

ABSTRACT
During infection, Neisseria gonorrhoeae senses and responds to stress; such responses may be modulated by MisRS (NGO0177 and NGO0176), a two-component system that is a homolog of CpxRA. In Escherichia coli, CpxRA senses and responds to envelope stress; CpxA is a sensor kinase/phosphatase for CpxR, a response regulator. When a cpxA mutant is grown in medium containing glucose, CpxR is phosphorylated by acetyl phosphate but cannot be dephosphorylated, resulting in constitutive activation. Kandler and coworkers (J. L. Kandler, C. L. Holley, J. L. Reimche, V. Dhulipala, J. T. Balthazar, A. Muszyński, R. W. Carlson, and W. M. Shafer, Antimicrob Agents Chemother 60:4690–4700, 2016, https://doi.org/10.1128/AAC.00823-16) showed that MisR (CpxR) is required for the maintenance of membrane integrity and resistance to antimicrobial peptides, suggesting a role in gonococcal survival in vivo. Here, we evaluated the contributions of MisR and MisS (CpxA) to gonococcal infection in a murine model of cervicovaginal colonization and identified MisR-regulated genes using RNA sequencing (RNA-Seq). The deletion of misR or misS severely reduced the capacity of N. gonorrhoeae to colonize mice or maintain infection over a 7-day period and reduced microbial fitness after exposure to heat shock. Compared to the wild type (WT), the inactivation of misR identified 157 differentially regulated genes, most of which encoded putative envelope proteins. The inactivation of misS identified 17 differentially regulated genes compared to the WT and 139 differentially regulated genes compared to the misR mutant, 111 of which overlapped those differentially expressed in the comparison of the WT versus the misR mutant. These data indicate that an intact MisRS system is required for gonococcal infection of mice. Provided the MisR is constitutively phosphorylated in the misS mutant, the data suggest that controlled but not constitutive activation is required for gonococcal infection in mice.
KEYWORDS: CpxRA, envelope stress, genes, infection, mice, MisRS, Neisseria gonorrhoeae, RNA-Seq
INTRODUCTION
Neisseria gonorrhoeae is the causative agent of gonorrhea, a sexually transmitted infection that is estimated to affect 820,000 Americans every year (1). Gonorrhea is a localized infection of the lower genital tract that primarily manifests as urethritis in men and cervicitis in women (2). A large proportion of gonococcal infections are asymptomatic, particularly in women, and therefore go undetected (2). If undetected or not appropriately treated, cervical infections can spread to the upper genital tract and cause pelvic inflammatory disease, resulting in infertility or ectopic pregnancy (2). In addition to genital tract infections, N. gonorrhoeae causes rectal and pharyngeal infections, which are frequently reported in men who have sex with men (2). Although N. gonorrhoeae is primarily associated with localized infections, systemic infections characterized by arthritis, dermatitis, and, rarely, endocarditis occur (2). Besides causing its own morbidities, N. gonorrhoeae facilitates the acquisition and transmission of human immunodeficiency virus type 1 (3–5). The high public health burden of gonorrhea is exacerbated by the lack of a vaccine and the emergence and spread of antimicrobial-resistant gonococcal strains (6, 7).
The success of N. gonorrhoeae as a human pathogen is in part attributed to its large repertoire of virulence and immune evasion mechanisms. These mechanisms can be broadly classified into four categories, those required for (i) attachment, (ii) local invasion, (iii) dissemination, and (iv) evasion of host immunity (8–10). After acquisition by sexual contact, the gonococcus attaches to genital epithelial cells; attachment is mediated by several bacterial factors, including type IV pili, opacity-associated (Opa) proteins, PorB, lipooligosaccharide (LOS), and outer membrane protein A (11–17). The gonococcus subsequently invades and survives and replicates within host cells, such as epithelial cells, neutrophils, and macrophages (13, 14, 18–21). Some gonococcal strains are intrinsically serum sensitive but become serum resistant by incorporating sialic acid into their LOS, which promotes the binding of factor H, an inhibitor of the alternative complement pathway (22, 23). Other gonococcal strains are intrinsically serum resistant because their porins bind C4B-binding protein and factor H; both mechanisms play a role in gonococcal dissemination (24, 25). The gonococcus also uses a variety of other mechanisms to evade the immune system, including antigenic and phase variation of Opa, pili, and LOS; molecular mimicry of human antigens by terminal LOS sugars; secretion of IgA1 protease; and prevention of the formation of phagolysosomes within neutrophils (26–32). Thus, to establish successful infection, the gonococcus must coordinate the expressions of various virulence/immune evasion factors at optimum levels and at the right time and place during infection.
In addition to coordinating virulence gene expression, the gonococcus must adapt to a variety of stresses and microenvironments in the host. For example, the gonococcus must contend with oxidative stress (from infiltrating neutrophils, which is a hallmark of symptomatic gonorrhea), nutrient limitation, anaerobiosis, temperature fluctuations, and low pH (9, 18, 33–37). The gonococcus also has to battle various host defense/immune mediators such as cytokines, chemokines, antimicrobial peptides, and free fatty acids (9, 38). Furthermore, the gonococcus must be able to acclimate to different microenvironments provided by different sites of infection such as the pharynx, rectum, cervix, and urethra; adapt to the lower and upper male and female genital tracts; and transition between biofilm and planktonic lifestyles (2, 9, 39). However, with the exception of iron and oxygen/nitrite regulation, little is known about how the gonococcus adapts to different stresses and microenvironments in the host.
Bacteria often use two-component systems (TCSs) to adapt to changing microenvironments. N. gonorrhoeae contains three TCSs: NarPQ, NtrXY, and MisRS (40–42). NarPQ and NtrXY regulate gonococcal adaptation to an anaerobic environment and the expression of respiratory genes, respectively. MisRS was originally characterized in the meningococcus and was named owing to its role in the regulation of the meningococcal LOS inner core structure (43). In the meningococcus, MisRS is required for virulence in a murine model and is involved in the regulation of over 78 genes belonging to various functional categories, such as protein folding and iron assimilation; a MisR-binding sequence was also identified (43–47). In N. gonorrhoeae strain FA19, MisR (NGEG_00293) is required for resistance to cationic antimicrobial peptides (CAMPs) and aminoglycosides as well as the maintenance of membrane integrity (42). Compared to its parent, 95 genes are differentially expressed in the misR mutant; these genes encode proteins involved in membrane permeability and redox control (42). MisRS has homology to multiple TCSs, such as PhoP/PhoQ and CpxRA (42). Based on the function or role of MisRS with respect to the maintenance of membrane integrity and the analysis of MisR-dependent targets, the authors of the latter study opined that MisRS functionally resembles CpxRA (42).
CpxRA is a TCS that regulates the response to envelope stress in Gram-negative bacteria. CpxA is a sensor kinase, and CpxR is its cognate response regulator (48–50). In response to envelope stress, CpxA autophosphorylates and transfers the phosphate group to CpxR. To alleviate envelope stress, phosphorylated CpxR increases the transcription of genes that encode factors involved in envelope protein folding and repair and decreases the expression of a majority of its target genes that encode envelope-localized proteins, perhaps in an effort to reduce nonessential protein traffic across the envelope (48). Consistent with the latter, the activation of CpxRA usually downregulates virulence determinants, which must traverse the envelope (48, 51). In addition to kinase activity, CpxA also possesses phosphatase activity; in the absence of envelope stress, the default state of CpxA is as an active phosphatase. When grown in the presence of glucose in organisms that have an intact Pta-AckA pathway (42), CpxR can accept phosphoryl groups from acetyl phosphate (52); thus, when a cpxA mutant is grown in the presence of glucose, CpxR is not readily dephosphorylated, and the system is constitutively activated (53).
In Haemophilus ducreyi, the constitutive activation of CpxRA through mutation of cpxA repressed the transcription of the majority of its targets that encode putative envelope proteins, including several established virulence determinants (51); accordingly, a cpxA mutant of Haemophilus ducreyi was attenuated for virulence in humans (54). Similarly, a cpxA mutant of Salmonella enterica serovar Typhimurium was attenuated for virulence in a murine oral challenge model (55). While the activation of CpxRA attenuates virulence in the above-mentioned pathogens, the inactivation of this system leads to variable phenotypes in vivo. cpxR mutants of S. Typhimurium and H. ducreyi were as virulent as their parents in murine and human infection models, respectively (55, 56). However, cpxR mutants of uropathogenic E. coli and Citrobacter rodentium were attenuated for virulence in their respective murine models (57, 58).
In the present study, we evaluated the contributions of the CpxR and CpxA homologs MisR and MisS to gonococcal fitness in a murine model and defined MisR- and MisS-dependent genes using RNA sequencing (RNA-Seq). This study was undertaken to address three important hypotheses. Given that a CpxR homolog is essential for the in vivo virulence of some bacterial pathogens, including meningococci, and MisR is needed for CAMP resistance and envelope integrity in the gonococcus (42, 47), we hypothesized that MisR would be required for gonococcal fitness in vivo. As the uncontrolled activation of CpxRA in a cpxA mutant attenuates the virulence of multiple pathogens (54, 55), our second hypothesis was that a gonococcal misS mutant would also be compromised for fitness in vivo. The activation of CpxRA downregulates virulence determinants and other envelope-localized proteins in other organisms (51); thus, our final hypothesis was that the activation of MisRS would downregulate gonococcal genes encoding known virulence determinants and other envelope-localized proteins.
RESULTS
Construction and characterization of misR and misS mutants.
