

Abstract
Background
As a superfamily of transcription factors (TFs), the basic helix-loop-helix (bHLH) proteins have been characterized functionally in many plants with a vital role in the regulation of diverse biological processes including growth, development, response to various stresses, and so on. However, no systemic analysis of the bHLH TFs has been reported in Brachypodium distachyon, an emerging model plant in Poaceae.
Results
A total of 146 bHLH TFs were identified in the Brachypodium distachyon genome and classified into 24 subfamilies. BdbHLHs in the same subfamily share similar protein motifs and gene structures. Gene duplication events showed a close relationship to rice, maize and sorghum, and segment duplications might play a key role in the expansion of this gene family. The amino acid sequence of the bHLH domains were quite conservative, especially Leu-27 and Leu-54. Based on the predicted binding activities, the BdbHLHs were divided into DNA binding and non-DNA binding types. According to the gene ontology (GO) analysis, BdbHLHs were speculated to function in homodimer or heterodimer manner. By integrating the available high throughput data in public database and results of quantitative RT-PCR, we found the expression profiles of BdbHLHs were different, implying their differentiated functions.
Conclusion
One hundred fourty-six BdbHLHs were identified and their conserved domains, sequence features, phylogenetic relationship, chromosomal distribution, GO annotations, gene structures, gene duplication and expression profiles were investigated. Our findings lay a foundation for further evolutionary and functional elucidation of BdbHLH genes.
Electronic supplementary material
The online version of this article (doi:10.1186/s12864-017-4044-4) contains supplementary material, which is available to authorized users.
Keywords: Genome-wide, bHLH, Brachypodium distachyon, Expression profile, Transcription factor
Background
Grasses (Poaceae), such as rice, maize, wheat, provide the bulk of nutrition and sustainable energy [1, 2]. Crop growth, development and productivity are continuously threatened by various adverse environmental factors including biotic and abiotic stresses for their sessile nature. They have evolved complicated physiological and biochemical responses by regulating the expression of a series of genes to survive and flourish under extreme living conditions.
Transcription factors (TFs) play key roles in the stress-related regulation network and signal pathways. Among them, basic helix-loop-helixes (bHLH) TFs constitute a large superfamily that has been identified in all eukaryotes including metazoans, plants, and fungi [3–5]. As the second largest class of plant TFs [6], bHLH was characterized with one specific bHLH domain, including a basic region and an HLH region [7]. The basic region, located at the N-terminus of the domain, consisting of approximately 17 amino acids, is a DNA-binding region that enables bHLH TFs to bind to E-box (CANNTG) [7, 8]; the HLH region includes two amphipathic α helices separated by a variable (both in length and primary sequence) loop and participates in the formation of homodimers or heterodimers [8, 9].
In metazoans, the bHLH TFs were divided into six groups (group A to F) based on their phylogenetic relationships, major functions and DNA-binding ability [8, 10–12]. Phylogenic tree of plant bHLHs was first constructed in Arabidopsis and AtbHLHs were divided into 12 subfamilies [13]. In rice, the phylogenic tree of bHLH TFs were divided into 22 subfamilies [14]. With more sequenced plant genomes, lots of bHLH proteins were identified. For example, genomes of Nicotiana tabacum, Daucus carota, Salvia miltiorrhiza, and Solanum lycopersicum contain 190, 146, 127, 159 bHLH genes respectively [15–18]. Researches in Arabidopsis revealed that bHLH TFs have versatile biological functions, such as regulating the seed germination [19], the development of epidermal cell [20], carpel [21] and anther [22], fruit dehiscence [23], responding to phytochrome [24] and phytohormone signal [25], stresses [26], etc.
Brachypodium distachyon, as the first sequenced species in Pooideae subfamily, has been proposed as a new model organism for functional genomics studies, due to the facility of cultivation and mature transformation system, short life cycle, small genome size and close relation to several cereals [1, 27, 28]. In this study, we identified 146 BdbHLH genes and conducted a genome-wide bioinformatics analysis based on the phylogenetic relationships. Meanwhile, the cis-elements in the promoter region, gene structure, conserved motifs, as well as chromosomal distribution, gene duplication and evolutionary mechanisms were investigated. Furthermore, the expression profiles of the BdbHLHs were investigated based on the published RNA-seq, microarray data and qRT-PCR. These results provide clues for functional elucidation of BdbHLHs.
Methods
Genome-wide identification, sequence alignment and phylogenetic analyses of BdbHLHs
Previous studies indicated that the bHLH domain in Arabidopsis contained 19 conserved amino acid residues distributed in the basic region (5), the first helix (5), the loop (1) and the second helix (8) (Additional file 1: Table S1) [29]. Among them, nine mismatches were allowed for the identification of bHLHs [7]. To identify candidate bHLH genes in Brachypodium distachyon, a BLAST of the bHLH domain was conducted based on the conserved bHLH motif in Arabidopsis and rice from the National Center of Biotechnology Information database (http://www.ncbi.nlm.nih.gov) and the Gramene database (http://www.gramene.org/). SMART [30] was applied to verify the candidate bHLH TFs. Proteins with less conserved bHLH domains or no bHLH domains were removed. The biochemical properties were predicted by ExPASy [31]. The GO (gene ontology) annotations of BdbHLHs were obtained from Gramene and Plant Transcriptional Regulatory Map [32], then were visualized by BGI WEGO website [33].
To investigate the phylogenetic relationship between bHLH proteins, protein sequence alignment was performed with default parameters and an un-rooted phylogenetic tree was constructed by MEGA (vision 6.0) [34] based on the neighbor joining (NJ) method with 1000 bootstrap replications and visualized by the EvolView [35].
Analysis of chromosomal distribution, gene duplication and synteny
The chromosomal distribution of BdbHLH genes was obtained from the Brachypodium distachyon genome annotations. Tandem duplication events were characterized as contiguous homologous genes on a single chromosome without any intervening gene and checked manually [36]. To analyze the collinear correlations between bHLHs in Brachypodium distachyon and rice, maize, sorghum, synteny blocks were downloaded from the Plant Genome Duplication Database [37]. The chromosomal distribution of BdbHLHs and the synteny relationships of related genes across the four species were visualized using Circos (vision 0.69) [38].
Analyses of promoters, gene structure, conserved motifs, and construction of the interaction network
The upstream 1500 bp genomic DNA sequences of BdbHLH genes were downloaded and submitted to the PlantCARE [39] to predict the putative cis-elements. The intron-exon organizations of BdbHLHs were displayed by the Gene Structure Display Server [40]. Conserved motifs of BdbHLHs were identified by MEME server [41] with maximum number of motifs set at 15 and optimum width of motifs from 5 to 200 amino acids. The interaction network was constructed based on homologs of BdbHLHs in Arabidopsis using the AraNet V2 tool [42] and visualized by Cytoscape (version 3.4.0) [43].
Analyzing the expression profiles of BdbHLHs
To analyze the expression profiles of BdbHLHs in different tissues and under phytohormone stresses, microarray data (SRP008505) [44] and high throughout RNA sequencing data (PRJDB2997) [45] were retrieved from EBI ArrayExpress (https://www.ebi.ac.uk/gxa/home) and DDBJ Sequence Read Archive (http://www.ddbj.nig.ac.jp/index-e.html) respectively, and then visualized by the MeV (version 4.9.0) [46].
For qRT-PCR, 2-week-old seedlings of Bd21 were used for different treatments. For salt, drought and plant hormone treatments, seedlings were treated in MS liquid medium containing 200 mM NaCl, 20% PEG6000 (to mimic drought stress), 100 μM MeJA, 100 μM ABA, 20 μM 6-BA and 1 mM SA for 2 h respectively and the roots were collected. For heat and cold treatments, seedlings were subjected to 45 °C and 4 °C respectively and the leaves were collected. Plants during heading stage were used for the collection of roots, stems, leaves and inflorescences. All materials were frozen in liquid nitrogen and stored at −80 °C for RNA isolation. RNA extraction, cDNA synthesis and qRT-PCR reaction were carried out as described previously [47]. The qRT-PCR reaction was performed in triplicate and data acquisition and analyses were performed using the QuantStudio™ Real-Time PCR Software (ThermoFisher Scientific). Samples were normalized using UBC18 (BRADI4G00660) expression [48] and relative expression levels were determined using the 2(−ΔΔCt) analysis method [49]. The primers used were listed in Additional file 1: Table S2.
Results and discussion
Identification, chromosomal distribution and physicochemical properties of BdbHLHs
With the criterion above, we searched proteins consisting of the conserved bHLH domain in the whole genome of Brachypodium distachyon. 146 BdbHLH proteins were identified. The ratio of bHLH genes in Brachypodium distachyon genome was about 0.55%, which is similar to Arabidopsis (0.59%) [29] and is more than rice (0.44%) [14] and poplar (0.40%) [4]. In order to verify the reliability of our identification, a BlastN program was used to search for all the expressed sequence tags (EST) in Brachypodium distachyon (Additional file 1: Table S3). 57.5% (84/146) of BdbHLHs were supported by the EST hits.
According to their physical positions (Additional file 1: Table S3), the 146 BdbHLH genes were mapped on five chromosomes (Fig. 1a): 47 (32.2%) on chromosome 1, 28 (19.2%) on chromosome 2, 39 (26.7%) on chromosome 3, 21 (14.4%) on chromosome 4, and 11 (7.5%) on chromosome 5. Similar to rice [14], tomato [18, 50] and the common bean [51], most BdbHLHs were found to be located at the both ends of chromosomes 1, 2, 3, 4 and the bottom of chromosome 5.
Fig. 1.
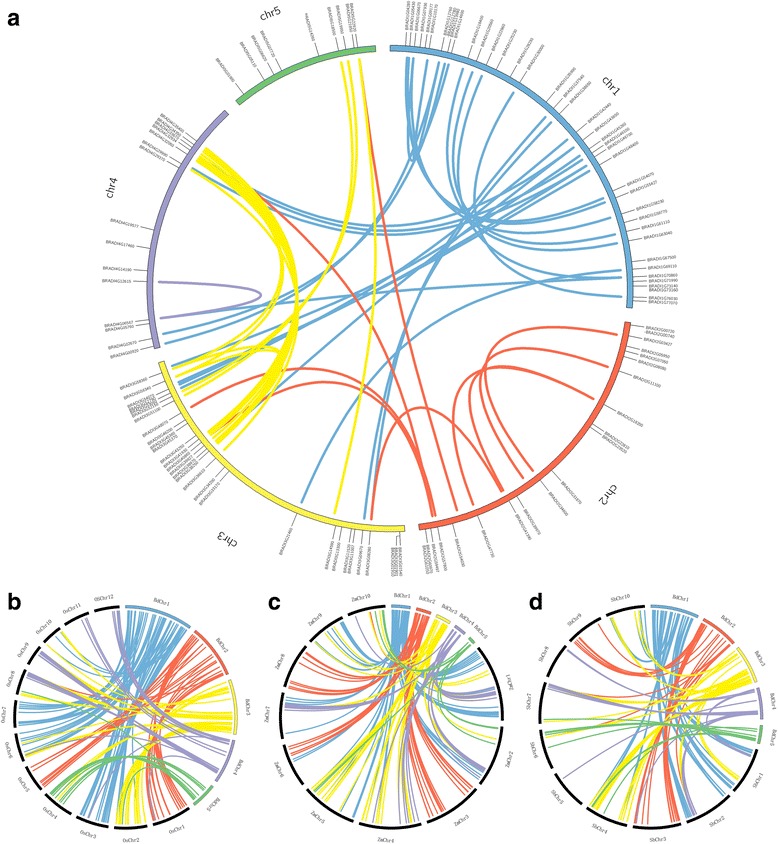
Genomic distribution of bHLH genes and the gene duplications in the Brachypodium distachyon (a), rice (b), maize (c), and sorghum (d) genome
To further characterize the BdbHLHs, we analyzed the physicochemical properties of the putative proteins (Additional file 1: Table S3). The Grand average of hydropathicity (GRAVY) of all the candidate BdbHLH proteins was predicted to be negative value ranging from −0.026 to −1.003, representing a hydrophilic characteristic. These proteins showed diversities in the length, molecular weight, theoretical isoelectric points (PI), number of negatively charged residues (Asp and Glu), number of positively charged residues (Arg and Lys).
Gene duplication and collinear correlations of bHLHs between Brachypodium distachyon and rice, maize and sorghum genomes
Among the 146 BdbHLHs, about two thirds were duplicated genes. A total of 19 (13.0%) BdbHLHs have been identified as tandem duplicated genes and distributed on chromosome 1, 2 and 3 (Fig. 1a, Additional file 1: Table S4), while BRADI1G12760 contains an incomplete bHLH domain and might lose the function during duplication [52]. Most of them were derived from the same subfamily with original genes (except for the pair BRADI3G52790 and BRADI3G53060 in chromosome 1). By contrast, 75 (58.2%) segmentally duplicated BdbHLHs were detected (Fig. 1a, Additional file 1: Table S5)
The substitution rates of non-synonymous versus synonymous (Ka/Ks) is an effective criterion to judge the selection pressure after gene duplications [53]. Thus, the Ka/Ks of duplicated BdbHLHs was calculated (Additional file 1: Tables S4 and S5). For most tandem duplicated gene pairs, the Ka/Ks value was less than 1, indicating a purifying selection during expansion, except for BRADI2G00730/BRADI2G00740, BRADI3G41940/BRADI3G41950 and BRADI3G52790/BRADI3G52790, the Ka/Ks ratio was 1.99, 1.63 and 1.15 respectively. This means accelerated evolution is accompanied with positive selection. For the segment duplicated gene pairs, all the Ka/Ks was less than 1 (ranging from 0.14 to 0.82) and the average is 0.46 (Additional file 1: Table S5), suggesting an intense purifying selection pressure during evolution. Meanwhile, the divergence time of the segment duplication event was predicted to take place around 76 Mya, which was much earlier than the tandem duplication (~42 Mya).
To further investigate the origin and evolutionary relationships of bHLH genes, comparative syntenic analyses at genome-wide level between Brachypodium distachyon and other grass species were conducted. Most BdbHLHs have orthologous in rice, maize and sorghum (80.8%, 69.9% and 72.6%, respectively) (Fig. 1b–d, Additional file 1: Tables S6–S8). The divergence time in rice, maize and sorghum was about 52 Mya, 56 Mya, and 59 Mya, respectively. The Ka/Ks ratio between Brachypodium distachyon and rice, maize, sorghum was 0.41, 0.41 and 0.32, implying these bHLH gene pairs have gone through strong purifying selection and there was an intimate correlation between Brachypodium distachyon and rice, maize, sorghum. In brief, gene duplication events including tandem duplication and segment duplication seemed likely to be essential for bHLH gene family expansion and functional conservation and divergence in the Poaceae.
Multiple sequence alignment, prediction of protein dimerization activity and DNA binding activity of BdbHLHs
As shown in Fig. 2 and Additional file 1: Table S1, 24 conserved amino acids were found in the bHLH domains (conservation more than 50%). Among them, Ile-20, Asn-21, Leu-24, Gln-28, Lys-36, Asp-38, Ile-43, Val-51 and Leu-54 were more conservative in plants [4, 54]. Some conserved amino acid residues such as Glu-13, Arg-14, Arg-16 and Leu-27 were not only detected in Brachypodium distachyon, but also in Arabidopsis and rice, suggesting they are essential to the biological function of bHLH proteins [3].
Fig. 2.
