

Abstract
Background
The suprachiasmatic nucleus (SCN) contains the master circadian clock that regulates daily rhythms of many physiological and behavioural processes in mammals. Previously we have shown that prokineticin 2 (PK2) is a clock-controlled gene that may function as a critical SCN output molecule responsible for circadian locomotor rhythms. As light is the principal zeitgeber that entrains the circadian oscillator, and PK2 expression is responsive to nocturnal light pulses, we further investigated the effects of light on the molecular rhythm of PK2 in the SCN. In particular, we examined how PK2 responds to shifts of light/dark cycles and changes in photoperiod. We also investigated which photoreceptors are responsible for the light-induced PK2 expression in the SCN. To determine whether light requires an intact functional circadian pacemaker to regulate PK2, we examined PK2 expression in cryptochrome1,2-deficient (Cry1-/-Cry2-/-) mice that lack functional circadian clock under normal light/dark cycles and constant darkness.
Results
Upon abrupt shifts of the light/dark cycle, PK2 expression exhibits transients in response to phase advances but rapidly entrains to phase delays. Photoperiod studies indicate that PK2 responds differentially to changes in light period. Although the phase of PK2 expression expands as the light period increases, decreasing light period does not further condense the phase of PK2 expression. Genetic knockout studies revealed that functional melanopsin and rod-cone photoreceptive systems are required for the light-inducibility of PK2. In Cry1-/-Cry2-/- mice that lack a functional circadian clock, a low amplitude PK2 rhythm is detected under light/dark conditions, but not in constant darkness. This suggests that light can directly regulate PK2 expression in the SCN.
Conclusion
These data demonstrate that the molecular rhythm of PK2 in the SCN is regulated by both the circadian clock and light. PK2 is predominantly controlled by the endogenous circadian clock, while light plays a modulatory role. The Cry1-/-Cry2-/- mice studies reveal a light-driven PK2 rhythm, indicating that light can induce PK2 expression independent of the circadian oscillator. The light inducibility of PK2 suggests that in addition to its role in clock-driven rhythms of locomotor behaviour, PK2 may also participate in the photic entrainment of circadian locomotor rhythms.
Background
Light is the principal zeitgeber that entrains circadian rhythms of physiology and behaviour [1,2]. The major light input pathway to the suprachiasmatic nucleus (SCN) is the retinohypothalamic tract [3], which arises from a population of retinal ganglion cells [4]. Recent studies have demonstrated that melanopsin-containing retinal ganglion cells, rods, and cones all convey photic information to the SCN, and mice lacking these photoreceptive systems cannot be entrained by light [5-11]. Excellent progress has been made in the understanding of circadian photic entrainment [12-15]. This includes light-induced transcriptional activation of core clock genes in the SCN, such as Per1 and Per2, as well as immediate-early gene c-fos. Exposure to light pulses at night induces expression of these genes in the SCN, and this light induction mechanism has been suggested as a critical pathway for the resetting of circadian clock in response to changes in light/dark conditions [16-19]. Intercellular signalling mechanisms between SCN neurons are also important in circadian photic entrainment, as mice with mutation in a neuropeptide receptor for VIP (Vasoactive Intestinal Peptide) and PACAP (Pituitary Adenylate Cyclase Activating Peptide) are unable to sustain normal circadian behaviour and exhibit loss of sensitivity to light [20].
In addition to the effect of light on circadian entrainment, light also has a direct effect on physiology and behaviour, generally termed as "masking" [21,22]. For instance, light pulses given at night acutely suppress the locomotor behaviour of nocturnal rodents [21,22], and this can occur without functional clockwork [23-27]. Masking may account for the maintenance under normal light/dark conditions of wheel-running rhythms in cryptochrome-deficient (Cry1-/-Cry2-/-) mice, which are behaviourally arrhythmic under constant darkness. The contribution of masking to normal locomotor activity rhythms is unclear, as is the participation of the SCN in masking effects of light. Vitaterna et al (1999) first observed a light-driven Per2 rhythm in the SCN in Cry1-/-Cry2-/- mice, and have suggested that the light-driven molecular rhythm in the SCN may be related to the preservation of their locomotor rhythm [25].
We previously found that prokineticin 2 (PK2) is a first order clock-controlled gene, whose expression in the SCN is regulated by CLOCK and BMAL1 acting on the E-boxes in the gene's promoter [28]. We have also demonstrated that PK2 may function as a SCN output molecule that transmits circadian locomotor rhythm via activation of a G protein-coupled receptor [28,29]. Interestingly, we also observed that PK2 expression is rapidly induced by light pulses administered at night [28], a characteristic that is usually seen with core clockwork genes but not clock-controlled genes. Here we further investigated the light regulation of the rhythm of PK2 expression in the SCN. In particular, we investigated the photoreceptive mechanisms responsible for the light-induced PK2 expression in the SCN. Utilizing Cry1-/-Cry2-/- mice, we also determined whether light can drive PK2 expression in the SCN independent of a functional circadian clock.
Results
PK2 responds differentially to the delay and advance of light/dark cycles
We first examined the effects of abrupt shifts of light/dark cycles on PK2 mRNA rhythm in the SCN. Animals were first entrained for two weeks under 12 hour light: 12 hour dark (LD), then subjected to either a 6 hour delay (6hrD) shift or 6 hour advance (6hrA) shift of light/dark cycles. We measured PK2 mRNA in the SCN of these animals to examine how quickly the PK2 mRNA rhythm re-entrains to the shifted light/dark cycles. Under LD, PK2 mRNA peaks during the day and remains low or undetectable during the night. During the first cycle of the delayed shift (6hrD), the PK2 mRNA rhythm responds quickly: the rising phase of PK2 expression adjusts rapidly to the delayed light/dark cycles, while the falling phase still resembles that of the unshifted light/dark cycles (Figure 1A). In contrast, the PK2 mRNA rhythm responds very little to a 6 hour advance shift (6hrA). During the first cycle of the advance shift, the PK2 oscillation pattern remains similar to that of the unshifted LD (Figure 1B). These changes in PK2 expression during 6hrD or 6hrA shift indicate that the endogenous circadian clock exerts dominant control over the PK2 rhythm, as PK2 expression cannot respond immediately and completely to the shifts of light/dark cycles.
Figure 1.
