

Abstract
Incentives play a crucial role in guiding behavior throughout our lives, but perhaps no more so than during the early years of life. The ventral striatum is a critical piece of an incentive-based learning circuit, sharing robust anatomical connections with subcortical (e.g., amygdala, hippocampus) and cortical structures (e.g., medial prefrontal cortex (mPFC), insula) that collectively support incentive valuation and learning. Resting-state functional connectivity (rsFC) is a powerful method that provides insight into the development of the functional architecture of these connections involved in incentive-based learning. We employed a seed-based correlation approach to investigate ventral striatal rsFC in a cross-sectional sample of typically developing individuals between the ages of 4.5 and 23-years old (n=66). Ventral striatal rsFC with the mPFC showed regionally specific linear age-related changes in connectivity that were associated with age-related increases in circulating testosterone levels. Further, ventral striatal connectivity with the posterior hippocampus and posterior insula demonstrated quadratic age-related changes characterized by negative connectivity in adolescence. Finally, across this age range, the ventral striatum demonstrated positive coupling with the amygdala beginning during childhood and remaining consistently positive across age. In sum, our findings suggest normative ventral striatal rsFC development is dynamic and characterized by early establishment of connectivity with medial prefrontal and limbic structures supporting incentive-based learning, as well as substantial functional reorganization with later developing regions during transitions into and out of adolescence.
Keywords: ventral striatum, development, resting-state, incentives, valuation
1.1 Introduction
Incentives play a crucial role in our lives, providing opportunities to learn about and adapt to the environment. Pursuit (or avoidance) of incentives is a powerful motivator of behavior particularly during development (e.g., early life), when less is known about the world and the value of environmental stimuli is being learned rapidly. Research across species highlights the ventral striatum as a key neural structure supporting incentive-based valuation by coding for expected and experienced incentives of varying valence and magnitude to inform learning and decision-making (Alexander et al., 1990; Delgado, 2007; Haber and Knutson, 2010; Haber and McFarland, 1999; Hare et al., 2008a; O’Doherty, 2004; Plassmann et al., 2007; Robbins and Everitt, 1996; Robbins et al., 1989). This structure is extensively connected to both cortical and subcortical structures involved in valuation and learning, forming components of functional neural circuits supporting motivated behavior (Alexander et al., 1990; Haber and Knutson, 2010; Haber and McFarland, 1999; Pennartz et al., 2011). Because of its extensive connectivity, the ventral striatum is considered a hub of cognitive and affective integration (Cohen et al., 2009; Delgado et al., 2000; Di Martino et al., 2008; Haber and Knutson, 2010; Hare et al., 2008a; Knutson et al., 2001; 2005; Leotti and Delgado, 2011a; 2011b; Li and Daw, 2011; Li et al., 2011; O’Doherty, 2004; Pennartz et al., 2011; Wimmer et al., 2012).
Ventral striatal function and connectivity patterns supporting incentive-based valuation and learning are well characterized in studies of adults (Cohen et al., 2009; Delgado et al., 2000; Di Martino et al., 2008; Hare et al., 2008a; Knutson et al., 2005; 2001; Leotti and Delgado, 2011b; 2011a; Li et al., 2011; Li and Daw, 2011; O’Doherty, 2004; Wimmer et al., 2012), which show that ventral striatum lies at the vertex of an incentive-based learning circuit that includes the amygdala, hippocampus, medial prefrontal cortex, and insula. Each of these regions contributes uniquely to incentive-based learning. The amygdala is highly involved in rapid evaluation of potentially incentivizing stimuli and affective learning across species (Belova et al., 2007; 2008; Campeau and Davis, 1995; Paton et al., 2006; Phelps and LeDoux, 2005). Hippocampal contributions to incentive-based decision-making involve incorporating contextual information (e.g., past experiences) to bias future decisions (Barron et al., 2013; Wimmer and Shohamy, 2012) and constructing representations of incentive value for novel stimuli based on memories of related component stimuli (Barron et al., 2013). Regions of medial prefrontal cortex (e.g., orbitofrontal cortex (OFC), ventromedial PFC (vmPFC)) represent expected and experienced incentive value (Alexander and Brown, 2011; Behrens et al., 2007; Hare et al., 2008a; Kennerley et al., 2006; Kringelbach et al., 2003; O’Doherty et al., 2001; Rolls, 2000), and support representation of outcome history and action outcome predictions (Alexander and Brown, 2011; Bartra et al., 2013; Behrens et al., 2007; Kennerley et al., 2006; Phelps et al., 2014). Finally, the insula, while functionally diverse, is a key component of a valuation system (Bartra et al., 2013; Kuhnen and Knutson, 2005; Niv et al., 2012; Phelps et al., 2014; Preuschoff et al., 2008), important in evaluating risk (Kuhnen and Knutson, 2005; Niv et al., 2012; Pessiglione et al., 2006; Preuschoff et al., 2008; Seymour et al., 2004), learning from aversive incentives (Chang et al., 2013; Pessiglione et al., 2006; Preuschoff et al., 2008; Seymour et al., 2004), and integrating cognitive, affective and interoceptive information (Chang et al., 2013; Chein et al., 2011; Preuschoff et al., 2008).
Characterization of this system’s development is an active area of investigation, with task-based functional magnetic resonance imaging (fMRI) studies largely focused on ventral striatal and prefrontal function within reward-seeking contexts (for reviews see Fareri et al., 2008; Galván, 2010; Richards et al., 2013; Somerville et al., 2010). A common thread within this body of work highlights large-scale functional changes occurring within the developing ventral striatum, such as heightened responses to monetary reward receipt and to reward anticipatory cues in adolescents compared to adults (Ernst et al., 2005; Somerville et al., 2011) and children (Somerville et al., 2011) (but see also (Bjork et al., 2004; 2010; Silvers et al., 2014)). More specifically, relative to children and adults, adolescents show enhanced sensitivity to larger versus smaller monetary rewards in the ventral striatum (Galván et al., 2006), enhanced learning-related signals (Niv and Schoenbaum, 2008) in the ventral striatum (Cohen et al., 2010), and stronger responses to positive affective cues (Somerville et al., 2011). Developmental differences in ventral striatal function during reward-seeking contexts are often observed in conjunction with differences in vmPFC and OFC recruitment, such as increased vmPFC activation in adolescents during high risk decisions (though see also Eshel et al., 2007; Van Leijenhorst et al., 2010a) and increased OFC activation in adults to omitted rewards (Van Leijenhorst et al., 2010b). A large region of vmPFC has also recently been implicated as showing a linear age-related increase in activation to the expected value associated with a decision (e.g., probability of receiving a reward * amount of reward) (van Duijvenvoorde et al., 2015). The differential functional developmental trajectories of the ventral striatum and related prefrontal cortical regions may in part underlie observed increases in reward-seeking during this developmental period.
In addition to these developmental changes, the amygdala, hippocampus and insula each have their own developmental timelines. Findings suggest the amygdala to be a functionally early developing region (reviewed in Tottenham and Sheridan, 2009), showing robust reactivity during childhood and adolescence which attenuates with age (Gee et al., 2013; Guyer et al., 2008; though see also Hare et al., 2008b; Swartz et al., 2014; Tottenham et al., 2012; Vink et al., 2014), whereas hippocampal development tends to be more protracted (Gogtay et al., 2006; Payne et al., 2010; Tottenham and Sheridan, 2009)(but see also (Wierenga et al., 2014)). The insula, which shares connectivity with cortical and subcortical structures including the ventral striatum (Reynolds and Zahm, 2005) is one of the first cortical structures to develop, beginning to differentiate prenatally (Alcauter et al., 2013), though refinement of organization, function and connectivity likely continues into early adulthood (Alcauter et al., 2013; Scherf et al., 2006). Recent studies report an adolescent peak in insula recruitment related to computations of risk (van Duijvenvoorde et al., 2015) and social learning signals (Jones et al., 2014) as compared to childhood and adulthood.
Ventral striatal development has been most commonly studied in the context of its association with prefrontal cortical development, which continues into early adulthood. Many structural neuroimaging studies suggest protracted prefrontal development in comparison to striatal development (Giedd, 2004; though see also Raznahan et al., 2014; Sowell et al., 1999), often supported by differential functional developmental trajectories of these regions (Casey et al., 2010; Somerville and Casey, 2010). It is often suggested that differences in reward-seeking behavior across development may result from these differential developmental trajectories. In terms of task-based connectivity between the ventral striatum and prefrontal cortex, age-related linear increases in ventral striatal-mPFC functional connectivity have been reported between late childhood through early adulthood when receiving positive incentives (van den Bos et al., 2012). Other studies suggest differential ventral striatal-mPFC recruitment during incentive-based learning depending on age; for example, representation of incentive-based learning signals in ventral striatum, subgenual anterior cingulate cortex (sgACC) and ventrolateral PFC is more robust at younger ages, and negatively correlated with performance, whereas by adulthood, performance improves with more focal representation of such signals in vmPFC (Christakou et al., 2013). Thus while it is clear that age-related changes in ventral striatal functional connectivity manifest across childhood into adulthood, functional relationships with prefrontal cortex seem to differ as a function of specific task-based contexts (Richards et al., 2013). Further, recent work has begun to investigate development of connectivity between the ventral striatum and the insula during incentive-based processes, though noting similar patterns of connectivity between these regions in adolescents and adults, potentially due to task parameters (Cho et al., 2013). In light of these discrepancies, we sought to characterize stable functional changes in the ventral striatal functional connectivity across this broad age range, from early childhood through young adulthood, in the absence of task-based demands.
One powerful approach to addressing the development of ventral striatal functional connectivity is the use of resting-state functional connectivity (rsFC), which has emerged as a non-invasive tool with which to investigate the stability of functional neural connections (Cole et al., 2010; Raichle, 2010; Raichle et al., 2001; Utevsky et al., 2014; Van Dijk et al., 2010). rsFC provides a means to characterize functional neural organization independently of differences in task demands and sensitivity to incentives (e.g., money) which may be confounded across development. rsFC is thought to reflect local oscillations of neuronal populations (Riedl et al., 2014) necessary for maintaining stable functional relationships between neural regions often involved in related processes (Buckner and Vincent, 2007). Successful implementations of rsFC have characterized the development of both functional neural networks (Fair et al., 2010; 2008; Pizoli et al., 2011) and connectivity of specific brain structures (Delmonte et al., 2013; Di Martino et al., 2011; Gabard-Durnam et al., 2014; Qin et al., 2012). In adulthood, striatal connectivity at rest shows widespread positive connectivity with mPFC, medial temporal lobe and posterior cortical structures (Di Martino et al., 2008), and network approaches have demonstrated connectivity between the ventral striatum and both association and limbic–related networks of regions (Choi et al., 2012). On the other hand, it has been demonstrated to date that children comparatively show less connectivity with medial prefrontal structures (Di Martino et al., 2011; Greene et al., 2014; but see also Porter et al., 2014). However, considering children and adults only makes it difficult to characterize the timing of neurodevelopmental changes in the construction and organization of ventral striatal functional connectivity. Delineating these changes across early childhood, adolescence and early adulthood will both help inform an understanding of functional neural development and provide a broader context within which to consider the timing of the developmental changes in incentive-based behavior.
We investigated ventral striatal rsFC in a cross-sectional sample of typically developing individuals aged 4–23 years old. We employed a seed-based correlation approach (Gabard-Durnam et al., 2014) to probe the nature of age-related changes in ventral striatal rsFC with targeted a priori ROIs involved in incentive-based valuation processes (Bartra et al., 2013; Phelps et al., 2014; Rangel et al., 2008), including the amygdala, hippocampus, mPFC, and insula. Based on prior investigations of amygdala development and the anatomical connectivity between the amygdala and ventral striatum (Gabard-Durnam et al., 2014; Gee et al., 2013; Groenewegen et al., 1999; Tottenham et al., 2012; Vink et al., 2014), we expected stable connectivity between the ventral striatum and amygdala to emerge early in life. Further, considering evidence of hippocampal development being more protracted and its involvement in more complex value-based processes (Barron et al., 2013; Gogtay et al., 2006; Payne et al., 2010), we expected later emergence of connectivity between the ventral striatum and hippocampus. We additionally expected diverse patterns of developmental change in ventral striatal connectivity with prefrontal cortex and insula, based on evidence suggesting divergent anatomical (Alcauter et al., 2013; Giedd et al., 1999; Sowell et al., 1999; 2002) and functional (Casey et al., 2010; Jones et al., 2014; Scherf et al., 2006; Somerville et al., 2010; van Duijvenvoorde et al., 2015) developmental timelines for these regions, particularly with respect to their roles in incentive-based processes. Additionally, the development of neural systems supporting incentive-based learning, particularly striatal-prefrontal circuits, has been linked to developmental changes in sex hormone levels (Casey et al., 2010; reviewed in Kuhn et al., 2010). Individual differences in testosterone levels significantly correlate with ventral striatal response to monetary reward receipt in adolescents, with increasing levels of testosterone associated with increased reward-related BOLD responses in the ventral striatum (Op de Macks et al., 2011). Testosterone is also related to: 1) prefrontal structural development within the anterior cingulate cortex (Koolschijn et al., 2014); 2) modulation of ventral striatal function in adults (Hermans et al., 2010; Rilling, 2013); and 3) ventral striatal-prefrontal structural and functional connectivity in adults (Peper et al., 2013; 2011). In light of this literature, as well as evidence pointing to the importance of sex hormones in behavioral development (i.e., novelty-seeking, risk-taking, sexual behavior) during the transitions to adulthood (Casey et al., 2010; Schulz et al., 2009a; 2009b; Spear, 2000), we examined changes in testosterone levels as one possible mechanism underlying significant age-related changes in ventral striatal connectivity at rest. Given that testosterone has been associated with both ventral striatal function across development as well as structural maturation of the anterior cingulate cortex, we expected to see associations between testosterone and ventral striatal connectivity with anterior cingulate cortex.
2.1 Methods and Materials
2.1.1 Participants
Resting-state fMRI data were acquired for 66 participants between the ages of 4.5 and 23 years old (35F/31M, mean = 13.03, sd = 4.89; see Figure 1 for a distribution of participants’ ages). 55 individuals in this sample were also included in another recent study from our group investigating the development of amygdala functional connectivity at rest (Gabard-Durnam et al., 2014). We recruited an ethnically diverse sample: European- (40.9%), Asian- (25.8%), African-American (21.2%), Native Hawaiian/Pacific Islander (7.6%), and American Indian/Alaska Native (4.5%). 16.7% of participants in this sample identified as Hispanic/Latino. All participants over the age of 18 were enrolled undergraduate students in Southern California, recruited through advertisements on campus. Children and adolescents were recruited for this study via local advertisements and state birth records obtained through the Developmental Subject Pool. All participants were screened over the phone for physical and psychological contraindications for MRI, including psychotropic medication status, developmental disabilities and neurological disorders. In lieu of a diagnostic interview to exclude individuals with a psychiatric disorder, we assessed participants to ensure they were within the normal range on clinical measures of socio-affective function, internalizing illness and intelligence. We screened child and adolescent participants in the laboratory with paper assessments of mental health including the Child Behavior Checklist (CBCL; (Achenbach, 1991)) and the Revised Children’s Anxiety and Depression Scale (Chorpita et al., 2000). All but one participant scored below the clinical cutoff T scores of 63 for CBCL total problems (mean = 44.98, sd = 10.08); however, we elected to include this individual in our final sample because they did not prove to be an outlier in any analysis, and because they were within normal limits on anxiety and internalizing scales of the RCADS. All participants were below borderline clinical cutoff T scores (65) on the RCADS total anxiety (mean = 34.75, sd = 4.35) and internalizing (mean = 33.86, sd = 4.73) scales. Measures of trait anxiety (STAI (Spielberger, 1983)) were collected for adult participants, with the mean score for our adult participants falling within normal limits (mean = 39.4, sd = 7.81). Adult participants or parents of child and adolescent participants provided written consent for participation in this study. Participants between the ages of 6 and 17 years old were assessed on cognitive ability via the Wechsler Abbreviated Scale of Intelligence (Weschsler, 1999). The average full-scale intelligence quotient of this sample was 113.2 (sd = 17.1). The data presented here were collected at the end of a larger experimental session examining neuroaffective development via the use of task-based functional neuroimaging; all participants received financial compensation ($80) for taking part in the study. This study was approved by local Institutional Review Boards.