Previous studies suggested that MisRS might play important roles in the pathogenesis of gonococcal infection (42). In FA1090, misR and misS appear to be located in an operon, with misS following misR. To better understand the role of this system in N. gonorrhoeae pathogenesis, we constructed insertionally inactivated mutants of misR and misS. PCR and sequence analysis confirmed that misS was insertionally inactivated by the kanamycin resistance marker and that this insertion was in frame. PCR and sequencing also confirmed that misR was inactivated by the erythromycin resistance gene and that this inactivation likely had a polar effect on the downstream gene misS. Both mutants grew similarly to the parental strain in broth, suggesting that misR and misS mutations had no effect on the growth kinetics of the gonococcus in vitro (see Fig. S1 in the supplemental material). In addition, there were no noticeable differences in the colony sizes of the three strains grown on plates.
MisR and MisS confer fitness and colonization advantages to N. gonorrhoeae during experimental murine genital infection.
To determine if MisR and MisS contributed to gonococcal fitness in vivo, we intravaginally inoculated groups of female BALB/c mice with suspensions containing equal numbers of FA1090 and either FA1090 misS::kan or FA1090 misR::erm bacteria. The relative number of CFU recovered from vaginal swabs (output) for each strain was determined every other day after inoculation for 8 days and normalized to the ratio of mutant to parent bacteria in the inocula (input). The misS mutant was recovered at averages of approximately 900-, 250-, 900-, and 3,000-fold-lower levels than the wild-type (WT) strain on days 2, 4, 6, and 8 postinoculation, respectively (Fig. 1A). Similarly, the misR mutant was recovered at 150-, 700-, 13,000-, and 900-fold-lower levels than the WT strain on days 2, 4, 6, and 8 postinoculation, respectively (Fig. 1B). No misS mutant bacteria were recovered from any mice 8 days after infection (Fig. 1A, open symbols), and no misR mutant bacteria were recovered from any mice 6 to 8 days after infection (Fig. 1B, open symbols). Consistent with the competitive index (CI) values, the average number of mutant CFU recovered from coinfected mice was markedly lower than the number of WT bacteria recovered (Fig. 1C and D). When cocultured in vitro through stationary phase, both the FA1090 misS::kan and FA1090 misR::erm mutants showed no difference in recovery relative to the WT strain (data not shown). Thus, in competition experiments, MisS and MisR conferred a strong fitness advantage to N. gonorrhoeae in the murine model.
FIG 1.

(A and B) Competitive infection of BALB/c mice with the FA1090 and FA1090 misS::kan (A) or the FA1090 and FA1090 misR::erm (B) strains. Solid circles indicate mice from which both WT and mutant bacteria were recovered, and open symbols indicate mice from which only WT bacteria were recovered. The competitive index was calculated as follows: [FA1090 misS::kan or FA1090 misR::erm/FA1090 (output)]/[FA1090 misS::kan or FA1090 misR::erm/FA1090 (input)]. Horizontal bars indicate geometric mean competitive indices. For cultures from which no mutant CFU were recovered, a value of 20 CFU (limit of detection) was used in the calculation. (C and D) Average numbers of mutant CFU and average calculated numbers of WT CFU recovered from coinfected mice over time.
The capacity of the WT strain to fully outcompete the mutant strains in vivo suggested that misS and misR are required for infection; noncompetitive infections are a more stringent test of this hypothesis. Accordingly, to determine whether MisR and MisS are required for N. gonorrhoeae infection in the mouse model, we intravaginally inoculated groups of female BALB/c mice with either FA1090, FA1090 misS::kan, or FA1090 misR::erm bacteria and quantitatively cultured vaginal swabs for N. gonorrhoeae over an 8-day period. Comparison of the percentages of mice colonized with each strain showed that the duration of infection was significantly shorter for the misR and misS mutants than for the parent strain (P = 0.002 and 0.004, respectively, as determined by a Mantel-Cox test) (Fig. 2A). Only 25% and 10% of mice inoculated with misS or misR mutant bacteria, respectively, remained infected at day 6 postinoculation; in contrast, 62% and 50% of mice inoculated with the WT strain were infected for 6 or 8 days, respectively. We also compared the numbers of CFU from mice that were colonized throughout the study period. These data showed a significantly lower colonization load for the misS and misR mutants than for the WT at all time points postinoculation (P = 0.005 and 0.003, respectively, as determined by 2-way analysis of variance [ANOVA] with Bonferroni's multiple-comparison test) (Fig. 2B). The demonstration that the misS and misR mutants are significantly attenuated relative to WT bacteria in noncompetitive infections strongly supports the importance of the MisS and MisR proteins in this surrogate animal model of female genital tract infection.
FIG 2.
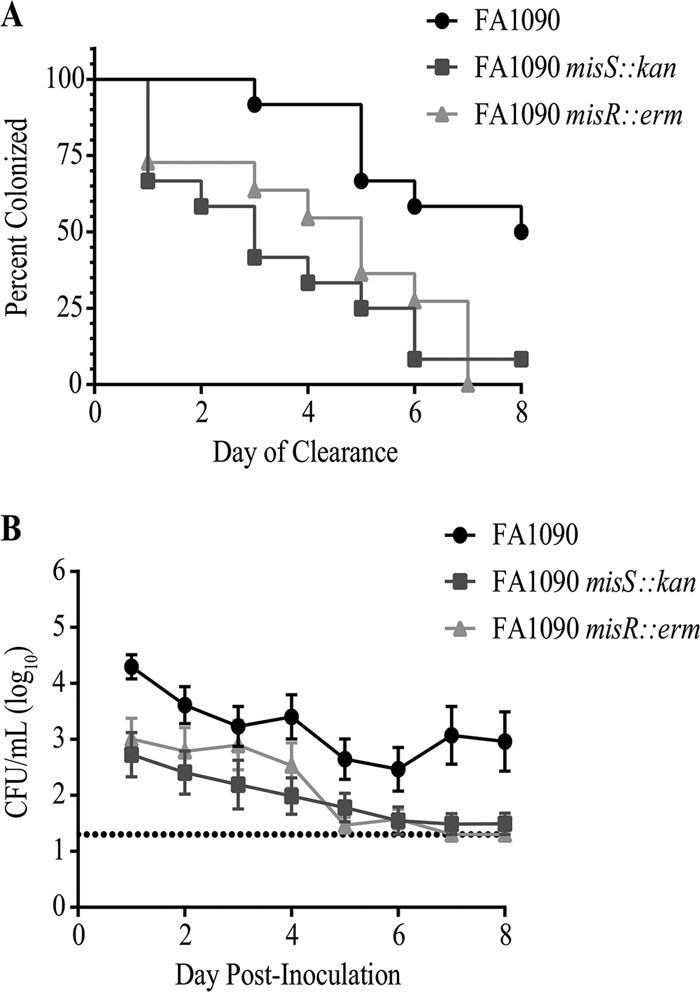
Noncompetitive infection of BALB/c mice with the FA1090, FA1090 misS::kan, and FA1090 misR::erm strains. (A) Percentage of BALB/c mice colonized with the indicated N. gonorrhoeae strains over time. Statistical significance was observed between the capacities of the WT strain and the misS (P = 0.004) and misR (P = 0.002) mutant strains to maintain colonization (as determined by a log rank [Mantel-Cox] test). (B) Colonization load in mice that maintained infection for 8 days, expressed as average log10 CFU per milliliter of vaginal swab suspension. Significantly fewer CFU were recovered over time from mice inoculated with either mutant than from mice inoculated with WT bacteria. (P = 0.005 and 0.003 for misR and misS mutant bacteria, respectively, as determined by 2-way ANOVA with Bonferroni's multiple-comparison test).
RNA-Seq analysis of MisR- and MisS-dependent transcriptomes.
To better understand the mechanisms underlying the reduced fitness of the misR and misS mutants in vivo, we next defined the transcriptomes of WT FA1090 (physiological level of MisRS activation) and the misR and misS mutants in glucose-containing medium. In other organisms, CpxRA is maximally activated in stationary phase (51, 52); as N. gonorrhoeae autolyses in stationary phase, we used late-log-phase organisms for the RNA-Seq experiments. To identify genes differentially expressed between different strains, we used a false discovery rate (FDR) of ≤0.1 and a 2-fold change as criteria. Using these criteria, 157, 139, and 17 genes belonging to different COG functional categories were differentially expressed between the WT and the misR mutant, between the misS mutant and the misR mutant, and between the misS mutant and the WT, respectively (Fig. 3 and Table 1; see also Tables S2 and S3 in the supplemental material). Of the 157 and 139 genes differentially expressed in comparisons of the WT versus the misR mutant and the misS mutant versus the misR mutant, 111 genes were shared (Fig. 3 and Table S2). Totals of 46 and 28 genes were uniquely differentially expressed in comparisons of the WT versus the misR mutant and the misS mutant versus the misR mutant, respectively; these differentially expressed genes belonged to various functional categories (Table S2). Taken together, these data suggest that the deletion of misR leads to widespread differential gene regulation compared to the WT, while the deletion of misS does not. This finding is opposite of those for the CpxRA systems in H. ducreyi and Vibrio cholerae, in which cpxA deletion leads to widespread differential gene regulation, while the deletion of cpxR does not (51, 59).
FIG 3.