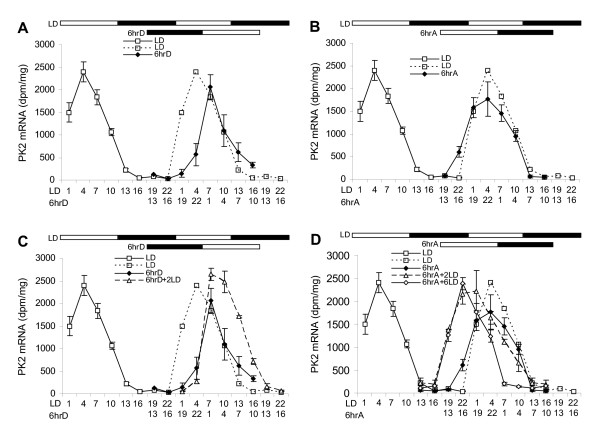
Temporal profiles of PK2 mRNA in the SCN in response to abrupt shifts of light/dark cycles. Animals were entrained to 12L:12D (LD) and subjected to either 6-hour delay of light/dark cycles (6hrD), 6-hour advance (6hrA), 6-hour delay followed by adaptation of 2 additional LD (6hrD+2LD), or 6-hour advance followed by adaptation of 2 additional LD (6hrA+2LD) or 6 additional LD (6hrA+6LD). Open and filled horizontal bars indicate light and dark periods, respectively. The LD data is doubled plotted as dashed line (open square) in all graphs. The zeitgeber time (ZT) on the x-axis reflects the timescale for LD, 6hrD or 6hrA. Please note that the additional LD adaptation groups use the same timescale as the 6hrD or 6hrA. (A) Temporal profiles of PK2 mRNA under 6hrD and LD. Note that PK2 mRNA responds quickly to the 6hrD shift. (B) Temporal profiles of PK2 mRNA rhythm under 6hrA and LD. Note that PK2 mRNA did not adjust to the 6hrA shift. (C) Temporal profiles of PK2 mRNA rhythm under 6hrD, 6hrD+2LD and LD to indicate adaptation of PK2 rhythm under 6hrD. Note that PK2 rhythm is stably entrained to 6hrD after two days. (D) Temporal profiles of PK2 mRNA rhythm under 6hrA, 6hrA+2LD, 6hrA+6LD and LD to illustrate adaptation of PK2 rhythm under 6hrA. Note that PK2 rhythm did not stably entrained to 6hrA until after 6 days. Each value is the mean ± SEM of 3 animals.
As it normally takes about 1–2 days for locomotor rhythms to stably entrain to phase delays and about 5–6 days to entrain to phase advances [30,31], we next examined the timecourse of shifts of the PK2 rhythm to 6 hour phase advances and delays. Consistent with the animal's locomotor behaviour, the PK2 mRNA rhythm reaches stable phase within 2 days of 6hrD shift (Figure 1C). In contrast, only the rise of PK2 reaches stable phase within 2 days of 6hrA shift, while the fall of PK2 takes longer (Figure 1D). Thus, we further examined whether the PK2 rhythm is stably entrained after 6 days of 6hrA shift. As expected, the PK2 rhythm is completely entrained to 6hrA shift after 6 days (Figure 1D). Together, the differential responses of PK2 rhythm to a 6hrD or 6hrA shift indicate that the endogenous circadian clock predominantly controls PK2 rhythm, as circadian oscillators typically show rapid phase delays but advance with transients [31,32]. The entrainment patterns of PK2 during phase shifts are consistent with behavioural studies in animals and human subjects [30,31].
PK2 rhythm is entrained by different photoperiods
We next examined the effect of photoperiod on the PK2 molecular rhythm in the SCN. PK2 mRNA was measured in the SCN of mice entrained under different photoperiods: 8 hour light: 16 hour dark (8L:16D), 16 hour light: 8 hour dark (16L:8D), or 20 hour light: 4 hour dark (20L:4D). During 12L:12D, PK2 mRNA is highly expressed during the 12 hour light phase with peak level at ZT4 (Figure 1A, Figure 3A). Under 16L:8D, PK2 mRNA expands to the entire 16 hour light phase and is essentially undetectable during the 8 hour dark period (Figure 2B). However, the expression of PK2 mRNA is not confined to the light phase of the shorter photoperiod (8L:16D), as PK2 mRNA rises before lights on and persists after lights off (Figure 2A). The temporal profile of PK2 mRNA under this short photoperiod (8L:16D) is very similar to that observed under 12L:12D (Figure 1A, Figure 3A) or constant darkness (2DD) [28]. Thus, although light can induce PK2 mRNA and expand the duration of PK2 expression, the phase angle of PK2 expression is determined by the circadian clock, and its duration cannot be further compressed under shorter photoperiods. Interestingly, the peak of PK2 mRNA expression was significantly higher in long days (16L:8D) than in shorter days (8L:16D) (Figure 2A–B), further indicate the enhancing effect of light on PK2 expression. However, a significant reduction in the PK2 peak level is observed under a very long photoperiod (20L: 4D) (Figure 2C). We also noticed that under 20L:4D, PK2 mRNA is further expanded and becomes detectable even in dark phase (Figure 2C). Under this long photoperiod (20L:4D), the difference between the peak and basal level of PK2 is only about 4 fold (Figure 2C). As it has been reported that the rhythms of mPer1 and mPer2 mRNAs in the SCN are also entrained with different phase angles under a variety of photoperiods [33-35], we have also examined Per1 and Per2 rhythm in our photoperiod studies (see Additional file 1). The Per1 and Per2 rhythm we observed under these photoperiods are consistent with previous findings [35]. Taken together, these results indicate that changes in photoperiod alter PK2 rhythm in the SCN, and the amplitudes of PK2 mRNA oscillation are greatly reduced in very long photoperiods.
Figure 3.
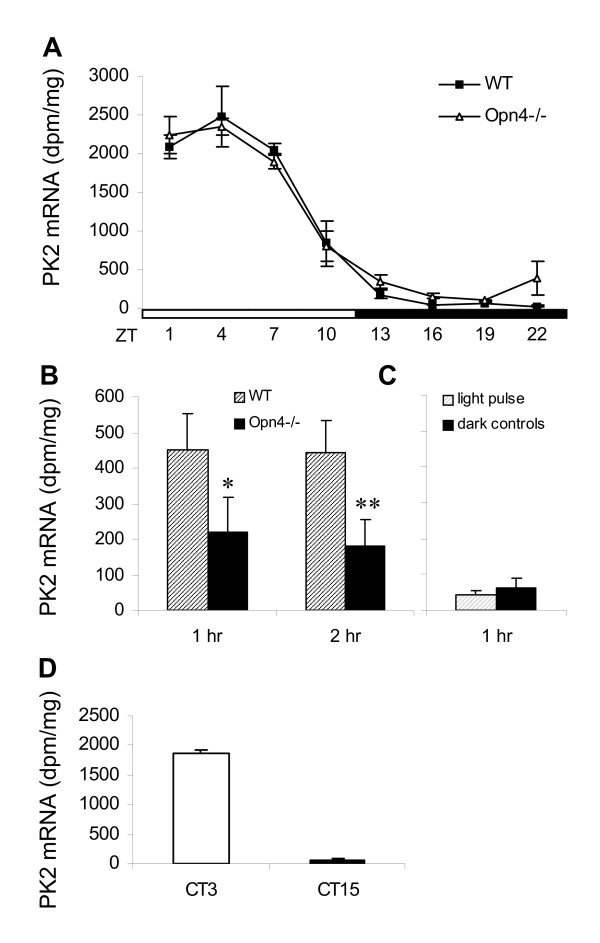
PK2 mRNA rhythm in the SCN of melanopsin-deficient (Opn4-/-) mice and triple knockout mice (Opn4-/- Gnat1-/- Cnga3-/- mice). (A) Temporal profiles of PK2 mRNA rhythm in wildtype (filled squares) and Opn4-/- mice (open triangles) under LD. Open and filled bars indicate light and dark periods, respectively. Each value represents the mean ± SEM of 3–4 animals. Two-way ANOVA indicated that there is no significant difference between genotypes. (B) Light pulse-induced PK2 mRNA expression in wildtype (shaded bars) and Opn4-/- mice (filled bars). PK2 mRNA was measured one and two hours after brief light pulse at ZT14. Each value represents the mean ± SEM of 6–8 animals. *p < 0.05, **p < 0.01, Student's t-test. (C) Light-pulse induced PK2 mRNA expression in triple knockout mice that lack melanopsin, rod and cone photoreceptive system (Opn4-/- Gnat1-/- Cnga3-/- mice). Dark controls received no light pulse. Each value represents the mean ± SEM of 3 animals. (D) PK2 mRNA expression in triple knockout mice at circadian time (CT) 3 and 15.