Figure 1.
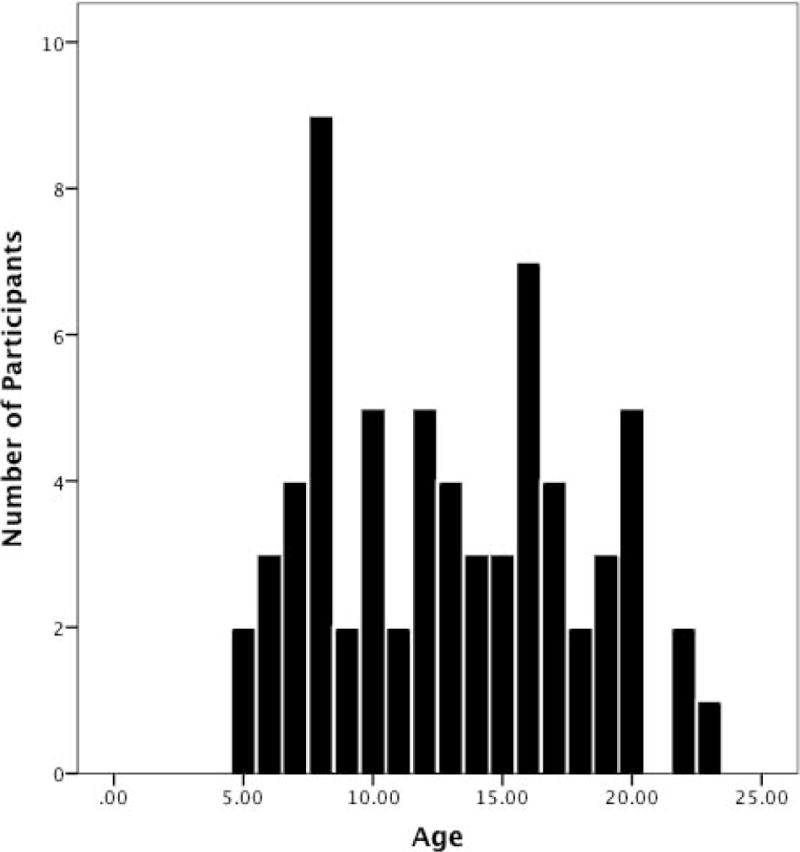
Distribution of participants’ age.
2.1.2 Procedures
2.1.2.1 fMRI data acquisition
Prior to the experimental session, all participants were acclimated to the scanner environment in a mock scanner. The resting state scan was performed at the end of a forty-five minute scanning session, which included task-based scanning. The resting state sequence was preceded by approximately fifteen minutes of structural neuroimaging sequences to minimize influence from the previous task-based sequences (Gabard-Durnam et al., 2014). As motion artifacts are a concern in both studies of resting-state connectivity and developmental neuroimaging studies, care was taken to minimize head motion at the outset of the experimental session, through the use of a vacuum-packed pillow and additional padding placed all around participants’ heads. Additional and extensive motion correction procedures were applied to the acquired data (see section 2.1.2.3 below). For the resting-state sequence, participants were instructed to remain still with their eyes closed (without falling asleep). We also presented participants with a white fixation cross on a plain black screen in the event they did not keep their eyes closed. Participants were able to view this display through MR-compatible video goggles (Resonance Technology, Inc). We assessed alertness via direct observation upon completion of the scan session and self-report.
Structural and functional data were acquired using a Siemens Trio 3T scanner with a 12-channel head coil. Anatomical images were collected using a T1-weighted MPRAGE sequence prior to the resting-state scan (in-plane resolution: 256 × 256, FOV: 256mm, 192 1 × 1 × 1mm slices). Resting-state data were acquired in a 6-minute, T2-weighted echoplanar imaging sequence in accordance with the following parameters: 33 oblique-axial slices, voxel size = 3.4 × 3.4 × 4.0 mm, slice thickness = 4mm, FOV = 220mm, 64×64 matrix, TR = 2000ms, TE = 30ms, flip angle = 75 degrees. Evidence suggests that a sequence of 6 minutes in length is sufficient for detection of reliable and stable functional correlations between regions at rest (Van Dijk et al., 2010). Oblique axial slices were acquired at an angle of approximately 20–30 degrees in order to maximize functional coverage while minimizing drop-out.
2.1.2.2 fMRI data preprocessing
Processing and analyses of neuroimaging data was performed using Analysis of Functional Neuroimages (AFNI) software (Cox, 1996). The first four volumes of the resting state run were discarded in order to account for the stabilization of the BOLD signal. Standard preprocessing steps were initially performed on functional data: slice-time correction, six parameter motion correction, and Gaussian spatial smoothing using a 6mm-FWHM smoothing kernel. Volumes of data exhibiting a change in motion of more than 2.5mm from the reference volume were excluded from analyses (i.e., censored). The mean length of participants’ resting state session was 168 volumes (sd = 19.04). One participant’s data included more than 3sd below the mean of 168 usable volumes; however, this participant proved not to be an outlier in any analysis, and as such was included in the final sample of data reported here. For all analyses, functional data were normalized to percent signal change, registered to anatomical images, warped to Talairach space (Talairach and Tournoux, 1988), and resampled to a resolution of 1mm3. We note that a number of studies have supported the notion that registration of children’s data to standard coordinate spaces based on adult brains (e.g., Talairach and Tournoux, MNI) is methodologically appropriate (Burgund et al., 2002; Kang et al., 2003). Previous work from our laboratory further supports this (Gabard-Durnam et al., 2014), demonstrating that anatomical averages of subcortical nuclei for developmental samples significantly overlap with the adult template, suggesting that differences in registration across development was not a confounding factor in analyses.
2.1.2.3 Motion Correction
BOLD signal fluctuations reflecting rsFC are highly susceptible to motion confounds (Power et al., 2012; Van Dijk et al., 2012), particularly for developmental samples (Cole et al., 2010; Satterthwaite et al., 2012; Van Dijk et al., 2012; 2010). Thus a number of additional processing steps were taken in order to guard against spurious influence of motion here. A conservative bandpass filter (0.009Hz<ƒ<0.08Hz) was applied to filter out high-frequency signals which may demonstrate a stronger sensitivity to motion artifacts in resting-state data (Satterthwaite et al., 2012). We performed standard motion correction (3 translation and 3 rotation parameters) and included these as regressors of no interest in our analyses at the single subject level. The first temporal derivatives associated with motion in each plane were also included as single subject regressors of no interest (Van Dijk et al., 2012; 2010). Influences of submillimeter motion and physiological effects are also potential confounds in resting-state fMRI, particularly in developmental samples. We accounted for these additional sources of spurious influence with regressors of no interest modeling white matter intensity, physiological noise in cerebrospinal fluid, and the mean global signal across the brain. These signals were regressed out at the single subject level using a simultaneous regression procedure (as opposed to bandpass filtering prior to regression) in order to avoid both inflated resting-state correlations and reintroducing noise into the signals of interest (Hallquist et al., 2013). We also included a group level motion regressor––mean framewise displacement––in our whole brain and ROI group analyses to further account for potential motion confounds. Mean framewise displacement (FD) was calculated by subtracting the translation values at every timepoint in each plane from the previous timepoint, taking the average of the absolute value of the change in motion at each timepoint, and then computing the square root of the sums of squares of these values (Gabard-Durnam et al., 2014; Van Dijk et al., 2012).
We note the current debate in the literature regarding the use and utility of regressing out the mean global signal at the individual participants level. While some evidence suggests that including global signal as a regressor of no interest in studies of rsFC may induce unwanted and spurious effects (Saad et al., 2013; Satterthwaite et al., 2013), additional recent evidence suggests that inclusion of the global signal may be useful (Miranda-Dominguez et al., 2014), particularly when additional physiological measures are not available (Chen et al., 2012). Because we did not collect physiological measures of heart rate and respiration, and because standard physiological toolboxes (e.g., PESTICA, IRF-RETROICOR (Beall, 2010; Beall and Lowe, 2007)) to generate physiological regressors are not optimized for developmental samples––i.e., physiological noise from heart rate and respiration may differ across development––, we chose to regress out the mean global signal at the single participant level. However, supplemental analyses were conducted without modeling global signal as a regressor of no interest to assess whether our results were influenced by spurious negative correlations (Gabard-Durnam et al., 2014; Hampson et al., 2010). For this supplemental analysis, we used estimates of whole brain correlation with the ventral striatum timeseries as a group level regressor. We computed the average correlation between the resting state timecourse of the bilateral ventral striatal ROI and the rest of the brain, obtaining a single correlation coefficient for each participant. These values were subsequently entered into our group analyses as a group level regressor of no interest. Results are presented in the Appendix A. We note, however, that this alternative analysis without global signal does not account for the influence of physiological noise in the resting-state data, leading to potential difficulties in interpretation and additional confounds that cannot be addressed.
2.1.2.4 Ventral striatal seed region and timeseries extraction
We chose a bilateral anatomical region of interest (ROI) encompassing the ventral striatum from the Oxford-GSK-Imanova structural striatal atlas (Tziortzi et al., 2011) included within the FSL software package (FMRIB, Oxford, UK). This anatomical ROI encompasses the nucleus accumbens, ventral caudate and ventral putamen (Figure 2). We chose this conservative ROI, as opposed to placing smaller seeds within different portions of the ventral striatum (Di Martino et al., 2011; 2008) as it can be difficult in human and non-human primates to clearly distinguish separable anatomical markers for ventral striatal subregions, with delineations most clearly characterized by differences in the regions from which afferents are received, and in the neurotransmitters and receptors present (Haber et al., 2006; Haber and Knutson, 2010; Haber and McFarland, 1999). We elected to choose a bilateral ROI because we had no a priori hypotheses regarding laterality differences in ventral striatal connectivity. Previous developmental studies of striatal connectivity (Porter et al., 2014) report results averaged across left and right ventral striatal seeds as minimal differences emerged in connectivity with the rest of the brain. This ROI was transformed to standard Talairach space in AFNI and overlaid onto each individual’s resting state functional data for timeseries extraction. To ensure that voxels containing white matter were not included in the analyses, individual white matter masks were created for each subject’s anatomical images using FSL’s FAST segmentation toolbox. A probability threshold of approximately 0.55 was employed by FAST to categorize voxels as white matter. We then subtracted out any voxels containing overlap between the anatomical ventral striatal ROI and an individual’s white matter mask using the 3dcalc function in AFNI. Resulting ventral striatal seed ROI masks were manually inspected for all subjects and manually adjusted within AFNI when necessary.
Figure 2.
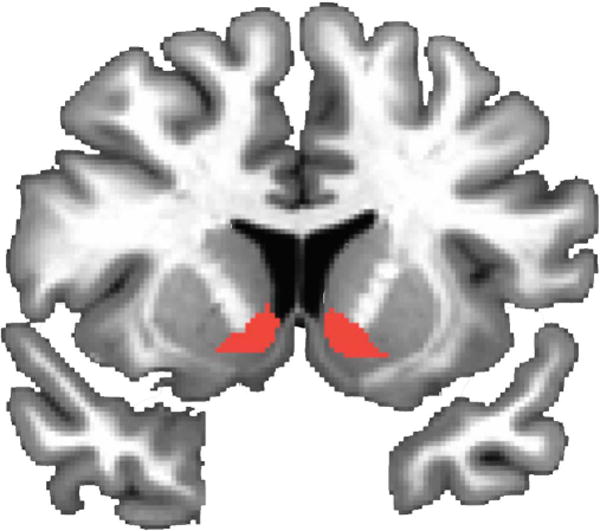
Bilateral ventral striatum seed region. A bilateral ventral striatum seed region encompassing the nucleus accumbens, ventral caudate and ventral putamen was chosen from the Oxford-GSK-Imanova Structural–Anatomical Striatal Atlas (Tziortizi et al., 2011). ROI masks were overlaid on individual participants’ brains; overlap between mask placement and white matter was subtracted out for each individual participant. ROI depicted at y = 7.
The timeseries was extracted from the ventral striatal ROI for all subjects after simultaneous regression to filter and remove effects of regressors of no interest. Whole brain multiple regression was performed at the single subject level using the 3dREMLfit program in AFNI to generate single subject whole brain maps showing regions demonstrating resting state functional connectivity with the ventral striatum. 3dREMLfit employs a least-squares restricted maximum likelihood estimation via an auto-regressive moving average (ARMA(1,1)) model to determine the best fit for each voxel and to correct for serial correlations within the residuals of the regression model.
2.1.2.5 Group Connectivity Analyses
2.1.2.5.1 Whole Brain Analysis: Age-constant connectivity
To characterize connectivity independent of any potential developmental differences, we conducted a whole-brain analysis holding age constant (Gabard-Durnam et al., 2014). Using the 3dttest++ program in AFNI, we performed a whole brain ANCOVA using individual participants’ ventral striatal timeseries as the dependent variable of interest, entering age as a mean centered regressor. This analysis highlights connectivity between the ventral striatal seed region and the rest of the brain that is significantly different from zero (i.e., either positive or negative connectivity). Mean framewise displacement (FD) served as a group level motion regressor. The resulting statistical parametric map (SPM) thus reveals significant connectivity with the ventral striatum when controlling for FD. Whole-brain results were set to an initial height threshold of p<.01 and whole-brain cluster corrected to p<.05 using the 3dClustSim program in AFNI, which conducts a series of Monte Carlo simulations over the whole brain to determine adequate correction for multiple comparisons and detection of false positive activations. Local maxima within activated clusters in this whole brain analysis were determined using the 3dExtrema program in AFNI.
2.1.2.5.2 Targeted ROI analyses
Targeted region of interest (ROI) analyses were conducted to specifically investigate the development of resting state connectivity within regions involved in incentive-based learning and valuation. We specifically targeted subcortical structures including the amygdala and hippocampus, as well as regions of prefrontal cortex––orbitofrontal cortex (OFC), ventromedial prefrontal cortex (vmPFC) and anterior cingulate cortex (ACC). As we had no a priori hypotheses regarding laterality differences, bilateral anatomical ROIs encompassing these regions were taken from the Harvard Oxford atlas in FSL (amygdala, hippocampus, OFC, vmPFC) and the Talairach Daemon atlas within AFNI (ACC). Because functional dissociations have been characterized in both human (anterior/posterior) and non-human animals (ventral/dorsal) (Fanselow and Dong, 2010; Moser and Moser, 1998; Satpute et al., 2012), we also examined connectivity with the anterior and posterior portions of the hippocampus separately based on coordinates from Kahn and colleagues (Kahn et al., 2008) by combining 3mm spheres of the two most anterior and two most posterior coordinates. The ACC mask was segmented into divisions roughly corresponding to the subgenual ACC/BA25, ventral ACC, perigenual ACC, and an anterior dorsal ACC region (Bush et al., 2000); these ACC subregions comprise portions of the ACC largely implicated in affective processing. The sgACC subregion was segmented based on overlap between the ACC mask and a mask encompassing BA25. All other ACC subregions were segmented based on divisions in the axial plane using coordinates from Bush, Luu and Posner (Chang et al., 2013; Pessiglione et al., 2006; Preuschoff et al., 2008; Singer et al., 2009): ventral ACC, z = −10:−1; pgACC, z = 1:9; adACC, z = 10:20). We additionally investigated ventral striatal connectivity with the insula, given the insula’s broad role in affective valuation and learning (Chang et al., 2013; Deen et al., 2011; Kelly et al., 2012). As evidence suggests functional subdivisions of the insula (Kelly et al., 2012), we segregated the insula into ventral anterior, dorsal anterior and mid-posterior clusters as per Kelly et al., (2012).