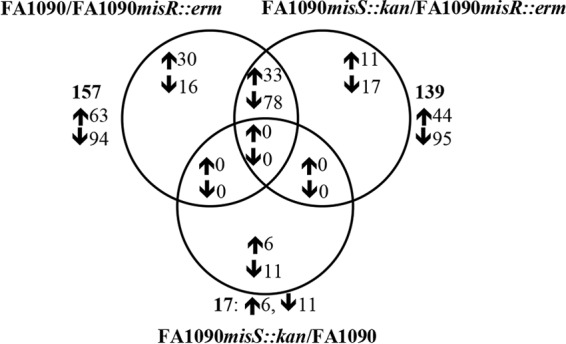
Venn diagram showing the numbers of genes differentially expressed in FA1090 versus FA1090 misR::erm, FA1090 misS::kan versus FA1090 misR::erm, and FA1090 misS::kan versus FA1090. The up- and downregulated genes or operons are indicated by up and down arrows, respectively. The total numbers of genes differentially expressed are indicated in boldface type outside the Venn diagram.
TABLE 1.
Summary of COG functional classifications of genes differentially expressed in the WT relative to the misR mutant, the misS mutant relative to the misR mutant, and the WT relative to the misS mutant
| COG functional class and category | No. of genes differentially expressed |
|||||
|---|---|---|---|---|---|---|
| WT/misR mutant |
misS mutant/misR mutant |
misS mutant/WT |
||||
| Up | Down | Up | Down | Up | Down | |
| Cellular processes and signaling | ||||||
| Cell cycle control, cell division, and chromosome partitioning | 0 | 0 | 0 | 0 | 0 | 0 |
| Cell wall/membrane/envelope biogenesis | 6 | 14 | 4 | 15 | 2 | 4 |
| Defense mechanisms | 2 | 6 | 1 | 7 | 0 | 0 |
| Intracellular trafficking, secretion, and vesicular transport | 0 | 1 | 0 | 2 | 0 | 0 |
| Posttranslational modification, protein turnover, and chaperones | 2 | 9 | 0 | 10 | 0 | 2 |
| Signal transduction mechanisms | 2 | 0 | 1 | 1 | 0 | 2 |
| Information storage and processing | ||||||
| Phage-derived proteins, transposases, and other mobilome components | 18 | 0 | 11 | 0 | 0 | 0 |
| Replication, recombination, and repair | 2 | 1 | 0 | 2 | 0 | 0 |
| RNA processing and modification | 0 | 1 | 0 | 1 | 0 | 0 |
| Transcription | 2 | 2 | 1 | 2 | 0 | 0 |
| Translation, ribosomal structure, and biogenesis | 1 | 4 | 0 | 4 | 1 | 0 |
| Metabolism | ||||||
| Amino acid transport and metabolism | 4 | 12 | 3 | 10 | 0 | 0 |
| Carbohydrate transport and metabolism | 2 | 3 | 1 | 1 | 0 | 0 |
| Coenzyme transport and metabolism | 0 | 1 | 1 | 1 | 0 | 1 |
| Energy production and conversion | 2 | 14 | 1 | 12 | 0 | 0 |
| Inorganic ion transport and metabolism | 1 | 6 | 2 | 3 | 3 | 0 |
| Lipid transport and metabolism | 2 | 3 | 2 | 3 | 0 | 0 |
| Nucleotide transport and metabolism | 1 | 1 | 1 | 2 | 0 | 0 |
| Secondary metabolite biosynthesis, transport, and catabolism | 0 | 0 | 1 | 0 | 0 | 0 |
| Poorly characterized | ||||||
| Function unknown | 1 | 10 | 3 | 10 | 0 | 0 |
| General function prediction only | 3 | 2 | 1 | 4 | 0 | 1 |
| Unclassified | 12 | 4 | 10 | 5 | 0 | 1 |
| Total | 63 | 94 | 44 | 95 | 6 | 11 |
qRT-PCR confirms RNA-Seq data.
As discussed below, the genes differentially expressed in the comparison of the misS mutant versus the misR mutant may best represent the activation versus the inactivation of the system; therefore, we concentrated on this data set for the remainder of this study. We validated the differentially regulated targets using quantitative reverse transcriptase PCR (qRT-PCR). Representative genes were selected based on their expression level, up- and downregulation, and fold change. A total of 13 genes were evaluated by qRT-PCR (Fig. 4A). qRT-PCR confirmed the expression of 13/13 targets identified by RNA-Seq (Fig. 4A). The fold changes derived from RNA-Seq were in good agreement with the fold changes derived from qRT-PCR (R2 = 0.89) (Fig. 4B).
FIG 4.
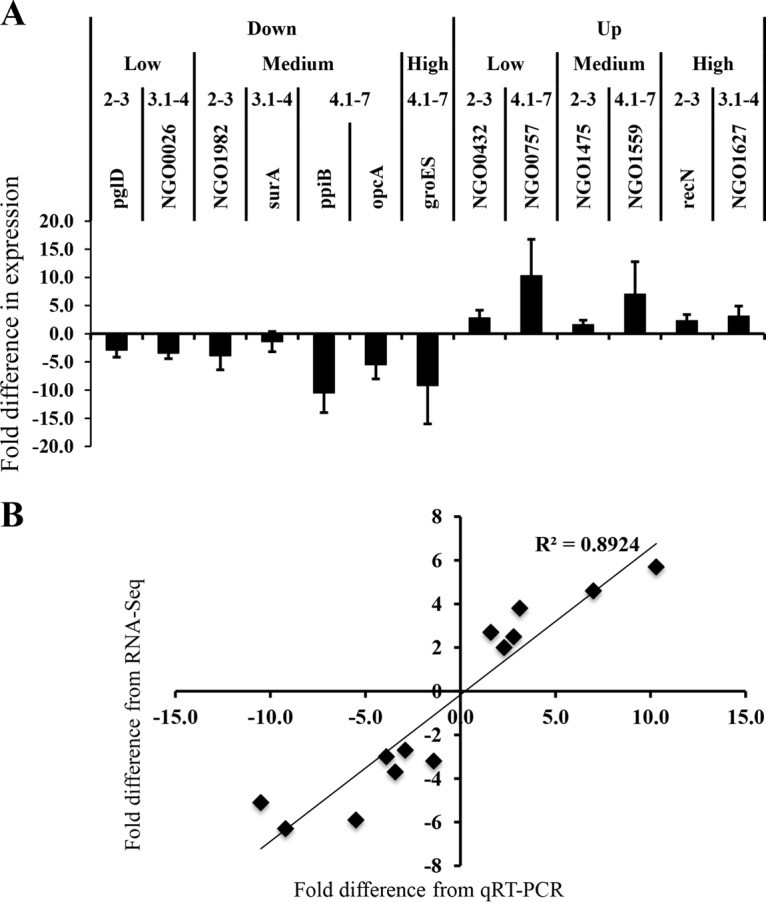
qRT-PCR validation of RNA-Seq data. (A) Fold change in expression levels of target genes in the misS mutant relative to the misR mutant. The expression levels of target genes were normalized to that of rps15. The data represent the means ± standard deviations from four independent experiments. (B) Correlation between the fold changes derived from qRT-PCR and those derived from RNA-Seq.
Inability to define a MisR-binding motif.
In E. coli, CpxR binds to its targets using the consensus sequence GTAAA(N5)GTAAA (60). However, in the meningococcus, MisR binds to its targets using the consensus sequence (A/T)(A/T)TGTAA(A/G/C)G (43). Our de novo motif analysis of the 450-bp putative promoter regions of MisRS-dependent targets did not yield any motif that was similar to that of either E. coli or N. meningitidis. A genome-wide search of the gonococcal genome using the position-specific scoring matrix derived from the N. meningitidis MisRS consensus sequence showed that only 16 out of 104 MisRS-dependent operons/genes contained the meningococcal motif (see Table S4 in the supplemental material), compared to 610 out of 1,274 non-MisRS-dependent operons/genes. There are three possible reasons for the lack of enrichment of the meningococcal MisR-binding motif in the gonococcal MisRS targets. First, gonococcal MisR may utilize a different consensus sequence than that of the meningococcus to bind to its targets. Second, unlike the CpxR-binding motif in E. coli, the majority of the residues in the meningococcal MisR-binding motif are not well conserved; this may explain why we were unable to find a motif similar to that of N. meningitidis in our de novo motif search. Finally, the set of differentially regulated genes identified in this study may represent both direct and indirect targets.
MisRS represses the majority of its targets that encode envelope-localized proteins and does not appear to upregulate genes encoding envelope protein folding and chaperoning factors.
One of the major functions of CpxRA is to downregulate protein traffic across the cell envelope (48, 51). Comparison of the transcriptome of the misS mutant to that of the misR mutant identified 139 differentially expressed genes; of these, 68% (95 genes) were downregulated (Table S2). The majority of the downregulated genes encode either envelope-localized proteins or proteins associated with envelope functions. This finding is very similar to those for H. ducreyi, in which the activation of CpxRA also leads to the downregulation of nearly 70% of its targets, particularly those encoding envelope-localized and/or -associated proteins (51). Taken together, these data suggest that, similarly to H. ducreyi, gonococcal MisRS represses the majority of its targets, especially those encoding envelope-localized and/or -associated proteins.
Another function of the CpxRA system in E. coli is to upregulate genes encoding envelope protein folding and chaperoning factors, such as degP, dsbA, ppiA, and fkpA (48, 49). Transcriptome analysis showed that the gonococcal degP, dsbA, ppiA, and fkpA genes were not differentially expressed in the misS mutant compared to the misR mutant (Table S2). This finding is very similar to those for H. ducreyi, in which the activation of CpxRA also does not upregulate genes encoding envelope protein folding and chaperoning factors. Together, these data suggest that, unlike E. coli and similar to H. ducreyi, gonococcal MisRS does not appear to increase the transcription of genes involved in envelope protein folding and chaperoning.
