00328.2006. [DOI] [PubMed] [Google Scholar]
- 13.Whitehead A, Dubansky B, Bodinier C, Garcia TI, Miles S, Pilley C, Raghunathan V, Roach JL, Walker N, Walter RB, Rice CD, Galvez F. Genomic and physiological footprint of the Deepwater Horizon oil spill on resident marsh fishes. Proc Nat Acad Sci USA. 2014;109:29298–29302. doi: 10.1073/pnas.1109545108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Dubansky B, Whitehead A, Miller JT, Rice CD, Galvez F. Multitissue molecular, genomic, and developmental effects of the Deepwater Horizon oil spill on resident Gulf killifish (Fundulus grandis) Environ Sci Technol. 47:5074–5082. doi: 10.1021/es400458p. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Bello SM, Franks DG, Stegeman JJ, Hahn ME. Acquired resistance to Ah receptor agonists in a population of atlantic killifish (Fundulus heteroclitus) inhabting a marine superfund site: in vivo and in vitro studies on the inducibility of xenobiotic metabolizing enzymes. Toxicol Sci. 2001;60:77–91. doi: 10.1093/toxsci/60.1.77. [DOI] [PubMed] [Google Scholar]
- 16.Meyer JN, Nacci DE, Di Giulio RT. Cytochrome P4501A (CYP1A) in killifish (Fundulus heteroclitus): heritability of altered expression and relationship to survival in contaminated sediments. Toxicol Sci. 2002;68:69–81. doi: 10.1093/toxsci/68.1.69. [DOI] [PubMed] [Google Scholar]
- 17.Wojdylo JV, Vogelbein W, Bain LJ, Rice CD. AHR-related activities in a creosote-adapted population of adult atlantic killifish, Fundulus heteroclitus, two decades post-EPA superfund status at the Atlantic Wood Site, Portsmouth, VA USA. Aquat Toxicol. 2016;177:77–85. doi: 10.1016/j.aquatox.2016.05.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Cotter KA, Nacci D, Champlin D, Yeo AT, Gilmore TD, Callard GV. Adaptive Significance of ERα Splice Variants in Killifish (Fundulus heteroclitus) Resident in an Estrogenic Environment. Endocrinology. 2016;157:2294–2308. doi: 10.1210/en.2016-1052. [DOI] [PubMed] [Google Scholar]
- 19.Helsen C, Claessens F. Looking at nuclear receptors from a new angle. Mol Cell Endocrinol. 2014;382:97–106. doi: 10.1016/j.mce.2013.09.009. [DOI] [PubMed] [Google Scholar]
- 20.Litoff EJ, Garriott TE, Ginjupalli GK, Butler L, Gay C, Scott K, Baldwin WS. Annotation of the Daphnia magna nuclear receptors: Comparison to Daphnia pulex. Gene. 2014;552:116–125. doi: 10.1016/j.gene.2014.09.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Hernandez JP, Mota LC, Baldwin WS. Activation of CAR and PXR by dietary, environmental and occupational chemicals alters drug metabolism, intermediary metabolism, and cell proliferation. Curr Pharmacog Personal Med. 2009;7:81–105. doi: 10.2174/187569209788654005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Thomson SA, Baldwin WS, Wang YH, Kwon G, LeBlanc GA. Annotation, phylogenetics, and expression of the nuclear receptors in Daphnia pulex. BMC Genomics. 2009;10:500. doi: 10.1186/1471-2164-10-500. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Cheng YY, Tao WJ, Chen J-J, Sun LN, Zhou LY, Song Q, Wang DS. Genome-wide identification, evolution and expression analysis of nuclear receptor superfamily in Nile tilapia, Oreochromis niloticus. Gene. 2015;569:141–142. doi: 10.1016/j.gene.2015.05.057. [DOI] [PubMed] [Google Scholar]
- 24.Chawla A, Repa JJ, Evans RM, Mangelsdorf DJ. Nuclear receptors and lipid physiology: opening the X-files. Science. 2001;294:1866–1870. doi: 10.1126/science.294.5548.1866. [DOI] [PubMed] [Google Scholar]
- 25.Kliewer SA, Lehmann JM, Willson TM. Orphan Nuclear Receptors: Shifting Endocrinology into Reverse. Science. 1999;284:757–760. doi: 10.1126/science.284.5415.757. [DOI] [PubMed] [Google Scholar]