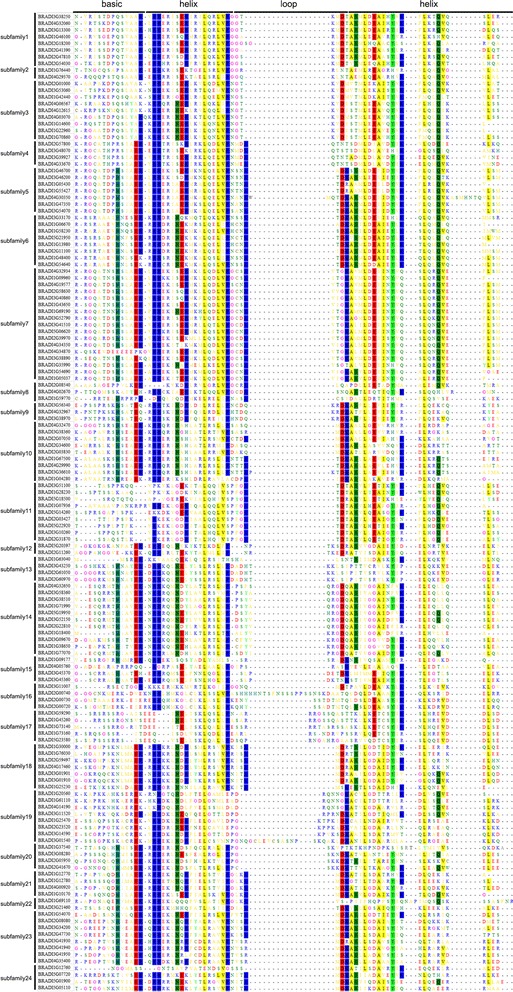
Multiple Sequence Alignment of the bHLH Domains. The amino acids with identity more than 50% are labeled with colored boxes
Previous studies indicated that the HLH domain was essential in both dimerization and DNA binding [9]. Especially, Leu-27 in helix 1 and Leu-73 in helix 2 were important for protein interaction [4]. In this study, 145 and 144 BdbHLHs were found to have Leu-27 and Leu-54 (corresponding to Leu-73 in AtbHLHs) respectively (Fig. 2). Recently, MYC2, MYC3 and MYC4 in Arabidopsis were reported to form homodimers through Leu, Ile and Val in the helixes [55]. In Brachypodium distachyon, including three homologues of AtMYC2, AtMYC3 and AtMYC4, the helixes in many BdbHLHs have these three kinds of amino acids simultaneously, implying the probability to form protein complexes (Fig. 2). In combination with GO annotation that all BdbHLHs showed protein dimerization activity (GO: 0046983, Additional file 1: Tables S9, S10 and Additional file 2: Figure S1), we speculated that BdbHLHs might function by forming protein complex and tried to construct the interaction network of BdbHLHs. Because of short of reported experiment data and databases, the interaction network was constructed based on the orthology analysis with AtbHLHs. According to the AraNet V2 [42], 57 BdbHLH proteins have orthologs in Arabidopsis. As a result, 660 interaction protein pairs were predicted (Fig. 3, Additional file 1: Table S11). It has been reported that members of bHLHs and MYB gene families might function cooperatively via physical interaction [6, 56–58], so the interaction network was further analyzed and a total of 16 MYB genes based on PlantTFDB database [59] were sought out (Fig. 3). The interaction network might provide some clues to investigate the molecular mechanism of bHLH.
Fig. 3.
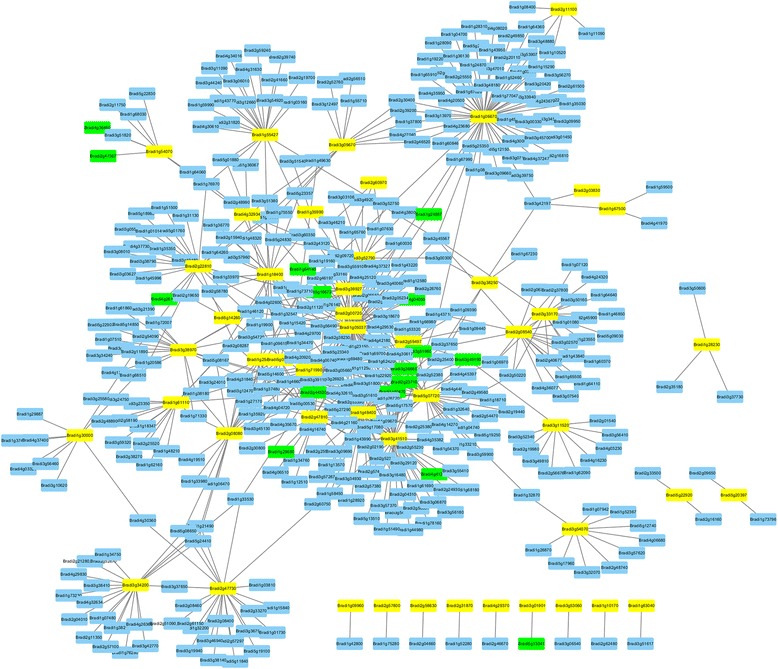
The interaction network of bHLHs in Brachypodium distachyon according to the orthologs in Arabidopsis. The BdbHLHs are in yellow blocks, the MYBs are in green blocks while other genes are in blue blocks
The BdbHLHs were grouped based on the amino acid sequence of the bHLH domain which determines the DNA binding activity [29]. Glu-13 is related to CA in the E-box DNA binding motif (CANNTG) and the substitution of it with other residues (Gln, Asp and Leu) abolishes the DNA binding activity [60–64]. Arg-16 could stabilize the position of Glu-13 and was essential in DNA binding [61, 62]. Based on the existence of Glu-13 and Arg-16 which play a key role in recognition of E-box, the BdbHLH proteins were divided into E-box binding and non-E-box binding (Additional file 1: Table S12) [62, 65]. In addition, His/Lys-9, Glu-13, and Arg-17 are responsible for the specificity to bind to G-box (CACGTG). Especially, Arg-17 directly interacts with the middle G and His-9 interacts with both the middle G and the first C of G-box [62, 63, 65]. So, the E-box-binding proteins were further divided into G-box-binding proteins and non-G-box binding proteins according to the presence of His/Lys-9, Glu-13 and Arg-17 residues or not. For example, it was reported that the G-box binding protein AtMYC2 was crystallized in complex with G-box DNA. Similar to mammalian bHLH TFs, further research showed that three conserved amino acids H453, E457 and R461 (corresponding to our His-9, Glu-13 and Arg-17, respectively, Fig. 2) were essential for the formation of the complex [55]. Meanwhile, the MYC2-DNA structures can further form homo-tetramer with significantly enhanced DNA binding affinity due to the interaction between conserved R458, Q459 and Q466 in one dimer with DNA in the other dimer [55]. In Brachypodium distachyon, three bHLHs, BRADI2G08080, BRADI3G34200 and BRADI2G47730 are highly conservative with AtMYC2 and possess Arg-14, Gln-15 and Gln-22 amino acids (corresponding to R458, Q459 and Q466, respectively), suggesting that they could form tetramers too. The bHLHs without predicting E-box-binding specific recognition residues but possessing additional basic amino acids in the basic region might be able to bind DNA without specificity for E-boxes were classified as non E-box DNA binders [62, 65]. According to the conservation of these residues, 102 BdbHLH proteins were predicted to be putative E-box-binding proteins wherein 78 belong to G-box-binding proteins, 25 as non-E-box-binding proteins for missing Glu-13/Arg-16 residues while 19 BdbHLHs containing less than six amino acid residues in the basic region fell into non-DNA-binding proteins (Fig. 4). Non-DNA-binding proteins, also known as HLH protein, might function like MYB-bHLH-WD40 which can interact with DNA binding proteins as negative regulators [66].
Fig. 4.
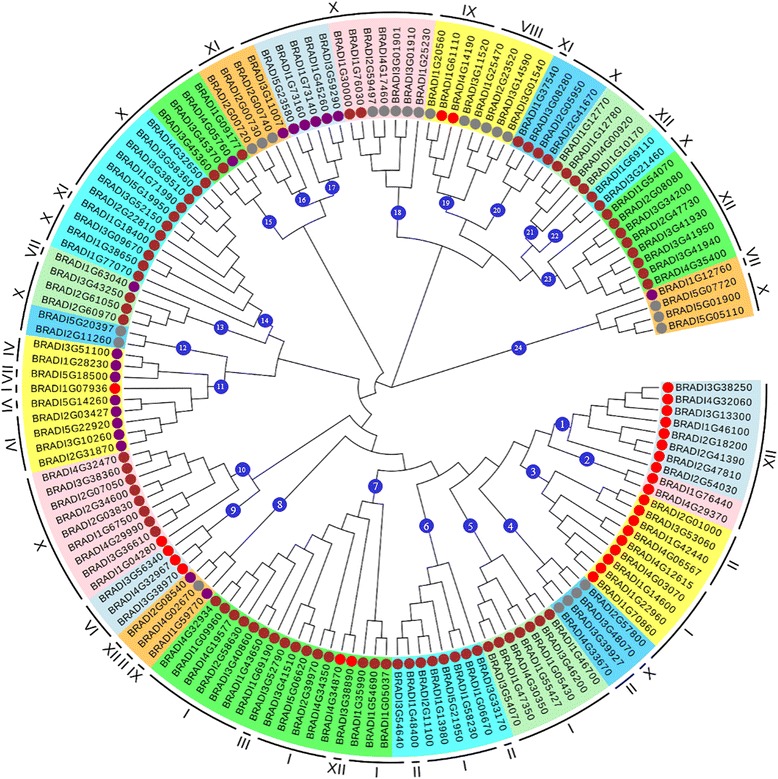
Phylogenetic tree constructed with the BdbHLH domains based on the neighbor-joining method. The tree shows the 24 phylogenetic groups (numbered within blue circles). The circles of different colors represent the predicted DNA-binding activity of each protein: G-box in brown, Non-G-box in gray, Non-E-box in red and No-DNA binding in purple
Gene structures, conserved motifs and functional prediction of BdbHLHs based on phylogenic analyses
A Neighbor Joining phylogenetic tree was constructed based on the bHLH domains of the BdbHLHs (Fig. 4). According to the values obtained in the bootstrap analysis, the bHLH domain in the outer clades had better resolution, permitting subfamilies of proteins to be delimited. The results showed that the BdbHLH amino acid sequences in the same subfamily were highly conserved, implying a strong evolutionary relationship among those members. Based on the statistical support of each branch, we selected those with a bootstrap value >50 to divide the BdbHLH proteins into 24 subfamilies. According to previous phylogenic classifications [4, 13], another phylogenic tree was constructed based on the full length alignments of the 146 BdbHLHs, 167 AtbHLHs and 39 functionally annotated OsbHLHs (Additional file 1: Table S13, Additional file 2: Figure S2). According to the evolutionary relationship, thirteen major subfamilies in Arabidopsis and rice were classified in consistent with the BdbHLH phylogenic tree except for subfamily VI and XIV (Additional file 2, Fig. S2) [4, 13].
Exon/intron organization, as a type of structural divergence, plays an important role in the evolution of gene families [67]. As shown in Fig. 5, 122 were found to possess introns in their bHLH domains among the 146 BdbHLH proteins. 9 conserved intron positions and 12 different intron distribution patterns (I ~ XII) were identified in our study and pattern IV (introns position in His-9 and Val-31) with 6 members was not present in Arabidopsis [26]. BdbHLHs possessing diverse gene structures in the conserved bHLH domains were regularly distributed in the phylogenic tree (Fig. 4). For example, the majority of pattern I and X members were found in the subfamilies 4–7 and 12–16 respectively. In contrast, the members in the subfamilies 11 and 17, which were almost the Non-DNA binding type proteins, had pattern IV and pattern X respectively.
Fig. 5.
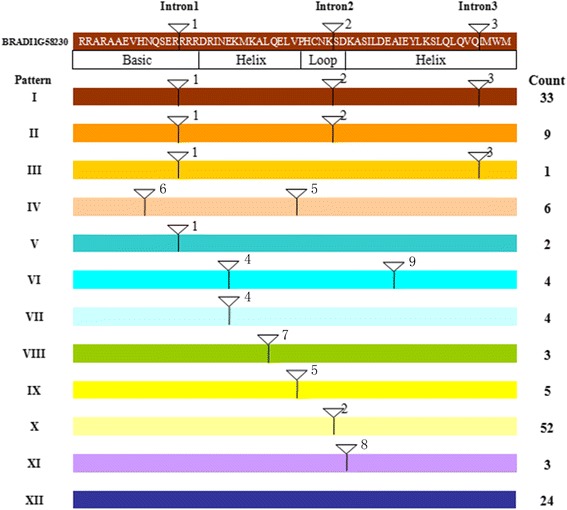
Intron distribution patterns in the coding sequence of the bHLH domain of BdbHLHs. The intron distribution patterns are marked with different colors, and position of introns is indicated by triangles. BRADI1G58230, as an example, is shown at top
Additionally, members of the same subfamily also displayed similar intron distribution patterns in view of the full-length genome sequences (Additional file 2: Fig. S3). For instance, all the BdbHLHs in subfamily 1 had only one exon, the whole members of subfamily 10 showed 2 exons while 7 out of 9 members in the group3 consisted of 5 exons which were in similar length and at similar positions.
In addition to bHLH domain, bHLH proteins in different subfamilies have different additional motifs which might be formed during evolution [7]. Totally, 15 conserved motifs were found (Additional file 2: Figure S3). Motif 1 and motif 2, located in bHLH domains, were found in almost all BdbHLHs (except for the BRADI4G05760 and BRADI1G12760). It is noteworthy that the BdbHLHs in the same subfamily were more likely to share same motif and location, which might imply similar biological functions [68]. For instance, a KRAAM motif before bHLH domain, which was reported to be involved in cold acclimation response [69], was found in BRADI2G59497 and BRADI4G17460 in subfamily18.