Comparison of MisRS-dependent genes to those required for infection in murine and human models.
Six genes, individually or in combination with other genes, increase colonization or survival (lst, expression of at least one opa, lptA, mtrA, and mntC) or are required for infection (mtrCDE) in the murine model (10, 61, 62). Similarly, five genes individually or in combination with other genes enhance infection (lptA, lst, pilE, and recA) or are required for infection (tbpA-tbpB in a lactoferrin receptor-deficient strain) in the human challenge model (8, 63–65). Except for opaD, which was downregulated, none of the genes that increase colonization or survival or that are required for infection in murine and human models were differentially expressed in the misS mutant compared to the misR mutant. These data suggest that MisRS does not appear to regulate established virulence determinants in N. gonorrhoeae but may regulate novel virulence factors that have not yet been characterized. The demonstration that mutants of the MisR-MisS system were highly attenuated during noncompetitive infections, as are MtrCDE mutants, strongly supports the importance of this two-component regulatory system in this infection model.
MisR contributes to FA1090 serum resistance.
Serum resistance plays an essential role in the pathogenesis of N. gonorrhoeae. FA1090 is intrinsically serum resistant because its porin protein binds C4B-binding protein and factor H (24, 25). Compared to the wild type, the porin protein P1 was downregulated 2-fold in the misR mutant but was not differentially regulated in the misS mutant (see Tables S2 and S3 in the supplemental material). Therefore, we assessed the survival rates of the FA1090, FA1090 misS::kan, and FA1090 misR::erm strains in 50% pooled complement-replete normal human serum (NHS). The misR mutant survived at a lower rate than the WT strain (P = 0.023), but the misS mutant did not (Fig. 5A). As controls for these experiments, all 3 strains grew (range, 105 to 135%) in heat-inactivated NHS. These results suggest that MisR contributes to serum resistance by upregulating porin expression.
FIG 5.

Effect of deletions of misR and misS on gonococcal survival in the presence of normal human serum (A) and heat shock (B). (A) Percent survival of FA1090, FA1090 misS::kan, and FA1090 misR::erm cells after exposure to 50% pooled normal human serum. Percent survival was calculated as follows: (geometric mean CFU after exposure to active NHS/geometric mean CFU before exposure to active NHS) × 100. (B) Percent survival of FA1090, FA1090 misS::kan, and FA1090 misR::erm cells after exposure to heat shock at 42°C. Percent survival was calculated as follows: (geometric mean CFU after heat shock/geometric mean CFU before heat shock) × 100. The data are means ± standard deviations from four independent experiments. *, P < 0.05; **, P < 0.01.
MisR and MisS are required for the gonococcus to resist heat shock but not oxidative stress.
The heat shock response is characterized by the upregulation of several proteases, chaperones, and regulatory proteins such as RpoH (37, 66). The gonococcus is likely to be exposed to febrile temperatures in some patients with upper reproductive tract infection or systemic infection. Comparison of the transcriptome of the misS mutant to that of the misR mutant showed that homologs of several genes involved in the heat shock response were differentially expressed (Table S2). Therefore, to determine if MisR and MisS contributed to gonococcal survival under heat shock, we compared the survival rates of the FA1090, FA1090 misS::kan, and FA1090 misR::erm strains after heat shock at 42°C. In these assays, both the misR and misS mutants survived at significantly lower rates than the WT strain (P < 0.01) (Fig. 5B). These results suggest that both MisR and MisS are required for the gonococcus to survive under heat shock.
A hallmark of symptomatic gonococcal infection is the local infiltration of polymorphonuclear neutrophils; thus, the organism is likely exposed to extracellular oxidative free radicals from neutrophils (13, 18). In addition, the gonococcus can also be exposed to oxidative free radicals generated intracellularly during metabolism. Extracellular and intracellular oxidative stresses can be mimicked in the laboratory by using hydrogen peroxide and paraquat, respectively. Homologs of several genes involved DNA damage and oxidative stress survival were differentially expressed in the misS mutant compared to the misR mutant (Table S2). Therefore, to determine if MisR and MisS contributed to gonococcal survival under oxidative stress, we compared the survival rates of the FA1090, FA1090 misS::kan, and FA1090 misR::erm strains after exposure to different concentrations of hydrogen peroxide and paraquat. In these assays, both the misR and misS mutants survived at rates similar to that of the WT strain (data not shown). Thus, we conclude that MisR and MisS do not appear to play a role in gonococcal resistance to H2O2- and paraquat-induced oxidative stress.
DISCUSSION
To establish successful infection, N. gonorrhoeae must be able to sense and respond to various stresses and microenvironments in its host. Gram-negative bacteria typically employ TCSs to sense and respond to stresses and changes in their microenvironments. Here, we explored the contributions of the inactivation of misR and misS to gonococcal infection in a murine model and defined MisR- and MisS-dependent genes using RNA-Seq.
The inactivation of MisRS (misR mutant) reduced gonococcal fitness and colonization in the murine genital tract. The in vivo defects correlated with the reduced ability of the misR mutant to survive heat shock and killing by pooled normal human serum. Compared to the WT, the inactivation of misR differentially regulated 157 genes; a large number of these genes encoded envelope-localized proteins and/or other proteins associated with envelope functions. Compared to the WT, an FA19 gonococcal misR mutant is more sensitive to CAMPs and aminoglycosides and exhibits increased membrane permeability (42). Based on these findings, we hypothesize that MisR increases gonococcal fitness in the murine genital tract by maintaining membrane integrity during times of stress, which could protect the gonococcus from host antimicrobial factors.
By comparing the transcriptome of a misR mutant to that of its parent strain, FA19, in late log phase, a previous RNA-Seq analysis identified 95 genes that were expressed at a ≥2-fold difference (42, 67, 68). Of these 95 genes, 55 were upregulated and 40 were downregulated. In contrast, examination of the 157 genes differentially expressed in the misR mutant compared to FA1090 showed that 63 genes were upregulated and 94 genes were downregulated. Comparison of the MisR-dependent genes identified in FA1090 to those identified in FA19 showed that only 15 genes were identified by both studies. Multiple differences between FA19 and FA1090 and other technical reasons may account for these discrepant results. For example, mtrCDE is inducible in FA19 by the transcriptional activator MtrA but is not inducible in FA1090 due to an 11-bp deletion in mtrA (69). Similarly, the promoter sequence of tbpBA (genes regulated by MisR in FA19 but not FA1090) differs between FA19 and FA1090 (67). To ameliorate a growth defect seen in the misR mutant, the FA19 strains were grown in the presence of 10 mM MgCl2; in our study, the misR mutant had no growth defect, and the FA1090 strains were grown in the absence of MgCl2. Each study used different techniques for library preparation and different sequencing platforms (67); even when the same strains are the source of RNA, these variables yield different RNA-Seq results (70). In addition, that study of FA19 used a t test with a P value of <0.05 after Bonferroni correction as a criterion for differential expression, while we used an FDR of <0.1. Finally, while complementation of the FA19 misR mutant restored the expression of selected genes and phenotypes to wild-type levels (42, 68), we did not complement the FA1090 misR or misS mutants; there is the possibility that some of the gene expression differences observed in our study could have been caused by secondary mutations. In both studies, the transcript levels of a subset of differentially expressed genes were verified by qRT-PCR, suggesting that both RNA-Seq data sets are valid.
The misS mutant differentially expressed only 17 genes compared to the WT. However, compared to the misR mutant, the misS mutant differentially regulated 139 genes. Comparison of the genes that were differentially expressed in the misS mutant versus the misR mutant with those that were differentially expressed in the WT versus the misR mutant showed that the majority of the genes were shared between the two comparisons. Analysis of the fold changes of the shared genes in the misS mutant versus the misR mutant and the WT versus the misR mutant also showed that they were highly correlated (R2 = 0.8). Thus, the deletion of misS does not lead to widespread changes in gene transcription compared to the WT under the growth conditions tested.
In contrast to these findings, when E. coli and H. ducreyi cpxA mutants are grown in the presence of glucose to early stationary phase, the CpxRA system is constitutively activated due to a lack of CpxA phosphatase activity and the ability of CpxR to accept phosphate groups from acetyl phosphate, which is made by the AckA-Pta pathway (51, 52). In this study, we grew gonococcal strains in the presence of 0.4% glucose. The FA1090 genome also contains an intact AckA-Pta pathway. Provided that MisR can accept phosphate groups from acetyl phosphate, the deletion of misS may lead to the constitutive activation of MisR. If this is the case, our data suggest that the MisRS system may be highly activated in FA1090 under the growth conditions tested. The deletion of H. ducreyi mtrC leads to the activation of the CpxRA regulon (72). Since FA1090 cannot induce the expression of mtrCDE, it is possible that the lack of induction leads to periplasmic membrane stress and activation of the system in late log phase. This could also account for the lack of overlap in the MisR-regulated genes between the 2 strains.
The deletion of misS also reduced gonococcal fitness and colonization in the murine model and impaired the ability of the organism to survive heat shock. Whether the levels of MisRS activation in the WT and the misS mutant in vivo are similar to those under laboratory growth conditions is unknown. The reduced fitness of the misS mutant in the murine genital tract may be due to the uncontrolled activation (“all-on” state) of MisRS in this mutant. Alternatively, the reduced fitness may simply reflect a lack of the ability of the misS mutant to sense and respond to environmental cues that activate or deactivate the system.