All ROI masks were resampled to 1 × 1 × 1 mm, and warped to Talairach atlas space in AFNI. Masks were tailored to individual participants’ anatomy by subtracting out any overlap between the anatomical ROI mask and an individual participant’s white matter mask. Parameter estimates (ß) indexing connectivity between each ROI and bilateral ventral striatum were extracted for each subject and entered into separate linear regression models in the R statistical language with age and FD as regressors. We also tested for quadratic effects of age with respect to ventral striatal connectivity with each of our targeted ROIs by including age, age-squared and FD as regressors. Quadratic analyses thus controlled for any linear effects. For tests of quadratic effects of age on connectivity only those that emerged as significant or trending are reported. Any age-related effects––linear or quadratic––emerge when controlling for effects of FD. For significant or trending linear effects, we also report the partial correlation between age and parameter estimates (ß) indexing connectivity between the VS and target ROIs, controlling for FD; for significant or trending quadratic effects, we performed partial correlations between age-squared and parameter estimates (ß) indexing connectivity between the VS and target ROIs, controlling for both linear effects of age and FD. The partial correlation value serves as an index of effect size. Because we tested multiple ROIs within larger regions (e.g., 6 mPFC ROIs), we applied a Sequential Bonferroni Correction (Holm, 1979; Rice, 1989). This correction method requires ordering all post-hoc comparisons within a family of tests in order of significance and seeing if the most significant meets a corrected p-value of .05/number of tests. Thus, within the mPFC, we tested 6 ROIs, so the most stringent test would have to meet a corrected level of significance of p<.05/6, or p<.0083. The next most significant test would be subject to a corrected p-value of .05/5, or p<.01, and so on. Within the insula and hippocampus, there were 3 regions tested in each, and so the most significant result within a family of tests would be subject to a corrected level of p<.05/3, or p<.016.
Statistical treatment of Age
Regression analyses were performed with age as a continuous predictor; however we additionally performed post-hoc one-sample t-tests within each age group of average parameter estimates (ß) reflecting connectivity against 0 (children: 4–10 (n=27); adolescents: 11–17 (n=28), adults: 18–23 (n=11)) for interpretation and display purposes. We note that significant results reported below do not change substantially when removing outliers whose rsFC parameter estimates were ±3sd outside of the sample mean (see Appendix A for additional results). To correct for multiple post-hoc comparisons, within each ROI, we applied a Sequential Bonferroni Correction to these analyses as well. In this case, because we were testing whether children, adolescents and adults each demonstrated rsFC with the ventral striatum that was significantly different from zero, the most significant result within an ROI needed to meet a corrected p-value of .05/3, or p<0.016. The next most significant test would be subject to a corrected p-value of .05/2, or p<.025, and the least significant test would have to meet a p-value of p<.05. For each ROI, if these criteria were not met, we have noted that results were trends or not significant after correction. For these post-hoc analyses, we also computed the correlation estimates (r) between nuisance residualized VS timeseries and target ROIs at the single subject level and report the average of these correlation estimates per age group to get an average measure of strength of coupling per age group.
2.1.2.5.3 Supplemental Whole-Brain Age-related Analyses
We additionally performed whole-brain voxelwise analyses examining linear and quadratic age-dependent changes in ventral resting-state connectivity. Methodological procedures and associated results for these whole brain analyses are described in full in Appendix A.
2.1.3 Hormonal Level Assessment
Recent evidence suggests that testosterone may play a role in development of ventral striatal function during reward-processing (Op de Macks et al., 2011), and in structural development (Koolschijn et al., 2014) of cortical components of incentive-based learning circuitry (Peper et al., 2013; 2011). Testosterone was assayed from saliva samples from a subset of participants (50 total, 23 female/27 male). Saliva (2 samples from the evening – 5:00PM and 8:00 PM) was assayed by Dresden Lab Service (Dresden, Germany). Salivary testosterone concentrations were measured using commercially available chemiluminescence-immuno-assays with high sensitivity (IBL International, Hamburg, Germany). The intra and interassay coefficients for testosterone are below 11%. We computed an average testosterone level for each subject. A log transformation was applied to account for positive skew in raw testosterone levels. We subsequently accounted for differences in average levels of testosterone production in males and females (t(47.5) = 1.95, p = .057; independent samples t-test, equal variances not assumed) by subtracting the minimum value within each group from individual testosterone levels for each subject, and then scaling this result by dividing by the difference between maximum and minimum values within each group (t(47.6) = 1.05, p > 0.3 after accounting for differences in production values) to obtain a percent of maximum possibility score for each participant (Cohen et al., 1999). Given effects of endogenous testosterone on structural development of ACC (Koolschijn et al., 2014), as well as on ventral striatal functional activation during reward-based tasks in developmental samples (Op de Macks et al., 2011), we had an a priori hypothesis that there would be an association between testosterone and ventral striatal connectivity with ACC subregions. We specifically looked in only in ACC regions that demonstrated a significant age-related change in connectivity with the ventral striatum. We conducted a non-parametric bootstrapping mediation analysis using the PROCESS macro in SPSS (Hayes, 2013; 2012) to assess whether circulating levels of salivary testosterone mediated significant age-related changes in connectivity between the ventral striatum and ACC subregions. Path coefficients, standard errors and significance levels are reported for the effects of age on salivary testosterone (path a), as well as for the effect of salivary testosterone on ventral striatal-sgACC connectivity (path b). Mediation was assessed via estimation of indirect and direct effects of age on ventral striatal-sgACC connectivity. Point estimates and 95% bias-corrected confidence intervals (Efron, 1987; Hayes and Scharkow, 2013) are reported for indirect and (ab) and direct effect (c′) paths. Confidence intervals not crossing zero indicate significant effects. Mean framewise displacement and gender were included as covariates of no interest in the mediation model.
3.1 Results
Whole Brain Analysis: Age Constant Connectivity
An ANCOVA examining resting state functional connectivity with bilateral ventral striatum revealed widespread positive coupling with a number of prefrontal cortical and subcortical regions when holding age constant (see Figure 3 and Table S1). Ventral striatal connectivity with medial prefrontal cortex was particularly robust, with positive connectivity emerging across our sample in multiple portions of the anterior cingulate and medial frontal gyrus, among other regions. Negative connectivity with the ventral striatum at rest was less widespread, but emerged primarily in lateral and posterior cortical regions such as superior temporal gyrus, fusiform gyrus, and precuneus, as well as in the including the cerebellum and extending to the brainstem (see Figure 3 and Table S1).
Figure 3.
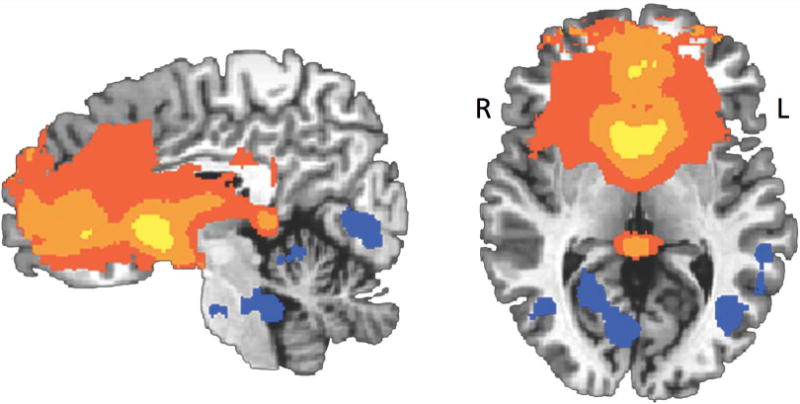
Age-constant connectivity. A whole brain ANCOVA holding age-constant (mean-centered) was conducted to highlight positive and negative connectivity that was significantly different from zero. Robust positive connectivity emerged between the ventral striatum and medial prefrontal cortex, as well as a number of subcortical structures including the brainstem, amygdala, and portions of parahippocampal gyrus.
Targeted ROI Analyses1
Our primary aim in this study was to probe age-related changes in ventral striatal connectivity with regions that have been shown to be highly connected during adulthood (including mPFC, amygdala, hippocampus and insula). We report age-related regression analyses performed on parameter estimates (ß) indexing connectivity between the ventral striatum and each ROI, and for all age-related analyses we also report results of a partial correlation (Partial r) between parameter estimates and age, controlling for FD, or between parameter estimates and age-squared, controlling for age (linear) and FD, which serves as a measure of effect size. We additionally report results of post-hoc t-tests against zero conducted on parameter estimates (ß), and as a measure of strength of coupling for these post-hoc tests we report the average correlation (r) between the ventral striatal and each ROI timeseries per age group. These results are also presented in Table 1. We note that supplemental whole brain age-related analyses are reported in Appendix A and generally support the results reported below.
Table 1.
Summary of ventral striatal rsFC with targeted regions of interest
|
|
||||||||
|---|---|---|---|---|---|---|---|---|
| Region of Interest | Subregion of Interest | Effect Size (Partial R) for Linear Age-Related Changes | Significance (p), Linear Age-Related Changes | Effect Size (Partial R) for Quadratic Age-Related Changes | Significance (p), Quadratic Age-Related Changes |
Average VS-ROI timeseries correlation by age group [r]
|
||
| Children | Adolescents | Adults | ||||||
| Medial Prefrontal Cortex | ||||||||
| Subgenual Anterior Cingulate Cortex (sgACC) | −0.39 | * | n.s. | 0.43 | 0.32 | 0.27 | ||
| Ventral Anterior Cingulate Cortex | −0.23 | †† | n.s. | 0.33 | 0.23 | 0.17 | ||
| Perigenual Anterior Cingulate Cortex (pgACC) | −0.22 | †† | n.s. | 0.35 | 0.23 | 0.28 | ||
| Anterior Dorsal Anterior Cingulate Cortex (adACC) | n.s. | n.s. | 0.29 | 0.17 | 0.26 | |||
| Orbitofrontal Cortex (OFC) | n.s. | n.s. | 0.18 | 0.13 | 0.09 | |||
| Ventromedial Prefrontal Cortex (vmPFC) | n.s. | n.s. | 0.20 | 0.14 | 0.10 | |||
| Hippocampus | 0.25 | † | n.s. | 0.03 | −0.02 | 0.15 | ||
| Anterior Hippocampus | 0.29 | † | n.s. | −0.04 | −0.03 | 0.12 | ||
| Posterior Hippocampus | n.s. | 0.33 | * | 0.02 | −0.14 | 0.05 | ||
| Amygdala | n.s. | n.s. | 0.15 | 0.06 | 0.16 | |||
| Insula | Ventral Anterior Insula | n.s. | 0.25 | † | 0.12 | −0.02 | 0.07 | |
| Dorsal Anterior Insula | −0.24 | †† | 0.23 | †† | 0.21 | 0.04 | 0.07 | |
| Mid/Posterior Insula | n.s. | 0.32 | * | 0.06 | −0.09 | 0.01 | ||
Note: Effect Sizes for age-related changes derived from partial correlations conducted on parameter estimates (ß) indexing an age-connectivity relationship.
Note: Average VS-ROI timeseries correlations reflect average of single subject VS-ROI timeseries correlations per age group.
denotes p<.05
denotes p<.10, trend after Sequential Bonferroni Correction
denotes p<.10, n.s. after Sequential Bonferroni Correction
3.1.1 Medial Prefrontal Cortex
We examined ventral striatal connectivity with six specific subregions of mPFC. There was significant positive coupling between the VS and all ROIs examined within the mPFC, and across most regions this coupling was robust and present in our youngest participants (see Figure 4). We observed significant age-related decreases in ventral striatum coupling with subgenual ACC/BA25 as a function of age (ß = −.03, SE = .008, t = −3.44, p<.001; Partial r = −0.39 Figure 4a), which was strongly positively coupled in childhood and became less coupled with increasing age (t-tests on ß vs.0, mean timeseries correlation: children: t(26) = 7.68, p < .001, r = 0.42; adolescents: t(27) = 5.39, p < .001, r = 0.32; adults: t(10) = 4.23, p < .005, r = 0.27). Similar trend level declines in coupling were observed in ventral ACC (ß = −0.02, SE = 0.01, t = −1.88, p = .06; not significant after Sequential Bonferroni Correction; Partial r = −.23 Figure 4b; children: t(26) =6.35, p < .001, r = .33; adolescents: t(27) = 4.74, p < .001, r = .23; adults: t(10) = 2.05, p =.07, r = .17) and perigenual ACC (ß = −.02, SE = .01, t = −1.81, p =.07; not significant after Sequential Bonferroni Correction; Partial r = −.22 Figure 4c; children: t(26) = 10.41, p < .001, r = .35; adolescents: t(27) = 5.62, p < .001, adults: t(10) = 6.46, p < .001, r = .27)).
Figure 4.
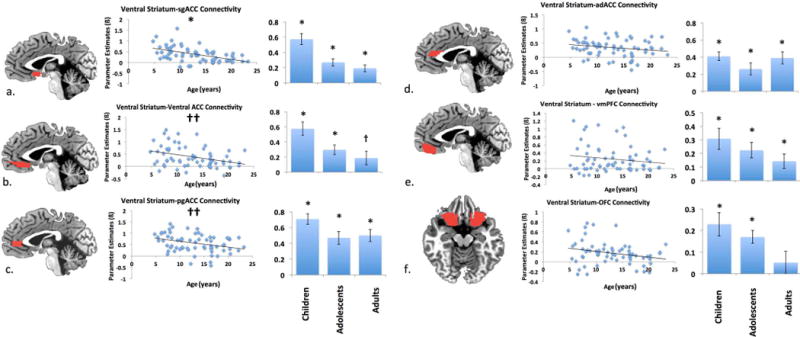
Ventral striatum-medial prefrontal cortex connectivity. Significant positive connectivity was present in all mPFC regions (subgenual ACC (a.), ventral ACC (b.), perigenual ACC (c.), anterior dorsal ACC (d.), vmPFC (e.), OFC (f.)) across age. Significant linear age-related decreases in connectivity emerged in ventral striatal-sgACC connectivity. Linear age-related decreases in connectivity with the ventral ACC and pgACC, become non-significant after Sequential Bonferroni Correction. * denotes significant differences from zero (p<.05). † denotes trend level differences from zero (p<.10). †† denotes not significant after Sequential Bonferroni Correction (p<.10). ROIs depicted at x = 2 (panels a-e) and z = −13 (panel f).
Consistent with our whole brain age-constant analysis, positive connectivity emerged between the ventral striatum and a number of mPFC regions which did not vary with age including adACC (ß = −.01, SE = .01, t = −1.11, p > 0.25; Figure 4d;), vmPFC (ß = −.006, SE = .009, t = −.70, p > 0.45; Figure 4e), and OFC (ß = −.002, SE = .005, t = −0.45, p > 0.6; Figure 4f;). In these regions, connectivity remained consistently positive across children, adolescents and adults [adACC: (children: t(26) = 8.01, p < .001, r = .29; adolescents: t(27) = 4.74, p < .001, r = .17; adults: t(10) = 5.64, p < .001, r = .26); vmPFC: (children: t(26) = 3.94, p < .001, r = .20; adolescents: t(27) = 3.95, p < .001, r = .14; adults: t(10) = 2.63, p < .025, r = .14); OFC: (children: t(26) = 4.21, p < .001, r = .18; adolescents: t(27) = 5.63, p < .001, r = .13; adults: t(10) = 1.00, p > 0.3, r = .09)]. No significant quadratic effects of age on ventral striatal-mPFC connectivity emerged.