In Arabidopsis and rice, functions of many bHLH proteins have been characterized. In the subgroup Ia, MUTE [70], FMA [71, 72], SPCH [73–75], bHLH071 [71] associated with stomatal development and HWS [76] was related to sepal fusion and organ size. Meanwhile, MUTE, FMA and SPCH2 in rice were also reported to play a role in the differentiation and development of stomatal cells [77], implying that members in the subgroup Ia might be mainly involved in cell division and organ differentiation. In subgroup Ib, ORG2, ORG3 [78–81], bHLH100 and bHLH101 in Arabidopsis [80–83] play a key role in response to iron-deficiency. Additionally, OsIRO2 is also an essential regulator of Fe uptake and iron utilization in rice [84–86]. One except is, AtRGE1 primarily functions in embryo growth [87–91]. In Arabidopsis, three subgroup II members bHLH010, bHLH089 and bHLH091 were reported to interact with DYT1 proteins and redundantly participate in the anther development [92]. Similarly, rice subgroup II members EAT1 and TIP2 play a key role in the anther development at early stage [93, 94]. In subgroup IIIa, AMS [95–98] and DYT1 were identified to be master regulators of pollen development [99, 100]. Consistently, rice UDT1 interacted with TDR to regulate early anther development [101–105]. AtICE1, AtICE2 and OsICE1, OsICE2 in IIIb were proved to participate in the response to deep freezing [69, 106–110] while another member NFL was involved in GA mediated control of flowering time [111]. In Arabidopsis, only one member of subgroup IIIc, At4g29930, was functionally characterized. It might regulate hypocotyl and root elongation [112] while one member of IIIc in rice showed a correlation to JA inducible transcriptional activation during wound and drought stresses [113]. In Arabidopsis, IIId members (JAM1-JAM3 and bHLH014) and IIIe bHLHs (MYC2-MYC4) were proposed to take part in JA-mediated plant development [114–119]. In rice, OsMYL1 and OsMYL2 interact with OsMYC2 to participate in the JA signalings [120–122]. Subgroup IIIf protein TT8 participates in anthocyanin and PA pathways [123], similar to IIIf members in rice that involved in anthocyanidin biosynthesis [124–126]. Besides, three other IIIf proteins in Arabidopsis were found to be involved in the development of epidermal cells (GL3, EGL3 and MYC1) [127–130]. IVb (PYE) and IVc proteins (bHLH034, bHLH104, bHLH115 and ILR3) were proved to modulate metal homeostasis [26, 131–133]. Similarly, rice OsIRO3 of subgroup IVb also regulates iron homeostasis [134]. In IVd, AtbHLH092 responds to osmotic stress and regulates circadian rhythms [135, 136]; OsDPF participates in the resistance to diseases [137]. Va members (BIM1, BIM2 and BIM3) were suggested to participate in brassinosteroid signal and positively modulate the shade avoidance syndrome in seedlings [138, 139] while Vb members showed regulatory capacity in diverse processes including vascular development (ABS5 and TMO) [140, 141], hypocotyl and root elongation (At2g40200) [112] and responses to abiotic stresses (STC8) [142]. MEE8 in subgroup VI was speculated to regulate genes necessary to embryo and endosperm biogenesis [143]. The reported VIIa members (PIF1, PIF3-PIF5, PIL1 and PIL2) showed a relationship with photo induced signal transduction [144–155]. The rice subgroup VII proteins (OsPIL1, OsPIL11, OsPIL12, OsPIL14, OsPIL15 and APG) were functional counter-parts of PILs in Arabidopsis and involved in red light-mediated signal transduction pathways [156–161]. In addition, VIIb members in Arabidopsis are involved in cotyledon expansion and regulated seed dormancy (SPT) [162, 163], cell separation in fruit dehiscence (ALC) and interact with phytochromes (UNE10, RSF1 and PIF7) [164–167]. Two subgroup VIIIa members PAR1 and PAR2 were reported to integrate hormone and shade transcriptional networks and redundantly function in the enhancement of seedling de-etiolation related to phytoreceptor signal [139, 154, 168–171]. In subgroup VIIIb, HEC1-HEC3 modulated the development of the transmitting tract and stigma [172], fruit opening (IND) [173] and axillary meristem formation (ROX) [174]. Three rice VIIIb proteins have been studied, i.e. OsbHLH120 might control root thickness and length [175]; OsLF negatively regulated flowering [159, 176] and LAX specified the terminal spikelet meristem [177–179]. Members in subgroup VIIIc (RHD6, RSL1, RSL2, RSL4 and At2g14760) were verified to be essential to root hair development [130, 180, 181], while OsbHLH133, one characterized subgroup VIIIc member in rice, was proved to regulate the iron distribution between root and shoot [182]. Subgroup IX bHLHs were proposed to be involved in photoperiodism flowering (FBH1-FBH4) [183] and facilitate stomatal opening through phosphorylation (AKS2) [184]. Two studied X proteins (bHLH068 and bHLH112) could response to abiotic stresses [185, 186]. Subgroup XI members in Arabidopsis (LRL1-LRL5) [130, 187, 188] and rice (OsRHL1) [189] regulate root hair development, while another protein OsPTF1 is involved in phosphate starvation tolerance [190]. Subgroup XII members displayed diverse regulation functions: to monitor brassinosteroid signaling, to respond to freezing tolerance (CESTA, BEE1-BEE3 and BHI1) [191–195] and to promote flower initiation and regulate cell elongation (CIL1, CIL2 and CIB1-CIB5) [196–198]. In rice, only one XII member, An-1, has been functionally characterized. It regulates awn development, grain size and grain number [199]. Subgroup XIII proteins (UPB1, LL1, LL2 and LHW) were mainly required for the establishment and maintenance of normal vascular differentiation and development [200–203]. Subgroup XIV members (SAC51 and SACL1–3) might be involved in the response to thermospermine and xylem differentiation [204–206]. Subfamily XV members (PRE1–6) tend to be take parts in light, brassinosteroid and gibberellin signaling and modulation of flowering time [207–211]. In rice, four proteins of subgroup XV were reported to be involved in brassinosteroid signal (ILI1 and BU1) [212, 213] and regulation of grain length and weight (PGL2) [214].
The analysis of the functional characterized bHLH proteins in different subgroups in Arabidopsis and rice above, indicate the conservative function of bHLH members from different species in the same subgroups. According to the phylogenic tree, 79 BdbHLHs distributed in 21 subfamilies were found to have functionally characterized homologous proteins in Arabidopsis and rice (Additional file 1: Table S13). Based on these results, the functions of these 79 BdbHLH TFs could be predicted to some extent. Researches on several functionally characterized BdbHLH proteins further support this opinion.
One example is, in Arabidopsis, two RSL class I proteins AtRHD6 (ROOT HAIR DEFECTIVE6, AtbHLH083) and AtRSL1 (ROOT HAIR DEFECTIVE 6-LIKE 1, AtbHLH086), were reported to regulate the expression of the RSL2 and RSL4 and function as positive regulators to regulate the development of root hair cells [180, 215–217]. In our phylogenic tree, BRADI2G01000, BRADI3G53060 and BRADI1G42440 and AtRHD6, AtRSL1 were tightly grouped within a subfamily, indicating high homology among them. As predicted, BdRSL1(BRADI2G01000), BdRSL2 (BRADI3G53060) and BdRSL3 (BRADI1G42440) do function in the development of root hair cells [180, 218].
The other example is, in Arabidopsis, group IIIb members AtICE1 (AtbHLH116) and AtSCRM (AtbHLH033) function together with group Ia member AtSPCH (AtbHLH098) to regulate stomatal development [106]. BdICE1 (BRADI4G17460) and BdSCRM2 (BRADI2G59497) were tightly grouped with AtICE1 and AtSCRM in subfamily 18 in our phylogenic tree. Consistently, BdICE1 and BdSCRM2 cooperated with BdSPCH1 (BRADI1G38650) and BdSPCH2 (BRADI3G09670) to regulate the stomatal development despite the differences of their individual roles [219].
Expression profiles of BdbHLHs
Since the functions of genes associated with their expression patterns, the expression profiles of BdbHLH genes were analyzed. According to the available microarray data, the expression levels of BdbHLH genes in 9 different tissues varied considerably (Fig. 6). Genes in the same subfamily showed similar expression profiles at some level. For example, majority of genes in subfamily 4, 5, 6, 7, 8, 9, 18 and 19 showed relatively high expression in all detected tissues while the bulk of genes from subfamily 1, 2, 3, 10 and 21 showed lower or no expression. In contrast, some subfamilies were found to be specifically expressed in certain tissues. For example, genes from subfamily 2 were specifically expressed in anther; genes from subfamily 12 were found only in inflorescences and anther; the expression of subfamily 14 members were mainly found in plant embryo, emerging inflorescences and early inflorescences, implying that their functions have been differentiated.
Fig. 6.
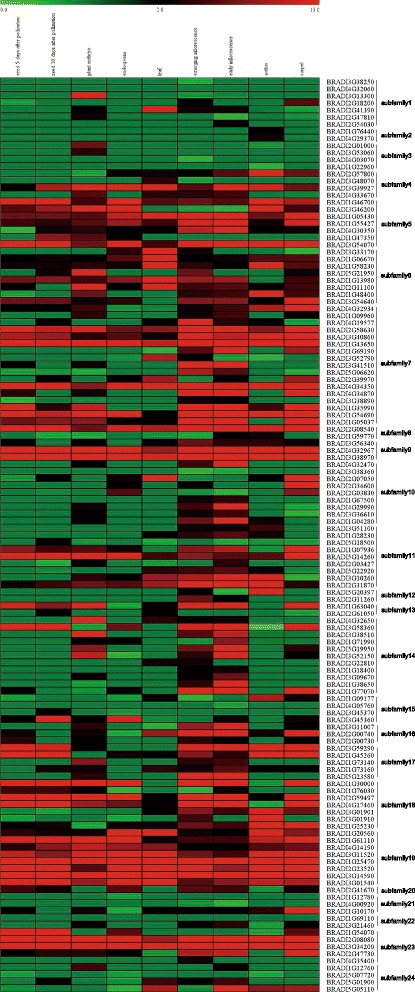
The expression profiles of BdbHLH genes in different tissues. The color scale is shown at the top. Higher expression levels are shown in red
Meanwhile, the expression level of different BdbHLHs was up or down-regulated by different phytohormones (Fig. 7). Genes in the same subfamily showed similar responses to exogenous phytohormones to some extent. For example, auxin down-regulated the expression of genes from subfamily 3 (except for BRADI3G53060 which was up-regulated by low concentration), 18 and 21 while high concentration of auxin up-regulated the expression of genes from subfamily 4; cytokine down-regulated the expression of genes in subfamily 20 while up-regulated the expression of genes in subfamily 21; SA down-regulated the expression of most genes in subfamily 9, 22, 3 (except for BRADI3G53060 which was up-regulated by low concentration) and subfamily 5 (at low concentration); ABA up-regulated the expression of subfamily 9 genes and down-regulated the expression of genes from subfamily 21 and subfamily 17 (except for that BRADI1G45260 was up-regulated by low concentration); JA down-regulated the expression of genes from subfamily 19, 20 and 5 (except for BRADI1G47350 was up-regulated at high concentration) while up-regulated the expression of genes from subfamily 9 and 21; GA down-regulated the expression of genes from subfamily 3, 9, 17 and 21; brassinosteroid down-regulated the expression of genes from subfamily 1 (low concentration) and subfamily 21; ethylene up-regulated the expression of genes from subfamily 4, 6 (except for BRADI5G21950 which was down-regulated by high concentration), 8 and 14 (low concentration) while down-regulated the expression of genes from subfamily16.
Fig. 7.
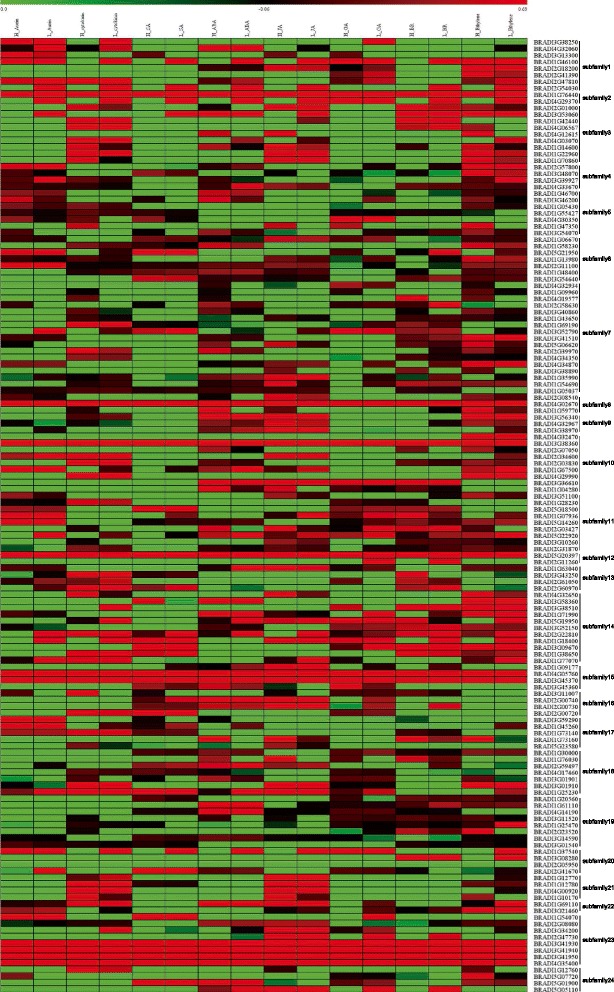
The expression profiles of BdbHLHs treated with high and low concentration of various phytohormones. The heatmap was generated with log2 based values. The color scale is shown at the top. Higher expression levels are shown in red while lower expression levels are shown in green. H stands for high concentration of phytohormone treatment while L stands for low concentration treatment
To further investigate the possible expression patterns of bHLHs in different organs and under abiotic stresses, the expression levels of 29 BdbHLHs randomly selected from 22 subfamilies were investigated using quantitative RT-PCR. As shown in Fig. 8a, during heading stage, 29 genes showed different expression patterns. For instance, BRADI2G54030 and BRADI2G07050 were primarily expressed in roots, while BRADI2G01000 showed a preferential expression in leaves. BRADI2G61050 was expressed high in root, stem and leaves but low in inflorescences. Genes from same subfamily probably display similar expression patterns. BRADI1G05037, BRADI3G41510, BRADI4G34350 and BRADI1G69190 from subfamily 7 showed low expression in four tested tissues and a relatively high expression level in leaves and inflorescences; BRADI1G28230 and BRADI5G14260 from subfamily 11 and BRADI1G71990 and BRADI3G09670 from subfamily 14 exhibited high expression in leaves while BRADI1G20560 and BRADI1G25470 from subfamily 19 were hardly detected in root. Taking the conserved motifs and similar gene structure into account, we predict genes in the same subfamily might play redundant roles to some extent.
Fig. 8.
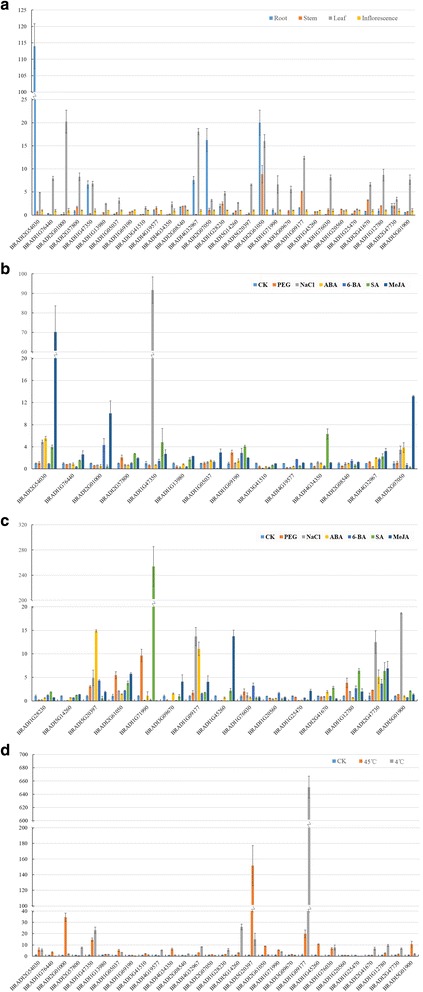
Quantitative RT-PCR analysis of 29 selected BdbHLH genes. The relative expression levels of the 29 genes in (a) different organs namely root, stem, leaf and inflorescence; (b and c) root with different treatments including 20% PEG6000, 200 mM NaCl, 100 μM MeJA, 100 μM ABA, 20 μM 6-BA and 1 mM SA; (d) leaf with treatments of high temperature (45 C) and low temperature (4 C)
Additionally, the expression profiles of 29 selected genes under different abiotic stresses were investigated too (Fig. 8b–d) and the cis-regulatory elements were analyzed to verify our results (Additional file 1: Table S14). In general, the expression patterns are consistent with the analysis of promoters. For example, BRADI1G47350 and BRADI2G54030 were strongly induced by NaCl and MeJA, respectively, consistent with the prediction that BRADI1G47350 contains 5 ABRE cis-elements (element respond to salt stress) and BRADI2G54030 contains 3 TGACG-motifs (cis-acting regulatory element involved in the MeJA-responsiveness) [220]; BRADI1G71990 containing 1 TCA-element (cis-acting element involved in salicylic acid responsiveness) was drastically up-regulated by SA. It might interact with BRADI3G51960 to respond to salicylic acid according to the interaction network. The expression of BRADI1G09177, BRADI5G01900 and BRADI5G20397 were up-regulated by NaCl and ABA, consistent with the presence of ABRE cis-elements in these genes. BRADI1G45260, BRADI2G01000 and BRADI2G07050, containing TGACG-motifs, showed higher expression under MeJA treatment. For extreme temperature including heat (45 °C) and cold (4 °C), the expression of BRADI1G09177 and BRADI5G20397 were strongly up-regulated by cold and heat treatment respectively, in accordance with the presence of DRE (regulatory element involved in cold- and dehydration-responsiveness) [221], or HSE (cis-acting element involved in heat stress responsiveness) [222].