Figure 2.
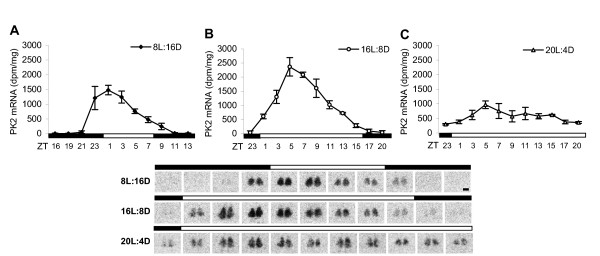
Effects of photoperiods on PK2 mRNA rhythm in the SCN. Temporal profiles of PK2 rhythm under 8L:16D (A), 16L:8D (B) and 20L:4D (C). Open and filled bars indicate light and dark periods, respectively. The zeitgeber time (ZT) on the x-axis reflects the timescale for each photoperiod. Each value represents the mean ± SEM of 3–4 animals. One-way ANOVA indicated that peak levels of all groups are significantly different from each other, p < 0.05–0.001 using Bonferroni's post-hoc test. Autoradiographic images show representative mRNA expression of PK2 during these photoperiods.
Light inducibility of PK2 is eliminated in mice that lack melanopsin, rod and cone phototransduction system (Opn4-/-, Gnat1-/- Cnga3-/- mice)
As melanopsin has been implicated in circadian photoreception [5-11], we examined whether the PK2 molecular rhythm is normally entrained in melanopsin-deficient (Opn4-/-) mice. Figure 3 shows that the oscillation profile of PK2 in the SCN of Opn4-/- mice is essentially identical to that observed in the wild type mice under LD. This normal temporal profile of PK2 mRNA corresponds with the normal locomotor rhythm of Opn4-/- mice under light/dark conditions [7,8]. As Opn4-/- mice display attenuated phase resetting in response to light pulses and exhibit impaired light masking responses to bright light [36], we also examined whether light inducibility of PK2 is blunted in Opn4-/- mice. Figure 3B shows that light pulse-induced PK2 in the SCN of Opn4-/- mice was significantly reduced by about 50% and 60%, one and two hours after the light pulse, respectively.
The Opn4-/- light pulse studies show that a residual PK2 expression is still present after a light pulse, suggesting that without melanopsin, other phototransduction system can still transmit light information to induce PK2 expression. Thus, we decided to examine the light inducibility of PK2 in triple knockout mice lacking melanopsin, rod and cone phototransduction systems (Opn4-/- Gnat1-/- Cnga3-/- mice), as these animals free run under light dark conditions (LD) and lack masking responses to light [10]. Figure 3C shows that the light pulse-induced PK2 in the SCN is completely eliminated in these triple knockout mice, consistent with their malfunctioning photoentrainment systems and their lack of masking responses to light [10]. In addition, we also observed that PK2 mRNA followed the free-running locomotor rhythms in these triple knockout mice (Figure 3D), with high levels of PK2 during the inactive phase (CT3) and low levels during active phase (CT15). Together, these results suggest that melanopsin contributes to the light inducibility of PK2, and intact melanopsin with functional rod/cone phototransduction systems are required for the light inducibility of PK2.
A low amplitude PK2 rhythm is preserved in cryptochrome-deficient (Cry1-/-Cry2-/-) mice under light/dark conditions
Previous studies have shown that the light-regulated Per2 rhythm is maintained in the SCN of cryptochrome-deficient (Cry1-/-Cry2-/-) mice that lack functional circadian clock [25,37]. In order to determine whether the regulation of PK2, Per1, Per2 and Bmal1 expression by light requires an intact circadian pacemaker, we systematically assessed the temporal mRNA profiles of clockwork genes in Cry1-/-Cry2-/- mice under both light/dark (LD) and constant dark (DD) conditions. Figure 4 shows that the molecular rhythm of Per2 remained largely intact in Cry1-/-Cry2-/- mice entrained under12L:12D, with levels about 4-fold higher during the light phase than the dark phase. This amplitude of the Per2 oscillation profile was similar to that observed in wild type mice [18,38]. A low amplitude Per1 rhythm in Cry1-/-Cry2-/ mice was also apparent under LD, but not DD (Figure 4B). We further detected a light-driven Bmal1 rhythm in the SCN of Cry1-/-Cry2-/- mice under LD, but not DD (Figure 4C). Interestingly, this Bmal1 rhythm in Cry1-/-Cry2-/- mice peaked during light phase, opposite from the Bmal1 rhythm in wild type mice and in phase with Per1 [39,40]. As it has been suggested that PER2 can positively regulate Bmal1 expression via inhibition of the orphan nuclear receptor REV-ERBα [41,42], it is possible that this Bmal1 rhythm is secondary to the light-driven Per2 rhythm. Further studies are required to clarify this observation.
Figure 4.