3.1.2 Amygdala
There were no significant age-related changes in ventral striatal-amygdala connectivity (ß = −.001, SE = 0.009 t = −.11, p > 0.90; Figure 5a). Significant positive coupling between the ventral striatum and bilateral amygdala emerged across all participants (children: t(26) = 3.17, p < .005, r = .15; adolescents: t(27) = 2.96, p < .01, r = .06; adults: t(10) = 3.73, p < .005, r = .18).
Figure 5.
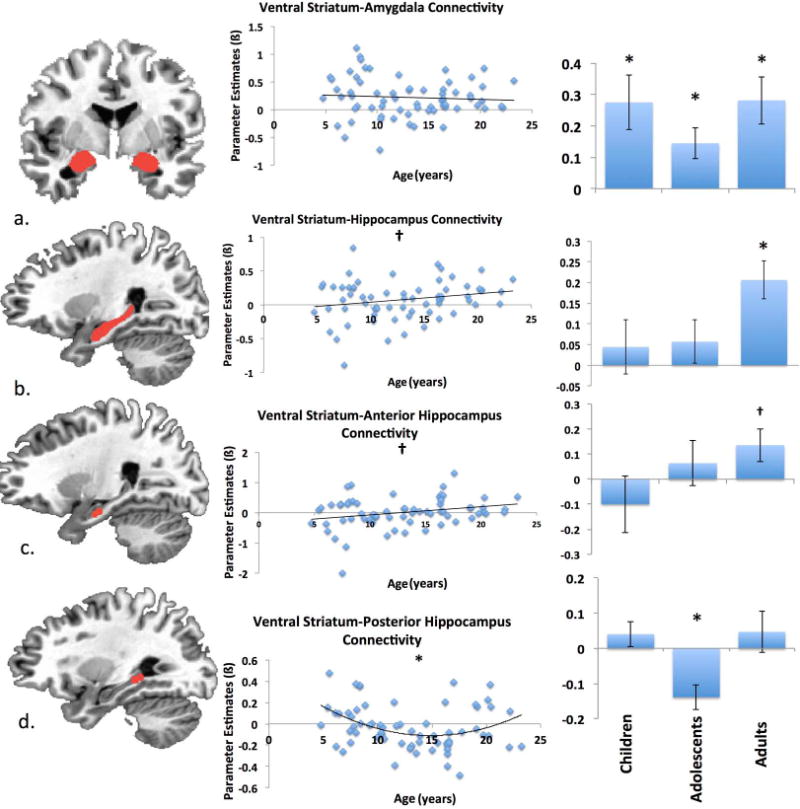
Ventral striatum-limbic connectivity. Ventral striatal-amygdala connectivity remained significantly positive across age (a). Positive connectivity between the ventral striatum and hippocampus was observed only in adulthood (trend after Sequential Bonferroni Correction) (b), which reflected the anterior portion’s connectivity changes (trend after Sequential Bonferroni Correction) (c) as opposed to posterior (d) portion of the hippocampus which demonstrated a significant quadratic age-related relationship. * denotes significant differences from zero (p<.05). † denotes trend level differences from zero (p<.10). ROIs depicted at y = −4 (panel a), x = 25 (panel b), x = 23 (panel c) x = 27 (panel d) .
3.1.3 Hippocampus
Significant positive age-related changes were observed in ventral striatal-hippocampal connectivity (ß = 0.02, SE = 0.008 t = 2.02, p <0.05; trend after Sequential Bonferroni Correction; Partial r = .25; Figure 5b) characterized by no significant coupling between these two regions in children (t(26) = 0.68, p > 0.50, r = .03) and adolescents (t(27) = 1.10, p > 0.20, r = −.02), but positive coupling emerging in adults (t(10) = 4.46, p <.002, r = .16). When examining the anterior and posterior hippocampus separately, age-related increases in connectivity with ventral striatum emerged in the anterior hippocampus (ß = .03, SE = .01, t = 2.40, p <.02; trend after Sequential Bonferroni Correction; Partial r = .29; Figure 5c), such that there was no significant connectivity in children (t(26) = −0.88, p > .39, r = −.04) and adolescents (t(27) = 0.71, p > 0.4, r = −.02), but these two regions were positively coupled in adulthood (t(10)= 2.07, p = .07; trend after Sequential Bonferroni Correction, r = .12).
Posterior hippocampus did not show significant linear age-related change (ß = −.005, SE = .006, t = −.94, p >.30; Figure 5d). However, adding a quadratic term to our model revealed a significant quadratic effect of age (ß = .003, SE = .001, t = 2.72, p <.009, Partial r = .33), such that adolescents demonstrated significant negative coupling between ventral striatum and posterior hippocampus (t(27) = −3.98, p <.001, r = −.14). Connectivity in children (t(26) = 1.14, p > 0.26, r = .02) and adults (t(10) = 0.81, p > 0.4, r = .05) was not significantly different than zero (see Appendix A for additional alternative global signal results).
3.1.4 Insula
Ventral striatal-insula connectivity demonstrated diverse patterns across three subdivisions of insular cortex. Analyses testing for linear effects of age on connectivity between the ventral striatum and insula revealed a trend of a relationship only in the dorsal anterior insula (ß = −.02, SE = .008, t = −1.92, p = .06; Partial r = −.24), which does not survive correction for Sequential Bonferroni Correction. No linear effects of age on connectivity between the ventral striatum and either ventral anterior insula (ß = −0.009, SE = .008, t = −1.15, p > 0.25) or posterior insula (ß = −0.01, SE = .007, t = −1.44, p > 0.15).
Adding a quadratic term to our analyses of ventral striatal-insula connectivity revealed significant quadratic effects (controlling for linear effects) of age on connectivity between the ventral striatum and posterior insula (ß = .004, SE = .001, t = 2.69, p < .01; Partial r = .32 Figure 6a). Similar quadratic patterns were also observed in ventral striatal-ventral anterior insula connectivity (ß = .003, SE = .002, t = 2.01, p < .05; trend after Sequential Bonferroni Correction; Partial r = .25; Figure 6b) and between the ventral striatum and dorsal anterior insula (ß = −.003, SE = .002, t = 1.88, p < .06; not significant after Sequential Bonferroni Correction; Partial r = .23; Figure 6c). Quadratic effects in ventral striatal-posterior insula connectivity were characterized by negative connectivity in adolescents (t(27) = −2.36, p < .03; trend after Sequential Bonferroni Correction, r = −.09), but no significant connectivity in children (t(26) = 1.66, p = .11, r = .06) or adults (t(10) = 0.36, p > 0.73, r = .01). Trend-level quadratic changes in ventral striatal connectivity with both the ventral and dorsal anterior insula were both characterized by positive connectivity in children (ventral anterior insula: t(26) = 3.11, p <.005, r = .11; dorsal anterior insula: t(26) = 4.03, p < .0005, r = .21), but not in adolescents (ventral anterior insula: t(26) = −.32, p >.70, r = −.02; dorsal anterior insula: t(27) = 1.17, p > 0.2, r = .04) or adults (ventral anterior insula: t(10) = 0.79, p > 0.40, r = .07; dorsal anterior insula: t(10) = 1.26, p > 0.2, r = .07) (see Appendix A for results of alternative global signal analysis).
Figure 6.
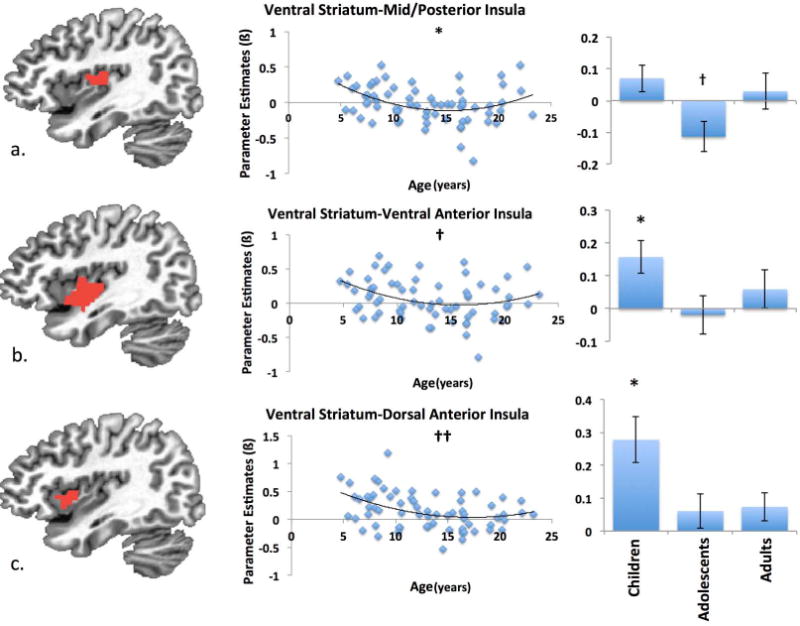
Ventral striatum-insula connectivity. Connectivity between the ventral striatum and mid/posterior insula (a.) demonstrated a significant quadratic relationship, with connectivity being negative during adolescence, but not in childhood or adulthood. Quadratic effects in connectivity between ventral striatum and ventral anterior insula (b.), and between ventral striatum and dorsal anterior insula became trends and non-significant, respectively, after Sequential Bonferroni Correction. (c.). * denotes significant differences from zero (p<.05). † denotes trend level differences from zero (p<.10). †† denotes not significant after Sequential Bonferroni Correction (p<.10). All ROIs depicted at in x = 35.
3.1.5 Associations with Age-related Change in Testosterone
We tested our a priori hypothesis that there would be an association between testosterone and significant age-related changes in ventral striatal connectivity with ACC subregions. We specifically tested whether age-related linear decreases in ventral striatal-sgACC connectivity, which were most significant between childhood and adolescence (t(45.67) = −3.39, p < .002) as opposed to between adolescence and adulthood (t(31.77) = 1.16, p > 0.25), were mediated by changes in circulating testosterone levels, controlling for mean framewise displacement and gender. A significant effect of age on salivary testosterone levels (a = 0.03, SE = 0.008, p<.001) (we note that separate linear regression analyses indicate that this effect was present in both males (ß = .038, SE = .01, t = 3.70, p < .001) and females (ß = .03, SE = .01, t = 2.70, p <.02)), and a significant effect of salivary testosterone levels on ventral striatal-sgACC connectivity (b = −0.42, SE = 0.20, p<.05; Figure 7a). To assess mediation, non-parametric bootstrapping revealed a significant indirect effect between age and ventral striatal-sgACC connectivity through salivary testosterone (ab = −0.01, BC CI: [−0.04, −0.001]), which weakened the direct effect of age on connectivity alone (c′ = −0.017, BC CI: [−0.04, 0.10]) (Figure 7b).
Figure 7.
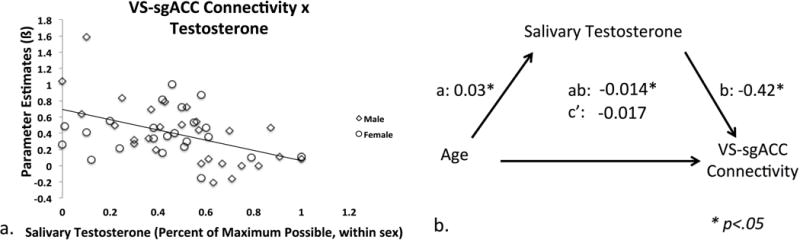
Mediation of age-related decreases in ventral striatal-sgACC connectivity by testosterone. (a.) Salivary testosterone levels demonstrated a significant negative linear relationship with ventral striatal-sgACC connectivity. (b.) The relationship between age and ventral striatal-sgACC connectivity was significantly mediated by salivary testosterone. * denotes p<.05.
4.1 Discussion
In this paper, we examined the development of the core neural circuits involved in incentive-based learning across childhood, adolescence and early adulthood (ages 4–23 years old) in a task-free manner. In general, our findings revealed that across these ages, significant linear and nonlinear changes were observed in resting state connectivity between the ventral striatum and the primary circuitry implicated in incentive-based learning (mPFC, hippocampus, and insula). Childhood is characterized by significant ventral striatal connections with mPFC and amygdala. Taking the stance that the ventral striatum is a hub for incentive value-related signals (Haber and Knutson, 2010), one interpretation of our data is that these regions all work together at early ages to aid in understanding environmental contingencies, both rewarding and aversive. With time, the types of value-related information communicated between the ventral striatum and these regions may shift, with those remaining consistently positively coupled (e.g., vmPFC, OFC, amygdala) more involved in stimulus, decision and outcome evaluation (Bartra et al., 2013; Rangel et al., 2008; Salzman et al., 2007), whereas those cortical regions demonstrating age-related shifts in connectivity strength and patterns become less involved in incentive-based learning, supporting more general affective processes (e.g., sgACC) (Christakou et al., 2013; Critchley et al., 2004). Further, quadratic (i.e., U-shaped) changes in connectivity characterized by negative connectivity patterns during adolescence between the ventral striatum and posterior hippocampus, as well as the posterior insula, potentially suggest opposing relationships between the ventral striatum and these regions during adolescence. Taken together our findings provide a novel context within which to consider how ventral striatal communication with cortical and subcortical regions across development can differentially impact incentive-based valuation and learning.
4.1.1 Linear Age-Related Changes in Ventral Striatal Connectivity
We observed diverse patterns of age-related changes in ventral striatal functional connectivity with a number of our targeted regions of interest. The sgACC exhibited robust positive connectivity in our youngest participants (i.e., prior to adolescence), suggesting early communication between these regions. Significant age-related decreases in connectivity were observed in ventral striatal connectivity with the subgenual anterior cingulate cortex, although connectivity remained significantly positive in adolescent and adult participants; correlation estimates support the notion that the strength of connectivity between these two regions declines with age. The sgACC is known to be highly involved in affective processes in adults, including mood regulation (Drevets et al., 1997; Price and Drevets, 2012), craving (Kober et al., 2010), fear learning (Dunsmoor et al., 2011) and extinction (Phelps et al., 2004). The sgACC supports complex, interactive value computations during decision-making (e.g., incorporating advantages and disadvantages of a potential option (Park et al., 2011)). During development, representation of incentive-based learning signals in sgACC in concert with ventral striatum and ventrolateral PFC at younger ages tends to relate to poorer performance, whereas by adulthood, representation of such signals in vmPFC relates to better performance (Christakou et al., 2013). Thus one interpretation of our findings is the decline in strength of positive coupling between these two regions across development reflects functional specialization of ventral striatum and sgACC over time to support differential affective valuation-based processes over time.
This age-related change in resting-state connectivity was most prominent during the transition between childhood and adolescence. To answer the question of whether these changes were related to changes in puberty, we observed that levels of the pubertal hormone testosterone, which, consistent with recent work (Koolschijn et al., 2014), increased with age for both males and females (albeit with different production ranges), were negatively associated with the strength of ventral striatal-sgACC connectivity and significantly mediated the negative relationship between age and connectivity. Androgen receptors are abundant throughout the mesolimbic dopaminergic system (Kritzer, 1997) and consistent with androgen and tyrosine hydroxylase co-localization findings, several human imaging studies have found interactions between the two systems. Testosterone has been associated with incentive-based processes and ventral striatal function during adolescence such that increasing levels of testosterone in males and females positively correlates with the ventral striatal response to reward (Op de Macks et al., 2011), and explained more of the variance in the ventral striatal reward response than did age. Testosterone has additionally been associated with volumetric changes in prefrontal structures (e.g., ACC) such that higher levels of testosterone predict gray matter maturation (volumetric decreases) in ACC, particularly in males (Koolschijn et al., 2014). Interpreted within this context, our findings suggest that testosterone might be an important developmental agent driving the functional specialization of the ventral striatum and sgACC to support different components of incentive-based valuation. In other words, given the relevance of testosterone to incentive-based behavior, neural function, and the integrity of striatal-prefrontal circuits in adulthood as well as during development, it is possible that early in life the sgACC co-activates with the ventral striatum to support incentive valuation, but that increasing levels of testosterone with age enhance the motivational relevance of the ventral striatum, reducing the functional relationship with sgACC, enabling the sgACC to convey more general affective and arousal based signals (reviewed in Critchley et al., 2004).