Conclusions
To study the bHLH gene family in the Brachypodium distachyon, an emerging model plant in grass, we identified 146 bHLH genes distributed in 5 chromosomes. Gene duplications showed that duplication events, especially segment duplications made up a large proportion in the expansion of BdbHLHs. Synteny analyses indicated that bHLHs in Brachypodium distachyon had close relationships with rice, maize and sorghum. GO analysis showed that the majority of BdbHLHs were involved in transcriptional regulation and displayed protein binding ability, suggesting that they might function in homodimer or heterodimer manners. According to phylogenetic analysis of the bHLH domains and the alignment with full-length sequences of Arabidopsis and rice, BdbHLH TFs were classified into 24 subfamilies. Based on the functional characterization of homologous genes in Arabidopsis and rice, the BdbHLHs were predicted to take part in various processes including growth and development, stress responses and so on. The expression profiles of BdbHLH genes in different tissues and under different phytohormones treatments were analyzed, and some tissue-specific and phytohormone-responsive genes were identified. Taken together, our results provide a solid foundation for further evolutionary and functional investigations on BdbHLHs.
Additional files
Consensus motifs of bHLH domains. Table S2. The primers used in the qRT-PCR. Table S3. Characteristic features of bHLH Transcription factor gene family identified in Brachypodium distachyon. Table S4. The Ka/Ks ratios and estimated divergence time for tandemly duplicated BdbHLH genes. Table S5. The Ka/Ks ratios and estimated divergence time for segmentally duplicated BdbHLH genes. Table S6. The Ka/Ks ratios and estimated divergence time for orthologous bHLH genes between Brachypodium distachyon and rice. Table S7. The Ka/Ks ratios and estimated divergence time for orthologous bHLH genes between Brachypodium distachyon and maize. Table S8. The Ka/Ks ratios and estimated divergence time for orthologous bHLH genes between Brachypodium distachyon and sorghum. Table S9. GO annotations of BdbHLHs. Table S10. GO descriptions of BdbHLHs. Table S11. Detailed information of interaction network of BdbHLH with other proteins. Table S12. Predicted DNA Binding Characteristics of the bHLH Domain in BdbHLH Proteins. Table S13. Summary of the predicted functions of BdbHLHs by comparative analysis with homologous bHLHs in Arabidopsis and rice. Table S14. The cis-regulatory elements in the promoter region of 29 BdbHLH genes. (XLSX 237 kb)
GO annotations of BdbHLHs. Figure S2. Phylogenic tree using alignments of bHLH proteins in Brachypodium distachyon, Arabidopsis and rice. Figure S3. Conserved elements and gene structures of BdbHLHs. (RAR 22526 kb)
The sequences of all the BdbHLH genes, including CDS, amino acids of the bHLH domains, protein and genomic DNA as well as 1500 bp upstream sequences. (RAR 398 kb)
Acknowledgements
Not applicable.
Funding
This work was supported by the Open Project Program of State Key Laboratory of Crop Stress Biology for Arid Areas, NWAFU (No. CSBAA2016016); Research Funds from Xinjiang Agricultural Vocational Technical College (No. XJNZYKJ201501) and the Fundamental Research Funds for the Central Universities (No. Z109021565). The funders had no role in the design of the study and collection, analysis, and interpretation of data and in writing the manuscript.
Availability of data and materials
The genome, cds, cDNA and protein sequences of bHLHs in Arabidopsis, rice and Brachypodium distachyon were obtained from the National Center of Biotechnology Information database (http://www.ncbi.nlm.nih.gov) and the Gramene database (http://www.gramene.org/) and were listed in Additional file 3. The phylogenetic trees of BdbHLHs and AtbHLHs, OsbHLHs and BdbHLHs generated in this study have been uploaded to TreeBASE with accession number NO. S21381. The access URL is (http://purl.org/phylo/treebase/phylows/study/TB2:S21381).
Abbreviations
- ABA
Abscisic acid
- bHLH
basic helix-loop-helix
- EST
Expressed sequence tag
- FPKM
Fragments kilobase of exon model per million mapped reads
- GO
Gene ontology
- GRAVY
Grand average of hydropathicity
- Ka
Substitution rate of non-synonymous
- Ks
Substitution rate of synonymous
- MeJA
Methyl jasmonate
- NJ
Neighbor joining
- PI
Isoelectric point
- SA
Salicylic acid
- TF
Transcription factor
Authors’ contributions
HL designed the experiments and revised the manuscript; XN, YG and SC collected and analyzed the public data and also drafted the manuscript; Manuscript preparation: HL, XN, YG; All the authors have read and approved the final manuscript.
Ethics approval and consent to participate
The plant materials used here were grown in the greenhouse of NWAFU for research use only. The seeds of Bd21 were presented by Dr. Jinrong Xu and Dr. Huiquan Liu (College of Plant Protection, NWAFU). Brachypodium distachyon is not listed in the Convention on International Trade in Endangered Species of Wild Fauna and Flora Appendices I, II and III (valid from 4 April 2017, https://cites.org/eng/app/appendices.php). Collection of plant materials complied with the institutional, national and international guidelines.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Footnotes
Electronic supplementary material
The online version of this article (doi:10.1186/s12864-017-4044-4) contains supplementary material, which is available to authorized users.
Contributor Information
Xin Niu, Email: 509489502@qq.com.
Yuxiang Guan, Email: 1026489469@qq.com.
Shoukun Chen, Email: 912872684@qq.com.
Haifeng Li, Email: lhf@nwsuaf.edu.cn.
References
- 1.Bevan MW, Garvin DF, Vogel JP. Brachypodium Distachyon genomics for sustainable food and fuel production. Curr Opin Biotech. 2010;21(2):211–217. doi: 10.1016/j.copbio.2010.03.006. [DOI] [PubMed] [Google Scholar]
- 2.Somerville C. The billion-ton biofuels vision. Science. 2006;312(5778):1277–1276. doi: 10.1126/science.1130034. [DOI] [PubMed] [Google Scholar]
- 3.Jones S. An overview of the basic helix-loop-helix proteins. Genome Biol. 2004;5(6):60–66. doi: 10.1186/gb-2004-5-6-226. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 4.Carretero-Paulet L, Galstyan A, Roig-Villanova I, Martínez-García JF, Bilbao-Castro JR, Robertson DL. Genome-wide classification and evolutionary analysis of the bHLH family of transcription factors in Arabidopsis, poplar, Rice, Moss, and algae. Plant Physiol. 2010;153(153):1398–1412. doi: 10.1104/pp.110.153593. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 5.Pires N, Dolan L. Origin and diversification of basic-helix-loop-helix proteins in plants. Mol Biol Evol. 2010;27(4):862–874. doi: 10.1093/molbev/msp288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 6.Feller A, Machemer K, Braun EL, Grotewold E. Evolutionary and comparative analysis of MYB and bHLH plant transcription factors. Plant J. 2011;66(1):94–116. doi: 10.1111/j.1365-313X.2010.04459.x. [DOI] [PubMed] [Google Scholar]
- 7.Atchley WR, Terhalle W, Dress A. Positional dependence, cliques, and predictive motifs in the bHLH protein domain. J Mol Evol. 1999;48(5):501–516. doi: 10.1007/PL00006494. [DOI] [PubMed] [Google Scholar]
- 8.Atchley WR, Fitch WM. A natural classification of the basic helix-loop-helix class of transcription factors. Proc Natl Acad Sci U S A. 1997;94(10):5172–5176. doi: 10.1073/pnas.94.10.5172. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Murre C, Mccaw PS, Baltimore D. A new DNA binding and dimerization motif in immunoglobulin enhancer binding, daughterless, MyoD, and myc proteins. Cell. 1989;56(5):777–783. doi: 10.1016/0092-8674(89)90682-X. [DOI] [PubMed] [Google Scholar]
- 10.Ledent V, Vervoort M. The basic helix-loop-helix protein family: comparative genomics and phylogenetic analysis. Genome Res. 2001;11(5):754–770. doi: 10.1101/gr.177001. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Ledent V, Paquet O, Vervoort M. Phylogenetic analysis of the human basic helix-loop-helix proteins. Genome Biol. 2002;3(6):1–18. doi: 10.1186/gb-2002-3-6-research0030. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 12.Fisher A, Caudy M. The function of hairy-related bHLH repressor proteins in cell fate decisions. BioEssays. 1998;20(4):298–306. doi: 10.1002/(SICI)1521-1878(199804)20:4<298::AID-BIES6>3.0.CO;2-M. [DOI] [PubMed] [Google Scholar]
- 13.Heim MA, Jakoby M, Werber M, Martin C, Weisshaar B, Bailey PC. The basic helix-loop-helix transcription factor family in plants: a genome-wide study of protein structure and functional diversity. Mol Biol Evol. 2003;20(5):735. doi: 10.1093/molbev/msg088. [DOI] [PubMed] [Google Scholar]
- 14.Li X, Duan X, Jiang H, Sun Y, Tang Y, Yuan Z, Guo J, Liang W, Chen L, Yin J. Genome-wide analysis of basic/helix-loop-helix transcription factor family in rice and Arabidopsis. Plant Physiol. 2006;141(4):1167–1184. doi: 10.1104/pp.106.080580. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 15.Rushton PJ, Timko MP. Tobacco transcription factors: novel insights into transcriptional regulation in the Solanaceae. Plant Physiol. 2008;147(1):280–295. doi: 10.1104/pp.107.114041. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 16.Chen YY, Li MY, Wu XJ, Huang Y, Ma J, Xiong AS. Genome-wide analysis of basic helix−loop−helix family transcription factors and their role in responses to abiotic stress in carrot. Mol Breeding. 2015;35(5):1–12. [Google Scholar]
- 17.Zhang X, Luo H, Xu Z, Zhu Y, Ji A, Song J, Chen S. Genome-wide characterisation and analysis of bHLH transcription factors related to tanshinone biosynthesis in Salvia miltiorrhiza. Sci Rep. 2015;5:11244. doi: 10.1038/srep11244. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 18.Sun H, Fan HJ, Ling HQ. Genome-wide identification and characterization of the bHLH gene family in tomato. BMC Genomics. 2015;16(1):1–16. doi: 10.1186/1471-2164-16-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Oh E, Kim J, Park E, Kim JI, Kang C, Choi G. PIL5, a phytochrome-interacting basic helix-loop-helix protein, is a key negative regulator of seed germination in Arabidopsis Thaliana. Plant Cell. 2004;16(11):3045–3058. doi: 10.1105/tpc.104.025163. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 20.Zhao H, Li X, Ma L. Basic helix-loop-helix transcription factors and epidermal cell fate determination in Arabidopsis. Plant Signal Behav. 2012;7(12):1556–1560. doi: 10.4161/psb.22404. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.Groszmann M, Bylstra Y, Lampugnani ER, Smyth DR. Regulation of tissue-specific expression of SPATULA, a bHLH gene involved in carpel development, seedling germination, and lateral organ growth in Arabidopsis. J Exp Bot. 2010;61(5):1495–1508. doi: 10.1093/jxb/erq015. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Farquharson KL. A domain in the bHLH transcription factor DYT1 is critical for anther development. Plant Cell. 2016;28(5):997–8. doi: 10.1105/tpc.16.00331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Rajani S, Sundaresan V. The Arabidopsis myc/bHLH gene ALCATRAZ enables cell separation in fruit dehiscence. Curr Biol. 2001;11(24):1914–1922. doi: 10.1016/S0960-9822(01)00593-0. [DOI] [PubMed] [Google Scholar]
- 24.Pablo Leivar EM, Oka Y, Liu T, Carle C, Castillon A, Huq E, Quail PH. Multiple phytochrome-interacting bHLH transcription factors repress premature photomorphogenesis during early seedling development in darkness. Curr Biol. 2008;18(23):1815–23. [DOI] [PMC free article] [PubMed]
- 25.Nakata M, Mitsuda N, Herde M, Koo AJK, Moreno JE, Suzuki K, Howe GA, Ohmetakagi M. A bHLH-type transcription factor, ABA-INDUCIBLE BHLH-TYPE TRANSCRIPTION FACTOR/JA-ASSOCIATED MYC2-LIKE1, acts as a repressor to negatively regulate jasmonate signaling in arabidopsis. Plant Cell. 2013;25(5):1641–1656. doi: 10.1105/tpc.113.111112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Long TA, Tsukagoshi H, Busch W, Lahner B, Salt DE, Benfey PN. The bHLH transcription factor POPEYE regulates response to iron deficiency in Arabidopsis roots. Plant Cell. 2010;22(7):2219–2236. doi: 10.1105/tpc.110.074096. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 27.Opanowicz M, Vain P, Draper J, Parker D, Doonan JH. Brachypodium Distachyon: making hay with a wild grass. Trends Plant Sci. 2008;13(4):172–177. doi: 10.1016/j.tplants.2008.01.007. [DOI] [PubMed] [Google Scholar]
- 28.Vogel JP, Garvin DF, Mockler TC, Schmutz J, Dan R, Bevan MW, Barry K, Lucas S, Harmonsmith M, Lail K. Genome sequencing and analysis of the model grass Brachypodium Distachyon. Nature. 2010;463(7282):763–768. doi: 10.1038/nature08747. [DOI] [PubMed] [Google Scholar]