- 26.Pascussi JM, Gerbal-Chaloin S, Duret C, Daujat-Chavanieu M, Vilarem MJ, Maurel P. The tangle of nuclear receptors that controls xenobiotic metabolism and transport: Crosstalk and consequences. Annu Rev Pharmacol Toxicol. 2008;48:1.1–1.31. doi: 10.1146/annurev.pharmtox.47.120505.105349. [DOI] [PubMed] [Google Scholar]
- 27.Mello T, Materozzi M, Galli A. PPARs and Mitochondrial Metabolism: From NAFLD to HCC. PPAR Res. 2017 doi: 10.1155/2016/7403230. epub ahead of print. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Yang H, Wang H. Signaling control of the constitutive androstane receptor (CAR) Protein Cell. 2014;5:113–123. doi: 10.1007/s13238-013-0013-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Kretschmer XC, Baldwin WS. CAR and PXR: Xenosensors of Endocrine Disrupters? Chem-Biol Interac. 2005;155:111–128. doi: 10.1016/j.cbi.2005.06.003. [DOI] [PubMed] [Google Scholar]
- 30.Evans RM. The nuclear receptor superfamily: A rosetta stone for physiology. Mol Endocrinol. 2005;19:1429–1434. doi: 10.1210/me.2005-0046. [DOI] [PubMed] [Google Scholar]
- 31.Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. doi: 10.1016/S0022-2836(05)80360-2. [DOI] [PubMed] [Google Scholar]
- 32.Marchler-Bauer A, Bryant SH. CD-Search: protein domain annotations on the fly. Nucleic Acids Res. 2004;32:W327–331. doi: 10.1093/nar/gkh454. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Hannas BR, Wang YH, Baldwin WS, Li Y, Wallace AD, LeBlanc GA. Interactions of the crustacean nuclear receptors HR3 and E75 in the regulation of gene transcription. Gen Comp Endocrinol. 2010;167:268–278. doi: 10.1016/j.ygcen.2010.03.025. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Marchler-Bauer A, Anderson JB, Derbyshire MK, DeWeese-Scott C, Gonzales NR, Gwadz M, Hao L, He S, Hurwitz DI, Jackson JD, Ke Z, Krylov D, Lanczycki CJ, Liebert CA, Liu C, Lu F, Lu S, Marchler GH, Mullokandov M, Song JS, Thanki N, Yamashita RA, Yin JJ, Zhang D, Bryant SH. CDD: a conserved domain database for interactive domain family analysis. Nucleic Acids Res. 2007;35:237–240. doi: 10.1093/nar/gkl951. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.Li Y, Ginjupalli GK, Baldwin WS. The HR97 (NR1L) Group of Nuclear Receptors: A New Group of Nuclear Receptors Discovered in Daphnia species. Gen Comp Endocrinol. 2014;206:30–42. doi: 10.1016/j.ygcen.2014.07.022. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Long E, Scott G, Kucklick J, Fulton M, Thompson B, Carr R, Scott K, Thursby G, Chandler G, Anderson J, Sloane G. NOAA Technical Memorandum NOS ORCA. 1997. Magnitude and extent of sediment toxicity in selected estuaries of South Carolina and Georgia; p. 178. [Google Scholar]
- 37.Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol Biol Evol. 2013;30:2725–2729. doi: 10.1093/molbev/mst197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Swofford DL. PAUP*: phylogenetic analysis using parsimony (*and other methods), version 4.0b10. Sinauer Associates; Sunderland, MA: 2001. [Google Scholar]
- 39.Maglich JM, Caravella JA, Lambert MH, Willson TM, Moore JT, Rammamurthy L. The first complete genome sequence from a teleost fish (Fugu rubripes) adds significant diversity to the nuclear receptor superfamily. Nucleic Acids Res. 2003;31:4051–4058. doi: 10.1093/nar/gkg444. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Metpally RPR, Vigneshwar R, Sowdhamini R. Genome inventory and analysis of nuclear hormone receptors in Tetraodon nigroviridis. J Biosci. 2007;32:43–50. doi: 10.1007/s12038-007-0005-4. [DOI] [PubMed] [Google Scholar]