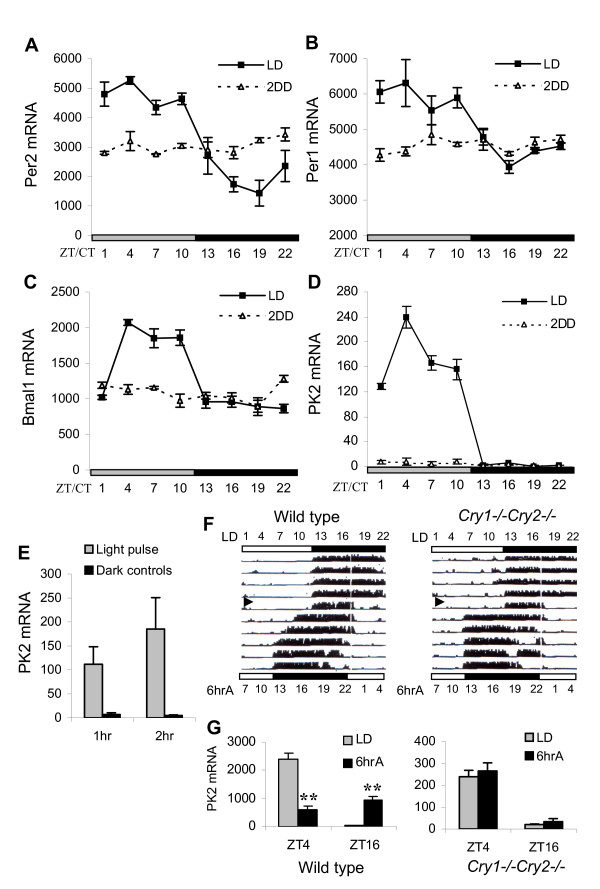
Light-driven molecular rhythms in the SCN of Cry1-/-Cry2-/- mice. Temporal mRNA profiles of Per2 (A), Per1 (B), Bmal1 (C) and PK2 (D) in Cry1-/-Cry2-/- mice under 12L:12D (LD) or two days constant darkness (2DD). Each value represents the mean ± SEM of 3–4 animals. Two-way ANOVA with Bonferroni's posthoc analysis was used to test for significant interactions between expression across time of sampling and under different lighting conditions (LD vs 2DD). p < 0.0001 (Per2), p < 0.002 (Per1), p < 0.0001 (Bmal1) and p < 0.0001 (PK2). (E) Light pulse-induced PK2 mRNA in Cry1-/-Cry2-/- mice. PK2 mRNA was measured one and two hours after brief light pulse at ZT14 (shaded bar). Black bar represents dark controls that did not receive light pulse. Each value represents the mean ± SEM of 5–6 animals. Two-way ANOVA indicates a significant difference in PK2 expression between light and dark treatment (p < 0.05), however, the PK2 induction is not significantly different between the two timepoints (1 hr vs 2 hr). (F) Locomotor behavioural rhythms of wild type (left) and Cry1-/-Cry2-/- mice (right) in response to 6 hour advance of light/dark cycle. Open and filled bars indicate light and dark periods, respectively. Black arrow indicates the day of 6 hour advance shift (6hrA). Numbers above and below the actograms represent timescale in zeitgeber time (ZT) for LD and 6hrA. (G) Rapid adjustment of PK2 rhythm in Cry1-/-Cry2-/- mice to 6 hour advance (6hrA). PK2 mRNA was quantitated in the SCN of wildtype and Cry1-/-Cry2-/- mice under LD (shaded) or 6hrA (black). Each value represents the mean ± SEM of 3–4 animals. Three-way ANOVA with Bonferroni's post hoc analysis indicates a significant interaction between light/dark cycle (LD vs 6hrA), timepoint (ZT4 vs ZT16) and genotype (wildtype vs Cry1-/-Cry2-/- mice), p < 0.001. Two-way ANOVA with Bonferroni's post hoc analysis show that there is significant difference in wildtype PK2 expression level between LD and 6hrA, (ZT4, ***p < 0.001; ZT16, **p < 0.01), but not in Cry1-/-Cry2-/- mice (ZT4, p = 1.000; ZT16, p = 1.000).
We also examined the molecular rhythm of PK2 in Cry1-/-Cry2-/- mice. Figure 4D shows that PK2 mRNA rhythm in the SCN of Cry1-/-Cry2-/- mice was apparent under LD, with the presence of a low level PK2 during light phase and absence of PK2 during dark phase (see Additional file 2). Similar to wild type mice, the peak level of this low amplitude PK2 rhythm was around ZT4, although its peak was only about 8% of that observed in wild type mice (Figure. 4D, Figure 1A, Figure 3A). No PK2 rhythm was evident when Cry1-/-Cry2-/- mice were placed under DD (Figure 4D). Furthermore, the inducibility of PK2 to nocturnal light pulses is also maintained in Cry1-/-Cry2-/- mice. PK2 mRNA increased one and two hours after a brief light pulse at ZT14 (Figure 4E). Nevertheless, light-induced PK2 was still detected in Per1,2,3-/- mice and Clk-/- mice that lack functional circadian clock (Cheng, Weaver & Zhou, unpublished observations). As PK2 remains responsive to light in these clock mutant mice that lack functional circadian clock, it is likely that the low amplitude PK2 rhythm in Cry1-/-Cry2-/- mice under LD is directly driven by light.
In order to test whether this light-driven PK2 rhythm may be related to the maintenance of behavioural rhythms observed in Cry1-/-Cry2-/- mice under LD, we studied the responses of Cry1-/-Cry2-/- mice to a 6 hour advance of lighting schedule. In contrast to the transients of entrainment of locomotor rhythms in wild type mice (which takes about 4-5 days to re-entrain to phase advance), the locomotor activity of Cry1-/-Cry2-/- mice adjusted rapidly to 6 hr advance (Figure 4F). Such a rapid response is characteristic of masking. A correlative rapid adjustment of PK2 was also observed in the SCN of Cry1-/-Cry2-/- mice (Figure 4G). As Cry1-/-Cry2-/- mice lack functional circadian clock and their locomotor behaviour and PK2 expression patterns are completely light driven, our results suggest that this low amplitude, light-driven rhythm of PK2 may contribute to or underlie the masking of locomotor behaviour in these animals.
Discussion
Our studies indicate that the molecular rhythm of PK2 in the SCN is predominantly controlled by the circadian clock, with light playing a modulatory role. Abrupt shifts of light/dark cycles significantly altered the phase of the PK2 rhythm. While PK2 expression re-entrained rapidly to phase delays, it takes several cycles of transients for PK2 to be stably entrained to phase advances (Figure 1). The rate of re-entrainment of PK2 molecular rhythms to these shifts is consistent with that of behavioural adaptation of animals and human subjects [30,31]. Our photoperiod studies indicate that PK2 expression in the SCN responds differentially to changes in photoperiod length (Figure 2). Although increasing light period can induce PK2 expression and expand the duration of PK2 rhythm (Figure 2B), shortening of the light period does not lead to corresponding reduction of the duration of PK2 expression (Figure 2A). It appears that a minimal duration of PK2 expression is maintained under short photoperiod (Figure 2A) and constant darkness [28], which further indicate the dominant control of PK2 expression by the circadian clock. Interestingly, the amplitude of the PK2 oscillation was greatly reduced under very long photoperiod (20L:4D) (Figure 2C). As the amplitude of both Per1 and Per2 rhythms were also reduced during 20L:4D (see Additional file 1), it is likely that these depressed rhythms of clockwork genes may contribute to the depressed PK2 rhythm observed. Whether reduction in the amplitude of expression in any of these genes is related to arrhythmicity in LL deserves further examination.