- 29.Toledo-Ortiz G, Quail PH. The Arabidopsis basic/helix-loop-helix transcription factor family. Plant Cell Plant Cell. 2003;15(8):1749–1770. doi: 10.1105/tpc.013839. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 30.Schultz J, Milpetz F, Bork P, Ponting CP. SMART, a simple modular architecture research tool: identification of signaling domains. Proc Natl Acad Sci U S A. 1998;95(11):5857–5864. doi: 10.1073/pnas.95.11.5857. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 31.Artimo P, Jonnalagedda M, Arnold K, Baratin D, Csardi G, Castro ED, Duvaud S, Flegel V, Fortier A, Gasteiger E. ExPASy: SIB bioinformatics resource portal. Nucleic Acids Res. 2012;40(Web Server issue):W597–W603. doi: 10.1093/nar/gks400. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Jin J, Feng T, Yang DC, Meng YQ, Lei K, Luo J, Ge G. PlantTFDB 4.0: toward a central hub for transcription factors and regulatory interactions in plants. Nucleic Acids Res. 2016;45(Database issue):D1040–D1045. doi: 10.1093/nar/gkw982. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Ye J, Fang L, Zheng H, Zhang Y, Chen J, Zhang Z, Wang J, Li S, Li R, Bolund L. WEGO: a web tool for plotting GO annotations. Nucleic Acids Research. 2006;34(Web Server issue):W293–W297. doi: 10.1093/nar/gkl031. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Mol Biol Evol. 2013;30(12):2725–2729. doi: 10.1093/molbev/mst197. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 35.He Z, Zhang H, Gao S, Lercher MJ, Chen WH, Hu S. Evolview v2: an online visualization and management tool for customized and annotated phylogenetic trees. Nucleic Acids Res. 2016;44(W1):W236–W241. doi: 10.1093/nar/gkw370. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 36.Chen L, Hu W, Tan S, Wang M, Ma Z, Zhou S, Deng X, Zhang Y, Huang C, Yang G. Genome-wide identification and analysis of MAPK and MAPKK gene families in Brachypodium Distachyon. PLoS One. 2012;7(10):440–444. doi: 10.1371/journal.pone.0046744. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Lee TH, Tang H, Wang X, Paterson AH. PGDD: a database of gene and genome duplication in plants. Nucleic Acids Res. 2013;41(Database issue):1152–1158. doi: 10.1093/nar/gks1104. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Krzywinski MI, Schein JE, Birol I, Connors J, Gascoyne R, Horsman D, Jones SJ, Marra MA. Circos: An information aesthetic for comparative genomics. Genome Res. 2009;19(9):1639–45. doi: 10.1101/gr.092759.109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Lescot M, Déhais P, Thijs G, Marchal K, Moreau Y, Yves VDP, Rouzé P, Rombauts S. PlantCARE, a database of plant cis-acting regulatory elements and a portal to tools for in silico analysis of promoter sequences. Nucleic Acids Res. 2002;30(1):325–327. doi: 10.1093/nar/30.1.325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Hu B, Jin JP, Guo AY, Zhang H, Luo JC, Gao G. GSDS 2.0: an upgraded gene feature visualization server. Bioinformatics. 2015;31(8):1296–1297. doi: 10.1093/bioinformatics/btu817. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 41.Bailey TL, Boden M, Buske FA, Frith M, Grant CE, Clementi L, Ren JY, Li WW, Noble WS. MEME SUITE: tools for motif discovery and searching. Nucleic Acids Res. 2009;37:W202–W208. doi: 10.1093/nar/gkp335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Lee T, Yang S, Kim E, Ko Y, Hwang S, Shin J, Shim JE, Shim H, Kim H, Kim C. AraNet v2: an improved database of co-functional gene networks for the study of Arabidopsis Thaliana and 27 other nonmodel plant species. Nucleic Acids Res. 2015;43(Database issue):D996. doi: 10.1093/nar/gku1053. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Shannon P, Markiel A, Ozier O, Baliga NS, Wang JT, Ramage D, Amin N, Schwikowski B, Ideker T. Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 2003;13(11):2498–2504. doi: 10.1101/gr.1239303. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Davidson RM, Gowda M, Moghe G, Lin H, Vaillancourt B, Shiu SH, Jiang N, Buell CR. Comparative transcriptomics of three Poaceae species reveals patterns of gene expression evolution. Plant J. 2012;71(3):492–502. doi: 10.1111/j.1365-313X.2012.05005.x. [DOI] [PubMed] [Google Scholar]
- 45.Kakei Y, Mochida K, Sakurai T, Yoshida T, Shinozaki K, Shimada Y. Transcriptome analysis of hormone-induced gene expression in Brachypodium Distachyon. Sci Rep. 2015;5:14476. doi: 10.1038/srep14476. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Chu VT, Gottardo R, Raftery AE, Bumgarner RE, Yeung KY. MeV+R: using MeV as a graphical user interface for Bioconductor applications in microarray analysis. Genome Biol. 2008;9(7):R118. doi: 10.1186/gb-2008-9-7-r118. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Liu M, Li H, Su Y, Li W, Shi C. G1/ELE Functions in the Development of Rice Lemmas in Addition to Determining Identities of Empty Glumes. Front Plant Sci. 2016;7:1006. doi: 10.3389/fpls.2016.01006. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 48.Hong SY, Seo PJ, Yang MS, Xiang F, Park CM. Exploring valid reference genes for gene expression studies in Brachypodium Distachyon by real-time PCR. BMC Plant Bio. 2008;8(1):112. doi: 10.1186/1471-2229-8-112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 49.Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2(−Delta Delta C(T)) method. Methods. 2001;25(4):402–408. doi: 10.1006/meth.2001.1262. [DOI] [PubMed] [Google Scholar]
- 50.Wang J, Hu Z, Zhao T, Yang Y, Chen T, Yang M, Yu W, Zhang B. Genome-wide analysis of bHLH transcription factor and involvement in the infection by yellow leaf curl virus in tomato ( Solanum Lycopersicum ) BMC Genomics. 2015;16(1):1–14. doi: 10.1186/1471-2164-16-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 51.Kavas M, Baloğlu MC, Atabay ES, Ziplar UT, Daşgan HY, Ünver T: Genome-wide characterization and expression analysis of common bean bHLH transcription factors in response to excess salt concentration. Mol Gen Genomics 2016, 291(1):1–15. [DOI] [PubMed]
- 52.Kent WJ, Baertsch R, Hinrichs A, Miller W, Haussler D. Evolution's cauldron: duplication, deletion, and rearrangement in the mouse and human genomes. Proc Natl Acad Sci U S A. 2003;100(20):11484–11489. doi: 10.1073/pnas.1932072100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Lynch M, Conery JS. The evolutionary fate and consequences of duplicate genes. Science. 2000;290(5494):1151–1155. doi: 10.1126/science.290.5494.1151. [DOI] [PubMed] [Google Scholar]
- 54.Brownlie P, Ceska TA, Lamers M, Romier C, Stier G, Teo H, Suck D. The crystal structure of an intact human max–DNA complex: new insights into mechanisms of transcriptional control. Structure. 1997;5(4):509–520. doi: 10.1016/S0969-2126(97)00207-4. [DOI] [PubMed] [Google Scholar]
- 55.Lian TF, Xu YP, Li LF, Su XD. Crystal structure of tetrameric Arabidopsis MYC2 reveals the mechanism of enhanced interaction with DNA. Cell Rep. 2017;19(7):1334–1342. doi: 10.1016/j.celrep.2017.04.057. [DOI] [PubMed] [Google Scholar]
- 56.Abe H, Urao T, Ito T, Seki M, Shinozaki K, Yamaguchishinozaki K. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell. 2003;15(1):63–78. doi: 10.1105/tpc.006130. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Schaart JG, Dubos C, Irene RDLF, Vos RCH, Jonker HH, Xu W, Routaboul JM, Lepiniec L, Bovy AG. Identification and characterization of MYB-bHLH-WD40 regulatory complexes controlling proanthocyanidin biosynthesis in strawberry (Fragaria×ananassa) fruits. New Phytol. 2012;197(2):454–467. doi: 10.1111/nph.12017. [DOI] [PubMed] [Google Scholar]
- 58.Ramsay NA, Glover BJ. MYB-bHLH-WD40 protein complex and the evolution of cellular diversity. Trends Plant Sci. 2005;10(2):63–70. doi: 10.1016/j.tplants.2004.12.011. [DOI] [PubMed] [Google Scholar]
- 59.Pérez-Rodríguez P, Riaño-Pachón DM, Corrêa LG, Rensing SA, Kersten B, Mueller-Roeber B. PlnTFDB: updated content and new features of the plant transcription factor database. Nucleic Acids Res. 2009;38(Database issue):D822–D827. doi: 10.1093/nar/gkp805. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.Fisher F, Goding CR. Single amino acid substitutions alter helix-loop-helix protein specificity for bases flanking the core CANNTG motif. EMBO J. 1992;11(11):4103–4109. doi: 10.1002/j.1460-2075.1992.tb05503.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 61.Ellenberger T, Fass D, Arnaud M, Harrison SC. Crystal-structure of transcription factor E47 - E-box recognition by a basic region helix-loop-helix dimer. Genes Dev. 1994;8(8):970–980. doi: 10.1101/gad.8.8.970. [DOI] [PubMed] [Google Scholar]
- 62.Shimizu T, Toumoto A, Ihara K, Shimizu M, Kyogoku Y, Ogawa N, Oshima Y, Hakoshima T. Crystal structure of PHO4 bHLH domain-DNA complex: flanking base recognition. EMBO J. 1997;16(15):4689–4697. doi: 10.1093/emboj/16.15.4689. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Fujii Y, Shimizu T, Toda T, Yanagida M, Hakoshima T. Structural basis for the diversity of DNA recognition by bZIP transcription factors. Nat Struct Biol. 2000;7(10):889–893. doi: 10.1038/82822. [DOI] [PubMed] [Google Scholar]
- 64.Ma PC, Rould MA, Weintraub H, Pabo CO. Crystal structure of MyoD bHLH domain-DNA complex: perspectives on DNA recognition and implications for transcriptional activation. Cell. 1994;77(3):451. doi: 10.1016/0092-8674(94)90159-7. [DOI] [PubMed] [Google Scholar]
- 65.Ferre-D'Amare AR, Pognonec P, Roeder RG, Burley SK. Structure and function of the b/HLH/Z domain of USF. EMBO J. 1994;13(1):180–189. doi: 10.1002/j.1460-2075.1994.tb06247.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 66.Fairman R, Beransteed RK, Anthonycahill SJ, Lear JD, Rd SW, Degrado WF, Benfield PA, Brenner SL. Multiple oligomeric states regulate the DNA binding of helix-loop-helix peptides. Proc Natl Acad Sci U S A. 1993;90(22):10429–10433. doi: 10.1073/pnas.90.22.10429. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 67.Xu G, Guo C, Shan H, Kong H. Divergence of duplicate genes in exon-intron structure. Proc Natl Acad Sci U S A. 2012;109(4):1187–1192. doi: 10.1073/pnas.1109047109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Kang WH, Kim S, Lee HA, Choi D, Yeom SI. Genome-wide analysis of Dof transcription factors reveals functional characteristics during development and response to biotic stresses in pepper. Sci Rep. 2016;6:33332. doi: 10.1038/srep33332. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Chinnusamy V, Ohta M, Kanrar S, Lee BH, Hong X, Agarwal M, Zhu JK. ICE1: a regulator of cold-induced transcriptome and freezing tolerance in Arabidopsis. Genes Dev. 2003;17(17):1043–1054. doi: 10.1101/gad.1077503. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 70.Pillitteri LJ, Sloan DB, Bogenschutz NL, Torii KU. Termination of asymmetric cell division and differentiation of stomata. Nature. 2007;445(7127):501–505. doi: 10.1038/nature05467. [DOI] [PubMed] [Google Scholar]
- 71.Ohashi-Ito K, Bergmann DC. Arabidopsis FAMA controls the final proliferation/differentiation switch during stomatal development. Plant Cell. 2006;18(10):2493–2505. doi: 10.1105/tpc.106.046136. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 72.Shirakawa M, Ueda H, Nagano AJ, Shimada T, Kohchi T, Haranishimura I. FAMA is an essential component for the differentiation of two distinct cell types, myrosin cells and guard cells, in Arabidopsis. Plant Cell. 2014;26(10):4039–4052. doi: 10.1105/tpc.114.129874. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.Marcos AD, Houbaert A, Triviño M, Delgado D, Martíntrillo M, Russinova E, Fenoll C, Mena M. A mutation in the bHLH domain of the SPCH transcription factor uncovers a BR-dependent mechanism for stomatal development. Plant Physiol. 2017;174(2):823. doi: 10.1104/pp.17.00615. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Macalister CA, Ohashiito K, Bergmann DC. Transcription factor control of asymmetric cell divisions that establish the stomatal lineage. Nature. 2006;445(7127):537–540. doi: 10.1038/nature05491. [DOI] [PubMed] [Google Scholar]
- 75.Lampard GR, Macalister CA, Bergmann DC. Arabidopsis stomatal initiation is controlled by MAPK-mediated regulation of the bHLH SPEECHLESS. Science. 2008;322(5904):1113. doi: 10.1126/science.1162263. [DOI] [PubMed] [Google Scholar]
- 76.Gonzalez-Carranza Z, Rompa U, Peters J, Bhatt A, Wagstaff C, Stead A, Roberts J. Hawaiian skirt : an F-box gene that regulates organ fusion and growth in Arabidopsis. Plant Physiol. 2007;144(3):1370. doi: 10.1104/pp.106.092288. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Liu T, Ohashiito K, Bergmann DC. Orthologs of Arabidopsis Thaliana stomatal bHLH genes and regulation of stomatal development in grasses. Development. 2009;136(13):2265–2276. doi: 10.1242/dev.032938. [DOI] [PubMed] [Google Scholar]
- 78.Youxi, Yuan, Huilan, Ning, Wang, Weina, Zhao, Juan, Daowen, Wang: FIT interacts with AtbHLH38 and AtbHLH39 in regulating iron uptake gene expression for iron homeostasis in Arabidopsis. Cell Res 2008, 18(3):385. [DOI] [PubMed]
- 79.Kang HG, Foley RC, Oñate-Sánchez L, Lin C, Singh KB. Target genes for OBP3, a Dof transcription factor, include novel basic helix-loop-helix domain proteins inducible by salicylic acid. Plant J. 2003;35(3):362–372. doi: 10.1046/j.1365-313X.2003.01812.x. [DOI] [PubMed] [Google Scholar]
- 80.Ning, Wang, Huajie, Juan, Zongan, Huang, Youxi, yuan, Huilan, Hong-Qing: Requirement and functional redundancy of Ib subgroup bHLH proteins for iron deficiency responses and uptake in Arabidopsis Thaliana. Mol Plant 2013, 6(2):503–513. [DOI] [PubMed]
- 81.Maurer F, Naranjo Arcos MA, Bauer P. Responses of a triple mutant Defective in three iron deficiency-induced BASIC HELIX-LOOP-HELIX genes of the subgroup Ib(2) to iron deficiency and salicylic acid. PLoS One. 2014;9(6):e99234. doi: 10.1371/journal.pone.0099234. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 82.Sivitz AB, Hermand V, Curie C, Vert G. Arabidopsis bHLH100 and bHLH101 control iron homeostasis via a FIT-independent pathway. PLoS One. 2012;7(9):e44843. doi: 10.1371/journal.pone.0044843. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Van DJ, Antoniou C, Filippou P, Pollier J, Gonzalez N, Dhondt S, Goossens A, Fotopoulos V, Inzé D. Strobilurins as growth-promoting compounds: How Stroby regulates Arabidopsis leaf growth. Plant Cell Environ. 2017.doi:10.1111/pce.12980. [DOI] [PubMed]
- 84.Ogo Y, Itai RN, Nakanishi H, Inoue H, Kobayashi T, Suzuki M, Takahashi M, Mori S, Nishizawa NK. Isolation and characterization of IRO2, a novel iron-regulated bHLH transcription factor in graminaceous plants. J Exp Bot. 2006;57(11):2867–2878. doi: 10.1093/jxb/erl054. [DOI] [PubMed] [Google Scholar]
- 85.Ogo Y, Itai R, Nakanishi H, Kobayashi T, Takahashi M, Mori S, Nishizawa N. The rice bHLH protein OsIRO2 is an essential regulator of the genes involved in Fe uptake under Fe-deficient conditions. Plant Cell Physiol. 2007;48:S44–SS4. doi: 10.1111/j.1365-313X.2007.03149.x. [DOI] [PubMed] [Google Scholar]