- 41.Robinson-Rechavi M, Marchand O, Escriva H, Bardet PL, Zelus D, Hughes S, Laudet V. Euteleost fish genomes are characterized by expansion of gene families. Genome Res. 2001;11:781–788. doi: 10.1101/gr.165601. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Glasauer SM, Neuhauss SC. Whole-genome duplication in teleost fishes and its evolutionary consequences. Mol Genet Genomics. 2014;289:1045–1060. doi: 10.1007/s00438-014-0889-2. [DOI] [PubMed] [Google Scholar]
- 43.Perl TN, Schmid BGM, Schwirz J, Chipman AD. The evolution of the knirps family of transcription factors in arthropods. Mol Biol Evol. 2013;30:1348–1357. doi: 10.1093/molbev/mst046. [DOI] [PubMed] [Google Scholar]
- 44.Laudet V. Evolution of the nuclear receptor superfamily: early diversification from an ancestral orphan receptor. J Mol Endocrinol. 1997;19:207–226. doi: 10.1677/jme.0.0190207. [DOI] [PubMed] [Google Scholar]
- 45.Bertrand S, Brunet F, Escriva H, Parmentier G, Laudet V, Robinson-Rechavi M. Evolutionary genomics of nuclear receptors: from twenty-five ancestral genes to derived endocrine systems. Mol Biol Evol. 2004;21:1923–1937. doi: 10.1093/molbev/msh200. [DOI] [PubMed] [Google Scholar]
- 46.Bridgham JT, Eick GN, Larroux C, Deshpande K, Harms MJ, Gauthier MEA, Ortlund EA, Degnan BM, Thornton JW. Protein Evolution by Molecular Tinkering: Diversification of the Nuclear Receptor Superfamily from a Ligand-Dependent Ancestor. PLoS Biology. 2010;8:e1000497. doi: 10.1371/journal.pbio.1000497. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Germain P, Staels B, Dacquet C, Spedding M, Laudet V. Overview of nomenclature of nuclear receptors. Pharmacol Rev. 2006;58:685–704. doi: 10.1124/pr.58.4.2. [DOI] [PubMed] [Google Scholar]
- 48.Bae Y, Kemper JK, Kemper B. Repression of CAR-mediated transactivation of CYP2B genes by the orphan nuclear receptor, short heterodimer partner (SHP) DNA Cell Biol. 2004;23:81–91. doi: 10.1089/104454904322759894. [DOI] [PubMed] [Google Scholar]
- 49.Bavner A, Sanyal S, Gustafsson JA, Treuter E. Transcriptional corepression by SHP: molecular mechanisms and physiological consequences. TRENDS Endocrinol Metab. 2005;16:478–488. doi: 10.1016/j.tem.2005.10.005. [DOI] [PubMed] [Google Scholar]
- 50.Johansson L, Thomsen JS, Damdimopoulos AE, Spyrou G, Gustafsson JA, Treuter E. The orphan nuclear receptor SHP inhibits agonist-dependent transcriptional activity of estrogen receptors ERa and ERb*. J Biol Chem. 1999;274:345–353. doi: 10.1074/jbc.274.1.345. [DOI] [PubMed] [Google Scholar]
- 51.Ehrlund A, Treuter E. Ligand-independent actions of the orphan receptors/corepressors DAX-1 and SHP in metabolism, reproduction and disease. J Steroid Biochem Mol Biol. 2012;130:169–179. doi: 10.1016/j.jsbmb.2011.04.007. [DOI] [PubMed] [Google Scholar]
- 52.Mark M, Ghyselinck NB, Chambon P. Function of retinoic acid receptors during embryonic development. Nucl Recept Signal. 2009;7:e002. doi: 10.1621/nrs.07002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Jetten AM. Retinoid-related orphan receptors (RORs): critical roles in development, immunity, circadian rhythm, and cellular metabolism. Nucl Recept Signal. 2009;7:e003. doi: 10.1621/nrs.07003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Ramakrishnan SN, Muscat GEO. The orphan Rev-erb nuclear receptors: a link between metabolism, circadian rhythm and inflammation? Nucl Recept Signal. 2006;4:e009. doi: 10.1621/nrs.04009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Forman BM, Chen J, Blumberg B, Kliewer SA, Henshaw R, Ong ES, Evans RM. Cross-talk among RORα1 and the Rev-erb family of orphan nuclear receptors. Mol Endocrinol. 1994;8:1253–1261. doi: 10.1210/mend.8.9.7838158. [DOI] [PubMed] [Google Scholar]