Our studies with Cry1-/-Cry2-/- mice revealed the presence of a light-driven PK2 molecular rhythm in the SCN under LD, indicating that light can drive PK2 rhythm independent of functional circadian clock. Interestingly, the molecular rhythms of some clockwork genes such as Per2, Per1, and Bmal1 were also partially maintained in the SCN of Cry1-/-Cry2-/- mice under LD (Figure 4). Thus, light-driven molecular oscillations of clockwork or clock-controlled output genes exist in the absence of functional circadian clock. Vitaterna et al (1999) first noticed such light-regulated Per2 molecular rhythm in the SCN of Cry1-/-Cry2-/- mice, and suggested the term of "light-driving" effect [25]. As Cry1-/-Cry2-/- mice lack functional circadian clocks and their locomotor behaviour remains rhythmic under LD, but not under DD conditions [24,25], it is likely that these light-driven molecular rhythms may drive the locomotor rhythms in these animals. As we have previously shown that PK2 may be a critical output molecule responsible for circadian locomotor rhythms, the presence of this light-driven PK2 rhythm in Cry1-/-Cry2-/- mice may thus contribute to or underlie masking as well as the free running behavioural rhythms in these animals. It is well established that an intact SCN is necessary for the preservation of free running locomotor rhythms [43]. The role of the SCN in masking of locomotor activity by light is controversial, with similar studies having produced contradictory results [23,44]. Thus, it is possible that there might be common signal molecule(s) that mediate(s) the light-masking and the circadian clock-controlled locomotor behaviour. Construction of PK2-deficient mice will be necessary to resolve the exact role of PK2 in the light-driven locomotor rhythms.
The light inducibility of PK2 in the SCN is an unusual characteristic for a clock-controlled gene. Our results demonstrate that melanopsin-positive retinal ganglion cells, in conjunction with rods and cones, are responsible for the light-inducibility of PK2 (Figure 3). The same photoreceptive system has been shown responsible for the entrainment of locomotor rhythm [5-11]. The light inducibility of PK2 may be related to the presence of a putative cyclic AMP response element (CRE) in the promoter of the PK2 gene [28]. CRE-dependent activation is critical for light-induced gene expression in the SCN [45-48]. The reduced light inducibility of PK2 in mutant mice that lack functional clock may indicate that CRE-dependent pathway and CLK/BMAL1 transcriptional factors may interact in the light-induced PK2 expression in the SCN. Accumulative data have implicated the photic regulation of the transcription of clock genes such as Per1 and Per2 in the entrainment of behavioural rhythms [30,34]. The phase of the core SCN clock gene expression determines the timing of clock-controlled SCN output signals that ultimately regulate physiology and behaviour. Unlike the Per1 promoter, whose activation in the SCN shifts rapidly when the LD cycle is advanced [31], PK2 exhibits transients during phase advance, more similar to those of Cry1 and Cry2 [30,31]. This is consistent with the role for PK2 as a clock-controlled gene and thus is downstream from the light-regulated expression of Per1 or Per2. The presence of E box motifs in the PK2 promoter suggests that light-regulated Per1 (and perhaps Per2) expression can influence PK2 expression. However, the light inducibility of PK2 indicates that PK2 may have a more direct and central role in entrainment in addition to its putative role as an SCN output signal. In other words, whether PK2 functions completely outside the central circadian loops or partly within them has yet to be determined. It is well established that the activation of glutamate receptor and its downstream actions are critical for the retinohypothalamic inputs of light to the SCN [49]. As receptor for PK2 is highly expressed in the SCN [28] and activation of the PK2 receptor triggers similar signalling pathways as that of glutamate receptors [29], it is possible that the circadian clock and/or light-driven PK2 may feed back to the core circadian loops in the SCN. In addition, PK2 has recently been shown to excite neurons that express PK2 receptor [50], further suggesting that PK2 may activate the firing of SCN neurons, and thus possibly participate in the synchronization of the circadian clock. Thus, the light inducibility of PK2 may be relevant to both the phase resetting of the core circadian loops and critical SCN output signals.
Conclusion
Our studies demonstrate that PK2 is predominantly driven by the circadian clock, as PK2 expression exhibits circadian transients in response to phase advances. Furthermore, shortening of the light period does not result in corresponding reduction of the phase of PK2 rhythm, also consistent with the dominant control from the circadian clock on PK2 expression. However, light also modulates PK2 rhythm. Nocturnal light pulses can directly induce PK2 expression in the SCN. Studies with Cry1-/-Cry2-/- mice revealed that light can drive a low amplitude PK2 molecular rhythm in the SCN in the absence of functional circadian oscillators. These studies demonstrate that PK2 molecular rhythm in the SCN is controlled by dual mechanisms: dominantly by the circadian transcriptional loops but also directly by light. The light inducibility of PK2 in the SCN suggest that in addition to PK2's role as a SCN output signal, PK2 may also participate in the photic entrainment of circadian clock and perhaps in masking.
Methods
Experiments of light/dark cycle shifts
Male adult C57BL/6 mice (Taconic Farms, New York) were entrained under 12 hour light: 12 hour dark (12L:12D, lights on at 0700 h) cycle for two weeks with food and water available ad libitum. Light phase was either delayed by 6 hours (lights on at 1300 h) or advanced by 6 hours (lights on at 0100 h) and samples were taken every three hours for the 24 hour period (Zeitgeber time, ZT, ZT1-22). To examine PK2 expression two days after the shift, animals were placed in two additional light/dark cycles and brain samples were collected. All animal procedures were approved by the Institutional Animal Care and Use Committee and consistent with Federal guidelines. In situ hybridization was used in all studies to examine PK2 mRNA expression in the SCN [28]. Antisense and sense riboprobes containing the coding region of mouse PK2 (accession number AF487280 1-528 nt), mouse Per1 (accession number AF022992 340-761nt), mouse Per2 (accession number AF035830 9-489 nt) and mouse Bmal1 (accession number AB015203 864-1362 nt) were generated.
Photoperiod studies
Animals were initially entrained under 12L:12D for one week, followed by placement in different photoperiods (light intensity ~400 lux) for three to four weeks: 8 hour light:16 hour dark (8L:16D, lights on at 0900 h, lights off at 1700 h), 16 hour light: 8 hour dark (16L:8D, lights on at 0500 h, lights off at 2100 h). For the 20 hour light: 4 hour dark (20L:4D, lights on at 0300 h, lights off at 2300 h), animals were first placed in 14L:10D for one week, transferred to 16L:8D for another week, followed by two weeks in 20L:4D. All brain samples were taken every two hours throughout the 24 hour cycle, except the first and the last two time points which were sampled every three hours.