- 86.Ogo Y, Itai RN, Kobayashi T, Aung MS, Nakanishi H, Nishizawa NK. OsIRO2 is responsible for iron utilization in rice and improves growth and yield in calcareous soil. Plant Mol Biol. 2011;75(6):593–605. doi: 10.1007/s11103-011-9752-6. [DOI] [PubMed] [Google Scholar]
- 87.Kondou Y, Nakazawa M, Kawashima M, T, Yoshizumi T, Suzuki K, Ishikawa A, Koshi T, Matsui R, Muto S: RETARDED GROWTH OF EMBRYO1, a new basic helix-loop-helix protein, expresses in endosperm to control EMBRYO growth. Plant Physiol 2008, 147(4):1924–1935. [DOI] [PMC free article] [PubMed]
- 88.Yang S, Johnston N, Talideh E, Mitchell S, Jeffree C, Goodrich J, et al. The endosperm-specific zhoupi gene of arabidopsis thaliana regulates endosperm breakdown and embryonic epidermal development. Development. 2008;135(21):3501–9. doi: 10.1242/dev.026708. [DOI] [PubMed] [Google Scholar]
- 89.Denay G, Creff A, Moussu S, Wagnon P, Thévenin J, Gérentes MF, Chambrier P, Dubreucq B, Ingram G. Endosperm breakdown in Arabidopsis requires heterodimers of the basic helix-loop-helix proteins ZHOUPI and INDUCER OF CBP EXPRESSION 1. Development. 2014;141(6):1222–1227. doi: 10.1242/dev.103531. [DOI] [PubMed] [Google Scholar]
- 90.Xing Q, Creff A, Waters A, Tanaka H, Goodrich J, Ingram GC. ZHOUPI controls embryonic cuticle formation via a signalling pathway involving the subtilisin protease ABNORMAL LEAF-SHAPE1 and the receptor kinases GASSHO1 and GASSHO2. Development. 2013;140(4):770–779. doi: 10.1242/dev.088898. [DOI] [PubMed] [Google Scholar]
- 91.Fourquin C, Beauzamy L, Chamot S, Creff A, Goodrich J, Boudaoud A, Ingram G. Mechanical stress mediated by both endosperm softening and embryo growth underlies endosperm elimination in Arabidopsis seeds. Development. 2016;143(18):3300–5. doi: 10.1242/dev.137224. [DOI] [PubMed] [Google Scholar]
- 92.Zhu E, You C, Wang S, Cui J, Niu B, Wang Y, Qi J, Ma H, Chang F. The DYT1-interacting proteins bHLH010, bHLH089 and bHLH091 are redundantly required for Arabidopsis anther development and transcriptome. Plant J. 2015;83(6):976. doi: 10.1111/tpj.12942. [DOI] [PubMed] [Google Scholar]
- 93.Fu ZZ, Yu J, Cheng XW, Zong X, Xu J, Chen MJ, Li ZY, Zhang DB, Liang WQ. The Rice basic helix-loop-helix transcription factor TDR INTERACTING PROTEIN2 is a central switch in early anther development. Plant Cell. 2014;26(4):1512–1524. doi: 10.1105/tpc.114.123745. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 94.Ko SS, Li MJ, Sun-Ben Ku M, Ho YC, Lin YJ, Chuang MH, Hsing HX, Lien YC, Yang HT, Chang HC, et al. The bHLH142 transcription factor coordinates with TDR1 to modulate the expression of EAT1 and regulate pollen development in Rice. Plant Cell. 2014;26(6):2486–2504. doi: 10.1105/tpc.114.126292. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 95.Sorensen AM, Kröber S, Unte US, Huijser P, Dekker K, Saedler H. The Arabidopsis ABORTED MICROSPORES (AMS) gene encodes a MYC class transcription factor. Plant J. 2003;33(2):413–423. doi: 10.1046/j.1365-313X.2003.01644.x. [DOI] [PubMed] [Google Scholar]
- 96.Xu J, Yang C, Yuan Z, Zhang D, Gondwe MY, Ding Z, Liang W, Zhang D, Wilson ZA. The ABORTED MICROSPORES regulatory network is required for postmeiotic male reproductive development in Arabidopsis Thaliana. Plant Cell. 2010;22(1):91–107. doi: 10.1105/tpc.109.071803. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 97.Xu J, Zhang D. ABORTED MICROSPORES acts as a master regulator of Pollen Wall formation in Arabidopsis. Plant Cell. 2014;26(4):1544. doi: 10.1105/tpc.114.122986. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 98.Ferguson AC, Pearce S, Band LR, Yang C, Ferjentsikova I, King J, Yuan Z, Zhang D, Wilson ZA. Biphasic regulation of the transcription factor ABORTED MICROSPORES (AMS) is essential for tapetum and pollen development in Arabidopsis. New Phytol. 2017;213(2):778. doi: 10.1111/nph.14200. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 99.Zhang W, Sun Y, Timofejeva L, Chen C, Grossniklaus U, Ma H. Regulation of Arabidopsis Tapetum development and function by DYSFUNCTIONAL TAPETUM1 (DYT1) encoding a putative bHLH transcription factor. Development. 2006;133(16):3085. doi: 10.1242/dev.02463. [DOI] [PubMed] [Google Scholar]
- 100.Feng B, Lu D, Ma X, Peng Y, Sun Y, Ning G, Ma H. Regulation of the Arabidopsis anther transcriptome by DYT1 for pollen development. Plant J. 2012;72(4):612–624. doi: 10.1111/j.1365-313X.2012.05104.x. [DOI] [PubMed] [Google Scholar]
- 101.Zhang DS, Liang WQ, Yuan Z, Li N, Shi J, Wang J, Liu YM, Yu WJ, Zhang DB. Tapetum degeneration retardation is critical for aliphatic metabolism and gene regulation during Rice pollen development. Mol Plant. 2008;1(4):599–610. doi: 10.1093/mp/ssn028. [DOI] [PubMed] [Google Scholar]
- 102.Li N, Zhang DS, Liu HS, Yin CS, Li XX, Liang WQ, Yuan Z, Xu B, Chu HW, Wang J. The Rice Tapetum degeneration retardation gene is required for Tapetum degradation and anther development. Plant Cell. 2006;18(11):2999. doi: 10.1105/tpc.106.044107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 103.Niu N, Liang W, Yang X, Jin W, Wilson ZA, Hu J, Zhang D. EAT1 promotes tapetal cell death by regulating aspartic proteases during male reproductive development in rice. Nat Commun. 2013;4(2):1445. doi: 10.1038/ncomms2396. [DOI] [PubMed] [Google Scholar]
- 104.Li L, Li YX, Song SF, Deng HF, Li N, Fu XQ, Chen GH, Yuan LP. An anther development F-box (ADF) protein regulated by tapetum degeneration retardation (TDR) controls rice anther development. Planta. 2015;241(1):157–166. doi: 10.1007/s00425-014-2160-9. [DOI] [PubMed] [Google Scholar]
- 105.Jung KH, Han MJ, Lee YS, Kim YW, Hwang IW, Kim MJ, Kim YK, Nahm BH, An GH. Rice undeveloped Tapetum1 is a major regulator of early tapetum development. Plant Cell. 2005;17(10):2705–2722. doi: 10.1105/tpc.105.034090. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 106.Kanaoka MM, Pillitteri LJ, Fujii H, Yoshida Y, Bogenschutz NL, Takabayashi J, Zhu JK, Torii KU. SCREAM/ICE1 and SCREAM2 specify three cell-state transitional steps leading to arabidopsis stomatal differentiation. Plant Cell. 2008;20(7):1775. doi: 10.1105/tpc.108.060848. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 107.Kim YS, Lee M, Lee JH, Lee HJ, Park CM. The unified ICE-CBF pathway provides a transcriptional feedback control of freezing tolerance during cold acclimation in Arabidopsis. Plant Mol Biol. 2015;89(1):187–201. doi: 10.1007/s11103-015-0365-3. [DOI] [PubMed] [Google Scholar]
- 108.Lee BH, Henderson DA, Zhu JK. The Arabidopsis cold-responsive transcriptome and its regulation by ICE1. Plant Cell. 2005;17(11):3155–3175. doi: 10.1105/tpc.105.035568. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 109.Kurbidaeva A, Ezhova T, Novokreshchenova M. Arabidopsis Thaliana ICE2 gene: phylogeny, structural evolution and functional diversification from ICE1. Plant Sci. 2014;229:10–22. doi: 10.1016/j.plantsci.2014.08.011. [DOI] [PubMed] [Google Scholar]
- 110.Nakamura J, Yuasa T, Huong TT, Harano K, Tanaka S, Iwata T, Phan T, Iwaya-Inoue M. Rice homologs of inducer of CBF expression (OsICE) are involved in cold acclimation. Plant Biotechnol. 2011;28(3):303–309. doi: 10.5511/plantbiotechnology.11.0421a. [DOI] [Google Scholar]
- 111.Sharma N, Xin R, Kim DH, Sung S, Lange T, Huq E: NO FLOWERING IN SHORT DAY (NFL) is a bHLH transcription factor that promotes flowering specifically under short-day conditions in Arabidopsis. Development 2016, 143(4):págs. 200501–201100. [DOI] [PMC free article] [PubMed]
- 112.Chen HJ, Chen CL, Hsieh HL. Far-Red Light-Mediated Seedling Development in Arabidopsis Involves FAR-RED INSENSITIVE 219/JASMONATE RESISTANT 1-Dependent and -Independent Pathways. PLoS One. 2015;10(7) doi: 10.1371/journal.pone.0132723. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 113.Miyamoto K, Shimizu T, Mochizuki S, Nishizawa Y, Minami E, Nojiri H, Yamane H, Okada K. Stress-induced expression of the transcription factor RERJ1 is tightly regulated in response to jasmonic acid accumulation in rice. Protoplasma. 2013;250(1):241–249. doi: 10.1007/s00709-012-0400-z. [DOI] [PubMed] [Google Scholar]
- 114.Song S, Qi T, Fan M, Zhang X, Gao H, Huang H, Wu D, Guo H, Xie D. The bHLH subgroup IIId factors negatively regulate Jasmonate-mediated plant defense and development. PLoS Genet. 2012;9(7):e1003653. doi: 10.1371/journal.pgen.1003653. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 115.Fonseca S, Fernández-Calvo P, Fernández GM, Díez-Díaz M, Gimenez-Ibanez S, López-Vidriero I, Godoy M, Fernández-Barbero G, Van LJ, De JG. bHLH003, bHLH013 and bHLH017 are new targets of JAZ repressors negatively regulating JA responses. PLoS One. 2014;9(1):e86182. doi: 10.1371/journal.pone.0086182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 116.Fernández-Calvo P, Solano R. The Arabidopsis bHLH transcription factors MYC3 and MYC4 are targets of JAZ repressors and act additively with MYC2 in the activation of jasmonate responses. Plant Cell. 2011;23(23):701–715. doi: 10.1105/tpc.110.080788. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 117.Sasakisekimoto Y, Jikumaru Y, Obayashi T, Saito H, Masuda S, Kamiya Y, Ohta H, Shirasu K. Basic helix-loop-helix transcription factors JASMONATE-ASSOCIATED MYC2-LIKE1 (JAM1), JAM2, and JAM3 are negative regulators of jasmonate responses in Arabidopsis. Plant Physiol. 2013;163(1):291–304. doi: 10.1104/pp.113.220129. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 118.Dombrecht B, Xue GP, Sprague SJ, Kirkegaard JA, Ross JJ, Reid JB, Fitt GP, Sewelam N, Schenk PM, Manners JM, et al. MYC2 differentially modulates diverse jasmonate-dependent functions in Arabidopsis. Plant Cell. 2007;19(7):2225–2245. doi: 10.1105/tpc.106.048017. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 119.Song S, Huang H, Gao H, Wang J, Wu D, Liu X, et al. Interaction between MYC2 and ETHYLENE INSENSITIVE3 Modulates Antagonism between Jasmonate and Ethylene Signaling in Arabidopsis. Plant Cell. 2014;26(1):263–79. doi: 10.1105/tpc.113.120394. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 120.Ogawa S, Miyamoto K, Nemoto K, Sawasaki T, Yamane H, Nojiri H, Okada K. OsMYC2, an essential factor for JA-inductive sakuranetin production in rice, interacts with MYC2-like proteins that enhance its transactivation ability. Sci Rep. 2017;7:40175. doi: 10.1038/srep40175. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 121.Uji Y, Akimitsu K, Gomi K. Identification of OsMYC2-regulated senescence-associated genes in rice. Planta. 2017;245(6):1241–1246. doi: 10.1007/s00425-017-2697-5. [DOI] [PubMed] [Google Scholar]
- 122.Uji Y, Taniguchi S, Tamaoki D, Shishido H, Akimitsu K, Gomi K. Overexpression of OsMYC2 results in the up-regulation of early JA-Rresponsive genes and bacterial blight resistance in Rice. Plant Cell Physiol. 2016;57(9):1814–1827. doi: 10.1093/pcp/pcw101. [DOI] [PubMed] [Google Scholar]
- 123.Nesi N, Debeaujon I, Jond C, Pelletier G, Caboche M, Lepiniec L. The TT8 gene encodes a basic helix-loop-helix domain protein required for expression of DFR and BAN genes in Arabidopsis siliques. Plant Cell. 2000;12(10):1863–1878. doi: 10.1105/tpc.12.10.1863. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 124.Sakamoto W, Ohmori T, Kageyama K, Miyazaki C, Saito A, Murata M, Noda K, Maekawa M. The purple leaf (Pl) locus of rice: the Pl(w) allele has a complex organization and includes two genes encoding basic helix-loop-helix proteins involved in anthocyanin biosynthesis. Plant Cell Physiol. 2001;42(9):982–991. doi: 10.1093/pcp/pce128. [DOI] [PubMed] [Google Scholar]
- 125.Oikawa T, Maeda H, Oguchi T, Yamaguchi T, Tanabe N, Ebana K, Yano M, Ebitani T, Izawa T. The birth of a black Rice gene and its local spread by introgression. Plant Cell. 2015;27(9):2401–2414. doi: 10.1105/tpc.15.00310. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 126.Sweeney MT, Thomson MJ, Pfeil BE, Mccouch S. Caught red-handed: Rc encodes a basic helix-loop-helix protein conditioning red pericarp in rice. Plant Cell. 2006;18(2):283–294. doi: 10.1105/tpc.105.038430. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 127.Payne CT, Zhang F, Lloyd AM. GL3 encodes a bHLH protein that regulates trichome development in arabidopsis through interaction with GL1 and TTG1. Genetics. 2000;156(3):1349–1362. doi: 10.1093/genetics/156.3.1349. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 128.Bernhardt C, Lee MM, Gonzalez A, Zhang F, Lloyd A, Schiefelbein J. The bHLH genes GLABRA3 (GL3) and ENHANCER OF GLABRA3 (EGL3) specify epidermal cell fate in the Arabidopsis root. Development. 2003;130(26):6431–6439. doi: 10.1242/dev.00880. [DOI] [PubMed] [Google Scholar]
- 129.Bernhardt C, Zhao MZ, Gonzalez A, Lloyd A, Schiefelbein J. The bHLH genes GL3 and EGL3 participate in an intercellular regulatory circuit that controls cell patterning in the Arabidopsis root epidermis. Development. 2005;132(2):291–298. doi: 10.1242/dev.01565. [DOI] [PubMed] [Google Scholar]
- 130.Bruex A, Kainkaryam RM, Wieckowski Y, Kang YH, Bernhardt C, Xia Y, Zheng X, Wang JY, Lee MM, Benfey P, et al. A gene regulatory network for root epidermis cell differentiation in Arabidopsis. PLoS Genet. 2012;8(1):e1002446. doi: 10.1371/journal.pgen.1002446. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 131.Li XL, Zhang HM, Ai Q, Liang G, Yu D. Two bHLH Transcription Factors, bHLH34 and bHLH104, Regulate Iron Homeostasis in Arabidopsis thaliana. Plant Physiol. 2016;170(4):2478–93. doi: 10.1104/pp.15.01827. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 132.Liang G, Zhang HM, Li XL, Ai Q, Yu DQ. bHLH transcription factor bHLH115 regulates iron homeostasis in Arabidopsis Thaliana. J Exp Bot. 2017;68(7):1743–1755. doi: 10.1093/jxb/erx043. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 133.Rampey AWW RA, Hobbs BN, Tierney MP, Lahner B, Salt DE, Bartel B. An Arabidopsis Basic Helix-Loop-Helix Leucine Zipper Protein Modulates Metal Homeostasis and Auxin Conjugate Responsiveness. Genetics. 2006;174(4):1841. doi: 10.1534/genetics.106.061044. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 134.Zheng LQ, Ying YH, Wang L, Wang F, Whelan J, Shou HX. Identification of a novel iron regulated basic helix-loop-helix protein involved in Fe homeostasis in Oryza sativa. BMC Plant Biol. 2010;10:166. doi: 10.1186/1471-2229-10-166. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 135.Jiang Y, Yang B, Deyholos MK. Functional characterization of the Arabidopsis bHLH92 transcription factor in abiotic stress. Mol Gen Genomics. 2009;282(5):503–516. doi: 10.1007/s00438-009-0481-3. [DOI] [PubMed] [Google Scholar]