We conclude that an intact MisRS system is critical for the gonococcus to establish successful infection in the mouse model of gonococcal infection of the female genital tract. CpxRA activation primarily affects genes encoding envelope-localized and/or -associated factors, many of which could be potential virulence determinants or vaccine targets; characterization of MisRS-downregulated targets could identify novel gonococcal virulence determinants and vaccine targets. Pharmacological activation of CpxRA is being pursued as a novel antimicrobial/antivirulence strategy for drug-resistant Gram-negative pathogens (73); as antimicrobial-resistant gonococcal strains are emerging at an alarming rate, it will be interesting to explore whether pharmacological activation or inhibition of MisRS could be an antimicrobial/antivirulence strategy for the gonococcus. Infection of mice with a meningococcal misR mutant stimulates a protective immune response, suggesting that a misR mutant could serve as a live attenuated vaccine for meningococcal infection (74); the potential of misR and misS mutants as live attenuated vaccines for gonococcal infection is also worth investigating.
MATERIALS AND METHODS
Bacterial strains and culture conditions.
The bacterial strains and plasmids used in the present study are listed in Table 2. N. gonorrhoeae strains were grown on gonococcal agar (GC agar) plates supplemented with Kellogg's supplement I and 12 μM Fe(NO3)3 at 37°C with 5% CO2 or in gonococcal broth supplemented with Kellogg's supplement I, 12 μM Fe(NO3)3, and 0.043% (wt/vol) sodium bicarbonate at 37°C with 5% CO2 and shaking at 100 rpm. Chemically competent cells of E. coli strains DH5α and One Shot Top10 (Invitrogen) were used for general cloning purposes. E. coli strains were grown in Luria-Bertani medium at 37°C except for strain DY380, which was maintained in L broth or agar and grown at 32°C or 42°C for the induction of the λ red recombinase. Where necessary, the media were supplemented with ampicillin (50 μg/ml for E. coli), kanamycin (50 μg/ml for both E. coli and N. gonorrhoeae), streptomycin (100 μg/ml for N. gonorrhoeae), and/or erythromycin (300 μg/ml for E. coli and 0.5 μg/ml for N. gonorrhoeae).
TABLE 2.
Bacterial strains and plasmids used in this study
| Strain or plasmid | Description | Source or reference |
|---|---|---|
| Strains | ||
| N. gonorrhoeae | ||
| FA1090 | Streptomycin-resistant WT strain | 80 |
| misS mutant | FA1090 misS::kan insertion mutant | This study |
| misR mutant | FA1090 misR::erm insertion mutant | This study |
| E. coli | ||
| DH5α and TOP10 | Strains used for general cloning procedures | Invitrogen |
| DY380 | DH10B derivative containing a defective λ prophage in which the red, bet, and gam genes are controlled by the temperature-sensitive λcI857 repressor | 81 |
| Plasmids | ||
| pGEM-T easy | Cloning vector | Invitrogen |
| pKF20 | pGEM-T Easy containing the misS gene along with flanking sequences | This study |
| pUC18,KmR | Vector containing the kanamycin resistance cassette | 82 |
| pKF21 | pKF20 with the misS gene insertionally inactivated by a kanamycin resistance cassette | This study |
| pKF22 | pGEM-T Easy containing the misR gene along with flanking sequences | This study |
| pGCC3 | Vector containing the erythromycin resistance cassette | 83 |
| pKF23 | pKF22 with the misR gene insertionally inactivated by an erythromycin resistance cassette | This study |
Construction of misS and misR mutants.
Insertional inactivation mutants of misS and misR were constructed in FA1090 as described below. All the primers used for the construction of mutants are listed in Table S1 in the supplemental material. The misS gene was amplified along with 432-bp upstream and 248-bp downstream sequences by using primers P1 and P2. The amplified fragment was cloned into the pGEM-T Easy vector, and the resulting construct was named pKF20. The kanamycin resistance cassette from pUC18,KmR, which is designed to introduce nonpolar insertions, was excised by using SmaI and ligated into similarly digested pKF20; the resulting construct was designated pKF21. pKF21 was linearized and transformed into FA1090; insertionally inactivated misS clones were selected on GC agar with kanamycin as described previously (61). For the construction of an insertional inactivation mutant of misR, the misR gene was amplified along with 345-bp upstream and 390-bp downstream sequences by using primers P3 and P4. The amplified fragment was cloned into the pGEM-T Easy vector, generating pKF22, which was then electroporated into DY380, a strain that contains the temperature-sensitive λ red recombinase. An erythromycin resistance cassette was amplified from pGCC3 by using primers P5 and P6, and the amplified fragment was electroporated into DY380 containing pKF22 for recombination. Following recombination, the misR gene in pKF22 was insertionally inactivated, and the resulting construct was named pKF23. pKF23 was linearized and transformed into FA1090, and the insertionally inactivated misR clones were selected on GC agar with erythromycin as described previously (61). The final misS::kan and misR::erm mutants were confirmed by sequencing.
Experimental murine infection.
All animal experiments were conducted according to the guidelines of the Association for the Assessment and Accreditation of Laboratory Animal Care and the Uniformed Services University Institutional Animal Care and Use Committee. Female BALB/c mice (6 to 8 weeks old; National Cancer Institute) were used for competitive-infection and noncompetitive-infection experiments. Mice were treated with water-soluble 17β-estradiol or implanted with a 21-day, slow-release 17β-estradiol pellet, respectively, to induce long-term susceptibility to N. gonorrhoeae and antibiotics (streptomycin, vancomycin, and trimethoprim) to reduce the overgrowth of commensal flora that occurs under the influence of estradiol, as described previously (10). For competitive infections, groups of mice (n = 7 per group) were inoculated intravaginally with 106 CFU of WT FA1090 bacteria combined with similar numbers of misS mutant or misR mutant bacteria (total dose, 2 × 106 CFU). Starting on day 2 postinoculation, vaginal swabs were collected every other day for 8 days and suspended in 100 μl of GC broth. To determine total CFU, vaginal swab suspensions and inocula were cultured quantitatively on GC agar with streptomycin; misS or misR mutant CFU were isolated on GC agar with streptomycin plus kanamycin or streptomycin plus erythromycin, respectively. The numbers of WT CFU were calculated by subtracting the number of kanamycin- or erythromycin-resistant CFU from the total number of CFU. The limit of detection (20 CFU) was used when no mutant bacteria were recovered from an animal. Data from mice from which cultures yielded <10 CFU (total number) were not included in the analysis. Results were expressed as the competitive index (CI) for individual mice by using the following equation: the number of mutant CFU over the number of WT CFU (output) divided by the number of mutant CFU over the number of WT CFU (input).
For noncompetitive infections, groups of mice (n = 11 to 12 per group) were inoculated vaginally with either FA1090, the misS mutant, or the misR mutant (∼0.5 × 106 to 1 × 106 CFU/mouse). Vaginal swabs were collected daily for 8 consecutive days postinoculation, and the swab material was suspended in 1 ml of phosphate-buffered saline (PBS). Serial dilutions were performed with GC broth, and dilutions were plated onto GC agar with streptomycin. Plates were incubated at 35°C with 7% CO2, and the number of CFU was determined after 48 h.
RNA-Seq analysis.
Total RNA was extracted from four independent cultures of the FA1090, FA1090 misS::kan, and FA1090 misR::erm strains grown to late log phase (optical density at 600 nm [OD600] of 0.8 to 1.0) by using TRIzol reagent (Invitrogen) according to the manufacturer's protocol. RNA was treated twice with Turbo DNA-free DNase (Ambion), and the efficacy of DNase treatment was confirmed by RT-PCR analysis of rps15 with primers P7 and P8 (see Table S1 in the supplemental material) and the QuantiTect SYBR green RT-PCR kit (Qiagen). The removal of rRNA from total RNA and the preparation of RNA-Seq libraries were performed by using the ScriptSeq Complete kit (Illumina, Inc.) according to the manufacturer's instructions. Libraries were sequenced on the Illumina NextSeq platform for single-end sequencing with read lengths of 75 bp at the Center for Genomics and Bioinformatics (Bloomington, IN). Illumina adapter sequences and low-quality base calls were removed with Trim Galore (http://www.bioinformatics.babraham.ac.uk/projects/trim_galore/). The resulting high-quality reads were aligned to the N. gonorrhoeae FA1090 reference genome (GenBank accession no. AE004969) by using bowtie2 (v2.2.6) (51, 75). Gene expression was quantified by using HTseq (76). Reads with a mapping quality of <10 were excluded during gene expression quantification. Differentially expressed genes were identified by using a prespecified FDR of ≤0.1 and a 2-fold change as the criteria and classified by using features from the annotated FA1090 genome and the COG database (77).
qRT-PCR.
qRT-PCR was performed as described previously, using the QuantiTect SYBR green RT-PCR kit (Qiagen) with an ABI Prism 7000 sequence detection system (Applied Biosystems) (51). Primers (P7 to P34) used for qRT-PCR are listed in Table S1 in the supplemental material. Expression levels of selected genes were normalized to that of rps15, which was not differentially expressed by the activation or inactivation of MisRS.
Motif identification.