Studies of melanopsin-deficient mice and mice that lack melanopsin, rods and cones
Wild type and melanopsin-deficient (Opn4-/-) mice (on C57BL/6:129 hybrid background) [5] were entrained to 12L:12D and sampled every three hours for the 24 hour period (ZT1-22). For light pulse studies, wild type, Opn4-/- mice and triple knockouts (Opn4-/- Gnat1-/- Cnga3-/- mice) that lack melanopsin, rod and cone phototransduction systems were used [10]. Animals received a 15 min light pulse (~200 lux) at ZT14 and brains were sampled one or two hours after light pulse. Dark control animals did not receive a light pulse.
Studies of cryptochrome-deficient (Cry1-/-Cry2-/-) mice
Cryptochrome-deficient (Cry1-/-Cry2-/-) mice on a C57BL/6:129 hybrid background were kindly provided by Dr. Aziz Sancar (University of North Carolina at Chapel Hill). Wild type and Cry1-/-Cry2-/- mice were entrained to 12L:12D and sampled every three hours for the 24 hour period (ZT1-22). A second group of Cry1-/-Cry2-/- mice were placed into two days of constant darkness (2DD) (Circadian time, CT, CT1-22). The mRNA levels of PK2, Per2, Per1 and Bmal1 were measured in the SCN. For light pulse experiments, Cry1-/-Cry2-/- mice received a 15 min light pulse (~400 lux) at ZT14, and sampled one or two hours after light pulse. Dark control Cry1-/-Cry2-/- mice did not receive a light pulse. For the shifting experiments, wildtype and Cry1-/-Cry2-/- mice were initially entrained under 12L:12D, then subjected to an acute 6 hour advance of lighting schedule. Running-wheel activities of these mice were monitored 10 days before and 10 days after the 6 hour advance shift. The 6 hour phase advance was then repeated and brains were collected at ZT4 and ZT16 on the day of the shift.
Authors' contributions
ELB sampled the Cry1-/-Cry2-/- mice and performed behavior experiments on Cry1-/-Cry2-/- mice. SH sampled the melanopsin-deficient mice and triple knockout mice. MYC performed the tissue sectioning, in situ hybridizations and all quantitative analyses. MYC, ELB and QYZ drafted the manuscript. ELB, SH, MYC and QYZ designed the studies. All authors read and approved the final manuscript.
Supplementary Material
Effect of different photoperiods on molecular rhythms in the SCN. Temporal profiles of Per1 (a) and Per2 (b) mRNA under 8L:16D, 16L:8D, 20L:4D. Open and filled bars indicate light and dark periods, respectively. The zeitgeber time (ZT) on the x-axis reflects the timescale for each photoperiod. Each value represents the mean ± SEM of 3–4 animals.
PK2 mRNA expression in Cry1-/-Cry2-/- and wildtype mice. Representative autoradiograms of PK2 mRNA in the SCN of Cry1-/-Cry2-/- mice (Cry) and wild type mice (WT) under LD (ZT1-22) are shown (top and bottom row, respectively). Scale bar = 1 mm.
Acknowledgments
Acknowledgements
We thank David R. Weaver and King-Wai Yau for providing helpful discussions, in situ probes and critical comments on the manuscript. We would also like to thank Aziz Sancar for providing access to Cry1-/-Cry2-/- mice, Frances Leslie for helpful discussions and equipment use, James Belluzzi for help with statistical analysis, Alex Lee for mice running-wheel setup and data analysis, Clayton Bullock and Jia-da Li for technical assistance and discussions. The research is partially supported by NIH grants (to QYZ and ELB). MYC was supported in part by a NIDA training grant and by a predoctoral fellowship from PhRMA Foundation.
Contributor Information
Michelle Y Cheng, Email: mycheng@uci.edu.
Eric L Bittman, Email: elb@bio.umass.edu.
Samer Hattar, Email: shattar@jhu.edu.
Qun-Yong Zhou, Email: qzhou@uci.edu.
References
- Reppert SM, Weaver DR. Molecular analysis of mammalian circadian rhythms. Annu Rev Physiol. 2001;63:647–676. doi: 10.1146/annurev.physiol.63.1.647. [DOI] [PubMed] [Google Scholar]
- Reppert SM, Weaver DR. Coordination of circadian timing in mammals. Nature. 2002;418:935–941. doi: 10.1038/nature00965. [DOI] [PubMed] [Google Scholar]
- Moore RY. Entrainment pathways and the functional organization of the circadian system. Prog Brain Res. 1996;111:103–119. doi: 10.1016/s0079-6123(08)60403-3. [DOI] [PubMed] [Google Scholar]
- Ebling FJ. The role of glutamate in the photic regulation of the suprachiasmatic nucleus. Prog Neurobiol. 1996;50:109–132. doi: 10.1016/S0301-0082(96)00032-9. [DOI] [PubMed] [Google Scholar]
- Hattar S, Liao HW, Takao M, Berson DM, Yau KW. Melanopsin-containing retinal ganglion cells: architecture, projections, and intrinsic photosensitivity. Science. 2002;295:1065–1070. doi: 10.1126/science.1069609. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Berson DM, Dunn FA, Takao M. Phototransduction by retinal ganglion cells that set the circadian clock. Science. 2002;295:1070–1073. doi: 10.1126/science.1067262. [DOI] [PubMed] [Google Scholar]
- Panda S, Sato TK, Castrucci AM, Rollag MD, DeGrip WJ, Hogenesch JB, Provencio I, Kay SA. Melanopsin (Opn4) requirement for normal light-induced circadian phase shifting. Science. 2002;298:2213–2216. doi: 10.1126/science.1076848. [DOI] [PubMed] [Google Scholar]
- Ruby NF, Brennan TJ, Xie X, Cao V, Franken P, Heller HC, O'Hara BF. Role of melanopsin in circadian responses to light. Science. 2002;298:2211–2213. doi: 10.1126/science.1076701. [DOI] [PubMed] [Google Scholar]
- Lucas RJ, Hattar S, Takao M, Berson DM, Foster RG, Yau KW. Diminished pupillary light reflex at high irradiances in melanopsin-knockout mice. Science. 2003;299:245–247. doi: 10.1126/science.1077293. [DOI] [PubMed] [Google Scholar]