- 56.Delezie J, Challet E. Interactions between metabolism and circadian clocks: reciprocal disturbances. Ann NY Acad Sci. 2011;1243:30–46. doi: 10.1111/j.1749-6632.2011.06246.x. [DOI] [PubMed] [Google Scholar]
- 57.Echeverría F, Ortiz M, Valenzuela R, Videla LA. Long-chain polyunsaturated fatty acids regulation of PPARs, signaling: Relationship to tissue development and aging. Prostaglandins Leukot Essent Fatty Acids. 2016;114:28–34. doi: 10.1016/j.plefa.2016.10.001. [DOI] [PubMed] [Google Scholar]
- 58.Leaver MJ, Boukouvala E, Antonopoulou E, Diez A, Favre-Krey L, Ezaz MT, Bautista JM, Tocher DR, Krey G. Three peroxisome proliferator-activated receptor isotypes from each of two species of marine fish. Endocrinology. 2005;146:3150–3162. doi: 10.1210/en.2004-1638. [DOI] [PubMed] [Google Scholar]
- 59.Leaver MJ, Ezaz MT, Fontagne S, Tocher DR, Boukouvala E, Krey G. Multiple peroxisome proliferator-activated receptor beta subtypes from Atlantic salmon (Salmo salar) J Mol Endocrinol. 2007;38:391–400. doi: 10.1677/JME-06-0043. [DOI] [PubMed] [Google Scholar]
- 60.Benton MJ. Vertebrate Paleontology. 4. Wiley-Blackwell; 2014. [Google Scholar]
- 61.Cortesi F, Musilova Z, Stieb SM, Hart NS, Siebeck UE, Malmstrøm M, Tørresen OK, Jentoft S, Cheney KL, Marshall NJ, Cheney KL, Marshall J, Carleton KL, Salzburgera W. Ancestral duplications and highly dynamic opsin gene evolution in percomorph fishes. Proc Natl Acad Sci USA. 2015;112:1493–1498. doi: 10.1073/pnas.1417803112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Baldwin WS, Marko PB, Nelson DR. The Cytochrome P450 (CYP) gene superfamily in Daphnia pulex. BMC Genomics. 2009;10:169. doi: 10.1186/1471-2164-10-169. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Nelson DR. Comparison of P450s from human and fugu: 420 million years of vertebrate P450 evolution. Arch Biochem Biophys. 2003;409:18–24. doi: 10.1016/s0003-9861(02)00553-2. [DOI] [PubMed] [Google Scholar]
- 64.Prindiville JS, Mennigen JA, Zamora JM, Moon TW, Weber JM. The fibrate drug gemfibrozil disrupts lipoprotein metabolism in rainbow trout. Toxicol Appl Pharmacol. 2011;251:201–208. doi: 10.1016/j.taap.2010.12.013. [DOI] [PubMed] [Google Scholar]
- 65.Cajaraville MP, Cancio I, Ibabe A, Orbea A. Peroxisome proliferation as a biomarker in environmental pollution assessment. Microsc Res Tech. 2003:61. doi: 10.1002/jemt.10329. [DOI] [PubMed] [Google Scholar]
- 66.Arnold H, Pluta HJ, Braunbeck T. Simultaneous exposure of fish to endosulfan and disulfoyon in vivo: ultrastructural, stereological and biochemical reactions in hepatocytes of male rainbow trout (Onchorhynchus mykiss) Aquat Toxicol. 1995;33:17–43. [Google Scholar]
- 67.Pedradas JR, Lopez-Barea J, Peinado J. Dieldrin induces peroxisomal enzymes in fish (Sparus aurata) liver. Comp Biochem Physiol C. 1996;115:125–131. doi: 10.1016/s0742-8413(96)00051-5. [DOI] [PubMed] [Google Scholar]
- 68.Ye T, Kang M, Huang Q, Fang C, Chen Y, Liu L, Dong S. Accumulation of di(2-ethylhexyl) phthalate causes endocrine-disruptive effects in marine medaka (Oryzias melastigma) embryos. Environ Toxicol. 2016;31:116–127. doi: 10.1002/tox.22028. [DOI] [PubMed] [Google Scholar]
- 69.Olivares-Rubio HF, Vega-López A. Fatty acid metabolism in fish species as a biomarker for environmental monitoring. Environ Pollut. 2016;218:297–312. doi: 10.1016/j.envpol.2016.07.005. [DOI] [PubMed] [Google Scholar]
- 70.Ackers JT, Johnston MF, Haasch ML. Immunodetection of hepatic peroxisomal PMP70 as an indicator of peroxisomal proliferation in the mummichog, Fundulus heteroclitus. Mar Environ Res. 2000;50:361–365. doi: 10.1016/s0141-1136(00)00061-1. [DOI] [PubMed] [Google Scholar]