4.1.2 Quadratic Changes in Ventral Striatal Connectivity
We observed significant quadratic age-related changes in connectivity between the ventral striatum and posterior hippocampus, characterized by negative connectivity during adolescence. Rodent analogues of the posterior hippocampus (i.e., dorsal hippocampus) have been implicated in supporting cognitive functions such as spatial learning and memory (Fanselow and Dong, 2010; McDonald and Hong, 2013). Additional recent rodent evidence reports signals associated with goal, decision and outcome values (Lee et al., 2012) and increased neural activity (i.e., theta oscillations) in the dorsal hippocampus of rodents during a reward decision-making task (Schmidt et al., 2013), suggesting that this region may play an important role in incentive valuation and decision-making. Corroborating human fMRI evidence supports a role for the posterior hippocampus in value-based decision-making as well, suggesting that it may inform or bias decisions based on the value of past experiences (Wimmer et al., 2012). Interpreted within this framework, the observed quadratic relationship ventral striatal rsFC with the posterior hippocampus may suggest that during adolescence these regions may exhibit an opposing relationship with each other such that the posterior hippocampus may either play less of a role in value-based decision-making or may communicate differential or competing value-based information during decision-making, making for less coherent communication between these regions and contributing to observed behavioral tendencies during (e.g., risk-taking) during this time period.
Quadratic age-related changes in connectivity also emerged with the mid/posterior insula, characterized by negative connectivity in adolescence as compared to no significant connectivity in childhood or adulthood. The mid-posterior subdivision of insular cortex has been linked to pain and somatosensory processes (Chang et al., 2013; Kelly et al., 2012), and integration of interoceptive signals to inform learning and decision-making (Chang et al., 2013; Kelly et al., 2012; Paulsen et al., 2011; Preuschoff et al., 2008) Decreased coupling with ventral striatum during adolescence could reflect a reorganization and/or lack of integrative communication between these regions during adolescence. Increases in risk-seeking during adolescence (Barkley-Levenson and Galvan, 2014; Chein et al., 2011; Somerville et al., 2010; Steinberg, 2008; Tymula et al., 2012), which are often associated with increases in ventral striatal response, may be a function of reduced ability to appraise interoceptive signals during this period of development. Other components of the insula—ventral/dorsal anterior insula––are often implicated in risk prediction and valuation in adults (Bartra et al., 2013; Preuschoff et al., 2008) and across development (Paulsen et al., 2011). Although we did not observe significant age-related changes in connectivity between the ventral striatum and these other subdivisions, future work should pursue investigations of the development specifically of ventral striatal-anterior insula connectivity, particularly in light of recent developmental models of anterior insula function in relation to both striatal and lateral prefrontal function (Smith et al., 2014), and in light of anatomical connectivity between the ventral striatum and the insula (Reynolds and Zahm, 2005).
4.1.3 Ventral Striatal-Amygdala/Hippocampal Connectivity
We had an a priori interest in characterizing the development of functional relationships between the ventral striatum with both the amygdala and hippocampus, based on the extensive literature across species implicating these regions in value-based learning and decision-making. We observed divergent patterns of connectivity between ventral striatum and these limbic structures. Significant positive coupling was present between the ventral striatum and amygdala in the youngest participants in our sample, and remained positive across adolescence and early adulthood, consistent with previous work (Gabard-Durnam et al., 2014). Amygdala function develops early in life so as to facilitate environmental adaptation and learning (Tottenham and Sheridan, 2009). The amygdala is crucial for rapid evaluation of both rewarding and aversive stimuli (Belova et al., 2008; 2007; Campeau and Davis, 1995; Davis, 1992; LaBar et al., 1998; LeDoux, 2000; Paton et al., 2006; Peck et al., 2013; Phelps and LeDoux, 2005), and this functionality develops early in life (Gee et al., 2013; see also Gilmore et al., 2012; Tottenham et al., 2012). In adulthood, the amygdala and ventral striatum support differential but complementary neural computations during learning (i.e., amygdala codes associability of a stimulus and outcome, ventral striatum codes for prediction errors) (Li et al., 2011). Thus, our resting-state data suggest that from an early age, the amygdala and ventral striatum operate in close concert to facilitate learning about rewarding and aversive incentives and patterns of resting state do not seem to change significantly from childhood to adulthood.
A markedly different pattern emerged regarding the development of ventral striatal-hippocampal connectivity. In addition to the quadratic age-related relationship between the ventral striatum and posterior hippocampus, linear patterns of age-related changes in ventral striatal connectivity with the hippocampus also emerged; although they did not remain significant after correction for multiple comparisons, given our a priori interest in the hippocampus, and the contrast with the amygdala, we believe it useful to briefly contextualize these patterns as potentially informative for future studies. Consistent with the notion that the hippocampus develops at later ages compared to the amygdala (Tottenham and Sheridan, 2009) in human (Gogtay et al., 2006) and non-human animals (Payne et al., 2010), linear age-related increases in ventral striatal-hippocampal connectivity (whole region and anterior subregion) at rest appeared to emerge in adulthood, but was not significantly present in childhood or adolescence. Incentive-based learning and decision-making indeed recruits the hippocampus in adults (Okatan, 2009), which may contribute by relaying more contextual and relational components of value-based information (Delgado and Dickerson, 2012; Pennartz et al., 2011). Rodent analogues of the human anterior hippocampus (ventral hippocampus) is implicated in supporting affective function. Human studies report significant correlations between BOLD activation in the anterior hippocampus during a contextual fear learning task and state anxiety (Satpute et al., 2012), as well as in coding the threat level present in the environment during decision-making (Bach et al., 2014), consistent with rodent literature implicating this region in affective function. The anterior hippocampus has also been implicated construction of value representations of novel stimuli based on memories of related components (Barron et al., 2013). Speculatively, our findings suggest that the linear development of positive connectivity between ventral striatum and hippocampus, and more specifically the anterior hippocampus, at later ages may reflect a later emergence of these regions working together to support more complex affective valuation processes. The development of ventral striatal-hippocampal connectivity remains an important question for future studies.
4.1.4 Relation to prior studies on striatal functional development
Our findings build upon recent investigations of striatal and basal ganglia rsFC development. Di Martino and colleagues (2011) report that typically developing children demonstrate more robust connectivity than typically developed adults between the ventral striatum and other striatal regions, middle cingulate cortex, precentral gyrus and insular cortex, but show less connectivity with medial prefrontal structures (vmPFC, OFC). The divergence between these latter results of Di Martino and colleagues and those presented here may be due to differences in analytic techniques (e.g., motion processing), or to the differences in the seed ROI specifications. Greene and colleagues (Greene et al., 2014) investigated basal ganglia connectivity in a similarly constructed sample (children: 7–12 years old, adults 19–31 years old) using partial correlation approaches and a winner take all analysis (see also Fair et al., 2010). Of note, connectivity between ventral striatum (comprised of a nucleus accumbens ROI within a larger basal ganglia ROI) and an orbitofrontal network emerged in both children and adults, consistent with the present study, and age-related decreases emerged in connectivity between the putamen/pallidum and somatomotor face network across childhood. We note that the consistencies with Greene and colleagues with respect to ventral striatal-orbitofrontal connectivity emerge in spite of differences our inclusion of global signal regression at the single subject level, while Greene and colleagues employed an alternative technique. Though some of our results may be influenced by the inclusion of global signal (see Appendix A), we note a large degree of consistency regardless of whether the global signal is included or not.
We also note a recent study examining developmental changes in ventral and dorsal striatal connectivity at rest in a developmental sample that extended into middle age (i.e., 9–44) (Porter et al., 2014). One finding of note concerns ventral striatal-insula connectivity. Porter and colleagues report age-related linear decreases in connectivity between the ventral striatum and anterior insula; the authors suggest that stronger coupling during adolescence than in adulthood could reflect a greater dependence on motivated behavior during this time period. While we did not observe significant age-related changes in ventral striatal-anterior insula connectivity, we did note significant quadratic age-related changes in connectivity between the ventral striatum and posterior insula. Differences in the construction of samples of participants, however, may underlie the differential patterns of results: the majority of participants were older than 19 years of age, and adolescents were considered in certain analyses to be all individuals under the age of 19 in the study by Porter and colleagues. Based on this, it is difficult to directly compare results, but we believe our observation of quadratic age-related changes in ventral striatal-insula connectivity may partially underlie developmental differences in incentive-based behaviors. These ideas should be more directly tested in future work.
4.1.5 Limitations
We believe the findings of the present study are an important contribution to the extant literature regarding neural systems supporting incentive-based valuation and their development. We do acknowledge some limitations of our work, however. First, while our overall sample size is similar to that of previous work (Cohen et al., 2010; Gabard-Durnam et al., 2014; van den Bos et al., 2013; van Duijvenvoorde et al., 2008), we do note that the adult subsample is somewhat small. We believe that a strength of our study is the large number of children and adolescents enrolled, because of the potential important behavioral and neural changes occurring during the early years of life (Casey et al., 2010; Somerville et al., 2010; Steinberg, 2008). Furthermore, though we present post-hoc t-tests for age groups to characterize connectivity that is significantly different from zero, these analyses were conducted primarily for interpretation purposes. Based on the fact that visual inspection of our data suggests that data from our adult participants were largely consistent, we believe that a larger adult subsample would have strengthened our findings.
Second, we acknowledge the ongoing debate in the literature regarding global signal regression in resting-state fMRI analyses. We chose to include this step in our analyses to account for physiological differences/artifacts that otherwise would not have been possible, as additional measures (e.g., heart rate, respiration) were not available. Not regressing out the global signal would have led our results to be potentially unduly confounded with physiological artifacts. Concern regarding the use of global signal regression in resting-state date stems from the idea that doing so can induce negative connectivity between regions, because this procedure necessitates that the sum of correlation coefficients across the brain with a seed region of interest must be zero. Thus, negative correlations must be present according to this view (Fox et al., 2009; Murphy et al., 2009). However, evidence also suggests that global signal regression not only improves the spatial specificity of functional connectivity maps via the removal of noise (Fox et al., 2009), but that negative or anti-correlations may be present regardless of whether the global signal is regressed out or not; this has been shown in both rodent (Liang et al., 2011) and human studies (Keller et al., 2013). Further, correspondence in negative correlations at rest both using fMRI and neuronal recordings in human patients (Keller et al., 2013), both with and without global signal regression, which suggests that negative connectivity may indeed have a neuronal basis. While our supplemental ROI analyses not employing global signal regression did reveal weaker relationships with the ventral striatum, the patterns were similar to those observed when including global signal regression, supporting the idea that this step may indeed help uncover existing effects. Negative connectivity suggests that regions or networks may be involved in opposing or competitive processes, as opposed to a zero correlation, which would suggest no relationship at all. The observation of negative connectivity between the ventral striatum and mid/posterior insula, as well as between the ventral striatum and the posterior hippocampus during adolescence, when interpreted within this light may suggest that these regions are performing opposing functions, underlying changes in incentive-based behavior during adolescence.
Third, we note the possibility some of our connectivity results in regions that were close to our ventral striatum ROI may have been due to signal bleed from our smoothing kernel. A recent study by Greene and colleagues (2014) noted a similar concern, choosing to regress out signal adjacent to their basal ganglia region of interest. It is noted in that study that doing so does leave open the possibility that true correlations between the region of interest are not possible to observe. Additionally, because of the focal location of the ventral striatum, and its connectivity to other regions of the striatum itself, including both the ventral striatal signal and that adjacent to it in regression analyses may lead to issues of co-linearity, making it potentially difficult to interpret connectivity estimates as well. While we chose not to take the approach of including adjacent signal as a regressor in our model, and note supplemental analyses in Appendix A in support of this, we do acknowledge this as a potential limitation of our study.
Fourth, while we note that our ROI results are generally supported by the whole-brain voxelwise results presented in Appendix A, there is the possibility that using anatomical ROIs particularly for significant portions of cortex may be problematic if the alignment of cytoarchitectonics with function is not specific enough. Future studies investigating the development of ventral striatal connectivity could employ functional parcellation approaches (Blumensath et al., 2013; Choi et al., 2012) to better address such issues.
Finally, while we took great lengths to communicate with our subjects throughout the scanning session, and in particular prior to the resting-state session to ensure alertness, it is certainly possible that some individuals may have experienced bouts of sleepiness during the resting-state sequence, given that it was at the end of the scanning session. Subject sleepiness in the fMRI environment is more often associated with older adults, college age students, and certain clinical populations (Duncan and Northoff, 2013; Raz et al., 2005). Nevertheless, it is possible that our youngest participants may have had difficulty remaining focused and awake by the end of the session.
4.1.6 Conclusions
Recent years have seen an increase in the use of resting-state fMRI techniques as a means toward understanding the organization of both neural networks and the connectivity between specific regions of interest. The present study examined the normative development of task-free ventral striatal connectivity across early childhood, adolescence and adulthood, which enables the characterization of continuous change in neurodevelopmental functional organization. Incentives are especially important for learning about environmental contingencies during the early periods of life––i.e., childhood and adolescence––when changes in incentive-based behavior and associated neural systems centered on the ventral striatum may be in a high state of flux. By focusing on this age range during which changes in ventral striatal functional communication may be most dynamic, our study may provide insight to the timing of normative developmental changes in behavior. Further, the rsFC findings presented here can help constrain interpretations of future developmental studies reporting task-based ventral striatal functional connectivity. Interpretations of accounts of no functional connectivity between the ventral striatum and, for example, mPFC during task in young children could be aided by the present results which would suggest that lack of task-based coupling may not necessarily mean the regions do not communicate early in life, but perhaps that development of the relationship for a particular process may not have yet emerged. Thus, the present findings provide a broader context within which to consider ventral striatal functional development and its behavioral implications.
Supplementary Material
Acknowledgments
This work was supported by the National Institute of Mental Health (R01MH091864 to N.T.) and the Dana Foundation.
Footnotes
The results presented in the main body of the manuscript include all 66 participants. We also present a number of supplemental analyses in Appendix A, one group of which is conducted with two participants removed––the participant who scored slightly above the clinical cutoff of CBCL scores, and the participant who contributed slightly less (107) than 3sd from the mean of usable TRs (110). The major results of interest do not change, but are presented in the Appendix for completeness.