- Hattar S, Lucas RJ, Mrosovsky N, Thompson S, Douglas RH, Hankins MW, Lem J, Biel M, Hofmann F, Foster RG, Yau KW. Melanopsin and rod-cone photoreceptive systems account for all major accessory visual functions in mice. Nature. 2003;424:76–81. doi: 10.1038/nature01761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Panda S, Provencio I, Tu DC, Pires SS, Rollag MD, Castrucci AM, Pletcher MT, Sato TK, Wiltshire T, Andahazy M, Kay SA, Van Gelder RN, Hogenesch JB. Melanopsin is required for non-image-forming photic responses in blind mice. Science. 2003;301:525–527. doi: 10.1126/science.1086179. [DOI] [PubMed] [Google Scholar]
- Dunlap JC. Genetics and molecular analysis of circadian rhythms. Annu Rev Genet. 1996;30:579–601. doi: 10.1146/annurev.genet.30.1.579. [DOI] [PubMed] [Google Scholar]
- Lowrey PL, Takahashi JS. Genetics of the mammalian circadian system: Photic entrainment, circadian pacemaker mechanisms, and posttranslational regulation. Annu Rev Genet. 2000;34:533–562. doi: 10.1146/annurev.genet.34.1.533. [DOI] [PubMed] [Google Scholar]
- Hastings M, Maywood ES. Circadian clocks in the mammalian brain. Bioessays. 2000;22:23–31. doi: 10.1002/(SICI)1521-1878(200001)22:1<23::AID-BIES6>3.0.CO;2-Z. [DOI] [PubMed] [Google Scholar]
- Cermakian N, Sassone-Corsi P. Environmental stimulus perception and control of circadian clocks. Curr Opin Neurobiol. 2002;12:359–365. doi: 10.1016/S0959-4388(02)00347-1. [DOI] [PubMed] [Google Scholar]
- Rusak B, Robertson HA, Wisden W, Hunt SP. Light pulses that shift rhythms induce gene expression in the suprachiasmatic nucleus. Science. 1990;248:1237–1240. doi: 10.1126/science.2112267. [DOI] [PubMed] [Google Scholar]
- Kornhauser JM, Nelson DE, Mayo KE, Takahashi JS. Photic and circadian regulation of c-fos gene expression in the hamster suprachiasmatic nucleus. Neuron. 1990;5:127–134. doi: 10.1016/0896-6273(90)90303-W. [DOI] [PubMed] [Google Scholar]
- Albrecht U, Sun ZS, Eichele G, Lee CC. A differential response of two putative mammalian circadian regulators, mper1 and mper2, to light. Cell. 1997;91:1055–1064. doi: 10.1016/S0092-8674(00)80495-X. [DOI] [PubMed] [Google Scholar]
- Shigeyoshi Y, Taguchi K, Yamamoto S, Takekida S, Yan L, Tei H, Moriya T, Shibata S, Loros JJ, Dunlap JC, Okamura H. Light-induced resetting of a mammalian circadian clock is associated with rapid induction of the mPer1 transcript. Cell. 1997;91:1043–1053. doi: 10.1016/S0092-8674(00)80494-8. [DOI] [PubMed] [Google Scholar]
- Harmar AJ, Marston HM, Shen S, Spratt C, West KM, Sheward WJ, Morrison CF, Dorin JR, Piggins HD, Reubi JC, Kelly JS, Maywood ES, Hastings MH. The VPAC(2) receptor is essential for circadian function in the mouse suprachiasmatic nuclei. Cell. 2002;109:497–508. doi: 10.1016/S0092-8674(02)00736-5. [DOI] [PubMed] [Google Scholar]
- Mrosovsky N. Masking: history, definitions, and measurement. Chronobiol Int. 1999;16:415–429. doi: 10.3109/07420529908998717. [DOI] [PubMed] [Google Scholar]
- Redlin U. Neural basis and biological function of masking by light in mammals: suppression of melatonin and locomotor activity. Chronobiol Int. 2001;18:737–758. doi: 10.1081/CBI-100107511. [DOI] [PubMed] [Google Scholar]
- Redlin U, Mrosovsky N. Masking by light in hamsters with SCN lesions. J Comp Physiol [A] 1999;184:439–448. doi: 10.1007/s003590050343. [DOI] [PubMed] [Google Scholar]
- van der Horst GT, Muijtjens M, Kobayashi K, Takano R, Kanno S, Takao M, de Wit J, Verkerk A, Eker AP, van Leenen D, Buijs R, Bootsma D, Hoeijmakers JH, Yasui A. Mammalian Cry1 and Cry2 are essential for maintenance of circadian rhythms. Nature. 1999;398:627–630. doi: 10.1038/19323. [DOI] [PubMed] [Google Scholar]
- Vitaterna MH, Selby CP, Todo T, Niwa H, Thompson C, Fruechte EM, Hitomi K, Thresher RJ, Ishikawa T, Miyazaki J, Takahashi JS, Sancar A. Differential regulation of mammalian period genes and circadian rhythmicity by cryptochromes 1 and 2. Proc Natl Acad Sci U S A. 1999;96:12114–12119. doi: 10.1073/pnas.96.21.12114. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Bae K, Jin X, Maywood ES, Hastings MH, Reppert SM, Weaver DR. Differential functions of mPer1, mPer2, and mPer3 in the SCN circadian clock. Neuron. 2001;30:525–536. doi: 10.1016/S0896-6273(01)00302-6. [DOI] [PubMed] [Google Scholar]
- Bunger MK, Wilsbacher LD, Moran SM, Clendenin C, Radcliffe LA, Hogenesch JB, Simon MC, Takahashi JS, Bradfield CA. Mop3 is an essential component of the master circadian pacemaker in mammals. Cell. 2000;103:1009–1017. doi: 10.1016/S0092-8674(00)00205-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Cheng MY, Bullock CM, Li C, Lee AG, Bermak JC, Belluzzi J, Weaver DR, Leslie FM, Zhou QY. Prokineticin 2 transmits the behavioural circadian rhythm of the suprachiasmatic nucleus. Nature. 2002;417:405–410. doi: 10.1038/417405a. [DOI] [PubMed] [Google Scholar]
- Lin DC, Bullock CM, Ehlert FJ, Chen JL, Tian H, Zhou QY. Identification and molecular characterization of two closely related G protein-coupled receptors activated by prokineticins/endocrine gland vascular endothelial growth factor. J Biol Chem. 2002;277:19276–19280. doi: 10.1074/jbc.M202139200. [DOI] [PubMed] [Google Scholar]
- Reddy AB, Field MD, Maywood ES, Hastings MH. Differential resynchronisation of circadian clock gene expression within the suprachiasmatic nuclei of mice subjected to experimental jet lag. J Neurosci. 2002;22:7326–7330. doi: 10.1523/JNEUROSCI.22-17-07326.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Yamazaki S, Numano R, Abe M, Hida A, Takahashi R, Ueda M, Block GD, Sakaki Y, Menaker M, Tei H. Resetting central and peripheral circadian oscillators in transgenic rats. Science. 2000;288:682–685. doi: 10.1126/science.288.5466.682. [DOI] [PubMed] [Google Scholar]