- 71.Lyssimachou A, Santos JG, André A, Soares J, Lima D, Guimarães L, Almeida CM, Teixeira C, Castro LF, Santos MM. The mammalian “obesogen” tributyltin targets hepatic triglyceride accumulation and the transcriptional regulation of lipid metabolism in the liver and brain of zebrafish. PLoS One. 2015;10:e0143911. doi: 10.1371/journal.pone.0143911. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Adeogun AO, Ibor OR, Regoli F, Arukwe A. Peroxisome proliferator-activated receptors and biotransformation responses in relation to condition factor and contaminant burden in tilapia species from Ogun River, Nigeria. Comp Biochem Physiol C Toxicol Pharmacol. 2016;183–184:7–19. doi: 10.1016/j.cbpc.2015.12.006. [DOI] [PubMed] [Google Scholar]
- 73.Milnes MR, Garcia A, Grossman E, Grun F, Shiotsugu J, Tabb MM, Kawashima Y, Katsu Y, Watanabe H, Iguchi T, Blumberg B. Activation of steroid and xenobiotic receptor (SXR, NR1I2) and its orthologs in laboratory, toxicologic, and genome model species. Environ Health Perspect. 2008;116:880–885. doi: 10.1289/ehp.10853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Gräns J, Wassmur B, Fernández-Santoscoy M, Zanette J, Woodin BR, Karchner SI, Nacci DE, Champlin D, Jayaraman S, Hahn ME, Stegeman JJ, Celander MC. Regulation of pregnane-X-receptor, CYP3A and P-glycoprotein genes in the PCB-resistant killifish (Fundulus heteroclitus) population from New Bedford Harbor. Aquat Toxicol. 2015;159:198–207. doi: 10.1016/j.aquatox.2014.12.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Moore LB, Maglich JM, McKee DD, Wisely B, Willson TM, Kliewer SA, Lambert MH, Moore JT. Pregnane X receptor (PXR), constitutive androstane receptor (CAR), and benzoate X receptor (BXR) define three pharmacologically distinct classes of nuclear receptors. Mol Endocrinol. 2002;16:977–986. doi: 10.1210/mend.16.5.0828. [DOI] [PubMed] [Google Scholar]
- 76.Krasowski MD, Yasuda K, Hagey LR, Schuetz EG. Evolutionary selection across the nuclear hormone receptor superfamily with a focus on the NR1I subfamily (vitamin D, pregnane X, and constitutive androstane receptors) Nucl Recept. 2005;3:2. doi: 10.1186/1478-1336-3-2. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Mathäs M, Burk O, Qiu H, Nußhag C, Gödtel-Armbrust U, Baranyai D, Deng S, Römer K, Nem D, Windshügel B, Wojnowski L. Evolutionary History and Functional Characterization of the Amphibian Xenosensor CAR. Mol Endocrinol. 2012;26:14–26. doi: 10.1210/me.2011-1235. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Zhao Y, Zhang K, Giesy JP, Hu J. Families of Nuclear Receptors in Vertebrate Models: Characteristic and Comparative Toxicological Perspective. Sci Rep. 2015;5:8554. doi: 10.1038/srep08554. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 79.Georges D, Blieck A. Rise of the earliest tetrapods: an early Devonian origin from marine environment. PLoS ONE. 2011;6:e221362011. doi: 10.1371/journal.pone.0022136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 80.Krasowski MD, Yasuda K, Hagey LR, Schuetz EG. Evolution of the pregnane X receptor: adaptation to cross-species differences in biliary bile salts. Mol Endocrinol. 2005;19:1720–1739. doi: 10.1210/me.2004-0427. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Zhang J, Huang W, Qatanani M, Evans RM, Moore DD. The constitutive androstane receptor and pregnane X receptor function coordinately to prevent bile acid-induced hepatotoxicity. J Biol Chem. 2004;279:49517–49522. doi: 10.1074/jbc.M409041200. [DOI] [PubMed] [Google Scholar]
- 82.Fiorucci S, Zampella A, Distrutti E. Development of FXR, PXR and CAR agonists and antagonists for treatment of liver disorders. Curr Top Med Chem. 2012;12:605–624. doi: 10.2174/156802612799436678. [DOI] [PubMed] [Google Scholar]