De novo identification of the MisR-binding consensus sequence was attempted by using the Multiple EM for Motif Elicitation (MEME) algorithm (78). Due to the lack of experimentally validated MisRS targets in N. gonorrhoeae, the 450-bp upstream putative promoter regions of 104 putative operons/genes differentially expressed in the comparison of the misS mutant versus the misR mutant were used for de novo motif identification by the MEME algorithm with the motif length restricted to 15 to 50 bp. For a genome-wide motif scan, a position-specific scoring matrix was generated from the 10 experimentally validated MisR-binding motifs of N. meningitidis and used to scan the 450-bp putative promoter regions of all the putative operons/genes in the gonococcal genome by using Virtual Footprint (79).
In vitro assays.
Serum bactericidal assays were performed as described previously (24, 54), except that pooled complement-replete NHS purchased from a commercial source (Quidel Corporation) was used in the assays. Briefly, the FA1090, FA1090 misS::kan, and FA1090 misR::erm strains were grown on GC agar plates for 16 to 18 h and harvested into GC broth. Approximately 2,000 CFU of each strain were mixed with either active or heat-inactivated (56°C for 35 min) NHS to a final concentration of 50%. After incubation at 37°C for 45 min, the samples were quantitatively cultured in triplicate. The data were reported as percent survival in NHS compared to that in the inoculum [(geometric mean CFU in NHS/geometric mean CFU in the inoculum) × 100]. For oxidative stress survival assays, approximately 107 cells of late-log-phase (OD600 = 0.8 to 1.0)-grown FA1090, FA1090 misS::kan, and FA1090 misR::erm strains were either treated or not treated with 0.2, 2.0, or 20.0 mM H2O2 (Sigma-Aldrich) or 0.2, 2.0, or 20.0 mM paraquat (Sigma-Aldrich) at 37°C for 15 min in GC broth and quantitatively cultured. For heat shock survival assays, approximately 107 late-log-phase cells of the FA1090, FA1090 misS::kan, and FA1090 misR::erm strains grown at 37°C were either shifted to 42°C or maintained at 37°C for 1 h in GC broth and quantitatively cultured. For both oxidative stress and heat shock survival assays, percent survival was determined by calculating the ratio of recovered CFU to initial CFU. All the phenotypic assays were repeated four times, and the data obtained were analyzed by using one-way analysis of variance (ANOVA) with Tukey's honestly significant different post hoc test. A P value of <0.05 was considered statistically significant.
Accession number(s).
Data from the RNA-Seq experiments were deposited at the NCBI Gene Expression Omnibus (GEO) database under accession numbers GSM2199875 to GSM2199886 (raw data) and GSE83341 (processed data).
Supplementary Material
ACKNOWLEDGMENTS
This work was supported by U.S. Public Health Service grants R01 AI42053 and U19 AI113170 to A.E.J. and by a Project Development Team within the Indiana CTSI NIH/NCRR grant number UL1TR001108 to S.M.S. RNA sequencing was performed by the Center for Genomics and Bioinformatics at Indiana University, Bloomington, IN, a core facility of the Indiana CTSI.
We thank Margaret Bauer for her thoughtful criticism of the manuscript.
Footnotes
Supplemental material for this article may be found at https://doi.org/10.1128/IAI.00307-17.
REFERENCES
- 1.Satterwhite CL, Torrone E, Meites E, Dunne EF, Mahajan R, Ocfemia MC, Su J, Xu F, Weinstock H. 2013. Sexually transmitted infections among US women and men: prevalence and incidence estimates, 2008. Sex Transm Dis 40:187–193. doi: 10.1097/OLQ.0b013e318286bb53. [DOI] [PubMed] [Google Scholar]
- 2.Hook EW, Handsfield HH. 2008. Gonococcal infection in the adult, p 627–645. In Holmes KK. (ed), Sexually transmitted diseases. McGraw-Hill, New York, NY. [Google Scholar]
- 3.Chen A, Boulton IC, Pongoski J, Cochrane A, Gray-Owen SD. 2003. Induction of HIV-1 long terminal repeat-mediated transcription by Neisseria gonorrhoeae. AIDS 17:625–628. doi: 10.1097/00002030-200303070-00019. [DOI] [PubMed] [Google Scholar]
- 4.Cohen MS, Hoffman RA, Royce RA, Kazembe P, Dyer JR, Daly CC, Zimba D, Vernazza PL, Maida M, Fiscus SA, Eron JJJ. 1997. Reduction of concentration of HIV-1 in semen after treatment of urethritis: implications for prevention of sexual transmission of HIV-1. Lancet 349:1868–1873. doi: 10.1016/S0140-6736(97)02190-9. [DOI] [PubMed] [Google Scholar]
- 5.Levine WC, Pope V, Bhoomkar A, Tambe P, Lewis JS, Zaidi AA, Farshy CE, Mitchell S, Talkington DF. 1998. Increase in endocervical CD4 lymphocytes among women with nonulcerative sexually transmitted diseases. J Infect Dis 177:167–174. doi: 10.1086/513820. [DOI] [PubMed] [Google Scholar]
- 6.Unemo M, Shafer WM. 2014. Antimicrobial resistance in Neisseria gonorrhoeae in the 21st century: past, evolution, and future. Clin Microbiol Rev 27:587–613. doi: 10.1128/CMR.00010-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 7.Jerse AE, Deal CD. 2013. Vaccine research for gonococcal infections: where are we? Sex Transm Infect 89(Suppl 4):iv63–iv68. doi: 10.1136/sextrans-2013-051225. [DOI] [PubMed] [Google Scholar]
- 8.Hobbs MM, Sparling PF, Cohen MS, Shafer WM, Deal CD, Jerse AE. 2011. Experimental gonococcal infection in male volunteers: cumulative experience with Neisseria gonorrhoeae strains FA1090 and MS11mkC. Front Microbiol 2:123. doi: 10.3389/fmicb.2011.00123. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 9.Edwards JL, Apicella MA. 2004. The molecular mechanisms used by Neisseria gonorrhoeae to initiate infection differ between men and women. Clin Microbiol Rev 17:965–981. doi: 10.1128/CMR.17.4.965-981.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 10.Jerse AE, Wu H, Packiam M, Vonck RA, Begum AA, Garvin LE. 2011. Estradiol-treated female mice as surrogate hosts for Neisseria gonorrhoeae genital tract infections. Front Microbiol 2:107. doi: 10.3389/fmicb.2011.00107. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 11.Rudel T, Boxberger HJ, Meyer TF. 1995. Pilus biogenesis and epithelial cell adherence of Neisseria gonorrhoeae pilC double knock-out mutants. Mol Microbiol 17:1057–1071. doi: 10.1111/j.1365-2958.1995.mmi_17061057.x. [DOI] [PubMed] [Google Scholar]
- 12.Sadarangani M, Pollard AJ, Gray-Owen SD. 2011. Opa proteins and CEACAMs: pathways of immune engagement for pathogenic Neisseria. FEMS Microbiol Rev 35:498–514. doi: 10.1111/j.1574-6976.2010.00260.x. [DOI] [PubMed] [Google Scholar]
- 13.Edwards JL, Butler EK. 2011. The pathobiology of Neisseria gonorrhoeae lower female genital tract infection. Front Microbiol 2:102. doi: 10.3389/fmicb.2011.00102. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 14.Harvey HA, Jennings MP, Campbell CA, Williams R, Apicella MA. 2001. Receptor-mediated endocytosis of Neisseria gonorrhoeae into primary human urethral epithelial cells: the role of the asialoglycoprotein receptor. Mol Microbiol 42:659–672. doi: 10.1046/j.1365-2958.2001.02666.x. [DOI] [PubMed] [Google Scholar]
- 15.Harvey HA, Porat N, Campbell CA, Jennings M, Gibson BW, Phillips NJ, Apicella MA, Blake MS. 2000. Gonococcal lipooligosaccharide is a ligand for the asialoglycoprotein receptor on human sperm. Mol Microbiol 36:1059–1070. doi: 10.1046/j.1365-2958.2000.01938.x. [DOI] [PubMed] [Google Scholar]
- 16.Swanson J. 1973. Studies on gonococcus infections. IV. Pili: their role in attachment of gonococci to tissue culture cells. J Exp Med 137:571–589. doi: 10.1084/jem.137.3.571. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 17.Serino L, Nesta B, Leuzzi R, Fontana MR, Monaci E, Mocca BT, Cartocci E, Masignani V, Jerse AE, Rappuoli R, Pizza M. 2007. Identification of a new OmpA-like protein in Neisseria gonorrhoeae involved in the binding to human epithelial cells and in vivo colonization. Mol Microbiol 64:1391–1403. doi: 10.1111/j.1365-2958.2007.05745.x. [DOI] [PubMed] [Google Scholar]
- 18.Chateau A, Seifert HS. 2016. Neisseria gonorrhoeae survives within and modulates apoptosis and inflammatory cytokine production of human macrophages. Cell Microbiol 18:546–560. doi: 10.1111/cmi.12529. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 19.Edwards JL, Apicella MA. 2005. I-domain-containing integrins serve as pilus receptors for Neisseria gonorrhoeae adherence to human epithelial cells. Cell Microbiol 7:1197–1211. doi: 10.1111/j.1462-5822.2005.00547.x. [DOI] [PubMed] [Google Scholar]