- Daan S, Pittendrigh C. A functional analysis of circadian pacemakers in nocturnal rodents. IV. Entrainment: pacemaker as clock. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 1976;106:291–331. [Google Scholar]
- Messager S, Ross AW, Barrett P, Morgan PJ. Decoding photoperiodic time through Per1 and ICER gene amplitude. Proc Natl Acad Sci U S A. 1999;96:9938–9943. doi: 10.1073/pnas.96.17.9938. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Albrecht U, Zheng B, Larkin D, Sun ZS, Lee CC. MPer1 and mper2 are essential for normal resetting of the circadian clock. J Biol Rhythms. 2001;16:100–104. doi: 10.1177/074873001129001791. [DOI] [PubMed] [Google Scholar]
- Steinlechner S, Jacobmeier B, Scherbarth F, Dernbach H, Kruse F, Albrecht U. Robust circadian rhythmicity of Per1 and Per2 mutant mice in constant light, and dynamics of Per1 and Per2 gene expression under long and short photoperiods. J Biol Rhythms. 2002;17:202–209. doi: 10.1177/07430402017003003. [DOI] [PubMed] [Google Scholar]
- Mrosovsky N, Hattar S. Impaired masking responses to light in melanopsin-knockout mice. Chronobiol Int. 2003;20:989–999. doi: 10.1081/CBI-120026043. [DOI] [PubMed] [Google Scholar]
- Okamura H, Miyake S, Sumi Y, Yamaguchi S, Yasui A, Muijtjens M, Hoeijmakers JH, van der Horst GT. Photic induction of mPer1 and mPer2 in cry-deficient mice lacking a biological clock. Science. 1999;286:2531–2534. doi: 10.1126/science.286.5449.2531. [DOI] [PubMed] [Google Scholar]
- Shearman LP, Zylka MJ, Weaver DR, Kolakowski LF, Jr, Reppert SM. Two period homologs: circadian expression and photic regulation in the suprachiasmatic nuclei. Neuron. 1997;19:1261–1269. doi: 10.1016/S0896-6273(00)80417-1. [DOI] [PubMed] [Google Scholar]
- Gekakis N, Staknis D, Nguyen HB, Davis FC, Wilsbacher LD, King DP, Takahashi JS, Weitz CJ. Role of the CLOCK protein in the mammalian circadian mechanism. Science. 1998;280:1564–1569. doi: 10.1126/science.280.5369.1564. [DOI] [PubMed] [Google Scholar]
- Hogenesch JB, Gu YZ, Jain S, Bradfield CA. The basic-helix-loop-helix-PAS orphan MOP3 forms transcriptionally active complexes with circadian and hypoxia factors. Proc Natl Acad Sci U S A. 1998;95:5474–5479. doi: 10.1073/pnas.95.10.5474. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Preitner N, Damiola F, Lopez-Molina L, Zakany J, Duboule D, Albrecht U, Schibler U. The orphan nuclear receptor REV-ERBalpha controls circadian transcription within the positive limb of the mammalian circadian oscillator. Cell. 2002;110:251–260. doi: 10.1016/S0092-8674(02)00825-5. [DOI] [PubMed] [Google Scholar]
- Ueda HR, Chen W, Adachi A, Wakamatsu H, Hayashi S, Takasugi T, Nagano M, Nakahama K, Suzuki Y, Sugano S, Iino M, Shigeyoshi Y, Hashimoto S. A transcription factor response element for gene expression during circadian night. Nature. 2002;418:534–539. doi: 10.1038/nature00906. [DOI] [PubMed] [Google Scholar]
- Stephan FK, Zucker I. Circadian rhythms in drinking behavior and locomotor activity of rats are eliminated by hypothalamic lesions. Proc Natl Acad Sci U S A. 1972;69:1583–1586. doi: 10.1073/pnas.69.6.1583. [DOI] [PMC free article] [PubMed] [Google Scholar]
- Li X, Gilbert J, Davis FC. Disruption of masking by hypothalamic lesions in Syrian hamsters. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2004. [DOI] [PubMed]
- Gau D, Lemberger T, von Gall C, Kretz O, Le Minh N, Gass P, Schmid W, Schibler U, Korf HW, Schutz G. Phosphorylation of CREB Ser142 regulates light-induced phase shifts of the circadian clock. Neuron. 2002;34:245–253. doi: 10.1016/S0896-6273(02)00656-6. [DOI] [PubMed] [Google Scholar]
- Obrietan K, Impey S, Smith D, Athos J, Storm DR. Circadian regulation of cAMP response element-mediated gene expression in the suprachiasmatic nuclei. J Biol Chem. 1999;274:17748–17756. doi: 10.1074/jbc.274.25.17748. [DOI] [PubMed] [Google Scholar]
- Tischkau SA, Mitchell JW, Tyan SH, Buchanan GF, Gillette MU. Ca2+/cAMP response element-binding protein (CREB)-dependent activation of Per1 is required for light-induced signaling in the suprachiasmatic nucleus circadian clock. J Biol Chem. 2003;278:718–723. doi: 10.1074/jbc.M209241200. [DOI] [PubMed] [Google Scholar]
- Yokota S, Yamamoto M, Moriya T, Akiyama M, Fukunaga K, Miyamoto E, Shibata S. Involvement of calcium-calmodulin protein kinase but not mitogen-activated protein kinase in light-induced phase delays and Per gene expression in the suprachiasmatic nucleus of the hamster. J Neurochem. 2001;77:618–627. doi: 10.1046/j.1471-4159.2001.00270.x. [DOI] [PubMed] [Google Scholar]
- Ding JM, Chen D, Weber ET, Faiman LE, Rea MA, Gillette MU. Resetting the biological clock: mediation of nocturnal circadian shifts by glutamate and NO. Science. 1994;266:1713–1717. doi: 10.1126/science.7527589. [DOI] [PubMed] [Google Scholar]
- Cottrell GT, Zhou QY, Ferguson AV. Prokineticin 2 modulates the excitability of subfornical organ neurons. J Neurosci. 2004;24:2375–2379. doi: 10.1523/JNEUROSCI.5187-03.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Effect of different photoperiods on molecular rhythms in the SCN. Temporal profiles of Per1 (a) and Per2 (b) mRNA under 8L:16D, 16L:8D, 20L:4D. Open and filled bars indicate light and dark periods, respectively. The zeitgeber time (ZT) on the x-axis reflects the timescale for each photoperiod. Each value represents the mean ± SEM of 3–4 animals.
PK2 mRNA expression in Cry1-/-Cry2-/- and wildtype mice. Representative autoradiograms of PK2 mRNA in the SCN of Cry1-/-Cry2-/- mice (Cry) and wild type mice (WT) under LD (ZT1-22) are shown (top and bottom row, respectively). Scale bar = 1 mm.