- 83.Howarth DL, Hagey LR, Law SH, Ai N, Krasowski MD, Ekins S, Moore JT, Kollitz EM, Hinton DE, Kullman SW. Two farnesoid X receptor alpha isoforms in Japanese medaka (Oryzias latipes) are differentially activated in vitro. Aquat Toxicol. 2010;98:245–255. doi: 10.1016/j.aquatox.2010.02.020. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 84.Krasowski MD, Ni A, Hagey LR, Ekins S. Evolution of promiscuous nuclear hormone receptors: LXR, FXR, VDR, PXR, and CAR. Mol Cell Endocrinol. 2011;334:39–48. doi: 10.1016/j.mce.2010.06.016. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 85.Krasowski MD, Ai N, Hagey LR, Kollitz EM, Kullman SW, Reschly EJ, Ekins S. The evolution of farnesoid X, vitamin D, and pregnane X receptors: insights from the greenspotted pufferfish (Tetraodon nigriviridis) and other non-mammalian species. BMC Biochemistry. 2011;12:5. doi: 10.1186/1471-2091-12-5. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 86.Reschly EJ, Ai N, Ekins S, Welsh WJ, Hagey LR, Hofmann AF, Krasowski MD. Evolution of the bile salt nuclear receptor FXR in vertebrates. J Lipid Res. 2008;49:1577–1587. doi: 10.1194/jlr.M800138-JLR200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 87.Kitambi SS, Hauptmann G. The zebrafish orphan nuclear receptor genes nr2e1 and NR2E3 are expressed in developing eye and forebrain. Gene Expr Patterns. 2007;7:521–528. doi: 10.1016/j.modgep.2006.10.006. [DOI] [PubMed] [Google Scholar]
- 88.Nelson SM, Frey RA, Wardwell SL, Stenkamp DL. The developmental sequence of gene expression within the rod photoreceptor lineage in embryonic zebrafish. Dev Dyn. 2008;237:2903–2917. doi: 10.1002/dvdy.21721. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 89.Wang T, Zhao LJ, Li P, Jiang H, Lu GC, Zhang WD, Li HL, Yuan BJ. Hepatoprotective effects and mechanisms of dehydrocavidine in rats with carbon tetrachloride-induced hepatic fibrosis. J Ethnopharmacol. 2011;138:76–84. doi: 10.1016/j.jep.2011.08.039. [DOI] [PubMed] [Google Scholar]
- 90.Amoutzias GD, Pichler EE, Mian N, De Graaf D, Imsiridou A, Robinson-Rechavi M, Bornberg-Bauer E, Robertson DL, Oliver SG. A protein interaction atlas for the nuclear receptors: properties and quality of a hub-based dimerisation network. BMC Systems Biology. 2007;1:34. doi: 10.1186/1752-0509-1-34. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 91.Aranguren XL, Beerens M, Vandevelde W, Dewerchin M, Carmeliet P, Luttun A. Transcription factor COUP-TFII is indispensable for venous and lymphatic development in zebrafish and Xenopus laevis. Biochem Biophys Res Commun. 2011;410:121–126. doi: 10.1016/j.bbrc.2011.05.117. [DOI] [PubMed] [Google Scholar]
- 92.Swift MR, Pham VN, Castranova D, Bell K, Poole RJ, Weinstein BM. SoxF factors and Notch regulate nr2f2 gene expression during venous differentiation in zebrafish. Dev Biol. 2014;390:116–125. doi: 10.1016/j.ydbio.2014.03.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 93.Tran P, Zhang XK, Salbert G, Hermann T, Lehmann JM, Pfahl M. COUP orphan receptors are negative regulators of retinoic acid response pathways. Mol Cell Biol. 1992;12:4666–4676. doi: 10.1128/mcb.12.10.4666. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Robinson-Rechavi M, Maina CV, Gissendanner CR, Laudet V, Sluder A. Explosive lineage-specific expansion of the orphan nuclear receptor HNF4 in nematodes. J Mol Evol. 2005;60:577–586. doi: 10.1007/s00239-004-0175-8. [DOI] [PubMed] [Google Scholar]
- 95.Cheng W, Guo L, Zhang Z, Soo HM, Wen C, Wu W, Peng J. HNF factors form a network to regulate liver-enriched genes in zebrafish. Dev Biol. 2006;294:482–496. doi: 10.1016/j.ydbio.2006.03.018. [DOI] [PubMed] [Google Scholar]