- 136.Hanano S, Stracke R, Jakoby M, Merkle T, Domagalska MA, Weisshaar B, Davis SJ. A systematic survey in Arabidopsis thaliana of transcription factors that modulate circadian parameters. BMC Genomics. 2008;9:182. doi: 10.1186/1471-2164-9-182. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 137.Yamamura C, Mizutani E, Okada K, Nakagawa H, Fukushima S, Tanaka A, Maeda S, Kamakura T, Yamane H, Takatsuji H, et al. Diterpenoid phytoalexin factor, a bHLH transcription factor, plays a central role in the biosynthesis of diterpenoid phytoalexins in rice. Plant J. 2015;84(6):1100–1113. doi: 10.1111/tpj.13065. [DOI] [PubMed] [Google Scholar]
- 138.Yin Y, Vafeados D, Tao Y, Yoshida S, Asami T, Chory J. A new class of transcription factors mediates brassinosteroid-regulated gene expression in Arabidopsis. Cell. 2005;120(2):249. doi: 10.1016/j.cell.2004.11.044. [DOI] [PubMed] [Google Scholar]
- 139.Cifuentes-Esquivel N, Bou-Torrent J, Galstyan A, Gallemí M, Sessa G, Salla Martret M, Roig-Villanova I, Ruberti I, Martínez-García JF. The bHLH proteins BEE and BIM positively modulate the shade avoidance syndrome in Arabidopsis seedlings. Plant J. 2013;75(6):989–1002. doi: 10.1111/tpj.12264. [DOI] [PubMed] [Google Scholar]
- 140.De RB, Möller B, Yoshida S, Grabowicz I, dRP B, Boeren S, Smith RS, Borst JW, Weijers D. A bHLH complex controls embryonic vascular tissue establishment and indeterminate growth in Arabidopsis. Dev Cell. 2013;24(4):426. doi: 10.1016/j.devcel.2012.12.013. [DOI] [PubMed] [Google Scholar]
- 141.Vera-Sirera F, De RB, Úrbez C, Kouklas E, Pesquera M, Álvarez-Mahecha JC, Minguet EG, Tuominen H, Carbonell J, Borst JW. A bHLH-based feedback loop restricts vascular cell proliferation in plants. Dev Cell. 2015;35(4):432. doi: 10.1016/j.devcel.2015.10.022. [DOI] [PubMed] [Google Scholar]
- 142.Ahmad A, Niwa Y, Goto S, Ogawa T, Shimizu M, Suzuki A, Kobayashi K, Kobayashi H. bHLH106 integrates functions of multiple genes through their G-box to confer salt tolerance on Arabidopsis. PLoS One. 2015;10(5):e0126872. doi: 10.1371/journal.pone.0126872. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 143.Pagnussat GC, Yu HJ, Ngo QA, Rajani S, Mayalagu S, Johnson CS, Capron A, Xie LF, Ye D, Sundaresan V. Genetic and molecular identification of genes required for female gametophyte development and function in Arabidopsis (vol 132, pg 603, 2005) Development. 2005;132(5):1161–1162. doi: 10.1242/dev.01595. [DOI] [PubMed] [Google Scholar]
- 144.Ni M, Tepperman JM, Quail PH. PIF3, a phytochrome-interacting factor necessary for normal photoinduced signal transduction, is a novel basic helix-loop-helix protein. Cell. 1998;95(5):657–667. doi: 10.1016/S0092-8674(00)81636-0. [DOI] [PubMed] [Google Scholar]
- 145.Huq E, Quail PH. PIF4, a phytochrome-interacting bHLH factor, functions as a negative regulator of phytochrome B signaling in Arabidopsis. EMBO J. 2002;21(10):2441–2450. doi: 10.1093/emboj/21.10.2441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 146.Hornitschek P, Kohnen MV, Lorrain S, Rougemont J, Ljung K, Lopez-Vidriero I, Franco-Zorrilla JM, Solano R, Trevisan M, Pradervand S, et al. Phytochrome interacting factors 4 and 5 control seedling growth in changing light conditions by directly controlling auxin signaling. Plant J. 2012;71(5):699–711. doi: 10.1111/j.1365-313X.2012.05033.x. [DOI] [PubMed] [Google Scholar]
- 147.Casson SA, Franklin KA, Gray JE, Grierson CS, Whitelam GC, Hetherington AM. Phytochrome B and PIF4 regulate stomatal development in response to light quantity. Comp Biochem Phys A. 2009;153A(2):S209–SS20. doi: 10.1016/j.cbpa.2009.04.579. [DOI] [PubMed] [Google Scholar]
- 148.Huq E, Alsady B, Hudson M, Kim C, Apel K, Quail PH. Phytochrome-interacting factor 1 is a critical bHLH regulator of chlorophyll biosynthesis. Science. 2004;305(5692):1937. doi: 10.1126/science.1099728. [DOI] [PubMed] [Google Scholar]
- 149.Oh E, Kang H, Yamaguchi S, Park J, Lee D, Kamiya Y, Choi G. Genome-wide analysis of genes targeted by PHYTOCHROME INTERACTING FACTOR 3-LIKE5 during seed germination in Arabidopsis. Plant Cell. 2009;21(2):403–419. doi: 10.1105/tpc.108.064691. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 150.Khanna R, Huq E, Kikis EA, Al-Sady B, Lanzatella C, Quail PH. A novel molecular recognition motif necessary for targeting Photoactivated Phytochrome signaling to specific basic helix-loop-helix transcription factors. Plant Cell. 2004;16(11):3033–3044. doi: 10.1105/tpc.104.025643. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 151.Luo Q, Lian HL, He SB, Li L, Jia KP, Yang HQ. COP1 and phyB physically interact with PIL1 to regulate its stability and Photomorphogenic development in Arabidopsis. Plant Cell. 2014;26(6):2441–2456. doi: 10.1105/tpc.113.121657. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 152.Li L, Zhang Q, Pedmale UV, Nito K, Fu W, Lin L, Hazen SP, Chory J. PIL1 participates in a negative feedback loop that regulates its own gene expression in response to shade (vol 7, pg 1582, 2014) Mol Plant. 2014;7(12):1805–1806. doi: 10.1093/mp/ssu128. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 153.Hwang YS, Quail PH. Phytochrome-regulated PIL1 derepression is developmentally modulated. Plant Cell Physiol. 2008;49(4):501–511. doi: 10.1093/pcp/pcn024. [DOI] [PubMed] [Google Scholar]
- 154.Roig-Villanova I, Bou J, Sorin C, Devlin PF, Martinez-Garcia JF. Identification of primary target genes of phytochrome signaling. Early transcriptional control during shade avoidance responses in Arabidopsis. Plant Physiol. 2006;141(1):85–96. doi: 10.1104/pp.105.076331. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 155.Ochoa-Fernandez R, Samodelov SL, Brandl SM, Wehinger E, Muller K, Weber W, Zurbriggen MD. Optogenetics in plants: red/far-red light control of gene expression. Methods Mol Biol. 2016;1408:125–139. doi: 10.1007/978-1-4939-3512-3_9. [DOI] [PubMed] [Google Scholar]
- 156.Nakamura Y, Kato T, Yamashino T, Murakami M, Mizuno T. Characterization of a set of phytochrome-interacting factor-like bHLH proteins in Oryza Sativa. Biosci Biotechnol Biochem. 2007;71(5):1183–1191. doi: 10.1271/bbb.60643. [DOI] [PubMed] [Google Scholar]
- 157.Zhu Y, Cai XL, Wang ZY, Hong MM. An interaction between a MYC protein and an EREBP protein is involved in transcriptional regulation of the rice Wx gene. J Biol Chem. 2003;278(48):47803–47811. doi: 10.1074/jbc.M302806200. [DOI] [PubMed] [Google Scholar]
- 158.Zhou J, Liu Q, Zhang F, Wang Y, Zhang S, Cheng H, Yan L, Li L, Chen F, Xie X. Overexpression of OsPIL15, a phytochrome-interacting factor-like protein gene, represses etiolated seedling growth in rice. J Integr Plant Biol. 2014;56(4):373–387. doi: 10.1111/jipb.12137. [DOI] [PubMed] [Google Scholar]
- 159.Zhao XL, Shi ZY, Peng LT, Shen GZ, Zhang JL. An atypical HLH protein OsLF in rice regulates flowering time and interacts with OsPIL13 and OsPIL15. New Biotechnol. 2011;28(6):788–797. doi: 10.1016/j.nbt.2011.04.006. [DOI] [PubMed] [Google Scholar]
- 160.Heang D, Sassa H. Antagonistic Actions of HLH/bHLH Proteins Are Involved in Grain Length and Weight in Rice. PLoS One. 2012;7(2) doi: 10.1371/journal.pone.0031325. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 161.Todaka D, Nakashima K, Maruyama K, Kidokoro S, Osakabe Y, Ito Y, Matsukura S, Fujita Y, Yoshiwara K, Ohme-Takagi M, et al. Rice phytochrome-interacting factor-like protein OsPIL1 functions as a key regulator of internode elongation and induces a morphological response to drought stress. Proc Natl Acad Sci U S A. 2012;109(39):15947–15952. doi: 10.1073/pnas.1207324109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 162.Josse EM, Gan YB, Bou-Torrent J, Stewart KL, Gilday AD, Jeffree CE, Vaistij FE, Martinez-Garcia JF, Nagy F, Graham IA, et al. A DELLA in disguise: SPATULA restrains the growth of the developing Arabidopsis seedling. Plant Cell. 2011;23(4):1337–1351. doi: 10.1105/tpc.110.082594. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 163.Makkena S, Lamb RS. The bHLH transcription factor SPATULA regulates root growth by controlling the size of the root meristem. BMC Plant Biol. 2013;13:1. doi: 10.1186/1471-2229-13-1. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 164.Leivar P, Quail PH. PIFs: pivotal components in a cellular signaling hub. Trends Plant Sci. 2011;16(1):19–28. doi: 10.1016/j.tplants.2010.08.003. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 165.Fairchild CD, Schumaker MA, Quail PH. HFR1 encodes an atypical bHLH protein that acts in phytochrome a signal transduction. Genes Dev. 2000;14(18):2377–2391. [PMC free article] [PubMed] [Google Scholar]
- 166.Fankhauser C, Chory J. RSF1, an Arabidopsis locus implicated in phytochrome a signaling. Plant Physiol. 2000;124(1):39–45. doi: 10.1104/pp.124.1.39. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 167.Leivar P, Monte E, Al-Sady B, Carle C, Storer A, Alonso JM, Ecker JR, Quail PH. The Arabidopsis Phytochrome-interacting factor PIF7, together with PIF3 and PIF4, regulates responses to prolonged red light by modulating phyB levels. Plant Cell. 2008;20(2):337–352. doi: 10.1105/tpc.107.052142. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 168.Hao YQ, Oh E, Choi G, Liang ZS, Wang ZY. Interactions between HLH and bHLH factors modulate light-regulated plant development. Mol Plant. 2012;5(3):688–697. doi: 10.1093/mp/sss011. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 169.Galstyan A, Cifuentesesquivel N, Boutorrent J, Martinezgarcia JF. The shade avoidance syndrome in Arabidopsis: a fundamental role for atypical basic helix-loop-helix proteins as transcriptional cofactors. Plant J. 2011;66(2):258. doi: 10.1111/j.1365-313X.2011.04485.x. [DOI] [PubMed] [Google Scholar]
- 170.Bou-Torrent J, Roig-Villanova I, Galstyan A, Martínez-García JF. PAR1 and PAR2 integrate shade and hormone transcriptional networks. Plant Signal Behav. 2008;3(7):453–454. doi: 10.4161/psb.3.7.5599. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 171.Zhou P, Song MF, Yang QH, Su L, Hou P, Guo L, Zheng X, Xi YL, Meng FH, Xiao Y, et al. Both PHYTOCHROME RAPIDLY REGULATED1 (PAR1) and PAR2 promote seedling Photomorphogenesis in multiple light signaling pathways. Plant Physiol. 2014;164(2):841–852. doi: 10.1104/pp.113.227231. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 172.Gremski K, Ditta G, Yanofsky MF. The HECATE genes regulate female reproductive tract development in Arabidopsis Thaliana. Development. 2007;134(20):3593–3601. doi: 10.1242/dev.011510. [DOI] [PubMed] [Google Scholar]
- 173.Liljegren SJ, Roeder AHK, Kempin SA, Gremski K, Ostergaard L, Guimil S, Reyes DK, Yanofsky MF. Control of fruit patterning in Arabidopsis by INDEHISCENT. Cell. 2004;116(6):843–853. doi: 10.1016/S0092-8674(04)00217-X. [DOI] [PubMed] [Google Scholar]
- 174.Yang F, Wang Q, Schmitz G, Muller D, Theres K. The bHLH protein ROX acts in concert with RAX1 and LAS to modulate axillary meristem formation in Arabidopsis. Plant J. 2012;71(1):61–70. doi: 10.1111/j.1365-313X.2012.04970.x. [DOI] [PubMed] [Google Scholar]
- 175.Li JZ, Han YC, Liu L, Chen YP, Du YX, Zhang J, Sun HZ, Zhao QZ. qRT9, a quantitative trait locus controlling root thickness and root length in upland rice. J Exp Bot. 2015;66(9):2723–2732. doi: 10.1093/jxb/erv076. [DOI] [PubMed] [Google Scholar]
- 176.Wang J, Hu J, Qian Q, Xue HW. LC2 and OsVIL2 promote Rice flowering by Photoperoid-induced epigenetic silencing of OsLF. Mol Plant. 2013;6(2):514–527. doi: 10.1093/mp/sss096. [DOI] [PubMed] [Google Scholar]
- 177.Oikawa T, Kyozuka J. Two-step regulation of LAX PANICLE1 protein accumulation in axillary meristem formation in Rice. Plant Cell. 2009;21(4):1095–1108. doi: 10.1105/tpc.108.065425. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 178.Komatsu K, Maekawa M, Ujiie S, Satake Y, Furutani I, Okamoto H, Shimamoto K, Kyozuka J. LAX and SPA: major regulators of shoot branching in rice. Proc Natl Acad Sci U S A. 2003;100(20):11765–11770. doi: 10.1073/pnas.1932414100. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 179.Komatsu M, Maekawa M, Shimamoto K, Kyozuka J. The LAX1 and FRIZZY PANICLE 2 genes determine the inflorescence architecture of rice by controlling rachis-branch and spikelet development. Dev Biol. 2001;231(2):364–373. doi: 10.1006/dbio.2000.9988. [DOI] [PubMed] [Google Scholar]
- 180.Masucci JD, Schiefelbein JW. The rhd6 mutation of Arabidopsis Thaliana alters root-hair initiation through an auxin- and ethylene-associated process. Plant Physiol. 1994;106(4):1335. doi: 10.1104/pp.106.4.1335. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 181.Vijayakumar P, Datta S, Dolan L. ROOT HAIR DEFECTIVE SIX-LIKE4 (RSL4) promotes root hair elongation by transcriptionally regulating the expression of genes required for cell growth. New Phytol. 2016;212(4):944–953. doi: 10.1111/nph.14095. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 182.Wang L, Ying YH, Narsai R, Ye LX, Zheng LQ, Tian JL, Whelan J, Shou HX. Identification of OsbHLH133 as a regulator of iron distribution between roots and shoots in Oryza Sativa. Plant Cell Environ. 2013;36(1):224–236. doi: 10.1111/j.1365-3040.2012.02569.x. [DOI] [PubMed] [Google Scholar]