References
- Achenbach TM. Manual for the Child Behavior Checklist/4-18 and 1991 Profile. University of Vermont, Department of Psychiatry; Burlington, VT: 1991. [Google Scholar]
- Alcauter S, Lin W, Keith Smith J, Gilmore JH, Gao W. Consistent Anterior-Posterior Segregation of the Insula During the First 2 Years of Life. Cereb Cortex. 2013 doi: 10.1093/cercor/bht312. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Alexander GE, Crutcher MD, DeLong MR. Basal ganglia-thalamocortical circuits: parallel substrates for motor, oculomotor, “prefrontal” and “limbic” functions. Prog Brain Res. 1990;85:119–146. [PubMed] [Google Scholar]
- Alexander WH, Brown JW. Medial prefrontal cortex as an action-outcome predictor. Nature Neuroscience. 2011;14:1338–1344. doi: 10.1038/nn.2921. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bach DR, Guitart-Masip M, Packard PA, Miró J, Falip M, Fuentemilla L, Dolan RJ. Human Hippocampus Arbitrates Approach-Avoidance Conflict. Curr Biol. 2014;24:541–547. doi: 10.1016/j.cub.2014.01.046. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barkley-Levenson E, Galvan A. Neural representation of expected value in the adolescent brain. Proc Natl Acad Sci USA. 2014;111:1646–1651. doi: 10.1073/pnas.1319762111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Barron HC, Dolan RJ, Behrens TEJ. Online evaluation of novel choices by simultaneous representation of multiple memories. Nature Neuroscience. 2013:1–9. doi: 10.1038/nn.3515. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bartra O, McGuire JT, Kable JW. The valuation system: a coordinate-based meta-analysis of BOLD fMRI experiments examining neural correlates of subjective value. NeuroImage. 2013;76:412–427. doi: 10.1016/j.neuroimage.2013.02.063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Beall EB. Adaptive cyclic physiologic noise modeling and correction in functional MRI. J Neurosci Methods. 2010;187:216–228. doi: 10.1016/j.jneumeth.2010.01.013. [DOI] [PubMed] [Google Scholar]
- Beall EB, Lowe MJ. Isolating physiologic noise sources with independently determined spatial measures. NeuroImage. 2007;37:1286–1300. doi: 10.1016/j.neuroimage.2007.07.004. [DOI] [PubMed] [Google Scholar]
- Behrens TEJ, Woolrich MW, Walton ME, Rushworth MFS. Learning the value of information in an uncertain world. Nature Neuroscience. 2007;10:1214–1221. doi: 10.1038/nn1954. [DOI] [PubMed] [Google Scholar]
- Belova M, Paton J, Morrison S, Salzman C. Expectation modulates neural responses to pleasant and aversive stimuli in primate amygdala. Neuron. 2007;55:970–984. doi: 10.1016/j.neuron.2007.08.004. doi:S0896-6273(07)00615-0 [pii] 10.1016/j.neuron.2007.08.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Belova M, Paton J, Salzman C. Moment-to-moment tracking of state value in the amygdala. J Neurosci. 2008;28:10023–10030. doi: 10.1523/JNEUROSCI.1400-08.2008. doi:28/40/10023 [pii] 10.1523/JNEUROSCI.1400-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bjork JM, Knutson B, Fong GW, Caggiano DM, Bennett SM, Hommer DW. Incentive-elicited brain activation in adolescents: similarities and differences from young adults. Journal of Neuroscience. 2004;24:1793–1802. doi: 10.1523/JNEUROSCI.4862-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bjork JM, Smith AR, Chen G, Hommer DW. Adolescents, adults and rewards: comparing motivational neurocircuitry recruitment using fMRI. PLoS ONE. 2010;5:e11440. doi: 10.1371/journal.pone.0011440. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Blumensath T, Jbabdi S, Glasser MF, Van Essen DC, Uğurbil K, Behrens TEJ, Smith SM. Spatially constrained hierarchical parcellation of the brain with resting-state fMRI. NeuroImage. 2013;76:313–324. doi: 10.1016/j.neuroimage.2013.03.024. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Buckner RL, Vincent JL. Unrest at rest: default activity and spontaneous network correlations. NeuroImage. 2007;37:1091–6. doi: 10.1016/j.neuroimage.2007.01.010. discussion 1097–9. [DOI] [PubMed] [Google Scholar]
- Burgund ED, Kang HC, Kelly JE, Buckner RL, Snyder AZ, Petersen SE, Schlaggar BL. The feasibility of a common stereotactic space for children and adults in fMRI studies of development. NeuroImage. 2002;17:184–200. doi: 10.1006/nimg.2002.1174. [DOI] [PubMed] [Google Scholar]
- Bush G, Luu P, Posner M. Cognitive and emotional influences in anterior cingulate cortex. Trends Cogn Sci (Regul Ed) 2000;4:215–222. doi: 10.1016/s1364-6613(00)01483-2. [DOI] [PubMed] [Google Scholar]
- Campeau S, Davis M. Involvement of the central nucleus and basolateral complex of the amygdala in fear conditioning measured with fear-potentiated startle in rats trained concurrently with auditory and visual conditioned stimuli. J Neurosci. 1995;15:2301–2311. doi: 10.1523/JNEUROSCI.15-03-02301.1995. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Casey BJ, Duhoux S, Malter Cohen M. Adolescence: what do transmission, transition, and translation have to do with it? Neuron. 2010;67:749–760. doi: 10.1016/j.neuron.2010.08.033. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chang LJ, Yarkoni T, Khaw MW, Sanfey AG. Decoding the Role of the Insula in Human Cognition: Functional Parcellation and Large-Scale Reverse Inference. Cereb Cortex. 2013;23:739–749. doi: 10.1093/cercor/bhs065. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chein J, Albert D, O’Brien L, Uckert K, Steinberg L. Peers increase adolescent risk taking by enhancing activity in the brain’s reward circuitry. Dev Sci. 2011;14:F1–10. doi: 10.1111/j.1467-7687.2010.01035.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chen G, Chen G, Xie C, Ward BD, Li W, Antuono P, Li SJ. A method to determine the necessity for global signal regression in resting-state fMRI studies. Magn Reson Med. 2012;68:1828–1835. doi: 10.1002/mrm.24201. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cho YT, Fromm S, Guyer AE, Detloff A, Pine DS, Fudge JL, Ernst M. Nucleus accumbens, thalamus and insula connectivity during incentive anticipation in typical adults and adolescents. NeuroImage. 2013;66:508–521. doi: 10.1016/j.neuroimage.2012.10.013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Choi EY, Yeo BTT, Buckner RL. The organization of the human striatum estimated by intrinsic functional connectivity. J Neurophysiol. 2012;108:2242–2263. doi: 10.1152/jn.00270.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Chorpita BF, Yim L, Moffitt C, Umemote LA, Francis SE. Assessment of symptoms of DSM-IV anxiety and depression in children: a revised child anxiety and depression scale. Behaviour Research and Therapy. 2000;38:835–855. doi: 10.1016/s0005-7967(99)00130-8. [DOI] [PubMed] [Google Scholar]
- Christakou A, Gershman SJ, Niv Y, Simmons A, Brammer M, Rubia K. Neural and Psychological Maturation of Decision-making in Adolescence and Young Adulthood. Journal of Cognitive Neuroscience. 2013;25:1807–1823. doi: 10.1037/0882-7974.20.2.220. [DOI] [PubMed] [Google Scholar]
- Cohen JR, Asarnow RF, Sabb FW, Bilder RM, Bookheimer SY, Knowlton BJ, Poldrack RA. A unique adolescent response to reward prediction errors. Nature Neuroscience. 2010;13:669–671. doi: 10.1038/nn.2558. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cohen MX, Schoene-Bake JC, Elger CE, Weber B. Connectivity-based segregation of the human striatum predicts personality characteristics. Nature Neuroscience. 2009;12:32–34. doi: 10.1038/nn.2228. [DOI] [PubMed] [Google Scholar]
- Cohen P, Cohen J, Aiken LS, West SG. The Problem of Units and the Circumstance for POMP. Multivariate Behavioral Research. 1999;34:315–346. doi: 10.1207/S15327906MBR3403_2. [DOI] [Google Scholar]
- Cole D, Smith SM, Beckmann CF. Advances and Pitfalls in the Analysis and Interpretation of Resting-State FMRI Data. Frontiers in Systems Neuroscience. 2010;4 doi: 10.3389/fnsys.2010.00008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cox RW. AFNI: Software for analysis and visualization of functional magentic resonance neuroimages. Computers and Biomedical Research. 1996;29:162–173. doi: 10.1006/cbmr.1996.0014. [DOI] [PubMed] [Google Scholar]
- Critchley HD, Wiens S, Rotshtein P, Ohman A, Dolan RJ. Neural systems supporting interoceptive awareness. Nature Neuroscience. 2004;7:189–195. doi: 10.1038/nn1176. [DOI] [PubMed] [Google Scholar]
- Davis M. The Role of the Amygdala in Fear and Anxiety. Annual Review of Neuroscience. 1992;15:353–375. doi: 10.1146/annurev.ne.15.030192.002033. [DOI] [PubMed] [Google Scholar]
- Deen B, Pitskel NB, Pelphrey KA. Three systems of insular functional connectivity identified with cluster analysis. Cereb Cortex. 2011;21:1498–1506. doi: 10.1093/cercor/bhq186. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delgado MR. Reward-related responses in the human striatum. Annals of the New York Academy of Sciences. 2007;1104:70–88. doi: 10.1196/annals.1390.002. [DOI] [PubMed] [Google Scholar]
- Delgado MR, Dickerson KC. Reward-related learning via multiple memory systems. Biological Psychiatry. 2012;72:134–141. doi: 10.1016/j.biopsych.2012.01.023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Delgado MR, Nystrom LE, Fissell C, Noll D, Fiez JA. Tracking the hemodynamic responses to reward and punishment in the striatum. J Neurophysiol. 2000;84:3072–3077. doi: 10.1152/jn.2000.84.6.3072. [DOI] [PubMed] [Google Scholar]
- Delmonte S, Gallagher L, O’Hanlon E, McGrath J, Balsters JH. Functional and structural connectivity of frontostriatal circuitry in Autism Spectrum Disorder. Front Hum Neurosci. 2013;7:430. doi: 10.3389/fnhum.2013.00430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Martino A, Kelly C, Grzadzinski R, Zuo XN, Mennes M, Mairena MA, Lord C, Castellanos FX, Milham MP. Aberrant striatal functional connectivity in children with autism. Biological Psychiatry. 2011;69:847–856. doi: 10.1016/j.biopsych.2010.10.029. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Di Martino A, Scheres A, Margulies DS, Kelly AMC, Uddin LQ, Shehzad Z, Biswal B, Walters JR, Castellanos FX, Milham MP. Functional Connectivity of Human Striatum: A Resting State fMRI Study. Cereb Cortex. 2008;18:2735–2747. doi: 10.1093/cercor/bhn041. [DOI] [PubMed] [Google Scholar]
- Drevets WC, Price JL, Simpson JR, Todd RD, Reich T, Vannier M, Raichle ME. Subgenual prefrontal cortex abnormalities in mood disorders. Nature. 1997;386:824–827. doi: 10.1038/386824a0. [DOI] [PubMed] [Google Scholar]
- Duncan N, Northoff G. Overview of potential procedural and participant-related confounds for neuroimaging of the resting state. J Psychiatry Neurosci. 2013;38:84–96. doi: 10.1503/jpn.120059. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Dunsmoor JE, Prince SE, Murty VP, Kragel PA, LaBar KS. Neurobehavioral mechanisms of human fear generalization. NeuroImage. 2011;55:1878–1888. doi: 10.1016/j.neuroimage.2011.01.041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Efron B. Better bootstrap confidence intervals. Journal of the American Statistical Association. 1987;82:171–185. [Google Scholar]
- Ernst M, Nelson EE, Jazbec S, McClure EB, Monk CS, Leibenluft E, Blair J, Pine DS. Amygdala and nucleus accumbens in responses to receipt and omission of gains in adults and adolescents. NeuroImage. 2005;25:1279–1291. doi: 10.1016/j.neuroimage.2004.12.038. [DOI] [PubMed] [Google Scholar]
- Eshel N, Nelson EE, Blair RJ, Pine DS, Ernst M. Neural substrates of choice selection in adults and adolescents: Development of the ventrolateral prefrontal and anterior cingulate cortices. Neuropsychologia. 2007;45:1270–1279. doi: 10.1016/j.neuropsychologia.2006.10.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fair DA, Bathula D, Mills KL, Dias TGC, Blythe MS, Zhang D, Snyder AZ, Raichle ME, Stevens AA, Nigg JT, Nagel BJ. Maturing thalamocortical functional connectivity across development. Frontiers in Systems Neuroscience. 2010;4:10. doi: 10.3389/fnsys.2010.00010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fair DA, Cohen AL, Dosenbach NUF, Church JA, Miezin FM, Barch DM, Raichle ME, Petersen SE, Schlaggar BL. The maturing architecture of the brain’s default network. Proc Natl Acad Sci USA. 2008;105:4028–4032. doi: 10.1073/pnas.0800376105. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fanselow MS, Dong HW. Are the Dorsal and Ventral Hippocampus Functionally Distinct Structures? Neuron. 2010;65:7–19. doi: 10.1016/j.neuron.2009.11.031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Fareri DS, Martin LN, Delgado MR. Reward-related processing in the human brain: developmental considerations. Dev Psychopathol. 2008;20:1191–1211. doi: 10.1017/S0954579408000576. [DOI] [PubMed] [Google Scholar]
- Fox MD, Zhang D, Snyder AZ, Raichle ME. The global signal and observed anticorrelated resting state brain networks. J Neurophysiol. 2009;101:3270–3283. doi: 10.1152/jn.90777.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gabard-Durnam LJ, Flannery J, Goff B, Gee DG, Humphreys KL, Telzer E, Hare T, Tottenham N. The development of human amygdala functional connectivity at rest from 4 to 23 years: A cross-sectional study. NeuroImage. 2014;95:193–207. doi: 10.1016/j.neuroimage.2014.03.038. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galván A. Adolescent development of the reward system. Front Hum Neurosci. 2010;4:6. doi: 10.3389/neuro.09.006.2010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Galván A, Hare TA, Parra CE, Penn J, Voss H, Glover G, Casey BJ. Earlier development of the accumbens relative to orbitofrontal cortex might underlie risk-taking behavior in adolescents. J Neurosci. 2006;26:6885–6892. doi: 10.1523/JNEUROSCI.1062-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gee DG, Humphreys KL, Flannery J, Goff B, Telzer EH, Shapiro M, Hare TA, Bookheimer SY, Tottenham N. A developmental shift from positive to negative connectivity in human amygdala-prefrontal circuitry. Journal of Neuroscience. 2013;33:4584–4593. doi: 10.1523/JNEUROSCI.3446-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Giedd JN. Structural magnetic resonance imaging of the adolescent brain. Annals of the New York Academy of Sciences. 2004;1021:77–85. doi: 10.1196/annals.1308.009. [DOI] [PubMed] [Google Scholar]
- Giedd JN, Blumenthal J, Jeffries NO, Castellanos FX, Liu H, Zijdenbos A, Paus T, Evans AC, Rapoport JL. Brain development during childhood and adolescence: a longitudinal MRI study. Nature Neuroscience. 1999;2:861–863. doi: 10.1038/13158. [DOI] [PubMed] [Google Scholar]
- Gilmore JH, Shi F, Woolson SL, Knickmeyer RC, Short SJ, Lin W, Zhu H, Hamer RM, Styner M, Shen D. Longitudinal development of cortical and subcortical gray matter from birth to 2 years. Cereb Cortex. 2012;22:2478–2485. doi: 10.1093/cercor/bhr327. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Gogtay N, Nugent TF, Herman DH, Ordonez A, Greenstein D, Hayashi KM, Clasen L, Toga AW, Giedd JN, Rapoport JL, Thompson PM. Dynamic mapping of normal human hippocampal development. Hippocampus. 2006;16:664–672. doi: 10.1002/hipo.20193. [DOI] [PubMed] [Google Scholar]