- 96.McNair A, Cereghini S, Brand H, Smith T, Breillat C, Gannon F. Synergistic activation of the Atlantic salmon hepatocyte nuclear factor (HNF) 1 promoter by the orphan nuclear receptors HNF4 and chicken ovalbumin upstream promoter transcription factor I (COUP-TFI) Biochem J, 352 Pt. 2000;2:557–564. [PMC free article] [PubMed] [Google Scholar]
- 97.Miyagawa S, Lange A, Tohyama S, Ogino Y, Mizutani T, Kobayashi T, Tatarazako N, Tyler CR, Iguchi T. Characterization of Oryzias latipes glucocorticoid receptors and their unique response to progestins. J Appl Toxicol. 2015;35:302–309. doi: 10.1002/jat.3020. [DOI] [PubMed] [Google Scholar]
- 98.Bury NR. The evolution, structure and function of the ray finned fish (Actinopterygii) glucocorticoid receptors. Gen Comp Endocrinol. 2017 doi: 10.1016/j.ygcen.2016.06.030. epub ahead of print. [DOI] [PubMed] [Google Scholar]
- 99.Kumai Y, Nesan D, Yijayan MM, Perry SF. Cortisol regulates Na+ uptake in zebrafish, Danio rerio, larvae via the glucocorticoid receptor. Mol Cell Endocrinol. 2012;364:113–125. doi: 10.1016/j.mce.2012.08.017. [DOI] [PubMed] [Google Scholar]
- 100.Notch EG, Shaw JR, Coutermarsh BA, Dzioba M, Stanton BA. Morpholino gene knockdown in adult Fundulus heteroclitus: Role of SGK1 in seawater acclimation. PLoS One. 2011;6:e29462. doi: 10.1371/journal.pone.0029462. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 101.Scott GR, Baker DW, Schulte PM, Wood CM. Physiological and molecular mechanisms of osmoregulatory plasticity in killifish after seawater transfer. J Exp Biol. 2008;211:2450–2459. doi: 10.1242/jeb.017947. [DOI] [PubMed] [Google Scholar]
- 102.Lin CH, Hu HJ, Hwang PP. Cortisol regulates sodium homeostasis by stimulating the transcription of sodium-chloride transporter (NCC) in zebrafish (Danio rerio) Mol Cell Endocrinol. 2016;422:93–102. doi: 10.1016/j.mce.2015.12.001. [DOI] [PubMed] [Google Scholar]
- 103.Flores AM, Shrimpton MJ. Differential physiological and endocrine responses of rainbow trout, Oncorhynchus mykiss, transferred from fresh water to ion-poor or salt water. Gen Comp Endocrinol. 2012;175:244–250. doi: 10.1016/j.ygcen.2011.11.002. [DOI] [PubMed] [Google Scholar]
- 104.Hoegg S, Brinkmann H, Taylor JS, Meyer A. Phylogenetic timing of the fish-specific genome duplication correlates with the diversification of teleost fish. J Mol Evol. 2004;59:190–201. doi: 10.1007/s00239-004-2613-z. [DOI] [PubMed] [Google Scholar]
- 105.Crollius HR, Weissenbach J. Fish genomics and biology. Genome Res. 2005;15:1675–1682. doi: 10.1101/gr.3735805. [DOI] [PubMed] [Google Scholar]
- 106.Douard V, Brunet F, Boussau B, Ahrens-Fath I, Vlaeminck-Guillem V, Haendler B, Laudet V, Guiguen Y. The fate of the duplicated androgen receptor in fishes: a late neofunctionalization event? BMC Evol Biol. 2008;8:336. doi: 10.1186/1471-2148-8-336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Thomas P, Sperry TS. Identificatin of two nuclear androgen receptors in kelp bass (Paralabrax clathratus) and their binding affinities for xenobiotics: Comparison with atlantic croaker (Micropogonias undulatus) androgen receptors. Biol Reprod. 1999;61:1152–1161. doi: 10.1095/biolreprod61.4.1152. [DOI] [PubMed] [Google Scholar]
- 108.Ogino Y, Kuraku S, Ishibashi H, Miyakawa H, Sumiya E, Miyagawa S, Matsubara H, Yamada G, Baker ME, Iguchi T. Neofunctionalization of Androgen Receptor by Gain-of-Function Mutations in Teleost Fish Lineage. Mol Biol Evol. 2016;33:228–244. doi: 10.1093/molbev/msv218. [DOI] [PubMed] [Google Scholar]