- 183.Ito S, Song YH, Josephson-Day AR, Miller RJ, Breton G, Olmstead RG, Imaizumi T. FLOWERING BHLH transcriptional activators control expression of the photoperiodic flowering regulator CONSTANS in Arabidopsis. Proc Natl Acad Sci U S A. 2012;109(9):3582–3587. doi: 10.1073/pnas.1118876109. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 184.Takahashi Y, Ebisu Y, Kinoshita T, Doi M, Okuma E, Murata Y, Shimazaki K. bHLH transcription factors that facilitate K+ uptake during stomatal opening are repressed by abscisic acid through phosphorylation. Sci Signal. 2013;6(280):ra48. doi: 10.1126/scisignal.2003760. [DOI] [PubMed] [Google Scholar]
- 185.Le Hir R, Castelain M, Chakraborti D, Moritz T, Dinant S, Bellini C. AtbHLH68 transcription factor contributes to the regulation of ABA homeostasis and drought stress tolerance in Arabidopsis thaliana. Physiol Plant. 2017;160(3):312–27. doi: 10.1111/ppl.12549. [DOI] [PubMed] [Google Scholar]
- 186.Liu YJ, Ji XY, Nie XG, Qu M, Zheng L, Tan ZL, Zhao HM, Huo L, Liu SN, Zhang B, et al. Arabidopsis AtbHLH112 regulates the expression of genes involved in abiotic stress tolerance by binding to their E-box and GCG-box motifs. New Phytol. 2015;207(3):692–709. doi: 10.1111/nph.13387. [DOI] [PubMed] [Google Scholar]
- 187.Karas B, Amyot L, Johansen C, Sato S, Tabata S, Kawaguchi M, Szczyglowski K. Conservation of lotus and Arabidopsis basic helix-loop-helix proteins reveals new players in root hair development. Plant Physiol. 2009;151(3):1175–1185. doi: 10.1104/pp.109.143867. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 188.Breuninger H, Thamm A, Streubel S, Sakayama H, Nishiyama T, Dolan L. Diversification of a transcription factor family led to the evolution of antagonistically acting genetic regulators of root hair growth. Curr Biol. 2016;26(12):1622–1628. doi: 10.1016/j.cub.2016.04.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 189.Ding WN, Yu ZM, Tong YL, Huang W, Chen HM, Wu P. A transcription factor with a bHLH domain regulates root hair development in rice. Cell Res. 2009;19(11):1309–1311. doi: 10.1038/cr.2009.109. [DOI] [PubMed] [Google Scholar]
- 190.Yi K, Wu Z, Zhou J, Du L, Guo L, Wu Y, Wu P. OsPTF1, a novel transcription factor involved in tolerance to phosphate starvation in rice. Plant Physiol. 2005;138(4):2087–2096. doi: 10.1104/pp.105.063115. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 191.Friedrichsen DM, Nemhauser J, Muramitsu T, Maloof JN, Alonso J, Ecker JR, Furuya M, Chory J. Three redundant Brassinosteroid early response genes encode putative bHLH transcription factors required for normal growth. Genetics. 2002;162(3):1445. doi: 10.1093/genetics/162.3.1445. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 192.Poppenberger B, Rozhon W, Khan M, Husar S, Adam G, Luschnig C, Fujioka S, Sieberer T. CESTA, a positive regulator of brassinosteroid biosynthesis. EMBO J. 2011;30(6):1149–1161. doi: 10.1038/emboj.2011.35. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 193.Eremina M, Unterholzner SJ, Rathnayake AI, Castellanos M, Khan M, Kugler KG, May ST, Mayer KFX, Rozhon W, Poppenberger B. Brassinosteroids participate in the control of basal and acquired freezing tolerance of plants. Proc Natl Acad Sci U S A. 2016;113(40):E5982–E5991. doi: 10.1073/pnas.1611477113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 194.Petridis A, Döll S, Nichelmann L, Bilger W, Mock HP. Arabidopsis Thaliana G2-LIKE FLAVONOID REGULATOR and BRASSINOSTEROID ENHANCED EXPRESSION1 are low-temperature regulators of flavonoid accumulation. New Phytol. 2016;211(3):912. doi: 10.1111/nph.13986. [DOI] [PubMed] [Google Scholar]
- 195.Malinovsky FG, Batoux M, Schwessinger B, Youn JH, Stransfeld L, Win J, Kim SK, Zipfel C. Antagonistic regulation of growth and immunity by the Arabidopsis basic helix-loop-helix transcription factor HOMOLOG OF BRASSINOSTEROID ENHANCED EXPRESSION2 INTERACTING WITH INCREASED LEAF INCLINATION1 BINDING bHLH1. Plant Physiol. 2014;164(3):1443–1455. doi: 10.1104/pp.113.234625. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 196.Liu Y, Li X, Li K, Liu H, Lin C. Multiple bHLH proteins form heterodimers to mediate CRY2-dependent regulation of flowering-time in Arabidopsis. PLoS Genet. 2013;9(10):e1003861. doi: 10.1371/journal.pgen.1003861. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 197.Ikeda M, Fujiwara S, Mitsuda N, Ohme-Takagi M. A Triantagonistic basic helix-loop-helix system regulates cell elongation in Arabidopsis. Plant Cell. 2012;24(11):4483–4497. doi: 10.1105/tpc.112.105023. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 198.Liu HT, Yu XH, Li KW, Klejnot J, Yang HY, Lisiero D, Lin CT. Photoexcited CRY2 interacts with CIB1 to regulate transcription and floral initiation in Arabidopsis. Science. 2008;322(5907):1535–1539. doi: 10.1126/science.1163927. [DOI] [PubMed] [Google Scholar]
- 199.Luo JH, Liu H, Zhou TY, Gu BG, Huang XH, Shangguan YY, Zhu JJ, Li Y, Zhao Y, Wang YC, et al. An-1 encodes a basic helix-loop-helix protein that regulates awn development, grain size, and grain number in Rice. Plant Cell. 2013;25(9):3360–3376. doi: 10.1105/tpc.113.113589. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 200.Tsukagoshi H, Busch W, Benfey PN. Transcriptional regulation of ROS controls transition from proliferation to differentiation in the root. Cell. 2010;143(4):606–616. doi: 10.1016/j.cell.2010.10.020. [DOI] [PubMed] [Google Scholar]
- 201.Ohashiito K, Bergmann DC. Regulation of the Arabidopsis root vascular initial population by LONESOME HIGHWAY. Development. 2007;134(16):2959–2968. doi: 10.1242/dev.006296. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 202.Li J, Dukowic-Schulze S, Lindquist IE, Farmer AD, Kelly B, Li T, Smith AG, Retzel EF, Mudge J, Chen C. The plant-specific protein FEHLSTART controls male meiotic entry, initializing meiotic synchronization in Arabidopsis. Plant J. 2015;84(4):659–671. doi: 10.1111/tpj.13026. [DOI] [PubMed] [Google Scholar]
- 203.Ohashi-Ito K, Matsukawa M, Fukuda H. An atypical bHLH transcription factor regulates early xylem development downstream of auxin. Plant Cell Physiol. 2013;54(3):398–405. doi: 10.1093/pcp/pct013. [DOI] [PubMed] [Google Scholar]
- 204.Cai Q, Fukushima H, Yamamoto M, Ishii N, Sakamoto T, Kurata T, Motose H, Takahashi T. The SAC51 family plays a central role in Thermospermine responses in Arabidopsis. Plant Cell Physiol. 2016;57(8):1583–1592. doi: 10.1093/pcp/pcw113. [DOI] [PubMed] [Google Scholar]
- 205.Yamamoto M, Takahashi T. Thermospermine enhances translation of SAC51 and SACL1 in Arabidopsis. Plant Signal Behav. 2017;12(1):e1276685. doi: 10.1080/15592324.2016.1276685. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 206.Katayama H, Iwamoto K, Kariya Y, Asakawa T, Kan T, Fukuda H, Ohashi-Ito K. A negative feedback loop controlling bHLH complexes is involved in vascular cell division and differentiation in the root apical meristem. Curr Biol. 2015;25(23):3144–3150. doi: 10.1016/j.cub.2015.10.051. [DOI] [PubMed] [Google Scholar]
- 207.Mara CD, Huang TB, Irish VF. The Arabidopsis floral homeotic proteins APETALA3 and PISTILLATA negatively regulate the BANQUO genes implicated in light signaling. Plant Cell. 2010;22(3):690. doi: 10.1105/tpc.109.065946. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 208.Lee S. Overexpression of PRE1 and its homologous genes activates gibberellin-dependent responses in Arabidopsis Thaliana. Plant Cell Physiol. 2006;47(5):591. doi: 10.1093/pcp/pcj026. [DOI] [PubMed] [Google Scholar]
- 209.Castelain M, Le Hir R, Bellini C. The non-DNA-binding bHLH transcription factor PRE3/bHLH135/ATBS1/TMO7 is involved in the regulation of light signaling pathway in Arabidopsis. Physiol Plant. 2012;145(3):450–460. doi: 10.1111/j.1399-3054.2012.01600.x. [DOI] [PubMed] [Google Scholar]
- 210.Wang H, Zhu Y, Fujioka S, Asami T, Li J, Li J. Regulation of Arabidopsis brassinosteroid signaling by atypical basic helix-loop-helix proteins. Plant Cell. 2009;21(12):3781–3791. doi: 10.1105/tpc.109.072504. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 211.Hyun Y, Lee I. KIDARI, encoding a non-DNA binding bHLH protein, represses light signal transduction in Arabidopsis Thaliana. Plant Mol Biol. 2006;61(1):283–296. doi: 10.1007/s11103-006-0010-2. [DOI] [PubMed] [Google Scholar]
- 212.Tanaka A, Nakagawa H, Tomita C, Shimatani Z, Ohtake M, Nomura T, Jiang CJ, Dubouzet JG, Kikuchi S, Sekimoto H, et al. BRASSINOSTEROID UPREGULATED1, encoding a helix-loop-helix protein, is a novel gene involved in Brassinosteroid signaling and controls bending of the lamina joint in Rice. Plant Physiol. 2009;151(2):669–680. doi: 10.1104/pp.109.140806. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 213.Zhang LY, Bai MY, Wu J, Zhu JY, Wang H, Zhang Z, Wang W, Sun Y, Zhao J, Sun X. Antagonistic HLH/bHLH transcription factors mediate brassinosteroid regulation of cell elongation and plant development in rice and Arabidopsis. Plant Cell. 2009;21(12):3767–3780. doi: 10.1105/tpc.109.070441. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 214.Heang D, Sassa H. An atypical bHLH protein encoded by POSITIVE REGULATOR OF GRAIN LENGTH 2 is involved in controlling grain length and weight of rice through interaction with a typical bHLH protein APG. Breeding Sci. 2012;62(2):133–141. doi: 10.1270/jsbbs.62.133. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 215.Menand B, Yi K, Jouannic S, Hoffmann L, Ryan E, Linstead P, Schaefer DG, Dolan L. An ancient mechanism controls the development of cells with a rooting function in land plants. Science. 2007;316(5830):1477–1480. doi: 10.1126/science.1142618. [DOI] [PubMed] [Google Scholar]
- 216.Yi K, Menand B, Bell E, Dolan L. A basic helix-loop-helix transcription factor controls cell growth and size in root hairs. Nat Genet. 2010;42(3):264–267. doi: 10.1038/ng.529. [DOI] [PubMed] [Google Scholar]
- 217.Rymen B, Kawamura A, Schäfer S, Breuer C, Iwase A, Shibata M, Ikeda M, Mitsuda N, Koncz C, Ohmetakagi M. ABA suppresses root hair growth via the OBP4 transcriptional regulator. Plant Physiol. 2017;173(3):1750–1762. doi: 10.1104/pp.16.01945. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 218.Min KC, Liam D. ROOT HAIR DEFECTIVE SIX-LIKE class I genes promote root hair development in the GrassBrachypodium distachyon. PLoS Genet. 2016;12(8):e1006211. doi: 10.1371/journal.pgen.1006211. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 219.Raissig MT, Abrash E, Bettadapur A, Vogel JP, Bergmann DC. Grasses use an alternatively wired bHLH transcription factor network to establish stomatal identity. Proc Natl Acad Sci U S A. 2016;113(29):201606728. doi: 10.1073/pnas.1606728113. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 220.Rouster J, Leah R, Mundy J, Cameron-Mills V. Identification of a methyl jasmonate-responsive region in the promoter of a lipoxygenase 1 gene expressed in barley grain. Plant J. 1997;11(3):513–523. doi: 10.1046/j.1365-313X.1997.11030513.x. [DOI] [PubMed] [Google Scholar]
- 221.Yamaguchishinozaki K, Shinozaki K. A novel cis-acting element in an Arabidopsis gene is involved in responsiveness to drought, low-temperature, or high-salt stress. Plant Cell. 1994;6(2):251–264. doi: 10.1105/tpc.6.2.251. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 222.Storozhenko S, Pauw PD, Montagu MV, Inzé D, Kushnir S. The heat-shock element is a functional component of the Arabidopsis APX1 gene promoter. Plant Physiol. 1998;118(3):1005–1014. doi: 10.1104/pp.118.3.1005. [DOI] [PMC free article] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.
Supplementary Materials
Consensus motifs of bHLH domains. Table S2. The primers used in the qRT-PCR. Table S3. Characteristic features of bHLH Transcription factor gene family identified in Brachypodium distachyon. Table S4. The Ka/Ks ratios and estimated divergence time for tandemly duplicated BdbHLH genes. Table S5. The Ka/Ks ratios and estimated divergence time for segmentally duplicated BdbHLH genes. Table S6. The Ka/Ks ratios and estimated divergence time for orthologous bHLH genes between Brachypodium distachyon and rice. Table S7. The Ka/Ks ratios and estimated divergence time for orthologous bHLH genes between Brachypodium distachyon and maize. Table S8. The Ka/Ks ratios and estimated divergence time for orthologous bHLH genes between Brachypodium distachyon and sorghum. Table S9. GO annotations of BdbHLHs. Table S10. GO descriptions of BdbHLHs. Table S11. Detailed information of interaction network of BdbHLH with other proteins. Table S12. Predicted DNA Binding Characteristics of the bHLH Domain in BdbHLH Proteins. Table S13. Summary of the predicted functions of BdbHLHs by comparative analysis with homologous bHLHs in Arabidopsis and rice. Table S14. The cis-regulatory elements in the promoter region of 29 BdbHLH genes. (XLSX 237 kb)
GO annotations of BdbHLHs. Figure S2. Phylogenic tree using alignments of bHLH proteins in Brachypodium distachyon, Arabidopsis and rice. Figure S3. Conserved elements and gene structures of BdbHLHs. (RAR 22526 kb)
The sequences of all the BdbHLH genes, including CDS, amino acids of the bHLH domains, protein and genomic DNA as well as 1500 bp upstream sequences. (RAR 398 kb)
Data Availability Statement
The genome, cds, cDNA and protein sequences of bHLHs in Arabidopsis, rice and Brachypodium distachyon were obtained from the National Center of Biotechnology Information database (http://www.ncbi.nlm.nih.gov) and the Gramene database (http://www.gramene.org/) and were listed in Additional file 3. The phylogenetic trees of BdbHLHs and AtbHLHs, OsbHLHs and BdbHLHs generated in this study have been uploaded to TreeBASE with accession number NO. S21381. The access URL is (http://purl.org/phylo/treebase/phylows/study/TB2:S21381).