- Greene DJ, Laumann TO, Dubis JW, Ihnen SK, Neta M, Power JD, Pruett JR, Black KJ, Schlaggar BL. Developmental Changes in the Organization of Functional Connections between the Basal Ganglia and Cerebral Cortex. Journal of Neuroscience. 2014;34:5842–5854. doi: 10.1523/JNEUROSCI.3069-13.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Groenewegen HJ, Wright CI, Beijer AVJ, Voorn P. Convergence and Segregation of Ventral Striatal Inputs and Outputs. Annals of the New York Academy of Sciences. 1999;877:49–63. doi: 10.1111/j.1749-6632.1999.tb09260.x. [DOI] [PubMed] [Google Scholar]
- Guyer AE, Monk CS, McClure-Tone EB, Nelson EE, Roberson-Nay R, Adler AD, Fromm SJ, Leibenluft E, Pine DS, Ernst M. A developmental examination of amygdala response to facial expressions. Journal of Cognitive Neuroscience. 2008;20:1565–1582. doi: 10.1162/jocn.2008.20114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haber SN, Kim KS, Mailly P, Calzavara R. Reward-related cortical inputs define a large striatal region in primates that interface with associative cortical connections, providing a substrate for incentive-based learning. Journal of Neuroscience. 2006;26:8368–8376. doi: 10.1523/JNEUROSCI.0271-06.2006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haber SN, Knutson B. The reward circuit: linking primate anatomy and human imaging. Neuropsychopharmacology. 2010;35:4–26. doi: 10.1038/npp.2009.129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Haber SN, McFarland NR. The concept of the ventral striatum in nonhuman primates. Annals of the New York Academy of Sciences. 1999;877:33–48. doi: 10.1111/j.1749-6632.1999.tb09259.x. [DOI] [PubMed] [Google Scholar]
- Hallquist MN, Hwang K, Luna B. The nuisance of nuisance regression: Spectral misspecification in a common approach to resting-state fMRI preprocessing reintroduces noise and obscures functional connectivity. NeuroImage. 2013;82C:208–225. doi: 10.1016/j.neuroimage.2013.05.116. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hampson M, Driesen N, Roth JK, Gore JC, Constable RT. Functional connectivity between task-positive and task-negative brain areas and its relation to working memory performance. Magnetic Resonance Imaging. 2010;28:1051–1057. doi: 10.1016/j.mri.2010.03.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hare TA, O’Doherty J, Camerer CF, Schultz W, Rangel A. Dissociating the role of the orbitofrontal cortex and the striatum in the computation of goal values and prediction errors. Journal of Neuroscience. 2008a;28:5623–5630. doi: 10.1523/JNEUROSCI.1309-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hare TA, Tottenham N, Galván A, Voss HU, Glover GH, Casey BJ. Biological substrates of emotional reactivity and regulation in adolescence during an emotional go-nogo task. Biological Psychiatry. 2008b;63:927–934. doi: 10.1016/j.biopsych.2008.03.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Hayes AF. PROCESS: A versatile computational tool for observed variable mediation, moderation and conditional process modeling [White paper] [WWW Document] 2012 www.afhayes.com. URL http://www.afhayes.com/public/process2012pdf (accessed 7.28.14)
- Hayes AF. Introductionto Mediation, Moderation and Conditional Process Analysis. Guilford Press; New York: 2013. [Google Scholar]
- Hayes AF, Scharkow M. The Relative Trustworthiness of Inferential Tests of the Indirect Effect in Statistical Mediation Analysis: Does Method Really Matter? Psychol Sci. 2013;24:1918–1927. doi: 10.1177/0956797613480187. [DOI] [PubMed] [Google Scholar]
- Hermans EJ, Bos PA, Ossewaarde L, Ramsey NF, Fernandez G, van Honk J. Effects of exogenous testosterone on the ventral striatal BOLD response during reward anticipation in healthy women. NeuroImage. 2010;52:277–283. doi: 10.1016/j.neuroimage.2010.04.019. [DOI] [PubMed] [Google Scholar]
- Holm S. A simple sequentially rejective multiple test procedure. Scandinavian Journal of Statistics. 1979;6:65–70. [Google Scholar]
- Jones RM, Somerville LH, Li J, Ruberry EJ, Powers A, Mehta N, Dyke J, Casey BJ. Adolescent-specific patterns of behavior and neural activity during social reinforcement learning. Cognitive, Affective, & Behavioral Neuroscience. 2014 doi: 10.3758/s13415-014-0257-z. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kahn I, Andrews-Hanna JR, Vincent JL, Snyder AZ, Buckner RL. Distinct Cortical Anatomy Linked to Subregions of the Medial Temporal Lobe Revealed by Intrinsic Functional Connectivity. J Neurophysiol. 2008;100:129–139. doi: 10.1152/jn.00077.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kang HC, Burgund ED, Lugar HM, Petersen SE, Schlaggar BL. Comparison of functional activation foci in children and adults using a common stereotactic space. NeuroImage. 2003;19:16–28. doi: 10.1016/s1053-8119(03)00038-7. [DOI] [PubMed] [Google Scholar]
- Keller CJ, Bickel S, Honey CJ, Groppe DM, Entz L, Craddock RC, Lado FA, Kelly C, Milham M, Mehta AD. Neurophysiological Investigation of Spontaneous Correlated and Anticorrelated Fluctuations of the BOLD Signal. Journal of Neuroscience. 2013;33:6333–6342. doi: 10.1523/JNEUROSCI.4837-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kelly C, Toro R, Di Martino A, Cox CL, Bellec P, Castellanos FX, Milham MP. A convergent functional architecture of the insula emerges across imaging modalities. NeuroImage. 2012;61:1129–1142. doi: 10.1016/j.neuroimage.2012.03.021. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kennerley SW, Walton ME, Behrens TEJ, Buckley MJ, Rushworth MFS. Optimal decision making and the anterior cingulate cortex. Nature Neuroscience. 2006;9:940–947. doi: 10.1038/nn1724. [DOI] [PubMed] [Google Scholar]
- Knutson B, Adams C, Fong G, Hommer D. Anticipation of increasing monetary reward selectively recruits nucleus accumbens. J Neurosci. 2001;21:RC159. doi: 10.1523/JNEUROSCI.21-16-j0002.2001. doi:20015472 [pii] [DOI] [PMC free article] [PubMed] [Google Scholar]
- Knutson B, Taylor J, Kaufman M, Peterson R, Glover G. Distributed neural representation of expected value. J Neurosci. 2005;25:4806–4812. doi: 10.1523/JNEUROSCI.0642-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kober H, Mende-Siedlecki P, Kross EF, Weber J, Mischel W, Hart CL, Ochsner KN. Prefrontal-striatal pathway underlies cognitive regulation of craving. Proc Natl Acad Sci USA. 2010;107:14811–14816. doi: 10.1073/pnas.1007779107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Koolschijn PCMP, Peper JS, Crone EA. The Influence of Sex Steroids on Structural Brain Maturation in Adolescence. PLoS ONE. 2014;9:e83929. doi: 10.1371/journal.pone.0083929.t002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kringelbach ML, O’Doherty J, Rolls ET, Andrews C. Activation of the human orbitofrontal cortex to a liquid food stimulus is correlated with its subjective pleasantness. Cerebral Cortex. 2003;13:1064–1071. doi: 10.1093/cercor/13.10.1064. [DOI] [PubMed] [Google Scholar]
- Kritzer MF. Selective colocalization of immunoreactivity for intracellular gonadal hormone receptors and tyrosine hydroxylase in the ventral tegmental area, substantia nigra, and retrorubral fields in the rat. Journal of Comparative Neurology. 1997;379:247–260. doi: 10.1002/(sici)1096-9861(19970310)379:2<247::aid-cne6>3.0.co;2-3. [DOI] [PubMed] [Google Scholar]
- Kuhn C, Johnson M, Thomae A, Luo B, Simon SA, Zhou G, Walker QD. Hormones and Behavior. Hormones and Behavior. 2010;58:122–137. doi: 10.1016/j.yhbeh.2009.10.015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Kuhnen CM, Knutson B. The neural basis of financial risk taking. Neuron. 2005;47:763–770. doi: 10.1016/j.neuron.2005.08.008. [DOI] [PubMed] [Google Scholar]
- LaBar KS, Gatenby JC, Gore JC, Ledoux JE, Phelps EA. Human amygdala activation during conditioned fear acquisition and extinction: a mixed-trial fMRI study. Neuron. 1998;20:937–945. doi: 10.1016/s0896-6273(00)80475-4. [DOI] [PubMed] [Google Scholar]
- LeDoux JE. Emotion circuits in the brain. Annual Review of Neuroscience. 2000;23:155–184. doi: 10.1146/annurev.neuro.23.1.155. [DOI] [PubMed] [Google Scholar]
- Lee H, Ghim JW, Kim H, Lee D, Jung M. Hippocampal neural correlates for values of experienced events. Journal of Neuroscience. 2012;32:15053–15065. doi: 10.1523/JNEUROSCI.2806-12.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Leotti LA, Delgado MR. Processing social and nonsocial rewards in the human brain. In: Decety J, Cacioppo J, editors. The Oxford Handbook of Social Neuroscience. Oxford University Press; New York: 2011a. pp. 178–194. [Google Scholar]
- Leotti LA, Delgado MR. The inherent reward of choice. Psychol Sci. 2011b;22:1310–1318. doi: 10.1177/0956797611417005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li J, Daw ND. Signals in human striatum are appropriate for policy update rather than value prediction. Journal of Neuroscience. 2011;31:5504–5511. doi: 10.1523/JNEUROSCI.6316-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li J, Schiller D, Schoenbaum G, Phelps EA, Daw ND. Differential roles of human striatum and amygdala in associative learning. Nature Neuroscience. 2011;14:1250–1252. doi: 10.1038/nn.2904. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Liang Z, King J, Zhang N. Anticorrelated resting-state functional connectivity in awake rat brain. NeuroImage. 2011:1–10. doi: 10.1016/j.neuroimage.2011.08.009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- McDonald RJ, Hong NS. How does a specific learning and memory system in the mammalian brain gain control of behavior? Hippocampus. 2013;23:1084–1102. doi: 10.1002/hipo.22177. [DOI] [PubMed] [Google Scholar]
- Miranda-Dominguez O, Mills BD, Grayson D, Woodall A, Grant KA, Kroenke CD, Fair DA. Bridging the Gap between the Human and Macaque Connectome: A Quantitative Comparison of Global Interspecies Structure-Function Relationships and Network Topology. Journal of Neuroscience. 2014;34:5552–5563. doi: 10.1523/JNEUROSCI.4229-13.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Moser MB, Moser EI. Functional differentiation in the hippocampus. Hippocampus. 1998;8:608–619. doi: 10.1002/(SICI)1098-1063(1998)8:6<608::AID-HIPO3>3.0.CO;2-7. [DOI] [PubMed] [Google Scholar]
- Murphy K, Birn RM, Handwerker DA, Jones TB, Bandettini PA. The impact of global signal regression on resting state correlations: are anti-correlated networks introduced? NeuroImage. 2009;44:893–905. doi: 10.1016/j.neuroimage.2008.09.036. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niv Y, Edlund JA, Dayan P, O’Doherty JP. Neural prediction errors reveal a risk-sensitive reinforcement-learning process in the human brain. Journal of Neuroscience. 2012;32:551–562. doi: 10.1523/JNEUROSCI.5498-10.2012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Niv Y, Schoenbaum G. Dialogues on prediction errors. Trends Cogn Sci. 2008;12:265–272. doi: 10.1016/j.tics.2008.03.006. doi:S1364-6613(08)00137-X [pii] 10.1016/j.tics.2008.03.006. [DOI] [PubMed] [Google Scholar]
- O’Doherty J, Kringelbach ML, Rolls ET, Hornak J, Andrews C. Abstract reward and punishment representations in the human orbitofrontal cortex. Nature Neuroscience. 2001;4:95–102. doi: 10.1038/82959. [DOI] [PubMed] [Google Scholar]
- O’Doherty JP. Reward representations and reward-related learning in the human brain: insights from neuroimaging. Current Opinion in Neurobiology. 2004;14:769–776. doi: 10.1016/j.conb.2004.10.016. [DOI] [PubMed] [Google Scholar]
- Okatan M. Correlates of reward-predictive value in learning-related hippocampal neural activity. Hippocampus. 2009;19:487–506. doi: 10.1002/hipo.20535. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Op de Macks ZA, Gunther Moor B, Overgaauw S, Güroglu B, Dahl RE, Crone EA. Testosterone levels correspond with increased ventral striatum activation in response to monetary rewards in adolescents. Developmental Brain Research. 2011;1:506–516. doi: 10.1016/j.dcn.2011.06.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Park SQ, Kahnt T, Rieskamp J, Heekeren HR. Neurobiology of Value Integration: When Value Impacts Valuation. Journal of Neuroscience. 2011;31:9307–9314. doi: 10.1523/JNEUROSCI.4973-10.2011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paton J, Belova M, Morrison S, Salzman C. The primate amygdala represents the positive and negative value of visual stimuli during learning. Nature. 2006;439:865–870. doi: 10.1038/nature04490. nature04490 [pii] 10.1038/nature04490. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Paulsen DJ, Carter RM, Platt ML, Huettel SA, Brannon EM. Neurocognitive development of risk aversion from early childhood to adulthood. Front Hum Neurosci. 2011;5:178. doi: 10.3389/fnhum.2011.00178. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Payne C, Machado CJ, Bliwise NG, Bachevalier J. Maturation of the hippocampal formation and amygdala in Macaca mulatta: a volumetric magnetic resonance imaging study. Hippocampus. 2010;20:922–935. doi: 10.1002/hipo.20688. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peck CJ, Lau B, Salzman CD. The primate amygdala combines information about space and value. Nature Neuroscience. 2013;16:340–348. doi: 10.1038/nn.3328. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Pennartz CMA, Ito R, Verschure PFMJ, Battaglia FP, Robbins TW. The hippocampal striatal axis in learning, prediction and goal-directed behavior. Trends Neurosci. 2011;34:548–559. doi: 10.1016/j.tins.2011.08.001. [DOI] [PubMed] [Google Scholar]
- Peper JS, Mandl RCW, Braams BR, de Water E, Heijboer AC, Koolschijn PCMP, Crone EA. Delay discounting and frontostriatal fiber tracts: a combined DTI and MTR study on impulsive choices in healthy young adults. Cereb Cortex. 2013;23:1695–1702. doi: 10.1093/cercor/bhs163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Peper JS, van den Heuvel MP, Mandl RCW, Pol HEH, van Honk J. Sex steroids and connectivity in the human brain: A review of neuroimaging studies. Psychoneuroendocrinology. 2011;36:1101–1113. doi: 10.1016/j.psyneuen.2011.05.004. [DOI] [PubMed] [Google Scholar]
- Pessiglione M, Seymour B, Flandin G, Dolan R, Frith C. Dopamine-dependent prediction errors underpin reward-seeking behaviour in humans. Nature. 2006;442:1042–1045. doi: 10.1038/nature05051. nature05051 [pii] 10.1038/nature05051. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Phelps EA, Delgado MR, Nearing KI, LeDoux JE. Extinction learning in humans: role of the amygdala and vmPFC. Neuron. 2004;43:897–905. doi: 10.1016/j.neuron.2004.08.042. [DOI] [PubMed] [Google Scholar]
- Phelps EA, LeDoux JE. Contributions of the amygdala to emotion processing: from animal models to human behavior. Neuron. 2005;48:175–187. doi: 10.1016/j.neuron.2005.09.025. [DOI] [PubMed] [Google Scholar]
- Phelps EA, Lempert KM, Sokol-Hessner P. Emotion and Decision Making: Multiple Modulatory Neural Circuits. Annual Review of Neuroscience. 2014;37:263–287. doi: 10.1146/annurev-neuro-071013-014119. [DOI] [PubMed] [Google Scholar]