- 20.Bos MP, Grunert F, Belland RJ. 1997. Differential recognition of members of the carcinoembryonic antigen family by Opa variants of Neisseria gonorrhoeae. Infect Immun 65:2353–2361. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 21.van Putten JP, Duensing TD, Carlson J. 1998. Gonococcal invasion of epithelial cells driven by P.IA, a bacterial ion channel with GTP binding properties. J Exp Med 188:941–952. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 22.Rice PA. 1989. Molecular basis for serum resistance in Neisseria gonorrhoeae. Clin Microbiol Rev 2(Suppl):S112–S117. doi: 10.1128/CMR.2.Suppl.S112. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 23.Welsch JA, Ram S. 2008. Factor H and neisserial pathogenesis. Vaccine 26(Suppl 8):I40–I45. doi: 10.1016/j.vaccine.2008.11.060. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 24.Ram S, McQuillen PD, Gulati S, Elkins C, Pangburn MK, Rice PA. 1998. Binding of complement factor H to loop 5 of porin protein 1A: a molecular mechanism of serum resistance of nonsialylated Neisseria gonorrhoeae. J Exp Med 188:671–680. doi: 10.1084/jem.188.4.671. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 25.Ram S, Cullinane M, Blom A, Gulati S, McQuillen D, Monks D, O'Connell C, Boden R, Elkins C, Pangburn M, Dahlback B, Rice P. 2001. Binding of C4b-binding protein to porin. A molecular mechanism of serum resistance of Neisseria gonorrhoeae. J Exp Med 193:281–296. doi: 10.1084/jem.193.3.281. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 26.Stern A, Brown M, Nickel P, Meyer TF. 1986. Opacity genes in Neisseria gonorrhoeae: control of phase and antigenic variation. Cell 47:61–71. doi: 10.1016/0092-8674(86)90366-1. [DOI] [PubMed] [Google Scholar]
- 27.Apicella MA, Shero M, Jarvis GA, Griffiss JM, Mandrell RE, Schneider H. 1987. Phenotypic variation in epitope expression of the Neisseria gonorrhoeae lipooligosaccharide. Infect Immun 55:1755–1761. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 28.Seifert HS, Wright CJ, Jerse AE, Cohen MS, Cannon JG. 1994. Multiple gonococcal pilin antigenic variants are produced during experimental human infections. J Clin Invest 93:2744–2749. doi: 10.1172/JCI117290. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 29.Griffiss JM, Schneider H, Mandrell RE, Yamasaki R, Jarvis GA, Kim JJ, Gibson BW, Hamadeh R, Apicella MA. 1988. Lipooligosaccharides: the principal glycolipids of the neisserial outer membrane. Rev Infect Dis 10(Suppl 2):S287–S295. [DOI] [PubMed] [Google Scholar]
- 30.Mulks MH, Plaut AG. 1978. IgA protease production as a characteristic distinguishing pathogenic from harmless neisseriaceae. N Engl J Med 299:973–976. doi: 10.1056/NEJM197811022991802. [DOI] [PubMed] [Google Scholar]
- 31.Plummer FA, Chubb H, Simonsen JN, Bosire M, Slaney L, Maclean I, Ndinya-Achola JO, Waiyaki P, Brunham RC. 1993. Antibody to Rmp (outer membrane protein 3) increases susceptibility to gonococcal infection. J Clin Invest 91:339–343. doi: 10.1172/JCI116190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 32.Johnson MB, Criss AK. 2013. Neisseria gonorrhoeae phagosomes delay fusion with primary granules to enhance bacterial survival inside human neutrophils. Cell Microbiol 15:1323–1340. doi: 10.1111/cmi.12117. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 33.Isabella VM, Clark VL. 2011. Deep sequencing-based analysis of the anaerobic stimulon in Neisseria gonorrhoeae. BMC Genomics 12:51. doi: 10.1186/1471-2164-12-51. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 34.Fisher SD, Reger AD, Baum A, Hill SA. 2005. RelA alone appears essential for (p)ppGpp production when Neisseria gonorrhoeae encounters nutritional stress. FEMS Microbiol Lett 248:1–8. doi: 10.1016/j.femsle.2005.05.014. [DOI] [PubMed] [Google Scholar]
- 35.Rohde KH, Dyer DW. 2003. Mechanisms of iron acquisition by the human pathogens Neisseria meningitidis and Neisseria gonorrhoeae. Front Biosci 8:d1186–d1218. doi: 10.2741/1133. [DOI] [PubMed] [Google Scholar]
- 36.St Amant DC, Valentin-Bon IE, Jerse AE. 2002. Inhibition of Neisseria gonorrhoeae by Lactobacillus species that are commonly isolated from the female genital tract. Infect Immun 70:7169–7171. doi: 10.1128/IAI.70.12.7169-7171.2002. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 37.Gunesekere IC, Kahler CM, Powell DR, Snyder LA, Saunders NJ, Rood JI, Davies JK. 2006. Comparison of the RpoH-dependent regulon and general stress response in Neisseria gonorrhoeae. J Bacteriol 188:4769–4776. doi: 10.1128/JB.01807-05. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 38.Bauer ME, Shafer WM. 2015. On the in vivo significance of bacterial resistance to antimicrobial peptides. Biochim Biophys Acta 1848:3101–3111. doi: 10.1016/j.bbamem.2015.02.012. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 39.Steichen CT, Shao JQ, Ketterer MR, Apicella MA. 2008. Gonococcal cervicitis: a role for biofilm in pathogenesis. J Infect Dis 198:1856–1861. doi: 10.1086/593336. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 40.Overton TW, Whitehead R, Li Y, Snyder LA, Saunders NJ, Smith H, Cole JA. 2006. Coordinated regulation of the Neisseria gonorrhoeae-truncated denitrification pathway by the nitric oxide-sensitive repressor, NsrR, and nitrite-insensitive NarQ-NarP. J Biol Chem 281:33115–33126. doi: 10.1074/jbc.M607056200. [DOI] [PubMed] [Google Scholar]
- 41.Atack JM, Srikhanta YN, Djoko KY, Welch JP, Hasri NH, Steichen CT, Vanden Hoven RN, Grimmond SM, Othman DS, Kappler U, Apicella MA, Jennings MP, Edwards JL, McEwan AG. 2013. Characterization of an ntrX mutant of Neisseria gonorrhoeae reveals a response regulator that controls expression of respiratory enzymes in oxidase-positive proteobacteria. J Bacteriol 195:2632–2641. doi: 10.1128/JB.02062-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 42.Kandler JL, Holley CL, Reimche JL, Dhulipala V, Balthazar JT, Muszynski A, Carlson RW, Shafer WM. 23 May 2016. The MisR response regulator is necessary for intrinsic cationic antimicrobial peptide and aminoglycoside resistance in Neisseria gonorrhoeae. Antimicrob Agents Chemother 60:4690–4700. doi: 10.1128/AAC.00823-16. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 43.Tzeng Y-L, Kahler CM, Zhang X, Stephens DS. 2008. MisR/MisS two-component regulon in Neisseria meningitidis. Infect Immun 76:704–716. doi: 10.1128/IAI.01007-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 44.Tzeng Y-L, Zhou X, Bao S, Zhao S, Noble C, Stephens DS. 2006. Autoregulation of the MisR/MisS two-component signal transduction system in Neisseria meningitidis. J Bacteriol 188:5055–5065. doi: 10.1128/JB.00264-06. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 45.Sannigrahi S, Zhang X, Tzeng YL. 2009. Regulation of the type I protein secretion system by the MisR/MisS two-component system in Neisseria meningitidis. Microbiology 155:1588–1601. doi: 10.1099/mic.0.023945-0. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 46.Zhao S, Montanez GE, Kumar P, Sannigrahi S, Tzeng YL. 2010. Regulatory role of the MisR/S two-component system in hemoglobin utilization in Neisseria meningitidis. Infect Immun 78:1109–1122. doi: 10.1128/IAI.00363-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 47.Jamet A, Rousseau C, Monfort JB, Frapy E, Nassif X, Martin P. 2009. A two-component system is required for colonization of host cells by meningococcus. Microbiology 155:2288–2295. doi: 10.1099/mic.0.027755-0. [DOI] [PubMed] [Google Scholar]
- 48.Raivio TL. 31 October 2013. Everything old is new again: an update on current research on the Cpx envelope stress response. Biochim Biophys Acta doi: 10.1016/j.bbamcr.2013.10.018. [DOI] [PubMed] [Google Scholar]
- 49.Raivio TL, Silhavy TJ. 1997. Transduction of envelope stress in Escherichia coli by the Cpx two-component system. J Bacteriol 179:7724–7733. doi: 10.1128/jb.179.24.7724-7733.1997. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 50.Vogt SL, Raivio TL. 2012. Just scratching the surface: an expanding view of the Cpx envelope stress response. FEMS Microbiol Lett 326:2–11. doi: 10.1111/j.1574-6968.2011.02406.x. [DOI] [PubMed] [Google Scholar]