- 109.Blin M, Norton W, Bally-Cuif L, Vernier P. NR4A2 controls the differentiation of selective dopaminergic nuclei in the zebrafish brain. Mol Cell Neurosci. 2008;39:596–604. doi: 10.1016/j.mcn.2008.08.006. [DOI] [PubMed] [Google Scholar]
- 110.Pei L, Castrillo A, Tontonoz P. Regulation of macrophage inflammatory gene expression by the orphan nuclear receptor Nur77. Mol Endocrinol. 2006;20:786–794. doi: 10.1210/me.2005-0331. [DOI] [PubMed] [Google Scholar]
- 111.Maxwell MA, Muscat GEO. The NR4A subgroup: immediate early response genes with pleiotropic physiological roles. Nucl Recept Signal. 2006;4:e002. doi: 10.1621/nrs.04002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 112.King-Jones K, Thummel CS. Nuclear receptors: A perspective from Drosophila. Nature Rev Genetics. 2005;6:311–323. doi: 10.1038/nrg1581. [DOI] [PubMed] [Google Scholar]
- 113.Dong RR, Yang SJ, Feng RJ, Fang LL, Sun YL, Zhang YG, Xie XJ, Wang DS. Complete feminization of catfish by feeding Limnodilus, an annelid worm collected in contaminated streams. Environ Res. 2014;133:371–379. doi: 10.1016/j.envres.2014.05.018. [DOI] [PubMed] [Google Scholar]
- 114.Sladek F. What are nuclear receptor ligands? Mol Cell Endocrinol. 2011;334:3–13. doi: 10.1016/j.mce.2010.06.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.LeBlanc GA, Norris DO, Kloas W, Kullman SW, Baldwin WS, Greally JM. Series on Testing & Assessment: No. 178. Organisation for Economic Co-operation and Development; Paris: 2012. Detailed Review Paper on the State of the Science on Novel In Vitro and In Vivo Screening and Testing Methods and Endpoints for Evaluating Endocdrine Disruptors; p. 213. [Google Scholar]
- 116.Kollitz EM, Zhang G, Hawkins MB, Whitfield GK, Reif DM, Kullman SW. Molecular Cloning, Functional Characterization, and Evolutionary Analysis of Vitamin D Receptors Isolated from Basal Vertebrates. PLOS ONE. 2015;10:e0122853. doi: 10.1371/journal.pone.0122853. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Reschly EJ, Krasowski MD. Evolution and function of the NR1I nuclear hormone receptor subfamily (VDR, PXR, and CAR) with respect to metabolism of xenobiotics and endogenous compounds. Curr Drug Metab. 2006;7:349–365. doi: 10.2174/138920006776873526. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Tzameli I, Chua SS, Cheskis B, Moore DD. Complex effects of rexinoids on ligand dependent activation or inhibition of the xenobiotic receptor, CAR. Nuclear Receptor. 2003:1. doi: 10.1186/1478-1336-1-2. http://www.nuclear-receptor.com/content/1/1/2. [DOI] [PMC free article] [PubMed]
- 119.Menuet A, Pellegrini E, Anglade I, Blaise O, Laudet V, Kah O, Pakdel F. Molecular characterization of three estrogen receptor forms in zebrafish: binding characteristics, transactivation properties, and tissue distributions. Biol Reprod. 2002;66:1881–1892. doi: 10.1095/biolreprod66.6.1881. [DOI] [PubMed] [Google Scholar]
- 120.Stolte EH, van Kemenade LV, Savelkoul HFJ, Flik G. Evolution of glucocorticoid receptors with different glucocorticoid sensitivity. J Endocrinol. 2006;190:17–28. doi: 10.1677/joe.1.06703. [DOI] [PubMed] [Google Scholar]
- 121.de Waal PP, Wang DS, Nijenhuis WA, Schulz RW, Bogerd J. Functional characterization and expression analysis of the androgen receptor in zebrafish (Danio rerio) testis. Reproduction. 2008;136:225–234. doi: 10.1530/REP-08-0055. [DOI] [PubMed] [Google Scholar]
- 122.Sablin EP, Blind RD, Krylova IN, Ingraham JG, Cai F, Williams JD, Fletterick RJ, Ingraham HA. Structure of SF-1 bound by different phospholipids; evidence for regulatory ligands. Mol Endocrinol. 2009;23:25–34. doi: 10.1210/me.2007-0508. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.