- Pizoli CE, Shah MN, Snyder AZ, Shimony JS, Limbrick DD, Raichle ME, Schlaggar BL, Smyth MD. Resting-state activity in development and maintenance of normal brain function. Proc Natl Acad Sci USA. 2011;108:11638–11643. doi: 10.1073/pnas.1109144108. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Plassmann H, O’Doherty J, Rangel A. Orbitofrontal cortex encodes willingness to pay in everyday economic transactions. J Neurosci. 2007;27:9984–9988. doi: 10.1523/JNEUROSCI.2131-07.2007. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Porter JN, Roy AK, Benson B, Carlisi C, Collins PF, Leibenluft E, Pine DS, Luciana M, Ernst M. Age-related changes in the intrinsic functional connectivity of the human ventral vs. dorsal striatum from childhood to middle age. Developmental Brain Research. 2014 doi: 10.1016/j.dcn.2014.08.011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Power JD, Barnes KA, Snyder AZ, Schlagger BL, Petersen SE. Spurious but systematic correlations in functional connectivity MRI networks arise from subject motion. NeuroImage. 2012;59:2142–2154. doi: 10.1016/j.neuroimage.2011.10.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Preuschoff K, Quartz SR, Bossaerts P. Human insula activation reflects risk prediction errors as well as risk. Journal of Neuroscience. 2008;28:2745–2752. doi: 10.1523/JNEUROSCI.4286-07.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Price JL, Drevets WC. Neural circuits underlying the pathophysiology of mood disorders. Trends Cogn Sci (Regul Ed) 2012;16:61–71. doi: 10.1016/j.tics.2011.12.011. [DOI] [PubMed] [Google Scholar]
- Qin S, Young CB, Supekar K, Uddin LQ, Menon V. Immature integration and segregation of emotion-related brain circuitry in young children. Proc Natl Acad Sci USA. 2012;109:7941–7946. doi: 10.1073/pnas.1120408109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raichle ME. Two views of brain function. Trends Cogn Sci (Regul Ed) 2010;14:180–190. doi: 10.1016/j.tics.2010.01.008. [DOI] [PubMed] [Google Scholar]
- Raichle ME, MacLeod AM, Snyder AZ, Powers WJ, Gusnard DA, Shulman GL. A default mode of brain function. Proc Natl Acad Sci USA. 2001;98:676–682. doi: 10.1073/pnas.98.2.676. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rangel A, Camerer C, Montague PR. A framework for studying the neurobiology of value-based decision making. Nat Rev Neurosci. 2008;9:545–556. doi: 10.1038/nrn2357. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Raz A, Lieber B, Soliman F, Buhle J, Posner J, Peterson BS, Posner MI. Ecological nuances in functional magnetic resonance imaging (fMRI): psychological stressors, posture, and hydrostatics. NeuroImage. 2005;25:1–7. doi: 10.1016/j.neuroimage.2004.11.015. [DOI] [PubMed] [Google Scholar]
- Raznahan A, Shaw PW, Lerch JP, Clasen LS, Greenstein D, Berman R, Pipitone J, Chakravarty MM, Giedd JN. Longitudinal four-dimensional mapping of subcortical anatomy in human development. Proc Natl Acad Sci USA. 2014;111:1592–1597. doi: 10.1073/pnas.1316911111. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Reynolds SM, Zahm DS. Specificity in the projections of prefrontal and insular cortex to ventral striatopallidum and the extended amygdala. Journal of Neuroscience. 2005;25:11757–11767. doi: 10.1523/JNEUROSCI.3432-05.2005. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rice W. Analyzing Tables of Statistical Tests. Evolution. 1989;43:223–225. doi: 10.1111/j.1558-5646.1989.tb04220.x. [DOI] [PubMed] [Google Scholar]
- Richards JM, Plate RC, Ernst M. A systematic review of fMRI reward paradigms used in studies of adolescents vs. adults: the impact of task design and implications for understanding neurodevelopment. Neuroscience and biobehavioral reviews. 2013;37:976–991. doi: 10.1016/j.neubiorev.2013.03.004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Riedl V, Bienkowska K, Strobel C, Tahmasian M, Grimmer T, Forster S, Friston KJ, Sorg C, Drzezga A. Local Activity Determines Functional Connectivity in the Resting Human Brain: A Simultaneous FDG-PET/fMRI Study. Journal of Neuroscience. 2014;34:6260–6266. doi: 10.1523/JNEUROSCI.0492-14.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Rilling JK. The neural and hormonal basis of human parental care. Neuropsychologia. 2013;51:731–747. doi: 10.1016/j.neuropsychologia.2012.12.017. [DOI] [PubMed] [Google Scholar]
- Robbins T, Everitt B. Neurobehavioural mechanisms of reward and motivation. Curr Opin Neurobiol. 1996;6:228–236. doi: 10.1016/s0959-4388(96)80077-8. doi::S0959-4388(96)80077-8 [pii] [DOI] [PubMed] [Google Scholar]
- Robbins TW, Cador M, Taylor JR, Everitt BJ. Limbic-striatal interactions in reward-related processes. Neuroscience and biobehavioral reviews. 1989;13:155–162. doi: 10.1016/s0149-7634(89)80025-9. [DOI] [PubMed] [Google Scholar]
- Rolls E. The orbitofrontal cortex and reward. Cerebral Cortex. 2000 doi: 10.1093/cercor/10.3.284. [DOI] [PubMed] [Google Scholar]
- Saad Z, Reynolds RC, Jo HJ, Gotts SJ, Chen G, Martin A, Cox R. Correcting Brain-Wide Correlation Differences in Resting-State FMRI. Brain Connect. 2013 doi: 10.1089/brain.2013.0156. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Salzman C, Paton J, Belova M, Morrison S. Flexible neural representations of value in the primate brain. Annals of the New York Academy of Sciences. 2007;1121:336–354. doi: 10.1196/annals.1401.034. annals.1401.034 [pii] 10.1196/annals.1401.034. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Satpute AB, Mumford JA, Naliboff BD, Poldrack RA. Human anterior and posterior hippocampus respond distinctly to state and trait anxiety. Emotion. 2012;12:58–68. doi: 10.1037/a0026517. [DOI] [PubMed] [Google Scholar]
- Satterthwaite TD, Elliott MA, Gerraty RT, Ruparel K, Loughead J, Calkins ME, Eickhoff SB, Hakonarson H, Gur RC, Gur RE, Wolf DH. An improved framework for confound regression and filtering for control of motion artifact in the preprocessing of resting-state functional connectivity data. NeuroImage. 2013;64:240–256. doi: 10.1016/j.neuroimage.2012.08.052. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Satterthwaite TD, Wolf DH, Loughead J, Ruparel K, Elliott MA, Hakonarson H, Gur RC, Gur RE. Impact of in-scanner head motion on multiple measures of functional connectivity: relevance for studies of neurodevelopment in youth. NeuroImage. 2012;60:623–632. doi: 10.1016/j.neuroimage.2011.12.063. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Scherf KS, Sweeney JA, Luna B. Brain basis of developmental change in visuospatial working memory. Journal of Cognitive Neuroscience. 2006;18:1045–1058. doi: 10.1162/jocn.2006.18.7.1045. [DOI] [PubMed] [Google Scholar]
- Schmidt B, Hinman JR, Jacobson TK, Szkudlarek E, Argraves M, Escabi MA, Markus EJ. Dissociation between Dorsal and Ventral Hippocampal Theta Oscillations during Decision-Making. Journal of Neuroscience. 2013;33:6212–6224. doi: 10.1523/JNEUROSCI.2915-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schulz KM, Molenda-Figueira HA, Sisk CL. Back to the future: The organizational-activational hypothesis adapted to puberty and adolescence. Hormones and Behavior. 2009a;55:597–604. doi: 10.1016/j.yhbeh.2009.03.010. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Schulz KM, Zehr JL, Salas-Ramirez KY, Sisk CL. Testosterone programs adult social behavior before and during, but not after, adolescence. Endocrinology. 2009b;150:3690–3698. doi: 10.1210/en.2008-1708. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Seymour B, O’Doherty JP, Dayan P, Koltzenburg M, Jones AK, Dolan RJ, Friston KJ, Frackowiak RS. Temporal difference models describe higher-order learning in humans. Nature. 2004;429:664–667. doi: 10.1038/nature02581. [DOI] [PubMed] [Google Scholar]
- Silvers JA, Insel C, Powers A, Franz P, Weber J, Mischel W, Casey BJ, Ochsner KN. Curbing Craving: Behavioral and Brain Evidence That Children Regulate Craving When Instructed to Do So but Have Higher Baseline Craving Than Adults. Psychol Sci. 2014;25:1932–1942. doi: 10.1177/0956797614546001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Singer T, Critchley HD, Preuschoff K. A common role of insula in feelings, empathy and uncertainty. Trends Cogn Sci (Regul Ed) 2009;13:334–340. doi: 10.1016/j.tics.2009.05.001. [DOI] [PubMed] [Google Scholar]
- Smith AR, Steinberg L, Chein J. The Role of the Anterior Insula in Adolescent Decision Making. Dev Neurosci. 2014;36:196–209. doi: 10.1159/000358918. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Somerville LH, Casey BJ. Developmental neurobiology of cognitive control and motivational systems. Curr Opin Neurobiol. 2010;20:236–241. doi: 10.1016/j.conb.2010.01.006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Somerville LH, Hare T, Casey BJ. Frontostriatal maturation predicts cognitive control failure to appetitive cues in adolescents. Journal of Cognitive Neuroscience. 2011;23:2123–2134. doi: 10.1162/jocn.2010.21572. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Somerville LH, Jones RM, Casey BJ. A time of change: behavioral and neural correlates of adolescent sensitivity to appetitive and aversive environmental cues. Brain Cogn. 2010;72:124–133. doi: 10.1016/j.bandc.2009.07.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Sowell ER, Thompson PM, Holmes CJ, Jernigan TL, Toga AW. In vivo evidence for post-adolescent brain maturation in frontal and striatal regions. Nature Neuroscience. 1999;2:859–861. doi: 10.1038/13154. [DOI] [PubMed] [Google Scholar]
- Sowell ER, Trauner DA, Gamst A, Jernigan TL. Development of cortical and subcortical brain structures in childhood and adolescence: a structural MRI study. Dev Med Child Neurol. 2002;44:4–16. doi: 10.1017/s0012162201001591. [DOI] [PubMed] [Google Scholar]
- Spear LP. The adolescent brain and age-related behavioral manifestations. Neuroscience and biobehavioral reviews. 2000;24:417–463. doi: 10.1016/s0149-7634(00)00014-2. [DOI] [PubMed] [Google Scholar]
- Spielberger C. Manual for the State-Trait Anxiety Inventory. rev. Consulting Psychologists Press; Palo Alto, CA: 1983. [Google Scholar]
- Steinberg L. A Social Neuroscience Perspective on Adolescent Risk-Taking. Dev Rev. 2008;28:78–106. doi: 10.1016/j.dr.2007.08.002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Swartz JR, Carrasco M, Wiggins JL, Thomason ME, Monk CS. Age-related changes in the structure and function of prefrontal cortex-amygdala circuitry in children and adolescents: a multi-modal imaging approach. NeuroImage. 2014;86:212–220. doi: 10.1016/j.neuroimage.2013.08.018. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Talairach J, Tournoux P. Co-planar Stereotaxic Atlas of the Human Brain. George Thieme Verlag; 1988. [Google Scholar]
- Tottenham N, Shapiro M, Telzer EH, Humphreys KL. Amygdala response to mother. Dev Sci. 2012;15:307–319. doi: 10.1111/j.1467-7687.2011.01128.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tottenham N, Sheridan MA. A review of adversity, the amygdala and the hippocampus: a consideration of developmental timing. Front Hum Neurosci. 2009;3:68. doi: 10.3389/neuro.09.068.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tymula A, Rosenberg Belmaker LA, Roy AK, Ruderman L, Manson K, Glimcher PW, Levy I. Adolescents’ risk-taking behavior is driven by tolerance to ambiguity. Proc Natl Acad Sci USA. 2012 doi: 10.1073/pnas.1207144109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Tziortzi AC, Searle GE, Tzimopoulou S, Salinas C, Beaver JD, Jenkinson M, Laruelle M, Rabiner EA, Gunn RN. Imaging dopamine receptors in humans with [11C]-(+)-PHNO: dissection of D3 signal and anatomy. NeuroImage. 2011;54:264–277. doi: 10.1016/j.neuroimage.2010.06.044. [DOI] [PubMed] [Google Scholar]
- Utevsky AV, Smith DV, Huettel SA. Precuneus Is a Functional Core of the Default-Mode Network. Journal of Neuroscience. 2014;34:932–940. doi: 10.1523/JNEUROSCI.4227-13.2014. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van den Bos W, Cohen MX, Kahnt T, Crone EA. Striatum-Medial Prefrontal Cortex Connectivity Predicts Developmental Changes in Reinforcement Learning. Cereb Cortex. 2012;22:1247–1255. doi: 10.1093/cercor/bhr198. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van den Bos W, Talwar A, McClure SM. Neural Correlates of Reinforcement Learning and Social Preferences in Competitive Bidding. Journal of Neuroscience. 2013;33:2137–2146. doi: 10.1523/JNEUROSCI.3095-12.2013. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Dijk KRA, Hedden T, Venkataraman A, Evans KC, Lazar SW, Buckner RL. Intrinsic functional connectivity as a tool for human connectomics: theory, properties, and optimization. J Neurophysiol. 2010;103:297–321. doi: 10.1152/jn.00783.2009. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Dijk KRA, Sabuncu MR, Buckner RL. The influence of head motion on intrinsic functional connectivity MRI. NeuroImage. 2012;59:431–438. doi: 10.1016/j.neuroimage.2011.07.044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Duijvenvoorde ACK, Huizenga HM, Somerville LH, Delgado MR, Powers A, Weeda WD, Casey BJ, Weber EU, Figner B. Neural Correlates of Expected Risks and Returns in Risky Choice across Development. Journal of Neuroscience. 2015;35:1549–1560. doi: 10.1523/JNEUROSCI.1924-14.2015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- van Duijvenvoorde ACK, Zanolie K, Rombouts SARB, Raijmakers MEJ, Crone EA. Evaluating the negative or valuing the positive? Neural mechanisms supporting feedback-based learning across development. Journal of Neuroscience. 2008;28:9495–9503. doi: 10.1523/JNEUROSCI.1485-08.2008. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Van Leijenhorst L, Gunther Moor B, Op de Macks ZA, Rombouts SARB, Westenberg PM, Crone EA. Adolescent risky decision-making: neurocognitive development of reward and control regions. NeuroImage. 2010a;51:345–355. doi: 10.1016/j.neuroimage.2010.02.038. [DOI] [PubMed] [Google Scholar]
- Van Leijenhorst L, Zanolie K, Van Meel CS, Westenberg PM, Rombouts SARB, Crone EA. What motivates the adolescent? Brain regions mediating reward sensitivity across adolescence. Cereb Cortex. 2010b;20:61–69. doi: 10.1093/cercor/bhp078. [DOI] [PubMed] [Google Scholar]
- Vink M, Derks JM, Hoogendam JM, Hillegers M, Kahn RS. Functional differences in emotion processing during adolescence and early adulthood. NeuroImage. 2014;91:70–76. doi: 10.1016/j.neuroimage.2014.01.035. [DOI] [PubMed] [Google Scholar]
- Weschsler D. Wechsler Abbreviated Scale of Intelligence. The Psychological Corporation: Harcourt Brace & Company; New York: 1999. [Google Scholar]
- Wierenga L, Langen M, Ambrosino S, van Dijk S, Oranje B, Durston S. Typical development of basal ganglia, hippocampus, amygdala and cerebellum from age 7 to 24. NeuroImage. 2014;96:67–72. doi: 10.1016/j.neuroimage.2014.03.072. [DOI] [PubMed] [Google Scholar]
- Wimmer GE, Daw ND, Shohamy D. Generalization of value in reinforcement learning by humans. Eur J Neurosci. 2012;35:1092–1104. doi: 10.1111/j.1460-9568.2012.08017.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Wimmer GE, Shohamy D. Preference by association: how memory mechanisms in the hippocampus bias decisions. Science. 2012;338:270–273. doi: 10.1126/science.1223252. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.