- 51.Gangaiah D, Zhang X, Fortney KR, Baker B, Liu Y, Munson RS Jr, Spinola SM. 2013. Activation of CpxRA in Haemophilus ducreyi primarily inhibits the expression of its targets, including major virulence determinants. J Bacteriol 195:3486–3502. doi: 10.1128/JB.00372-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 52.Wolfe AJ, Parikh N, Lima BP, Zemaitaitis B. 2008. Signal integration by the two-component signal transduction response regulator CpxR. J Bacteriol 190:2314–2322. doi: 10.1128/JB.01906-07. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 53.Lima BP, Antelmann H, Gronau K, Chi BK, Becher D, Brinsmade SR, Wolfe AJ. 2011. Involvement of protein acetylation in glucose-induced transcription of a stress-responsive promoter. Mol Microbiol 81:1190–1204. doi: 10.1111/j.1365-2958.2011.07742.x. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 54.Spinola SM, Fortney KR, Baker B, Janowicz DM, Zwickl B, Katz BP, Blick RJ, Munson RS Jr. 2010. Activation of the CpxRA system by deletion of cpxA impairs the ability of Haemophilus ducreyi to infect humans. Infect Immun 78:3898–3904. doi: 10.1128/IAI.00432-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 55.Humphreys S, Rowley G, Stevenson A, Anjum MF, Woodward MJ, Gilbert S, Kormanec J, Roberts M. 2004. Role of the two-component regulator CpxAR in the virulence of Salmonella enterica serotype Typhimurium. Infect Immun 72:4654–4661. doi: 10.1128/IAI.72.8.4654-4661.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 56.Labandeira-Rey M, Dodd D, Fortney KR, Zwickl B, Katz BP, Janowicz DM, Spinola SM, Hansen EJ. 2011. A Haemophilus ducreyi cpxR deletion mutant is virulent in human volunteers. J Infect Dis 203:1859–1865. doi: 10.1093/infdis/jir190. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 57.Debnath I, Norton JP, Barber AE, Ott EM, Dhakal BK, Kulesus RR, Mulvey MA. 2013. The Cpx stress response system potentiates the fitness and virulence of uropathogenic Escherichia coli. Infect Immun 81:1450–1459. doi: 10.1128/IAI.01213-12. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 58.Thomassin JL, Giannakopoulou N, Zhu L, Gross J, Salmon K, Leclerc JM, Daigle F, Le Moual H, Gruenheid S. 2015. The CpxRA two-component system is essential for Citrobacter rodentium virulence. Infect Immun 83:1919–1928. doi: 10.1128/IAI.00194-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 59.Slamti L, Waldor MK. 2009. Genetic analysis of activation of the Vibrio cholerae Cpx pathway. J Bacteriol 191:5044–5056. doi: 10.1128/JB.00406-09. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 60.De Wulf P, McGuire AM, Liu X, Lin ECC. 2002. Genome-wide profiling of promoter recognition by the two-component response regulator CpxR-P in Escherichia coli. J Biol Chem 277:26652–26661. doi: 10.1074/jbc.M203487200. [DOI] [PubMed] [Google Scholar]
- 61.Hobbs MM, Anderson JE, Balthazar JT, Kandler JL, Carlson RW, Ganguly J, Begum AA, Duncan JA, Lin JT, Sparling PF, Jerse AE, Shafer WM. 2013. Lipid A's structure mediates Neisseria gonorrhoeae fitness during experimental infection of mice and men. mBio 4:e00892-13. doi: 10.1128/mBio.00892-13. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 62.Packiam M, Yedery RD, Begum AA, Carlson RW, Ganguly J, Sempowski GD, Ventevogel MS, Shafer WM, Jerse AE. 2014. Phosphoethanolamine decoration of Neisseria gonorrhoeae lipid A plays a dual immunostimulatory and protective role during experimental genital tract infection. Infect Immun 82:2170–2179. doi: 10.1128/IAI.01504-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 63.Cohen MS, Cannon JG. 1999. Human experimentation with Neisseria gonorrhoeae: progress and goals. J Infect Dis 179:S375–S379. doi: 10.1086/513847. [DOI] [PubMed] [Google Scholar]
- 64.Schneider H, Schmidt KA, Skillman DR, Van De Verg L, Warren RL, Wylie HJ, Sadoff JC, Deal CD, Cross AS. 1996. Sialylation lessens the infectivity of Neisseria gonorrhoeae MS11mkC. J Infect Dis 173:1422–1427. doi: 10.1093/infdis/173.6.1422. [DOI] [PubMed] [Google Scholar]
- 65.Cornelissen CN, Kelley M, Hobbs MM, Anderson JE, Cannon JG, Cohen MS, Sparling PF. 1998. The transferrin receptor expressed by gonococcal strain FA1090 is required for the experimental infection of human male volunteers. Mol Microbiol 27:611–616. doi: 10.1046/j.1365-2958.1998.00710.x. [DOI] [PubMed] [Google Scholar]
- 66.Yura T, Nagai H, Mori H. 1993. Regulation of the heat-shock response in bacteria. Annu Rev Microbiol 47:321–350. doi: 10.1146/annurev.mi.47.100193.001541. [DOI] [PubMed] [Google Scholar]
- 67.Velez Acevedo RN, Ronpirin C, Kandler JL, Shafer WM, Cornelissen CN. 2014. Identification of regulatory elements that control expression of the tbpBA operon in Neisseria gonorrhoeae. J Bacteriol 196:2762–2774. doi: 10.1128/JB.01693-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 68.Kandler JL, Acevedo RV, Dickinson MK, Cash DR, Shafer WM, Cornelissen CN. 2016. The genes that encode the gonococcal transferrin binding proteins, TbpB and TbpA, are differentially regulated by MisR under iron-replete and iron-depleted conditions. Mol Microbiol 102:137–151. doi: 10.1111/mmi.13450. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 69.Rouquette C, Harmon JB, Shafer WM. 1999. Induction of the mtrCDE-encoded efflux pump system of Neisseria gonorrhoeae requires MtrA, an AraC-like protein. Mol Microbiol 33:651–658. doi: 10.1046/j.1365-2958.1999.01517.x. [DOI] [PubMed] [Google Scholar]
- 70.Thomason MK, Bischler T, Eisenbart SK, Forstner KU, Zhang A, Herbig A, Nieselt K, Sharma CM, Storz G. 2015. Global transcriptional start site mapping using differential RNA sequencing reveals novel antisense RNAs in Escherichia coli. J Bacteriol 197:18–28. doi: 10.1128/JB.02096-14. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 71.Reference deleted.
- 72.Rinker SD, Trombley MP, Gu X, Fortney KR, Bauer ME. 2011. Deletion of mtrC in Haemophilus ducreyi increases sensitivity to human antimicrobial peptides and activates the CpxRA regulon. Infect Immun 79:2324–2334. doi: 10.1128/IAI.01316-10. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 73.van Rensburg JJ, Fortney KR, Chen L, Krieger AJ, Lima BP, Wolfe AJ, Katz BP, Zhang ZY, Spinola SM. 2015. Development and validation of a high-throughput cell-based screen to identify activators of a bacterial two-component signal transduction system. Antimicrob Agents Chemother 59:3789–3799. doi: 10.1128/AAC.00236-15. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 74.Newcombe J, Eales-Reynolds LJ, Wootton L, Gorringe AR, Funnell SG, Taylor SC, McFadden JJ. 2004. Infection with an avirulent phoP mutant of Neisseria meningitidis confers broad cross-reactive immunity. Infect Immun 72:338–344. doi: 10.1128/IAI.72.1.338-344.2004. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 75.Langmead B, Salzberg SL. 2012. Fast gapped-read alignment with Bowtie 2. Nat Methods 9:357–359. doi: 10.1038/nmeth.1923. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 76.Anders S, Pyl PT, Huber W. 2015. HTSeq—a Python framework to work with high-throughput sequencing data. Bioinformatics 31:166–169. doi: 10.1093/bioinformatics/btu638. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 77.Galperin MY, Makarova KS, Wolf YI, Koonin EV. 2015. Expanded microbial genome coverage and improved protein family annotation in the COG database. Nucleic Acids Res 43:D261–D269. doi: 10.1093/nar/gku1223. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 78.Bailey TL, Elkan C. 1994. Fitting a mixture model by expectation maximization to discover motifs in biopolymers, p 28–36. In Altman R, Brutlag D, Karp P, Lathrop R, Searls D (ed), Proceedings of the Second International Conference on Intelligent Systems for Molecular Biology. AAAI Press, Menlo Park, CA. [PubMed] [Google Scholar]
- 79.Munch R, Hiller K, Grote A, Scheer M, Klein J, Schobert M, Jahn D. 2005. Virtual Footprint and PRODORIC: an integrative framework for regulon prediction in prokaryotes. Bioinformatics 21:4187–4189. doi: 10.1093/bioinformatics/bti635. [DOI] [PubMed] [Google Scholar]
- 80.Nachamkin I, Cannon JG, Mittler RS. 1981. Monoclonal antibodies against Neisseria gonorrhoeae: production of antibodies directed against a strain-specific cell surface antigen. Infect Immun 32:641–648. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 81.Lee EC, Yu D, Martinez de Velasco J, Tessarollo L, Swing DA, Court DL, Jenkins NA, Copeland NG. 2001. A highly efficient Escherichia coli-based chromosome engineering system adapted for recombinogenic targeting and subcloning of BAC DNA. Genomics 73:56–65. doi: 10.1006/geno.2000.6451. [DOI] [PubMed] [Google Scholar]
- 82.Menard R, Sansonetti PJ, Parsot C. 1993. Nonpolar mutagenesis of the ipa genes defines IpaB, IpaC, and IpaD as effectors of Shigella flexneri entry into epithelial cells. J Bacteriol 175:5899–5906. doi: 10.1128/jb.175.18.5899-5906.1993. [DOI] [PMC free article] [PubMed] [Google Scholar]
- 83.Mehr IJ, Seifert HS. 1997. Random shuttle mutagenesis: gonococcal mutants deficient in pilin antigenic variation. Mol Microbiol 23:1121–1131. doi: 10.1046/j.1365-2958.1997.2971660.x. [DOI] [PubMed] [Google Scholar]
Associated Data
This section collects any data citations, data availability statements, or supplementary materials included in this article.